1 miniuib, a novel minitransposon-based system for stably inserting
TRANSCRIPT

MiniUIB, a Novel Minitransposon-Based System for Stable Insertionof Foreign DNA into the Genomes of Gram-Negative and Gram-Positive Bacteria
Joseph Alexander Christie-Oleza,a* Isabel Brunet-Galmés,a Jorge Lalucat,a,b Balbina Nogales,a Rafael Boscha
Microbiologia, Departament de Biologia, Universitat de les Illes Balears (UIB),a and Institut Mediterrani d’Estudis Avançats (IMEDEA, CSIC-UIB),b Palma de Mallorca, Spain
Transposition of the insertion sequence (IS) ISPpu12 is actively induced after conjugative interaction. The transposase ofthis IS can act in trans on structures flanked by inverted repeats similar to those of the transposon. Based on that fact, anISPpu12-based minitransposon, miniUIB, has been constructed in order to biotechnologically exploit the self-regulationof ISPpu12 and its increased activity after conjugative interaction. Mobilization of the miniUIB structure into the genomeof Pseudomonas stutzeri AN10 after conjugative interaction was demonstrated. A single gene, i.e., the kanamycin resis-tance determinant, or large genetic structures of >12 kb, i.e., alkBFGHJKL and alkST operons of Pseudomonas putidaTF4-1L (GPo1), have been easily integrated in P. stutzeri AN10 by an RP4-based delivery system. Therefore, the integrationof the alk determinants by use of the miniUIB system has extended the biodegradation capabilities of this strain. PlasmidpJOC100, containing the transposase and regulator genes of ISPpu12 adjacent to the miniUIB structure, was constructedin order to extend the host range of this biotechnologically useful genetic tool to other model and real-world bacteria. Theeffectiveness of the system for random mutagenesis in a phylogenetic wide range of bacteria and for the insertion of novelfunctions has been demonstrated, even in successive steps.
Minitransposon-based insertion systems have proven use-ful for inserting DNA fragments into a host chromosome.
These biotechnological tools consist in nucleic acid sequencesflanked by the inverted repeats (IR) of a transposon. These IRsare recognized by the transposase, an enzyme encoded outsideof the mobile structure, mobilizing the IR-IR structure to an-other DNA region or replicon. Since the transposase gene is notmobilized with the minitransposon, the outcome is a stableinsertion.
As revealed by literature, the developed minitransposon sys-tems mini-Tn5 and mini-Tn10 (1, 2) are by far the most popularlyused mainly for random transposon-mutagenesis (up to 975 citesaccording to the ISI Web of Knowledge at the end of September2012). Their biotechnological use lies on the simplicity of theirgenetic structure together with the R6K-based suicidal plasmid (3)in RP4 conjugation delivery-based Escherichia coli �pir strains (2,4). Since their description, several interesting innovations havebeen applied to these systems, such as the addition of novel resis-tance determinants (5), the insertion of an R6K replicase for rapididentification of the flanking region of insertion (6), or the intro-duction of the sacB marker to force the loss of antibiotic-resistancedeterminants by homologous recombination (7). However, theinstability of these minitransposon structures in the absence of aninsert between the IRs requires the use of intermediate plasmids inorder to perform the constructions (2). Also, in some recipientstrains, transposition frequencies of these systems are as low as thefrequency of cointegration of the whole vector by homologousrecombination (2), not allowing to easily discern true transposi-tion events.
An alternative to the random insertion of mini-Tn5 are themini-Tn7 minitransposons, which specifically integrates at theattTn7 site, located downstream the highly conserved glmS gene(8). It has been proven that the Tn7-based system is a useful tool tointegrate DNA sequences into the chromosomes of different
Gram-negative bacteria (9). In this sense it might be a betterchoice than random strategies (i.e., miniTn5) because the inser-tion site is known and, consequently, it is not necessary to evaluatethe fitness of the recombinant (9). However, the absence of thetransposition target, or even the presence of a badly conservedtransposition target, could complicate obtaining recombinants. Inaddition, it is well known that Tn7 displays transposition immu-nity (i.e., Tn7 does not integrate into DNA molecules already har-boring Tn7) (10). This phenomenon may complicate the use ofminiTn7-derivatives in bacteria harboring Tn7 and may prevent asecond integration event with the same Tn7-based system, if nec-essary.
ISPpu12 is an ISL3-like IS that was first described in the TOLplasmid pWW0 of Pseudomonas putida mt-2 (11). In a recentstudy, we showed that ISPpu12 was also present in P. stutzeri AN10(12), in addition to another ISL3-like IS named ISPst9 with similarflanking IRs. Both ISL3-like ISs transpose use a cut-and-pastetransposition mechanism by means of circular intermediates (12,13). In addition, we showed that ISPpu12 was responsible of thetransposition activation of these two ISL3-like ISs that occur in P.stutzeri AN10 cells that undertake conjugative interaction (inter-action with a conjugative strain such as E. coli S17-1, probably
Received 18 October 2012 Accepted 21 December 2012
Published ahead of print 28 December 2012
Address correspondence to Rafael Bosch, [email protected].
* Present address: Joseph Alexander Christie-Oleza, School of Life Sciences,University of Warwick, Coventry, United Kingdom.
Supplemental material for this article may be found at http://dx.doi.org/10.1128/AEM.03214-12.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AEM.03214-12
March 2013 Volume 79 Number 5 Applied and Environmental Microbiology p. 1629–1638 aem.asm.org 1629
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 20
0.12
6.83
.173
.

mediated by the conjugative pili, with or without horizontal genetransfer) (13). Thus, we demonstrated that activation of transpo-sition was due to TnpR, the transcriptional regulator encodedbetween the IRs of ISPpu12, and that the in trans activity ofISPpu12 transposase on ISPst9 was responsible for ISPst9 transpo-sition in response to that stimulus (12). In fact, ISPst9 was not ableto activate its own transposition after conjugative interaction be-cause it lacks the regulatory tnpR gene (12).
We describe here the construction of the miniUIB minitrans-poson and a set of miniUIB-based tools designed to exploit bio-technologically the self-regulation of ISPpu12 and its increasedactivity after conjugative interaction. This reliable and ready-to-
use system has proven effective for inserting genetic sequences inrecipients compatible with the RP4 conjugation process.
