НАГЛЯДНАЯ БИОХИМИЯbinom-press.ru/books_1/biohim/naglyadnaya_biohim_frag.pdf ·...
TRANSCRIPT

Я. Кольман, К.Г. Рём
НАГЛЯДНАЯ БИОХИМИЯ

НАГЛЯДНАЯ БИОХИМИЯ

Jan Koolman, ScDProfessor
Philipps University MarburgInstitute of Physiological Chemistry
Marburg, Germany
Klaus-Heinrich Roehm, ScDProfessor (retired)
Philipps University MarburgInstitute of Physiological Chemistry
Marburg, Germany
Third edition, revised and updated
With 220 color plates prepared by Juergen Wirth
COLOR ATLAS OF BIOCHEMISTRY
ThiemeStuttgart • New York

МоскваЛаборатория знаний
Я. Кольман, К.�Г. Рём
НАГЛЯДНАЯ БИОХИМИЯ
Перевод с английского канд. хим. наук Т. П. Мосоловой
5�е издание,переработанное и дополненное

УДК 571.1ББК 28.072
К62
Кольман Я.
К62 Наглядная биохимия / Я. Кольман, К.-Г. Рём ; пер. с англ.Т. П. Мосоловой. — 5-е изд., перераб. и доп. — М. : Лабораториязнаний, 2018. — 509 с. : ил.
ISBN 978-5-906828-11-8
Существенно переработанное и дополненное справочное издание в на-глядной форме — в виде цветных схем — описывает все биохимическиепроцессы. Рассмотрены биохимически важные соединения, их строениеи свойства, основные процессы с их участием, а также механизмыи биохимия важнейших процессов в живой природе. Новое издание сделанос нового английского издания тех же немецких авторов.
Для студентов и преподавателей химических, биологических и ме-дицинских вузов, биохимиков, биологов, медиков, а также широкогокруга читателей, интересующихся процессами, происходящими в живоморганизме.
УДК 571.1
ББК 28.072
Справочное издание
Кольман Ян
Рём Клаус-Генрих
НАГЛЯДНАЯ БИОХИМИЯ
Ведущий редактор канд. биол. наук Н. Г. Иванова. Художник В. А. Прокудин
Технический редактор Т. Ю. Федорова. Корректор Е. В. Барановская
Компьютерная верстка: Т. Э. Внукова
Подписано в печать 18.10.17. Формат 60×90/16.
Усл. печ. л. 32,00. Заказ
Издательство «Лаборатория знаний»
125167, Москва, проезд Аэропорта, д. 3
Телефон: (499) 157-5272
e-mail: [email protected], http://www.pilotLZ.ru
ISBN 978-5-906828-11-8
c©Copyright 2013 of the original English languageedition by Georg Thieme Verlag KG,Stuttgart, Germany. Original title: “ColorAtlas of Biochemistry”, 3rd edition,by Jan Koolman, Klaus-Heinrich Roehm
c© Лаборатория знаний, 2018

Об авторах 5
Ян Кольман (слева) родился в Любеке (Герма-ния) и рос на обдуваемых ветрами берегах Бал-тийского моря. Полученное Кольманом класси-ческое образование отразилось на его мировоз-зрении. С 1963 по 1969 г. он изучал биохимию в университете Тюбингена. Диссертационную ра-боту, посвященную биохимическим процессам в организме насекомых и других беспозвоночных, Кольман выполнял в университете Марбурга под руководством Петера Карлсона. Защитил докторскую диссертацию по медицине в 1977 г. и получил должность профессора в 1984 г. В 2010 г. вышел на пенсию. Область интересов охватывает биохимическую эндокринологию и методы преподавания биохимии. Кольман же-нат, его жена — преподаватель живописи.
Клаус-Генрих Рём (справа) родился в Штут-гарте. Окончил Евангелическую теологическую семинарию в Урахе, где получил классическое образование, некоторое время посвятил заня-тиям физикой, а затем получил диплом биохи-мика в университете Тюбингена, где впервые встретился с Яном Кольманом. С 1970 г. также работал в Марбурге и выполнял диссертаци-онную работу под руководством Фридхельма
Шнейдера. Защитил докторскую диссертацию на Химическом факультете университета в 1980 г. С 1986 г. до выхода на пенсию в 2008 г. был профессором Медицинского факультета в Марбурге. Научные интересы Рёма лежат в об-ласти структуры и функций ферментов и соз-дания лекарственных препаратов. Его жена — биолог, у них двое детей.
Юрген Вирт (в центре) учился в Берлине и Оффенбахе (Германия), специализируясь на свободной графике и иллюстрациях. С 1963 по 1977 г. участвовал в оформлении экспозиции Франкфуртского музея естествознания Шенкен-берга и в это же время как внештатный сотруд-ник выполнял заказы нескольких издательств, иллюстрируя школьные учебники, научно-попу-лярные и научные книги. За оформление книг награжден несколькими дипломами и премия-ми. С 1978 г. Вирт преподавал в Художествен-ном училище в Швабиш-Гмюнде, а в 1986 г. по-лучил должность профессора информационного дизайна в Художественном училище Дармштад-та. Область интересов Вирта включает в себя научную и информационную графику и изобра-зительные методы.
ОБ АВТОРАХ

6 Предисловие
ПРЕДИСЛОВИЕ
Биохимия — динамичная, быстро развивающая-ся область знаний, что и демонстрирует данная книга. Очень сложно точно определить границы между биохимией и смежными науками, такими как клеточная биология, анатомия, физиология, генетика или фармакология. По этой причине данное наглядное пособие является централь-ным среди целой серии подобных книг, выпу-щенных издательством Thieme.Из-за недостатка места основное внимание в данной книге уделяется биохимии человека, хотя биохимия животных, растений и микроор-ганизмов не менее интересна. Обсуждаемые в книге вопросы в первую очередь интересны для студентов-медиков. Опять-таки из-за недостат-ка места невозможно отразить полностью весь объем имеющейся информации. Таким обра-зом, необходимо подчеркнуть, что данная книга не может заменить хороший и полный учебник по биохимии. Как и во всех других карманных справочниках, выпускаемых издательством Thieme, важнейшая роль в данной книге отводится цветным иллю-страциям. Авторы уделили большое внимание графическому представлению материала: текст следует рассматривать в качестве описания и дополнения к иллюстрациям. Как и в предыду-щих изданиях, для облегчения восприятия ис-пользуются цветные символы и коды, расшиф-
рованные на форзацах. Авторы старались стро-го придерживаться данной кодировки на всем протяжении книги.При подготовке к четвертому изданию книга была значительно переработана и расширена с целью более полного соответствия интересам студентов медицинских специальностей. Уда-лена часть материала из области естественных наук, но при этом расширен объем информа-ции, касающейся патологических нарушений биохимических процессов. При этом авторы постарались не изменить исходной концепции книги.Авторы выражают благодарность издательству Thieme, особенно Анжелике Финдготт, Анни Хол-линс и Софии Хенгст за помощь и поддержку. Мы также очень признательны многим нашим коллегам, соавторам и заинтересованным чита-телям за конструктивную критику, комментарии и рекомендации. Мы и в дальнейшем будем благодарны за любые комментарии и предло-жения, которые позволили бы улучшить книгу. Свои письма вы можете отправлять по адресу [email protected] или [email protected].
Ян Кольман,Клаус-Генрих Рём,
Юрген Вирт

Оглавление 7
ОГЛАВЛЕНИЕ
Основы биохимии I 11
Химия
Периодическая система элементов Д.И. Менделеева ............................................12Изомерия ........................................................14Классы соединений I ......................................16Классы соединений II......................................18Химические реакции .......................................20Окислительно-восстановительныепроцессы ........................................................22Кислоты и основания ......................................24
Физическая химия
Энергетика ......................................................26Термодинамика ...............................................28Катализ ...........................................................30Вода как растворитель ...................................32Гидрофобные взаимодействия .......................34
Биомолекулы I 37
Углеводы
Химия сахаров ................................................38Моносахариды и дисахариды .........................40Полисахариды .................................................42Гликопротеины и гликозаминогликаны ............44
Липиды
Общая информация ........................................46Жирные кислоты и жиры ................................48Глицеролипиды ...............................................50Сфинголипиды ................................................52Изопреноиды ..................................................54Стероиды ........................................................56
Аминокислоты
Аминокислоты: свойства .................................58Протеиногенные аминокислоты ......................60Селеноцистеини непротеиногенные аминокислоты ................62
Пептиды и белки
Пептиды и белки: общая информация............64Структура белков и пептидов .........................66Структурные белки .........................................68Растворимые белки ........................................70Модификация белков ......................................72
Нуклеотиды и нуклеиновые кислоты
Азотистые основания и нуклеотиды ...............74Рибонуклеиновые кислоты (РНК) ....................76Дезоксирибонуклеиновые кислоты (ДНК) .......78
Метаболизм I 81
Ферменты
Основные закономерности .............................82Ферментативный катализ ...............................84Ферментативная кинетика I ............................86Ферментативная кинетика II ...........................88Аллостерическая регуляция ............................90Ингибиторы .....................................................92Ферментативный анализ .................................94Коферменты I ..................................................96Коферменты II .................................................98Коферменты III ..............................................100Коферменты IV ..............................................102Патологические нарушения ..........................104
Метаболизм
Промежуточный метаболизм I ......................106Промежуточный метаболизм II .....................108Регуляция метаболизма I ..............................110Регуляция метаболизма II .............................112
Энергетический метаболизм
АТФ ...............................................................114Энергетическое сопряжение.........................116Сохранение энергии на мембранах ..............118Энергетический метаболизм ........................120Дегидрогеназы кетокислот ...........................122Реакции цикла трикарбоновых кислот ..........124Метаболические функции циклатрикарбоновых кислот ..................................126Митохондриальный транспорт ......................128Дыхательная цепь .........................................130Синтез АТФ ...................................................132Регуляция энергетическогометаболизма .................................................134Нарушения энергетическогометаболизма .................................................136
Метаболизм углеводов
Общие сведения ...........................................138Гликолиз ........................................................140

