© 2012 pearson education, inc. enzymes chapter 6
TRANSCRIPT

© 2012 Pearson Education, Inc.
ENZYMESChapter 6

© 2012 Pearson Education, Inc.
Enzymes: The Catalysts of Life
• Enzyme catalysis: virtually all cellular processes or reactions are mediated by protein (sometimes RNA) catalysts called enzymes
• The presence of the appropriate enzyme makes the difference between whether a reaction can take place and whether it will take place

© 2012 Pearson Education, Inc.
Activation Energy and the Metastable State
• Many thermodynamically feasible reactions in a cell that could occur do not proceed at any appreciable rate (on their own)
• For example, the hydrolysis of ATP has G = –7.3 kcal/mol or oxidation of glucose
• ATP + H2O ADP + Pi
• However, ATP dissolved in water remains stable for several days

© 2012 Pearson Education, Inc.
Before a Chemical Reaction Can Occur, the Activation Energy Barrier Must Be Overcome
• Molecules that could react with one another often do not because they lack sufficient energy
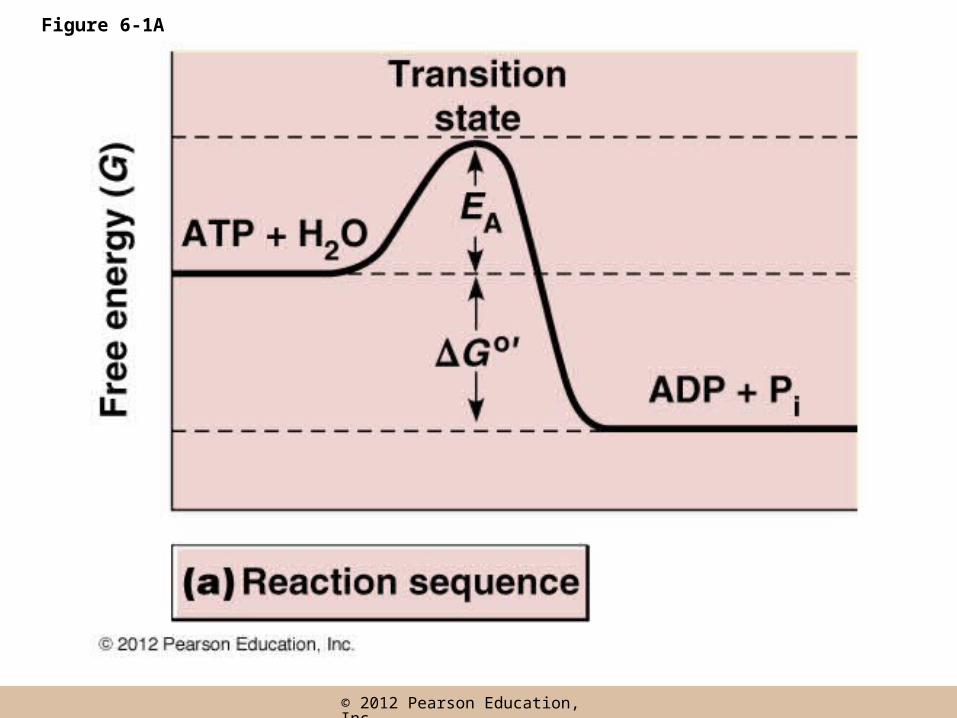
• Each reaction has a specific activation energy, EA
• EA: the minimum amount of energy required before collisions between the reactants will give rise to products

© 2012 Pearson Education, Inc.
Transition state
• Reactants need to reach an intermediate chemical stage called the transition state
• The transition state has a higher free energy than that of the initial reactants
© 2012 Pearson Education, Inc.
Figure 6-1A

© 2012 Pearson Education, Inc.
Activation energy barrier
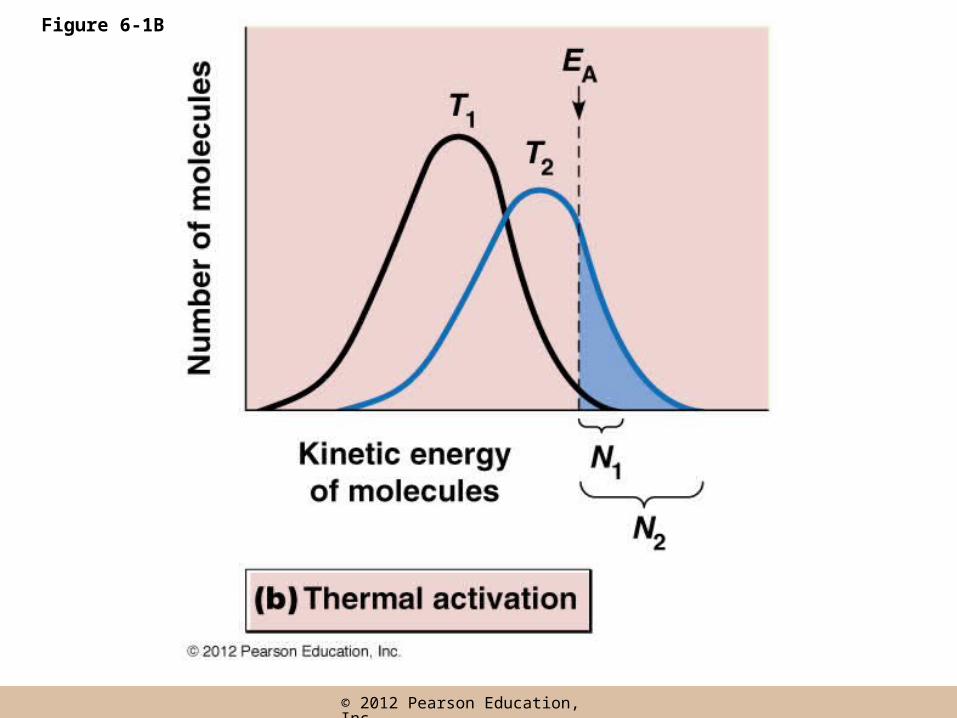
• The rate of a reaction is always proportional to the fraction of molecules with an energy equal to or greater than EA
• The only molecules that are able to react at a given time are those with enough energy to exceed the activation energy barrier, EA
© 2012 Pearson Education, Inc.
Figure 6-1B

© 2012 Pearson Education, Inc.
The Metastable State Is a Result of the Activation Barrier
• For most reactions at normal cell temperature, the activation energy is so high that few molecules can exceed the EA barrier
• Reactants that are thermodynamically unstable (seemingly stable), but lack sufficient EA, are said to be in a metastable state
• Life depends on high EAs that prevent most reactions in the absence of catalysts-critical why?

© 2012 Pearson Education, Inc.
Catalysts Overcome the Activation Energy Barrier
• The EA barrier must be overcome in order for needed reactions to occur
• This can be achieved by either increasing the energy content of molecules or by lowering the EA requirement

© 2012 Pearson Education, Inc.
Increasing the energy content of a system
• The input of heat can increase the kinetic energy of the average molecule, ensuring that more molecules will be able to take part in a reaction- in lab we heat solutions
• This is not useful in cells, however, which are isothermal
• Isothermal: constant in temperature

