τελικη εκθεση...
TRANSCRIPT

ΤΕΛΙΚΗ ΕΚΘΕΣΗ ΜΕΛΕΤΗΣ
Ο γενετικός και μοριακός ρόλος των πρωτεινών AIF (Apoptosis Inducing Factor) στην ενεργοποίηση του εγγενούς
ανοσοποιητικού συστήματος των φυτών
Χρηματοδοτούμενη από το Κοινωφελές Ίδρυμα Ιωάννη Σ. Λάτση
Επιστημονικός Υπεύθυνος: Δημήτριος Τσιτσιγιάννης Λέκτορας Φυτοπαθολογίας, Τμήμα Επιστήμης Φυτικής Παραγωγής
Γεωπονικό Πανεπστήμιο Αθηνών \
Δεκέμβριος 2010

2
Κοινωφελές Ίδρυμα Ιωάννη Σ. Λάτση – ΜΕΛΕΤΕΣ 2010
ΤΙΤΛΟΣ ΜΕΛΕΤΗΣ
Ο γενετικός και μοριακός ρόλος των πρωτεινών AIF (Apoptosis Inducing Factor) στην ενεργοποίηση του εγγενούς ανοσοποιητικού συστήματος των φυτών
Επιστημονικός Υπεύθυνος - Συντονιστής: Δημήτριος Τσιτσιγιάννης Λέκτορας Φυτοπαθολογίας, Τμήμα Επιστήμης Φυτικής Παραγωγής
Γεωπονικό Πανεπστήμιο Αθηνών
Μέλη της Ομάδας׃ Σοφία Κουντούρη Μεταπτυχιακή Φοιτήτρια, Τμήμα Επιστήμης Φυτικής Παραγωγής, Εργαστήριο Φυτοπαθολογίας, Γεωπονικό Πανεπστήμιο Αθηνών
ΠΕΡΙΛΗΨΗ
Τα φυτά, όπως όλοι οι οργανισμοί, έχουν αναπτύξει ένα εγγενές «ανοσοποιητικό» σύστημα που τα επιτρέπει να αντιμετωπίζουν μολύνσεις από διάφορα παθογόνα. Η πρώτη γραμμή άμυνας είναι η αναγνώριση των επιτιθέμενων μικροοργανισμών και η παραγωγή διαφόρων ουσιών με στόχο την καταστροφή τους. Η δεύτερη γραμμή άμυνας είναι ο προγραμματισμένος θάνατος των κυττάρων που έχουν προσβληθεί με αποτέλεσμα να περιοριστεί η εξάπλωση της προσβολής. Ο προγραμματισμένος κυτταρικός θάνατος (ΠΚΘ) είναι μια διαδικασία που πραγματοποιείται φυσιολογικά κατά τη διάρκεια της ανάπτυξης και άμυνας των πολυκύτταρων οργανισμών αλλά μπορεί να συνδέεται και με σοβαρές παθήσεις όπως καρκίνος και εκφυλιστικές ασθένειες. Έρευνες των τελευταίων χρόνων έχουν αποδείξει την ύπαρξη κοινών βιοχημικών μονοπατιών του ΠΚΘ μεταξύ φυτικών, ζωικών και μικροβιακών κυττάρων αλλά σε αντίθεση με τους άλλους οργανισμούς, η διαδικασία αυτή στα φυτά δεν έχει εκτενώς μελετηθεί. Στόχος της παρούσης μελέτης ήταν η διαλεύκανση μιας ομάδας γονιδίων που εντοπίστηκε στο γενετικό φυτό μοντέλο Arabidopsis thaliana, που έχουν ομοιότητες με το γονίδιο των θηλαστικών ΑIF (Apoptosis Inducing Factor), μια φλαβοπρωτεΐνη που σχετίζεται με ασθένειες συνδεδεμένες με φαινόμενα αυξημένης απόπτωσης όπως η μόλυνση από τον ιό HIV, νευροδιαταραχές, καρδιακά επεισόδια. Κατά την παρούσα μελέτη διαπιστώθηκε για πρώτη φορά ότι τα At-AIF γονίδια παίζουν σημαντικό ρόλο στην ικανότητα ενεργοποίησης του εγγενούς ανοσοποιητικού συστήματος των φυτών και της ευαισθησίας ή ανθεκτικότητάς τους σε μια σειρά σημαντικών παθογόνων των καλλιεργειών.

3
ABSTRACT
Plants, like all organisms, have developed an innate immune system that allows them to defeat infections from various pathogens. The first line of defense is to identify the invasive microorganisms and to produce various molecules in order to destroy them. The second line of defense is the programmed death of infected cells, thereby limiting the spread of infection. The programmed cell death (PCD) is a process that normally takes place during development and defense of multicellular organisms and it may be linked to serious diseases like cancer and degenerative diseases. Research in recent years has demonstrated the existence of common biochemical pathways of PCD among plant, animal and microbial cells but, unlike other organisms, this process in plants has not been well characterised. The aim of this study was to investigate a family of genes identified in the genetic model plant Arabidopsis thaliana, that have similarities with the mammalian AIF gene (Apoptosis Inducing Factor), a phylogenetically old, 57 kDa flavoprotein, which shares similarity to bacterial, fungus and plant oxidoreductases. AIF is associated with diseases related to increased apoptotic events such as infection with HIV, neurodegenerations, heart attacks. The present study showed for the first time that the At-AIF genes play an important role in the activation of the plant immune system and the resistance or susceptibility to a number of important crop pathogens.

4
ΕΙΣΑΓΩΓΗ
Τα φυτά εκτίθενται συνεχώς σε ένα μεγάλο αριθμό παθογόνων. Kατά τη διάρκεια της εξελικτικής τους πορείας ανέπτυξαν όμως διάφορα συστήματα άμυνας ώστε να μπορέσουν να προστατευτούν από ενδεχόμενη μόλυνση από διάφορους παθογόνους μικροοργανισμούς. Για να αποβούν μολυσματικά τα περισσότερα παθογόνα πρέπει να αποκτήσουν πρόσβαση στο εσωτερικό του φυτού, είτε διαπερνώντας απευθείας τη φυλλική ή ριζική επιφάνεια, είτε εισερχόμενα από πληγές ή φυσικά ανοίγματα στα φύλλα όπως είναι τα στομάτια. Τα φυτά σε αντίθεση με τα θηλαστικά, δεν διαθέτουν κινούμενα αμυντικά κύτταρα. Για να αποφύγουν την προσβολή, βασίζονται στον αμυντικό μηχανισμό κάθε μεμονωμένου κυττάρου καθώς και στην παραγωγή διασυστηματικών σημάτων προερχόμενα από το σημείο της αρχικής προσβολής. Τα φυτά έχουν αναπτύξει έναν παθητικό μηχανισμό και έναν ιδιαίτερα πολύπλοκο εγγενή μηχανισμό άμυνας. Ο πρώτος αποτελείται από φυσικούς παράγοντες άμυνας των φυτών δηλαδή ειδικές κατασκευές όπως φυσικοί φραγμοί της εφυμενίδας (κυτταρικά τοιχώματα, στομάτια, κηροί, τοξικοί μεταβολίτες). Ο δεύτερος έχει τη δυνατότητα να ενεργοποιείται μέσα σε λίγα λεπτά έως ώρες από την πρώτη επαφή του φυτού με το παθογόνο το οποίο έχει τη δυνατότητα παραγωγής εξειδικευμένων μορίων όπως είναι οι διεγέρτες που ενεργοποιούν με τη σειρά τους εγγενείς μηχανισμούς ανθεκτικότητας των φυτών. Ο εγγενής ή ενεργητικός μηχανισμός άμυνας των φυτών, στον οποίο περιλαμβάνονται διάφοροι ιστολογικοί και βιοχημικοί μηχανισμοί, με τους οποίους τα φυτά αντιδρούν στην προσβολή από τα παθογόνα διαφοροποιείται βιοχημικά σε 3 συστήματα: α) στο βασικό ανοσοποιητικό σύστημα που ενεργοποιείται μέσα σε 10-30 min από την επαφή του παθογόνου με το φυτό, β) στο ανοσοποιητικό σύστημα των γονιδίων ανθεκτικότητας που ενεργοποιείται 2-3 h μετά την επαφή και την απελευθέρωση των διεγερτών από το παθογόνο στο φυτικό κύτταρο και γ) στην επαγόμενη και επίκτητη διασυστηματική αντοχή (Systemic Acquired Resistance και Induced Systemic Resistance) δηλαδή στη βιολογική, βιοχημική ή χημική διέγερση λανθανόντων μηχανισμών αντοχής των φυτών σε παθογόνα στα οποία τα επαγόμενα φυτά εμφανίζουν ευαισθησία πριν από τη δράση των παραγόντων διέγερσης.
Ο Προγραμματισμένος Κυτταρικός Θάνατος (ΠΚΘ-Programmed Cell Death) είναι μια φυσιολογική διαδικασία του κύκλου ζωής των πολυκύτταρων οργανισμών (1,2). Οι «ευεργετικές» συνέπειες του θανάτου δεν είναι εξαίρεση αλλά κανόνας στην βιολογία. Περίπου 50 δισεκατομμύρια κύτταρα πεθαίνουν καθημερινά στο σώμα μας και αυτό μεταξύ άλλων μας προστατεύει από σοβαρές ασθένειες όπως ο καρκίνος, ενώ ο προγραμματισμένος κυτταρικός θάνατος είναι σημαντικός για τη σωστή λειτουργικά διαμόρφωση του σώματός μας. Ο ΠΚΘ αποτελεί επίσης ένα είδος θανάτου που τα κύτταρα έχουν προγραμματίσει να εκτελούν με σκοπό να παρεμποδιστεί μια επικείμενη εισβολή ενός παθογόνου το οποίο επιδιώκει να διεισδύσει και να εγκατασταθεί στον ξενιστή του. Στα θηλαστικά ο προγραμματισμένος κυτταρικός θάνατος διαχωρίζεται σε τρεις μορφές: την απόπτωση, την αυτοφαγία και τη νέκρωση. Στα φυτά η πιο μελετημένη μορφή ΠΚΘ είναι η “αντίδραση υπερευαισθησίας-ΑΥ)” που μοιάζει με την απόπτωση στα θηλαστικά και εκδηλώνεται ως η ακραία έκφραση ανθεκτικότητας σε μυκητολογικές, βακτηριολογικές και ιολογικές προσβολές και αποτελεί μέρος του εγγενούς ενεργητικού ανοσοποιητικού συστήματος που συνήθως εκδηλώνεται 2-3 h μετά την επαφή του φυτού με το παθογόνο. Η ΑΥ περιλαμβάνει κυτταρολογικές και βιοχημικές αλλοιώσεις και ταχύτατη νέκρωση των προσβαλλομένων κυττάρων με επακόλουθο τον εγκλωβισμό του παθογόνου στην αρχική θέση διείσδυσής του και στην αποφυγή ανάπτυξης της ασθένειας (3,4,5). Το φυτό προκαλώντας τοπική νέκρωση των κυτττάρων που έχει καταφέρει να προσβάλλει το παθογόνο, προσπαθεί να το αποτρέψει να πολλαπλασιαστεί περαιτέρω και να μολύνει τον

