22nd biennial evergreen international phage...
TRANSCRIPT

22nd Biennial Evergreen International
Phage Meeting
Aug. 6-11, 2017
The Evergreen State College, Olympia, WA

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
22nd Biennial Evergreen International Phage Meeting – Aug. 6-11, 2017
The Evergreen State College, Olympia, WA
Organizing committee: Elizabeth Kutter (chair), Jason Gill, Susan Lehman,
Krystyna Dabrowska, Peter Fineran, Debbie Hinton, Ramy Aziz, Paul Hyman, Dan Nelson, Joana
Azeredo, Bob Blasdel, Martha Clokie, Sandra Morales, and Martha Vives
Daniel Bryan, co-coordinator
Meeting goals: Building on the original Cold Spring Harbor phage meetings, the Evergreen
meeting’s primary goal is to provide an opportunity for members of the ever-broadening phage
community from various backgrounds, ages and places to build strong working relationships and
share skills and ideas, leading to new insights and collaborations and, in recent years, helping spur
on many kinds of applications. People are encouraged to present work that is still in progress.
Posters are strongly encouraged, both as independent presentations with their own abstracts
(grouped by general topic) and as a way of letting participants more closely examine the detailed
data of those giving talks. They remain up throughout the meeting, rather than having specific
presentation times for individual groups of posters.
Note that presenting at this meeting does not constitute a publication. Abstracts will not be
published online, and permission must be obtained from the respective authors before
quoting any of their abstracts or data, or applying it in a way that may be inappropriate.
Thanks to contributors: The Evergreen State College and the Phagebiotics Research Foundtion,
cosponsors and planners; Evergreen grants writer Dorothea Collins; Matthew Stidham, digital
support; the TESC Longhouse crew for taking over our traditional Native American salmon bake;
and the many of you who have helped with meeting planning. Special thanks to the following
organizations for helping support young scholars and participants from underdeveloped countries:
AmpliPhi Biosciences Inc (APHB): AmpliPhi is a biopharmaceutical company focused on the
development of an internally generated pipeline of bacteriophage-based products for human
therapeutic use. The company operates from laboratories in Australia, the USA and Slovenia and
has a production facility exclusively dedicated to manufacture of therapeutic phage products under
current Good Manufacturing Practices (cGMP) standards.
The Capitol Aeroporter has gone from simply being the means by which most participants
reached Evergreen to becoming an active supporter of our work toward phage applications. This
year, they have donated transportation for a number of the participants in our Africa product, as
well as making special efforts to make the trip to Evergreen as positive as possible in terms of
paying attention to the schedules of all of the meeting attendants in making van assignments.
EpiBiome, Inc., based in South San Francisco, CA, is a venture-backed precision microbiome
engineering company that employs a genomics approach to understand complex microbial
communities and deploys bacteriophages to specifically eliminate problematic bacteria while
leaving the rest of the community intact.
Intralytix, Inc., based in Baltimore, has been successfully exploring phage applications to food
safety and human health since the 1990s. Their product targeting Listeria in ready to eat meats and

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
cheeses was the first FDA approved phage product for human ingestion, and they are now
producing approved products for a number of other livestock and human pathogens.
Jafral is a small contract development and manufacturing organization (CDMO) which provides
support with process development, analytical development and manufacturing for bacteriophage
products. They are set up to manufacture phages for almost any purpose, including environmental,
agricultural, food safety, diagnostic and veterinary applications.
Synthetic Genomics: Synthetic Genomics Inc. (SGI) uses their pioneering and proprietary science
and technology to develop products to positively impact the world. From new vaccines and
therapeutics, food and nutritional products, humanized organs for transplant, biofuels, biobased-
chemicals, and agricultural solutions, they are advancing products through their own programs and
with industry leading partners. They are fully leveraging their synthetic biology, genomics,
bioinformatics and viral engineering toolbox to engineer bacteriophage with improved therapeutic
potential against medically important MDR pathogens such as P. aeruginosa.
The Bill and Melinda Gates Foundation, for providing travel support for scientists from various
African countries who are working toward using phage to increase food safety, and also for helping
support the recent 11-day training led by Martha Clokie and Janet Nole at the Makerere University
College of Veterinary Medicine, in which a number of the participants here took part.
Thanks also to the National Institutes of Health and the US Department of Agriculture for their
many years-long history of supporting this collaborative Evergreen meeting through grant funding.
This abstract book is not a publication. Information can not be referenced without explicit permission of the author(s).
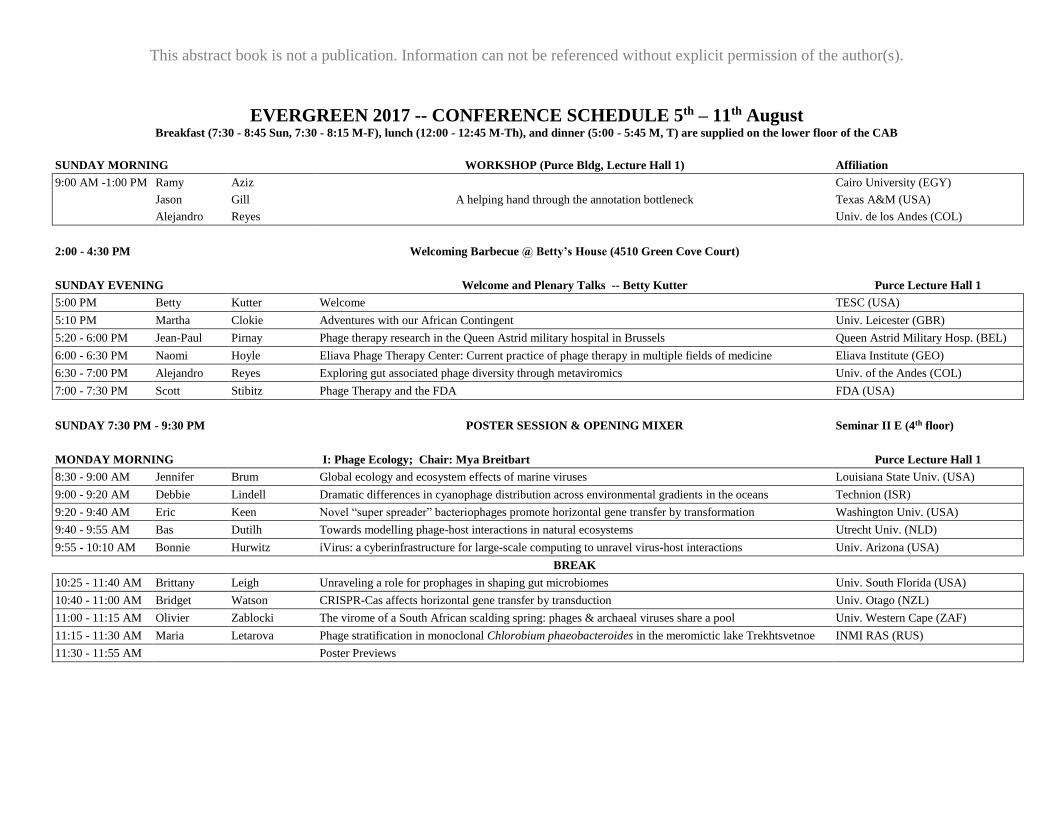
EVERGREEN 2017 -- CONFERENCE SCHEDULE 5th – 11th August Breakfast (7:30 - 8:45 Sun, 7:30 - 8:15 M-F), lunch (12:00 - 12:45 M-Th), and dinner (5:00 - 5:45 M, T) are supplied on the lower floor of the CAB
SUNDAY MORNING
WORKSHOP (Purce Bldg, Lecture Hall 1) Affiliation
9:00 AM -1:00 PM Ramy Aziz Cairo University (EGY)
Jason Gill A helping hand through the annotation bottleneck Texas A&M (USA)
Alejandro Reyes Univ. de los Andes (COL)
2:00 - 4:30 PM
Welcoming Barbecue @ Betty’s House (4510 Green Cove Court)
SUNDAY EVENING
Welcome and Plenary Talks -- Betty Kutter Purce Lecture Hall 1
5:00 PM Betty Kutter Welcome TESC (USA)
5:10 PM Martha Clokie Adventures with our African Contingent Univ. Leicester (GBR)
5:20 - 6:00 PM Jean-Paul Pirnay Phage therapy research in the Queen Astrid military hospital in Brussels Queen Astrid Military Hosp. (BEL)
6:00 - 6:30 PM Naomi Hoyle Eliava Phage Therapy Center: Current practice of phage therapy in multiple fields of medicine Eliava Institute (GEO)
6:30 - 7:00 PM Alejandro Reyes Exploring gut associated phage diversity through metaviromics Univ. of the Andes (COL)
7:00 - 7:30 PM Scott Stibitz Phage Therapy and the FDA FDA (USA)
SUNDAY 7:30 PM - 9:30 PM POSTER SESSION & OPENING MIXER Seminar II E (4th floor)
MONDAY MORNING
I: Phage Ecology; Chair: Mya Breitbart Purce Lecture Hall 1
8:30 - 9:00 AM Jennifer Brum Global ecology and ecosystem effects of marine viruses Louisiana State Univ. (USA)
9:00 - 9:20 AM Debbie Lindell Dramatic differences in cyanophage distribution across environmental gradients in the oceans Technion (ISR)
9:20 - 9:40 AM Eric Keen Novel “super spreader” bacteriophages promote horizontal gene transfer by transformation Washington Univ. (USA)
9:40 - 9:55 AM Bas Dutilh Towards modelling phage-host interactions in natural ecosystems Utrecht Univ. (NLD)
9:55 - 10:10 AM Bonnie Hurwitz iVirus: a cyberinfrastructure for large-scale computing to unravel virus-host interactions Univ. Arizona (USA)
BREAK
10:25 - 11:40 AM Brittany Leigh Unraveling a role for prophages in shaping gut microbiomes Univ. South Florida (USA)
10:40 - 11:00 AM Bridget Watson CRISPR-Cas affects horizontal gene transfer by transduction Univ. Otago (NZL)
11:00 - 11:15 AM Olivier Zablocki The virome of a South African scalding spring: phages & archaeal viruses share a pool Univ. Western Cape (ZAF)
11:15 - 11:30 AM Maria Letarova Phage stratification in monoclonal Chlorobium phaeobacteroides in the meromictic lake Trekhtsvetnoe INMI RAS (RUS)
11:30 - 11:55 AM Poster Previews
This abstract book is not a publication. Information can not be referenced without explicit permission of the author(s).
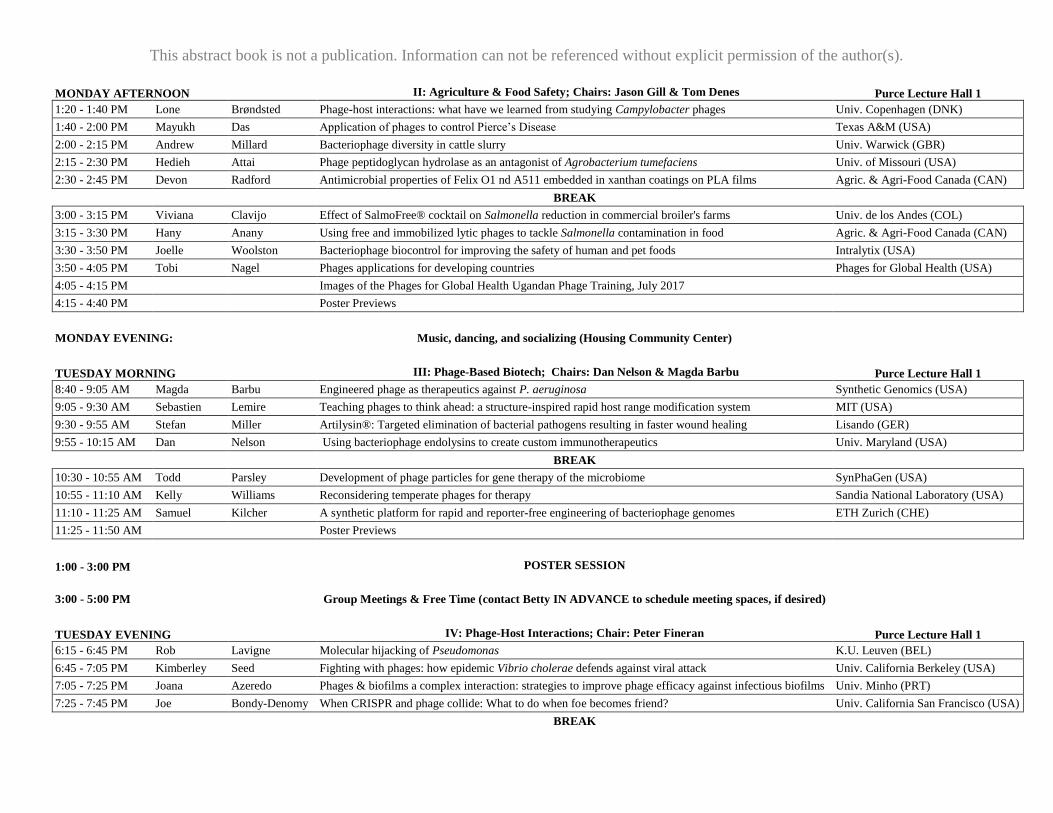
MONDAY AFTERNOON
II: Agriculture & Food Safety; Chairs: Jason Gill & Tom Denes Purce Lecture Hall 1
1:20 - 1:40 PM Lone Brøndsted Phage-host interactions: what have we learned from studying Campylobacter phages Univ. Copenhagen (DNK)
1:40 - 2:00 PM Mayukh Das Application of phages to control Pierce’s Disease Texas A&M (USA)
2:00 - 2:15 PM Andrew Millard Bacteriophage diversity in cattle slurry Univ. Warwick (GBR)
2:15 - 2:30 PM Hedieh Attai Phage peptidoglycan hydrolase as an antagonist of Agrobacterium tumefaciens Univ. of Missouri (USA)
2:30 - 2:45 PM Devon Radford Antimicrobial properties of Felix O1 nd A511 embedded in xanthan coatings on PLA films Agric. & Agri-Food Canada (CAN)
BREAK
3:00 - 3:15 PM Viviana Clavijo Effect of SalmoFree® cocktail on Salmonella reduction in commercial broiler's farms Univ. de los Andes (COL)
3:15 - 3:30 PM Hany Anany Using free and immobilized lytic phages to tackle Salmonella contamination in food Agric. & Agri-Food Canada (CAN)
3:30 - 3:50 PM Joelle Woolston Bacteriophage biocontrol for improving the safety of human and pet foods Intralytix (USA)
3:50 - 4:05 PM Tobi Nagel Phages applications for developing countries Phages for Global Health (USA)
4:05 - 4:15 PM Images of the Phages for Global Health Ugandan Phage Training, July 2017
4:15 - 4:40 PM Poster Previews
MONDAY EVENING:
Music, dancing, and socializing (Housing Community Center)
TUESDAY MORNING
III: Phage-Based Biotech; Chairs: Dan Nelson & Magda Barbu Purce Lecture Hall 1
8:40 - 9:05 AM Magda Barbu Engineered phage as therapeutics against P. aeruginosa Synthetic Genomics (USA)
9:05 - 9:30 AM Sebastien Lemire Teaching phages to think ahead: a structure-inspired rapid host range modification system MIT (USA)
9:30 - 9:55 AM Stefan Miller Artilysin®: Targeted elimination of bacterial pathogens resulting in faster wound healing Lisando (GER)
9:55 - 10:15 AM Dan Nelson Using bacteriophage endolysins to create custom immunotherapeutics Univ. Maryland (USA)
BREAK
10:30 - 10:55 AM Todd Parsley Development of phage particles for gene therapy of the microbiome SynPhaGen (USA)
10:55 - 11:10 AM Kelly Williams Reconsidering temperate phages for therapy Sandia National Laboratory (USA)
11:10 - 11:25 AM Samuel Kilcher A synthetic platform for rapid and reporter-free engineering of bacteriophage genomes ETH Zurich (CHE)
11:25 - 11:50 AM Poster Previews
1:00 - 3:00 PM
POSTER SESSION
3:00 - 5:00 PM
Group Meetings & Free Time (contact Betty IN ADVANCE to schedule meeting spaces, if desired)
TUESDAY EVENING
IV: Phage-Host Interactions; Chair: Peter Fineran Purce Lecture Hall 1
6:15 - 6:45 PM Rob Lavigne Molecular hijacking of Pseudomonas K.U. Leuven (BEL)
6:45 - 7:05 PM Kimberley Seed Fighting with phages: how epidemic Vibrio cholerae defends against viral attack Univ. California Berkeley (USA)
7:05 - 7:25 PM Joana Azeredo Phages & biofilms a complex interaction: strategies to improve phage efficacy against infectious biofilms Univ. Minho (PRT)
7:25 - 7:45 PM Joe Bondy-Denomy When CRISPR and phage collide: What to do when foe becomes friend? Univ. California San Francisco (USA)
BREAK
This abstract book is not a publication. Information can not be referenced without explicit permission of the author(s).
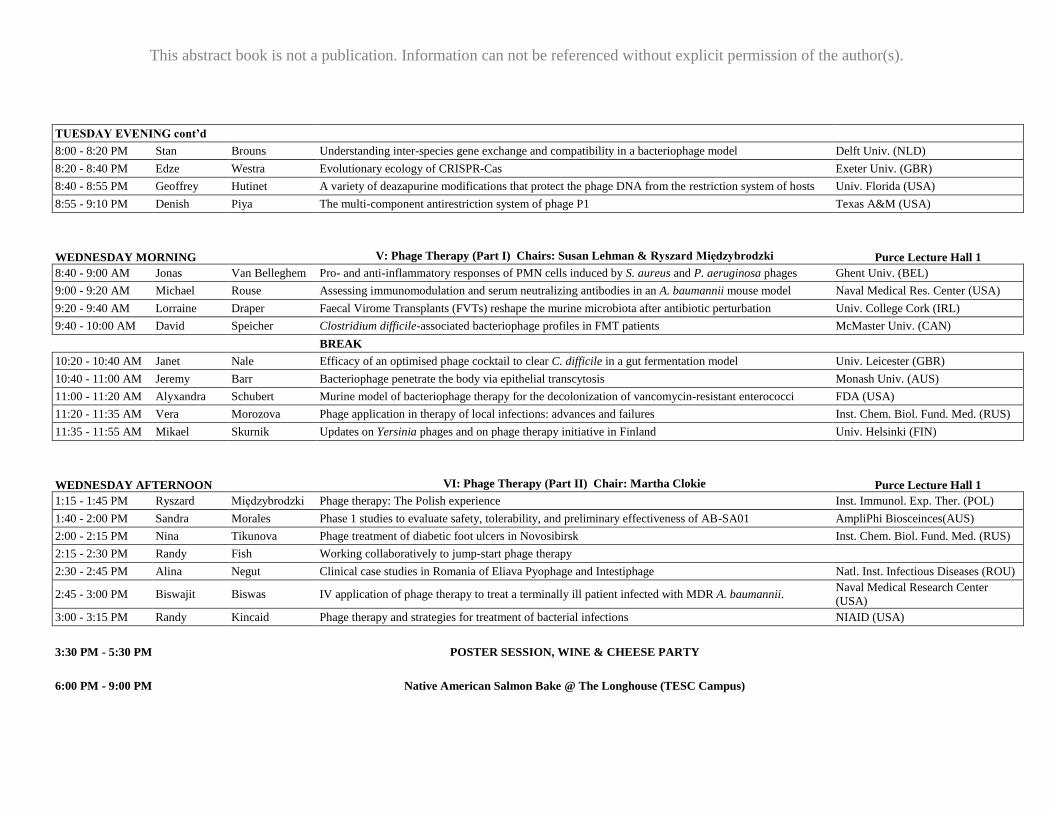
TUESDAY EVENING cont’d
8:00 - 8:20 PM Stan Brouns Understanding inter-species gene exchange and compatibility in a bacteriophage model Delft Univ. (NLD)
8:20 - 8:40 PM Edze Westra Evolutionary ecology of CRISPR-Cas Exeter Univ. (GBR)
8:40 - 8:55 PM Geoffrey Hutinet A variety of deazapurine modifications that protect the phage DNA from the restriction system of hosts Univ. Florida (USA)
8:55 - 9:10 PM Denish Piya The multi-component antirestriction system of phage P1 Texas A&M (USA)
WEDNESDAY MORNING
V: Phage Therapy (Part I) Chairs: Susan Lehman & Ryszard Międzybrodzki Purce Lecture Hall 1
8:40 - 9:00 AM Jonas Van Belleghem Pro- and anti-inflammatory responses of PMN cells induced by S. aureus and P. aeruginosa phages Ghent Univ. (BEL)
9:00 - 9:20 AM Michael Rouse Assessing immunomodulation and serum neutralizing antibodies in an A. baumannii mouse model Naval Medical Res. Center (USA)
9:20 - 9:40 AM Lorraine Draper Faecal Virome Transplants (FVTs) reshape the murine microbiota after antibiotic perturbation Univ. College Cork (IRL)
9:40 - 10:00 AM David Speicher Clostridium difficile-associated bacteriophage profiles in FMT patients McMaster Univ. (CAN)
BREAK
10:20 - 10:40 AM Janet Nale Efficacy of an optimised phage cocktail to clear C. difficile in a gut fermentation model Univ. Leicester (GBR)
10:40 - 11:00 AM Jeremy Barr Bacteriophage penetrate the body via epithelial transcytosis Monash Univ. (AUS)
11:00 - 11:20 AM Alyxandra Schubert Murine model of bacteriophage therapy for the decolonization of vancomycin-resistant enterococci FDA (USA)
11:20 - 11:35 AM Vera Morozova Phage application in therapy of local infections: advances and failures Inst. Chem. Biol. Fund. Med. (RUS)
11:35 - 11:55 AM Mikael Skurnik Updates on Yersinia phages and on phage therapy initiative in Finland Univ. Helsinki (FIN)
WEDNESDAY AFTERNOON VI: Phage Therapy (Part II) Chair: Martha Clokie Purce Lecture Hall 1
1:15 - 1:45 PM Ryszard Międzybrodzki Phage therapy: The Polish experience Inst. Immunol. Exp. Ther. (POL)
1:40 - 2:00 PM Sandra Morales Phase 1 studies to evaluate safety, tolerability, and preliminary effectiveness of AB-SA01 AmpliPhi Biosceinces(AUS)
2:00 - 2:15 PM Nina Tikunova Phage treatment of diabetic foot ulcers in Novosibirsk Inst. Chem. Biol. Fund. Med. (RUS)
2:15 - 2:30 PM Randy Fish Working collaboratively to jump-start phage therapy
2:30 - 2:45 PM Alina Negut Clinical case studies in Romania of Eliava Pyophage and Intestiphage Natl. Inst. Infectious Diseases (ROU)
2:45 - 3:00 PM Biswajit Biswas IV application of phage therapy to treat a terminally ill patient infected with MDR A. baumannii. Naval Medical Research Center
(USA)
3:00 - 3:15 PM Randy Kincaid Phage therapy and strategies for treatment of bacterial infections NIAID (USA)
3:30 PM - 5:30 PM
POSTER SESSION, WINE & CHEESE PARTY
6:00 PM - 9:00 PM
Native American Salmon Bake @ The Longhouse (TESC Campus)

This abstract book is not a publication. Information can not be referenced without explicit permission of the author(s).