MATERIALS AND METHODSBacterial strains, media, and culture conditions. P. stutzeri strains AN10(Aps Cms Kms Sms nah� Alk� ISPpu12�) and AN11 (Kms nah� Alk�
ISPpu12-free) (14), Klebsiella pneumoniae CMD1 (15), and P. putida IS�,an ISPpu12-free derivative of P. putida mt-2 (16) obtained in our labora-tory, were used as recipients of the minitransposons used in the presentstudy. A total of 50 phylogenetically distinct environmental isolates fromour collection of 292 previously isolated strains (17) from polluted beach-sand samples after the Prestige oil spill (Galicia, Spain) were also used. E.coli S17-1�pir (2) was used for hosting pGP704 plasmid (4) and its deriv-
TABLE 1 Plasmids, primers, and probes used in this study
Plasmid, primer, orprobe Description or sequence (5=–3=)
Source orreference
PlasmidspBBR1MCS-1 Cmr; broad-host-range cloning vector 18pBC SK(�) Cmr; ori pBBR322 StratagenepCSI1 Apr Cmr; source of the Cmr gene used in this study 19pCSI2 Apr Kmr; source of the Kmr gene used in this study 19pGP704 Apr; ori R6K, mob RP4 4pGP704Km Apr Kmr; pGP704-derivative containing the Kmr gene at the EcoRI site of its MCS This studypKNG101 Smr; ori R6K, mob RK2 20pJOC21 Apr; pGP704-derivative containing miniUIB (IRL-MCS-IRR of ISPst9) This studypJOC22Km Apr Kmr; pJOC21-derivative containing miniUIB-Km (1.6 kb, IRL-Kmr-IRR) This studypJOC22Cm Apr Cmr; pJOC21-derivative containing miniUIB-Cm (2.1 kb, IRL-Cmr-IRR) This studypJOC22BC Apr Cmr; pJOC21-derivative containing miniUIB-BC (3.6 kb, IRL-pBC SK-IRR) This studypJOC22KmBC Apr Cmr Kmr; pJOC21-derivative containing miniUIB-KmBC (5.0 kb, IRL-Kmr-pBC SK-IRR) This studypJOC22KNG Apr Smr; pJOC21-derivative containing miniUIB-KNG (7.0 kb, IRL-pKNG101-IRR) This studypJOC22KmKNG Apr Kmr Smr; pJOC21-derivative containing miniUIB-KmKNG (8.4 kb, IRL-Kmr-pKNG101-IRR) This studypJOC22BCKmKNG Apr Kmr Smr; pJOC21-derivative containing miniUIB-BCKmKNG (11.8 kb, IRL-pBC
SK-Kmr-pKNG101-IRR)This study
pALKB Apr; pJOC21-derivative containing miniUIB-ALKB (8.5 kb, IRL-alkBFGHJKL-IRR) This studypALK Apr; pJOC21-derivative containing miniUIB-ALK (12.8 kb, IRL-alkST-alkBFGHJKL-IRR) This studypJOC100 Apr; pJOC21-derivative containing miniUIB100 (IRL-MCS-IRR-ISPpu12 without IRs) This studypJOC100Km Apr Kmr; pJOC100-derivative containing miniUIB100-Km (IRL-Kmr-IRR-ISPpu12 without IRs) This studypJOC100ALK Apr; pJOC100-derivative containing miniUIB100-ALK (IRL-alkST-alkBFGHJKL-IRR-ISPpu12 without IRs) This study
PrimersKM-F AAA CGT CTT GCT CGA GGC C This studyKM-R GGA GAA AAC TCA CCG AGG C This studyPCR3-F GAG ATC TTC GGG TAT GCG GAT TTA ATG This studyPCR3-R GAT CTA GAC CCG GGC TAT TGT CAA GAC AG This studyPCR1-F CAG GTA CCG CAT GAC CGA AAT GCC CGA This studyPCR1-R GGA ATT CGT GGG TAT GCG GAT TTA ATG G This studyALK1-F AAT CTA GAT TTC CAG CAG ACG ACG GAG C This studyALK1-R CGC GCC GAG CTC CAG CGT TGT CC This studyALK2-F GGC GAG TAC CAG GAC GGC GTA GGG This studyALK2-R TTG AGC TCT TAG AAA ACA TAT GAC GCA CC This studyALKRsma-F AAC CCG GGG CAC GTA CGG AGT GCG GG This studyALKRsma-R AAC CCG GGG GCG AAG GCC GAA GTC GGC This studyISPPU25Eco-F TTG AAT TCT TAC CTT TGC ATG AGA GTG AG This studyISPPU3348Eco-R TTG AAT TCG GGG CAC CTT CAC CCC ATC This study
ProbesTNPA tnpA of ISPst9 and ISPpu12 13TNPR tnpR of ISPpu12 12KM Kmr gene of pCSI2, PCR using KM-F and KM-R primers This studyALK alkBFGH genes of P. putida TF4-1L (GPo1), PCR using ALK1-F and ALK1-R primers This studyPGP Entire EcoRI-digested pGP704 plasmid This study
a Cmr, chloramphenicol resistance; Apr, ampicillin resistance; Smr, streptomycin resistance; Kmr, kanamycin resistance.
Christie-Oleza et al.
1630 aem.asm.org Applied and Environmental Microbiology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 20
0.12
6.83
.173
.

atives (Table 1) and for RP4-based conjugations. Conjugations were car-ried out as described previously (13). Briefly, aliquots of late-exponential-phase cultures of donor and recipient strains were spotted together (cellratio of 1:1) onto a membrane filter (nitrocellulose, 0.22 �m; Millipore)and incubated at 30°C for 7 h (or 30 min, when indicated in the text) onthe surface on a Luria-Bertani (LB) agar plate. P. putida TF4-1L (GPo1)(21) was used as source of alkane degradation determinants (alkBFGHJKLand alkST). When not specified, strains were cultured on LB broth at30°C. Mineral basal medium (MBM) (22) supplemented with succinate0.5% (wt/vol) and the appropriate antibiotics (ampicillin [Ap], 100 �g/ml; kanamycin [Km], 50 �g/ml; chloramphenicol [Cm], 40 �g/ml;and/or streptomycin [Sm], 50 �g/ml) were used for transconjugant selec-tion. MBM supplemented with n-octane and n-hexadecane, at 2% (wt/vol) each, was used for growing alk� strains.
DNA manipulations. All DNA manipulations were done as previ-ously described (12, 13). Restriction endonuclease digestions (Roche andGE Healthcare) and ligations with T4 DNA ligase (Invitrogen) were per-formed as recommended by each manufacturer. PCR amplifications weredone using Taq DNA polymerase (GE Healthcare). Plasmid DNA wasisolated by alkaline lysis using the QIAprep spin miniprep kit (Qiagen).Southern blot hybridization against EcoRI-digested genomic DNA wasperformed as described previously (23). Enhanced chemiluminescencedirect labeling (ECL direct nucleic acid labeling and detection system; GEHealthcare) was used for hybridization.
Primers and probes used in the present study are listed in Table 1.Probes, their targets, and the procedure for their synthesis were as follows:TNPA, for the detection of tnpA genes of both ISPst9 and ISPpu12 ISs,obtained as previously described (13); TNPR, for the detection of tnpRgene of ISPpu12, obtained as previously described (12); KM, for the de-tection of Kmr-gene, obtained by PCR amplification using primers KM-Fand KM-R on plasmid pCSI2 (19); ALK, for alkBFGH detection, obtainedby PCR amplification using primers ALK1-F and ALK1-R using genomicDNA obtained from P. putida TF4-1L (GPo1); and PGP, for pGP704detection, obtained by direct DNA labeling of EcoRI-digested plasmid.
Plasmids and miniUIB-derivatives constructed in this work. Plas-mids used in the present study are listed in Table 1. Plasmid pJOC21,carrying the basic miniUIB genetic structure (Fig. 1A), was obtained afterinserting the two IRs (IRL and IRR) of ISPst9 into pGP704. IRR was firstamplified by PCR using primers PCR1F and PCR1R from P. stutzeri AN10genomic DNA. The resulting PCR product was further digested with SacI andEcoRI and cloned between SacI and EcoRI restriction sites in pGP704 plas-mid. In a second step, IRL was obtained by PCR amplification using primersPCR3F and PCR3R and cloned at the BglII and XbaI restriction sites.