5
ξενιστή. Πρόκειται δηλαδή για μια μορφή «αυτοκτονίας» ενός ή περισσοτέρων κυττάρων του φυτού με σκοπό την παρεμπόδιση του παθογόνου στις θέσεις διείσδυσης και την αποτροπή προέλασης του παθογόνου, πολλαπλασιασμού και μετακίνησης του μέσα στα κύτταρα καθώς και ανάπτυξης της ασθένειας. H ΑΥ ενεργοποιείται συνήθως από προϊόντα μεταβολισμού του παθογόνου (διεγέρτες) και προϋποθέτει μοριακή επικοινωνία αλληλοαναγνώρισης μεταξύ φυτού-ξενιστή και παθογόνου. Το παθογόνο, αφού προκαλέσει τον ξενιστή του να εκδηλώσει ΑΥ, αποτυγχάνει να πολλαπλασιαστεί σε μεγάλους πληθυσμούς και έτσι δεν εκφράζονται τα συμπτώματα της ασθένειας. Η εξέλιξη του φαινομένου αυτού συμβαίνει μέσα σε λίγες ώρες μετά την επαφή του ξενιστή με το παθογόνο και συμβαίνει μόνο στα σημεία της προσβολής στα ανθεκτικά φυτά (6,7) (Εικ. 1).
Έρευνες των τελευταίων χρόνων έχουν αποδείξει την ύπαρξη κοινών βιοχημικών μονοπατιών του ΠΚΘ μεταξύ φυτικών, ζωικών και μικροβιακών κυττάρων. Η διαδικασία αυτή στα φυτά δεν έχει εκτενώς μελετηθεί. Σε προηγούμενη μελέτη (Programmed Cell Death across the Eucaryotic Kingdoms, FP6-2003-LIFESCIHEALTH), ο Επ. Υπεύθυνος της μελέτης πραγματοποίησε μια λεπτομερή βιοπληροφορική ανάλυση του γενώματος του φυτού Arabidopsis thaliana για την εύρεση ορθόλογων γονιδίων που υπεισέρχονται στον ΠΚΘ σε άλλους οργανισμούς (θηλαστικά, μύκητες, έντομα, νηματώδεις). Εκτεταμένες αναλύσεις οδήγησαν στην ανακάλυψη μια σειράς γονιδίων που δεν είχαν χαρακτηρισθεί στα φυτά και πιθανόν να υπεισέρχονται στον κυτταρικό θάνατο και στην ευαισθησία της A.thaliana σε διάφορα παθογόνα. Το φυτό A. thaliana είναι εξαιρετικό ερευνητικό εργαλείο για τη Φυτοπαθολογία λόγω του ευρέως φάσματος παθογόνων που το προσβάλλουν, της πλήρους γνώσης του γονιδιώματος του, της εύκολης παραγωγής διαγονιδιακών φυτών και της ταχύτητας πολλαπλασιασμού του λόγω του μικρού βιολογικού του κύκλου.
Μια οικογένεια ιδιαίτερα ενδιαφέροντων γονιδίων που εντοπίστηκε και πιθανόν να εμπλέκεται στην ενεργοποίηση του αμυντικού συστήματος των φυτών είναι τα ορθόλογα γονίδια της πρωτεΐνης των θηλαστικών AIF-Apoptosis Inducing Factor με ουσιαστικό ρόλο στον ΠΚΘ. Η AIF των θηλαστικών είναι μια 57 kDa φλαβοπρωτεΐνη, η οποία διαθέτει συντηρημένες περιοχές και ομοιότητες με οξειδορεδουκτάσες των βακτηρίων, μυκήτων και φυτών (8). Η AIF πρωτεΐνη στον άνθρωπο σχετίζεται με ασθένειες αυξημένης απόπτωσης όπως η μόλυνση από τον ιό HIV, νευροδιαταραχές, ισχαιμία και καταστροφή του εγκεφάλου καθώς και καρδιακά επεισόδια. Ακολουθώντας το αποπτωτικό αποτέλεσμα και τη μιτοχονδριακή ομοιότητα, η
Figure 1: A. Μακροσκοπική εκδήλωση της αντίδρασης υπερευαισθησίας σε φύλλο καπνού μετά από εφαρμογή μέσω διήθησης του διεγέρτη Avr9 του μύκητα Cladosporium fulvum που αναγνωρίζεται από την πρωτείνη ανθεκτικότητας Cf9. B. Μικροσκοπική εκδήλωση της αντίδρασης υπερευαισθησίας σε επιδερμικό κύτταρο Αραβίδοψης ως αποτέλεσμα της παραγωγής H2O2 (ανίχνευση με diamino benzidine ως καστανό προιόν της αντίδρασης) μετά τη διείσδυση του παθογόνου Εrysiphe cichoracearum (μπλε υφές) που αναγνωρίζεται από την πρωτείνη ανθεκτικότητας RPW8 (2).

6
πρωτεΐνη AIF εδράζει στα κυτοχρώματα και στον πυρήνα από όπου ελέγχεται η σύνθεση χρωματίνης και ο σχηματισμός του DNA. Η AIF παίρνει μέρος στην οξείδωση του NAD(P)H και στις δραστηριότητες της μονο-δευδρο-ασκορβικής ρεδουκτάσης. Ομόλογα γονίδια της AIF έχουν βρεθεί σε νηματώδεις, μύκητες και φυτά. Το γενετικό φυτό μοντέλο Arabidopsis thaliana διαθέτει μια ομάδα 5 γονιδίων, ορθόλογων της πρωτεΐνης των θηλαστικών AIF (Apoptosis Inducing Factor) με κρίσιμο ρόλο στον ΠΚΘ.
ΣΚΟΠΟΣ ΤΗΣ ΜΕΛΕΤΗΣ
Βασικός σκοπός της παρούσας μελέτης ήταν η αξιολόγηση των At-AIF γονιδίων του φυτού A.thaliana, ως προς την ικανότητά εκδήλωσης του ΠΚΘ και της ευαισθησίας ή ανθεκτικότητάς τους σε μια σειρά σημαντικών παθογόνων των καλλιεργειών. Επίσης σκοπός ήταν να γίνει μια πρώτη προσέγγιση στη λειτουργική ανάλυση των At-AIF μέσω της έκφρασης διαφόρων γονιδίων που εμπλέκονται στην ενεργοποίηση του εγγενούς ανοσοποιητικού συστήματος των φυτών.
ΜΕΘΟΔΟΛΟΓΙΑ - ΑΠΟΤΕΛΕΣΜΑΤΑ
Προηγούμενες μελέτες του Επιστημονικού Υπεύθυνου οδήγησαν στην εύρεση των μεταλλαγμένων σειρών Α. thaliana στα γονίδια At-AIF2, At-AIF3 και At-AIF5. Η παρούσα μελέτη είχε σκοπό την ανεύρεση μεταλλαγμένων σειρών Α. thaliana στα γονίδια At-AIF1 και At-AIF4 και στον περαιτέρω χαρακτηρισμό τους. Η μελέτη πραγματοποιήθηκε στα πλαίσια των παρακάτω φάσεων των οποίων τα αποτελέσματα παρουσιάζονται:
1. Γονιδιακή σίγηση του At-AIF1
Εκτεταμένη γενετική ανάλυση των μεταλλαγμένων σειρών Α. thaliana (SALK_114156) στο γονίδιο At-AIF1 (η προμήθεια των T-DNA μεταλλαγμένων σειρών έγινε από την τράπεζα σπόρων European Arabidopsis Stock Centre - NASC) απέδειξε ότι δεν ήταν δυνατό να απομονωθούν ομοζυγωτικές σειρές με απενεργοποιημένο το At-AIF1. Αυτό υποδηλώνει ότι η πρωτείνη At-AIF1 είναι πιθανώς απαραίτητη για την ανάπτυξη του φυτού και απενεργοποίηση του γονιδίου καθιστά τις συγκεκριμένες σειρές πιθανόν μη βιώσιμες. Για να ξεπεραστεί αυτή η δυσκολία και να εξεταστεί ο ρόλος του At-AIF1 στη φυσιολογία και αμυντικό μηχανισμό των φυτών, πραγματοποιήθηκε γονιδιακή σίγηση με την πρωτοποριακή μέθοδο των “artificial mRNAs” όπως περιγράφεται στην Εικ. 2. Με αυτόν τον τρόπο η έκφραση του γονιδίου δεν θα είναι μηδενική και υποθέσαμε ότι τα μεταλλαγμένα aif1 φυτά θα επιβιώσουν.
Το artificial At-AIF1 microRNA κλωνοποιήθηκε στο φορέα pBIN19 υπό τον έλεγχο του υποεκκινητή 35S και του γονιδίου ανθεκτικότητας της καναμυκίνης (για γενετική επιλογή in planta) και στη συνέχεια με τη βοήθεια του Agrobacterium tumefaciens GV3101 μετασχηματίστηκε σε φυτά A. thaliana (Εικ. 3). Μεταλλαγμένοι σπόροι του A. thaliana με το artificial At-AIF1 microRNA επωάστηκαν σε θρεπτικό υπόστρωμα MS με την προσθήκη καναμικίνης ώστε να επιλεχθούν μόνο εκείνα τα φυτά τα οποία είχαν μετασχηματιστεί με το φορέα. Ακολούθησε επιλογή 50 μεταλλαγμένων φυτών και στη συνέχεια γενετική επιλογή με βάση τα επίπεδα του At-AIF1 mRNA. Τελικά επιλέχτηκαν 3 ομοζυγωτικές σειρές, aif1-C1, aif1-C2 και aif1-F6, που εκφράζαν το γονίδιο At-AIF1 σε χαμηλά ποσοστά συγκριτικά με το μάρτυρα, όπως φαίνεται στην Εικ. 4. Ακολούθησαν πειράματα θερμοκηπίου στα μεταλλαγμένα aif1 φυτά ώστε να προσδιοριστεί αν το γονίδιο At-AIF1 εμπλέκεται στο αμυντικό σύστημα του φυτού.