© 2012 Pearson Education, Inc.
Lowering activation energy
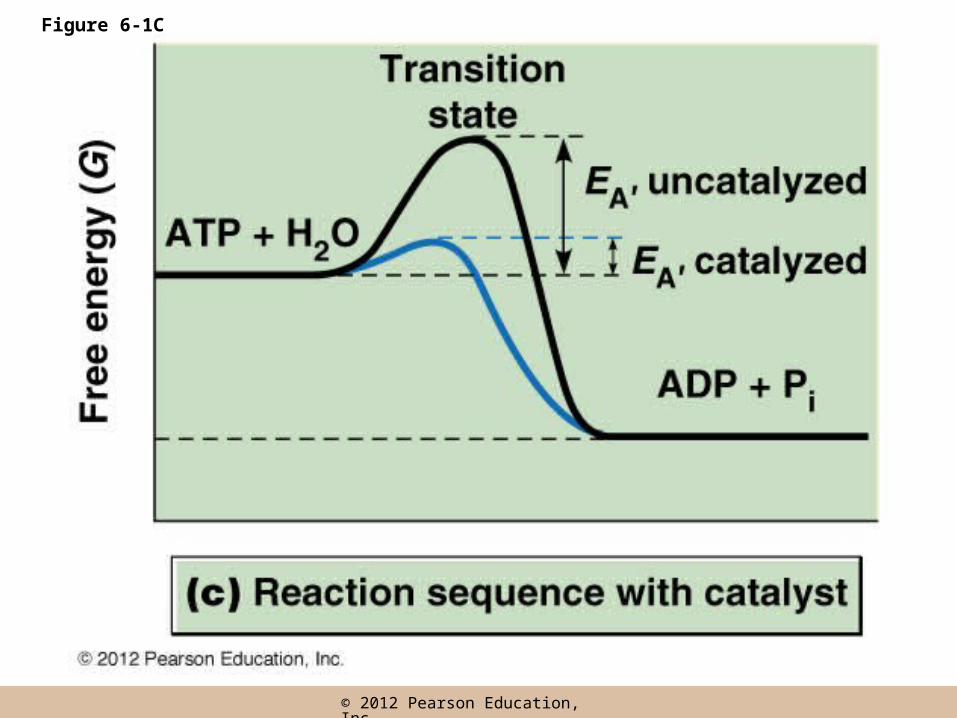
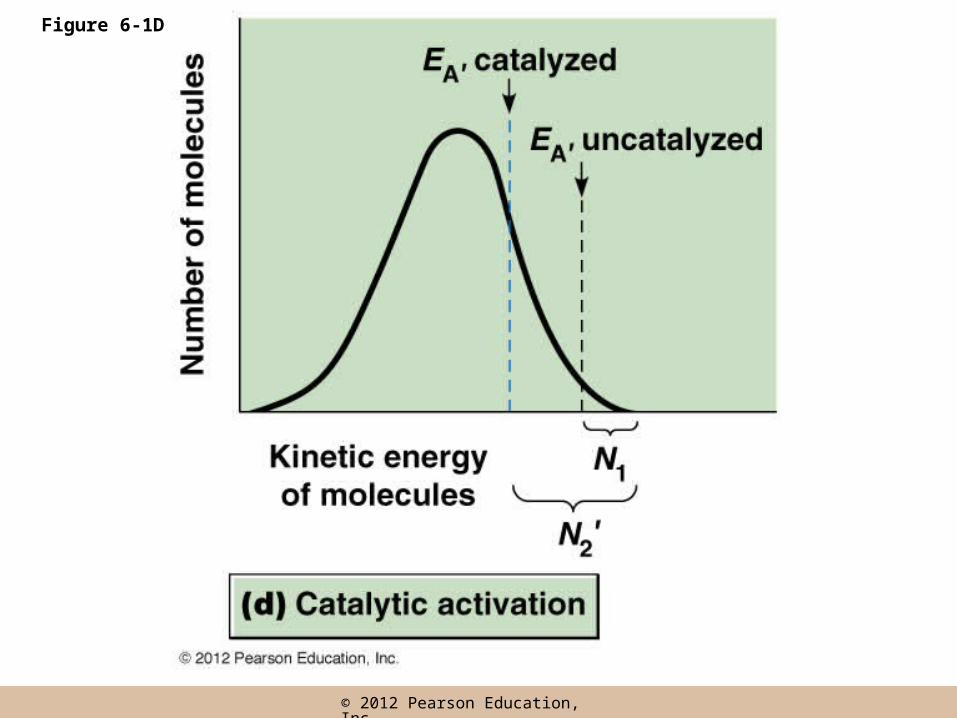
• If reactants can be bound on a surface and brought close together, their interaction will be favored and the required EA will be reduced
• A catalyst enhances the rate of a reaction by providing such a surface and effectively lowering EA
• Catalysts themselves proceed through the reaction unaltered, not permanently changes or consumed as the reaction proceeds
© 2012 Pearson Education, Inc.
Figure 6-1C
© 2012 Pearson Education, Inc.
Figure 6-1D

© 2012 Pearson Education, Inc.
The reaction of catalase (enzyme) in the decomposition of living tissue:
2 H2O2 → 2 H2O + O2
© 2012 Pearson Education, Inc.
What is quantum tunneling???

© 2012 Pearson Education, Inc.
Enzymes as Biological Catalysts
• All catalysts share three basic properties
– They increase reaction rates by lowering the EA required (is thermal activation required?)
– They form transient, reversible complexes with substrate molecules
– They change only the rate at which equilibrium is achieved, not the position of the equilibrium. Cannot change G´
• Organic catalysts are enzymes

© 2012 Pearson Education, Inc.
Most Enzymes Are Proteins
• Most enzymes are known to be proteins– Fermentation studies Buchner 1897– Urease was first enzyme crystallized
• However, recently, it has been discovered that some RNA molecules also have catalytic activity
• These are called ribozymes and will be discussed later

© 2012 Pearson Education, Inc.
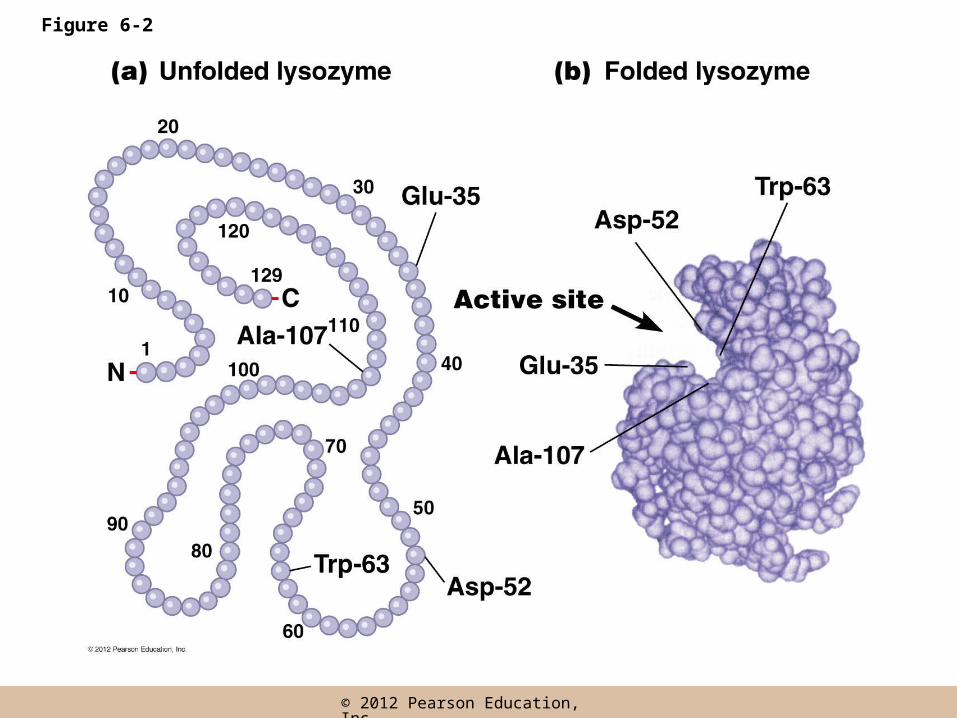
The Active Site
• Every enzyme contains a characteristic cluster of amino acids that forms the active site
• This results from the three dimensional folding of the protein, and is where substrates bind and catalysis takes place
• The active site is usually a groove or pocket that accommodates the intended substrate(s) with high affinity
© 2012 Pearson Education, Inc.
Figure 6-2

© 2012 Pearson Education, Inc.
Amino acids involved in the active site
• Substrate binding can depend on amino acids at various positions. Catalysis is usually very specific amino acids– Of the 20 different amino acids, only a few are
involved in the active site– These are cysteine, histine, serine, asparate,
glutamate, and lysine
• These can participate in binding the substrate and several serve as donors or acceptors of protons

© 2012 Pearson Education, Inc.
Cofactors
• Some enzymes contain nonprotein cofactors needed for catalytic activity, often because they function as electron acceptors
• These are called prosthetic groups and are usually metal ions (Mg, Zn, Fe) or small organic molecules called coenzymes
• Coenzymes are derivatives of vitamins (niacin, riboflavin, vit. B)

© 2012 Pearson Education, Inc.
Prosthetic groups
• Prosthetic groups are located at the active site and are indispensable for enzyme activity
• Each molecule of the enzyme catalase has a multimeric structure called a porphyrin ring to which a necessary iron atom is bound
• The requirement for certain prosthetic groups on some enzymes explains our requirements for trace amounts of vitamins and minerals

© 2012 Pearson Education, Inc.
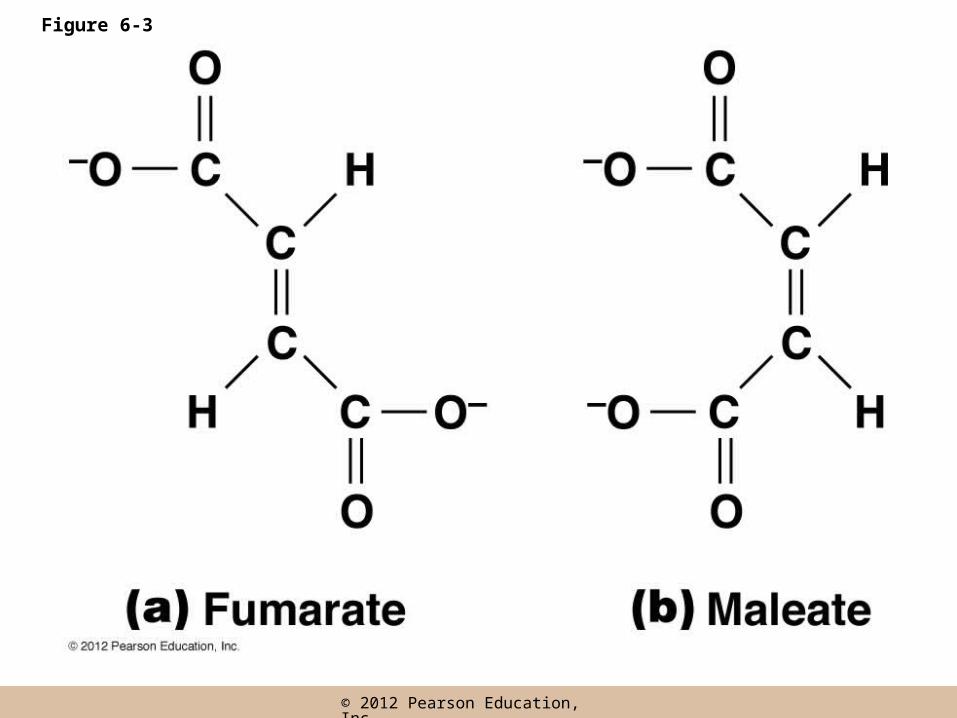
Enzyme Specificity
• Due to the shape and chemistry of the active site, enzymes have a very high substrate specificity
• Inorganic catalysts are very nonspecific whereas similar reactions in biological systems generally have a much higher level of specificity (trans fat)
© 2012 Pearson Education, Inc.
Figure 6-3