Diverse genetic material was cloned between the IRs of pJOC21 givingthe miniUIB derivatives represented in Fig. 1. Plasmids pJOC22Km, har-boring miniUIB-Km (Fig. 1B), and pJOC22Cm, containing miniUIB-Cm(Fig. 1C), were obtained by cloning the Kmr and the Cmr determinants ofpCSI2 and pCSI1 plasmids (19), respectively, into the XbaI (Kmr) orEcoRV (Cmr) restriction sites of pJOC21. Plasmid pJOC22BC, harboringminiUIB-BC (Fig. 1D), was obtained by cloning the SmaI-linearized pBCSK(�) cloning vector (Cmr; Stratagene) into the SmaI restriction site ofpJOC21. Plasmid pJOC22BCKm, harboring miniUIB-BCKm (Fig. 1E),was obtained by cloning the Kmr determinant of pCSI2 into the XbaIrestriction site of pJOC22BC. Plasmid pJOC22KNG, harboring miniUIB-KNG (Fig. 1F), was obtained by cloning the SmaI-linearized pKNG101plasmid (Smr [20]) between the SmaI and EcoRV restriction sites ofpJOC21. Plasmid pJOC22KmKNG, harboring miniUIB-KmKNG(Fig. 1G), was obtained by cloning the Kmr determinant of pCSI2 into theXbaI restriction site of pJOC22KNG. Plasmid pJOC22BCKmKNG, har-boring miniUIB-BCKmKNG (Fig. 1H), was obtained by cloning theSmaI-linearized pBC SK(�) cloning vector (Cmr; Stratagene) into theSmaI restriction site of pJOC22KmKNG.
Plasmid pALK, containing the miniUIB-ALK minitransposon(Fig. 1J) with the alkST and alkBFGHJKL determinants of P. putidaTF4-1L (GPo1), was constructed in three steps. First, the alkBFGH deter-
minants were inserted between the XbaI and SacI restriction sites ofpJOC21 after PCR amplification with primers ALK1-F and ALK1-R. ThealkJKL genes were then amplified by PCR using primers ALK2-F andALK2-R and further cloned in the SacI restriction site, resulting in thepALKB plasmid that harbors the miniUIB-ALKB minitransposon (Fig.1I). Finally, the regulator operon alkST was amplified with primersALKRsma-F and ALKRsma-R and inserted in the SmaI restriction site ofpALKB, resulting in pALK plasmid.
Finally, pJOC100 (Fig. 1K) and pJOC100ALK (Fig. 1M) plasmids,both containing the transposition machinery of ISPpu12 without the IRs,were obtained by PCR amplification of IR-free ISPpu12 of P. stutzeriAN10 with primers ISPPU-25F-eco and ISPPU-3348R-eco, followed bycloning in the EcoRI restriction site of pJOC21 and pALK, respectively.Plasmid pJOC100Km (Fig. 1L) was obtained after cloning the Kmr frompCSI2 in the EcoRV restriction site of pJOC100.
Nucleotide sequence accession number. The sequence of the pJOC21plasmid has been deposited in GenBank under accession numberKC295444.
RESULTS AND DISCUSSIONActivated ISPpu12 has an in trans effect on genetic structuresflanked by ISPpu12-like IRs. Previous experiments revealed thatISPpu12 activates its own transposition, and it was also responsi-ble for the transposition of ISPst9 in P. stutzeri AN10 after conju-gative interaction (12). This in trans effect on ISPst9, an element ofthe same ISL3 family as ISPpu12 (17), was explained by the highsimilarity between their IRs and therefore, the plausible recogni-tion by the transposase of ISPpu12. An ISPpu12-derived mini-transposon, designated as miniUIB, was designed to explore the intrans activity of ISPpu12 on genetic structures flanked by similarIRs to those found in the transposon (Fig. 1A). The miniUIBstructure was constructed on the R6K-replicase suicidal plasmidpGP704 giving pJOC21. To follow the mobilization of miniUIB,the Kmr gene of pCSI2 plasmid was cloned between the IRs,producing miniUIB-Km (Fig. 1B). The resulting plasmid,pJOC22Km, was transferred into P. stutzeri AN10 (original strainwith ISPpu12 encoded in its genome) and P. stutzeri AN11 (neg-ative control, lacking ISPpu12 in its genome) by a 7-h RP4-basedconjugation using E. coli S17-1�pir as donor strain. Conjugativeinteraction was expected to activate the transposition mechanismof ISPpu12 in P. stutzeri AN10, having an in trans effect on theentering miniUIB-Km structure located on plasmid pJOC22Km.The plasmid pGP704Km, similar to pJOC22Km but with no IRsflanking the Kmr determinant, was used as a negative control. Asshown in Table 2, the frequency of Kmr acquisition was almost 500times higher in P. stutzeri AN10 when the antibiotic resistancecassette was flanked by the IRs. Also, in all analyzed cases, the Kmr
transconjugants did not acquire the Apr determinant of the vector.On the other hand, when conjugating with pGP704Km, the Kmr
gene was acquired at a low frequency, similar to that seen for theApr determinant harbored in the plasmid (Table 2), indicatingthat the entire plasmid which is unable to replicate in Pseudomo-nas, was incorporated into the genome of P. stutzeri AN10 by agenetic process not related to transposition (i.e., recombination).As expected, no differences were observed between both plasmids(pJOC22Km and pGP704Km) when P. stutzeri AN11 (strain with-out ISPpu12 encoded in its genome) was used as conjugative re-cipient strain (Table 2).
Ten of the P. stutzeri AN10 transconjugants (P. stutzeriAN10Km) which had acquired the Kmr determinant frompJOC22Km were further analyzed by genomic DNA digestion fol-
MiniUIB Minitransposon
March 2013 Volume 79 Number 5 aem.asm.org 1631
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 20
0.12
6.83
.173
.

lowed by Southern blotting hybridization. Between one and threehybridization bands were obtained when a specific probe for theKmr gene (KM probe) was used (Fig. 2A), whereas no hybridiza-tion signal was obtained when pGP704 was used as a probe (resultnot shown). As expected, hybridizations with TNPA, a probe forthe transposase gene of both ISPst9 and ISPpu12, and TNPR, aprobe specific for the regulator gene of ISPpu12, revealed transpo-sition of both ISL3-like elements of P. stutzeri AN10 (Fig. 2B).
In order to reduce the number of miniUIB copies acquired by therecipient strain, conjugation experiments between E. coli S17-1�pir
(pJOC22Km) and P. stutzeri AN10 using reduced mating times wereperformed. Maximum frequency of Kmr acquisition was obtainedjust after 30 min of mating time (Fig. 2C). As previously done, 10 P.stutzeri AN10 transconjugants that acquired the Kmr determinantfrom pJOC22Km were further analyzed by genomic DNA digestionwith EcoRI, followed by Southern blotting hybridization. A singlehybridization band was obtained in 9 of them when the KM probewas used (see Fig. S1 in the supplemental material). Due to this, fur-ther mating experiments were carried out with only 30 min of conju-gative interaction and not the 7 h used previously.
FIG 1 Physical and genetic maps of the miniUIB structures. Genetic structures harbored on pGP704 plasmid are shown. Gray and white triangles represent theleft and right IRs of ISPst9, respectively. Unique restriction sites (B, BglII; E, EcoRI; K, KpnI; M, SmaI; N, NdeI; R, EcoRV; S, SacI; X, XbaI) are indicated. (A)miniUIB in pJOC21; (B) miniUIB-Km (1.6 kb) in pJOC22Km; (C) miniUIB-Cm (2.1 kb) in pJOC22Cm; (D) miniUIB-BC (3.6 kb) in pJOC22BC; (E)miniUIB-BC/Km (5.0 kb) in pJOC22BCKm; (F) miniUIB-KNG (7.0 kb) in pJOC22KNG; (G) miniUIB-Km/KNG (8.4 kb) in pJOC22KmKNG; (H) miniUIB-BC/Km/KNG (11.8 kb) in pJOC22BCKmKNG; (I) miniUIB-ALKB (8.5 kb) in pALKB; (J) miniUIB-ALK (12.8 kb) in pALK; (K) miniUIB beside IR-deletedISPpu12 in pJOC100; (L) miniUIB-Km beside IR-deleted ISPpu12 in pJOC100Km; (M) miniUIB-ALK beside IR-deleted ISPpu12 in pJOC100ALK.
Christie-Oleza et al.
1632 aem.asm.org Applied and Environmental Microbiology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 20
0.12
6.83
.173
.