8 Оглавление
Пентозофосфатный путь ...............................142Глюконеогенез ..............................................144Метаболизм гликогена ..................................146Регуляция метаболизма углеводов I .............148Регуляция метаболизма углеводов II ............150Патологические нарушения ..........................152
Метаболизм липидов
Общие сведения ...........................................154Расщепление жирных кислот:
-окисление ..................................................156Другие пути расщепления жирных кислот ....158Биосинтез жирных кислот .............................160Метаболизм жирных кислот:другие реакции .............................................162Биосинтез сложных липидов ........................164Биосинтез холестерина ................................166Патологические нарушения ..........................168
Метаболизм белков
Общие сведения ...........................................170Протеолиз .....................................................172Метаболизм азота ........................................174Трансаминирование и дезаминирование ......176Расщепление аминокислот I .........................178Расщепление аминокислот II ........................180Цикл мочевины .............................................182Биосинтез аминокислот ................................184Патологические нарушения ..........................186
Метаболизм нуклеотидов
Общие сведения ...........................................188Расщепление нуклеотидов ............................190Биосинтез пуринов и пиримидинов ..............192Биосинтез нуклеотидов ................................194Патологические нарушения ..........................196
Метаболизм порфиринов
Биосинтез гема .............................................198Расщепление гема ........................................200
Клеточные органеллы I 203
Основные представления
Структура клетки ...........................................204Клеточные компоненты и цитоплазма ..........206
Цитоскелет
Компоненты ..................................................208Структура и функции ....................................210Двигательные белки .....................................212
Ядро
Ядро ..............................................................214
Митохондрии
Структура и функции ....................................216
Мембраны
Структура и компоненты мембран ................218
Транспортные процессы ...............................220Транспортные белки .....................................222Эндоцитоз и экзоцитоз .................................224
ЭПР и аппарат Гольджи
Структура и функции ....................................226Сортировка белков .......................................228Синтез белка в шероховатом эндоплазматическом ретикулуме .................230Созревание белка .........................................232
Лизосомы
Лизосомы......................................................234
Пероксисомы
Пероксисомы ................................................236
Молекулярная генетика I 239
Общие сведения
Общие сведения ...........................................240Геном
Гены и геномы ...............................................242Хроматин
Хроматин ......................................................244Ферменты, модифицирующиенуклеиновые кислоты ....................................246Репликация ...................................................248Транскрипция ................................................250Контроль транскрипции ................................252Созревание РНК ...........................................254
Генетический код
Генетический код ..........................................256Трансляция I: инициация...............................258Трансляция II .................................................260Антибиотики ..................................................262Мутации и репарация ...................................264
Генная инженерия
Клонирование ДНК .......................................266Секвенирование ДНК ....................................268Полимеразная цепная реакция (ПЦР) ...........270Генная инженерия в медицине ......................272
Ткани и органы I 275
Система пищеварения
Общая информация ......................................276Пищеварительные железы и их секреты ......278Процесс пищеварения ..................................280Всасывание I .................................................282Всасывание II ................................................284Патологические нарушения ..........................286
Кровь
Кровь: состав и функции ..............................288Белки плазмы крови .....................................290Липопротеины I .............................................292
Оглавление 9
Липопротеины II ............................................294Гемоглобин и транспорт газов ......................296Реактивные формы кислорода .....................298Эритроциты ..................................................300Кислотно-основной баланс ...........................302Свертывание крови .......................................304Ингибирование свертывания крови. Фибринолиз ..................................................306Группы крови ................................................308Патологические нарушения ..........................310
Иммунная система
Иммунная система ........................................312Специфический иммунный ответ ..................314Активация T-клеток .......................................316Система комплемента ..................................318Антитела .......................................................320Патологические нарушения ..........................322
Печень
Функции ........................................................324Метаболизм углеводов .................................326Метаболизм липидов ....................................328Желчные кислоты ..........................................330Биотрансформация .......................................332Система цитохрома Р450..............................334Метаболизм этилового спирта .....................336Нарушения функции печени..........................338
Жировая ткань
Функции ........................................................340Патологические нарушения ..........................342
Почки
Функции ........................................................344Возврат воды и электролитов .......................346Метаболизм ..................................................348
Мышцы
Сокращение мышц ........................................350Регуляция мышечных сокращений ................352Энергетический метаболизммышечной ткани ...........................................354Патологические нарушения ..........................356
Соединительная ткань
Кости и зубы .................................................358Коллагены .....................................................360Внеклеточный матрикс I................................362Внеклеточный матрикс II ...............................364Патологические нарушения ..........................366
Головной мозг и органы восприятия
Передача сигнала в центральнойнервной системе (ЦНС) ................................368Потенциал покоя и потенциал действия .......370Нейромедиаторы ..........................................372Рецепторы нейромедиаторов .......................374Метаболические процессыв головном мозге ..........................................376
Зрение ..........................................................378Патологические нарушения ..........................380
Интеграция метаболизма
Интеграция метаболизма I ...........................382Интеграция метаболизма II ...........................384Интеграция метаболизма III ..........................386Интеграция метаболизма IV ..........................388
Питание I 391
Питательные вещества
Органические соединения ............................392Минеральные веществаи следовые элементы ...................................394Метаболизм кальция.....................................396Метаболизм железа ......................................398Патологические нарушения ..........................400
Витамины
Витамины I ....................................................402Витамины II ...................................................404
Сигнальные системы I 407
Механизмы передачи сигнала
Передача сигнала .........................................408Мембранные рецепторы ...............................410Ионные каналы .............................................412ГТФ-связывающие белки ..............................414Вторичные посредники I ...............................416Вторичные посредники II ..............................418Протеинкиназы и фосфатазы .......................420Сигнальные каскады .....................................422
Гормоны
Общие сведения ...........................................424Содержание в плазме и иерархиягормонов .......................................................426
Липофильные сигнальные
вещества
Механизм действия ......................................428Кортикостероиды ..........................................430Половые стероидные гормоныи менструальный цикл ..................................432Метаболизм стероидных гормонов ..............434Гормоны щитовидной железы .......................436
Гидрофильные сигнальные
вещества
Инсулин ........................................................438Сахарный диабет ..........................................440Другие гормоны ............................................442Катехоламины ...............................................444Тканевые гормоны и медиаторы ...................446Эйкозаноиды .................................................448Цитокины ......................................................450

10 Оглавление
Рост и развитие I 453
Пролиферация клеток
Клеточный цикл I...........................................454Клеточный цикл II ..........................................456Апоптоз .........................................................458Онкогены ......................................................460Образование опухолей .................................462Цитостатические препараты .........................464Вирусы ..........................................................466
Приложения I 469
Часто используемыесокращения ...................................................470Физические величиныи единицы измерения ...................................471Источники иллюстраций ...............................473Дополнительная литература .........................474
Предметный указатель I 475