© 2012 Pearson Education, Inc.
Group specificity
• Not all enzymes are quite specific– Some enzymes will accept a number of
closely related substrates– Others accept any of an entire group of
substrates sharing a common feature
• This group specificity is most often seen in enzymes involved in degradation of polymers (carboxypeptidase A)

© 2012 Pearson Education, Inc.
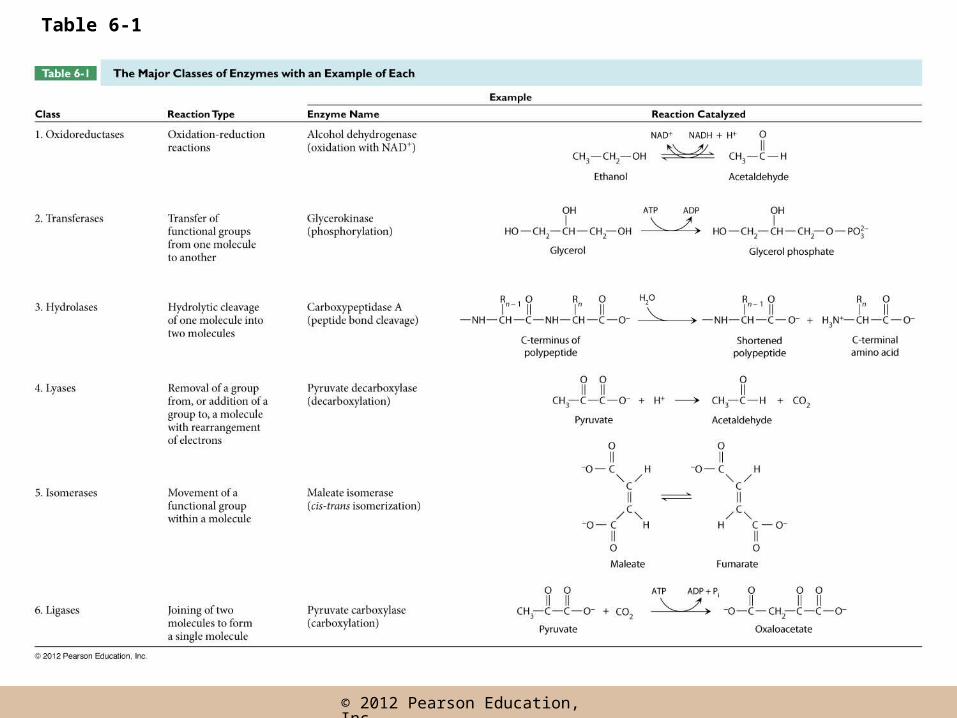
Enzyme Diversity and Nomenclature
• Thousands of different enzymes have been identified, with enormous diversity
• Names have been given to enzymes based on substrate (protease, ribonuclease, amylase), or function (trypsin, catalase)
• Under the Enzyme Commission (EC), enzymes are divided into six major classes based on general function

© 2012 Pearson Education, Inc.
Six classes of enzymes
• Oxidoreductases• Transferases• Hydrolases• Lysases• Isomerases• Ligases
© 2012 Pearson Education, Inc.
Table 6-1

© 2012 Pearson Education, Inc.
Sensitivity to Temperature
• Enzymes are characterized by their sensitivity to temperature
• This is not a concern in homeotherms, birds and mammals, that maintain a constant body temperature (independent of environment)
• However, many organisms function at their environmental temperature, which can vary widely (insects, reptiles, worms, plants)

© 2012 Pearson Education, Inc.
Enzyme activity and temperatures
• At low temperatures, the rate of enzyme activity increases with temperature due to increased kinetic activity of enzyme and substrate molecules
• However, beyond a certain point, further increases in temperature result in denaturation of the enzyme molecule and loss of enzyme activity

© 2012 Pearson Education, Inc.
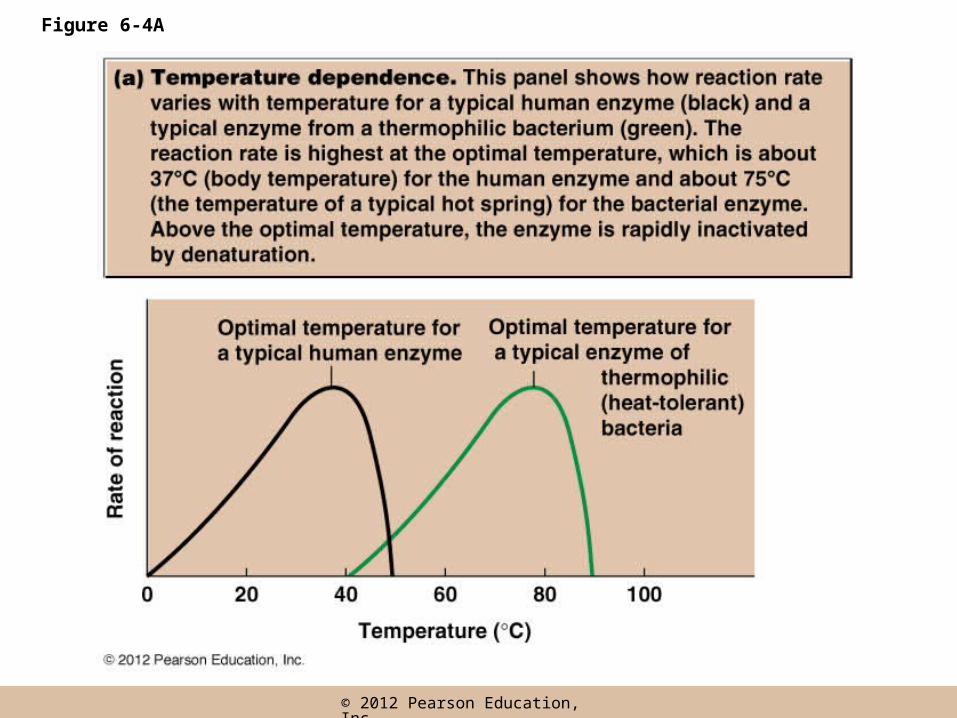
Optimal temperature
• The temperature range over which an enzyme denatures varies among enzymes and organisms
• The reaction rate of human enzymes is maximum at 37oC (the optimal temperature), the normal body temperature
• Most enzymes of homeotherms are inactivated by temperatures above 50–55oC
© 2012 Pearson Education, Inc.
Figure 6-4A

© 2012 Pearson Education, Inc.
Ranges of heat sensitivity
• Some enzymes are unusually sensitive and will denature at temperatures as low as 40oC
• Some enzymes retain activity at unusually high temperatures, such as the enzymes of archaea that live in acidic hot springs
• Enzymes of cryophilic (cold-loving) organisms such as Listeria bacteria (yeast/molds) can function at low temperatures, even under refrigeration

© 2012 Pearson Education, Inc.
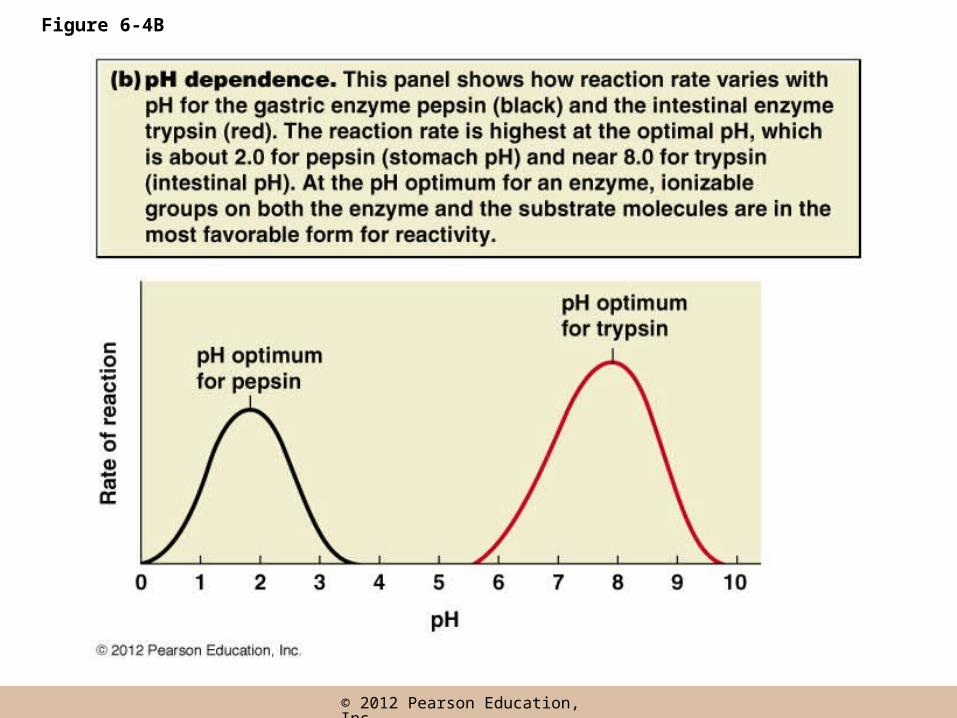
Sensitivity to pH
• Most enzymes are active within a pH range of about 3–4 units
• pH dependence is usually due to the presence of charged amino acids at the active site or on the substrate
• pH changes affect the charge of such residues, and can disrupt ionic and hydrogen bonds
© 2012 Pearson Education, Inc.
Figure 6-4B