THURSDAY MORNING
VII: Molecular Biology; Chair: Debbie Hinton Purce Lecture Hall 1
8:40 - 9:10 AM Bob Blasdel The implications of the dynamic interchange between killing a cell and letting it hang itself for picking the
right phages for therapy K.U. Leuven (BEL)
9:10 - 9:25 AM Debbie Hinton T4 MotA contains a novel DNA binding motif that specifically recognizes modified DNA NIH (USA)
9:25 - 9:45 AM Jennifer West The bacteriophage T4 MotB protein, a DNA binding protein, boosts the level of T4 late gene expression NIH (USA)
9:45 - 10:05 AM Scott Nelson The regulation and mechanism of the bacteriophage T4 MR complex (gp46/47) Iowa State Univ. (USA)
10:05 - 10:20 AM Betty Kutter Hot tales of T4’s transition from host to phage metabolism TESC (USA)
BREAK
10:40 - 10:55 AM Angela Makumi Superinfection exclusion systems as molecular drivers of lactococcal 936 phage evolution Univ. College Cork (IRL)
10:55 - 11:15 AM Nikolai Prokhorov Host recognition by podoviruses G7C and Alt Univ. Texas Medical Branch (USA)
11:15 - 11:30 AM Ines Staes Phages that farm: insights into the phage-carrier state dynamics K.U. Leuven (BEL)
11:30 - 11:45 AM Lanying Zeng Virus interactions inside the cell: competition or cooperation? Texas A&M (USA)
11:45 - 12:00 AM Junjie Zhang Asymmetric cryo-EM structure of the canonical Allolevivirus Qβ reveals a single maturation protein and
the genomic ssRNA in situ Texas A&M (USA)
1:00 - 3:00 PM
FINAL POSTER SESSION
THURSDAY AFTERNOON
VIII: Open Section; Chair: Paul Hyman Purce Lecture Hall 1
3:00 - 3:20 PM Krystyna Dabrowska Review of factors determining phage penetration in animals and humans, 1940-2016 Inst. Immunol. Exp. Ther. (POL)
3:20 - 3:35 PM Rachel Chang Anti-pseudomonal activity of dry powder PEV20 phage in mouse lung infection Univ. Sydney, (AUS)
3:35 - 3:50 PM Cas Mosterd CRISPR-Cas and phage-host interactions in Streptococcus mutans Laval Univ. (CAN)
3:50 - 4:10 PM Sydney Hayes Mucosal vaccine delivery for peptide antigens via lambda display phage Univ. Saskatchewan (CAN)
4:10 - 4:25 PM Dwayne Roach Immunophage synergy Pasteur Inst. (FRA)
4:25 - 4:40 PM Marcin Los Phage for construction of biosensors Phage Consultants (POL)
4:40 - 4:55 PM Amelia McKitterick Elucidating the specificity determinants of anti-phage genomic island excision in Vibrio cholera Univ. California Berkeley (USA)
4:55 - 5:10 PM Paul Hyman Is more better? Use of multiple hosts in phage isolation Ashland Univ. (USA)
5:10 - 5:20 PM Betty Kutter Final words, looking forward TESC (USA)
THURSDAY EVENING
FREE EVENING -- DINNER IN TOWN
Dancing and Socializing in the Housing Community Center
FRIDAY
8:45 AM: 2 Buses: trip to Mt Rainier (meet in dorm loop)
Timing may be extended by taking private cars.
Ride direct to airport for night flights is possible.
EVENING: Housing Community Center available

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
ORAL PRESENTATION ABSTRACTS
Abstracts are arranged in the order they appear on the presentation schedule. Sections are
as follows:
Sunday Genomics & Annotation Workshop
Plenary Talks
Section I: Phage Ecology
Section II: Agriculture & Food Safety
Section III: Phage-Based Biotech
Section IV: Phage-Host Interactions
Section V: Phage Therapy (Part I)
Section VI: Phage Therapy (Part II)
Section VII: Molecular Mechanisms
Section VIII: Open Session
The presenting author’s name appears in bold text. The names of other authors who are
attending the meeting have been underlined.
E-mail addresses are provided for the corresponding author, which may or may not be the
same as the presenting author.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sunday Workshop 9
From Sequence to Knowledge: Assembly, Annotation, and Analysis
of Phage Genomes from Genomic and Metagenomic Data Sets
Alejandro Reyes1, Jason Gill2, Ramy K. Aziz3
1 Department of Biological Sciences, Universidad de los Andes, Bogotá, Colombia.
2 Department of Animal Science, Texas A&M University, College Station, TX
3 Faculty of Pharmacy, Cairo University, Egypt and member of the SEED/PATRIC team
Bios:
● Alejandro Reyes is an Associate professor at the Department of Biological Sciences,
School of Sciences, Universidad de los Andes, Bogotá, Colombia. He is also Adjunct
Assistant Professor at the Department of Pathology and Immunology, Washington
University School of Medicine, St Louis, MO, and the Group leader of the Max Planck
tandem group in Computational Biology, Universidad de los Andes, Bogotá, Colombia.
● Jason Gill is an Assistant Professor in the Department of Animal Science and the Faculty
of Genetics at Texas A&M University, College Station, Texas USA. He is also Associate
Director of the Center for Phage Technology (CPT) at Texas A&M, a state-funded center
devoted to the study of phages for applications in human health and agriculture.
● Ramy Aziz is a Professor and Acting Chair at the Department of Microbiology and
Immunology, Faculty of Pharmacy, Cairo University, Egypt. He is also a member of the
SEED/PATRIC team at the Argonne National Lab (where RAST is hosted) and has
formerly been involved with the PhAnToMe project for phage genome annotation.
Unprecedented advances in genome sequencing improved sequence quality and accuracy,
increased throughput, while decreasing cost and time. This sequencing revolution resulted in tens
of thousands of bacterial genomes that are more or less fully sequenced. Surprisingly, although
phages are an order of magnitude more abundant than bacteria, and although their genomes are
smaller and easier to sequence, fewer phage genomes are in public databases, and consequently
bacteriophage biodiversity is undersampled.
Currently, hundreds of phages are being sequenced from genomes and metagenomes, but
more sequences are needed, and more phage-specific tools are being developed to fill the gap in
phage sequence assembly, interpretation, annotation, and comparative genomics.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
10 Sunday Workshop
This year, the “Phage Genomics Workshop” will have a dual focus: (i) Annotation of assembled
phage genome sequences (from isolated phages); (ii) Analysis, Assembly, and Annotation of
phages from environmental or human-associated metagenomes/microbiomes.
Optional hands-on and problem-solving activities will be suggested.
Major toolkits that will be demonstrated in this workshop:
- PhAnToMe (Phage Annotation Tools and Methods) developed by the laboratory of
Robert Edwards at SDSU in collaboration with Mya Breitbart, Jeff Elhai, and Matt
Sullivan (http://www.phantome.org)
- The SEED and RAST toolkits, including the latest development: RAST toolkit (RAST-
tk), developed by an international team lead by the Argonne National Laboratory, IL, USA
(http://www.theseed.org)
- The CPT deployments of Galaxy and WebApollo, which are customized for the analysis
and annotation of whole phage genomes. This system is still under development and
supports customizable workflows to facilitate many kinds of DNA and protein sequence
analysis. (https://cpt.tamu.edu/galaxy-pub/)
- GenSeed-HMM, developed by the laboratory of Arthur Gruber at USP, Brazil, in
collaboration with Alejandro Reyes and Alan Durham. This package is designed for
targeted assembly from metagenomes using specific motifs (represented as HMMs) from
variable proteins.
Links:
Tool or Database URL
RAST http://rast.nmpdr.org
RASTtk To download: https://github.com/TheSEED/RASTtk-Distribution/releases/
For tutorials: http://tutorial.theseed.org
MyRAST http://blog.theseed.org/servers/installation/distribution-of-the-seed-server-
packages.html
Download: http://blog.theseed.org/downloads/myRAST-Intel.dmg
Prokka http://www.vicbioinformatics.com/software.prokka.shtml
phAST http://www.phantome.org/PhageSeed/Phage.cgi?page=phast
BASys http://basys.ca/
GenSAS v3.0 http://gensas.bioinfo.wsu.edu/
IGS Prokaryotic Annotation
Pipeline
http://www.igs.umaryland.edu/research/bioinformatics/analysis/whole_ge
nome.php
MAKER Web Annotation
Service (MWAS)
http://www.yandell-lab.org/software/mwas.html
GenSEED HMM https://sourceforge.net/projects/genseedhmm/
Copies of the workshop presentations and related tutorials can be found online:
http://bit.ly/phigenomics2017; http://bit.ly/phantome2017; http://egybio.net/tutorial/

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Plenary Talks 11
Phage therapy research in the Queen Astrid military hospital in Brussels
Jean-Paul Pirnay
Queen Astrid Military Hospital, Brussels, Belgium
The worldwide emergence of “superbugs” and a dry pipeline for new antibiotics threaten modern
medicine with a return to the pre antibiotic era. Phages - the viruses of bacteria - could help fight
antimicrobial resistance. In 2003, a first phage therapy related study proposal was submitted to the
R&D department of Belgian Defense. It was dismissed as mere “science fiction” with a score of
4/20. Today, phage therapy research has become commonplace in the Queen Astrid military
hospital and encompasses different aspects:
i) The isolation, selection and characterization of candidate therapeutic phages active against
clinically important pathogens such as Acinetobacter baumannii, which is often associated with
military operations in the Middle East (PMID: 25111143), Pseudomonas aeruginosa,
Staphylococcus aureus, Klebsiella pneumoniae and Escherichia coli, including the O104:H4 strain
from the 2011 foodborne EAHEC outbreak in Germany (PMID: 23285164).
ii) Clinical trials:
A small clinical safety study (PMID: 25356373): 10 applications of phage cocktail BFC 1
(PMID: 19300511), active against P. aeruginosa and S. aureus, in burn wound infections.
PhagoBurn (www.phagoburn.eu), funded by the European Commission: Evaluating phage
therapy for the treatment of burn wounds, infected with E. coli and P. aeruginosa, through
a randomized controlled trial (results pending).
iii) Study of the bacterium-phage (host-parasite) relationship, with an emphasis on bacterial phage
resistance evolution and the development of adequate treatment protocols (PMID: 22660719,
PMID: 26476097).
iv) Elaboration of a dedicated regulatory framework for phage therapy (www.P-H-A-G-E.org)
(PMID: 21063753).
v) Development of realistic production and QC/QA regimens for therapeutic phage products
(PMID: 25585954).
In the margin of these studies, and under the umbrella of article 37 (unproven interventions) of
“The Declaration of Helsinki,” eleven patients with multidrug resistant infections were treated with
phages in the Queen Astrid military hospital.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
12 Plenary Talks
Eliava Phage Therapy Center: Current Practice of Phage Therapy in Multiple
Fields of Medicine
N. Hoyle*, L. Nadareishvili, P. Zhvania, L. Pipia, G. Khvichia, I. Tedoradze, N. Odishelidze, N.
Pruidze, D. Nizharadze
Eliava Phage Therapy Center, Tbilisi, Republic of Georgia 0160
*E-mail: [email protected]
Since Felix d’Herelle and George Eliava co-founded it in the 1930’s, what is now the Eliava
Institute has developed and produced therapeutic phage cocktails, building on the broad
explorations the two began at the Pasteur Institute 100 years ago. The current Eliava
Biopreparations LLD makes commercial cocktails such as Pyophage and Intestiphage which are
used throughout Georgia as a first line of defense where appropriate. The Institute also prepares
individual phages from their vast collection when called for by clinical testing. They have always
worked very closely with many Georgian clinics, hospitals and individual physicians and have their
own pharmacy. However, until a few years ago, we did not have our own clinic, specifically
focused on conditions where the use of phage can be particularly beneficial and sometimes using
modern tools to measure key parameters. Indeed, this Phage Therapy Center is now able to
leverage the Eliava Institute’s significant collection of phage as well as its extensive scientific
experience working with phage, collecting crucial clinical data to inform future formal trials.
In our clinical practice, we have gained unique experience in using phage therapy to treat various
infectious complications of genetic diseases, as well as other antibiotic resistant and refractory
infections. As allopathic physicians, we understand the importance of objective examination and
practice according to evidence based medicine. While the need for large scale clinical trials
(RCT’s) is indisputable, phage therapy is complex and diverse enough that choosing the right
conditions for a double-blind approach has proven very tricky in major trials to date, while demand
for an alternative to complement current approaches is ever increasing, as can be seen through our
broad patient and physician contacts. Patients from countries all over the world travel to our clinic
for treatment with phage therapy. This treatment is not considered experimental by the Georgian
Ministry of Health, and phage preparations are registered drugs in Georgia.
Many patients have chronic infections which have been refractory to antibiotic therapy. Infection
control and prevention is a priority for patients with genetic conditions such as cystic fibrosis as
well as conditions such as diabetes that predispose to infections that are very difficult to treat.
Other frequent problems treated at our center include chronic bacterial prostatitis, chronic sinusitis,
lung infections, irritable bowel syndrome, and infected wounds. Pre-operative elimination of
antibiotic resistant bacteria in patient carriers is another useful application. We would like to share
our experiences via a few case reports which represent interesting and important outcomes in phage
therapy. Some involve straightforward use of the standard commercial Eliava cocktails, while
others have required preparation of appropriate phage from Institute collections or fresh isolation of
phage specifically targeting the patient’s bacteria, through the mediation of the Eliava’s other
research laboratories and subsidiary companies.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Plenary Talks 13
Exploring gut associated phage diversity through metaviromics
Alejandro Reyes
Department of Biological Sciences, Max Planck tandem group in Computational Biology,
Universidad de los Andes, Bogotá, Colombia;
Center for Genome Sciences and Systems Biology, Washington University School of Medicine, St
Louis, MO, USA
E-mail: [email protected]
Recent advances studying the microbial ecology of different environments have shown that any
microbial environment is not fully characterized until we understand its viral component. Viruses
are the most abundant and diverse biological entity found in any microbial environment, however,
their function and role on any of those environments is far from being elucidated. Current advances
in DNA sequencing has allowed for the first time, the characterization of the viral component
(virome) associated to environments without requiring culturing the cellular component. However,
to date, our capacity to annotate and understand the functional potential of the virome is limited
due to the minute component of viral diversity fully characterized in public databases and our
restriction to computational tools based on sequence comparison. Recently we surveyed over 140
samples originated from 59 individuals of 22 families, sequencing effort reached an average of
46,000 reads per sample. However, only 30% had significant similarity to public databases queried.
De novo assembly of the reads allowed the usage of 92.3% of the reads in a total of 16,766 contigs,
some of those reaching over 150,000 nucleotides. Even though it is widely known that with longer
reads (or contigs) more chances of identifying a significant similarity in databases, still less than
50% of the contigs showed significant similarity with known sequences. In parallel, looking for
novel strategies for contig annotation that go beyond sequence similarity, we generated clusters of
orthologous phage and viral domains (ViPhOGs), that regardless of a known function associated,
can be used with high sensitivity and specificity as classifiers for different viral taxonomical levels.
Furthermore, coupling the ViPhOGs with the generation of pHMM constitute a tool capable of
identifying and annotating far more viral diversity than previously possible. This methodology,
together with machine learning algorithms such as Random Forest, opens a way to identify
signature viral domains from viral metagenomic studies increasing our capacity to characterize and
annotate that huge challenge of the viral dark matter.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
14 Plenary Talks
Phage Therapy and the FDA
Scott Stibitz
US Food and Drug Administration
This presentation is intended to give an overview of the US FDA’s regulatory approach
towards bacteriophage therapy. It will provide a brief description of the FDA’s regulatory
organization, and describe which division is responsible for regulation of products for
Phage Therapy. The presentation will also provide an overview of standard FDA
biologics development pathways with regards to investigational new drugs (IND), and
discuss FDA’s expanded access (compassionate use) programs. Finally, chemistry,
manufacturing, and control (CMC) issues specific to bacteriophage will be covered, along
with FDA’s expectations for characterization of phage for use in clinical trials

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section I: Phage Ecology 15
Global Ecology and Ecosystem Effects of Marine Viruses
Jennifer Brum
Louisiana State University, Baton Rouge, LA
E-mail: [email protected]
Marine viruses have important roles in microbial mortality, gene transfer, metabolic
reprogramming and biogeochemical cycling. However, methodological limitations have
previously prevented a quantitative assessment of their community structure and ecosystem
impacts. Recent transformative advances have enabled quantitative assessments of environmental
viral communities using metagenomic techniques, facilitating a rapid increase in knowledge of
environmental viral ecology and their impacts on ecosystem function. In this presentation, I will
focus on how we are using advanced metagenomic-based analyses from global-scale datasets to
connect viruses with ecosystem function, including an exploration of the roles of marine viruses in
the oceanic carbon cycle.
.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
16 Section I: Phage Ecology
Dramatic differences in cyanophage distribution patterns
across environmental gradients in the oceans Debbie Lindell, Ilia Maidanik, Nava Baran, Michael Carlson, Shay Kirzner, Irena Pekarsky, Sveta
Goldin
Faculty of Biology, Technion – Israel Institute of Technology, Haifa, Israel.
*E-mail:[email protected]
Viruses are globally abundant and extremely diverse in their genetic make-up and in the hosts they
infect. They influence the abundance, diversity and evolution of their hosts as well as the
biogeochemical cycling of matter in the oceans. Yet current methods are inadequate for gaining a
quantitative understanding of their impact on these processes. Here, we employ a new culture-
independent, single phage, solid-phase PCR method, polonies, to gain the first quantitative view of
cyanophages at the phage family level. Dramatic differences in abundances and distribution
patterns were found for different cyanophage families along a transect traversing large
environmental gradients in the North Pacific Ocean as well as over the seasonal cycle in the Red
Sea. In addition, stark differences in the abundances of different lineages (clades) of T7-like
cyanophages were found in both oceanic regions: clade B T7-like cyanophages that encode host-
derived photosynthesis genes and infect either Synechococcus or Prochlorococcus hosts were
considerably more abundant than clade A T7-like cyanophages that lack these photosynthesis
genes and primarily infect Synechococcus hosts. Intriguingly, laboratory studies show that the more
abundant clade B phages have a longer lytic cycle, are less virulent and produce fewer phages per
burst than the clade A phages. These findings highlight that phylogenetic diversity within a single
phage family reflect biologically and ecologically meaningful differences.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section I: Phage Ecology 17
Novel “super spreader” bacteriophages promote horizontal gene transfer by
transformation
Eric Keen*, Valery Bliskovsky, Francisco Malagon, James Baker, Jeffrey Prince, James
Klaus, and Sankar Adhya
*Center for Genome Sciences, Washington University in St. Louis, St. Louis, MO 63108
E-mail: [email protected]
Bacteriophages infect an estimated 1023 to 1025 bacterial cells each second, many of which carry
physiologically relevant plasmids (e.g., those encoding antibiotic resistance). However, even
though phage-plasmid interactions occur on a massive scale and have potentially significant
evolutionary, ecological, and biomedical implications, plasmid fate upon phage infection and lysis
has not been investigated to date. Here we show that a subset of the natural lytic phage population,
which we dub “superspreaders,” releases substantial amounts of intact, transformable plasmid
DNA upon lysis, thereby promoting horizontal gene transfer by transformation. Two novel
Escherichia coli phage superspreaders, SUSP1 and SUSP2, liberated four evolutionarily distinct
plasmids with equal efficiency, including two close relatives of prominent antibiotic resistance
vectors in natural environments. SUSP2 also mediated the extensive lateral transfer of antibiotic
resistance in unbiased communities of soil bacteria from Maryland and Wyoming. Furthermore, the
addition of SUSP2 to cocultures of kanamycin-resistant E. coli and kanamycin-sensitive Bacillus
simplex resulted in roughly 1,000-fold more kanamycin-resistant Bacillus simplex than arose in
phage-free controls. Unlike many other lytic phages, neither SUSP1 nor SUSP2 encodes homologs
to known hydrolytic endonucleases, suggesting a simple potential mechanism underlying the
superspreading phenotype. Consistent with this model, the deletion of endonuclease IV and the
nucleoid-disrupting protein ndd from coliphage T4, a phage known to extensively degrade
chromosomal DNA, significantly increased its ability to promote plasmid transformation. Taken
together, our results suggest that phage superspreaders may play key roles in microbial evolution
and ecology but should be avoided in phage therapy and other medical applications.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
18 Section I: Phage Ecology
Towards modelling phage-host interactions in natural ecosystems
Bas Dutilh
Utrecht University, The Netherlands
Phages are diverse and abundant in all ecosystems and influence microbial community dynamics.