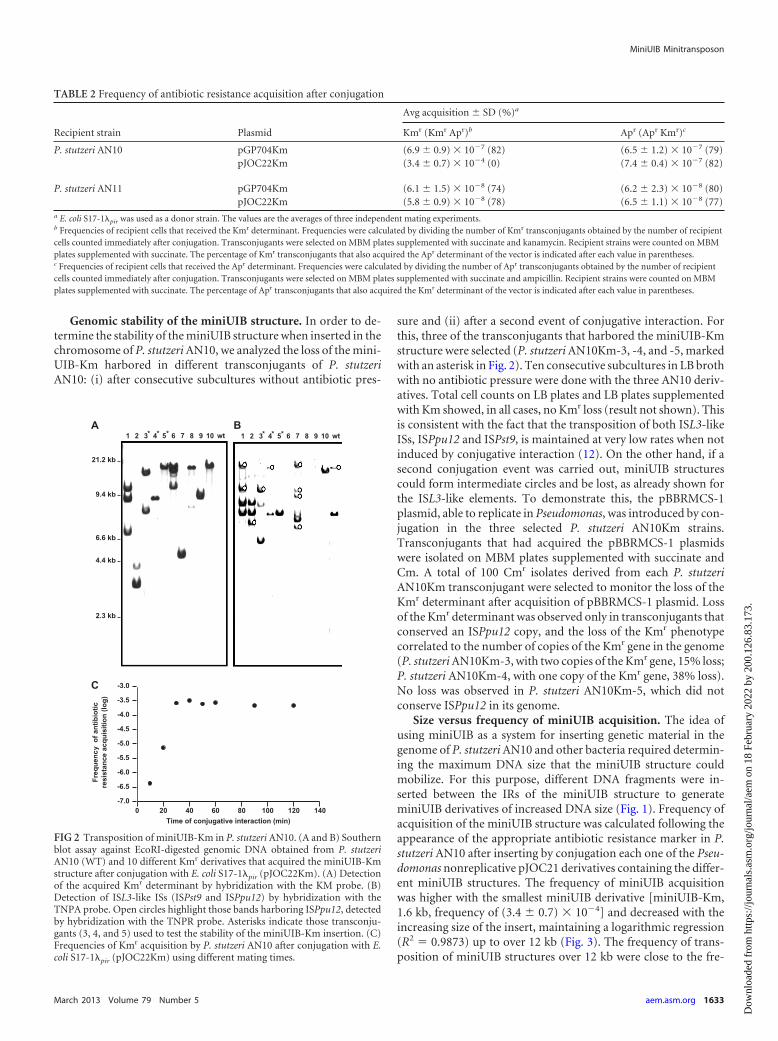
Genomic stability of the miniUIB structure. In order to de-termine the stability of the miniUIB structure when inserted in thechromosome of P. stutzeri AN10, we analyzed the loss of the mini-UIB-Km harbored in different transconjugants of P. stutzeriAN10: (i) after consecutive subcultures without antibiotic pres-
sure and (ii) after a second event of conjugative interaction. Forthis, three of the transconjugants that harbored the miniUIB-Kmstructure were selected (P. stutzeri AN10Km-3, -4, and -5, markedwith an asterisk in Fig. 2). Ten consecutive subcultures in LB brothwith no antibiotic pressure were done with the three AN10 deriv-atives. Total cell counts on LB plates and LB plates supplementedwith Km showed, in all cases, no Kmr loss (result not shown). Thisis consistent with the fact that the transposition of both ISL3-likeISs, ISPpu12 and ISPst9, is maintained at very low rates when notinduced by conjugative interaction (12). On the other hand, if asecond conjugation event was carried out, miniUIB structurescould form intermediate circles and be lost, as already shown forthe ISL3-like elements. To demonstrate this, the pBBRMCS-1plasmid, able to replicate in Pseudomonas, was introduced by con-jugation in the three selected P. stutzeri AN10Km strains.Transconjugants that had acquired the pBBRMCS-1 plasmidswere isolated on MBM plates supplemented with succinate andCm. A total of 100 Cmr isolates derived from each P. stutzeriAN10Km transconjugant were selected to monitor the loss of theKmr determinant after acquisition of pBBRMCS-1 plasmid. Lossof the Kmr determinant was observed only in transconjugants thatconserved an ISPpu12 copy, and the loss of the Kmr phenotypecorrelated to the number of copies of the Kmr gene in the genome(P. stutzeri AN10Km-3, with two copies of the Kmr gene, 15% loss;P. stutzeri AN10Km-4, with one copy of the Kmr gene, 38% loss).No loss was observed in P. stutzeri AN10Km-5, which did notconserve ISPpu12 in its genome.
Size versus frequency of miniUIB acquisition. The idea ofusing miniUIB as a system for inserting genetic material in thegenome of P. stutzeri AN10 and other bacteria required determin-ing the maximum DNA size that the miniUIB structure couldmobilize. For this purpose, different DNA fragments were in-serted between the IRs of the miniUIB structure to generateminiUIB derivatives of increased DNA size (Fig. 1). Frequency ofacquisition of the miniUIB structure was calculated following theappearance of the appropriate antibiotic resistance marker in P.stutzeri AN10 after inserting by conjugation each one of the Pseu-domonas nonreplicative pJOC21 derivatives containing the differ-ent miniUIB structures. The frequency of miniUIB acquisitionwas higher with the smallest miniUIB derivative [miniUIB-Km,1.6 kb, frequency of (3.4 � 0.7) � 10�4] and decreased with theincreasing size of the insert, maintaining a logarithmic regression(R2 � 0.9873) up to over 12 kb (Fig. 3). The frequency of trans-position of miniUIB structures over 12 kb were close to the fre-
TABLE 2 Frequency of antibiotic resistance acquisition after conjugation
Recipient strain Plasmid
Avg acquisition � SD (%)a
Kmr (Kmr Apr)b Apr (Apr Kmr)c
P. stutzeri AN10 pGP704Km (6.9 � 0.9) � 10�7 (82) (6.5 � 1.2) � 10�7 (79)pJOC22Km (3.4 � 0.7) � 10�4 (0) (7.4 � 0.4) � 10�7 (82)
P. stutzeri AN11 pGP704Km (6.1 � 1.5) � 10�8 (74) (6.2 � 2.3) � 10�8 (80)pJOC22Km (5.8 � 0.9) � 10�8 (78) (6.5 � 1.1) � 10�8 (77)
a E. coli S17-1�pir was used as a donor strain. The values are the averages of three independent mating experiments.b Frequencies of recipient cells that received the Kmr determinant. Frequencies were calculated by dividing the number of Kmr transconjugants obtained by the number of recipientcells counted immediately after conjugation. Transconjugants were selected on MBM plates supplemented with succinate and kanamycin. Recipient strains were counted on MBMplates supplemented with succinate. The percentage of Kmr transconjugants that also acquired the Apr determinant of the vector is indicated after each value in parentheses.c Frequencies of recipient cells that received the Apr determinant. Frequencies were calculated by dividing the number of Apr transconjugants obtained by the number of recipientcells counted immediately after conjugation. Transconjugants were selected on MBM plates supplemented with succinate and ampicillin. Recipient strains were counted on MBMplates supplemented with succinate. The percentage of Apr transconjugants that also acquired the Kmr determinant of the vector is indicated after each value in parentheses.