© 2012 Pearson Education, Inc.
Sensitivity to Other Factors
• Enzymes are sensitive to factors such as molecules and ions that act as inhibitors or activators
• Most enzymes are also sensitive to ionic strength (dissolved ions) of the environment
• This affects hydrogen bonding and ionic interactions needed to maintain tertiary conformation
• Magnesium

© 2012 Pearson Education, Inc.
Substrate Binding, Activation, and Catalysis Occur at the Active Site
• Because of the precise chemical fit between the active site of the enzyme and its substrates, enzymes are highly specific

© 2012 Pearson Education, Inc.
Substrate Binding
• Once at the active site, the substrate molecules are bound to the enzyme surface in the right orientation to facilitate the reaction
• Substrate binding usually involves hydrogen bonds, ionic bonds, or both
• Substrate binding is readily reversible

© 2012 Pearson Education, Inc.
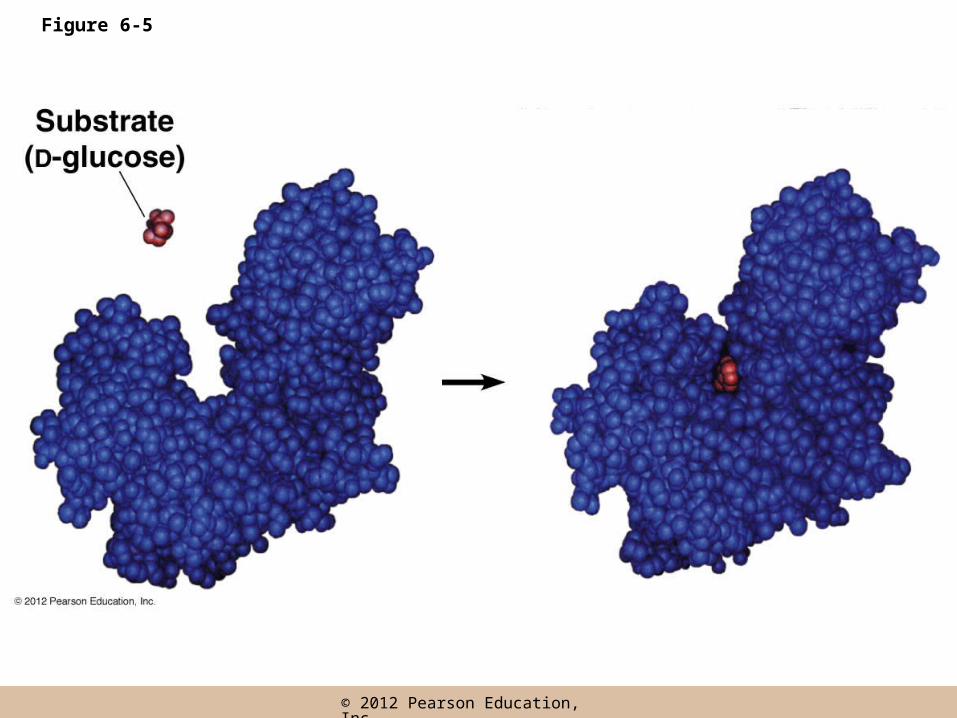
The induced-fit model
• In the past, the enzyme was seen as rigid, with the substrate fitting into the active site like a key in a lock (lock-and-key model)
• A more accurate view is the induced-fit model, in which substrate binding at the active site induces a conformational change in the shape of the enzyme
© 2012 Pearson Education, Inc.
Figure 6-5

© 2012 Pearson Education, Inc.
Video: Closure of hexokinase via induced fit

© 2012 Pearson Education, Inc.
Conformational change
• The induced conformational change brings needed amino acid side chains into the active site, even those that are not nearby
• Sometimes these are not nearby unless the substrate is bound to the active site

© 2012 Pearson Education, Inc.
Substrate Activation
• The role of the active site is to recognize and bind the appropriate substrate and also to activate it by providing the right environment for catalysis
• This is called substrate activation, which proceeds via several possible mechanisms

© 2012 Pearson Education, Inc.
Three common mechanisms of substrate activation
• Bond distortion, making it more susceptible to catalytic attack
• Proton transfer, which increases reactivity of substrate
• Electron transfer, resulting in temporary covalent bonds between enzyme, substrate

© 2012 Pearson Education, Inc.
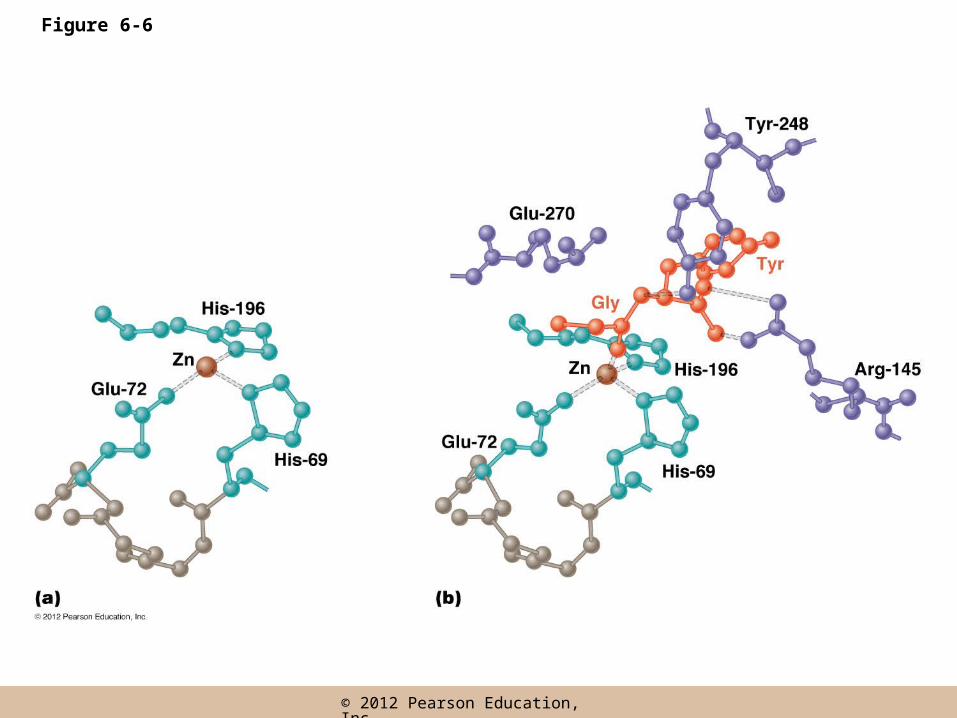
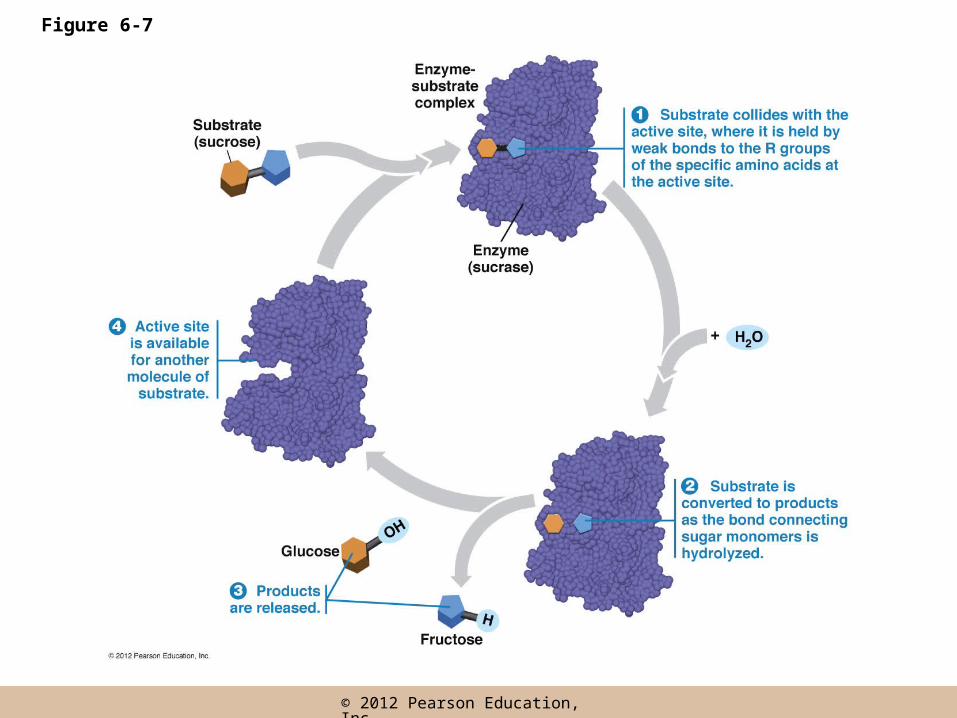
The Catalytic Event
• The sequence of events
– 1. The random collision of a substrate molecule with the active site results in it binding there
– 2. Substrate binding induces a conformational change that tightens the fit, facilitating the conversion of substrate into products
© 2012 Pearson Education, Inc.
Figure 6-6