As metagenomics is unveiling the composition of microbial ecosystems at an unprecedented rate, a
major remaining question is whether we understand enough of phage-host interactions to model
microbial community dynamics using an eco-systems biology approach. To do this, we should first
accurately link phages to their hosts using metagenomic data, a question that implies several others:
(i) How many hosts can a phage infect, how stable are phage-host interactions in evolution, and at
what taxonomic level can phage-host interactions be predicted the best? (ii) What aspects of the
phage life cycle leave measurable signals in the respective genome sequences, and which genomic
signals are the strongest predictors of phage-host interaction? Next, given that phage-host
interactions can be predicted, what dynamics do we see, and how can they be explained? Based on
the methods we developed to address the questions above, I will discuss a recent example showing
initially counter-intuitive dynamics, that left us with more questions than answers in the quest for a
mechanistic model of phages in microbial ecosystems.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section I: Phage Ecology 19
iVirus: a cyberinfrastructure for large-scale computing to unravel virus-host
interactions
Bonnie Hurwitz
University of Arizona, Tuscon, AZ
E-mail: [email protected]
Currently, a number of tools exist for analyzing viromes and finding viruses in both bacterial
genomes and metagenomes. Yet, most of these tools are available through disparate website or
code repositories that can be challenging to install and/or are not optimized for high performance
computing to meet scale of modern virome and microbiome datasets. To enhance reproducibility
and methods development we are building iVirus, a data repository and optimized high-
performance computing infrastructure for elucidating viral-host interactions. iVirus is built on the
existing CyVerse cyberinfrastructure and provides tools, data and metadata resources specific to
virus ecology. Over the next year, the iVirus project is adding new metadata search capabilities for
users to discover data in the iVirus Data Commons, add these data (and/or their own private data)
to a shopping cart, and launch CyVerse tools (built by the iVirus team or others) directly from the
iVirus site. All analyses will be output to the user’s private CyVerse account, and accessible either
through CyVerse or in the iVirus “sandbox”. The iVirus website will become a central hub for
discovering massive viral ‘omics datasets stored in CyVerse, and running high-performance
computing analyses at CyVerse using XSEDE high-performance computing resources.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
20 Section I: Phage Ecology
Unraveling a role for prophages in shaping gut microbiomes
Brittany A. Leigh1, Zachary Graham2, Assunta Liberti3, Mya Breitbart1* and Larry J. Dishaw3
1. University of South Florida, College of Marine Sciences, 2. St Petersburg College, 3.
University of South Florida, College of Medicine, Department of Pediatrics
*E-mail: mya@usf@edu
Phages play important roles in shaping microbial communities. Phages integrated into bacterial
genomes are predicted to play equally important roles, however, to date very little is known about
how either influence in vivo systems, primarily due to a lack of tractable model systems. Here, we
present evidence that the recently developed model system for gut host-microbial interactions,
Ciona intestinalis, can provide information on the role that phages play in shaping the outcome of
colonization and in influencing homeostasis in gut microbiomes. As a protochordate, Ciona
possesses only an innate immune system, and sequencing has shown that it maintains a core
community of both bacteria and viruses in the gut. In addition, metavirome and 16S rDNA
amplicon sequencing also reveals compartmentalization (stomach, midgut, hindgut) of distinct
communities. We have successfully cultured approximately 80 bacteria that encompass 9 of the 13
core families and tested each isolate for inducible prophages using mitomycin C. At least 1/3 of the
cultured bacteria contain inducible prophages, and the number of lysogens in vivo could be even
higher. The morphology of each of the induced phages was determined using TEM and their
complete genomes were sequenced and annotated. In vitro assays demonstrate an increase in the
formation of biofilms by some lysogens in the presence of a Ciona secretory immune protein, the
Variable region-containing Chitin Binding Protein (VCBP) known to directly interact with
bacteria; this outcome appears to be coupled to the induction of virus-like particles. Each of the
induced prophages was also assessed for lytic activity against all other cultured isolates. We found
that a number of the induced prophages are capable of infecting other members of the Ciona
microbiome, suggesting that induced prophages have the capacity to shape bacterial community
structure within an animal. Furthermore, preliminary in vivo experiments also suggest that
colonization of germ-free Ciona with a single bacterial lysogen results in increased immune
activity and subsequent prophage induction early in the colonization process. Thus, Ciona affords
the unique opportunity to dissect the complex tripartite dynamics involving host, bacterial, and
phage interactions during colonization of mucosal surfaces.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section I: Phage Ecology 21
CRISPR-Cas affects horizontal gene transfer by transduction
Bridget N. J. Watson1, Raymond H. J. Staals1,† and Peter C. Fineran1,2*
1Department of Microbiology and Immunology, University of Otago, PO Box 56, Dunedin 9054,
New Zealand; 2Bio-Protection Research Centre, University of Otago, PO Box 56, Dunedin 9054,
New Zealand; †Present Address: Laboratory of Microbiology, Department of Agrotechnology and
Food Sciences, Wageningen University, 6708 WE Wageningen, The Netherlands.
*E-mail: [email protected]
A powerful contributor to prokaryotic evolution is horizontal gene transfer (HGT) through
transformation, conjugation and transduction. Gene gain can be advantageous, neutral or
detrimental to bacterial fitness. Bacteria and archaea control both HGT and phage infection through
CRISPR-Cas adaptive immunity. Although the beneficial effects from resisting phage infection are
evident, the net result of CRISPR-Cas on HGT is currently enigmatic, due to the lack of detectable
effects over evolutionary timescales. The established ability of CRISPR-Cas to limit HGT through
conjugation and transformation has even been considered an evolutionary downside. However, the
role of CRISPR-Cas in controlling transduction is largely overlooked. Transduction is the phage-
mediated transfer of bacterial DNA between cells and arguably has the greatest impact on HGT.
Here we addressed the role of the Pectobacterium atrosepticum type I-F CRISPR-Cas system on
horizontal gene transfer via generalised transduction. By assaying the effects of spacers with
different targets, we reach an unexpected conclusion regarding the role of CRISPR-Cas on the
transduction of plasmids and chromosomal loci.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
22 Section I: Phage Ecology
The virome of a South African scalding spring: bacteriophages and archaeal
viruses share the pool
Olivier Zablocki*, Lonnie van Zyl, Bronwyn Kirby, Marla Trindade
Institute for Microbial Biotechnology and Metagenomics, University of the Western Cape, South
Africa
*E-mail: [email protected]
Virus communities in terrestrial boiling/acidic hot springs are typically composed of a low
diversity of archaeal virus genera with distinctive morphologies. However, limited virome data are
available from scalding springs (≥50°C - ≤90°C), which may allow for a broader selection of
microbial consortia given intermediate thermal regimes. We hypothesized that scalding
environments may contain a more heterologous virus community, composed of both
bacteriophages and archaeal viruses. In this study, the viral assemblage of the Brandvlei hot spring
(South Africa) was examined through electron microscopy and metagenomics. The majority of
viral morphotypes were tailed. Siphoviridae-like virus particles, jumbo-sized myoviruses and
lemon-shaped virions (Fuselloviridae) were all observed. Using metavirome data, homologous
sequence to the polymerase family B gene (polB2) was used to provide additional support for the
presence of closely related salterprovirus-like sequences in the sample. Taxonomic classification of
predicted virus genes showed a dominance of tailed cyanophages in the spring. A large number of
predicted CRISPR loci with no database homologs hinted at a novel pool of archaeal viruses,
further supported by a large number of archaea-derived genes with archaeal virus-specific functions
but with no reported viruses. The second most abundant predicted phage genome had host
assignment to the Planctomycetes species, Gemmata, which had not previously isolated from
thermal environments, and with no isolated viruses identified to date. This study demonstrated that
a scalding spring environment contained a complex viral diversity, encompassing both archaeal
viruses and bacteriophages, whilst showing evidence for previously unknown viruses associated
with a member of the phylum Planctomycetes.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section I: Phage Ecology 23
Phage stratification in monoclonal Chlorobium phaeobacteroides in the
meromictic lake Trekhtsvetnoe
Maria Letarova
INMI, Russian Academy of Sciences
E-mail: [email protected]
The metabolic activity of prokaryotic life has been one of the dominant forces shaping the
landscapes and chemistry of the surface of our planet since the Biosphere emerged ca. 3 billion
years ago. The bacterial communities in free living ecosystems, such as in water and in soils, are
generally very complex and diverse, with every component comprising only for a small fraction of
total microbial population. Microbial populations that develop at the sites with steep transition of
physic-chemical conditions, specially of red-ox potential (Eh), may feature much higher density
and reduced complexity. The metabolic work of such populations contributes to the large scale
convertion of organic and inorganic substance at these biogeochemical barriers.
In our study we focused on the microbial communities of a sharp biogeochemical barriers
developed in liquid environment of the stratified water column in the meromictic lakes separating
from the White sea. We analysed in substantial detail the matter and energy flow in Trekhcvetnoe
Lake located in Kandalaksha bay near the White Sea Biological Station of MSU. Stratification of
the Trekhcvetnoe Lake as well as of the other lakes separating from the White Sea is imposed by a
sharp salinity gradient. The energy flow in this lake is mainly based on the sulfur cycle. Hydrogen
sulfide flow from the bottom part of the water column is intercepted by the microbial community
of 'bacterial plate' or biofilter layer that is about 30 cm wide and situated at approximately 2 m
depth. This layer is dominated by green sulfur bacteria that use energy of the light to oxidise H2S
and produce organic matter in anoxigenic photosynthesis process. The total microbial cell count in
the thin biofilter layer is 2-3 orders of magnitude higher than in surrounding water layers and
reaches 2x108 cells/ml. Metagenomic analysis revealed that 70-95% of this biomass is represented
by Chlorobium phaeovibrioides. Interestingly genetic diversity of the dominant strain population in
Trekhcvetnoe Lake is extremely low and close to that of the clonal bacterial populations.
Nevertherless the viruse like particles (VLP) observed in biofilter layer water samples taken with
the intervals as small as 2.5 cm are significantly different suggesting the ultra-fine stratification
within this narrow horizon. Taking into account high optical absorbance (OD700 > 0,8), one can
expect that steep light intensity decline over first top 2 cm should impose marked differences in
physiological state of the dominant microbial population. The H2S gradient within the biofilter is
also very steep with the concentration dropping from 6 mM to zero within 30 cm distance. This
ultra-fine stratification is reflected in the composition of associated viral community suggesting
substantial differences in virus effects such as mortality and lateral gene transfer over genetically
similar bacteria within cm distances in the liquid environment. Despite high VLP concentration
associated with high density homogenous host population the biofilter layer is very stable over
many years. This suggests that some mechanisms limiting lytic phage growth should exist in this
habitat.
The comparative study of neighboring meromictic lakes revealed that they also feature the
highly populated microbial layers that differ however by their location in the water column, by the
community composition and stability. So the comparative biogeography of the viruses and bacteria
between this closely located objects may reveal the mechanisms of the virus – bacteria coexistence
in the communities of the biogeochemical barriers.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
24 Section II: Agriculture & Food Safety
Phage-host interactions: what have we learned from studying Campylobacter
phages
Martine Holst Sørensen, Y. Emre Gencay and Lone Brøndsted*
Department of Veterinary and Animal Sciences, University of Copenhagen, Stigbøjlen 4, 1870,
Frederiksberg C, Denmark
*E-mail:[email protected]
Campylobacter jejuni remains to be the leading cause of bacterial foodborne illness in the
western world and is seriously affecting child health and mortality in developing countries. C.
jejuni encodes a unique and highly variable surface and while some diversity is due to variation in
genetic content, multiple phase variable genes are found in genetic loci encoding CPS, lipo-
oligosaccharides and O-linked glycosylation of flagella. In C. jejuni phase variable genes encode a
polyG tract that causes slipped strand mispairing during replication, leading to frame shift
mutations, thus turning expression of the gene ON or OFF.
Campylobacter phages are members of the Myoviridae and belong to the two genera of the
Eucampyvirinae subfamily; Cp220virus and Cp8virus. Using our large phage collection, we
showed that Campylobacter phages are either dependent on CPS or motile flagella for infection.
Interestingly, the receptor type dependency correlated with the phage genus; Cp220virus are
dependent on motile flagella, whereas Cp8virus rely on CPS for infection. Analysis of
flagellotropic phage F341 showed a reversible, yet specific binding to the flagellum, and
demonstrated that C. jejuni becomes phage resistant when motility is lost. Overall, our data suggest
that the expression of surface structures influence phage sensitivity and provide the bacterium with
an efficient defense mechanism against phage infection.
While the actual receptor for the flagelloptropic phages is under investigation, further
analysis of the CPS-dependent phage F336 identified the MeOPN modification of GalfNAc present
in the CPS as a phage receptor in strain NCTC11168. This lead to the hypothesis that MeOPN is a
common receptor for all CPS-dependent phages. To test this, we deleted the only MeOPN
transferase gene in strain NCTC12662 sensitive to all our CPS-dependent phages. By HR-MAS
NMR we showed that the mutant is deficient of MeOPN of the CPS and seven phages did not form
any plaques on this strain. On the other hand, 33 phages infected the mutant, although at a lower
efficiency compared to the wild type. Interestingly, the different levels of infectivity of the CPS-
dependent phages neither correlate with the time of isolation nor the origin of phages. To
investigate the phage diversity further and identify receptor-binding proteins all our CPS phages
are currently being sequenced using PacBio. Nevertheless, all our CPS-dependent phages were
affected by the lack of MeOPN, indicating the vast importance of this unique surface modification
for phage infection of C. jejuni.
Resistance to CPS phage F336 develops due to loss of the receptor by phase variation of a
polyG tract in the MeOPN-GalfNAc transferase gene cj1421. However, phage resistance is also
associated with phase variable expression of other CPS genes in vitro and in a chicken model; ON
expression states of cj1422 (attaching MeOPN to heptose) and cj1426 (attaching 6-O-Me to
heptose). Population analyses investigating all phase variable genes in C. jejuni NCTC11168
demonstrated a highly specific phase variable response after phage F336 exposure, only selecting
for specific phase variants of cj1421, cj1422 and cj1426. We hypothesize that the dynamic changes
of these modifications influence the conformation of CPS, hence impacting phage binding
depending on the interaction with the specific receptor binding protein. Finally, multiple phase
variation events led to phage resistance, thus increasing the chance of phage resistant sub-
populations present in a growing culture.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section II: Agriculture & Food Safety 25
Application of Phages to Control Pierce’s Disease
Mayukh Das, Tushar Suvra Bhowmick and Carlos F. Gonzalez
Department of Plant Pathology & Microbiology,
Center for Phage Technology, Texas A&M University, College Station, Texas
Pierce's Disease (PD), caused by a xylem-blocking, insect-transmitting bacterium Xylella fastidiosa
subsp. fastidiosa, is a major threat to the wine industry in the USA. PD is prevalent from Florida to
California and causes major damage in wine-producing regions. Current approaches for control are
only partially successful and in most cases include the use of chemical pesticides. Phage therapy is
an alternative that could provide a treatment for PD. Phages are the most abundant and ubiquitous
genetic entity on earth. Besides ubiquity, two major properties of phages, specificity and
exponential propagation, make them attractive as antibacterial agents. A phage therapy system
could offer a novel control strategy that also provides high specificity and non-toxicity to animals,
plants or non-target bacteria that may be beneficial to plants against the PD pathogen. Specifically,
a cocktail of phages exhibiting broad host range activity that reflects a diversity of receptors would
maximize the potency of the treatment and minimize the possibility for development of resistance.
A cocktail of four virulent phages (Sano, Salvo, Prado, Paz) were both tested for the therapeutic
and prophylactic efficacy in Vitis vinifera (variety Cabernet Sauvignon) in greenhouse
experiments. Vines were inoculated with bacteria and/or phage(s) and were evaluated for symptom
development for 12 weeks. During the 12-week period, triplicate vines (two cordons each) were
harvested. Plants were segmented and individual segments were assayed using Real Time PCR
(qRTPCR) to quantify Xylella and/or phages. qRTPCR results showed movement of pathogen
and/or phage(s) in vines over a 12-week period. Typical PD symptoms were visible by week 8 in
pathogen inoculated control vines. PD symptoms ceased to progress one week post-therapeutic
treatment and symptoms were not observed in prophylactically treated grapevines. In therapeutic
and prophylactic studies, the PD pathogen declined to almost non-detectable levels as compared to
vines not challenged or pre-treated with phage(s), respectively. Phage(s) were able to replicate in
vines in the presence of the host. Field trials are being conducted in Texas and California to
determine field efficacy. Successful application of phages as biocontrol agents for X. fastidiosa will
offer a novel alternative method to wine industry for the treatment and/or prevention of PD that is
effective, sustainable and environmentally safe.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
26 Section II: Agriculture & Food Safety
Bacteriophage diversity in cattle slurry
Pavelas Sazinas1, Tamsin Redgwell2, Branko Rihtman2, Aurelija Grigonyte2 & Andrew Millard1*
1 Warwick Medical School, University of Warwick, Coventry, UK; 2 School of Life Sciences,
University of Warwick, Coventry, UK
*E-mail: [email protected]
There are ~1.8 million dairy cows in the UK, with the dairy farming industry contributing £3.8
billion to the economy. The UK dairy herd produces ~15 billion litres of milk per year and ~30
million tonnes of cattle slurry. This slurry contains a mixture of bacteria, including potential human
pathogens, antibiotics, and co-selective anti-microbials, such as ionic copper and zinc. Current
legalisation requires that farms have the capacity to store five months worth of slurry, meaning that
millions of litres are stored in tanks before being used as fertiliser. The diversity of bacteriophage
within these slurry tanks is poorly characterised. In this study we have begun to characterise the
bacteriophage community, combining genomics of individual isolates along with metagenomics on
the total viral community over a three-year period. To date, greater than 100 bacteriophages
infecting Escherichia coli have been isolated and had their genome sequenced. The most
commonly isolated bacteriophages fall within the genus T4virus. However, discrete populations
can be identified based on the year of isolation. Furthermore, a large number of bacteriophages
have been isolated that fall into poorly represented genera. Exemplified by the isolation of 14
bacteriophages that fall within the genus Seuratvirus, that previously contained only two
representatives.
Analysis of the viral metagenome has allowed a many of what are thought to be complete
bacteriophage genomes to be assembled. These genomes share little similarity with known
bacteriophage isolates, yet are in high abundance within cattle slurry. In common with most viral
metagenomes, the vast majority of the metagenome has limited similarity to known phage isolates.