7
a b c
100
bpla
dder
1000 bp500 bp
d
100
bpla
dder
1000 bp
500 bp
Εικόνα 3: Επιλογή μεταλλαγμένων φυτών A. thaliana με το artificial At-AIF1 microRNA σε θρεπτικό υπόστρωμα MS+kanamycin που αποδεικνύει την παρουσία του φορέα pBIN19+At-AIF1 microRNA στο γενετικό τους υλικό.
Εικόνα 4: RT-PCR που δείχνει τα επίπεδα έκφρασης του γονιδίου At-AIF1 και του σταθερά εκφραζόμενου γονιδίου RPL4 (ribosomal protein L4: housekeeping gene) σε σειρές A. thaliana αγρίου τύπου Col-0 και μεταλλαγμένες aif1 σειρές με το artificial At-AIF1 microRNA (C1, C2, D1 και F6). Οι σειρές C2 και D1 δείχνουν τα χαμηλότερα ποσοστά έκφρασης του γονιδίου At-AIF1. H C2 είναι ομοζυγωτική ενώ η D1 είναι ετεροζυγωτική σειρά ως προς το At-AIF1 microRNA.
Εικόνα 2: Χρήση αλληλοεπικαλυπτόμενης PCR για τη δημιουργία των artificial AIF1 miRNAs. Οι εκκινητές I μέχρι IV σχεδιάστηκαν με τη βοήθεια του Web microRNA designer (http://wmd.weigelworld.org) και χρησιμοποιήθηκαν για να αντικαταστήσουν το miRNA (μπλε) του φορέα RS300 με τις αλληλουχίες των artificial miRNA (κόκκινο) του AIF1 (Α). Οι εκκινητές A και Β βασίζονται στην αλληλουχία του φορέα RS300 (miR319a pBSK). Με τη βοήθεια συνδυασμένης PCR τα προϊόντα a (A-IV), b (II-III) και c (I-B) που ενισχύθηκαν αρχικά (Β), χρησιμοποιήθηκαν ως εκμαγεία για την περαιτέρω ενίσχυση με τη βοήθεια των εκκινητών A και B λειτουργικών miRNAs (Γ). Η διαδικασία περιγράφεται πιο αναλυτικά στη δημοσίευση των Schwab et al., 2006 (9).
AIF1
RPL4
Col-0 C1 C2 D1 F6
Α
Β Γ

8
2. Ταυτοποίηση των aif4 μεταλλαγμένων σειρών
Προμηθευτήκαμε 3 μεταλλαγμένες σειρές Αραβίδοψης (SALK_030775; SALK_068667; SALK_087231) στις οποίες έχει γίνει ένθεση του T-DNA στο γονίδιο At-AIF4. Έγινε γονοτυπική ανάλυση των φυταρίων με τη βοήθεια της μεθόδου της PCR όπως έχει περιγραφεί (10) με σκοπό να βρεθούν ομοζυγωτικές σειρές στις οποίες το γονίδιο At-AIF4 έχει απενεργοποιηθεί. Μετά από εκτεταμένες μοριακές αναλύσεις καταφέραμε να βρούμε 3 σειρές (C1-B6-6, C1-B6-8 και C1-B6-9) στις οποίες γονοτυπικά το γονίδιο At-AIF4 διαπιστώθηκε ότι έχει απενεργοποιηθεί. Μετά από εξαγωγή γενωματικού DNA από τις διάφορες σειρές που προέκυψαν έγινε PCR με τους εκκινητές RP και LP (εξειδικευμένοι για το γονίδιο At-AIF4) (Εικ. 5Α) που δεν ενίσχυσαν μέρος του γονιδίου At-AIF4 στις σειρές 6, 8 και 9 (Εικ. 5Β-Ι.) λόγω της παρουσίας του ιδιαίτερα μεγάλου μεγέθους του Τ-DNA αλλά ενισχύθηκε το προιόν των ~500 βάσεων από τους εκκινητές LBb1 και RP (Εικ. 5Β-ΙΙ.) (Ο εκκινητής LBb1 βρίσκεται μέσα στο Τ-DNA).
3. Δοκιμές αντίδρασης υπερευαισθησίας στις aif σειρές
Στα πλαίσια της φάσης αυτής έγιναν πειράματα αλληλεπίδρασης των μεταλλαγμένων aif σειρών και του μάρτυρα Col-0 με τη φυλή Emwa1 του φυτοπαθογόνου ωομύκητα Hyaloperonospora arabidopsis (προηγούμενη ονομασία H. parasitica). Η φυλή Emwa1 δεν προκαλεί ασθένεια στον οικότυπο Col-0 της Αραβίδοψης λόγω της αναγνώρισης του γονιδίου ανθεκτικότητας RPP5 (μια TIR-NB-LRR πρωτείνη) και οδηγεί στην εκδήλωση της αντίδρασης υπερευαισθησίας (ΑΥ) και στην ανάπτυξη ανθεκτικότητας. Φυτά τριών εβδομάδων ψεκάστηκαν με αιώρημα κονιδίων του ωομύκητα σε συγκέντρωση 4x104 κονίδια/ml, καλύφθηκαν με πλαστική σακούλα για την διατήρηση υψηλής υγρασίας καθόλη τη διάρκεια της μόλυνσης και μετά από 6 ημέρες συλλέχθηκαν και ακολούθησε η βιοδοκιμή της ΑΥ που πραγματοποιείται με ένα ειδικό πρωτόκολλο που βασίζεται στη χρώση των νεκρών κυττάρων με Trypan Blue. Η παρατήρηση έγινε σε οπτικό μικροσκόπιο. Τα συγκεκριμένα πειράματα έδειξαν ότι μειώνεται σημαντικά η ανάπτυξη της ΑΥ (μη εμφάνιση νεκρών κυττάρων-μπλε μετά από χρώση με Trypan Blue) στις σειρές aif3 και aif5, υποδηλώνοντας ότι τα γονίδια At-AIF3 και At-AIF5 παίζουν σημαντικό ρόλο στην εκδήλωση της ΑΥ (Εικ. 6). (11). Οι σειρές aif1 και aif2 φαίνεται επίσης
RP
Α
LP
RP LP
LBb1
1 2 6 7 8 9Β
Ι.
ΙΙ.
Εικόνα 5: Α. Διάγραμμα σχεδιασμού γονοτυπικής ανάλυσης T-DNA μεταλλαγμένων σειρών A. thaliana. Β. PCR που ενίσχυσε μέρος του γονιδίου At-AIF4 (Ι) με τους εκκινητές RP-AIF4 και LP-AIF4 ή μέρος του T-DNA και του AIF4 με τους εκκινητές RP-AIF4 και LΒ1b σε μεταλλαγμένες σειρές A. thaliana. Οι σειρές 6, 8 και 9 είναι μεταλλαγμένες και ομοζυγωτικές σειρές στο γονίδιο At-AIF4.