© 2012 Pearson Education, Inc.
The Catalytic Event (continued)
• The sequence of events
– 3. The products are then released from the active site
– 4. The enzyme molecule returns to the original conformation with the active site available for another molecule of substrate
© 2012 Pearson Education, Inc.
Figure 6-7

© 2012 Pearson Education, Inc.
Enzyme Kinetics
• Enzyme kinetics describes the quantitative aspects of enzyme catalysis and the rate of substrate conversion into products
• Reaction rates are influenced by factors such as the concentrations of substrates, products, and inhibitors

© 2012 Pearson Education, Inc.
Initial reaction rates
• Initial reaction rates are measured over a brief time, during which the substrate concentration has not yet decreased enough to affect the rate of reaction

© 2012 Pearson Education, Inc.
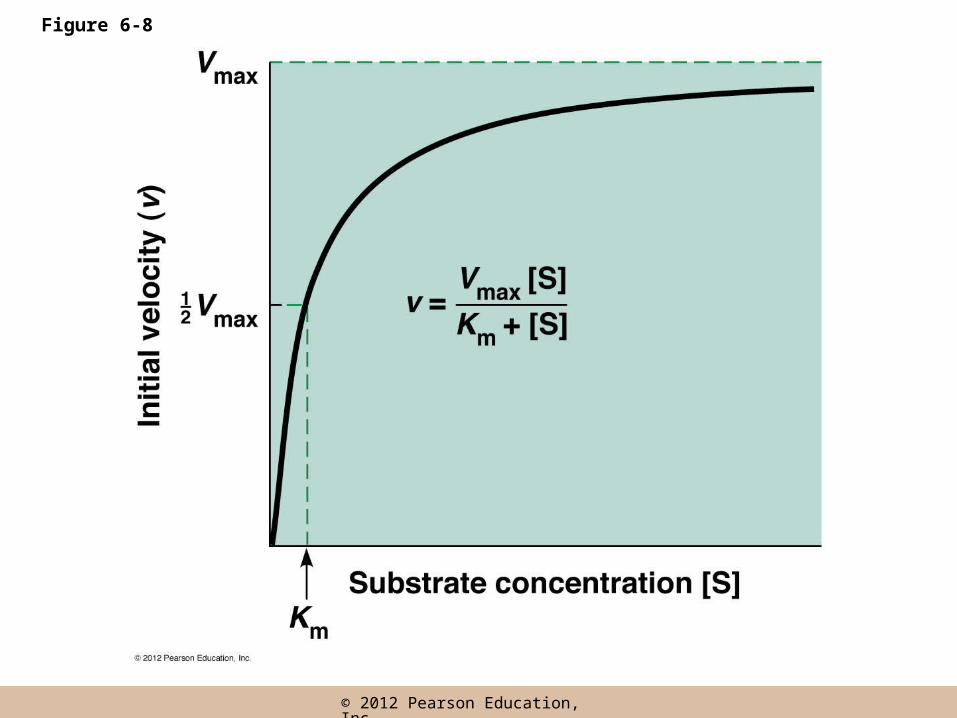
Most Enzymes Display Michaelis–Menten Kinetics
• Initial reaction velocity (v), the rate of change in product concentration per unit time, depends on the substrate concentration [S]
• At low [S], doubling [S] will double v, but as [S] increases each additional increase in [S] results in a smaller increase in v
• When [S] becomes very large the value of v reaches a maximum
© 2012 Pearson Education, Inc.
Figure 6-8

© 2012 Pearson Education, Inc.
Vmax and saturation
• As [S] tends toward infinity, v approaches an upper limiting value, maximum velocity (Vmax)
• The value of Vmax can be increased by adding more enzyme
• The inability of increasingly higher substrate concentrations to increase the reaction velocity beyond a finite upper value is called saturation

© 2012 Pearson Education, Inc.
The Michaelis–Menten Equation
• Michaelis and Menten postulated a theory of enzyme action
• Enzyme E first reacts with the substrate, to form a transient complex, ES
• ES then undergoes the catalytic reaction to generate E and P

© 2012 Pearson Education, Inc.
The Michaelis–Menten Equation (continued)
•
• The above model, under steady state conditions gives the Michaelis–Menten equation
•
• Km (the Michaelis constant) = the concentration of substrate that gives half maximum velocity

© 2012 Pearson Education, Inc.
What Is the Meaning of Vmax and Km?
• We can understand the relationship between v and [S], and the meaning of Vmax and Km by considering three cases regarding [S]

© 2012 Pearson Education, Inc.
Case 1: Very Low Substrate Concentration ([S] << Km)
• If [S] << Km
• Then, Km + [S] = [Km]
•
• So at very low [S], the initial velocity of the reaction is roughly proportional to [S]

© 2012 Pearson Education, Inc.
Case 2: Very High Substrate Concentration ([S] >> Km)
• If [S] >> Km
• Then, Km + [S] = [S]
•
• So at very high [S], the initial velocity of the reaction is independent of variation in [S] and Vmax is the velocity at saturating substrate concentrations

© 2012 Pearson Education, Inc.
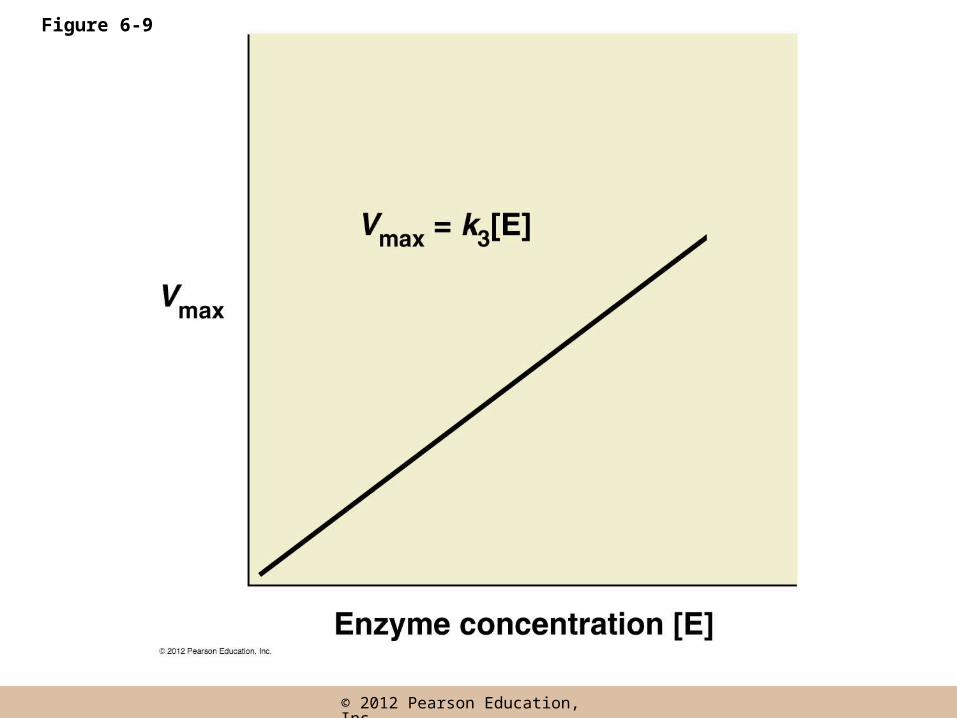
Vmax
• Vmax is an upper limit determined by
– The time required for the actual catalytic reaction
– How many enzyme molecules are present
• The only way to increase Vmax is to increase enzyme concentration
© 2012 Pearson Education, Inc.
Figure 6-9