Основы биохимии

12 Химия
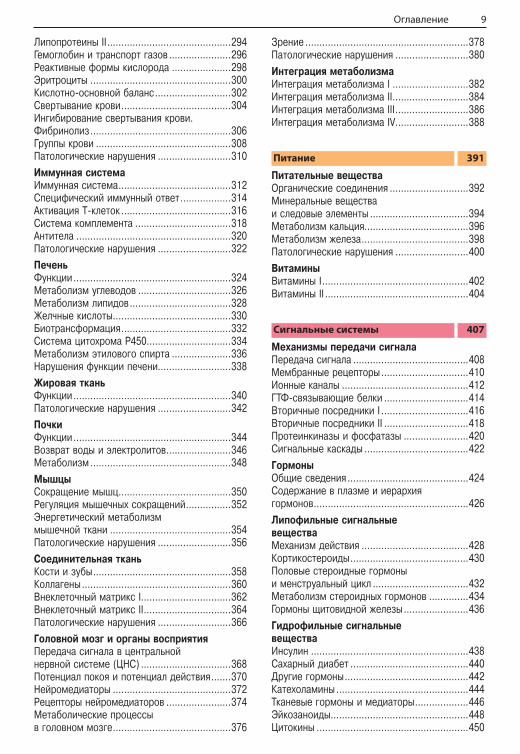
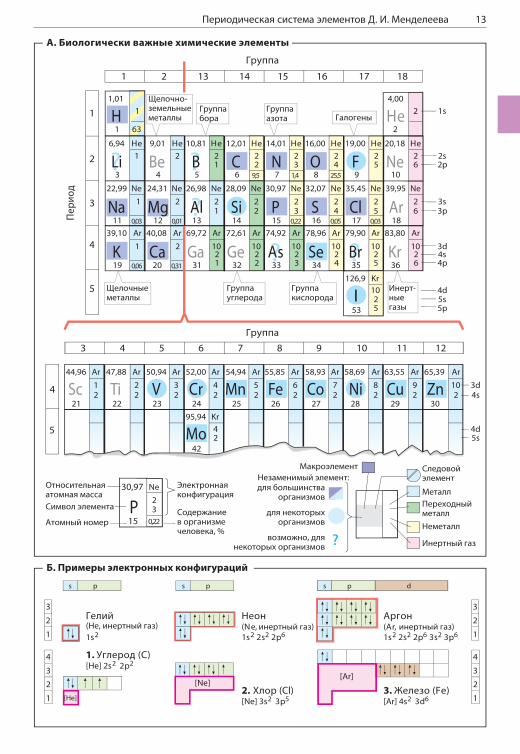
ПЕРИОДИЧЕСКАЯ СИСТЕМА ЭЛЕМЕНТОВ Д. И. МЕНДЕЛЕЕВА
А. Биологически важныехимические элементыВ природе встречается 81 стабильный химиче-ский элемент. 15 из них входят в состав всех живых существ, а еще 8–10 обнаружены лишь в некоторых организмах. На рисунке представ-лена первая половина Периодической таб-лицы, в которой и сосредоточены все важные для биологических систем элементы. Кроме фи-зических и химических характеристик (атомный номер, относительная атомная масса, название группы и электронная конфигурация), здесь приводится информация о распределении этих элементов в живых организмах и их содержании в организме человека.Тела животных на 99% состоят из четырех эле-ментов — водорода (H), кислорода (O), углерода (C) и азота (N). Водород и кислород входят в со-став воды, на долю которой приходится 60–70% от массы клетки (см. с. 206). Вместе с углеро-дом и азотом эти элементы являются основны-ми составляющими органических веществ, определяющих многие биологические процессы в организме. Во многих молекулах, кроме того, содержатся сера (S) и фосфор (P). Эти макро-элементы жизненно необходимы для всех жи-вых существ.Другая группа важных для биологических си-стем элементов, в сумме составляющих не бо-лее 0,5% от массы тела, представлена главным образом неорганическими ионами. Это так на-зываемые электролиты, среди которых щелоч-ные металлы натрий (Na) и калий (K) и щелоч-ноземельные металлы магний (Mg) и кальций (Ca). Галоген хлор (Cl) тоже всегда находится в клетке в ионизированном состоянии. Все дру-гие важные для жизни элементы присутствуют в организме в столь малом количестве, что их называют следовыми элементами (микроэле-ментами, с. 394). К этой группе относятся та-кие переходные металлы, как железо (Fe), цинк (Zn), медь (Cu), кобальт (Co) и марганец (Mn), а также некоторые неметаллы, такие как йод (I) и селен (Se).
Б. Примеры электронных конфигурацийХимические свойства элементов и типы связей, которые они образуют друг с другом, опреде-ляются строением их электронной оболочки. На рисунке А отражена электронная конфи-гурация некоторых элементов, а на рисунке Б расшифрованы использованные символы и со-
кращения. Более подробную информацию об электронных конфигурациях атомов можно по-черпнуть из учебников химии.Электроны в атомах могут занимать различные орбитали. Эти орбитали характеризуются глав-ным квантовым числом (1, 2, 3 и т. д.) и обо-значаются буквами s, p или d. По мере увели-чения числа электронов в атомах происходит постепенное заполнение орбиталей. На каждой орбитали может находиться не более двух элек-тронов, которые должны иметь противоположно направленные спины. На рисунке А отражено распределение электронов по орбиталям для всех представленных элементов. Например, шесть электронов углерода (Б1) занимают ор-битали 1s, 2s и две орбитали 2p. Заполненная орбиталь 1s имеет такую же электронную кон-фигурацию, как у инертного газа гелия (He). По-этому эта часть электронной оболочки углерода на рисунке А обозначена как «He». Ниже, спра-ва от символа элемента, указано количество электронов на каждой следующей заполненной орбитали (в случае углерода это орбитали 2s и 2p). Так, электронная оболочка хлора (Б2) состоит из электронной оболочки неона (Ne) и еще семи электронов на орбиталях 3s и 3p. В атоме железа (Б3) — переходного металла восьмой группы четвертого периода — электро-ны занимают орбиталь 4s, хотя орбитали 3d остаются частично свободными. Многие реак-ции переходных металлов, например окисли-тельно-восстановительные реакции или образо-вание комплексов с основаниями, происходят с участием незаполненных d-орбиталей.Наиболее устойчивое электронное состояние у атомов второго и третьего периодов достига-ется при наличии на их внешнем электронном уровне восьми электронов («правило октета»). Такое состояние реализуется, например, в ато-мах инертных газов, а также в таких ионах, как Cl– (3s23p6) и Na+ (2s22p6). Только в случае во-дорода и гелия устойчивая электронная конфи-гурация достигается при наличии на внешней орбитали 1s всего лишь двух электронов.
Периодическая система элементов Д. И. Менделеева 13
??
?
??
1s
2s2p
3s3p
3d4s4p
4d5s5p
3d4s
4d5s
4
5
44,96
Sc21
Ar
12
47,88
Ti22
Ar
22
50,94
V23
Ar
32
52,00
Cr24
Ar
42
54,94
Mn25
Ar
52
55,85
Fe26
Ar
62
58,93
Co27
Ar
72
58,69
Ni28
Ar
82
63,55
Cu29
Ar
92
65,39
Zn30
Ar
102
3 4 5 6 7 8 9 10 11 12
1,01
H1
1
63
4,00
He2
2
6,94
Li3
19,01
Be 2
4
10,81
B5
21
12,01
C6
He22
14,01
N7
He23
1,4
16,00
O8
He24
25,5
19,00
F9
He25
20,18 He
Ne10
26
HeHe He
22,99 Ne
Na 1
11 0,03
24,31
Mg12
Ne2
0,01
26,98 Ne
Al13
21
28,09
Si14
Ne22
30,97 Ne
P15
23
0,22
32,07
S16
Ne24
0,05
35,45
Cl17
Ne25
0,03
39,95
Ar18
26
39,10 Ar
K19
1
0,06
40,08 Ar
Ca20
2
0,31
69,72
Ga31
Ar
1021
72,61
Ge32
Ar
1022
74,92
As33
Ar
1023
78,96
Se34
Ar
10
79,90
BrAr
1025
83,80
Kr36
Ar
1026
126,9
I53
Kr1025
24 35
Ne
1
2
3
4
5
1 2 13 14 15 16 17 18
30,97
P15 0,22
?
Ne23
9,5
95,94
Mo42
Kr42
s p ds p s p
[Ne][Ar]
4
3
2
1
3
2
1
4
3
2
1
3
2
1
[He]
Щелочно-земельныеметаллы Галогены
Щелочныеметаллы
Инерт-ныегазы
Группа
Группа
Относительнаяатомная массаСимвол элемента
Атомный номер
Электроннаяконфигурация
Содержаниев организмечеловека, %
для большинстваорганизмов
Макроэлемент Следовойэлемент
МеталлПереходныйметаллНеметалл
Инертный газ
Пер
иод
возможно, длянекоторых организмов
для некоторыхорганизмов
Незаменимый элемент:
Группабора
Группаазота
Группауглерода
Группакислорода
А. Биологически важные химические элементы
Б. Примеры электронных конфигураций
Гелий(He, инертный газ)1s2
Неон(Ne, инертный газ)1s2 2s2 2p6
Аргон(Ar, инертный газ)1s2 2s2 2p6 3s2 3p6
1. Углерод (C)[He] 2s2 2p2
2. Хлор (Cl)[Ne] 3s2 3p5
3. Железо (Fe)[Ar] 4s2 3d6