FIG 2 Transposition of miniUIB-Km in P. stutzeri AN10. (A and B) Southernblot assay against EcoRI-digested genomic DNA obtained from P. stutzeriAN10 (WT) and 10 different Kmr derivatives that acquired the miniUIB-Kmstructure after conjugation with E. coli S17-1�pir (pJOC22Km). (A) Detectionof the acquired Kmr determinant by hybridization with the KM probe. (B)Detection of ISL3-like ISs (ISPst9 and ISPpu12) by hybridization with theTNPA probe. Open circles highlight those bands harboring ISPpu12, detectedby hybridization with the TNPR probe. Asterisks indicate those transconju-gants (3, 4, and 5) used to test the stability of the miniUIB-Km insertion. (C)Frequencies of Kmr acquisition by P. stutzeri AN10 after conjugation with E.coli S17-1�pir (pJOC22Km) using different mating times.
MiniUIB Minitransposon
March 2013 Volume 79 Number 5 aem.asm.org 1633
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 20
0.12
6.83
.173
.

quencies obtained for the control plasmid pGP704Km (Table 2),unable to transpose and replicate in Pseudomonas. These data sug-gest that longer inserts may undergo transposition at lower fre-quencies, similar to the frequency of insertion by random recom-bination. This was corroborated by the increased detection of Apr
colonies (up to 50%), the resistance determinant marker ofpGP704 plasmid, when large miniUIB structures were used(Fig. 3).
Biotechnological applications of miniUIB in P. stutzeriAN10. Here, we present two different strategies in which theminiUIB element was used to transform P. stutzeri AN10 into analkane-degrading bacterium.
The first strategy consisted in introducing the alkane determi-nants in a single conjugation step. For this, we constructed thepALK plasmid (Table 1), which harbors the miniUIB-ALK mini-transposon (Fig. 1J). The miniUIB-ALK contains the two alkanedegradation operons of P. putida TF4-1L (GPo1) (21): the 4.3-kbregulation operon alkST, and the 8.3-kb catabolic operon alkBF-
GHJKL. Plasmid pALK was introduced into AN10 by conjugation.The mixture of mated cells were collected and incubated in MBMsupplemented with n-octane and n-hexadecane (each at 2% [wt/vol])as the sole carbon and energy source in order to select for transcon-jugants carrying the miniUIB-ALK structure. Genes alkBFGHJKL en-code the functions necessary for the first steps of C5-C12 n-alkanesdegradation such as uptake, hydroxylation, and dehydrogenation ofthe compounds (24). Thus, n-octane was used as carbon and energysource. The substrate can be mineralized when the cell has an active�-oxidation pathway for fatty acids. The fatty acid degradation path-way of P. stutzeri could be constitutively expressed as in P. putida (24)or could be induced only by fatty acids longer than C12-like in E. coli(25). To ensure the induction of the fatty acid degradation genes and,by this way, not to be dependent on the appearance of a spontaneousconstitutive mutant, we added 2% of n-hexadecane to the media, aspreviously done by Eggink et al. (25). After incubation in MBM sup-plemented with n-octane and n-hexadecane, growth was only ob-tained in AN10 cells mated with E. coli S17-1�pir (pALK) and not inthe three negative controls used: (i) P. stutzeri AN10, (ii) E. coli S17-1�pir (pALK), and (iii) AN10 cells mated with E. coli S17-1�pir har-boring plasmid pALKB, which contains the catabolic operon alkBF-GHJKL but lacks the regulatory operon alkST. Growth also failed inAN10 cells mated with E. coli S17-1�pir (pALK) but incubated inMBM without alkane supplementation.
An aliquot of the culture grown in alkanes was serially dilutedand plated on MBM, and further incubated with n-octane andn-hexadecane in vapor phase as unique carbon and energysources. Eight transconjugants (P. stutzeri AN10-ALK) were iso-lated and proved to be AN10 derivatives by 16S rRNA gene se-quence analysis. In order to discriminate whether the miniUIB-ALK transposed into the AN10 genome or whether pALK wascaptured, we cultivated all eight AN10-ALK transconjugants inthe presence of Ap. Only one of the eight isolates analyzed (P.stutzeri AN10-ALK1) was Apr, indicating that in this transconju-gant the whole plasmid had been acquired. This was further con-firmed by Southern blotting hybridization analysis using a specificprobe for the alk determinants and for linearized pGP704(Fig. 4A). Hybridization with the ALK probe revealed the presenceof the alk determinants in all transconjugants, whereas the PGPprobe (for pGP704 plasmid) gave a hybridization band only withthe AN10-ALK-1 transconjugant. In addition, hybridization alsorevealed that only one of the transconjugants (AN10-ALK8) har-bored more than one copy of the miniUIB-ALK structure. Thus, it
FIG 3 Frequency of miniUIB acquisition by P. stutzeri AN10 versus size ofminiUIB structure. Strain AN10 was conjugated with E. coli S17-1�pir harbor-ing different pJOC21 derivatives. Dots and error bars indicate averages andstandard deviations of three independent experiments, respectively. Percent-ages revealed entire plasmid acquisition, evaluated by acquired Apr phenotypeof isolated transconjugants. Dashed line shows the frequency of entirepGP704Km acquisition by P. stutzeri AN10 after conjugation with E. coli S17-1�pir harboring this plasmid. Continuous line shows the logarithmical regres-sion between both parameters. Plasmids, miniUIB derivatives, and miniUIBlength are as follows: A, pJOC22Km, miniUIB-Km, 1.6 kb; B, pJOC22Cm,miniUIB-Cm, 2.1 kb; C, pJOC22BC, miniUIB-BC, 3.6 kb; D, pJOC22BCKm,miniUIB-BC/Km, 5.0 kb; E, pJOC22KNG, miniUIB-KNG, 7.0 kb; F,pJOC22KmKNG, miniUIB-Km/KNG, 8.4 kb; G, pJOC22BCKmKNG, mini-UIB-BC/Km/KNG, 11.8 kb.
FIG 4 Alkane degradation by P. stutzeri AN10 and it derivatives harboring the miniUIB-ALK structure. (A) Southern blot hybridization with the ALK probeagainst EcoRI-digested genomic DNA obtained from P. stutzeri AN10 (WT) and 8 different AN10-ALK derivatives that acquired the miniUIB-ALK structureafter conjugation with E. coli S17-1�pir (pALK). Open circle highlight the unique bands harboring the entire pALK plasmid detected by hybridization with thePGP probe. Asterisks indicate those transconjugants (3, AN10-ALK3; 8, AN10-ALK8) used to evaluate the acquired alkane degradation phenotype. (B) Growth(maximum cell density) of P. stutzeri AN10, AN10-ALK3, and AN10-ALK8 on MBM supplemented with n-octane (black bars), n-hexadecane (gray bars), anda mixture of both n-alkanes (white bars). Values are represented as the averages and standard deviations of three independent experiments.
Christie-Oleza et al.
1634 aem.asm.org Applied and Environmental Microbiology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 20
0.12
6.83
.173
.

was demonstrated that the alk determinants were acquired by truetransposition of the 12.8-kb miniUIB-ALK structure in 7 of the 8transconjugants tested. P. stutzeri strains AN10-ALK3 and AN10-ALK8 were selected to carry out further growth experiments (Fig.4B). Both AN10-ALK transconjugants could grow with only n-oc-tane although absorbance measurements (600 nm) of the culturesalmost doubled at the stationary phase when hexadecane wasadded. No growth was detected when hexadecane was used as solecarbon and energy source, probably because the alk genes onlyallow the degradation of C5-C12 n-alkanes (24). No differences ingrowth were detected due to the presence of one or two copies ofthe miniUIB-ALK structure (Fig. 4B), probably due to the enrich-ment strategy used for the selection of transconjugants.