9
ότι μειώνουν την εκδήλωση της ΑΥ ενώ η σειρά aif4 δεν παρουσιάζει κάποια διαφοροποίηση συγκριτικά με το Col-0.
Μάρτυρας Col-0 Μεταλλαγμένες σειρές aif3
Μάρτυρας Col-0 Μεταλλαγμένες σειρές aif5
Εικόνα 6. Άγριο στέλεχος Columbia (Col-0) και μεταλλαγμένες aif σειρές Αραβίδοψης, μολύνθηκαν σε ηλικία 3 εβδομάδων από τη βλάστησή τους, με κονίδια του ωομύκητα H. arabidopsis φυλή Εmwa2. 4 ημέρες μετά τη μόλυνση, έγινε συλλογή και χρώση των φύλλων με Trypan Blue, χρωστική που βάφει μπλε τα νεκρά κύτταρα και τις υφές των μυκήτων.
Μάρτυρας Col-0 Μεταλλαγμένες σειρές aif1
Μεταλλαγμένες σειρές aif2 Μεταλλαγμένες σειρές aif4

10
Πραγματοποιήθηκαν επίσης πειράματα αλληλεπίδρασης των μεταλλαγμένων aif σειρών με το μη μολυσματικό βακτηριακό στέλεχος Pseudomonas syringae pv. tomato που φέρει το γονίδιο αμολυσματικότητας AvrRpt2 και οδηγεί στην εκδήλωση της ΑΥ. Φυτά τεσσάρων εβδομάδων ψεκάστηκαν με αιώρημα βακτηρίων σε συγκέντρωση 108 βακτήρια/ml, καλύφθηκαν με πλαστική σακούλα για 2 ημέρες και μετά από 4 ημέρες συλλέχθηκαν και ακολούθησε η βιοδοκιμή της ΑΥ με τη χρώση των νεκρών κυττάρων με Trypan Blue. Τα συγκεκριμένα πειράματα έδειξαν ότι μειώνεται η εκδήλωση της ΑΥ στις aif3 και aif5 σειρές ενώ στις υπόλοιπες σειρές η μείωση της ΑΥ είναι αρκετά πιο μικρή.
Εικόνα 7. Άγριο στέλεχος Columbia (Col-0) και μεταλλαγμένες aif σειρές Αραβίδοψης, μολύνθηκαν σε ηλικία 4 εβδομάδων από τη βλάστησή τους, με βατήρια Pseudomonas syringae pv. tomato AvrRpt2. 4 ημέρες μετά τη μόλυνση, έγινε συλλογή και χρώση των φύλλων με Trypan Blue, χρωστική που βάφει μπλε τα νεκρά κύτταρα του φυτού.
Αμόλυντα
Col -0
aif1
aif2
aif3 aif4
aif5
Αμόλυντα
Col -0
aif1
aif2
aif3 aif4
aif5

11
Φ4. Δοκιμές παθογένειας στις aif σειρές
Πραγματοποιήθηκαν πειράματα παθογένειας των aif σειρών με διάφορα παθογόνα: α) τη φυλή Noco2 του φυτοπαθογόνου ωομύκητα που προκαλεί ασθένεια στην Αραβίδοψη (Col-0) β) το παθογόνο βακτήριο φυλλώματος Pseudomonas syringae pv. tomato DC3000, γ) τον εδαφογενή φυτοπαθογόνο μύκητα Verticillium dahliae, δ) τον εδαφογενή φυτοπαθογόνο μύκητα Fusarium oxysporum f.sp. raphani και δ) το φυτοπαθογόνο μύκητα φυλλώματος Alternaria brassicicola.
Μολύνσεις με το παθογόνο H. arabidopsis έδειξαν ότι τα μεταλλαγμένα φυτά aif5 και aif3 είναι πιο ευαίσθητα συγκριτικά με το μάρτυρα Col-0 (Εικ. 8A, 8B).
Μολύνσεις με το βακτήριο Pseudomonas syringae pv. tomato έδειξαν ότι τα μεταλλαγμένα φυτά aif3 είναι πιο ανθεκτικά στην ασθένεια από την αρχή της μόλυνσης, τα aif4 παρουσιάζουν ανθεκτικότητα μετά την 4η ημέρα ενώ σε όλες τις σειρές παρατηρείται μείωση του παθογόνου την 6η ημέρα υποδηλώνοντας ότι πιθανόν να ενεργοποιούνται μηχανισμοί αντοχής στα φυτά αυτά. (Εικ. 9).
0,0
0,5
1,0
1,5
2,0
2,5
Day 2 Day 4 Day 6
Log cfu/cm
2
Col‐0 aif1 aif2 aif3 aif4 aif5
Εικ. 8. Α. Μόλυνση φυτών Αραβίδοψης σε ηλικία 3 εβδομάδων με 4x104 κονίδια/ml του ωομύκητα H. arabidopsis φυλή Noco2 (συμβατή σχέση: ασθένεια). Καταμέτρηση των παραγομένων κονιδίων στα φύλλα, 7 ημέρες μετά τη μόλυνση. Β. Μόλυνση των aif μεταλλαγμένων φυτών Αραβίδοψης με τον ωομύκητα H. arabidopsis.
0
20
40
60
80
100
120
140
Κονίδια/φυτό
Κονίδια
X 1
000
/ φυτό
Col-0 aif2-1 aif3-1 aif5-1 aif5-2
Α
Εικ. 9. Α. Μόλυνση φυτών Αραβίδοψης σε ηλικία 5 εβδομάδων με ψεκασμό αιωρήματος βακτηρίων (108 βακτήρια/ml) του Pseudomonas syringae pv. tomato DC3000. Καταμέτρηση των βακτηρίων που αναπτύχθηκαν στα φύλλα, 3 ημέρες μετά τη μόλυνση. Β. Μόλυνση φυτού Αραβίδοψης με το βακτήριο Pseudomonas syringae pv. tomato DC3000
Α
Col-0 aif-2
aif-3 aif-5
Β
Β
Hyaloperonospora arabidopsis
Pseudomonas syringae pv. tomato DC3000

12
Μολύνσεις με το εδαφογενές παθογόνο V. dahliae έδειξαν ότι αυξάνεται η ευαισθησία των φυτών στις σειρές που είναι απενεργοποιημένο το γονίδιο At-AIF5 (Εικ. 10) που υποθέτουμε ότι οφείλεται στην εξασθένιση των αμυντικών μηχανισμών του φυτού. Αντίθετα, οι σειρές aif3 (Εικ. 11), aif1 (Εικ. 12) και aif4 (Εικ. 12) (21-25 ημέρες μετά τη μόλυνση) εμφανίζουν μεγαλύτερη ανθεκτικότητα ενώ οι σειρές aif2 (Εικ. 11) δεν διαφέρουν σημαντικά συγκριτικά με το μάρτυρα.
Εικ. 10. Α. Μόλυνση φυτών Αραβίδοψης σε ηλικία 5 εβδομάδων με ριζοπότισμα 10 ml αιωρήματος κονιδίων/φυτό (107 κονίδια/ml) Verticillium dahliae. H γραφική παράσταση δείχνει τα ποσοστά της ασθένειας από 19-33 ημέρες μετά τη μόλυνση. Β. Συμπτώματα του μύκητα Verticillium dahliae σε φυτά Αραβίδοψης Col-0 και aif5 21 (πάνω) και 33 (κάτω) ημέρες μετά τη μόλυνση με το εδαφογενές παθογόνο V. dahliae.
Col-0 aif5
21 ημ.
33 ημ.
Β
Α
Ημέρες μετά τη μόλυνση
Ποσοστό
ασθένειας
%
Col-0 aif5
Εικ. 11. Α. Μόλυνση φυτών Αραβίδοψης σε ηλικία 5 εβδομάδων με ριζοπότισμα 10 ml αιωρήματος κονιδίων/φυτό (107 κονίδια/ml) Verticillium dahliae. H γραφική παράσταση δείχνει τα ποσοστά της ασθένειας από 19-33 ημέρες μετά τη μόλυνση. Β. Συμπτώματα του μύκητα Verticillium dahliae σε φυτά Αραβίδοψης Col-0, aif3 και aif2, 25 ημέρες μετά τη μόλυνση με το εδαφογενές παθογόνο V. dahliae
Verticillium dahliae
Col-0 aif3 aif2
B Ημέρες μετά τη μόλυνση
Ποσοστό
ασθένειας
Α Col-0 aif3 aif2
Verticillium dahliae