14 Химия
ИЗОМЕРИЯ
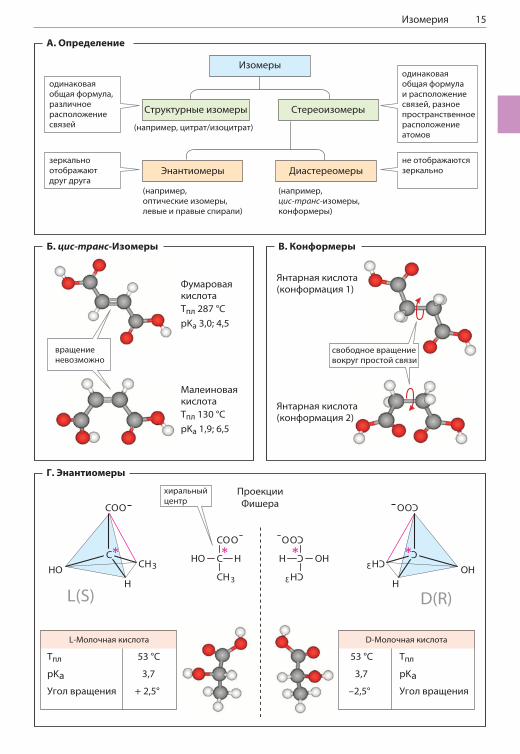
А. Определение
Изомерами называют молекулы, имеющие одинаковое количество атомов (и одинаковую молекулярную формулу), но различающиеся по структуре. Если изомеры различаются поряд-ком соединения атомов в молекуле, их назы-вают структурными изомерами. Примерами структурных изомеров могут служить лейцин и изолейцин (с. 60) или цитрат и изоцитрат (с. 124). Стереоизомерия может быть резуль-татом различного пространственного располо-жения заместителей относительно химической связи (Б, В) или наличия в молекуле хирального центра (Г). Если стереоизомеры различаются, как предмет и его зеркальное отражение, их на-зывают оптическими изомерами; все другие стереоизомеры называют диастереомерами.
Б. цис-транс-Изомеры
Вращение атомов вокруг двойной связи затруд-нено, поэтому у атомов, соединенных двойной связью, разные заместители могут распола-гаться в пространстве двумя способами. Так, в молекуле фумаровой кислоты (промежуточ-ное соединение в цикле трикарбоновых кислот, с. 124) карбоксильные группы располагаются по разные стороны от двойной связи (транс-, или E-положение). Изомер фумаровой кислоты, ма-леиновая кислота, не синтезируется в живых организмах. В этой молекуле карбоксильные группы находятся по одну сторону от двойной связи — это цис-, или Z-изомер. Цис-транс-изомеры (геометрические изомеры) различа-ются по химическим и физическим свойствам (температура плавления, Тпл, значение pKa). Превращение одного соединения в другое про-исходит только в ходе химической реакции.Особую роль цис-транс-изомерия играет в ме-таболизме жиров. Например, заместители при двойной связи в молекулах природных жирных кислот (с. 48) обычно находятся в цис-кон-фигурации, а ненасыщенные промежуточные соединения в процессе -окисления имеют транс-конфигурацию.
В. Конформеры
Формы молекул, образующиеся при свобод-ном вращении заместителей относительно про-стых связей (например, одинарных связей C–C),называют конформерами. В растворе даже небольшие молекулы могут иметь несколько конформаций. В представленных на рисунке конформерах янтарной кислоты атомы рас-
положены таким же образом, как в молекулах фумаровой и малеиновой кислот. В растворе су-ществуют обе формы молекулы наряду со мно-гими другими, однако конформация 1 (верхняя) энергетически более выгодна, что объясняется большим расстоянием между карбоксильными группами, и поэтому встречается чаще. Био-логически активные молекулы, такие как белки и нуклеиновые кислоты, содержат тысячи меж-атомных связей, допускающих свободное враще-ние заместителей, и поэтому теоретически мо-гут принимать множество конформаций. Однако обычно они встречаются в природе лишь в виде совершенно определенной (нативной) конфор-мации, стабилизированной внутримолекулярны-ми взаимодействиями (с. 70 и 78). Если нативная конформация макромолекулы нарушается в про-цессе денатурации, исчезает и биологическая активность молекулы.
Г. Энантиомеры
Еще один тип изомерии возникает в молекулах, имеющих хиральный центр, или в полностью хиральных молекулах. Хиральность (от греч. chier — рука) объясняет существование струк-тур, которые нельзя наложить друг на друга, поскольку они различаются, как предмет и его зеркальное отражение (зеркальные изомеры). Чаще всего причиной этого типа изомерии яв-ляется наличие в молекуле асимметрического атома углерода — атома углерода с четырьмя разными заместителями. Такие соединения су-ществуют в виде двух форм (энантиомеров) с различной конфигурацией. Часто энантиомер-ные формы называют L- и D-формами.Для однозначного обозначения конфигурации молекул используют R/S-номенклатуру (см. учебник химии). Для изображения структуры хирального центра применяют проекционные формулы, или проекции, Фишера (с. 58). Энан-тиомеры обладают очень похожими химически-ми свойствами, поэтому их достаточно сложно разделить химическим путем. Для разделения смесей таких соединений можно использовать их способность вращать плоскость поляризации плоскополяризованного света в противополож-ных направлениях (оптическая активность). Например, такой способностью обладают энан-тиомеры молочной кислоты. Правовращающая L-молочная кислота содержится в мышцах и крови животных, а левовращающая D-форма синтезируется микроорганизмами и содержит-ся, например, в молоке.
Изомерия 15
HOH
COO
CH3C
HOH
COO
CH3C
L(S) D(R)
COO
C
CH3
HO H
COO
C
CH3
HOH
не отображаютсязеркально
одинаковаяобщая формула,различноерасположениесвязей
(например,оптические изомеры,левые и правые спирали)
(например,цис-транс-изомеры,конформеры)
одинаковаяобщая формулаи расположениесвязей, разноепространственноерасположениеатомов
зеркальноотображаютдруг друга
В. Конформеры
Янтарная кислота(конформация 1)
Янтарная кислота(конформация 2)
cвободное вращениевокруг простой связи
А. Определение
D-Молочная кислота
53 °C
3,7
–2,5°
Тпл
pKa
Угол вращения
L-Молочная кислота
53 °C
3,7
+ 2,5°
Тпл
pKa
Угол вращения
ПроекцииФишера
хиральныйцентр
Изомеры
Структурные изомеры
(например, цитрат/изоцитрат)
Стереоизомеры
Энантиомеры
вращениеневозможно
Б. цис-транс-Изомеры
Г. Энантиомеры
Диастереомеры
ФумароваякислотаТпл 287 °СpKa 3,0; 4,5
МалеиноваякислотаТпл 130 °СpKa 1,9; 6,5