© 2012 Pearson Education, Inc.
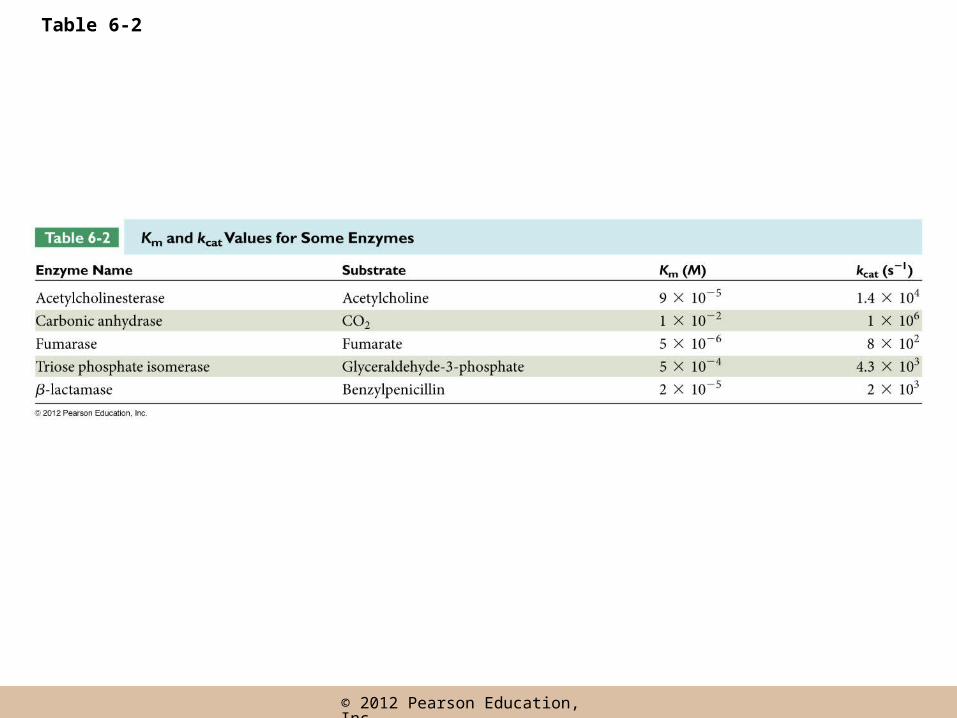
Why Are Km and Vmax Important to Cell Biologists?
• The lower the Km value for a given enzyme and substrate, the lower the [S] range in which the enzyme is effective
• Vmax is important, as a measure of the potential maximum rate of the reaction
• By knowing Vmax, Km, and the in vivo substrate concentration, we can estimate the likely rate of the reaction under cellular conditions
© 2012 Pearson Education, Inc.
Table 6-2

© 2012 Pearson Education, Inc.
Enzyme Inhibitors Act Either Irreversibly or Reversibly
• Enzymes are influenced (mostly inhibited) by products, alternative substrates, substrate analogs, drugs, toxins, and allosteric effectors
• The inhibition of enzyme activity plays a vital role as a control mechanism in cells
• Drugs and poisons frequently exert their effects by inhibition of specific enzymes

© 2012 Pearson Education, Inc.
Inhibitors important to enzymologists
• Inhibitors of greatest use to enzymologists are substrate analogs and transition state analogs
• These are compounds that resemble real substrates or transition states closely enough to occupy the active state but not closely enough to complete the reaction
• Substrate analogs are important tools in fighting infectious diseases

© 2012 Pearson Education, Inc.
Reversible and irreversible inhibition
• Irreversible inhibitors, which bind the enzyme covalently, cause permanent loss of catalytic activity and are generally toxic to cells
– For example, heavy metal ions, nerve gas poisons, some insecticides
• Reversible inhibitors bind enzymes noncovalently and can dissociate from the enzyme

© 2012 Pearson Education, Inc.
Reversible inhibition (continued)
• The fraction of enzyme available for use in a cell depends on the concentration of the inhibitor and how easily the enzyme and inhibitor can dissociate
• The two forms of reversible inhibitors are competitive inhibitors and noncompetitive inhibitors

© 2012 Pearson Education, Inc.
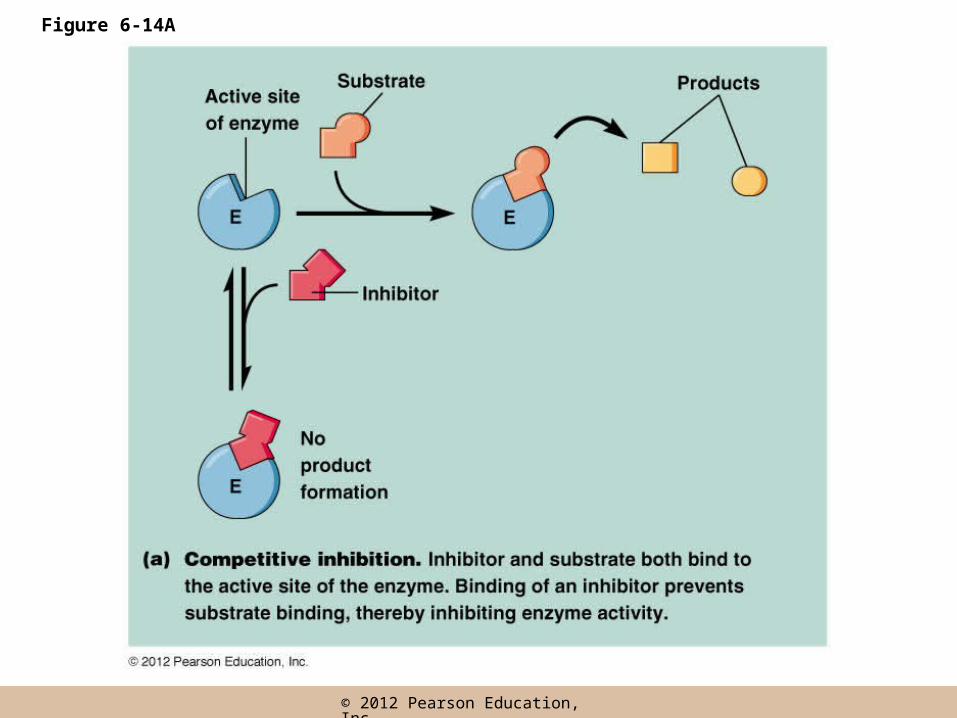
Competitive inhibition
• Competitive inhibitors bind the active site of an enzyme and so compete with substrate for the active site
• Enzyme activity is inhibited directly because active sites are bound to inhibitors, preventing the substrate from binding
© 2012 Pearson Education, Inc.
Figure 6-14A

© 2012 Pearson Education, Inc.
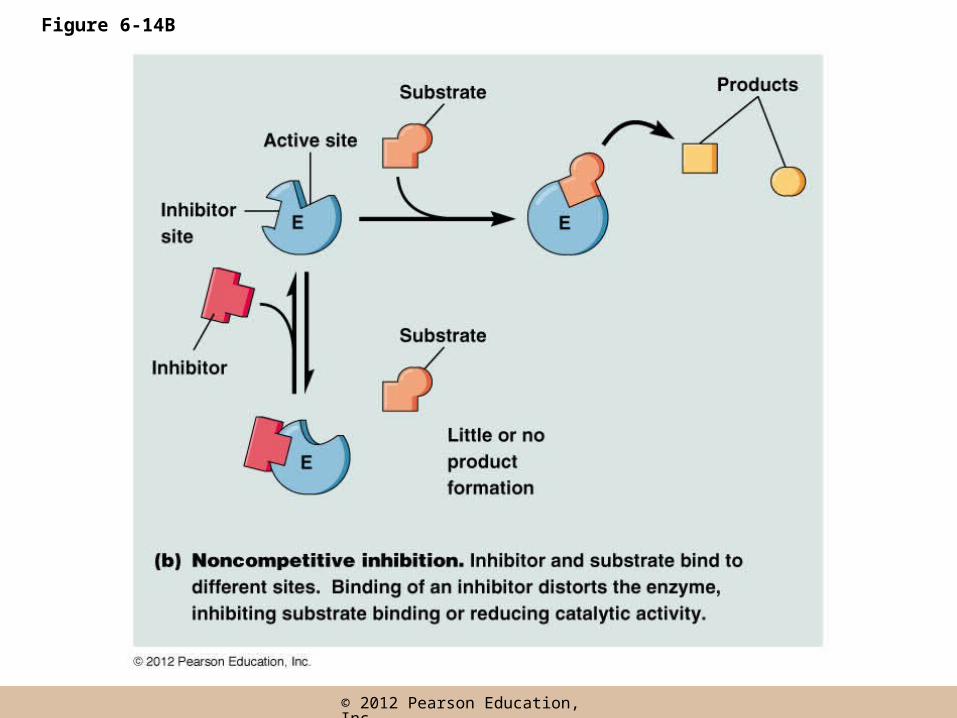
Noncompetitive inhibition
• Noncompetitive inhibitors bind the enzyme molecule outside of the active site
• They inhibit activity indirectly by causing a conformation change in the enzyme that
– Inhibits substrate binding at the active site, or
– Reduces catalytic activity at the active site
© 2012 Pearson Education, Inc.
Figure 6-14B

© 2012 Pearson Education, Inc.
Enzyme Regulation
• Enzyme rates must be continuously adjusted to keep them tuned to the needs of the cell
• Regulation that depends on interactions of substrates and products with an enzyme is called substrate-level regulation
• Increases in substrate levels result in increased reaction rates, whereas increased product levels lead to lower rates