13
Μολύνσεις με το εδαφογενές παθογόνο Fusarium oxysporum f.sp. raphani έδειξαν ότι οι σειρές aif2, aif3 και aif5 είναι πιο ευαίσθητες στο παθογόνο (Εικ. 13)
0
10
20
30
40
50
1 2 3 4 5 6
Ημέρες μετά τη μόλυνση
Ποσ
οστό ασθ
ενείας (%)
Col‐o aif3 aif2 aif5
10 12 14 16 18 20
0
50
100
150
200
250
300
350
400
450
Col‐o aif3 aif2 aif5
AUDPC
Ποσ
ό ασθ
ένειας
Εικ. 13. Α. Μόλυνση φυτών Αραβίδοψης σε ηλικία 5 εβδομάδων με ριζοπότισμα 10 ml αιωρήματος κονιδίων/φυτό (107 κονίδια/ml) Fusarium oxysporum f.sp. raphani. H γραφική παράσταση δείχνει τα ποσοστά της ασθένειας από 10-20 ημέρες μετά τη μόλυνση. Β. Τελικό ποσό ασθένειας του μύκητα Fusarium oxysporum f.sp. raphani σε φυτά Αραβίδοψης Col-0, aif2, aif3 και aif3 εκφρασμένο ως το εμβαδό AUDPC (Area Under Disease Progress Curve).
0
10
20
30
40
50
60
15ημ. 17ημ. 19ημ. 21ημ. 23ημ. 25ημ.
Col‐o aif1 aif4
Ποσ
οστό α
σθένειας (%
)
Ημέρες μετά τη μόλυνση
Εικ. 12. Α. Μόλυνση μεταλλαγμένων σειρών aif1 και aif4 Αραβίδοψης σε ηλικία 5 εβδομάδων με ριζοπότισμα 10 ml αιωρήματος κονιδίων/φυτό (107 κονίδια/ml) Verticillium dahliae. H γραφική παράσταση δείχνει τα ποσοστά της ασθένειας από 15-25 ημέρες μετά τη μόλυνση.
Verticillium dahliae
Fusarium oxysporum f.sp. raphani
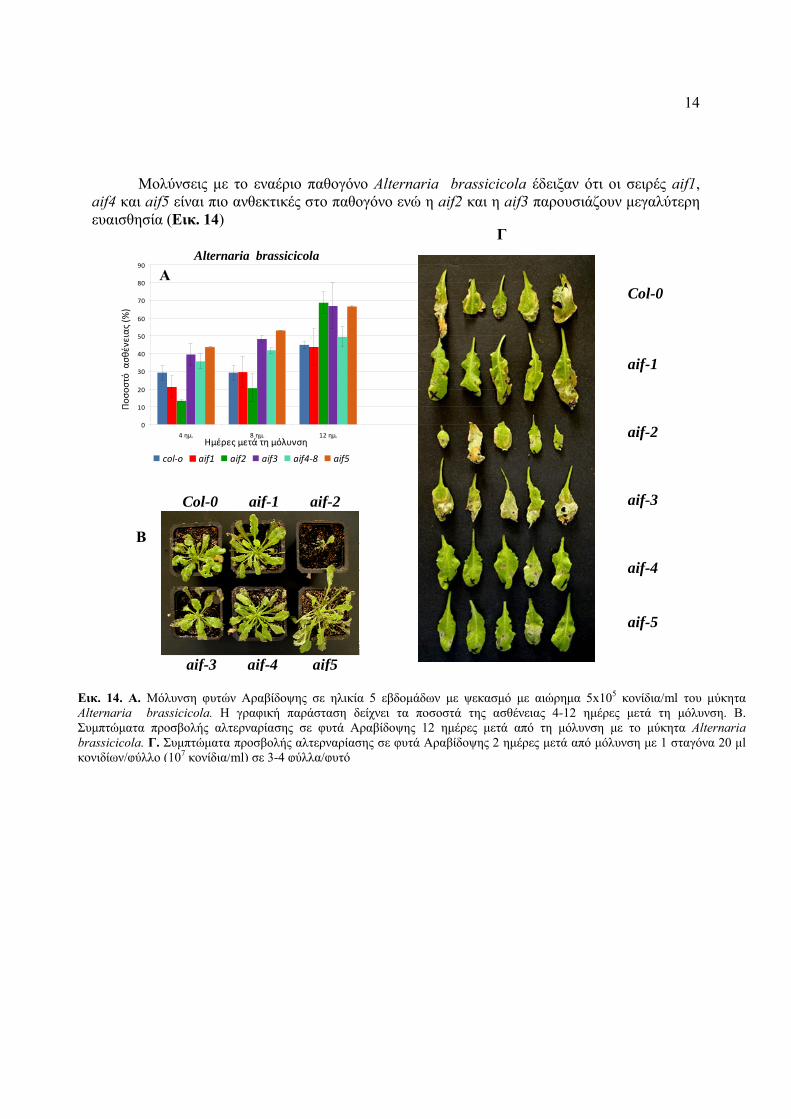
14
Μολύνσεις με το εναέριο παθογόνο Alternaria brassicicola έδειξαν ότι οι σειρές aif1, aif4 και aif5 είναι πιο ανθεκτικές στο παθογόνο ενώ η aif2 και η aif3 παρουσιάζουν μεγαλύτερη ευαισθησία (Εικ. 14)
Col-0
aif-1
aif-2
aif-3
aif-4
aif-5
Col-0 aif-1 aif-2
aif-3 aif-4 aif5
Εικ. 14. Α. Μόλυνση φυτών Αραβίδοψης σε ηλικία 5 εβδομάδων με ψεκασμό με αιώρημα 5x105 κονίδια/ml του μύκητα Alternaria brassicicola. H γραφική παράσταση δείχνει τα ποσοστά της ασθένειας 4-12 ημέρες μετά τη μόλυνση. Β. Συμπτώματα προσβολής αλτερναρίασης σε φυτά Αραβίδοψης 12 ημέρες μετά από τη μόλυνση με το μύκητα Alternaria brassicicola. Γ. Συμπτώματα προσβολής αλτερναρίασης σε φυτά Αραβίδοψης 2 ημέρες μετά από μόλυνση με 1 σταγόνα 20 μl κονιδίων/φύλλο (107 κονίδια/ml) σε 3-4 φύλλα/φυτό
Α
Β
Γ
0
10
20
30
40
50
60
70
80
90
4 ημ. 8 ημ. 12 ημ.
col‐o aif1 aif2 aif3 aif4‐8 aif5
Ημέρες μετά τη μόλυνση
Ποσ
οστό α
σθένειας (%
)
Alternaria brassicicola

15
Φ5. Έκφραση γονιδίων του αμυντικού μηχανισμού στα aif μεταλλαγμένα φυτά
Τα φυτά μέσω του εγγενούς ενεργητικού ανοσοποιητικού συστήματος αντιλαμβάνονται τα αρχικά στάδια προσβολής από τα διάφορα παθογόνα και μεταφέρουν αυτή την επαφή σε μια σειρά βιοχημικών αντιδράσεων που οδηγούν στην ΑΥ. Η αναγνώριση του παθογόνου γίνεται άμεσα με διάφορες αλλαγές που συμβαίνουν στη βιοχημική σύσταση των κυττάρων όπως στη σύσταση των μεμβρανών σε ιόντα, τοπική συσσώρευση μικρομοριακών ενώσεων (σαλικυλικό οξύ, οξείδια του αζώτου (ΝΟ), ενεργές ρίζες οξυγόνου (ROS) οι οποίες είναι σε θέση να κατευθύνουν τα κύτταρα σε ΠΚΘ (2, 3, 5). Επιπλέον, τα φυτά προσπαθούν να ενισχύσουν το αμυντικό τους σύστημα και με άλλους τρόπους όπως είναι η εναπόθεση στα κυτταρικά τους τοιχώματα στρωμάτων λιγνίνης και καλλόζης για ενίσχυση των φυσικών φραγμών του φυτού και τη σύνθεση αντιμικροβιακών μεταβολιτών όπως οι φυτοαλεξίνες για να εμποδίσουν την ανάπτυξη των παθογόνων.
Μετά την πάροδο λίγων λεπτών/ωρών από την αλληλεπίδραση του παθογόνου με τον ξενιστή, η μεταγωγή του σήματος της επαγωγής της AY οφείλεται σε μια σειρά πρωτεινών παθογένειας (Pathogenesis Related – PR γονίδια), πρωτεινικών κινασών, λιποξυγενασών (LOX), μεταγραφικών παραγόντων (π.χ WRKY 33 και 53), της πρωτείνης PDF1.2. (plant defencin) και άλλων πρωτεϊνών. Οι διεγέρτες επίσης των παθογόνων συμβάλλουν στην ενεργοποίηση των πρωτεϊνών παθογένειας, στη σύνθεση φυτοαλεξινών και στην ενεργοποίηση των βιοχημικών μονοπατιών του σαλικυλικού οξέος, του ιασμονικού οξέος και του αιθυλενίου. Η επαγωγή των γονιδίων PR-1, PR-2 (β-1,3-glucanase) και PR-5 (thaumatin-like protein) χρειάζεται τη μεσολάβηση του σαλικυλικού οξέος ως σήματος ενώ η επαγωγή των γονιδίων PR-3, PR-4 (endochitinases) και VSP1 (Vegetative Storage Protein 1) εξαρτάται από το ιασμονικό οξύ. Οι μεταγραφικοί παράγοντες WRKY παίζουν σημαντικό ρόλο στην ενεργοποίηση του αμυντικού συστήματος των φυτών (Εικ. 15).
Κατά τη φάση αυτή της μελέτης πραγματοποιήθηκε ανάλυση των aif σειρών μετά την μόλυνση τους με το παθογόνο Pseudomonas syringae pv. tomato DC3000, ώστε να αξιολογηθεί η ικανότητα μεταβίβασης του σήματος σε γονιδιακό επίπεδο στις διάφορες aif σειρές με τη βοήθεια της μεθόδου RT- PCR.
Συγκεκριμένα, φυτά τεσσάρων εβδομάδων ψεκάστηκαν με αιώρημα βακτηρίων σε συγκέντρωση 107 βακτήρια/ml και μετά από 2 ημέρες συλλέχθηκαν φύλλα από κάθε aif σειρά και το μάρτυρα Col-0, που είχαν μολύνθεί με το παθογόνο βακτήριο ή ήταν αμόλυντα (για να πραγματοποιηθεί σύγκριση ως την έκφραση διαφόρων γονιδίων). Ακολούθησε εξαγωγή RNA και χρησιμοποιώντας 1 μg RNA από κάθε δείγμα έγινε σύνθεση cDNA (με Αντίστροφη Μεταγραφάση) και ακολούθησε PCR με εκκινητές που αντιστοιχούσαν σε κάθε υπό εξέταση γονίδιο. Ως γονίδιο αναφοράς και χρήσης της ίδιας ποσότητας cDNA σε όλα τα δείγματα
Εικ. 15. Βασική σχηματική απεικόνιση κύριων μονοπατιών και γονιδίων που ενεργοποιούνται μετά την αλληλεπίδραση παθογόνων/εντόμων και του αμυντικού συστήματος του φυτού Arabidopsis.