However, the simultaneous isolation of bacteriophage from the same sample that is used for the
construction of a viral metagenome, has proved fruitful in increasing the fraction of a viral
metagenome that can be assigned to known viruses.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section II: Agriculture & Food Safety 27
Phage Peptidoglycan Hydrolase as an Antagonist of Agrobacterium
tumefaciens
Hedieh Attai, Kenya Phillips, Jeanette Rimbey, George Smith, Pamela Brown
Division of Biological Sciences, University of Missouri, Columbia, MO
*E-mail: [email protected]
Bacteriophages can be used as biocontrol agents to protect plants from phytopathogens such as
Agrobacterium tumefaciens. We have thus isolated five lytic bacteriophages from environmental
sources (AP 2, 3, 4, 7, 8) with narrow-host ranges. Preliminary results indicate that coinoculation
of A. tumefaciens with phage limits the effects of Crown Gall disease. In order to better understand
the mechanism of phage-mediated killing, we have sequenced the bacteriophage genomes and have
begun to characterize the phage endolysins, or proteins that cleave the bacterial peptidoglycan cell
wall. The genomes of AP2 and AP3 contain a putative endolysin, Phage Peptidoglycan Hydrolase
(PPH) with an atypical domain structure. PPH contains a predicted peptidoglycan-binding region, a
transmembrane domain and a positively charged C-terminal tail. Thus, PPH is predicted to be a
transmembrane protein with its N-terminus in the periplasm, suggesting that PPH may function
independent of accessory proteins to mediate host cell lysis. Expression of PPH from an inducible
promoter inhibits cell growth in A. tumefaciens. Time-lapse microscopy shows that PPH-
expressing A. tumefaciens exhibit a branching morphology, normally only observed when the
divisome is perturbed. In addition, expression of PPH also inhibits E. coli growth and triggers cell
filamentation. Zymography, an SDS-PAGE with peptidoglycan as a substrate, confirms that PPH
cleaves peptidoglycan. Together, these observations suggest that PPH is capable of gaining access
to the periplasm where it may interact with cell division machinery, cleave the peptidoglycan, and
ultimately lyse the host cell. Further characterization of PPH includes site-directed mutagenesis of
functional residues and purification of His-tagged PPH.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
28 Section II: Agriculture & Food Safety
Characterization of Antimicrobial Properties of Salmonella Phage Felix O1
and Listeria phage A511 Embedded in Xanthan Coatings on Poly(lactic acid)
Films
Devon Radford a*, Brandon Guild b, Philip Strange a, Rafath Ahmed a, Loong-Tak Lim b,
Sampathkumar Balamurugan a
aGuelph Research and Development Centre, Agriculture and Agri-Food Canada, 93 Stone Road
West, Guelph, Ontario, N1G 5C9, Canada bDepartment of Food Science, University of Guelph, Guelph, Ontario, N1G 2W1, Canada
*E-mail: [email protected]
Beyond simply providing a barrier between food and external contaminants, active packaging
technologies aim to inhibit pathogen survival and growth within the packaged environment.
Bacteriophages have a proven track record as targeted antimicrobials but have yet to be
successfully integrated in active packaging without serious loss of activity. We have developed two
bacteriophage based xanthan coatings on poly(lactic acid) (PLA) film which significantly inhibits
Salmonella Typhimurium and Listeria monocytogenes growth in culture (P < 0.01), and
significantly reduces survival and growth of diverse cocktails of Salmonella sp. and L.
monocytogenes respectively on precooked sliced turkey breast over 30 days of anaerobic packaging
at 4 or 10°C (P < 0.05). Specifically, reductions of 0.832 log at 4°C and 1.30 log at 10°C for
Salmonella sp., and 6.31 log at 4°C and 1.52 log at 10°C for L. monocytogenes were observed. The
coating containing Listeria phage A511 also significantly inhibited growth of L. monocytogenes
over 14 days in aerobic packaging (3.79 log at 4 oC, 2.17 log at 10°C, P < 0.05). These coatings
showed 99.99% phage release within 30 minutes for both phages. Similar approaches could be
used to develop packaging inhibitory to other significant foodborne pathogens such as
Campylobacter, and Escherichia coli, as well as spoilage bacteria.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section II: Agriculture & Food Safety 29
Effect of the phage cocktail SalmoFree(R) on Salmonella reduction in
commercial broiler's farms
Viviana Clavijo, Alejandro Reyes, Diana Baquero, Alejandra Arevalo, Pilar Donado, Martha
Josefina Vives*
Universidad de los Andes.Bogotá, Colombia.
*E-mail: [email protected]
The World Health Organization considers Salmonella one of the most important zoonotic
foodborne pathogens. Bacteriophages, acting as host-specific parasites of bacterial cells, are today
one of the possible alternatives to antibiotics in animal therapy which can also contribute to food
safety and security. Researchers at Universidad de los Andes developed a phage cocktail against
Salmonella strains, focusing on those of veterinarian and human importance. The product was
named SalmoFree® and its phages have been completely characterized by host range, infection
assays, stability in chlorine, transmission electron microscopy, genome sequencing and a safety
trial in broilers kept in cage batteries. Due to the lack of knowledge in Colombia of phage therapy
and the consequent distrust among poultry farmers, the technology needs to be tested at the
production scale to be considered as viable, safe and effective, especially when there are no prior
reports on use by commercial broiler farms. The present study aimed to evaluate SalmoFree® in a
commercial broiler farm which belongs to an integrated Colombian poultry company. The
objective of the assay was to test the effectiveness of SalmoFree® in controlling Salmonella and
also to assess the relationship between the use of phages and such productivity parameters as feed
conversion, weight gain and homogeneity. We selected four production houses in a farm which had
a record of the presence of Salmonella during two previous production cycles. The selected
production-houses had between 6100 and 13400 broilers each (Ross). A two-phase feeding
program was used in this experiment: a standard commercial starter diet from day 0 to day
21, and a grower diet from day 22 to day 42. Broilers in two of the production houses were
supplemented with SalmoFree® in drinking water at days 18, 27 and 34 and those in the other two
houses were supplemented with a suspension without phages for the same days. Weight and feed
intake were recorded for the four houses. The presence of Salmonella was determined in cloacal
swabs, feces and sampling-shoes samples, following standard protocols. Data of Salmonella
incidence, feed conversion and homogeneity were calculated for all production houses and
statistical analysis was performed to compare these parameters between the houses treated with
SalmoFree® and the control houses. All together, the results represent important information for
the development of phage therapy in Columbia.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
30 Section II: Agriculture & Food Safety
Using Free and Immobilized lytic Bacteriophages to Tackle Salmonella
Contamination in Food
Hany Anany1,3, Noha Eldougdoug2, Hajar Hawsawi3, Stevan Cucic3, Vince Leung4, Monsur Ali4,
Carlos Filip4, Mansel Griffiths3
1Agriculture and Agri-Food Canada, Guelph, ON, Canada; 2Microbiology Department, Benha
University, Benha, Egypt; 3Canadian Research Institute for Food Safety, Food Science Department,
University of Guelph, Guelph, ON, Canada; 4Chemical Engineering Department, McMaster
University, Hamilton, ON, Canada
Bacteriophages have been envisioned as a novel and safe tool to control different types of
foodborne pathogenic bacteria. Salmonella is one of the most important foodborne pathogens that
are associated with various outbreaks around the world. Contaminated tomatoes and raw chicken
were linked to some of these outbreaks. Hence, the objective of this work was to isolate and
characterize different lytic bacteriophages against Salmonella Newport and apply these phages to
control the growth of Salmonella Newport and enhance the safety of cherry tomato and raw
chicken breast. Local sewage samples were used for the isolation of Salmonella Newport lytic
phages. The morphology of the isolated phages was determined by TEM and their restriction
digestion pattern was characterized using different restriction enzymes. Their stability, host range,
and ability to control the growth of Salmonella in broth were investigated.
For food application, phages were applied by dipping contaminated cherry tomato in a phage
cocktail solution or coated on the raw chicken breast packaging material (immobilized).
Polysaccharides were used to stabilize immobilized phages on the packaging material. Four lytic
phages, belonging to family Myoviridae (CGG4-1, CGG4-2) and Siphoviridae (CGG3-1, CGG3-
2), were selected from 15 isolated phages based on their broad host range patterns against 26
tested Salmonella serovars. The four phages behaved differently when stored at various
environmental conditions. One phage, CGG 4-1, was sequenced and showed no lysogenic or
virulent coding sequences in its genome. The isolated phages have a latent period of around 50 min
and burst size of around 100. When a cocktail of the isolated phages was used to control the growth
of Salmonella Newport in broth medium, complete inhibition of bacterial growth was observed at
25 and 12°C for 24 h. On the other hand, a 4.5 log CFU/g reduction in the bacterial count was
observed when applying the phage cocktail at MOI of 105 on contaminated tomato fruits stored at
25°C for 3 days. Pullulan and trehalose were able to stabilize dried phage cocktail and maintain its
infectivity on coated food packaging material. When phage cocktail coated on butcher’s paper and
applied on contaminated raw chicken breast samples, 2.1 log CFU/g reduction in Salmonella count
was achieved after 4 days storage at 12°C. These finding support the notion of considering lytic
phages as a biocontrol option for Salmonella spp contamination in different food matrices.
Furthermore, developing stable and infective phage-based bioactive packaging materials could
broaden phage applications as biocontrol agent and might alleviate some concerns of using
spraying/dipping approach to apply phages during food production.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section II: Agriculture & Food Safety 31
Bacteriophage biocontrol for improving the safety of human and pet foods
Joelle Woolston
Intralytix, Inc., Baltimore, MD
E-mail: [email protected]
Bacteriophages are naturally part of the normal microflora of many foods, and the 'phage
biocontrol' approach is based on the concept of using the right phage, in the right place, in the right
concentration to eliminate or significantly reduce pathogenic bacteria. Interest in using
bacteriophages to improve food safety has been driven by both the continued occurrence of
foodborne outbreaks worldwide and the desire of consumers for natural foods. The bacteriophage
biocontrol approach has been applied to three main areas of food safety: (i) pre-harvest treatment of
livestock, (ii) decontamination of inanimate surfaces in the processing environment, and (iii) post-
harvest treatment (i.e. direct food applications), the area which has received the most attention.
Bacteriophages can reduce levels of the targeted bacterial pathogen on a variety of foods,
including, but not limited to, dairy products, fruits and vegetables, and poultry. Because of the
specificity of bacteriophages, their application only affects the target bacteria (the pathogen) but
will not affect the other naturally present and potentially beneficial microflora. This presentation
will review the use of bacteriophage biocontrol as a food safety measure, in both human and pet
foods, as well as discuss regulatory and safety issues concerning their use.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
32 Section II: Agriculture & Food Safety
Phage Applications for Developing Countries
Tobi Nagel1, Doudou Batumbo2, Didier Bompangue2,3,4, Nicholas Carrigy5, Benjamin Chan6,
Martha Clokie7, Ian Connerton8, Daniel De Vos9, Fitriya Dewi10, Ayman El-Shibiny11, Lasha
Gogokhia12, Diah Iskandriati10, Adamu Ahmad Kaikabo13, Guyguy Kamwiziku2, Erastus
Kang'ethe14, Samuel Kariuki15, Rudovick Kazwala16, Alice Maestri17, Alice Nyambura Maina14,15,
Angela Makumi15, Jesca Nakavuma18, Janet Nale7, George Nasinyama18,19, Julien Ntaongo2, Joko
Pamungkas10, Jean-Paul Pirnay9, Paul Turner6, and Reinhard Vehring5
1Phages for Global Health (USA), 2University of Kinshasa (DR Congo), 3Ministry of Public Health
(DR Congo), 4University of Franche-Comte (France), 5University of Alberta (Canada), 6Yale
University (USA), 7University of Leicester (UK), 8University of Nottingham (UK), 9Queen Astrid
Military Hospital (Belgium), 10Primate Research Center at Bogor Agricultural University
(Indonesia), 11University of Science and Technology (Egypt), 12University of Utah (USA), 13National Veterinary Research Institute (Nigeria), 14University of Nairobi (Kenya), 15Kenya
Medical Research Institute (Kenya), 16Sokoine University of Agriculture (Tanzania), 17University
of Turin (Italy), 18Makerere University (Uganda), 19Kampala International University (Uganda)
Phages for Global Health is a nonprofit organization that facilitates the development of phages for
applications in developing countries. We accomplish this by bringing together global phage experts
and developing world infectious disease specialists, including food safety scientists, veterinarians,
clinicians, regulatory advisors, and stakeholder engagement professionals. With this mix of
specialists, we create international, multidisciplinary teams that have both the technical expertise
necessary to develop phage products and the local scientific and cultural knowledge to deliver
products that will be socially accepted. Our growing consortium of partners includes leaders from
nonprofit organizations, universities and governmental institutions across Africa, Asia, Europe and
North America. We have also created a series of short-term laboratory training programs to teach
the key essentials of phage biology to scientists in developing countries. Current projects include:
(1) Training program (East Africa, West Africa, Indonesia)
We have developed a 2-week, hands-on laboratory workshop through which we can teach scientists
in developing countries how to isolate and characterize phages in their own regions. The first of
these workshops will be held in East Africa during July 2017 (hosted at Makerere University in
Uganda), and plans are underway for subsequent workshops in West Africa (at the National
Veterinary Research Institute in Nigeria) and Indonesia (at the Primate Research Center at Bogor
Agricultural University).
(2) Campylobacter phages for decontamination of retail poultry meat (Kenya, Egypt)
Campylobacter is the leading bacterial cause of gastroenteritis worldwide, and recent data
demonstrate that it is particularly detrimental for children under age 2 in developing countries.
Globally, Kenya has the highest case fatality rate of Campylobacteriosis, with 8.8% of infected
people dying, mostly children. In terms of overall incidence of Campylobacter-associated diarrhea,
Egypt has the highest reported rate of any developing country. To address this public health issue,
we have undertaken to develop phage products to decrease Campylobacter levels on retail poultry.
(3) Cholera phages for water decontamination and prophylactic treatment (DR Congo)
According to the World Health Organization, there are an estimated 1.4 - 4.3 million cholera cases
and up to 142,000 associated deaths worldwide each year. Over the past decade >60% of the
reported cholera cases have occurred in Africa, with the largest percentage of those in the
Democratic Republic of Congo. We are developing phage products to test for both water
decontamination and prophylactic treatment of people during cholera outbreaks.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section III: Phage-Based Biotech 33
Engineered phage as therapeutics against P. aeruginosa
Yvette Del Rosario, Matt LaFave, Srividya Akella, Toby H. Richardson, Todd C. Peterson, Magda
Barbu*
Synthetic Genomics Inc., La Jolla, California, USA.
*E-mail: [email protected]
The emergence of bacterial clinical isolates resistant to nearly all-current use antibiotics has
renewed interest in the development of lytic bacteriophage (or phage) therapeutic and prophylactic
agents. The advantages of using phage to combat infectious diseases include minimal disruption of
resident flora, lack of cross-resistance with antibiotics, low toxicity and self-limiting dosing.
Despite these attractive features, translational development of natural phage has been hindered
mainly by the difficulty of accessing bacterial hosts within biofilms, the rapid emergence of
resistant bacteria to a single phage, and above all, by the narrow host specificity of phage compared
to antibiotics. The need for customized and complex combinations of natural phage to achieve
adequate host range activity has made their development as licensed therapeutics very difficult.
Genetic engineering of phage genomes can overcome these hurdles; however, broadly applicable
methods for efficient construction of defined mutations in virulent phage genomes are still in their
infancy. Using multidrug-resistant Pseudomonas aeruginosa (MDR PA) as proof-of-principle, we
developed a completely cell-free phage engineering platform that allows rapid and iterative editing
of viral genomic DNA. In parallel, we sequenced and annotated the genomes of several hundred
MDR P. aeruginosa clinical isolates, and subsequently determined the susceptibility of each isolate
to specific phages. Through bioinformatics analysis in combination with our engineering platform,
we have successfully collapsed the host range of a family into a representative phage. We further
engineered wide host range phage to express secondary payloads, such as biofilm degrading
enzymes and antimicrobial moieties, and demonstrated that these phages have improved activity
compared to natural phage in P. aeruginosa models of infection. We predict that our engineering
platform may be adapted for more refined applications, such as construction of viruses with
multiple receptor specificity, which may accelerate both development of phage therapeutics and
functional genomics.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
34 Section III: Phage-Based Biotech
Teaching phages to think ahead: a structure-inspired rapid host range
modification system
Sébastien Lemire*, Kevin Yehl, Andrew Yang, Hiroki Ando and Timothy Lu
Research Laboratory of Electronics, Center for Synthetic Biology, Massachusetts Institute of
Technology
*E-mail: [email protected]
Bacterial antibiotic resistance development will be one of the most serious public health threat of
the 21st century. Still, few solutions have been put forth and antibiotic development is largely at a
standstill. Bacteriophages have a tremendous potential in the fight against antibiotic resistant
bacteria; however, naturally sourced bacteriophages often display a limited host range and bacteria
readily evolve resistance against them. While assembling cocktails of bacteriophages that
collectively provide the desired pathogen strain coverage is a convenient ad hoc solution to the
problem, current drug approval schemes make it hard for such cocktails to pass testing and become
routinely available for the treatment of patients.
Inspired by antibody specificity engineering principles, we have created vast libraries of phage
mutants (up to ~107 different mutants) by targeting the regions of the phage T3 tail fiber protein
most likely to direct receptor recognition. This approach maximizes the functional diversity while
minimizing the risk of crippling the protein. We call these mutants phagebodies and screened the
libraries for host-range alteration with a focus on isolating phagebodies capable of infecting
mutants E. coli BL21, the normal T3 host, that are resistant to T3. Despite sampling a very small
fraction of the total phagebody diversity we have created, we isolated phagebodies that were
capable of curbing resistance development in its host on plates and in liquid cultures for an
extended period and some even appeared to completely prevent resistance development, a feat we
could not reproduce through natural evolution of T3. We also demonstrate that the genetic diversity
we generated translates into an equally wide array of different host range phenotypes including
phagebodies that have not simply expanded their host range beyond binding to wild-type E. coli
BL21 but have actually lost the ability to grow on the wild-type bacterium and instead prefer the
T3-resistant mutants.
Our work shows that therapeutic phages can be programmed to brace for resistance and proposes a
pre-emptive design as opposed to deploy reactive coping strategies after resistance becomes an
issue. We also show that expanding host range to target receptor mutants does not necessarily come
at the expense of species selectivity, an advantage of phages over chemical antibiotics.
This is an emerging concept that we hope will attract attention to the possibilities of genetic
engineering and synthetic biology in promoting phage therapy as a viable therapeutic option for
bacterial pathogen control. We anticipate that our phagebody library strategy will help, not only to
create better therapeutic phages, but also explore the link between structure and function in phage-
host interactions.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section III: Phage-Based Biotech 35
Artilysin®: Targeted elimination of bacterial pathogens resulting in a faster
wound healing
Stefan Miller*
Lisando GmbH, Am BioPark 13, D-93053 Regensburg, Germany
*E-mail: [email protected]
Chronic wounds are becoming a growing problem for patients, healthcare professionals, and the
health care system. For example, the US health care system this means affecting about 5.7 million
patients and costing an estimated 20 billion dollars annually. Decubitus wounds e.g. are often
infected by multi-resistant strains of Pseudomonas aeruginosa, Staphylococcus aureus or
Acinetobacter baumanii. To accelerate wound healing and protecting the skin microbiome, there is
a clear need for effective antimicrobials, which show a targeted mode of action.
Artilysins display a new, targeted mode of action. Artilysins are recombinant fusion proteins
consisting of an endolysin combined with a targeting peptide that transfers the endolysin through
the outer membrane of Gram-negatives. Art-175 kills P. aeruginosa, including multidrug-resistant
strains, by puncturing the peptidoglycan layer within a minute. Artilysin® Art-175 is highly
effective against highly antibiotic resistant strains of P. aeruginosa and A. baumanii, without
observable resistance development. As Artilysins activity does not require an active bacterial
metabolism, they show a superior bactericidal effect against persisters of P. aeruginosa and A.
baumanii.
Preclinical data underline the broad applicability of Artilysins against bacterial infections being
non-cytotoxic and non-haemolytic. Clinical observation studies with several patients using standard
(homecare) or compassionate use (hospital) application show that on wounds, were conservative
treatments fail, Artilysins application results in removing the persisting bacterial pathogens and a
clearly accelerated wound healing. By removing pathogens, Artilysin facilitates the commensal
bacteria re-colonising skin and wound, thereby preventing pathogens from re-infecting. In
summary, Artilysins are novel antibacterial proteins for targeted elimination of bacterial infections
that favour the natural microbiome and thereby accelerate wound healing.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
36 Section III: Phage-Based Biotech
Using Bacteriophage Endolysins to Create Custom Immunotherapeutics
Daniel C. Nelson*
Institute for Bioscience and Biotechnology Research, University of Maryland, Rockville, MD
*E-mail: [email protected]
In the era of growing antibiotic resistance, life-threatening infections by several bacterial pathogens
are a cause for major concern. While the lytic activity of bacteriophage endolysins offers an
untapped reservoir of potential therapeutics, bacteriolytic therapies are not without concern,
including release of bacterial toxins, lipopolysaccharide, or other inflammatory mediators.
Additionally, as challenges with vaccines continue to rise, non-traditional concepts that engage the
innate and/or adaptive immune responses will be needed to meet these challenges. Toward this end,
we are developing two independent platform technologies that exploit the species-specific affinity
of endolysin cell wall binding domains (CBDs) for the bacterial surface of target organisms. In one
approach, CBDs are fused to specific antitoxin neutralizing monoclonal antibodies to generate
Infection Site Targeted Antitoxin antibodies, or ISTAbs. ISTAbs accumulate at the site of
infection where they are needed most and sequester the toxins at the bacterial surface, thus
immediately neutralizing the effects of the toxins and preventing their release into circulation.
More importantly, since the antitoxin antibodies are humanized, anchoring them to the bacterial
surface via the CBDs functionally opsonizes the bacterium targeting it for opsonophagocytic
killing. In a second approach, CBDs are fused to an antigen against which most people have
existing antibodies (i.e. childhood vaccines such as tetanus or diphtheria toxoid). The CBD acts as
an “ImmunoBridge”, anchoring the antigen to the bacterial surface thereby redirecting pre-existing
immunity against the new invading pathogen resulting in opsonophagocytic clearance of infection.