16 Химия
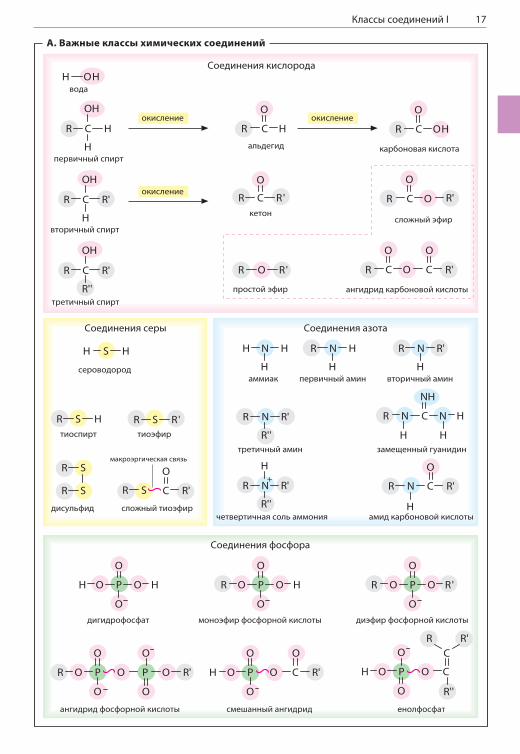
КЛАССЫ СОЕДИНЕНИЙ IА. Важные классы химических соединенийБольшинство биологических молекул — это производные простых соединений углерода (C), кислорода (O), водорода (H), азота (N), серы (S) и фосфора (P). Многие важные для биохимиче-ских процессов соединения кислорода, азота и серы можно рассматривать как производные их соединений с водородом (H2O, NH3, H2S). Фосфор в биологических системах практически всегда встречается в виде производных фос-форной кислоты (H3PO4).Если один или несколько атомов водорода в со-единениях водорода с неметаллами заменяется другой группой (например, алкильной), образуют-ся производные типа R—XHn-1, R—XHn-2—R и т. д. Таким образом, спирты (R—OH) и простые эфи-ры (R—O—R) являются производными воды (H2O), первичные (R—NH2), вторичные (R—NH—R) и
третичные амины — производными ам-
миака (NH3), а тиоспирты (R—SH) и тиоэфи-ры (R—S—R’) — производными сероводорода (H2S). Такие полярные группы, как —OH и —NH2, встречаются во многих органических со-единениях. Поскольку эти заместители гораздо активнее вступают в химические реакции, чем углеводородная основа молекулы, их называют функциональными группами.Новые функциональные группы получаются в результате окисления перечисленных выше классов соединений. Например, окисление тиоспиртов приводит к образованию дисуль-фидных связей (R—S—S—R). Окисление пер-вичных спиртов (R—CH2—OH) приводит снача-ла к образованию альдегидов (R—C(O)—H), а затем карбоновых кислот (R—C(O)—OH). При окислении вторичных спиртов образуются ке-тоны (R—C(O)—R). Альдегиды и кетоны содер-жат карбонильную группу .Присоединение имина (=NH) по карбонильной группе альдегида приводит к удалению моле-кулы воды и образованию альдимина (не по-казано). Альдимины являются промежуточными соединениями в реакциях метаболизма амино-кислот и служат, среди прочего, для связыва-ния альдегидов с аминогруппами белков (с. 72). Присоединение спирта по карбонильной группе альдегида приводит к образованию полуацета-ля (R—O—C(H)OH—R). Известные примеры по-луацеталей — циклические сахара (с. 38). При окислении полуацеталей образуются эфиры карбоновых кислот.Очень важную роль в биологических системах играют карбоновые кислоты и их производ-
ные, у которых в карбоксильной группе —OH заменяется на какую-либо другую группу. Та-кие производные карбоновых кислот образу-ются в результате нуклеофильного замещения в активированных промежуточных соединениях с отщеплением молекулы воды (с. 20). Эфиры карбоновых кислот (R—O—CO—R’) образуют-ся в ходе подобных реакций между карбоновы-ми кислотами и спиртами. В частности, к этой группе веществ относятся жиры (с. 48). Анало-гичным образом, при взаимодействии карбоно-вых кислот и тиоспиртов образуются тиоэфиры (R—S—CO—R’), которые играют чрезвычайно важную роль в метаболизме карбоновых кис-лот. Одно из наиболее известных соединений из этой группы — ацетил-кофермент А (с. 18). При взаимодействии карбоновых кислот и пер-вичных аминов получаются амиды карбоновых кислот (R—NH—CO—R’). Поскольку в пептидах и белках аминокислотные остатки связаны между собой амидной связью, связь такого типа назы-вают пептидной связью (с. 66).Фосфорная кислота H3PO4 является трехоснов-ной кислотой, т. е. содержит три гидроксильные группы, способные отдавать ионы водорода. В нормальных физиологических условиях как минимум одна из этих групп находится в дис-социированном состоянии, а две другие могут взаимодействовать со спиртами. В результате таких взаимодействий образуются монозаме-щенные и дизамещенные эфиры фосфорной кислоты (соответственно R—O—P[O]O—OH и R—O—P[O]O—O—R’). Моноэфиры фосфорной кислоты участвуют в метаболизме углеводов (с. 38), а фосфодиэфирные связи присутству-ют в фосфолипидах (с. 50) и нуклеиновых кис-лотах (с. 74). Соединения двух кислот называют ангидридами. Для образования ангидридной связи требуются большие затраты энергии. По этой причине фосфоангидридные связи играют чрезвычайно важную роль в запасании и исполь-зовании химической энергии в клетке (с. 114, 132). Смешанные ангидриды карбоновых кислот и фосфорной кислоты наряду с енолфосфатами относят к высокоэнергетическим (макроэрги-ческим) клеточным метаболитам.
Классы соединений I 17
O
PO
O
OH H
O
PO
O
OR P R'
O
O
O
O
PO
O
OR H
O
PO
O
OR R'
O
PO
O
OH C R'
O C
R''
O
PO
O
OH C
R'R
NR
H
H NR
H
R'NH
H
HSH H
SR H SR R'
SR
SR R'C
O
SR+
H
NR
R''
R'
NR
R''
R'
NH
CN
H
NR H
H
R'C
O
H
NR
H OH
OH
CR
R''
R'
OH
CR
H
R'
OH
CR
H
H
O
CR OH
R'
O
CR O
R'
O
CR O C
O
O
CR H
O
CR R'
OR R'
вода
моноэфир фосфорной кислоты
тиоэфир
первичный аминаммиак
третичный амин
вторичный амин
дигидрофосфат
ангидрид фосфорной кислоты смешанный ангидрид
тиоспирт
дисульфид
енолфосфат
диэфир фосфорной кислоты
сложный тиоэфирчетвертичная соль аммония амид карбоновой кислоты
замещенный гуанидин
сероводород
А. Важные классы химических соединений
Соединения серы Соединения азота
Соединения кислорода
Соединения фосфора
сложный эфир
карбоновая кислота
ангидрид карбоновой кислоты
окисление
окисление
окисление
макроэргическая связь
первичный спирт
вторичный спирт
третичный спирт
альдегид
кетон
простой эфир

18 Химия
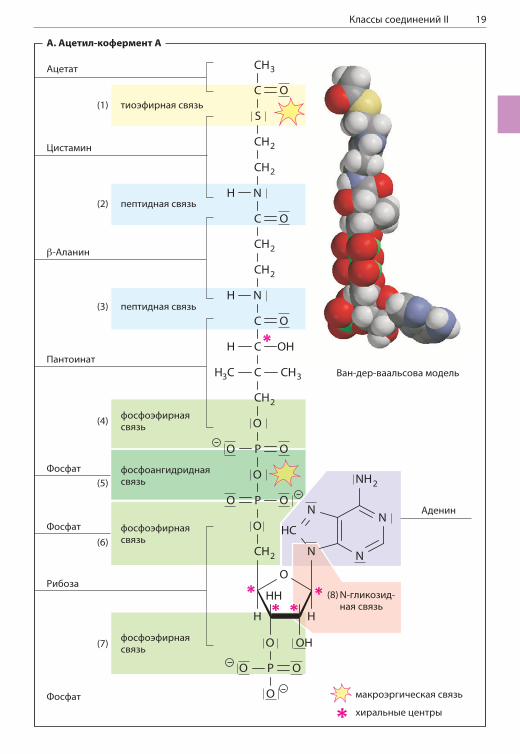
КЛАССЫ СОЕДИНЕНИЙ II
Многие биомолекулы имеют модульную струк-туру и построены из множества более мелких фрагментов, на которые их можно расщепить. Сборка биомолекул обычно происходит за счет реакций конденсации, протекающих с выделе-нием молекул воды. Напротив, расщепление таких молекул — гидролитический процесс, т. е. процесс, протекающий за счет присоединения молекулы воды. Модульное строение биомо-лекул проиллюстрировано на примере важного кофермента.
А. Ацетил-кофермент А
Кофермент А (коэнзим А, КоА, CoA; с. 98) — ну-клеотид с достаточно сложной структурой. Функ-ция этой молекулы заключается в активации ацильных групп (остатков карбоновых кислот). В результате связывания карбоксильной группы карбоновой кислоты с тиогруппой кофермен-та возникает тиоэфирная связь (—S—CO—R; с. 18), в которой ацильная группа имеет высокий химический потенциал. По этой причине эта группа может быть передана другим молекулам в ходе экзергонических реакций. Этот процесс имеет особенно большое значение для метабо-лизма жиров (с. 328) и для двух реакций цикла трикарбоновых кислот (с. 124).Как обсуждается на с. 28, потенциал пере-носа групп соответствует изменению энергии Гиббса ( G) при гидролитическом расщеплении молекулы. Это произвольное определение, но оно позволяет оценить химическую энергию расщепляемой связи. Отщепление ацетильной группы от молекулы ацетил-кофермента А мож-но записать в следующем виде:
Ацетил-кофермент А + H2O ацетат + кофермент А
В стандартных условиях и при pH 7 изменение хи-мического потенциала G ( G°; с. 28) в ходе данной реакции составляет –32 кДж/моль, что сравнимо с G° в реакции гидролиза АТФ (с. 114). Кроме богатой энергией тиоэфирной связи, ацетил-ко-фермент А содержит несколько других гидроли-зуемых связей с разной степенью устойчивости. Эти связи и образующиеся при их расщеплении фрагменты перечислены ниже.(1) Реакционноспособная тиольная группа в тио-
эфирной связи кофермента А происходит из цистамина. Этот биогенный амин (с. 62) образуется в результате декарбоксилирова-ния аминокислоты цистеина.
(2) Аминогруппа цистамина связана с карбо-ксильной группой другого биогенного ами-на, -аланина, через пептидную связь
(—CO—NH—). -Аланин образуется при де-карбоксилировании аминокислоты аспарта-та, а также при расщеплении пиримидино-вых оснований (с. 190).
(3) Еще одна амидная связь в молекуле КоА связывает следующий модуль молекулы — остаток пантоевой кислоты. Это соедине-ние содержит хиральный центр и поэтому может существовать в двух энантиомерных формах (с. 14). В природном КоА встреча-ется лишь одна из двух форм — (R)-пантои-нат. В организме человека это вещество не синтезируется, а поступает с пищей в виде витамина В5, пантотеновой кислоты, — соединения -аланина и пантоевой кислоты (с. 404).
(4) Гидроксильная группа при атоме C4 панто-ината связана с остатком фосфорной кис-лоты посредством сложноэфирной связи.
Описанная выше часть молекулы представ-ляет собой функциональное звено. В клетке эта часть молекулы КоА образуется из пан-тотеновой кислоты. Это звено также присут-ствует в связанной форме (в форме 4’-фос-фопантетеина) в молекуле фермента, назы-ваемого синтазой жирных кислот (с. 160). В молекуле КоА этот элемент связан с 3 ,5 -аденозиндифосфатом.
(5) При соединении двух остатков фосфорной кислоты образуется не эфирная, а макро-эргическая фосфоангидридная связь (как и в других нуклеозидфосфатах). А вот сле-дующие связи (6) и (7) вновь представляют собой сложноэфирные связи.
(8) Основание аденин связано с атомом C1 ри-бозы посредством N-гликозидной связи (с. 44, 74). Атом C1, как и атомы C2, C3 и C4 в остатке рибозы, тоже представляет собой хиральный центр (с. 14).
Классы соединений II 19
CH3
C
S
O
CH2
CH2
N
C
CH2
H
O
CH2
NH
C
C
O
H OH
C
CH2
CH3H3C
O
P
O
OO
P
O
OO
CH2
O
H
HH
O OH
H
N
NN
HC
N
NH2
P
O
OO
(8)
А. Ацетил-кофермент А
β-Аланин
N-гликозид-ная связь
(3)
(1)
(2)
(4)
(5)
(6)
(7)
Ацетат
Цистамин
Пантоинат
Фосфат
Фосфат
Рибоза
Фосфат
тиоэфирная связь
пептидная связь
пептидная связь
фосфоэфирнаясвязь
фосфоэфирнаясвязь
фосфоэфирнаясвязь
фосфоангидриднаясвязь
Ван-дер-ваальсова модель
Аденин
макроэргическая связь
хиральные центры