The second strategy consisted in introducing the alkane deter-minants in two separate conjugation steps. The rationale of thisexperiment was to demonstrate that, although a second conjuga-tion event could result in the loss of the first miniUIB structureinserted, a sufficiently high percentage of the population will endwith both miniUIB structures introduced in successive events. Amulticonjugation approach to introduce different DNA frag-ments in sequential transposition steps could also be of great in-terest for future biotechnological applications. This strategy ofusing multiple steps of DNA integration with the same minitrans-poson is not possible with minitransposons with transpositionimmunity, like Tn7 derivatives (10). For this two-step process, weused the previously constructed plasmid pALKB (harboringminiUIB-ALKB, which contains the alkBFGHJKL operon)(Fig. 1I) and a newly constructed plasmid pALKC (harboringminiUIB-ALKC, which contains the regulatory operon alkST anda chloramphenicol resistance determinant) (see Fig. S2 in the sup-plemental material). P. stutzeri AN10 was first mated with E. coliS17-1�pir (pALKC). Mated cells were then grown in liquid MBMsupplemented with succinate 0.5% (wt/vol) and Cm to selectthose AN10 transconjugants that had acquired miniUIB-ALKC.The grown culture was used directly in a second conjugation ex-periment with E. coli S17-1�pir (pALKB). As in the first strategy,the resultant second mating event was grown in MBM supple-mented with n-octane and n-hexadecane as sole carbon andenergy sources. Successful growth was only obtained in double-conjugated AN10 cells and not in the negative controls (see Fig. S3in the supplemental material).
Extending the use of the miniUIB system to ISPpu12-freemodel microorganisms. The use of the miniUIB structure in bac-teria lacking ISPpu12 was tested in another member of P. stutzerispecies and in two different widely used model species: Klebsiellapneumoniae, related to respiratory infections (up to 14,795 refer-ences in PubMed on 1 December 2012) and P. putida, involved inhydrocarbon biodegradation (up to 5,566 references in PubMedon the same date). The strategy used was started by introducingboth tnpA and tnpR genes of ISPpu12 in the suicidal vector, out-side of the miniUIB structure. For this, the complete ISPpu12sequence (without the IRs) was inserted in the EcoRI restrictionsite of pJOC21, obtaining pJOC100 (Fig. 1K). The gene structureof the IS was conserved in order to maintain its activation pheno-type after conjugative interaction. Unlike the instability reportedfor mini-Tn5 (1), plasmid pJOC100 was stable in E. coli, probablydue to the transcriptional repression of tnpA mediated by TnpR inthe absence of stimuli (12).
In order to test the integration of miniUIB in ISPpu12-lackingstrains, the miniUIB-Km structure was reconstructed by cloning
the Kmr gene of pCSI2 between the IRs, giving pJOC100Km (Fig.1L). This construction was introduced in three different bacterialstrains lacking ISPpu12 in their genome: K. pneumoniae CMD1, P.stutzeri AN11, and P. putida IS�. The results with miniUIB-Kmwere compared to the ones obtained with its homolog in theminiTn5 series: the miniTn5-Km1 minitransposon (1, 2). Allmating experiments were performed in triplicate. In all cases, Kmr
acquisition frequencies were higher when miniUIB-Km was used[K. pneumoniae CMD1, (4.91 � 0.6) � 10�5 for miniUIB-Km,(1.4�0.7)�10�5 for miniTn5-Km1; P. stutzeri AN11, (1.1�0.3)�10�4 for miniUIB-Km, (3.9 � 1.2) � 10�8 for miniTn5-Km1; P.putida IS�, (9.7 � 0.8) � 10�5 for miniUIB-Km, (5.2 � 2.8) � 10�6
for miniTn5-Km1].In order to determine whether miniUIB-Km was randomly
inserted in single copy, eight P. stutzeri AN11 Kmr derivatives wereselected, and a Southern blot hybridization was performed againsttheir EcoRI-digested genomic DNAs with the KM probe. All Kmr
AN11 derivatives showed different single hybridization bands(Fig. 5), suggesting that miniUIB-Km was randomly inserted bytransposition in the genome of P. stutzeri AN11 and, thus, it can beused as a genetic tool for random transposon mutagenesis inISPpu12-lacking microorganisms.
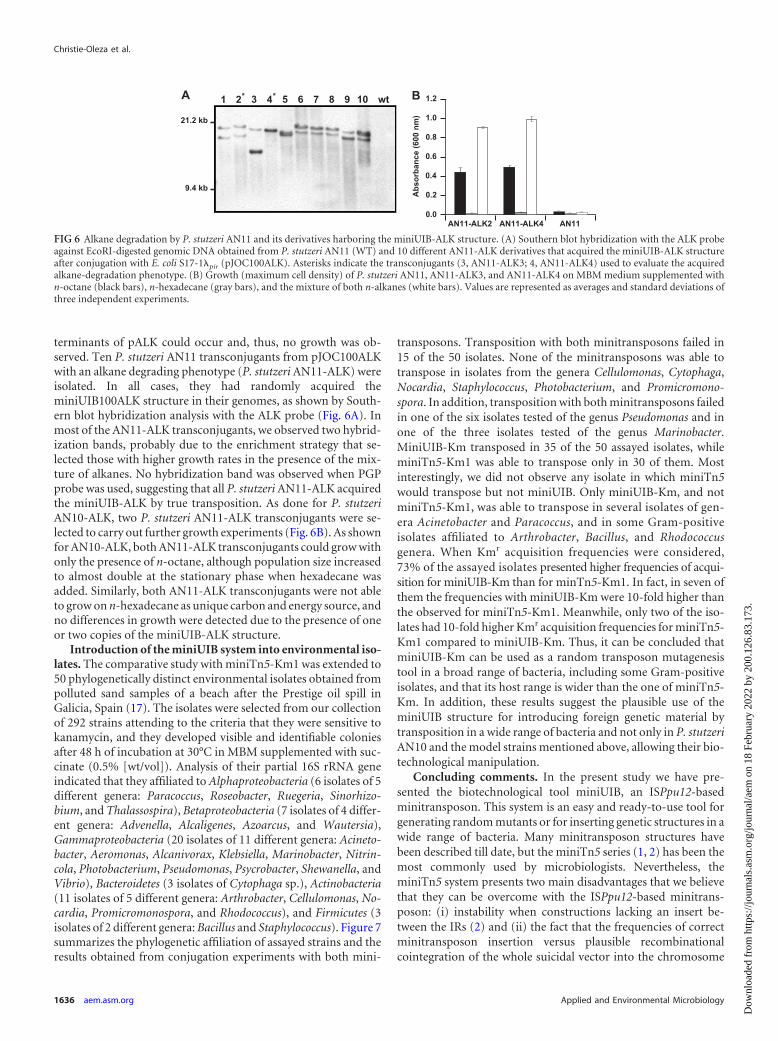
In order to prove that the miniUIB structure could be usefulfor introducing biotechnologically relevant foreign-DNA inISPpu12-free bacteria, we transformed P. stutzeri AN11 into analkane-degrading bacterium by introducing the miniUIB-ALKstructure. For this purpose we constructed pJOC100ALK (Fig.1M) by cloning the IR-free ISPpu12 of P. stutzeri AN10 into theEcoRI site of pALK. Following the strategy previously used for P.stutzeri AN10, we obtained a P. stutzeri AN11 derivative cultureable to grow using n-octane and n-hexadecane as unique carbonand energy source (Fig. 6). No growth was observed in controlcultures. A negative control using pALK (which previously gave apositive growth in AN10) was also included. As expected, due tothe absence of ISPpu12 in the genome of P. stutzeri AN11 genome,no transposition and, consequently, no acquisition of the alk de-
FIG 5 Southern blot hybridization with the KM probe against EcoRI-digestedgenomic DNA obtained from eight different Kmr derivatives of P. stutzeriAN11 that acquired the miniUIB-Km structure after conjugation with E. coliS17-1�pir (pJOC100Km).