In this respect, the ImmunoBridge acts as a post-exposure vaccine. Both ISTAbs and
ImmunoBridges possess the potential to have broad application across multiple pathogens based on
an array of available endolysin CBDs.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section III: Phage-Based Biotech 37
Development of Phage-derived Particles for Gene Therapy of the Microbiome
Todd B. Parsley*
SynPhaGen, LLC. Gaithersburg, MD USA.
*E-mail: [email protected]
Objective. At SynPhaGen we are developing a platform technology for in vivo transduction of the
microbiome. The platform uses non-lytic, non-bactericidal and replication-deficient bacteriophage-
derived particles (BP) that are engineered to target and deliver their therapeutic genetic cargo to
specific bacteria inhabiting the commensal microflora, transforming the bacterial cell into a local
protein factory. We hypothesize that the close association of the transduced bacteria with host cells
will deliver the therapy to the site of greatest medical need, thus increasing the efficacy of the
therapeutic and decreasing the potential adverse systemic side-effects. The objective of the
presented studies is to demonstrate proof-of-concept using BPs for the in vitro and in vivo
transduction of bacteria and expression of therapeutic sequences.
Methods. BPs were synthesized in an E. coli packaging cell line using a two-plasmid system: one
plasmid encoding an engineered filamentous phage genome and the other a therapeutic plasmid
composed of the gene(s) of interest and a modified receptor binding protein engineered to
selectively bind to and transduce specific strains of E. coli or Pseudomonas aeruginosa. BPs
programmed with a nanoluciferase reporter gene were harvested from packaging cell line culture
supernatants and characterized for selective packaging of the therapeutic plasmid sequences,
examined for transduction specificity, titered in susceptible bacteria in vitro, and subsequently used
in vivo in murine models. BPs programmed to deliver genes encoding functional sRNA gene
sequences, an immunogenic peptide (TARP), or soluble therapeutic protein (rhIL-10), were also
engineered and evaluated in vitro.
Results. Characterization of BPs by PCR revealed selective packaging of the therapeutic plasmid
sequences. In vitro analysis of BPs encoding nanoluciferase demonstrated that the transduction
specificity could be tuned for transduction of specific strains of E. coli or P. aeruginosa by
modification of the N-terminus of the receptor binding protein. This was shown both in vitro as
well as in vivo where BPs could successfully transduce E. coli cells in the gastrointestinal tract of
CD-1 mice. Data on the in vivo transduction of P. aeruginosa in the lungs of Balb/c mice is in
progress and will be presented. Expression of TARP peptide and soluble rhIL-10 was achieved in
E. coli transduced with BPs programmed to deliver the respective genes. Transduction of E. coli
with BPs engineered to deliver genes encoding functional sRNA targeting lacZ resulted
suppression of endogenous ß-galactosidase expression.
Conclusions. As proof of concept, we have engineered BPs that selectively transduced specific
strains of E. coli or Pseudomonas aeruginosa. Selectivity was achieved by modification of the N-
terminus of the BP receptor binding protein. BPs can be programmed to deliver genes encoding
functional sRNA gene sequences, immunogenic peptides (TARP), or soluble therapeutic proteins
(rhIL-10).

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
38 Section III: Phage-Based Biotech
Reconsidering temperate phages for therapy
Britney Y. Lau, Raga Krishnakumar, Julian M. Wagner, Corey M. Hudson, Joseph S. Schoeniger,
Kelly P. Williams*
Sandia National Laboratories, Livermore CA 94551
*E-mail: [email protected]
Temperate phages grow lytically in most of the bacterial cells they infect, yet produce a small
fraction of lysogen cells by repressing lytic growth and establishing stable inheritance, usually
through integration into the chromosome. Temperate phages have rightly been shunned for therapy
because the lysogens i) are not killed, ii) are resistant to reinfection by the original phage, iii) may
have gained virulence/resistance/toxin genes enhancing the pathogenicity phenotype (lysogenic
conversion), and iv) can occasionally transduce chromosomal markers flanking the integration site
[1]. However, this guideline had set in before the development of a general PCR-based approach
for multilocus engineering of bacteriophage genomes [2]. Engineering knockout mutations in
integrase or repressor genes essential for lysogeny (and in virulence/resistance/toxin genes if any)
would remove the above objections to temperate phages.
This lysogeny-knockout approach would allow exploitation of the vast number of temperate phages
that are integrated, in prophage form, within sequenced bacterial genomes. Prophages are the main
class (ICEs being the other major class) within the larger category of genomic islands, which can
be defined as mobile DNAs that integrate site-specifically into bacterial (or archaeal) genomes due
to an integrase (of the tyrosine recombinase family, or less frequently, of the serine recombinase
family). We have developed tools that identify and precisely map such genomic islands. Our
experimental method Juxtaposer [3] can detect mobility events due to genomic islands (and
insertion sequences) in subpopulations of a culture, through the circularization and deletion
junctions they produce among next-generation sequencing reads. For higher throughput, we have
developed two bioinformatic methods, and applied them to a large set of sequenced genomes.
Islander is based on the preference of genomic islands for inserting into tRNA genes [4].
Comparator is based on two principles: the cohesion of the integration module, which practically
means that the island’s integrase gene will be found near one end of the integrated island, and a
negative comparative genomic approach, meaning a search for a reference genome in which the
integration site of the candidate island is uninterrupted. These complementary bioinformatic tools
find large numbers of genomic islands, and Comparator can also be configured to find insertion
sequences. In a test set of 2168 genomes we found 7949 islands. We are scaling up the application
of these algorithms to the >80000 prokaryotic genomes at GenBank. This database will allow
selection/production of custom phage cocktails from very close relatives of nearly any target
bacterium, which are more likely to be efficacious at killing the target than phages “fished” from
environmental or clinical samples.
1. Abedon ST, Kuhl SJ, Blasdel BG, Kutter EM. 2011. Bacteriophage 1:66-85.
2. Ando H, Lemire S, Pires DP, Lu TK. 2015. Cell Syst. 2015 1(3):187-196.
3. Schoeniger JS, Hudson CM, Bent ZW, Sinha A, Williams KP. 2016. Nucleic Acids Res 44:6830-6839.
4. Hudson CM, Lau BY, Williams KP. 2015. Nucleic Acids Res 43:D48-D53.
Sandia National Laboratories is a multimission laboratory managed and operated by National Technology
and Engineering Solutions of Sandia, LLC., a wholly owned subsidiary of Honeywell International, Inc., for
the U.S. Department of Energy’s National Nuclear Security Administration under contract DE-NA0003525.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section III: Phage-Based Biotech 39
A Synthetic Platform for Rapid and Reporter-Free Engineering of
Bacteriophage Genomes
Samuel Kilcher*, Patrick Studer, Christina Muessner, Jochen Klumpp, and Martin J. Loessner
ETH Zurich; Institute of Food, Nutrition and Health
*E-mail: [email protected]
Targeted modification of virulent bacteriophage genomes is frequently achieved using homology-
directed allele replacement. This approach is difficult, time-consuming, and often requires the
incorporation of phage-selectable markers such as fluorescence or luminescence. Therefore, the use
of engineered phages in biotechnology and fundamental research has been limited to phages for
which efficient cloning systems are available. Our objective was to develop a fast, accurate, and
reporter-free method for targeted engineering of a wide range of phage genomes that is broadly
applicable to viruses of Gram-positive bacteria. To this end, we used L-form bacteria as recipients
of synthetic phage genomes: L-forms are cell wall-deficient variants of otherwise walled bacteria
that indefinitely proliferate in osmotically stabilized media. They lack the multi-layered
peptidoglycan envelope, which restricts transformation of Gram-positive bacteria with large DNA
molecules. We show that the Listeria monocytogenes L-form strain Rev2L can be transformed with
full-length bacteriophage DNA, resulting in genome rebooting and production of infectious viruses
from naked DNA. Listeria L-forms did not only reboot Listeria phage genomes, but also supported
reactivation of Bacillus and Staphylococcus phage DNA, effectively bypassing the evolutionary
conserved, strict genus barriers controlling virus infection. Based on these findings, we developed a
unique platform technology for phage engineering that uses Rev2L cells as rebooting
compartments for fully synthetic, in vitro-assembled genomes, allowing rapid, accurate and
reporter-free construction of tailored phage infecting Gram-positive pathogens. We used this
approach to produce strictly lytic bacteriophages by targeted modification of temperate phage
genomes, and demonstrated strongly enhanced killing efficacy of the synthetic viruses. Lytic
activity was further expanded by incorporation of heterogeneous pathogen-specific cell wall-
hydrolases into recombinant phages as genetic payload. Altogether, L-form bacteria as rebooting
tools in synthetic phage biology open unprecedented avenues not only for fundamental research,
but also for therapeutic and biotechnological phage development.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
40 Section IV: Phage-Host Interactions
Molecular hijacking of Pseudomonas
Rob Lavigne
Laboratory of Gene Technology - KU Leuven, Leuven, Belgium.
Bacteriophages infecting Pseudomonas aeruginosa mirror the widespread and diverse nature of
their hosts. Tailed Pseudomonas phages are represented in different taxonomic clades and share
little or no sequence similarity between them, with dsDNA genomes ranging between 40kb to over
300kb. Most of this genomic diversity is found within a high number of functionally unknown
open reading frames (ORFans) that are present in hypervariable regions, which are often involved
in adaptation to host-specific conditions. Therefore, these phages are an ideal system to study the
molecular interactions that govern the virus-host interplay.
Our research aims to understand this virus-host interplay, elucidating the role of ORFans which
mediate the metabolic take-over of the bacterial cell. This subversion of host functions is incredibly
diverse, impacting replication, transcription, RNA turnover, (post)-translational modifications and
metabolic pathways. This lecture will focus on different examples which illustrate these hijacking
mechanisms and links them to potential applications in antibacterial design strategies and
biotechnological applications.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section IV: Page-Host Interactions 41
Fighting with phages: how epidemic Vibrio cholerae defends against viral
attack
Kim Seed
Department of Plant & Microbial Biology, University of California, Berkeley
*E-mail: [email protected]
Vibrio cholerae is a globally important water-borne pathogen that is the causative agent of the
severe acute diarrheal disease cholera. Successful epidemic strains of V. cholerae must defend
against predatory phages in the aquatic environment and within the human intestinal tract. Our
work suggests that a key weapon for phage defense in epidemic V. cholerae are mobile genetic
elements called PLEs (for phage-inducible chromosomal island-like elements). We have identified
and characterized five unique PLEs in V. cholerae. PLE+ isolates have been recovered from
patients between 1949-2011 (spanning the entire collection period for which strains were
available), from different locations including Egypt, Mozambique and Bangladesh. All PLEs,
regardless of temporal or geographic origin, show specific activity towards a prominent phage
called ICP1. Our efforts to understand the molecular mechanisms underpinning PLE activity,
including how specificity is imparted onto PLE-phage interactions and how PLE blocks phage
production, will be discussed.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
42 Section IV: Phage-Host Interactions
Phages and biofilms a complex interaction: strategies to improve phage
efficacy against infectious biofilms
Joana Azeredo, Priscila Pires, Luis Melo and Sanna Sillankorva
CEB – Centre of Biological Engineering, LIBRO - Laboratory of Research in Biofilms Rosário
Oliveira, University of Minho, 4710-057 Braga, Portugal
The intricate heterogeneous structure of biofilms confers to bacteria an important survival strategy
in times of adversities even against their natural predators – the phages. In these structures, phages
and cells establish a complex relationship to guarantee the long term survival of the progeny of
both entities. Theoretically, the close proximity of cells within the biofilm structure could enhance
phage-host interaction and facilitate phage infection. Conversely, the biofilm structure and
composition as well as the physiological state of the biofilm cells may be an obstacle to phage
infection. Nonetheless, phages have developed mechanisms to overcome biofilm barriers in a
natural evolutionary prey-predator model. A thorough characterisation of biofilm/phage interaction
and the identification of the weak aspects of biofilms and the strong features of phages are thus
important to develop efficient phage-based biofilm control strategies. In this presentation studies
involving the use of phages for the treatment or prevention of bacterial biofilms will be presented
highlighting the biofilm features that difficult phage infection and the phage characteristics that
enhances biofilm control. Also, some strategies based on combined therapies that can be used to
enhance phage therapy against infectious biofilms will be presented (Figure 1).
Figure 1. Schematic representation of strategies to overcome biofilm barriers using phages and
combined therapies. The biofilm is represented by cells under different metabolic conditions and
susceptibility to phages

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section IV: Page-Host Interactions 43
When CRISPR and phage collide: What to do when foe becomes friend?
Benjamin J. Rauch, Adair L. Borges, Joseph Bondy-Denomy*
Department of Microbiology & Immunology and Quantitative Biosciences Institute, University of
California San Francisco
*E-mail: [email protected]
Bacterial CRISPR-Cas systems utilize sequence-specific RNA-guided nucleases to defend against
bacteriophage infection. As a counter-measure, numerous phages produce proteins to block the
function of the Class 1 CRISPR-Cas systems, which utilize multi-subunit protein complexes to
enact immunity. To find inhibitors of Class 2 systems (e.g. Cas9), we took a bioinformatics
approach to identify candidate microbes for downstream investigation. We searched bacterial
genomes for the co-existence of a CRISPR spacer and its target, a potential indicator for CRISPR
inhibition. This “self-targeting” is lethal if the CRISPR-Cas system is functional, as the bacterial
genome is cleaved. This analysis led to the discovery of four unique and pervasive type II-A
CRISPR-Cas9 inhibitor proteins encoded by Listeria monocytogenes prophages. Despite the utility
of genomic self-targeting as a discovery tool, in the Pseudomonas aeruginosa Class 1 CRISPR-Cas
system, we find that self-targeting (i.e. the acquisition of a prophage possessing a CRISPR target
and encoding an anti-CRISPR) is not a stable situation. Self-targeting P. aeruginosa strains have
growth defects, which can be suppressed by the deletion of cas genes. Therefore, our understanding
of self-targeting is currently incomplete and we are now investigating how bacteria cope with the
concomitant presence of a CRISPR guide RNA and its target in the same cell.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
44 Section IV: Phage-Host Interactions
Understanding inter-species gene exchange and compatibility in a
bacteriophage model
F.L. Nobrega 1, Y. Liu2, N.M. Edner3, S.J.J. Brouns1-2
1Bionanoscience, TU Delft, Delft 2Wageningen UR, Wageningen, The Netherlands 3Division of
Infection and Immunity, University College London, London, United Kingdom
Bacteriophages are highly diverse viral entities that strongly influence the evolution of bacterial
communities. While the study of phages has provided significant knowledge in other fields, phages
themselves remain relatively obscure entities. Many important questions linger regarding phage
modular genome organization, rapid phage evolution, and host-specific interactions. Here we show
a tool to study gene function and essentiality, host specificity and evolution. This technique is
based on bacteriophage genome shuffling for the creation of chimeric phages. By applying this,
from all 3 major families of tailed phages (E. coli phage Myoviridae S2-36s, Siphoviridae S2-55s
and Podoviridae S1-55L) we were able to obtain many different recombinant phages with distinct
features from the parental phages. Most interestingly, we have sequenced and annotated the
genome of one recombinant phage (chimera phiChi3) using Nanopore sequencing, and this
revealed rearrangement of genes from the three parental phages. Sixty-five percent of the
recombinant phage genes derived from S2-36s, and 29 % and 7 % from phages S2-55s and S1-55l
respectively. Although most derived from S2-36s, the recombinant phage surprisingly does not
display the same morphology. Instead, it has a siphovirus morphology like phage S2-55s. This
morphology is mostly derived from the contribution of phage S2-55s to the tail structure of the
recombinant phage, while S2-55s provided capsid genes, and S2-36s donated the host recognition
structures. However, the host range of the recombinant phage is limited when compared to that of
S2-36s, which we discuss to be related to the absence of important receptor binding proteins, as tail
fibers with enzymatic activity. This confirm that phages have high levels of flexibility and
adaptability to accommodate and re-arrange genetic information from other phages, even from
distinct families, posing the hypothesis of exchangeable protein modules complementarity among
microbial populations.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section IV: Page-Host Interactions 45
Evolutionary ecology of CRISPR-Cas
Edze R. Westra
Environment and Sustainability Institute, Centre for Ecology and Conservation, University of
Exeter, Biosciences, Penryn, Cornwall, UK.
*E-mail: [email protected]
Bacteria have a range of sophisticated immune mechanisms to protect against virus infections, but
it is unclear why all these different mechanisms evolved in the first place. Under laboratory
conditions, bacteria typically evolve de novo virus resistance using either surface modification or
CRISPR-Cas adaptive immune systems. In this talk I will discuss ecological factors that can tip the
balance in the evolution of these two immune mechanisms and examine their distinct co-
evolutionary implications.
Recent publications: van Houte S, Buckling A, Westra ER. Evolutionary Ecology of Prokaryotic Immune
Mechanisms. 2016 Microbiol Mol Biol Rev. 80(3):745-63.
van Houte S, Ekroth AK, Broniewski JM, Chabas H, Ashby B, Bondy-Denomy J, Gandon S, Boots
M, Paterson S, Buckling A, Westra ER. 2016 The diversity-generating benefits of a prokaryotic
adaptive immune system. Nature 532(7599):385-8.
Westra ER, van Houte S, Oyesiku-Blakemore S, Makin B, Broniewski JM, Best A, Bondy-
Denomy J, Davidson A, Boots M, Buckling A. 2015 Parasite Exposure Drives Selective Evolution
of Constitutive versus Inducible Defense. Curr Biol. 25(8):1043-9.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
46 Section IV: Phage-Host Interactions
A variety of deazapurine modifications that protect the phage DNA from the
restriction system of the hosts.
Geoffrey Hutinet1, Witold Kot, Roman Hillebrand, Alexander B. Carstens, Sylvain Moineau,
Graham F. Hatfull, Peter C. Dedon, Lars H. Hansen, Valérie de Crécy-Lagard.
1 Department of Microbiology and Cell Science, University of Florida, Gainesville, FL, USA.
*E-mail: [email protected]
Escherichia coli phage 9g is resistant to many restriction enzymes and we recently discovered its
DNA was heavily modified with 2’-deoxyarchaeosine (dG+), a novel complex modification. This
7-deazaguanine-derived base (G+) is normally found in tRNA of Archaea. Both G+ and queuosine
(Q), another 7-deazaguanine derivative found in bacterial and eukaryotic tRNA, are synthesized
from GTP through the 7-cyanodeazaguanine (preQ0) intermediate. Phage 9g genome encodes
homologs of the preQ0 synthesis proteins folE, queD and queE. These are functional when
expressed in trans on multicopy plasmids and complement the Q- phenotype of the respective E.
coli mutants. A homolog of the archaeal Gat-QueC protein, previously shown to be involved in G+
synthesis, is also encoded in phage 9g genome and we propose it is involved in converting preQ0 to
the archaeosine base before or after its insertion in phage DNA. In Archaea, preQ0 is inserted into
tRNA by the tRNA-guanine transglycosylase (aTGT). A TGT-like enzyme is encoded in 9g
genome, and we renamed it DpdA for DeoxyPurine in DNA. Detailed multiple alignment analysis
revealed that all the DpdA proteins identified in phages contain aTGT catalytic and preQ0 binding
residues but lack the predicted Zinc binding residues. In addition, a histidine residue is highly
conserved in all these proteins, at position 196 of 9g DpdA, but is absent from the homologs that
modify tRNAs. Expression of the phage 9g dpdA gene did not complement the Q- of an E. coli tgt
strain, confirming that it has functionally diverged. Phage genomes encoding G+ biosynthesis
pathway proteins can be classified as follow: 1) those encoding both DpdA and other proteins of
the G+ or Q biosynthetic pathways; 2) those who only harbor DpdA; 3) those encoding no DpdA
nor TGT but some G+ biosynthetic pathway proteins. A Mass Spectrometry (MS) analysis of
genomic DNA from a subset of these phages showed that phages from the first group harbored dG+
or other 2’-deoxyribose-7-deazaguanine derivatives. For example, dpreQ0, the deoxynucleotide
form of the preQ0 base, was detected in the Mycobacterium Rosebush phage, whereas the
Streptococcus Dp-1 phage harbored dpreQ1 (the deoxynucleotide form of 7-aminomethyl-7-
deazaguanine or preQ1 base). Interestingly, these phages are also protected from various restriction
enzymes. Heterologous expression in E. coli of the two 9g genes thought to be required for
insertion of G+ in DNA, gat-queC and dpdA, conferred partial resistance to cleavage by EcoRI to
the expression plasmids. MS analysis of the corresponding plasmid DNA showed that it was
modified with 2’-deoxyribose-7-amido-7-deazaguanine (dADG) and not dG+, as the parent 9g
phage. This work extends the discovery of G+ in DNA to other 7-deazaguanine derivatives that
protect phages from a wide variety of restriction enzymes encoded by the hosts.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section IV: Page-Host Interactions 47
The multi-component antirestriction system of phage P1
Denish Piya, Leonardo Vara, William K. Russell, Ry Young, Jason J. Gill
Texas A&M University, College Station, TX
*E-mail: [email protected] / [email protected]
Bacterial Type I restriction-modification (R-M) systems present a major barrier to foreign DNA
entering the bacterial cell. The temperate phage P1 packages several proteins into the virion that
protect the phage DNA from host restriction. Isogenic P1 deletion mutants were used to
reconstitute the previously described restriction phenotypes associated with darA and darB. While
P1ΔdarA and P1ΔdarB produced the expected phenotypes, deletions of adjacent genes hdf and
ddrA also produced darA-like phenotypes and deletion of ulx produced a darB-like phenotype,
implicating several new proteins of previously unknown function in the P1 dar antirestriction
system. Interestingly, disruption of ddrB decreased P1’s sensitivity to EcoB and EcoK restriction.