20 Химия
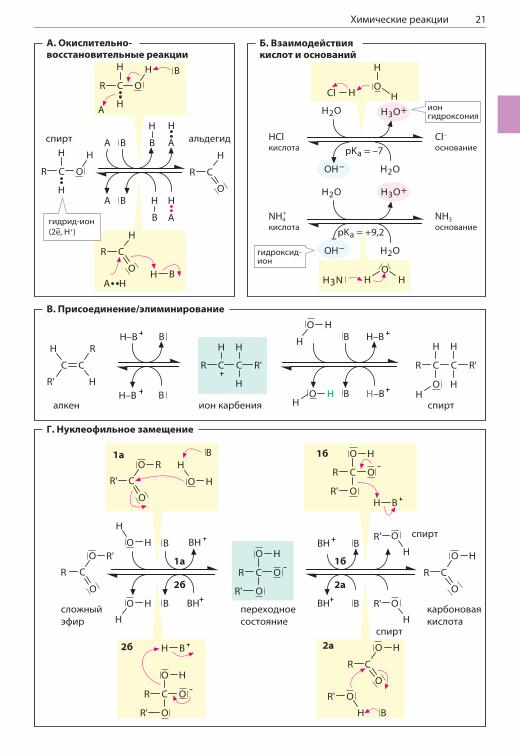
ХИМИЧЕСКИЕ РЕАКЦИИ
При химических реакциях электроны или группы атомов связываются молекулами, обменивают-ся между молекулами или перемещаются вну-три молекул. В этом смысл химических реакций. Ниже на простых примерах проиллюстрированы наиболее важные реакции органической химии. Красными стрелками обозначен процесс пере-дачи электронов.
А. Окислительно-восстановительные реакцииВ окислительно-восстановительных реакциях (с. 22) электроны передаются от одной мо-лекулы (восстановителя) к другой молекуле (окислителю). Процесс часто сопровождается передачей одного или двух протонов, однако определяющим критерием окислительно-восста-новительной реакции служит перенос электро-нов, когда восстановитель окисляется, а окисли-тель восстанавливается.На схеме А отражено окисление спирта до аль-дегида и восстановление альдегида до спирта. В данном случае происходит передача одного гидрид-иона (двух электронов и одного прото-на; см. с. 22) от молекулы спирта к окислите-лю А. Лишний протон связывается с основани-ем B, которое выступает в роли катализатора. В реакции восстановления альдегида соедине-ние A—H играет роль восстановителя, а H—B служит катализатором.
Б. Взаимодействия кислоти основанийВ отличие от окислительно-восстановительных реакций, при взаимодействии кислот и осно-ваний происходит только перенос протонов (ионов H+) без переноса электронов (с. 24). При диссоциации кислоты (здесь это соляная кисло-та HCl) вода выступает в роли акцептора про-тонов и превращается в ион гидроксония H3O+. В обратной реакции вода выполняет функцию кислоты и протонирует сопряженное основание Cl–. С водой взаимодействует основание NH3 (аммиак), а образуется гидроксид-ион (OH–) и сопряженная кислота — ион аммония (NH4
+).
В. Присоединение/элиминирование
Реакции, в которых атомы или молекулы при-соединяются к другим молекулам по кратным связям, называются реакциями присоедине-ния. Обратный процесс — удаление группы ато-мов с образованием кратной связи — называют элиминированием.
При присоединении воды к алкенам сначала происходит атака иона водорода по двойной связи алкена. Образуется нестабильный ион карбения, или карбкатион, который притягива-ет молекулу воды, а в результате отщепления протона образуется спирт. Обратная реакция дегидратации (отщепление молекулы воды от молекулы спирта) также катализируется кисло-той и протекает с образованием того же про-межуточного соединения.
Г. Нуклеофильное замещение
Реакция, в которой одна функциональная группа (с. 16) замещается на другую, называется реак-цией замещения. Такие реакции могут проте-кать по механизму нуклеофильного или электро-фильного замещения (см. учебник химии). При нуклеофильном замещении (SN2) происхо-дит присоединение одной молекулы к другой, а затем отщепление уходящей группы. Гидролиз сложного эфира с образованием спирта и кислоты и этерификация (взаимодей-ствие кислоты со спиртом) — примеры реакций, протекающих по механизму SN2. Протекание той и другой реакций возможно благодаря поляр-ности двойной связи C=O. При гидролизе эфира от молекулы воды протон отщепляется при уча-стии основания B. Образующийся ион OH– яв-ляется сильным нуклеофильным агентом и ата-кует положительно заряженный карбонильный углерод в молекуле эфира (1а), что приводит к возникновению неустойчивого переходного со-стояния с sp3-гибридизацией. Далее возможно либо удаление молекулы воды (2б) с образо-ванием эфира, либо удаление молекулы спирта (ROH) (1б) с образованием свободной кислоты. Реакция этерификации (2) протекает через те же стадии, но в обратном направлении.При перегруппировке, или изомеризации (на схеме не показано), группы атомов (функцио-нальных групп) переносятся внутри одной и той же молекулы. Из биохимических процессов к этим реакциям относятся изомеризация саха-рофосфатов (с. 138) и изомеризация метилма-лонил-КоА в сукцинил-КоА (с. 180).
Химические реакции 21
H3O+
OH–
H B
R C O
O
OR'
H
CR
O
R'B
B
HO
H
HO
H
BH
BH
R C O
O
OR'
H
B R' O
H
BH
BR' O
HBH
1б
2aR C
O
O H
H B
R C O
O
OR'
H
B
R C
O
O H
R' O
H
OB
C
H
R
H
HA
R C
O
H
OC
H
R
H
H
HO
HB1a 1б
1a
2б
2б 2a
C C
R'
H
H
R
R C C
H
R'
H
H
B
B
HO
H
R C C
H
R'
H
HOH
H–B
H–B
B
B
H
B
O
H3O+
H
BBA A
H
BA H
A
R C
O
H
H BHA
OH
H
Cl H
H2O
pKa = –7
H2O
OH– H2O
pKa = +9,2
H2O
OHHH3N
HO H–B
H–B
H
CR'
O
RO
Б. Взаимодействиякислот и оснований
А. Окислительно-восстановительные реакции
В. Присоединение/элиминирование
спирталкен ион карбения
спирт альдегид
гидрид-ион(2e–, H+)
NH3основание
Cl–
основание
Г. Нуклеофильное замещение
переходноесостояние
карбоноваякислота
спирт
сложныйэфир
спирт
NH4+
кислота
HClкислота
гидроксид-ион
ионгидроксония