16
χρησιμοποιήθηκε το γονίδιο RPL4 (ribosomal protein L4: housekeeping gene) (Εικ. 16) που η έκφραση του δεν επηρεάζεται από τις μεταλλάξεις των aif σειρών.
Με βάση τα αποτελέσματα της μεταγραφικής ανάλυσης (Εικ. 16) διαπιστώνεται ότι στις σειρές aif3, aif4 και aif5 επάγεται σημαντικά το γονίδιο PR-1 (β-1,3-glucanase) μετά τη μόλυνση ενώ μειώνεται η έκφρασή του στη σειρά aif2. Τα γονίδια PR-2 (β-1,3-glucanase), PR-4 και PR-5 (thaumatin-like protein) δεν φαίνεται να επηρεάζονται από τις διάφορες μεταλλάξεις εκτός από μια αυξημένη επαγωγή του γονιδίου PR-4 στην aif1. Ιδιαίτερα ενδιαφέρουσα είναι η επαγωγή της λιποξυγενάσης LOX3 σε όλες τις aif σειρές (εκτός της aif3) μετά τη μόλυνση με το βακτήριο ενώ παρατηρείται μια υπερέκφραση του γονιδίου στις σειρές aif3, aif4 ακόμα και χωρίς την παρουσία του παθογόνου. Η έκφραση του γονιδίου VSP1 καταστέλλεται στις σειρές aif1, aif3 και aif4 χωρίς την παρουσία του παθογόνου ενώ μετά την μόλυνση με το παθογόνο επάγεται σε μικρό βαθμό μόνο στις σειρές aif3 και aif4. Όσον αφορά την έκφραση των WRKY μεταγραφικών παραγόντων, τα γονίδια WRKY33 και WRKY53 εμφανίζουν αυξημένη έκφραση σε όλες τις aif σειρές (ιδιαίτερα στην περίπτωση του WRKY53) χωρίς την παρουσία του παθογόνου ενώ μετά τη μόλυνση το γονίδιο WRKY33 επάγεται μόνο στην aif1 σειρά ενώ μειώνεται η έκφραση του στις σειρές aif3, aif4 και aif5. Η έκφραση τo γονιδίου WRKY53 μειώνεται σημαντικά στις σειρές aif3, aif4 και aif5 ενώ δεν παρατηρείται επαγωγή του γονιδίου στις σειρές aif1 και aif2 όπως παρατηρείται στο Col-0. Το γονίδιο PDF1.2 εμφανίζει μειωμένη έκφραση στη σειρά aif1 χωρίς την παρουσία του παθογόνου ενώ επάγεται μετά τη μόλυνση σε αντίθεση με το Col-0 που η έκφρασή του καταστέλλεται. Στις άλλες aif σειρές δεν παρατηρούνται σημαντικές διαφοροποιήσεις όσον αφορά την έκφραση του γονιδίου PDF1.2.
RPL4
PR-1
PR-2
PR-4
PR-5
LOX3-1
PDF-1.2
WRKY33
VSP1
WRKY53
Ladd
erCo
l-0aif
-1aif
-2aif
-3aif
-4aif
-5
Ladd
erCo
l-0aif
-1aif
-2aif
-3aif
-4aif
-5
Μάρτυρας Pseudomonas syringae pv. tomato
RPL4
PR-1
PR-2
PR-4
PR-5
LOX3-1
PDF-1.2
WRKY33
VSP1
WRKY53
Ladd
erCo
l-0aif
-1aif
-2aif
-3aif
-4aif
-5
Ladd
erCo
l-0aif
-1aif
-2aif
-3aif
-4aif
-5
Ladd
erCo
l-0aif
-1aif
-2aif
-3aif
-4aif
-5
Ladd
erCo
l-0aif
-1aif
-2aif
-3aif
-4aif
-5
Μάρτυρας Pseudomonas syringae pv. tomato
Εικ. 16. Έκφραση διαφόρων γονιδίων που εμπλέκονται στο αμυντικό σύστημα του φυτού Arabidopsis στις aif σειρές σε αμόλυντα φυτά ή μετά τη μόλυνση με το παθογόνο βακτήριο Pseudomonas syringae pv. tomato DC3000.

17
Σε μια περαιτέρω προσπάθεια να αναλυθεί η έκφραση των AIF γονιδίων μετά από την αλληλεπίδραση φυτών Αραβίδοψης με διάφορα παθογόνα, διεγέρτες και ορμόνες έγινε χρήση των προγραμμάτων γονιδιακής ανάλυσης Arabidopsis eFP Browser (http://www.bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi) και Genevestigator (www.genevestigator. com) και διαπιστώθηκαν τα εξής (Εικ. 17,18):
1) η έκφραση του γονιδίου At-AIF1 καταστέλλεται μετά από 24 h από τη μόλυνση φυτών Arabidopsis με τα παθογόνα Pseudomonas syringae pv. tomato DC3000 και Phytophthora infestans ενώ φαίνεται να επάγεται μετά από 5 ημέρες μόλυνση με το μύκητα Erysiphe orontii που προκαλεί την ασθένεια ωίδιο.
2) το γονίδιο At-AIF2 υπερεκφράζεται κυρίως μετά από μόλυνση με το μύκητα B. cinerea (αύξηση 2,46 φορές), με τον ωομύκητα P. infestans (αύξηση 2,57 φορές), με μολυσματικά και αμολυσματικά στελέχη του Pseudomonas syringae pv. tomato καθώς και μετά από εφαρμογή των διεγερτών Flg22 (φλατζελίνη, PAMP διεγέρτης) και συριγκολίνη (syringolin) που εκκρίνεται από το βακτήριο P. syringae pv. syringae (αύξηση 2,37 φορές).
3) η έκφραση του γονιδίου At-AIF3 αυξάνεται κυρίως μετά από μόλυνση με το μύκητα B. cinerea και λιγότερο μετά από μόλυνση με το βακτήριο Pseudomonas syringae pv. tomato DC3000 και μετά από 5 ημέρες μόλυνση με το μύκητα Erysiphe orontii.
4) η έκφραση του γονιδίου At-AIF4 καταστέλλεται μετά από μόλυνση με το βακτήριο Pseudomonas syringae pv. tomato και τον ωομύκητα Phytophthora infestans.
5) το γονίδιο At-AIF5 υπερεκφράζεται σημαντικά μετά από μόλυνση με το μύκητα Botrytis cinerea, P. syringae, του διεγέρτη Flg22 και της ορμόνης methyl jasmonate.
Εικ. 17. Αποτελέσματα έκφρασης των γονιδίων At-AIF5 και At-AIF2 μετά την αλληλεπίδραση φυτών Αραβίδοψης με διάφορα παθογόνα ή ορμόνες. Το κόκκινο χρώμα υποδηλώνει αύξηση και το πράσινο ελάττωση της έκφρασης του γονιδίου. Η ανάλυση έγινε με τη βοήθεια των αποτελεσμάτων ενός μεγάλου αριθμού μικροσυστοιχιών που επεξεργάζεται το πρόγραμμα Genevestigator.
Treatment # of Chips Mean Std-
Err Ratio Std-Err Mean # of
Chips Control
Biotic: B. cinerea (+) 6 2938 1270 5.6 92 525 6 Biotic: B. cinerea (-)
Biotic: E. cichoracearum (+) 4 570 57 2.6 20 219 4 Biotic: E. cichoracearum (-)
Biotic: E. orontii (+) 24 210 37 1.86 19 113 24 Biotic: E. orontii (-)
Biotic: P. infestans (+) 6 202 23 2.62 19 77 6 Biotic: P. infestans (-)
Biotic: P. syringae (+) 9 3998 1051 2.25 561 1779 9 Biotic: P. syringae (-)
Chemical: syringolin (+) 3 260 22 1.34 117 194 3 Chemical: syringolin (-)
Hormone: ABA (+) 4 812 171 0.63 94 1290 4 Hormone: ABA (-)
Hormone: ACC (+) 4 1547 231 1.2 94 1290 4 Hormone: ACC (-)
Hormone: BL (+) 4 1865 368 1.45 94 1290 4 Hormone: BL (-)
Hormone: GA3 (+) 4 1533 173 1.19 94 1290 4 Hormone: GA3 (-)
Hormone: IAA (+) 4 1392 197 1.08 94 1290 4 Hormone: IAA (-)
Hormone: MJ (+) 4 12328 1523 9.56 94 1290 4 Hormone: MJ (-)
Hormone: zeatin (+) 4 1191 153 0.92 94 1290 4 Hormone: zeatin (-)
At-A
IF5
Treatment # of Chips Mean Std-
Err Ratio Std-Err Mean # of
Chips Control
Biotic: B. cinerea (+) 6 12505 2188 2.46 693 5088 6 Biotic: B. cinerea (-)
Biotic: E. cichoracearum (+) 4 3386 172 1.43 242 2368 4 Biotic: E. cichoracearum (-)
Biotic: E. orontii (+) 24 4370 292 1.12 211 3906 24 Biotic: E. orontii (-)
Biotic: P. infestans (+) 6 9615 1050 2.57 679 3744 6 Biotic: P. infestans (-)
Biotic: P. syringae (+) 9 8324 1299 1.53 300 5434 9 Biotic: P. syringae (-)
Chemical: syringolin (+) 3 5453 1013 2.37 302 2303 3 Chemical: syringolin (-)
Hormone: ABA (+) 4 4848 402 0.83 293 5876 4 Hormone: ABA (-)
Hormone: ACC (+) 4 6152 552 1.05 293 5876 4 Hormone: ACC (-)
Hormone: BL (+) 4 5133 414 0.87 293 5876 4 Hormone: BL (-)
Hormone: GA3 (+) 4 5575 310 0.95 293 5876 4 Hormone: GA3 (-)
Hormone: IAA (+) 4 5913 392 1.01 293 5876 4 Hormone: IAA (-)
Hormone: MJ (+) 4 7725 799 1.31 293 5876 4 Hormone: MJ (-)
Hormone: zeatin (+) 4 4721 195 0.8 293 5876 4 Hormone: zeatin (-)
At-A
IF2