Proteomic analysis of purified virions suggests that packaging of antirestriction components into
P1 virions follows a distinct pathway that begins with the incorporation of DarA and Hdf and
concludes with DarB and Ulx. Electron microscopy analysis showed that hdf and darA mutants
also produce abnormally high proportions of virions with aberrant small heads, which suggests Hdf
and DarA play a role in capsid morphogenesis. The P1 antirestriction system is more complex than
previously realized and is comprised of multiple proteins including DdrA, DdrB, Hdf, and Ulx in
addition to DarA and DarB.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
48 Section V: Phage Therapy (Part I)
Pro- and anti-inflammatory responses of peripheral blood mononuclear cells
induced by Staphylococcus aureus and Pseudomonas aeruginosa phages
Jonas D Van Belleghem1*, Maia Merabishvili2,1, Frédéric Clement3, Rob Lavigne4 and Mario
Vaneechoutte1.
1Laboratory Bacteriology Research, Department of Chemistry, Microbiology and Immunology,
University Ghent, 9000, Ghent, Belgium, 2Laboratory for Molecular and Cellular Technology
(LabMCT), Queen Astrid Military Hospital, 1120 Brussels, Belgium, 3Centre for Vaccinology,
Ghent University and Hospital, 9000, Ghent, Belgium, 4Division of Gene Technology, Katholieke
Universiteit Leuven, 3001, Leuven, Belgium
*E-mail: [email protected]
The ability of bacteriophages to kill bacteria is well known as well as their potential use to combat
bacterial infections, as an alternative to antibiotics. Bacteria that inhabit the intestine and skin are
generally regarded as stable residents that may confer metabolic and/or immune benefits to their
hosts. The host immune system has evolved mechanisms to tolerate these commensal organisms
while at the same time providing protection for the host from pathogens. Similarly, metagenomic
studies have revealed that a vast variety of bacteriophages are associated with healthy humans. It
has been demonstrated that oral uptake of phages by animals results in the translocation of phages
to systemic tissues. This suggests that mammals have mechanisms for the uptake and delivery of
phages and may allow intestinal phages to elicit innate and adaptive immune responses. We show
that individual phages can induce an immune response.
In order to assess a possible immune response against phages, we conducted a whole transcriptome
analysis of human peripheral blood mononuclear cells (PBMCs) stimulated with either
Pseudomonas aeruginosa or a P. aeruginosa phage PNM lysate. The phage lysate most closely
reflects the immunological state obtained during phage therapy, when the phage titer is the highest
and predominantly bacterial fragments, from the lysed bacterial cells, are present.
Next, to understand to what extent the phage particles interact with the human immune system, the
immune response induced by two different phage purification strategies (i.e. either a phage lysate
or a highly-purified phage) was compared. Using twelve immunity related genes, the immune
response induced by five different phages, representing the three major phage morphologies, were
compared, four infecting P. aeruginosa and one infecting S. aureus. This might give us an idea
whether the immune response might be phage host specific or phage morphology dependent.
We found that for all five phages, the overall immune response, as determined by RT-qPCR is very
comparable and largely in correspondence with the transcriptome analysis: down-regulation of LYZ
and TGFBI, and up-regulation of CXCL1, CXCL5, IL1A, IL1B, IL1RN, IL6, SOCS3 and TNFA.
Moreover, the immune response induced by a large number of phages (i.e. 1011 pfu/ml) was found
to be endotoxin independent, since addition of 106 EU/ml (or 1 EU/PBMC) to the highly-purified
S. aureus phage ISP or P. aeruginosa phage PNM did not subvert the immune response to the one
induced by the corresponding phage lysate.
The influence of phages on e.g. cytokine production, as observed in our study, may not be all that
unexpected, since there are several indications for other interactions of phages with human cells.
For example, it has been shown that phages are able to interact with human mucosal surfaces and
form a non-host derived immune barrier, as well as that they may play a significant role in clinical
transplantation as it was shown in mice that they reduce cellular infiltration of allogenic skin
allografts.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section V: Phage Therapy (Part I) 49
Assessment of immunomodulation and serum neutralizing antibodies during
bacteriophage therapy in an A. baumannii mouse wound infection model
Michael D. Rouse1*, Anna Jacobs2, Biswajit Biswas3, LT James Regeimbal4, Matthew Henry3,
LCDR Theron Hamilton3, Jessica A. Roman1, Josh Stanbro1, Yonas Alemnah2, MAJ Jonathan
Shearer2, MAJ Samandra Demons2, LTC Stuart D. Tyner2, LT Chaselynn Watters1, CDR Michael
G. Stockelman1, LCDR Mark P. Simons1
1Naval Medical Research Command Silver Spring, MD, 2 Walter Reed Army Institute of Research
Silver Spring, MD, 3 Biological Defense Research Detachment FT Detrick, MD, 4 Naval Medical
Research Unit – 6 Lima, Peru
*E-mail: [email protected]
Objectives: The spread of multidrug antibiotic resistance (MDR) is a widely-recognized crisis in
the treatment of bacterial infections, and ranks in top tier of high operational risk and infectious
disease threats. Recently, the World Health Organization published its first ever list of antibiotic-
resistant "global priority pathogens": a catalogue of 12 genera of bacteria that pose the greatest
threat to human health, with Acinetobacter baumannii listed in the “Priority 1: Critical” category of
pathogens. Lytic bacteriophage cocktails specifically targeted against each of the high priority
bacterial pathogens are promising antimicrobial candidates; however, the possibility of host
immune responses interfering with phage therapy is a concern that must be investigated.
Methods: BALB/c mice were pre-immunized with control (PBS) or bacteriophage cocktail of 5
diverse phages previously shown to lyse A. baumannii strain AB5075. After 3 weeks, mice were
immunosuppressed with cyclophosphamide before wounding (dorsal full thickness injury) and
challenged topically with AB5075-lux. Following infection, mice were treated with PBS or the
phage at 4h post-infection and every 24h for 2 days. Mice were then evaluated for 3 more weeks to
assess the safety and efficacy of the bacteriophage treatment relative to the placebo. We assessed
mortality, bacterial burden, wound closure, systemic and local cytokine profiles, and alterations in
host cellular immunity, including serum neutralizing antibodies.
Results: While pre-immunization with bacteriophage cocktail did not modulate circulating
cytokines, it did lead to a significant increase in total immunoglobulins, specifically Ig2A and
IgG2b compared to control. Furthermore, we found that these immunoglobulins were capable of
specifically binding and neutralizing our phage cocktail from lysing the targeted bacteria. Our data
demonstrate that bacteriophage administration also decreased circulating neutrophils, while
elevating monocytes within 24h. Such effects were observed to last for 2 weeks following the first
phase of pre-immunization; however, no significant changes were observed in immune cell
populations of secondary lymphoid organs, including spleen and lymph nodes. Administration of
bacteriophage treatment effectively prevented wound expansion as assessed by bioluminescence.
Importantly, despite moderate immunological changes associated with pre-immunization, there was
no statistical difference observed in therapeutic efficacy of phage cocktail between phage-
immunized or control-immunized groups.
Conclusion: Our data demonstrate that wound infection outcome does not appear to be impaired
by prior exposure to the therapeutic phage. These findings are an important step in development
of personalized phage therapy that will continue to be studied in other animal infection models.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
50 Section V: Phage Therapy (Part I)
Faecal Virome Transplants (FVTs) reshape the murine microbiota after
antibiotic perturbation
Lorraine A. Draper1,2, Feargal J. Ryan1,2, Marion Dalmasso1,2,†, Pat G. Casey1,2, Angela
McCann1,2, Vimalkumar Velayudhan 1,2, R. Paul Ross1,3, Colin Hill1,2*
1 APC Microbiome Institute, University College Cork, Cork, Ireland. 2 School of Microbiology, University College Cork, Cork, Ireland
3 Teagasc Food Research Centre, Moorepark, Fermoy, Co. Cork, Ireland
†Present address: Normandie Univ, UNICAEN, ABTE, 14000 Caen, France
*E-mail: [email protected]
Background: It has become increasingly apparent that creating and maintaining a complex and
diverse gut microbiome is fundamental to human health. Thus, there are increasing efforts to
identify methods that can modulate and influence the microbiome, especially in those who, due to
disease or circumstance, have had their native microbiome disrupted. Faecal microbial
transplantation (FMT) is one such method and it has been shown to replenish the microbiome of an
individual with that of a healthy donor’s. In which case, the microbiome as a whole is reintroduced
into the recipient i.e. bacterial, archaeal, fungal and viral species. Within this study we determined
if the faecal virome of which bacteriophages predominate could alone transform an individual’s
microbiome via a Faecal Virome Transplant (FVT).
Methods: The current study describes how the administration of virus-like particles (VLPs) in the
form of an FVT influenced and reshaped the murine microbiome of test mice following microbial
perturbation with antibiotics. Subsequent to administration of penicillin and streptomycin, test mice
(n=8) received a bacteria free-VLP enriched faecal transplants, while control mice (n=8) received a
heat-treated nuclease treated version of the same.
Results: Mice that received FVTs, in which bacteriophages predominate, had a bacteriome (as
determined via 16S rRNA sequencing) that separated into distinct groups via principle co-ordinate
analysis (PCoA) and contained differentially abundant taxa when compared to the control group.
Additionally through metagenomic sequencing of Virome DNA, we were able to obtain a snapshot
of the putative viruses present in the gut of test and control mice at various time points. It was
observed that viral content appeared to differ in both abundance and diversity.
Conclusions: This study has validated the role of FVTs in gut microbiome population dynamics.
There is potential to take advantage of such, especially with the increasing trend toward performing
FMTs. Up until now these have focussed on transferring living bacteria and spores; as
bacteriophage are non-living proteinaceous entities they could form a robust, inexpensive
alternative that is susceptible to standardisation and could be delivered as a freeze dried
formulation.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section V: Phage Therapy (Part I) 51
Clostridium difficile-associated Bacteriophage profiles in Faecal Microbiota
Transplant Patients
David Speicher
McMaster University, Hamilton, Ontario, Canada
Clostridium difficile infection (CDI) is a serious hospital-associated infection, and relapsing or
refractory CDI may respond only to faecal microbiota transplantation (FMT). At St. Joseph’s
Healthcare Hamilton, we have performed over 1000 FMTs since 2010. In a recent study
comparing fresh vs frozen faecal samples delivered rectally, FMT was successful in 80% of
patients with 2 to 6 treatments, but 20% of cases never resolve, even after multiple FMTs.
Comparable results have been obtained using lyophilised faecal samples. The rapid clinical
response, usually in 2-5 days but in as little as 24 hours, and the rapid clearance of C. difficile
following FMT suggests that something other than bacterial recolonization is at work, presumably
phages. To elucidate the role of phages in recovery, we are currently comparing whole genome
sequences from C. difficile isolates from diagnostically confirmed CDI and asymptomatic carriers,
and scrutinizing the sequences for prophage DNA. We have also identified several myoviruses
sequences in samples from diagnostically confirmed CDI, albeit at much lower concentrations than
C. difficile. Understanding the role and profiles of phages may be critical to further improving
FMT efficacy and for making it available more widely in future.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
52 Section V: Phage Therapy (Part I)
Efficacy of an optimised phage cocktail to clear Clostridium difficile in a gut
fermentation model
Janet Y. Nale1, Tamsin T. Redgwell2, Andrew Millard2 and Martha R. J. Clokie1*
1Department of Infection, Immunity and Inflammation, University of Leicester, Leicester, LE1
9HN, UK, 2Division of Biomedical Sciences, Unit of Microbiology and Infection, University of
Warwick, Coventry, CV4 7AL, UK
*E-mail: [email protected]
Clostridium difficile infection (CDI) is a significant cause of infectious diarrhoea and is responsible
for death in 10% of cases, and treatment is limited. Conventional antibiotics are ineffective for all
ribotypes or could trigger dysbiosis and resistance leading to recurrent infection. There is also
significant cost associated with the newest available antibiotic that has limited its use. Thus, there
has been a shift of interest towards exploring novel antimicrobials to either replace or supplement
antibiotics.
We have previously isolated a large bank of bacteriophages and described the activity of an
optimised 4-phage cocktail on C. difficile in vitro using cultures and biofilms, and in vivo using
hamster and wax moth larvae CDI models. Although the data obtained from these models provides
novel insights into the therapeutic applications of the phages, more information is needed to
determine their specificity and efficacy in the presence of human gut microbiome.
Here, we describe the activity of the cocktail using an in vitro human gut fermentation CDI model
with freshly voided fecal samples from 4 healthy volunteers covering diverse ethnicity and age
groups. The bacterial communities present in each sample were assayed on selective media prior to
mixing in equal proportions. The combined fecal slurry was added to a minimal medium and
treated prophylactically or remedially with phages and bacteria over a time course anaerobically.
We observed 6 and 1-log reductions in C. difficile counts in the prophylaxis and remedial regimens
respectively within the first 5h post-infection and complete eradication of bacteria at 24h in both
regimens. C. difficile remained undetected from this time till the experiment was terminated at 72
hour. Commensal Enterococci, Bifidobacteria, Lactobacilli, total Anearobes and
Enterobacteriaceae were not affected by both regimens. However, the phage control showed ~2
logs increase of the total Anaerobes and Enterobactericeae counts compared to the regimens and
the bacterial/untreated controls. Comparable impact of the phage treatments on culturable and
unculturable components of the microbiota was ascertained using metagenomics analysis.
This data supports the prophylactic application of the phages to treat CDI. The elevated levels of
specific commensals in the phage-treated control could be used to prevent colonisation of C.
difficile and provide protection from the infection.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section V: Phage Therapy (Part I) 53
Bacteriophage penetrate the body via epithelial transcytosis
Sophie Nguyen1, Kristi Baker2, Benjamin S. Padman3, Thomas A. Weston1, Kyle Schlosser1,
Barbara Bailey4,5, Michael Lazarou3, Antoni Luque4,5,6, Forest Rohwer1,5, Richard Blumberg2,
Jeremy J. Barr1,5,7*
1 San Diego State University, Department of Biology, San Diego, CA 92182, USA 2 Division of Gastroenterology, Hepatology and Endoscopy, Brigham and Women’s Hospital,
Harvard Medical School, Boston, MA 02115, USA 3 Monash University, Department of Biochemistry and Molecular Biology, Biomedicine Discovery
Institute, Melbourne, VIC 3800, Australia 4 San Diego State University, Department of Mathematics & Statistics, San Diego, CA 92182,
USA; 5 Viral Information Institute, San Diego State University, San Diego, CA 92182, USA 6 Computational Science Research Center, San Diego State University, San Diego, CA 92182,
USA; 7 Monash University, School of Biological Sciences, Melbourne, VIC 3800, Australia
*E-mail: [email protected]
Bacterial viruses are amongst the most numerous biological entities within the human body. These
viruses are found within regions of the body that have conventionally been considered sterile,
including the blood, lymph and organs. However, the primary mechanism that bacterial viruses use
to access the body remains unknown. Here we use in vitro studies to demonstrate the rapid and
directional transcytosis of diverse bacteriophages across confluent cell layers originating from the
gut, lung, liver, kidney and brain. Bacteriophage transcytosis across cell layers had a significant
preferential directionality for apical-to-basolateral transport, with approximately 0.1% of total
bacteriophage applied being transcytosed over a two hour period. Microscopy and subcellular
fractionation revealed bacteriophages accessed microsomal and cytosolic compartments of the
eukaryotic cell, passaging via the Golgi apparatus before being exocytosed. We posit that the
human body is continually absorbing phages from the gut and transporting them throughout the
epithelial cell structure and subsequently the body. From our experimental results we estimate that
thirty-one billion bacteriophage particles are transcytosed from the gut into the average human
body each day. The transcytosis of bacteriophage is a natural and ubiquitous process that enables
direct interactions between bacteriophages and the cells, organs, and immune systems of the human
body.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
54 Section V: Phage Therapy (Part I)
Murine Model of Bacteriophage Therapy for the Decolonization of
Vancomycin-Resistant Enterococci
Alyxandria M. Schubert, and Paul E. Carlson Jr.
Division of Bacterial, Parasitic and Allergenic Products, Office of Vaccines Research and Review,
Center for Biologics Evaluation and Research, U.S. Food and Drug Administration, Silver Spring,
MD
*E-mail: [email protected]
Since the advent of antibiotics in the 1920s, these drugs have saved millions of people from
diseases such as pneumonias, healthcare associated infections, and foodborne illnesses. However,
the continued use of antibiotics has led to several unintended consequences, including disruption of
the indigenous beneficial gut bacteria and a rise in antibiotic-resistant bacteria. The Centers for
Disease Control and Prevention (CDC) estimates that 23,000 deaths are caused by antibiotic-
resistant bacteria each year in the United States, and these organisms constitute a growing problem
worldwide. Vancomycin-resistant enterococci (VRE), which have been classified as a serious
threat by the CDC and World Health Organization, are responsible for 20,000 U.S. infections
annually. The inability to treat these infections with common antibiotics necessitates the
development of alternative methods of intervention.
The objective of this investigation was to develop and characterize an effective bacteriophage
therapy against VRE, including both vancomycin-resistant E. faecalis and E. faecium. To that end,
several naturally occurring phage isolates with activity against a range of VRE strains were isolated
from sewage. In a mice model, various antibiotics were used to disrupt the normal gut microbiota
and allow VRE to colonize and persist. A cocktail of lytic phages with high activity against the E.
faecalis strain used to infect mice. This cocktail and the constituent individual phages were
administered to VRE colonized mice to study their efficacy and the dynamics of phage resistance.
Preliminary data suggests that a phage cocktail could be successfully utilized to decolonize VRE-
infected mice. This treatment will be compared with a standard antibiotic intervention, ampicillin,
for differences in efficacy as well as their effects on the microbiota. We hypothesize that
bacteriophage therapy will have minimal effects on the microbiota compared with antibiotic
treatment. These bacteriophage therapy investigations will have a significant impact on a largely
understudied field and contribute to solving the antibiotic-resistant bacteria problem.
This work was funded by CBER/FDA and an Interagency Agreement with DMID/NIAID
(AAI15021-001-02000).

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section V: Phage Therapy (Part I) 55
Phage application in therapy of different local infections: advances and
failures
V.V. Morozova, Yu.N. Kozlova, E.A. Yakovets2, N.V. Tikunova
Institute of Chemical Biology and Fundamental Medicine SB RAS, Novosibirsk, Russia
*E-mail: [email protected]
The traditional delivery method for phage therapy is direct application to the local site of infection.
Here, we describe four successful cases of localized phage therapy and two cases of ineffective
local phage application.
Two cases of local phage application to treat a nasopharyngeal S. aureus infection led to clinical
cure of patients. One more case of successful phage therapy was the treatment of a newborn girl
suffering from bronchititis associated with P. aeruginosa infection. Piobacteriophage and
Sextaphage preparations were applied together by inhalation. Pseudomonas aeruginosa was
eliminated and now the newborn recovered.
The treatment of polymicrobial urinary tract infection was carried out in cooperation with the
Centre of New Medical Technologies (Novosibirsk). A patient (51 years old woman) suffered from
a chronic urinary tract infection after surgical operation to eradicate a cancer. Multiple courses of
antibiotics were applied, and as a result, multidrug resistance strains were acquired. Five clinical
strains were cultured from the urine sample and phages were obtained to kill all the strains. Phages
were applied by urinary catheter and per rectal for seven days. Two strains (E. coli and K.
pneumoniae) were revealed a week after treatment: the K. pneumoniae strain was highly sensitive
to Piobacteriophage and the E. coli was sensitive to amoxiclav. A complex treatment with
piobacteriophage (per os + per rectum) in combination with amoxiclav was carried out for the next
10 days. A week after treatment no microbes were cultured from the urinary samples.
A patient (75 years old man) suffering from otitis showed only partial microbiological cure. Two
strains of P. aeruginosa were cultured from his ear, both sensitive to P. aeruginosa phage.
However, the treatment eliminated only one of the strains. The application of phage treatment after
an infection developed post orthopedic surgery was also unsuccessful. The negative outcome
suggests this direct method of application was inadequate.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
56 Section V: Phage Therapy (Part I)
Updates on Yersinia phages and on phage therapy initiative in Finland
Mikael Skurnik
Department of Bacteriology and Immunology, Medicum, and Research Programs Unit,
Immunobiology, University of Helsinki, and Helsinki University Central Hospital Laboratory
Diagnostics, Helsinki, Finland
E-mail: [email protected]
Genus Yersinia belongs to Enterobacteriaeae and includes 17 species of which 4 cause diseases to
animals. Most famous and notorious pathogen is Y. pestis the causative agent of bubonic and
pneumonic plague, while Y. pseudotuberculosis and Y. enterocolitica cause mainly enteric
infections. Y. ruckeri is a fish pathogen causing the red mouth disease. We have isolated Yersinia –
specific phages from various sources and sequenced altogether 34 phages.