22 Химия
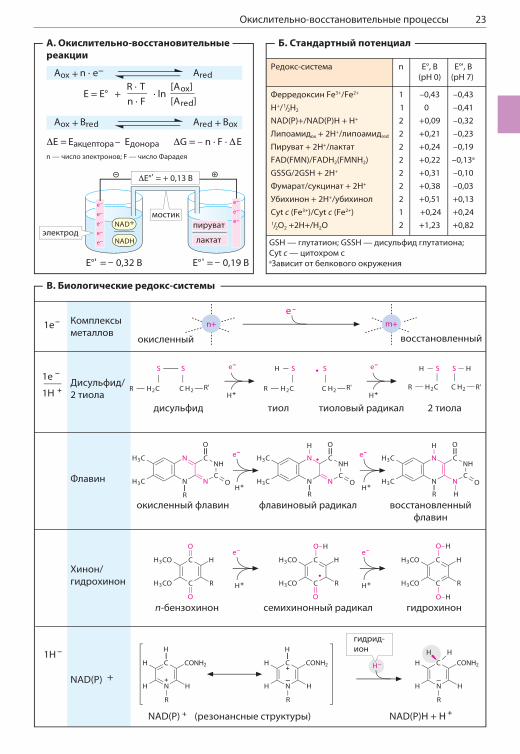
ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫЕ ПРОЦЕССЫ
А. Окислительно-восстановительные реакцииОкислительно-восстановительными реакция-ми, или редокс-реакциями, называют реакции, в которых происходит перенос электронов между реагирующими веществами (с. 20). Как и при реакциях между кислотой и основанием, редокс-реакции тоже всегда проходят между двумя соединениями, которые в данном слу-чае называются редокс-парой. Главное разли-чие между двумя компонентами редокс-пары (Aox и Ared) заключается в том, что они содер-жат разное число электронов. Компонент (Ared) у которого больше электронов, называется восстановленной формой соединения, а другой компонент (Aox) называется окислен-ной формой.Окислительно-восстановительный потен-циал (редокс-потенциал, E) системы — это способность компонентов этой системы при-нимать и отдавать электроны. Уравнение Нерн-ста связывает потенциал редокс-системы E со стандартным потенциалом E0 (постоянная, которая определяется экспериментально) ком-понентов и с концентрациями обоих компонен-тов системы. Редокс-потенциал измеряется в вольтах (В) и может быть положительным или отрицательным.В окислительно-восстановительных реакциях восстановленная форма одного соединения (восстановитель, Bred) передает электроны окисленной форме другого соединения (окис-литель, Aox). В результате восстановитель окисляется, а окислитель восстанавливается. Каждый восстановитель способен проявлять активность только в определенных окисли-тельно-восстановительных реакциях. Разность потенциалов между двумя редокс-системами можно определить с помощью гальваниче-ской ячейки. На рисунке это показано на при-мере следующей реакции:
пируват + NADH + H+ лактат + NAD+
В нормальных условиях система NAD+/NADH + H+ обладает более высоким отрицательным по-тенциалом. По этой причине обратная реакция (лактат + NAD+ пируват + NADH + H+) про-текать не может. В соответствии со значением восстановительных потенциалов все редокс-пары можно расположить в ряд. Спонтанный перенос электронов (например, в дыхательной цепи, с. 130) возможен только в том случае, если редокс-потенциал донора более отрицате-лен, чем потенциал акцептора.
Б. Стандартный потенциалВ таблице представлены значения стандартных потенциалов важных биологических систем. Ус-ловились, что для редокс-пары 2H+/H2 в стан-дартных условиях (когда концентрации всех ком-понентов системы, включая концентрацию H3O+, равны 1 моль/л) стандартный потенциал имеет нулевое значение, E0(2H+/H2) = 0. В биохимии обычно считают, что при рH 7 E0(2H+/H2) = 0.
В. Биологические редокс-системыБольшинство окислительно-восстановитель-ных процессов в клетке протекает при уча-стии ферментов и растворимых или связанных кофакторов.В некоторых кофакторах содержатся активные редокс-компоненты — ионы металлов. Реак-ция обычно сопровождается переносом одного электрона (e–), при этом металл изменяет свою степень окисления. В этих реакциях часто обра-зуются неспаренные электроны, но они локали-зованы на d-орбиталях (с. 12) и поэтому менее опасны, чем неспаренные электроны в атомах неметаллов («свободные радикалы», см. ниже).Среди других редокс-систем важное значение имеют дисульфиды (R—S—S—R) и соответству-ющие им тиоспирты, или тиолы (R—SH). Для восстановления дисульфида требуются 2e– и 2H+. Реакция протекает в две стадии с образованием очень активного тиорадикала в качестве промежу-точного соединения. В клетках существуют специ-альные защитные системы, предназначенные для обезвреживания свободных радикалов (с. 298).Для полного восстановления флавинов (FMN и FAD, с. 96) требуются 2e– и 2H+, промежуточный продукт реакции — флавиновый радикал.В окислительно-восстановительных реакциях хинонов и хинолов также образуются свобод-ные радикалы, например семихинонный ради-кал, но они менее реакционноспособны, чем флавиновые радикалы.Пиридиннуклеотиды NAD+ и NADP+ (с. 96) всегда действуют в растворимой форме. Окис-ленные коферменты содержат ароматическое никотинамидное кольцо, в котором делокали-зован положительный заряд. На схеме справа представлена резонансная структура, в которой обедненный электронами положительно за-ряженный атом углерода находится в пара-по-ложении к атому азота. Если к этой структуре присоединяется гидрид-ион H–, возникает вос-становленная форма NADH или NADPH без об-разования промежуточного радикала. Поскольку процесс сопровождается высвобождением иона H+, восстановленную форму пиридиннуклеотида правильно записывать как NAD(P)H + H+, а не NAD(P)H2.
Окислительно-восстановительные процессы 23
1e
1HC
N
CONH
H
HH 2
H
R
H
H–
e
C
N
CONH
H
H
H 2
H
R
C
N
CONH
H
H
H 2
H
R
AredAox
E = E° + · lnR · Tn · F [Ared]
[Aox]
e–
e–
e–
e–
e–
e–
e–
e–
NAD+
NADH
Ared + BoxAox + Bred
+ n · e–
N
N NC
NHCH3C
H3C
R
O
O
N
N NC
NHCH3C
H3C
R
O
O
H
H
N
N NC
NHCH3C
H3C
R
O
O
He
e e
H
e
H
C
C
H
R
O
H3CO
H3CO
O
C
C
H
R
O–H
H3CO
H3CO
O O–H
C
C
H
R
O–H
H3CO
H3CO
e– e–
H H
H H
1e
1H
m+n+
R'R H2CH2C
S S SH
R H2C R'H2C
S• SH
R H2C R'H2C
S H
2e–,1 H
•
•
А. Окислительно-восстановительныереакции
В. Биологические редокс-системы
Комплексыметаллов
окисленный восстановленный
Флавин
Хинон/гидрохинон
ΔG = – n · F · ΔEΔE = Eакцептора – Eдонора
NAD(P) +
п-бензохинон семихинонный радикал гидрохинон
дисульфид тиол тиоловый радикал 2 тиола
Дисульфид/2 тиола
Б. Стандартный потенциал
NAD(P) (резонансные структуры) NAD(P)H + H
гидрид-ион
восстановленныйфлавин
флавиновый радикалокисленный флавин
мостикпируват
лактатэлектрод
'
' ' E° = – 0,32 В E° = – 0,19 В
ΔE° = + 0,13 В
n — число электронов; F — число Фарадея
Редокс-система n E°, В E°', В (pH 0) (pH 7)
Ферредоксин Fe3+/Fe2+ 1 –0,43 –0,43H+/1/2H2 1 0 –0,41NAD(P)+/NAD(P)H + H+ 2 +0,09 –0,32Липоамидox + 2H+/липоамидred 2 +0,21 –0,23Пируват + 2H+/лактат 2 +0,24 –0,19FAD(FMN)/FADH2(FMNH2) 2 +0,22 –0,13а
GSSG/2GSH + 2H+ 2 +0,31 –0,10Фумарат/сукцинат + 2H+ 2 +0,38 –0,03Убихинон + 2H+/убихинол 2 +0,51 +0,13Cyt с (Fe3+)/Cyt с (Fe2+) 1 +0,24 +0,241/2O2 +2H+/H2O 2 +1,23 +0,82
GSH — глутатион; GSSH — дисульфид глутатиона;Cyt с — цитохром саЗависит от белкового окружения