18
At-AIF1

19At-AIF2

20At-AIF3

21At-AIF4

22
At-AIF5
Εικ. 18. Αποτελέσματα έκφρασης των 5 γονιδίων At-AIF μετά την αλληλεπίδραση φυτών Αραβίδοψης με διάφορα παθογόνα, διεγέρτες ή ορμόνες. Το κόκκινο χρώμα υποδηλώνει αύξηση και το μπλε ελάττωση της έκφρασης του γονιδίου. Η ανάλυση έγινε με τη βοήθεια των αποτελεσμάτων ενός μεγάλου αριθμού μικροσυστοιχιών που επεξεργάζεται το πρόγραμμα Arabidopsis eFP Browser .

23
Στα παρακάτω σχηματικά διαγράμματα (Εικ. 19) με τη βοήθεια του προγράμματος Arabidopsis eFP Browser (http://www.bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi) μελετήθηκε η έκφραση των διαφόρων At-AIF γονιδίων στους διάφορους ιστούς του φυτού Arabidospis κατά την ανάπτυξη του υπό φυσιολογικές συνθήκες. Όπως φαίνεται η έκφραση του γονιδίου At-AIF1 εστιάζεται κυρίως στα φύλλα της ροζέτας των φύλλων του φυτού, του At-AIF2 στο άνθος και φύλλα της ροζέτας που βρίσκονται στο στάδιο της ωρίμανσης-νέκρωσής τους, του At-AIF3 σε όλα τα φύλλα και άνθος και του At-AIF4 στις κοτυληδόνες, στα φύλλα του ανθικού στελέχους και στα φύλλα της ροζέτας που βρίσκονται στο στάδιο της ωρίμανσης-νέκρωσης. Ιδιαίτερο ενδιαφέρον παρουσιάζει το γονίδιο At-AIF5 το οποίο δεν φαίνεται να εκφράζεται κάτω από φυσιολογικές συνθήκες ανάπτυξης του φυτού παρά μόνο σε χαμηλά επίπεδα στο ριζικό σύστημα και στις κοτυληδόνες του.
At-AIF1

24
At-AIF2
At-AIF3

25
At-AIF4
At-AIF5
Εικ. 19. Αποτελέσματα έκφρασης των 5 γονιδίων At-AIF στους διάφορους ιστούς του φυτού Arabidospis thaliana κατά την ανάπτυξη του υπό φυσιολογικές συνθήκες.. Η ανάλυση έγινε με τη βοήθεια των αποτελεσμάτων ενός μεγάλου αριθμού μικροσυστοιχιών που επεξεργάζεται το πρόγραμμα Arabidopsis eFP Browser .

26
6. Δημιουργία διπλών μεταλλαγμένων σειρών
Πραγματοποιήθηκαν αλλεπάλληλες γενετικές διασταυρώσεις των α) aif2 Χ aif3 και β) αif2 Χ aif5 με τα συνήθη πρωτόκολλα (11) και ακολούθησε γονοτυπική ανάλυση των σπόρων που παρήχθησαν από τις διασταυρώσεις για την επιβεβαίωση των διπλά μεταλλαγμένων σειρών (Εικ. 20). Έγινε εξαγωγή DNA από τα φυτά που προέκυψαν και ακολούθησε PCR με εξειδικευμένους εκκινητές. Η γονοτυπική ανάλυση που παρουσιάζεται στην Εικ. 21 δείχνει την επιτυχή δημιουργία των διπλά μετταλλαγμένων ετεροζυγωτικών σειρών aif2 X aif5 –4 (Εικ. 21Β) και aif2 X aif3 –9 (Εικ. 21Γ) λόγω της παρουσίας των ζωνών με τους εξειδικευμένους εκκινητές RP+LP και RP+LBb1 για τα γονίδια At-AIF5 (Εικ. 21Β) και At-AIF3 (Εικ. 21Γ) και της ζώνης με τους εκκινητές RP+LBb1 για το γονίδιο At-AIF2 (Εικ. 21Δ).
Εικ. 20. Α. Αφαίρεση των στημόνων και εμφάνιση του στύλου του άνθους πριν τη γονιμοποίηση. Β. Στήμονες άνθους Αραβίδοψης που χρησιμοποιούνται για τη γονιμοποίηση του στύλου. Γ. Γονιμοποιημένο άνθος και εμφάνιση του καρπού (κέρας) της διασταύρωσης aif2 X aif3
Α Β
Γ aif2 X aif5
RP+L
P
RP+L
Bb1
RP+L
P
RP+L
Bb1
RP+L
PRP
+LBb
1
Αif5 εκκινητές
3 4 5
aif2 X aif3
RP+L
P
RP+L
Bb1
RP+L
P
RP+L
Bb1
RP+L
PRP
+LBb
1
Αif3 εκκινητές
9 10 11
AIF2 RP+LBb1
Col-0 9 10 11 3 4 5 ladder
Α
Β
Γ
Δ
aif2 X aif3 aif2 X aif5
aif2 X aif5
RP+L
P
RP+L
Bb1
RP+L
P
RP+L
Bb1
RP+L
PRP
+LBb
1
Αif5 εκκινητές
3 4 5
aif2 X aif3
RP+L
P
RP+L
Bb1
RP+L
P
RP+L
Bb1
RP+L
PRP
+LBb
1
Αif3 εκκινητές
9 10 11
aif2 X aif3
RP+L
P
RP+L
Bb1
RP+L
P
RP+L
Bb1
RP+L
PRP
+LBb
1
Αif3 εκκινητές
9 10 11
AIF2 RP+LBb1
Col-0 9 10 11 3 4 5 ladder
Α
Β
Γ
Δ
aif2 X aif3 aif2 X aif5
Εικ. 21. Α. Σχηματική ανάλυση γονοτυπικής ανάλυσης με τους εκκινητές RP, LP και LBb1 και παρουσία μονών ή διπλών ζωνών για την εξακρίβωση αν μια σειρά είναι ετεροζυγωτική ή ομοζυγωτική για το υπό μελέτη γονίδιο. Β. Ανάλυση προιόντων PCR με εξειδικευμένους εκκινητές για το γονίδιο At-AIF5 για τις σειρές aif2 x aif5. Γ. Ανάλυση προιόντων PCR με εξειδικευμένους εκκινητές για το γονίδιο At-AIF3 για τις σειρές aif2 x aif3 Δ. Ανάλυση προιόντων PCR με εξειδικευμένους εκκινητές για το γονίδιο At-AIF2 για τις σειρές aif2 x aif3 και aif2 x aif5