I will present new data on a couple of the phages and on application of phages in food biocontrol. I
will also summarize our efforts towards first phage therapy trials in Finland

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VI: Phage Therapy (Part II) 57
Phage Therapy: the Polish Experience
Ryszard Międzybrodzki1,2,3, Beata Weber-Dąbrowska1,2, Sławomir Letkiewicz1, Wojciech
Fortuna1,2, Paweł Rogóż1, Andrzej Górski1,2,,3*
1Bacteriophage Laboratory, Hirszfeld Institute of Immunology and Experimental Therapy,
Polish Academy of Sciences, Wrocław, Poland 2Phage Therapy Unit, Hirszfeld Institute of Immunology and Experimental Therapy,
Polish Academy of Sciences, Wroclaw, Poland 3Department of Clinical Immunology, Medical University of Warsaw, Warsaw, Poland
*E-mail: [email protected]
Tradition of the therapeutic use of bacteriophages in Poland is almost as long as their discovery by
Twort and d’Hereulle. The first report on their therapeutic use in the treatment of diarrhea dates to
1923. Since that time phage therapy was undergoing continuous evolution from standard through
nonstandard to therapeutic treatment due to increasing standards of current pharmaceutical law and
ethical regulations. Since 1970s Hirszfeld Institute became a main center supplying phages and
supervising the phage therapy in Poland. Reports of its phage group, published between 1980 and
2000, on the clinical effects of the phage therapy of hundreds of patients were the most cited papers
on the clinical phage therapy in the English literature.
At the end of 2005 Hirszfeld Institute opened its own Phage Therapy Unit, so far the only such
center among the European Union countries. Since that time almost 600 patients with different
chronic bacterial infections resistant to antibiotic treatment were qualified for the phage therapy
and treated according to a detailed protocol approved by an independent bioethical committee. This
protocol was subjected to periodical changes including the most recent one applying phage therapy
in children older than 6 years, and the possibility of application of phage preparations by
intravesical instillations in patients with urinary tract infections. Monitoring of the production of
anti-phage antibodies was also introduced. We have possibility to apply in our patients not only
monovalent phage lysate but also cocktails of staphylococcal phages as well as their purified
preparations. Our general results show that a good response to the treatment could be achieved in
approximately 40% of the patients which we consider as very encouraging result for future clinical
trials, since it was achieved in a group of patients with complex clinical problems. We follow the
principle that our primary aim is to treat the patient and not only try to eradicate infection at all
cost. Moreover, our recent studies open new perspectives for phage therapy in immunomodulation.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
58 Section VI: Phage Therapy (Part II)
Phase 1 studies to evaluate the safety, tolerability and preliminary
effectiveness of a novel bacteriophage-based treatment, AB-SA01
Sandra Morales
On behalf of the team members at AmpliPhi Biosciences, Queen Elizabeth Hospital, Adelaide, SA,
Australia, Walter Reed Army Institute of Research, Silver Spring, MD, USA
*E-mail: [email protected]
In 2016, AmpliPhi completed two clinical studies assessing the safety, tolerability and preliminary
effectiveness of a novel bacteriophage-based treatment, AB-SA01, when applied to the skin of
healthy volunteer subjects and to the nasal cavities and sinuses of subjects with chronic
rhinosinusitis (CRS) associated with Staphylococcus aureus. AB-SA01 is a therapeutic
bacteriophage mixture of three well-characterised individual S. aureus phages produced under
current good manufacturing practice (cGMP) in AmpliPhi’s own GMP certified facility.
The study on the skin of healthy volunteers was conducted in the USA, in collaboration with
colleagues at the Walter Reed Army Institute of Research, under an Investigational New
Drug Application (IND#16820). The study on patients suffering from CRS associated with S.
aureus, (PJPhage_SA01), was conducted at the Queen Elizabeth Hospital in Adelaide, South
Australia. Overall, AB-SA01 has been shown to be safe and well tolerated in the populations and
dose levels tested in the studies conducted to date. In the CRS study of patients with S. aureus,
reductions in S. aureus counts were observed over the course of treatment. A summary of the
results and practical lessons learnt from these studies will be presented.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VI: Phage Therapy (Part II) 59
Our experience in personalized phage therapy of patients with diabetic
ulcers
Tikunova, N.V.,* Morozova, V.V., Kozlova, Yu.N., Ganichev, D.A., Vlassov, V.V.
Institute of chemical biology and fundamental medicine SB RAS, Novosibirsk, Russia;
630090, Lavrent’ev av. 8, Novosibirsk,
*E-mail: [email protected]
Antibiotic-resistant bacteria become a global threat now. A promising approach to solving the
problem is the use of bacteriophages - viruses that can destruct bacteria, but not able to infect
humans. Bacteriophages do not cause side effects and are not toxic. Unlike antibiotics,
bacteriophages are mostly species- and strain-specific and therefore do not disrupt the normal
human microbiome. In Russia, there are approved phage preparations and cocktails, but most of
them were developed in the last century and their characteristics do not meet recent requirements.
In addition, the main criterion for the application of therapeutic bacteriophage should be sensitivity
of bacterial agent to the bacteriophage. This requires a fast and qualitative microbiological analysis,
rapid selection of the appropriate therapeutic phage or a cocktail, and microbiological control of
treatment. In ICBFM SB RAS, the approach to personalized implementation of phage therapy was
developed and tested. A number of original and unique bacteriophages that are effective against a
broad spectrum of bacteria were isolated and characterized. Several new phage cocktails, including
unique polyspecific cocktail containing bacteriophages against 11 species of pathogenic and
nosocomial bacteria were elaborated. Clinical data on personalized application of phage
preparations for the treatment of diabetic foot ulcers were analyzed. The use of phage therapy in
the clinic provided the healing of most patients, whose diseases were caused by resistant bacteria
and previous antibiotic therapy was unsuccessful.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
60 Section VI: Phage Therapy (Part II)
Working Collaboratively to Jump-start Phage Therapy
E. M. Kutter1,2*, R. Fish2, S. Langevin3 G. Wheat2, B. Blasdel2, D. Bryan1,2, M. Kutateladze4, R.
Adamia4, Sarah Kuhl5
Evergreen State College1 and Phagebiotics Research Foundation2, Olympia, WA; University of
Washington3; Eliava Institute, Tbilisi, Republic of Georgia4;
*E-mail: [email protected]
Phage are increasingly being considered as part of a desperately needed solution to the
growing crisis of antibiotic resistance around the world. Progress toward successful clinical trials
and governmental approval is still frustratingly slow for a variety of reasons. A major public-health
problem lending itself to physician-initiated double-blind Staphylococcus phage trials is the
treatment of diabetic foot infections, where vascular and immune problems limit antibiotic
effectiveness and treatment frequently fails even with non-MRSA Staphylococcus. Infection often
extends into the bone and leads to amputation, even when treatment with antibiotics begins early.
Here we present our data on a set of compassionate-use cases in which the sequenced,
highly purified commercial Eliava Institute Staphylococcus phage Sb1 was added to standard
treatment after the failure of appropriate antibiotics for diabetic toe ulcers. As antibiotic treatment
and good surgical debridement had failed, amputation was the only standard alternative. The fact
that all cases of previously intransigent wounds, most with bone involvement, went on to heal and
none required amputation is a remarkable result for clinicians who know the unfortunate natural
history of such infections. The treatment consists of infusing packing gauze with the
Staphylococcus phage product after routine debridement. Localized injections were also
incorporated for those cases involving osteomyelitis or joint infection. It is remarkably easy to
apply phage treatment in routine practice and such treatment appears to be safe and leads to healing
of intransigent chronic wounds, reducing the rate of amputation. Phage treatment appears to have
the potential to improve the quality of care and decrease the costs of amputation, rehabilitation and
chronic wound management. We are now beginning to use metagenomics to analyze the microbial
composition of such diabetic toe wounds and try to understand why using phage targeting only
Staphylococcus has been so successful.
We propose that S. aureus phage targeting diabetic foot ulcers be the subject of a public-
private-academic initiative, moving quickly toward controlled clinical trials. Much data is already
available on Staphylococcus phage wound treatment, from the 1921 first phage therapy paper, to
Ward MacNeal’s US work in the 1940’s, to the extensive French, Polish and Georgian clinical
work.
Diabetic ulcer treatment occurs largely in civilian and military wound-care centers where
standardized monitoring and well-defined protocols facilitate cross-institutional data collection for
double-blind trials. No other variation from standard of care would be needed. Particular features
of the large, purely lytic Staphylococcus phage K genus are ideal for phage therapy. Many hit over
90% of clinical Staphylococcus isolates and all tested MRSA strains, provoke very little resistance,
and have been widely used both as monophages and in French, Georgian, Russian and Belgian
therapeutic cocktails. Two such phages were included in the first US FDA-approved phase 1
phage trial, in Lubbock, Texas. Broad US safety data comes from the decades-long human use of
Staphylococcus Phage Lysate (SPL), still marketed as a veterinary immune stimulant against
Staphylococcus. We have found SPL to contain as high a phage concentration as commercial phage
cocktails.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VI: Phage Therapy (Part II) 61
Clinical case studies in Romania of Eliava Pyophage and Intestiphage
Alina Cristina Neguț, Oana Săndulescu, Maria Magdalena Motoi, Anca Streinu-Cercel, Adrian
Streinu-Cercel
National Institute for Infectious Diseases Prof. Dr. Matei Balş, Romania
*E-mail: [email protected]
Background: Chronic infections have become of worldwide interest not only due to antibiotic
resistance and biofilm formation but also due to the presence of persister cells that are tolerant to
antimicrobials. New antimicrobials for combating this phenotype are needed. Can bacteriophages
be a solution?
Methods: We have performed an experimental study in the National Institute for Infectious
Diseases. Our group, in collaboration with Prof. Dr. Matei Balș have treated 7 patients with chronic
infections using antibiotics and bacteriophages in combination. The patients included in this study
suffered hard-to-treat infections due to antibiotic resistant bacteria, biofilm formation or hard to
sterilize infection sites and received antibiotics and commercial Georgian phage cocktails:
PyoPhage or IntestiPhage (Eliava BioPreparations, Tbilisi, Georgia). Before initiating therapy, we
evaluated the isolated microorganism for both antibiotic and phage susceptibility.
Results: We have included into the study the following cases: recurrent endocarditis with
Staphylococcus aureus, chronic hip osteomyelitis with S. aureus and P. aeruginosa, periprosthetic
dorsolumbar soft-tissue infection with S. aureus, chronic cutaneous infection with S. simulans, E.
coli and P. aeruginosa, axillary hidradenitis with Proteus mirabilis and S. epidermidis, chronic
osteomyelitis of the thigh-bone with S. aureus and chronic osteomyelitis of the humerus with S.
aureus. The combined therapy proved to be safe and the patients had no adverse reactions or
changes in laboratory safety parameters. A total of 6 patients had negative cultures during therapy
but only one case achieved sustained bacteriological response with clinical cure, the others relapsed
after stopping the therapy. For the patient with recurrent endocarditis the timespan between
relapses was longer when receiving combined therapy compared with antimicrobial treatment
alone. The sustained bacteriological response was achieved for the patient with periprosthetic
dorsolumbar soft-tissue infection with methicillin susceptible S. aureus. The case was considered
hard-to-treat due to a history of multiple drug allergy syndrome, presence of multiple foreign
bodies (metallic rods and screws), with biofilm formation and unsuccessful long-term antibiotic
therapy.
Conclusions: This is the first Romanian pilot study, since 1975, treating patients with combined
therapy antibiotics and bacteriophages. The results confirmed the safety and the importance of
future research in this domain.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
62 Section VI: Phage Therapy (Part II)
Intravenous Application of Phage Therapy to Treat a Terminally Ill Patient
Who Was Infected with Multi Drug Resistant A. baumannii.
Biswajit Biswas1,2*, Robert Schooley4, Matthew Henry1,2, Javier Quinones1,2, Luis Estrella1, David
Wolfe1, Robert K. Pope5, Matthew Doan1, James Regeimbal3, Eric Hall3, Alfred Joseph Mateczun1
and Theron Hamilton1
1 Biological Defense Research Directorate, Naval Medical Research Center – Frederick, 84000
Research Plaza, Fort Detrick, MD; 2 Henry M. Jackson Foundation for the Advancement of
Military Medicine, 6720-A Rockledge Dr, Suite 100, Bethesda, MD; 3 Naval Medical Research
Center, Infectious Diseases Directorate 503 Robert Grant Blvd, Silver Spring, MD 20910; 4
University Of California, San Diego, CA; 5 National Bioforensic Analysis & Countermeasures
Center, Fredrick, MD.
*E-mail: [email protected]
Current global surveillance indicates that multi drug resistant (MDR) bacteria are emerging at an
alarming rate. There is also a significant concern about the potential of generating highly virulent
microorganisms using genetic engineering and synthetic biology. With the growing problem of
rapidly occurring and spreading antimicrobial resistance, alternative treatment must be sought and
developed. Phages or bacterial viruses are the most abundant biomolecules on the surface of the
earth and known to be bacterial killers. Furthermore without any partiality phage will kill both
MDR and non-MDR bacterial pathogens. In this regard a properly formulated phage cocktail
would be highly effective for overcoming antimicrobial resistance (AMR) of any bacteria.
Currently we are developing phage therapy to treat MDR bacterial infections in human. In this
project we invented a rapid process to select combination of broad spectrum phages which can
overcome the emergence of phage resistance bacteria and ultimately enhance the therapeutic
efficacy of the phages. Recently we received a request from University of California, San Diego
(UCSD) to provide cocktail phages which are enabled to kill an MDR A. baumannii clinical isolate
called the TP strain. We immediately engaged to evaluate the killing efficacy of 98 A. baumannii
phages from our library on the TP strain using our proprietary liquid assay system. Our rapid assay
results indicated that the phage library contained 10 very virulent phages which very effectively
killed the TP strain. After further analysis using our proprietary system we selected 4 phages to
prepare cocktail which can overcome the emergence of phage resistance when used of TP strain.
Selected phages were amplified on TP strain and purified using cesium chloride density gradient
technique to prepare phage cocktail. The cocktail was sent to UCSD where physician used the
phage cocktail to treat a terminally ill patient who was infected with MDR A. baumannii. This
phage therapy was done as an FDA-approved, off-use intravenous infusion to treat patient’s
systemic infection. Within 48 hours of intravenous administration of phage cocktail the patient
came out of his coma and within 7 days patient’s temperature and white blood cell count became
normal and his creatinine level improved significantly. Additionally phage therapy also re-
sensitizes the TP strain to several antibiotics to which it was previously acquired resistant. The
detail of this phage therapy study will be presented here.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VI: Phage Therapy (Part II) 63
Phage Therapy and Strategies for Treatment of Bacterial Infections
Randall L. Kincaid
National Institute of Allergy and Infectious Diseases, NIH Rockville, MD 20892
E-mail: [email protected]
Clinical interventions using bacteriophages have been employed in some parts of the world for
more than 80 years, although such use has not been widely adopted by Western medicine.
Nonetheless, phage therapy is being viewed increasingly as a viable alternative strategy for
treatment of antibiotic-resistant infections, owing to the fundamentally different mechanisms by
which phage kill bacteria. Phages have co-evolved with our commensal bacteria and may also
provide protection from infection through their localization on mucosal surfaces; thus, they can be
viewed as a part of our natural microbiome and are unlikely to pose substantial safety risks to the
patient. Phages represent the most abundant form of “life” and constitute an almost inexhaustible
source of anti-infective options – a virtual pharmacopeia of medically-relevant solutions.
There are several challenges to broad adoption of phage therapy: 1) well-controlled clinical studies
are needed to establish efficacy of phage products, 2) the processes for the manufacture, assay and
formulation of phage preparations require characterization and standardization to ensure products
that are reliable and 3) medical indications for phage therapy that represent clear unmet needs must
be identified. These are not insurmountable issues but they require longer-term strategies to create
products that will meet regulatory expectations and provided clear medical benefit over existing
treatments. These challenges are very similar to those see for all new therapeutic and prophylactic
agents, and require product-specific planning and the design of appropriate clinical studies.
An overview of these planning needs will be presented at the meeting, with emphasis on the role
that the government may play. This includes a discussion of fundamentally different models for
phage therapy (treatment centers vs products), strategies for optimizing the effectiveness of phage
therapy formulations, product standards that might be needed to create repositories of phage and
the specific indications that would be of most value to the infectious disease community. We will
also consider paradigms for prevention of infectious disease, both in medical and community
settings. Lastly, we will consider the prospects for future investment in phage-based solutions,
both from the commercial and from government sectors.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
64 Section VII: Molecular Biology
The implications of the dynamic interchange between killing a cell and letting
it hang itself for picking the right phages for therapy.
Bob Blasdel1,Hanne Hendrix1, Jeroen De Smet1, Rob Lavigne1*
1Laboratory of Gene Technology - KU Leuven, Leuven, Belgium
*E-mail: [email protected]
By performing RNA-Seq on the non-ribosomal RNA of synchronously infected Pseudomonas
aeruginosa cells we are able to compare host transcript data for infected cells to an uninfected
control with eight fundamentally different lytic phages. With data for so many distantly related
phages, we were able to highlight the host-mediated transcriptional stress responses to phage
infection by defining the genes that are significantly differentially expressed in common between
all of the different infections. Within this stress response we have discovered a number of
fundamentally new defenses used by P. aeruginosa PAO1 to inhibit phage infection. These include
a quorum sensing based mechanism using the Pseudomonas Quinolone Signal (PQS), has been
previously demonstrated to control an oxidative stress response encouraging metabolic dormancy,
to reduce infection vigor in sister bacteria by daughter phage.
At the same time, having defined the host-mediated transcriptional stress response to phage
infection, we can also define the phage-specific differential expression resulting from phage-
mediated stresses or molecular mechanisms. Indeed, we have found that by leaving the host
transcriptional apparatus intact, each of these phages appears to be able to direct the expression of
the host genome towards phage ends, using the host genome as if it were an auxiliary phage
genome. For example, we have found that PAK_P3 appears to rely on host machinery that it
expresses to manipulate the RNA metabolism of the phage-infected cell, while both YuA and
PAK_P4 impose an iron stress transcriptional response on their hosts. Allowing transcription of the
host genome to proceed during infection likely allows phage-infected cells to adapt to new
metabolic conditions, reduces latent periods, and increases burst sizes as evidenced by the impact
of rifampicin on infections by giant phage that do not require the host RNA polymerase to
transcribe their own genomes but appear to be aided by the transcription of the hosts’.
However, they may also have reduced host ranges due to the host defenses they allow to function,
and could also lead to less predictable results in patients, leading to interesting questions about
whether they may be the best fit for phage therapy. Indeed, like some antibiotics, we have observed
that each of the eight phages we have studied activates the transcription of at least one prophage in
P. aeruginosa PAO1 or PAK. Similarly, we have observed that infection by LUZ19 upregulates the
expression of the hcnABC operon that is responsible for the production of hydrogen cyanide, one
of the primary virulence factors of P. aeruginosa. By choosing phages that inhibit the expression of
the host, as well as other competing genetic elements within the cell, or by manipulating phages to
do so we can hope to gain more control over what happens within the infected cell – reducing the
chance of encouraging horizontal gene transfer or the activation of virulence factors with phage
therapy.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VII: Molecular Biology 65
The phage T4 MotA transcription factor contains a novel DNA binding motif
that specifically recognizes modified DNA
Maxime G. Cuypers1, Rosanna M. Robertson1, Leslie Knipling2, M. Brett Waddell1, Deborah M.
Hinton2* and Stephen W. White1
1Department of Structural Biology, St. Jude Children’s Research Hospital, Memphis, TN 38105,
USA 2Gene Expression and Regulation Section, Laboratory of Cell and Molecular Biology,
NIDDK, National Institutes of Health, Bethesda, Maryland 20892, USA
*E-mail: [email protected]
Bacteriophage T4 temporally controls expression of its genome, giving rise to early,
middle, and late transcripts. T4 uses E. coli RNA polymerase throughout this process, but encodes
factors that change the specificity of the polymerase, switching recognition from early to middle to
late promoters. Like all bacterial polymerases, E. coli RNA polymerase is composed of a core that
synthesizes RNA (beta, beta’, 2 alphas, and omega) plus a sigma specificity subunit that recognizes
the promoter sequence. While recognition of T4 early promoters does not require additional
factors, activation of middle promoters requires the T4 co-activator AsiA and the T4 activator
MotA in a process called sigma appropriation.
AsiA remodels the C-terminal region of sigma, MotANTD interacts with sigma C-terminal
residues, and MotACTD binds to the middle promoter motif, the MotA box,
5’(a/t)(a/t)(a/t)TGCTTtA3’ centered at -30. Using structures (RNA polymerase, MotACTD,
MotANTD, and AsiA/sigma) and cleavage analyses obtained with FeBABE–conjugated MotA, we
generated a structure-based model for sigma appropriation, which indicates the positions of the
proteins relative to each other and to the DNA within the competent transcription complex (James
et al. NAR 2016 44, 7974). We have now determined the crystal structure of the MotAlinker-CTD with
MotA box DNA. The structure reveals a new type of protein/DNA interaction, with the saddle-
like, double-wing motif of MotACTD lying within the major groove and the arginine-rich MotAlinker
lying within the upstream minor groove. MotACTD binds DNA primarily through non-specific
interactions with the DNA backbone although there are two specific interactions with the MotA
box in the major groove. The MotAlinker plays a key role in stabilizing the complex via minor
groove interactions. The structure is remarkably similar to that of our transcription model,
justifying the use of FeBABE analyses to map protein/DNA interactions at a high resolution.