© 2012 Pearson Education, Inc.
Allosteric regulation and covalent modification
• Cells can turn enzymes on and off as needed by two mechanisms: allosteric regulation and covalent modification
• Usually enzymes regulated this way catalyze the first step of a multi-step sequence
• By regulating the first step of a process, cells are able to regulate the entire process

© 2012 Pearson Education, Inc.
Allosteric Enzymes Are Regulated by Molecules Other than Reactants and Products
• Allosteric regulation is the single most important control mechanism whereby the rates of enzymatic reactions are adjusted to meet the cell’s needs

© 2012 Pearson Education, Inc.
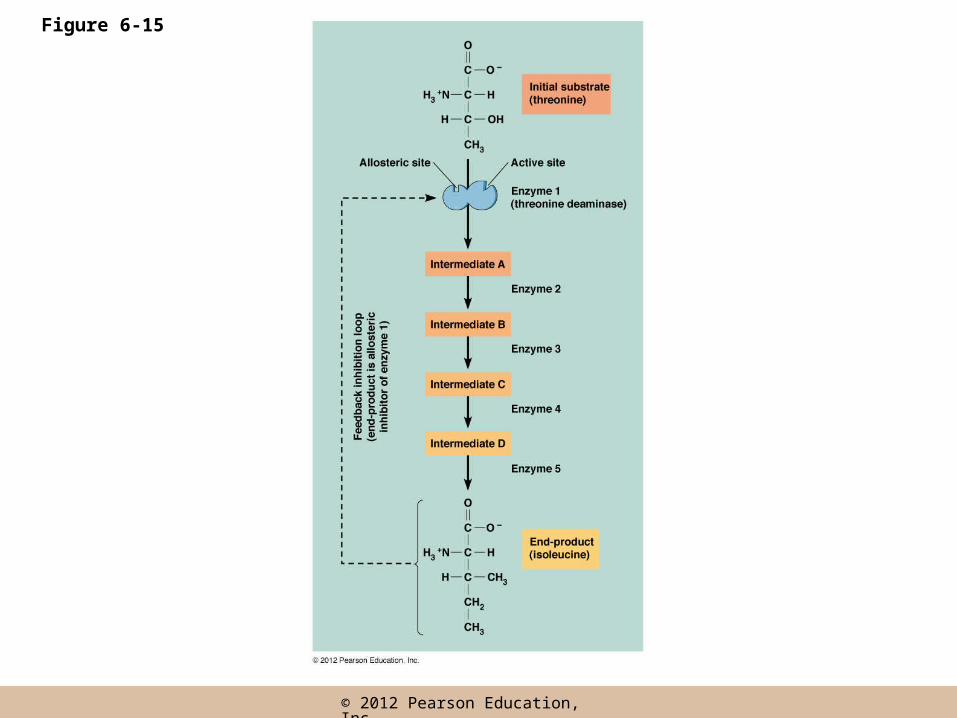
Feedback Inhibition
• It is not in the best interests of a cell for enzymatic reactions to proceed at the maximum rate
• In feedback (or end-product) inhibition, the final product of an enzyme pathway negatively regulates an earlier step in the pathway
•
© 2012 Pearson Education, Inc.
Figure 6-15

© 2012 Pearson Education, Inc.
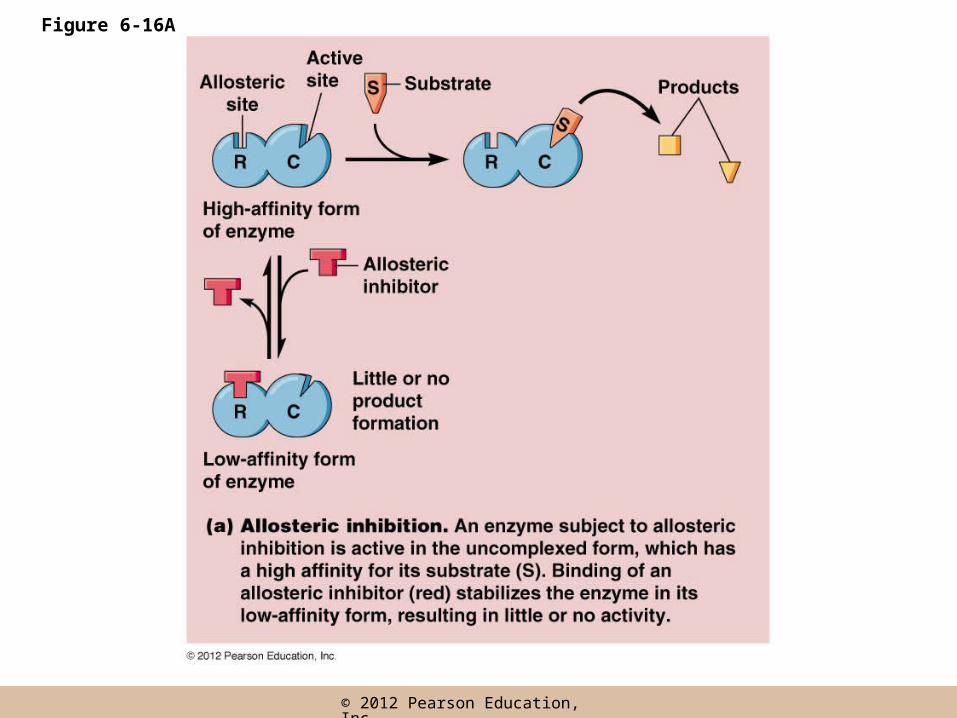
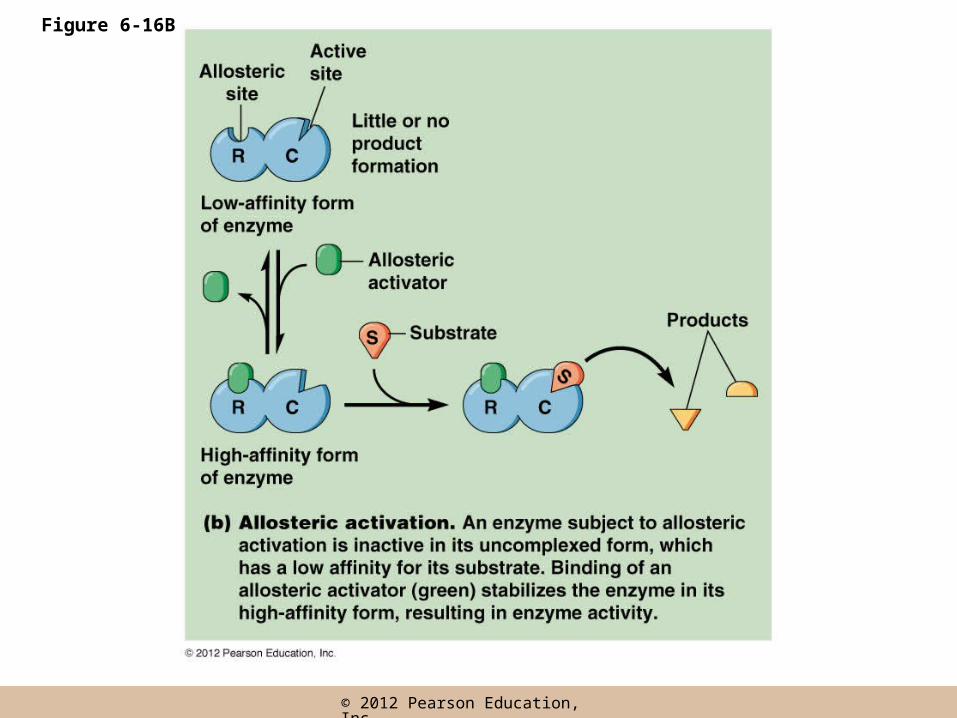
Allosteric Regulation
• Allosteric enzymes have two conformations, one in which it has affinity for the substrate(s) and one in which it does not
• Allosteric regulation makes use of this property by regulating the conformation of the enzyme
• An allosteric effector regulates enzyme activity by binding and stabilizing one of the conformations

© 2012 Pearson Education, Inc.
Allosteric regulation (continued)
• An allosteric effector binds a site called an allosteric (or regulatory) site, distinct from the active site
• The allosteric effector may be an activator or inhibitor, depending on its effect on the enzyme
• Inhibitors shift the equilibrium between the two enzyme states to the low affinity form; activators favor the high affinity form
© 2012 Pearson Education, Inc.
Figure 6-16A
© 2012 Pearson Education, Inc.
Figure 6-16B

© 2012 Pearson Education, Inc.
Allosteric enzymes
• Most allosteric enzymes are large, multisubunit proteins with an active or allosteric site on each subunit
• Active and allosteric sites are on different subunits, the catalytic and regulatory subunits, respectively
• Binding of allosteric effectors alters the shape of both catalytic and regulatory subunits

© 2012 Pearson Education, Inc.
Allosteric Enzymes Exhibit Cooperative Interactions Between Subunits
• Many allosteric enzymes exhibit cooperativity
• As multiple catalytic sites bind substrate molecules, the enzyme changes conformation, which alters affinity for the substrate
• In positive cooperativity the conformation change increases affinity for substrate; in negative cooperativity, affinity for substrate is decreased

© 2012 Pearson Education, Inc.
Enzymes Can Also Be Regulated by the Addition or Removal of Chemical Groups
• Many enzymes are subject to covalent modification
• Activity is regulated by addition or removal of groups, such as phosphate, methyl, acetyl groups, etc.