27
ΣΥΜΠΕΡΑΣΜΑΤΑ
Κατά την παρούσα μελέτη έγινε χαρακτηρισμός των 5 μεταλλαγμένων aif σειρών και διαπιστώθηκε για πρώτη φορά ότι τα At-AIF γονίδια παίζουν σημαντικό ρόλο στην ικανότητα ενεργοποίησης του εγγενούς ανοσοποιητικού συστήματος του φυτού Arabidopsis thaliana και της ευαισθησίας ή ανθεκτικότητάς τους σε μια σειρά σημαντικών παθογόνων των καλλιεργειών.
Συγκεκριμένα τα γονίδια At-AIF3 και At-AIF5 και σε μικρότερο βαθμό τα γονίδια At-AIF1 και At-AIF2 έχουν την ικανότητα να καταστέλλουν τον κυτταρικό θάνατο (αντίδραση υπερευαισθησίας) μετά από μόλυνση με μη συμβατά παθογόνα (Hyaloperonospora arabidopsis Emwa2 και Pseudomonas syringae pv. tomato AvrPt2) χωρίς όμως να «σπάνε» τελείως την ανθεκτικότητα του ξενιστή.
Τα αποτελέσματα των δοκιμών παθογένειας δείχνουν ότι οι 5 aif σειρές συμπεριφέρονται διαφορετικά ανάλογα με το παθογόνο, με μείωση ή αύξηση της ευαισθησίας-ανθεκτικότητάς τους γεγονός που υποδηλώνει ότι τα At-AIF γονίδια εμπλέκονται σε διαφορετικά μονοπάτια του εγγενούς ανοσοποιητικού συστήματος. Από όλες τις σειρές συμπεραίνουμε ότι η aif5 παρουσιάζει μια μεγαλύτερη ευαισθησία στα περισσότερα παθογόνα που εξετάσαμε.
Τα φυτά μέσω του εγγενούς ενεργητικού ανοσοποιητικού συστήματος αντιλαμβάνονται τα αρχικά στάδια προσβολής από τα διάφορα παθογόνα και μεταφέρουν αυτή την επαφή σε μια σειρά βιοχημικών αντιδράσεων που οδηγούν στην ΑΥ. Μετά την πάροδο λίγων λεπτών/ωρών από την αλληλεπίδραση του παθογόνου με τον ξενιστή, η μεταγωγή του σήματος της επαγωγής της AY οφείλεται σε μια σειρά γονιδίων - πρωτεϊνών. Η αναλυτική έκφραση γονιδίων που σχετίζονται με τον αμυντικό μηχανισμό στα aif μεταλλαγμένα φυτά έδειξε ότι όλα τα At-AIF γονίδια παίζουν σημαντικό ρόλο στην έκφραση διαφόρων γονιδίων του αμυντικού συστήματος του φυτού A. thaliana σε διαφορετικό βαθμό το καθένα και ακολουθώντας διαφορετικό μονοπάτι.
Περαιτέρω έρευνες κρίνονται απαραίτητες για την αξιολόγηση των At-AIF γονιδίων και θα εστιαστούν σε α) πειράματα έκφρασης γονιδίων του αμυντικού συστήματος του φυτού A. thaliana μετά από μόλυνση με διάφορα συμβατά και μη συμβατά παθογόνα β) πειράματα παθογένειας με τις διπλά μεταλλαγμένες σειρές, γ) πειράματα δημιουργίας μεταλλαγμένων φυτών στα οποία τα At-AIF γονίδια υπερεκφράζονται και δ) πειράματα εντοπισμού των At-AIF γονιδίων σε κυτταρικό και ιστολογικό επίπεδο με τη χρήση των γονιδίων GFP και GUS.

28
ΠΑΡΑΔΟΤΕΑ
Τα παραδοτέα της συγκεκριμένης μελέτης είναι: • η δημιουργία διαγονιδιακών μεταλλαγμένων σειρών στα AIF γονίδια που μελετώνται για
πρώτη φορά παγκοσμίως στα φυτά
o Κατά τη φάση 1 και 2 δημιουργήθηκαν οι σειρές aif1 και aif4, οπότε ολοκληρώθηκε η δημιουργία όλων των διαγονιδιακών μεταλλαγμένων aif σειρών
• η διαλεύκανση του ρόλου των AIF πρωτεϊνών στην εκδήλωση του προγραμματισμένου κυτταρικού θανάτου (αντίδραση υπερευαισθησίας)
o Στη φάση 3 περιγράφεται ο ρόλος των των AIF γονιδίων στην εκδήλωση αντίδραση υπερευαισθησίας
• η συμβολή του ρόλου των AIF πρωτεϊνών στην παθογένεια διαφόρων φυτοπαθογόνων
o Στη φάση 4 περιγράφεται η συμβολή του ρόλου των AIF πρωτεϊνών στην παθογένεια διαφόρων φυτοπαθογόνων και συγκεκριμένα: α) τη φυλή Noco2 του φυτοπαθογόνου ωομύκητα H. arabidopsis β) το παθογόνο βακτήριο φυλλώματος Pseudomonas syringae pv. tomato DC3000, γ) τον εδαφογενή φυτοπαθογόνο μύκητα Verticillium dahliae, δ) τον εδαφογενή φυτοπαθογόνο μύκητα Fusarium oxysporum f.sp. raphani και δ) το φυτοπαθογόνο μύκητα φυλλώματος Alternaria brassicicola.
• η αναλυτική έκφραση των γονιδίων του αμυντικού μηχανισμού στα aif μεταλλαγμένα φυτά και στην καλύτερη κατανόηση της αλληπίδρασης ξενιστή-παθογόνου (καταστολή ή επαγωγή του ΠΚΘ)
o Στη φάση 5 περιγράφεται α) η αναλυτική έκφραση των At-AIF γονιδίων στα διάφορα στάδια ανάπτυξης του φυτού A. thaliana και β) η αναλυτική έκφραση διαφόρων γονιδίων που σχετίζονται με τον αμυντικό μηχανισμό στα aif μεταλλαγμένα φυτά
• η δημιουργία διπλών και τριπλών μεταλλαγμένων aif2, aif3 και aif5 σειρών
o Στη φάση 6 περιγράφεται η δημιουργία των aif2 x aif5 και aif2 x aif3 ετεροζυγωτικών σειρών. Η διπλή μεταλλαγμένη σειρά aif3 x aif5 παρόλο το μεγάλο αριθμό προσπαθειών που πραγματοποιήθηκαν δεν μπόρεσε να δημιουργηθεί (ίσως αυτό να σημαίνει ότι η συγκεκριμένη διπλή σειρά δεν μπορεί να επιβιώσει). Ως εκ τούτου και η δημιουργία της τριπλής σειράς δεν ήταν δυνατόν να πραγματοποιηθεί.
• η πιθανή χρησιμοποίηση των AIF πρωτεϊνών σε πειράματα γενετικής βελτίωσης των φυτών για ανθεκτικότητα σε ασθένειες
o περαιτέρω έρευνα των ιδιαίτερα ενδιαφέροντων αποτελεσμάτων αυτής της μελέτης θα συμβάλλει στην πιθανή χρήση των AIF πρωτεϊνών σε πειράματα γενετικής βελτίωσης των φυτών για ανθεκτικότητα σε ασθένειες
• η ευρύτερη κατανόηση της λειτουργίας και της εξέλιξης του προγραμματισμένου κυτταρικού θανάτου των φυτών συγκριτικά με τα θηλαστικά.

29
o Τα αποτελέσματα της μελέτης οδηγούν στο συμπέρασμα ότι τα At-AIF γονίδια παίζουν σημαντικό ρόλο στη λειτουργία και εξέλιξη του προγραμματισμένου κυτταρικού θανάτου των φυτών όπως και στα θηλαστικά.
ΔΙΑΧΥΣΗ ΤΩΝ ΑΠΟΤΕΛΕΣΜΑΤΩΝ
Τα αποτελέσματα της παρούσης μελέτης παρουσιάστηκαν α) στο 12ο Μεσογειακό Φυτοπαθολογικό Συνέδριο στη Ρώμη (20-25 Ιουνίου 2010) από τον Επιστημονικό Υπεύθυνο και β) στο 15ο Πανελλήνιο Φυτοπαθολογικό Συνέδριο στην Κέρκυρα (5-8 Οκτωβρίου 2010, Ξενοδοχείο Aquis Corfu Holiday Palace) από την κα Σ. Κουντούρη και τον Επιστημονικό Υπεύθυνο της παρούσης μελέτης.
ΒΙΒΛΙΟΓΡΑΦΙΑ
1. Baehrecke, E.H. How death shapes life during development, (2002) Nature Reviews in Molecular and Cellular
Biology, 3 pp. 779–787. 2. Hofius, D., Tsitsigiannis, D.I., Jones, J.D. and Mundy, J. (2007). Inducible cell death in plant immunity.
Seminars in Cancer Biology, 17(2):166-187 3. Lam, E. (2004) Controlled cell death, plant survival and development. Nature Reviews in Molecular and
Cellular Biology, 5: 305-315. 4. Pennell, R.I., and Lamb, C. (1997) Programmed Cell Death in Plants. Plant Cell 9: 1157-1168. 5. Jones, J.D., and Dangl, J.L. (2006) The plant immune system. Nature 444: 323-329. 6. Chisholm, S.T., Coaker, G., Day, B., and Staskawicz, B.J. (2006) Host-microbe interactions: shaping the
evolution of the plant immune response. Cell 124: 803-814. 7. Cande, C., Cecconi, F., Dessen, P., and Kroemer, G. (2002) Apoptosis-inducing factor (AIF): key to the
conserved caspase-independent pathways of cell death? Journal of Cell Science 115: 4727-4734. 8. Penninger, J.M., and Kroemer, G. (2003) Mitochondria, AIF and caspases--rivaling for cell death execution.
Nature Cell Biology 5: 97-99. 9. Schwab R, Ossowski S, Riester M, Warthmann N, Weigel D. (2006) Highly specific gene silencing by artificial
microRNAs in Arabidopsis. Plant Cell. 18(5):1121-1133. 10. http://signal.salk.edu/Help/iSectToolHelp.html 11. Weigel D. and Glazebrook J. (2002) Arabidopsis: A Laboratory Manual, Cold Spring Harbor Laboratory Press,
NY, p. 41-53