24 Химия
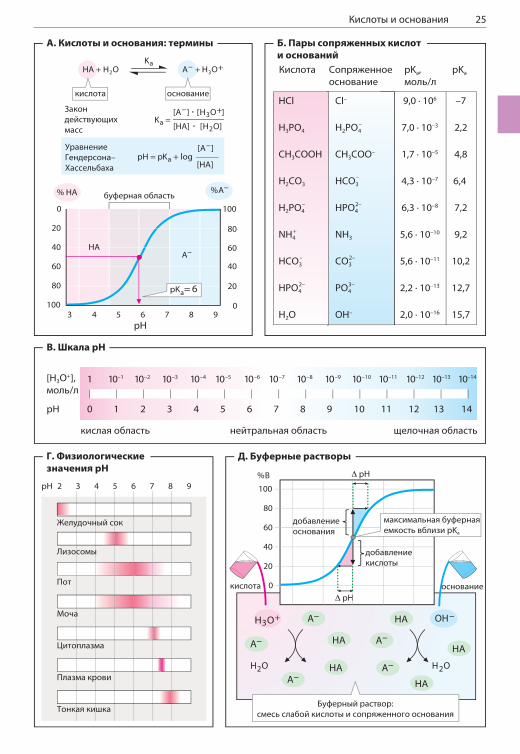
КИСЛОТЫ И ОСНОВАНИЯ
А. Кислоты и основания:термины
В соответствии с определением Брёнстеда кислотами называют вещества, способные от-давать ионы водорода (протоны), а основани-ями называют вещества, способные принимать протоны.Вода усиливает кислотно-основные свойства растворенных веществ, поскольку сама мо-жет выступать в роли кислоты или основания. В водных растворах кислота HA отдает протон растворителю. В результате образуется ани-он кислоты A– и протонированная молекула воды — ион гидроксония H3O+, который обыч-но упрощенно обозначают H+. В свою очередь, основания принимают ионы H+ от молекул воды, что приводит к образованию гидроксид-ионов OH– и протонированных оснований (на схеме не показано; с. 20). Если для реакции с участием кислот и осно-ваний записать закон действующих масс, это равновесие удобно характеризовать констан-той кислотной диссоциации Ka. Если вместо концентрации протонов и Ka использовать на-туральные логарифмы этих величин, получим уравнение Гендерсона–Хассельбаха. Это уравнение описывает уровень диссоциации кислоты в зависимости от pH. Если построить график соответствующей зависимости, можно получить кривую диссоциации кислоты (ниж-ний рисунок). pH в точке перегиба будет соот-ветствовать pKa системы.
Б. Пары сопряженных кислоти основанийВ результате реакции между кислотой и основа-нием всегда образуются сопряженные с ними соответственно основание и кислота. Силу кис-лоты характеризует ее константа кислотной дис-социации Ka. Чем меньше pKa, тем сильнее кис-лота. Сильным кислотам соответствуют слабые сопряженные основания, и наоборот. Например, очень сильной соляной кислоте соответствует слабое основание хлорид-ион, а слабой кислоте H2O — очень сильное основание OH–.
В. Шкала pH
Для воды pKa равно 15,7 (Б), а произведение [H+]•[OH–] называется ионным произведени-ем воды. [H+]•[OH–] = 10–14; это величина по-стоянная, не изменяется даже при растворении в воде других кислот или оснований. Это озна-чает, что в чистой воде при 25 °C концентрации
ионов H+ и OH– равны. [H+] = [OH–] = 10–7 моль/л. Чистая вода имеет нейтральную кислотность pH 7. Водные растворы с более высоким содер-жанием ионов H+ (0 < pH < 7) кислые, а с более низким (7 < pH < 14) — щелочные.
Г. Физиологические значения pH
Значения pH в клетках и во внеклеточной жид-кости варьируют лишь в узких пределах. В нор-ме pH крови может изменяться от 7,35 до 7,45 (с. 302), при этом содержание протонов в крови изменяется не более чем на 30%. В цитоплазме pH чуть ниже, чем pH крови (примерно 7,0–7,3). В лизосомах (pH 4,5–5,5; с. 234) концентрация протонов в сотни раз выше, чем в цитоплазме. Экстремальные значения кислотности характер-ны для желудка (pH 2) и тонкой кишки (pH > 8). Поскольку почки способны выводить как кис-лоты, так и основания (с. 346), pH мочи может изменяться в широком интервале значений (от 4,8 до 7,5).
Д. Буферные растворы
Краткосрочные изменения кислотности в орга-низме сглаживаются благодаря действию бу-ферных систем (с. 302). Буферный раствор представляет собой смесь слабой кислоты (HA) и сопряженного с ней основания (A–) или сла-бого основания с сопряженной кислотой. В бу-ферной системе нейтрализуется влияние как протонов, так и гидроксид-ионов.В первом случае (слева на схеме) основание A– связывает значительное количество до-бавленных протонов H+, в результате чего об-разуются кислота HA и вода. Если в буферный раствор добавляют гидроксид-ионы (OH–), они взаимодействуют с HA и образуют основание A– и воду (справа). В обоих случаях отношение [HA]/[A–] изменяется, тогда как pH изменяется очень мало. Как показывает кривая диссоциа-ции (вверху), буферные системы наиболее эф-фективно действуют в диапазоне pH, близком к pKa кислоты. Именно в этой области кривая поднимается наиболее резко, что соответствует наименьшему изменению pH ( pH) при соответ-ствующем изменении концентрации ( c) прото-нов или гидроксид-ионов. Другими словами, буферная емкость c/ pH данной системы максимальна вблизи значения pKa.
Кислоты и основания 25
Δ pH
pH 2 3 4 5 6 7 8 9
H2O H2O
pH3 4 5 6 7 8 9
100
80
60
40
20
0
20
40
60
80
100
0
% HA % A–
HA + H2O A–Ka
pH = pKa + log[HA]
[A–]
Ka =[A–] · [H3O+]
[HA] · [H2O]
[H3O+],моль/л
10–1 –210 –310 –410 –510 –610 –710 –810 –910 –1010 –1110 –1210 –1310 –14101
pH 1 2 3 4 5 6 7 8 9 10 11 12 13 140
A– HA OH–H3O+
A–
A–
A–HA
HA A–
HA
HA
pKa= 6
HAA–
+ H3O+
Δ pH% B
100
80
60
40
20
0
Д. Буферные растворыГ. Физиологическиезначения pH
А. Кислоты и основания: термины
В. Шкала pH
Б. Пары сопряженных кислоти оснований
буферная область
Кислота Сопряженноеоснование
рKa,моль/л
рKa
Буферный раствор:смесь слабой кислоты и сопряженного основания
добавлениеоснования
максимальная буфернаяемкость вблизи pKa
добавлениекислоты
Желудочный сок
Лизосомы
Пот
Моча
Цитоплазма
Плазма крови
Тонкая кишка
Закондействующихмасс
УравнениеГендерсона–Хассельбаха
кислая область нейтральная область щелочная область
кислота
кислота основание
основание
HCl Cl– 9,0 · 106 –7
H3PO4 H2PO4– 7,0 · 10–3 2,2
CH3COOH CH3COO– 1,7 · 10–5 4,8
H2CO3 HCO3– 4,3 · 10–7 6,4
H2PO4– HPO4
2– 6,3 · 10–8 7,2
NH4+ NH3 5,6 · 10–10 9,2
HCO3– CO3
2– 5,6 · 10–11 10,2
HPO42– PO4
3– 2,2 · 10–13 12,7
H2O OH– 2,0 · 10–16 15,7
[ . . . ]
ATP
КоА
NAD
ТДФ ПЛФ
FMN
ТГФ
FAD
ADP
P
dTMP
NADP
AMP
1
1
1
H
C
N
37
77
70
110
S 104
P
O 66
Символ Ван-дер-в.радиус,
пм
Отн.мол.
масса
Ковал.радиус,
пм
Водород
Углерод
Азот
Кислород
Фосфор
Сера
Металлы Me
100
170
150
190
180
140
1,008
12,011
14,007
30,974
32,060
15,999мембрана
углевод сигнальнаямолекула
внешняяповерхность
внутренняяповерхность
белок
ДНКРНК
Б. БиомолекулыА. Важные элементы
запасное вещество
В. Процессы, реакции, метаболиты
Г. Коферменты и нуклеотиды (примеры)
другие коферменты
нуклеотиды
окислительно-восстановительныекоферменты
комплексноедействие
ингибирование
транспорт
катализ
многостадийныйпроцесс
реакция
вторичный посредник
фосфат илифосфатная группа
макроэргическая связь возможная патология
хиральный центр
сигнальные вещества
активацияметаболический
процесс
общий процесс
метаболит
углевод
нуклеотид
липид
белок/аминокислота
патологический процесс
процесс не идет,молекула неактивна
процесс идет,молекула активна
незаменим для человека
транспортный процесс
метаболический процесс
другой процесс
уменьшение концентрации
рост концентрации
катаболический путь
анаболический путь
1
NADH
11
1
эффектор
действие
кофермент
А. Схема ферментативной реакции
фермент
Классы ферментов:
1 Оксидоредуктазы2 Трансферазы3 Гидролазы4 Лиазы5 Изомеразы6 Лигазы
субстрат
этиловый спирт
продукт
ацетальдегид
ферментномер на схеме кофактор
метиловыйспирт
протеинкиназа
протеинфосфатаза
[Zn2+]1 Алкогольдегидрогеназа
ядерная клетка
аппарат Гольджи
митохондрия
эритроцит
легкие
кишечник
жировая ткань
почки
мышцы
печень
рибосома
транскрипционныйфактор
антитело
мембрана
переносчик
ионный канал
транспортная АТФаза
односпиральныймембранный рецептор
семиспиральный мембранный рецептор
В. Клеточные структурыи сложные молекулы
Б. Ткани, органы, органеллы
NAD