MiniUIB Minitransposon
March 2013 Volume 79 Number 5 aem.asm.org 1635
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 20
0.12
6.83
.173
.
terminants of pALK could occur and, thus, no growth was ob-served. Ten P. stutzeri AN11 transconjugants from pJOC100ALKwith an alkane degrading phenotype (P. stutzeri AN11-ALK) wereisolated. In all cases, they had randomly acquired theminiUIB100ALK structure in their genomes, as shown by South-ern blot hybridization analysis with the ALK probe (Fig. 6A). Inmost of the AN11-ALK transconjugants, we observed two hybrid-ization bands, probably due to the enrichment strategy that se-lected those with higher growth rates in the presence of the mix-ture of alkanes. No hybridization band was observed when PGPprobe was used, suggesting that all P. stutzeri AN11-ALK acquiredthe miniUIB-ALK by true transposition. As done for P. stutzeriAN10-ALK, two P. stutzeri AN11-ALK transconjugants were se-lected to carry out further growth experiments (Fig. 6B). As shownfor AN10-ALK, both AN11-ALK transconjugants could grow withonly the presence of n-octane, although population size increasedto almost double at the stationary phase when hexadecane wasadded. Similarly, both AN11-ALK transconjugants were not ableto grow on n-hexadecane as unique carbon and energy source, andno differences in growth were detected due to the presence of oneor two copies of the miniUIB-ALK structure.
Introduction of the miniUIB system into environmental iso-lates. The comparative study with miniTn5-Km1 was extended to50 phylogenetically distinct environmental isolates obtained frompolluted sand samples of a beach after the Prestige oil spill inGalicia, Spain (17). The isolates were selected from our collectionof 292 strains attending to the criteria that they were sensitive tokanamycin, and they developed visible and identifiable coloniesafter 48 h of incubation at 30°C in MBM supplemented with suc-cinate (0.5% [wt/vol]). Analysis of their partial 16S rRNA geneindicated that they affiliated to Alphaproteobacteria (6 isolates of 5different genera: Paracoccus, Roseobacter, Ruegeria, Sinorhizo-bium, and Thalassospira), Betaproteobacteria (7 isolates of 4 differ-ent genera: Advenella, Alcaligenes, Azoarcus, and Wautersia),Gammaproteobacteria (20 isolates of 11 different genera: Acineto-bacter, Aeromonas, Alcanivorax, Klebsiella, Marinobacter, Nitrin-cola, Photobacterium, Pseudomonas, Psycrobacter, Shewanella, andVibrio), Bacteroidetes (3 isolates of Cytophaga sp.), Actinobacteria(11 isolates of 5 different genera: Arthrobacter, Cellulomonas, No-cardia, Promicromonospora, and Rhodococcus), and Firmicutes (3isolates of 2 different genera: Bacillus and Staphylococcus). Figure 7summarizes the phylogenetic affiliation of assayed strains and theresults obtained from conjugation experiments with both mini-
transposons. Transposition with both minitransposons failed in15 of the 50 isolates. None of the minitransposons was able totranspose in isolates from the genera Cellulomonas, Cytophaga,Nocardia, Staphylococcus, Photobacterium, and Promicromono-spora. In addition, transposition with both minitransposons failedin one of the six isolates tested of the genus Pseudomonas and inone of the three isolates tested of the genus Marinobacter.MiniUIB-Km transposed in 35 of the 50 assayed isolates, whileminiTn5-Km1 was able to transpose only in 30 of them. Mostinterestingly, we did not observe any isolate in which miniTn5would transpose but not miniUIB. Only miniUIB-Km, and notminiTn5-Km1, was able to transpose in several isolates of gen-era Acinetobacter and Paracoccus, and in some Gram-positiveisolates affiliated to Arthrobacter, Bacillus, and Rhodococcusgenera. When Kmr acquisition frequencies were considered,73% of the assayed isolates presented higher frequencies of acqui-sition for miniUIB-Km than for minTn5-Km1. In fact, in seven ofthem the frequencies with miniUIB-Km were 10-fold higher thanthe observed for miniTn5-Km1. Meanwhile, only two of the iso-lates had 10-fold higher Kmr acquisition frequencies for miniTn5-Km1 compared to miniUIB-Km. Thus, it can be concluded thatminiUIB-Km can be used as a random transposon mutagenesistool in a broad range of bacteria, including some Gram-positiveisolates, and that its host range is wider than the one of miniTn5-Km. In addition, these results suggest the plausible use of theminiUIB structure for introducing foreign genetic material bytransposition in a wide range of bacteria and not only in P. stutzeriAN10 and the model strains mentioned above, allowing their bio-technological manipulation.
Concluding comments. In the present study we have pre-sented the biotechnological tool miniUIB, an ISPpu12-basedminitransposon. This system is an easy and ready-to-use tool forgenerating random mutants or for inserting genetic structures in awide range of bacteria. Many minitransposon structures havebeen described till date, but the miniTn5 series (1, 2) has been themost commonly used by microbiologists. Nevertheless, theminiTn5 system presents two main disadvantages that we believethat they can be overcome with the ISPpu12-based minitrans-poson: (i) instability when constructions lacking an insert be-tween the IRs (2) and (ii) the fact that the frequencies of correctminitransposon insertion versus plausible recombinationalcointegration of the whole suicidal vector into the chromosome
FIG 6 Alkane degradation by P. stutzeri AN11 and its derivatives harboring the miniUIB-ALK structure. (A) Southern blot hybridization with the ALK probeagainst EcoRI-digested genomic DNA obtained from P. stutzeri AN11 (WT) and 10 different AN11-ALK derivatives that acquired the miniUIB-ALK structureafter conjugation with E. coli S17-1�pir (pJOC100ALK). Asterisks indicate the transconjugants (3, AN11-ALK3; 4, AN11-ALK4) used to evaluate the acquiredalkane-degradation phenotype. (B) Growth (maximum cell density) of P. stutzeri AN11, AN11-ALK3, and AN11-ALK4 on MBM medium supplemented withn-octane (black bars), n-hexadecane (gray bars), and the mixture of both n-alkanes (white bars). Values are represented as averages and standard deviations ofthree independent experiments.
Christie-Oleza et al.
1636 aem.asm.org Applied and Environmental Microbiology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 20
0.12
6.83
.173
.

are insufficient in some strains due to the dependence of hostregulation for transposition.
Plasmid pJOC100 harbors the exclusive self-regulated transpo-sition mechanism of ISPpu12, which is strongly activated onlyafter conjugative interaction (12, 13). This converts the minitrans-poson miniUIB into a host-independent system that, as intended,is highly active only during the conjugation process. UnlikeminiTn5, which is unstable when nothing is cloned between itsIRs (2), miniUIB is a ready-to-use system with no need of auxiliaryplasmids. In this sense, miniUIB is a stable structure ready forcloning DNA fragments up to 12 kb in length into its multicloningsite. The high frequency of transposition allows selecting for realtransconjugants (i.e., the alk determinants) by their phenotype,without the use of nondesirable markers (i.e., antibiotic resistancedeterminants).
Finally, we have also demonstrated that it is possible to intro-duce distinct DNA-fragments in the same recipient cell by usingdifferent miniUIB structures in successive conjugation steps. Thisprobably makes the miniUIB more attractive for its biotechnolog-ical use than other systems, such as the miniTn7-based minitrans-posons which display transposition immunity (9, 10).
ACKNOWLEDGMENTS
J.A.C.-O was supported by grants from the Government of the BalearicIslands, Ramon Areces Foundation, and Marie Curie Actions of the ECFP7. I.B.-G. was supported by a fellowship from the Government of theBalearic Islands (with FSE cofunding). Funds were obtained from projectsCTM2008-02574/MAR, CSD2009-00006 and CTM2011-24886 (FEDERcofunding) from the Spanish Ministry of Economy and Competitivity(MINECO). Funds for competitive research groups from the Govern-ment of the Balearic Islands (FEDER cofunding) are also acknowledged.