© 2012 Pearson Education, Inc.
Phosphorylation and Dephosphorylation
• The reversible addition of phosphate groups is a common covalent modification
• Phosphorylation occurs most commonly by transfer of a phosphate group from ATP to the hydroxyl group of Ser, Thr, or Tyr residues in a protein
• Protein kinases catalyze the phosphorylation of other proteins

© 2012 Pearson Education, Inc.
Dephosphorylation
• Dephosphorylation, the removal of phosphate groups from proteins, is catalyzed by protein phosphatases
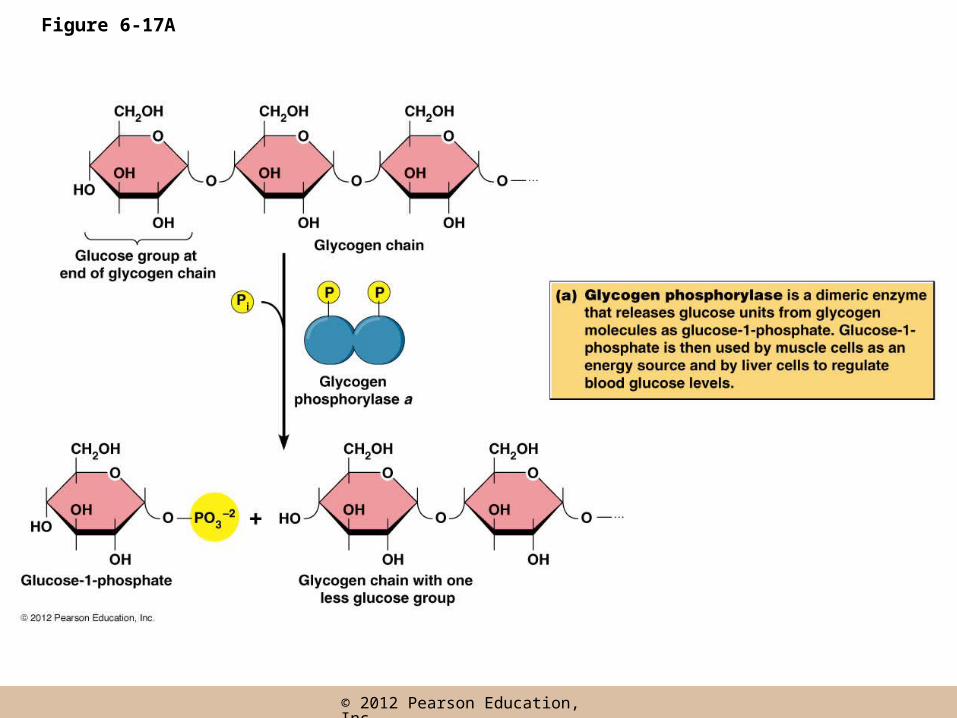
• Depending on the enzyme, phosphorylation may be associated with activation or inhibition of the enzyme
• Fisher and Krebs won the Nobel prize for their work on glycogen phosphorylase
© 2012 Pearson Education, Inc.
Figure 6-17A

© 2012 Pearson Education, Inc.
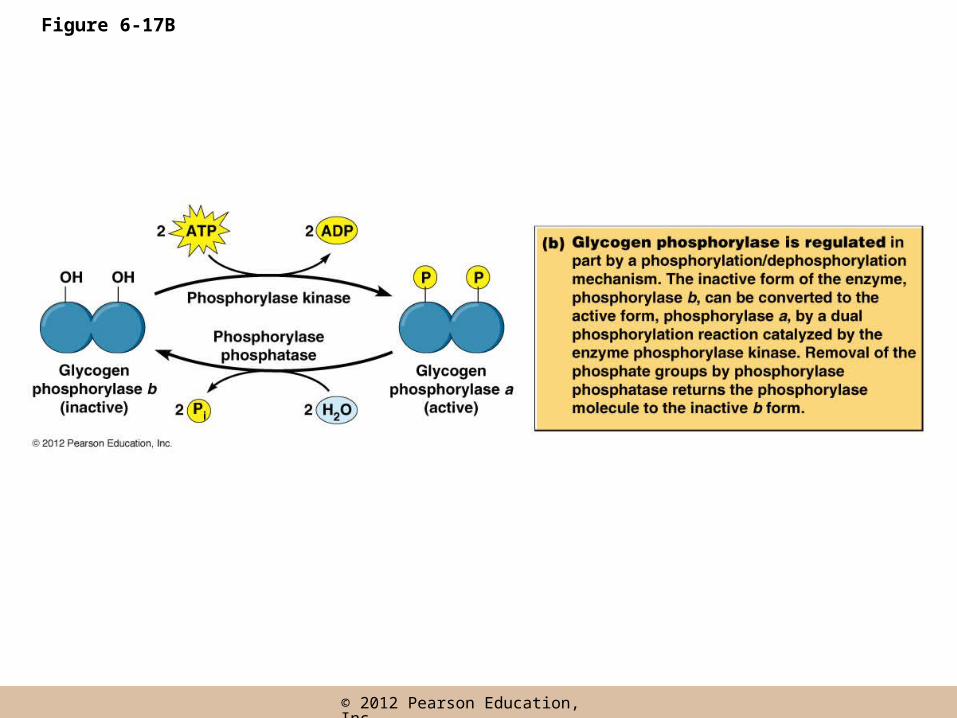
Regulation of glycogen phosphorylase
• Glycogen phosphorylase exists as two inter-convertible forms
– An active, phosphorylated form (glycogen phosphorylase-a)
– An inactive, non-phosphorylated form (glycogen phosphorylase-b)
• The enzymes responsible
– Phosphorylase kinase phosphorylates the enzyme
– Phosphorylase phosphatase removes the phosphate
© 2012 Pearson Education, Inc.
Figure 6-17B

© 2012 Pearson Education, Inc.
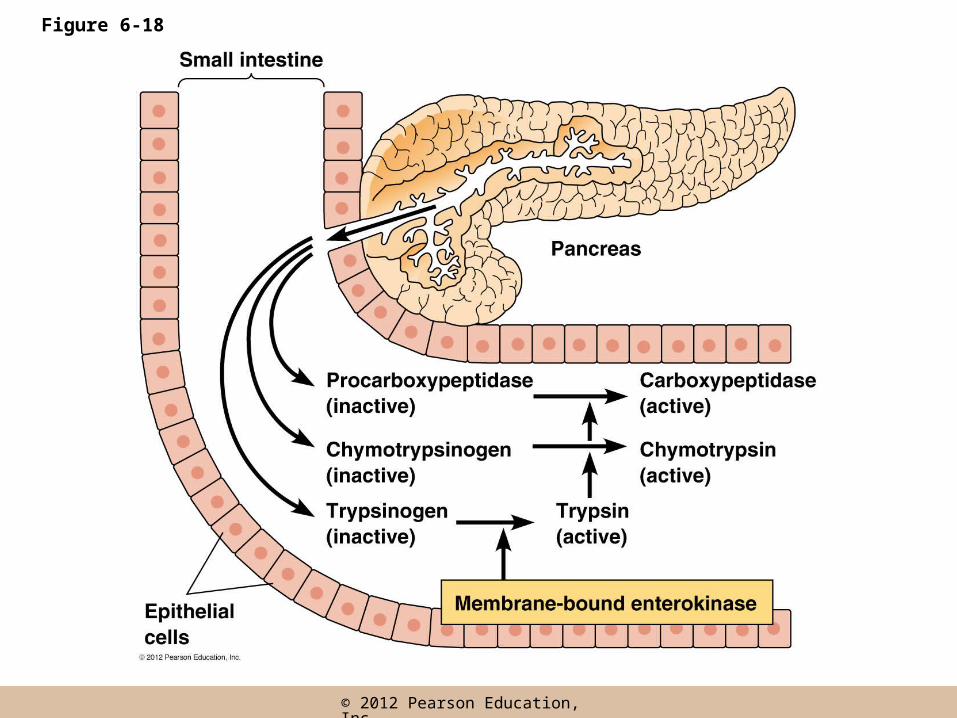
Proteolytic Cleavage
• The activation of a protein by a one-time, irreversible removal of part of the polypeptide chain is called proteolytic cleavage
• Proteolytic enzymes of the pancreas, trypsin, chymotrypsin, and carboxypeptidase, are examples of enzymes synthesized in inactive form (as zymogens) and activated by cleavage as needed
© 2012 Pearson Education, Inc.
Figure 6-18

© 2012 Pearson Education, Inc.
RNA Molecules as Enzymes: Ribozymes
• Some RNA molecules have been found to have catalytic activity; these are called ribozymes
• Self-splicing rRNA from Tetrahymena thermophila and ribonuclease P are examples
• It is thought by some that RNA catalysts predate protein catalysts, and even DNA