A patent has been submitted to the Spanish Office for Patents andTrademarks (reference number P201230697, 09/05/2012), which coversthe work described in this article.
REFERENCES1. de Lorenzo V, Herrero M, Jakubzik U, Timmis KN. 1990. Mini-Tn5
transposon derivatives for insertion mutagenesis, promoter probing, andchromosomal insertion of cloned DNA in gram-negative eubacteria. J.Bacteriol. 172:6568 – 6572.
2. Herrero M, de Lorenzo V, Timmis KN. 1990. Transposon vectors con-taining non-antibiotic resistance selection markers for cloning and stablechromosomal insertion of foreign genes in gram-negative bacteria. J. Bac-teriol. 172:6557– 6567.
3. Kolter R, Inuzuka M, Helinski DR. 1978. Trans-complementation-dependent replication of a low molecular weight origin fragment fromplasmid R6K. Cell 15:1199 –1208.
4. Miller VL, Mekalanos JJ. 1988. A novel suicide vector and its use inconstruction of insertion mutations: osmoregulation of outer membraneproteins and virulence determinants in Vibrio cholerae requires toxR. J.Bacteriol. 170:2575–2583.
5. Taghavi S, Delanghe H, Lodewyckx C, Mergeay M, van der Lelie D.2001. Nickel-resistance-based minitransposons: new tools for genetic ma-nipulation of environmental bacteria. Appl. Environ. Microbiol. 67:1015–1019.
6. Rossignol M, Basset A, Espéli O, Boccard F. 2001. NKBOR, a mini-Tn10-based transposon for random insertion in the chromosome of Gram-negativebacteria and the rapid recovery of sequences flanking the insertion sites inEscherichia coli. Res. Microbiol. 152:481–485.
7. Li R, Wang G, Shen B, Wang R, Song Y, Li S, Jiang J. 2009. Randomtransposon vectors pUTTns for the markerless integration of exogenousgenes into gram-negative eubacteria chromosomes. J. Microbiol. Meth-ods 79:220 –226.
8. Peters JE, Craig NL. 2001. Tn7: smarter than we thought. Nat. Rev. Mol.Cell. Biol. 2:806 – 814.
9. Choi KH, Gaynor JB, White KG, Lopez C, Bosio CM, Karkhoff-
FIG 7 Comparison of the transposition frequencies of miniUIB-Km and miniTn5-Km1 in all phylogenetically different isolates used. The phylogenetic tree(neighbor-joining dendrogram based on Jukes-Cantor 16S rRNA gene distance matrix) shows relationships between the type strains of each represented generaused in the present study. First column (isolates) indicates the number of isolates used that affiliated to each genus (28 total genera). Second and third columnindicate the number of isolates that acquired the Kmr determinant of miniUIB-Km and miniTn5-Km1, respectively. Plotted comparisons (frequency of Kmr
acquisition of miniUIB-Km divided by the one obtained for miniTn5-Km1) were calculated only with those 33 isolates that were able to receive both minitrans-posons. Black circles indicate the 22 isolates that presented higher frequencies for miniUIB-Km than for miniTn5-Km1. White circles indicate the eight isolateswith higher frequencies for miniUIB-Tn5 than for miniUIB-Km. Gray circles indicate the ratio of transposition obtained for P. stutzeri AN11 (number 1), P.putida IS� (number 2), and K. pneumoniae CMD1 (number 3).
MiniUIB Minitransposon
March 2013 Volume 79 Number 5 aem.asm.org 1637
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 20
0.12
6.83
.173
.

Schweizer RR, Schweizer HP. 2005. A Tn7-based broad-range bacterialcloning and expression system. Nat. Methods 2:443– 448.
10. Arciszewska LK, Drake D, Craig NL. 1989. Transposon Tn7 cis-actingsequences in transposition and transposition immunity. J. Mol. Biol. 207:35–52.
11. Williams PA, Jones RM, Shaw LE. 2002. A third transposable element,ISPpu12, from the toluene-xylene catabolic plasmid pWW0 of Pseudomo-nas putida mt-2. J. Bacteriol. 184:6572– 6580.
12. Christie-Oleza JA, Nogales B, Lalucat J, Bosch R. 2010. TnpR encodedby an ISPpu12 isoform regulates transposition of two different ISL3-likeinsertion sequences in Pseudomonas stutzeri after conjugative interaction.J. Bacteriol. 192:1423–1432.
13. Christie-Oleza JA, Lanfranconi MP, Nogales B, Lalucat J, Bosch R.2009. Conjugative interaction induces transposition of ISPst9 in Pseu-domonas stutzeri AN10. J. Bacteriol. 191:1239 –1247.
14. García-Valdés E, Cozar E, Rotger R, Lalucat J, Ursing J. 1988. Newnaphthalene degrading marine Pseudomonas strains. Appl. Environ. Mi-crobiol. 54:2478 –2485.
15. van de Krundert JA, van Gestel MH, Meerdink G, de Marie S. 1988.Emergence of bacterial resistance to cefamandole in vivo due to outermembrane protein deficiency. Eur. J. Clin. Microbiol. Infect. Dis. 7:776 –778.
16. Williams PA, Murray K. 1974. Metabolism of benzoate and the methyl-benzoates by Pseudomonas putida (arvilla) mt-2: evidence for the exis-tence of a TOL plasmid. J. Bacteriol. 120:416 – 423.
17. Christie-Oleza JA, Nogales B, Martín-Cardona C, Lanfranconi MP,
Albertí S, Lalucat J, Bosch R. 2008. ISPst9, an ISL3-like insertion se-quence from Pseudomonas stutzeri AN10 involved in catabolic gene inac-tivation. Int. Microbiol. 11:101–110.
18. Kovach ME, Phillips RW, Elzer PH, Roop RM, II, Peterson KM. 1994.pBBR1MCS: a broad-host-range cloning vector. Biotechniques 16:800 –802.
19. Elhai J, Wolk CP. 1988. A versatile class of positive-selection vectorsbased on the nonviability of palindrome-containing plasmids that allowscloning into long polylinkers. Gene 68:119 –138.
20. Kaniga K, Delor I, Cornelis GR. 1991. A wide-host-range suicide vectorfor improving reverse genetics in Gram-negative bacteria: inactivation ofthe blaA gene of Yersinia enterocolitica. Gene 109:137–141.
21. Schwartz RD. 1973. Octene epoxidation by a cold-stable alkane-oxidizingisolate of Pseudomonas oleovorans. Appl. Microbiol. 25:574 –577.
22. Aragno M, Schlegel HG. 1981. The hydrogen-oxidizing bacteria, p 865–893. In Starr MP, Stolp H, Trüper HG, Balows A (ed), The prokaryotes: ahandbook on habitats, isolation and identification of bacteria. Springer-Verlag, Berlin, Germany.
23. Sambrook J, Russell DW. 2001. Molecular cloning: a laboratory manual,3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
24. van Beilen JB, Wubbolts MG, Witholt B. 1994. Genetics of alkaneoxidation by Pseudomonas oleovorans. Biodegradation 5:161–174.
25. Eggink G, Lageveen RG, Altenburg B, Witholt B. 1987. Controlled andfunctional expression of the Pseudomonas oleovorans alkane utilizing sys-tem in Pseudomonas putida and Escherichia coli. J. Biol. Chem. 262:17712–17718.
Christie-Oleza et al.
1638 aem.asm.org Applied and Environmental Microbiology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 20
0.12
6.83
.173
.