Furthermore, the structure predicted that the hydroxymethyl, glucosylation modification present on
cytosines within T4 DNA might provide an important MotA/DNA interaction. Using EMSAs and
SPR, we find that this modification on the nontemplate strand substantially improves binding,
primarily by significantly decreasing the off rate.
Although AsiA and MotA analogs have been identified in dozens of T4-like phages, a
process similar to sigma appropriation has not been observed outside phage. However, the novel
double-wing structure has been reported for two proteins of unknown function, the conserved E.
coli YjbR and the Pseudomonas syringae protein Pspto_3016. Thus, understanding how MotA
interacts with its partner DNA may yield insights into the functions of these proteins and other
unidentified members of the family. Another protein that contains the double wing motif is the T4
SF2 helicase UvsW, which has a key role in T4 recombination and is functionally and structurally
related to the eukaryotic Rad54 protein. The MotAlinker/CTD/DNA complex suggests a role for the
UvsW double wing structure. We speculate that the largely non-specific interactions allow both
MotA and UvsW to search for their motifs (the MotA box motif and 3- and 4-way junction DNA,
respectively) by ‘riding along’ the DNA via the saddle-like surface of the double-wing domain.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
66 Section VII: Molecular Biology
The bacteriophage T4 MotB protein, a DNA binding protein, boosts the level
of T4 late gene expression
Jennifer Patterson West1, L. Iyer2, and Deborah M. Hinton1
1Gene Expression and Regulation Section, Laboratory of Cell and Molecular Biology, NIDDK,
National Institutes of Health, Bethesda, Maryland 20892, USA, 2Computational Biology Branch,
National Library of Medicine, National Institutes of Health, Bethesda, Maryland 20892, USA
E-mail: [email protected]; [email protected]
Condensation and organization of genomic DNA is crucial in all cells for orderly replication and
gene expression. In bacteria, histone-like proteins such as the abundant H-NS and its less abundant
homolog StpA, are DNA binding proteins that form higher-order nucleoprotein complexes needed
for DNA condensation. H-NS targets AT-rich DNA sequences and condenses genomic DNA
through a proposed looping mechanism that typically represses transcription at the affected region.
As phage genomes and xenogeneic sequences acquired from horizontal gene transfer often display
a high AT content, H-NS can protect bacteria from the expression of foreign genes by
preferentially binding these sequences.
Bacteriophage T4 (65.5% AT) is a lytic virus that infects E. coli, (45% AT) resulting in cell lysis
after ~20 min. The T4 genome is expressed temporally from early, middle and late promoters. The
T4 late promoter, TATAAATA, is strikingly similar to the H-NS binding motif, TCGATAAATT.
Thus, it is not surprising that T4 may need to overcome H-NS repression. T4 Arn, a protein that
structurally mimics DNA has been shown to bind H-NS, preventing its interaction with DNA and
formation of higher order structures. However, Arn is not essential.
The T4 motB gene encodes a highly conserved T4 early protein whose function has not been
characterized previously. We find that expression of plasmid-borne motB is highly toxic to E. coli,
resulting in decondensation of host DNA, cell lengthening, significant reduction in actively
dividing cells compared to a vector control, and cell lysis. MotB co-purifies with DNA, H-NS, and
StpA. Electrophoresis mobility shift assays indicate that H-NS and MotB bind similar AT-rich
sequences. However, MotB binds with 10-100-fold higher affinity than H-NS depending on the
sequence. Although a T4 motB amber mutant has no noticeable phenotype in a plaque assay, the
mutant has a 2-fold reduction in burst size and RNA-seq analyses indicate that the expression of
several T4 late genes are significantly reduced. Expression of early and middle transcripts are not
reduced. These findings are consistent with MotB relieving H-NS binding to and repression of late
promoters and/or a direct activation of late gene expression by MotB. We hypothesize that the
interaction of MotB with either H-NS (and StpA), DNA, or both is part of a mechanism used by T4
to disrupt H-NS dependent repression leading to optimal expression of its late genes. Determining
why and how MotB is toxic can provide paths to the formation of anti-bacterials that work by a
mechanism not previously targeted by antibiotics.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VII: Molecular Biology 67
The regulation and mechanism of the bacteriophage T4 MR complex
(gp46/47)
Scott Nelson
Iowa State University, Ames, IA
E-mail: [email protected]
Bacteriophage T4 gp46 (Rad50 homolog) and gp47 (Mre11 homolog) are central players in DNA
repair and are implicated in the processing of DNA ends caused by DNA double strand breaks
(DSBs). Mre11 and Rad50 form a stable complex (MR) and work cooperatively in repairing
DSBs. Due to difficulties in the expression and purification of the full-length Rad50 protein, a
detailed mechanistic understanding of the MR complex has been lacking. We have established a
tractable system for the biochemical characterization of T4 Mre11 and Rad50. Rad50 enhances the
nuclease activity of Mre11, while DNA and Mre11 in combination stimulate the ATPase activity of
Rad50. The structural basis for the cross activation of MR complex has been elusive. Recent
crystal structures of truncated forms MR complex suggest that several conformational
rearrangements may occur during its ATP hydrolysis and nuclease cycle. These conformational
changes likely transmit signals between active sites and appear to affect the dimer interface of
Mre11 and the coiled-coil domain of Rad50 to the greatest extent. We have used a combination of
biochemical and biophysical experiments to explore the connections between these large-scale
movements and the various activities of the MR complex. These results have been incorporated
into a comprehensive model that describes the entire catalytic cycle of the MR complex during
processive nuclease activity.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
68 Section VII: Molecular Biology
Hot Tales of T4’s Transition from Host to Phage Metabolism
Elizabeth Kutter
The Evergreen State College, Olympia, WA
E-mail: [email protected]
Much of what we know about phage-host interactions was originally discovered by introducing
radioactively labeled molecules into T4-infected E. coli. While biologists today have access to
tools like RNAseq, the old standby of radiolabeling is still uniquely useful in exploring some
details of the process.
Shutoff of host protein synthesis: 14C-labeled mixed amino acids were used to show that
synthesis of all host proteins ceases within 1-3 min after infection, while 2-D gels showed the
detailed pattern of synthesis of each protein. The very extensive and well-studied collection of T4
amber and deletion mutants facilitated extensive identification of many of the genes.
Effects on host lipid synthesis: Measurements involving 14C – acetate incorporation showed that
T4 quite surprisingly stimulates phosphatidyl glycerol (PG) synthesis, while phosphatidyl ethanol
(PE) continues to be made at the pre-infection rate. Both the stimulation of PG and the ongoing
synthesis of PE and PG are under specific control of genes lying in the deletable region between
rIIB and gp52, which encodes one component of the topoisomerase; thus, it is probably not simply
a passive coincidence for the phage.
Host DNA breakdown and non-canonical nucleotide use explored using 3HdT: For many
years, our year-long, full-time “Molecule to Organism” program as well as the phage lab used
radiolabeled dT to explore consequences of the phage infection process through looking at phage
DNA synthesis and host DNA breakdown, in a very safe and highly reproducible manner teaching
many aspects of phage biology and learning about the principles and practice of working with
radionucleotides.
A “hibernation” mode of T4 infection of stationary-phase E. coli: When T4 infects cells that
have been growing at least 24-48 hours, it generally kills the host it is attacking quite efficiently,
but produces no progeny phage until after nutrients are added. We have determined that the host
DNA is largely broken down to mononucleotides over just a few hours and the nucleotides are
effectively re-incorporated into phage DNA, indicating that the infection proceeds at least through
production of the phage’s “new” phage nucleotide complex, but no new phage are made until
nutrient addition at 24 or 48 hours – then rapidly producing and releasing nearly 1011 phage per ml.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VII: Molecular Biology 69
Superinfection exclusion systems as molecular drivers of lactococcal 936
phage evolution Jennifer Mahony1, 3, Angela Makumi1, Irma van Rijswick1, Christian Cambillau2 and Douwe van
Sinderen1, 3.
1School of Microbiology, University College Cork, Western Road, Cork; 2 Architecture et Fonction
des Macromolécules Biologiques, UMR 7257 CNRS & Aix-Marseille University, Campus de
Luminy, Case 932, 13288 Marseille Cedex 09, France; 3Alimentary Pharmabiotic Centre,
Biosciences Institute, University College Cork, Western Road, Cork, Ireland.
Bacteriophages have an overpowering abundance in the biosphere having a profound impact on the
conduct and ecology of their hosts. The two best described and understood routes in phage
propagation includes the (i) lytic cycle, which can affect the structure of microbial corporation as
well as the host mutation rates and (ii) the lysogenic cycle in which the phage genome is integrated
into the host chromosome (prophage). Such prophage integration leads to an expanded genome
providing a further potential increase in host adaptive abilities and fitness. Furthermore, bacterial
hosts may benefit from the protection conferred by prophage-encoded immunity and/or phage-
resistance against homologous and heterologous phages. One such group of phage-resistance
systems, termed superinfection exclusion (Sie) systems, provide protection against certain
members of heterologous phages. In this study, we have focused on the characterization of Sie
systems derived from dairy lactococcal strains with potential implications for the dairy
fermentation industry. Among the ten known groups of lactococcal phages, members of the so-
called 936 phage species are the most frequently encountered and are among the most problematic
to such fermentations.
Here, we present data that demonstrates that these Sie systems are powerful evolutionary drivers of
members of the 936 phage species. Mutants of representative members of the 936 phage species
capable of bypassing the Sie system exhibited an extensive set of mutations in the gene that
specifies the tail tape measure protein (TMP), highlighting not only the role of TMP in evading the
action of these Sie systems, but also its possible involvement in the DNA injection process of 936
phages. To scrutinize this further, we also present data at single cell level bringing such elusive
insights within reach, shedding light on the regulation and modus operandus involved in the Sie
system.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
70 Section VII: Molecular Biology
Host recognition by podoviruses G7C and Alt
Nikolai S. Prokhorov1,2,3, Sergey Nazarov2, Cristian Riccio2, Ricardo Guerrero-Ferreira2, Alla K.
Golomidova3, Evelina L. Zdorovenko4, Yuriy A. Knirel4,
Andrey V. Letarov3 & Petr G. Leiman1,2
1Department of Biochemistry & Molecular Biology,
University of Texas Medical Branch, Galveston, Texas 2École Polytechnique Fédérale de Lausanne (EPFL), Lausanne, Switzerland
3Winogradsky Institute of Microbiology,
Research Center of Biotechnology, RAS, Moscow, Russia 4Zelinsky Institute of Organic Chemistry, RAS, Moscow, Russia
The O22 E. coli strain 4s populates the lower intestine of a horse. It is a host to a number of
bacteriophages that compete and, possibly, help each other to survive in this complex and dense
microbiological environment. N4-like phages G7C and Alt are close relatives with essentially
identical genomes, except for a single gene the product of which is responsible for host recognition
and forms a tailspike on the phage particle. We used a combination of X-ray crystallography and
cryo-electron microscopy to determine the structure of G7C tail in atomic details. It carries twelve
copies of two different tailspikes that form a branched structure. The longer tailspike - gp66 (1063
aa) - contains a phage binding domain, a flexible linker, a branched domain responsible for the
attachment of the other spike, and a C-terminal ß-helical receptor-binding domain. This protein is
identical in both phages. The shorter spikes of G7C and Alt - gp63.1 (851 aa) and gpA63.1 (854 aa)
- have nearly identical N-terminal domains that interact with gp66 and strikingly different C-
terminal receptor-binding domains. The short spike of Alt is a lyase. It degrades the E. coli 4s O-
antigen into small fragments. The short spike of G7C acts differently. It keeps the backbone of the
E. coli 4s O-antigen polymer intact but removes one O-acetyl group per its repeating unit. Thus,
G7C and Alt utilize non-homologous domains and different types of enzymatic activities to
recognize and bind to the same cellular receptor molecule. Functional analysis of host recognition
by G7C, Alt, and their tailspike mutants revealed that O-antigen binding and modification are
critically important for productive infection. We found that the kinetics of O-antigen processing by
the tailspikes appears to control the transition from primary host recognition to irreversible
adsorption and subsequent DNA release. Our findings demonstrate that phages with identical
particle structure and tail architecture can utilize strikingly different biochemical processes to
efficiently recognize, bind, and infect their host cells.
Prokhorov, N.S., C. Riccio, E.L. Zdorovenko, M.M. Shneider, C. Browning, Y.A. Knirel, P.G.
Leiman & A.V. Letarov, (2017) Function of bacteriophage G7C esterase tailspike in host cell
adsorption. Mol Microbiol. doi: 10.1111/mmi.13710.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VII: Molecular Biology 71
Phages that farm: insights into the phage-carrier state dynamics
Ines Staes1, William Cenens1, Nick de la Croix1, Angela Makumi1, Sanne Wolput1, Rob Lavigne2,
Abram Aertsen1
1Laboratory of Food Microbiology, Department of Microbial and Molecular Systems (M2S),
Faculty of Bioscience Engineering, KU Leuven, Belgium, 2Laboratory of Gene Technology,
Department of Biosystems, Faculty of Bioscience Engineering, KU Leuven, Belgium
E-mail: [email protected]
Bacteria are condemned to an eternal battle with their viral predators, bacteriophages, as the latter
can only multiply and spread their progeny at the cost of their bacterial host. Phages with a strict
lytic lifestyle inexorably kill the infected cell, consequently leading to a drastic reduction of
available hosts. A safer approach is opted by temperate phages that insert their DNA into the
bacterial chromosome and replicate along with it, although this protection comes at the expense of
an impairment in spread of progeny. A possible solution by which a phage can circumvent this
dilemma, is the formation of a phage carrier state in which a delay of the integration event allows
the growth of a phage free subpopulation that is transient resistant to superinfection. This strategy
might contribute to a stable phage-host co-existence in nature as it allows these farming phages to
foster a reservoir of bacterial cells that afterwards can be “harvested”. In the temperate phage P22 –
Salmonella Typhimurium model system, we previously observed that phage carrier cells
overproduce the P22 pid ORFan gene, which leads to derepression of the host’s dgo-operon. In the
search for phage-encoded factors involved in pid-regulation, expression or function, a screening
was performed using a library of DES-mutagenized P22-phages. This yielded interesting
mutations in the secondary immunity region of P22 that provide further insight into its infection
dynamics.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
72 Section VII: Molecular Biology
Virus Interactions inside the Cell: Competition or Cooperation?
Lanying Zeng
Department of Biochemistry and Biophysics, Center for Phage Technology, Texas A&M
University, 300 Olsen Blvd, College Station, TX 77843
E-mail: [email protected]
Living cells make fate-determining decisions based on signals from their environment.
Understanding the decision making process is essential for unveiling the mysteries of life and for
improving human health. I will discuss our recent study of a paradigmatic system of cell-fate
determination, the bacterium E. coli and its virus – phage lambda, using a live-cell 4-color system
combined with computational modeling at the single-virus/single-virus-DNA resolution. The
textbook picture is: upon infection by phage lambda, E. coli undergoes one of two alternate
pathways - lytic (violent) or lysogenic (dormant). With the high-resolution studies, this
paradigmatic system has been revealed to be more complicated than previously thought. Our study
suggests that individual phages vote and interact within the cell: they cooperate during
lysognization, compete among each other during lysis, and confusion or coexistence between the
two pathways occasionally occurs. I will also discuss the sources/mechanisms for phages to make
different decisions and utilize variable strategies for their development.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VII: Molecular Biology 73
Asymmetric cryo-EM structure of the canonical Allolevivirus Qβ reveals a
single maturation protein and the genomic ssRNA in situ
Junjie Zhang
Center for Phage Technology, Department of Biochemistry and Biophysics, Texas A&M
University, College Station, TX 77843
Single-stranded (ss) RNA viruses infect all domains of life. To date, for most ssRNA virions, only
the structures of the capsids and their associated protein components have been resolved to high
resolution. Qβ, an ssRNA phage specific for the conjugative F-pilus, has a T = 3 icosahedral lattice
of coat proteins assembled around its 4,217 nucleotides of genomic RNA (gRNA). In the mature
virion, the maturation protein, A2, binds to the gRNA and is required for adsorption to the F-pilus.
Here, we report the cryo-electron microscopy (cryo-EM) structures of Qβ with and without
symmetry applied. The icosahedral structure, at 3.7-Å resolution, resolves loops not previously
seen in the published X-ray structure, whereas the asymmetric structure, at 7-Å resolution, reveals
A2 and the gRNA. A2 contains a bundle of α-helices and replaces one dimer of coat proteins at a
twofold axis. The helix bundle binds gRNA, causing denser packing of RNA in its proximity,
which asymmetrically expands the surrounding coat protein shell to potentially facilitate RNA
release during infection. We observe a fixed pattern of gRNA organization among all viral
particles, with the major and minor grooves of RNA helices clearly visible. A single layer of RNA
directly contacts every copy of the coat protein, with one-third of the interactions occurring at
operator-like RNA hairpins. These RNA–coat interactions stabilize the tertiary structure of gRNA
within the virion, which could further provide a roadmap for capsid assembly.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
74 Section VIII: Open Section
What factors determine phage penetration in animals and humans? Systematic
and critical review of reports published in the years 1940-2016
Krystyna Dąbrowska
Ludwik Hirszfeld Institute of Immunology and Experimental Therapy, Polish Academy of
Sciences, R. Weigla 12, 53-114 Wrocław, Poland
E-mail: [email protected]
The efficacy of medical applications of bacteriophages, including antibacterial therapy,
vaccines and other types of nanocarriers rely on phage ability to reach target organs or tissues. This
ability of phages to “travel” inside human or animal body is variable. Reports from multiple human
and animal studies present differentiated or even contradictory conclusions on how a phage may
penetrate in vivo. Many factors that may potentially determine phage penetration, e.g. the route of
administration, phage type including phage morphology and host range, dose and application
schedule, characteristics of treated individuals (species, age), presence or lack of sensitive bacteria,
and many others. Mostly due to the multiplicity of these factors, their true impact on phage
penetration is still unclear.
Here I present a systematic review of phage-related literature available in PubMed (up to
Dec 31st 2016) focusing on in vivo experiments that involved phage penetration into blood or
tissues, and gut transit of phage. Systematic review was further developed by a critical review of
related mechanisms. Criteria for the systematic search were: ((phage) OR bacteriophage) AND
((per os) OR (gut) OR (intestine) OR (fecal recovery) OR (route of administration) OR (oral) OR
(inhalation) OR (intranasal) OR (intraperitoneal) OR (intramuscular) OR (subcutaneous) OR
(intracranial) OR (intravenous) OR (parenteral) OR (experimental infection) OR (skin penetration)
OR (wound) OR (transdermal) OR (penetration) OR (cutaneous) OR (circulation)), followed by
pre-defined inclusion and exclusion criteria.
The search resulted in 3279 titles that were analyzed in two-steps; eventually 216
publications were selected for full-text analysis; 119 individual experiments were extracted for
phage penetration analysis and 84 individual experiments for phage gut transit analysis. They were
quantified for the strength of the effect and investigated factors were categorized as follow: route of
administration, phage morphology, phage host, dose, species of treated individuals, their age,
presence/lack of sensitive bacteria. The most explicit correlations of phage penetration as revealed
by the statistical analysis of sample distribution in groups were:
1. route of administration, with the lowest efficacy of oral administration (96.3%) and with
equal efficacy of others (i.p., i.v., s.c., i.m., intranasal)
2. phage type, with the better penetration of podoviruses and microviruses (weaker
penetration: myoviruses, siphoviruses and inoviruses)
3. presence of a sensitive bacterial host in the system.
No important impact was noted for the factors like bacterial host type, animal species or age. Once
entering the system, phage was able to arrive to the most of organs and tissues, including spleen,
liver, kidneys and urinary bladder, thymus, hearth, lungs, muscle, and even brain and bone marrow.
Gut transit of phage depended mostly on the presence of a sensitive bacterial host in situ.
Surprisingly, no marked impact of stomach acidity neutralization was revealed. Applied phage
dose correlated to phage recovery level.
This work was supported by the National Science Centre in Poland, grant UMO-
2012/05/E/NZ6/03314 and by Wroclaw Centre of Biotechnology, programme The Leading
National Research Centre (KNOW) for years 2014-2018

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VIII: Open Section 75
Anti-Pseudomonal activity of phage PEV20 in a dry powder formulation
in a murine lung infection model
Yoon-Kyung (Rachel) Chang1, Ke Chen2, Jiping Wang2,3, Jian Li3, Sandra Morales4, Elizabeth
Kutter5 and Hak-Kim Chan1*
1Faculty of Pharmacy, University of Sydney, Sydney, Australia, 2Monash Institute of
Pharmaceutical Sciences, Monash University, Melbourne, Australia, 3Department of Microbiology,
Monash University, Melbourne, Australia, 4 AmpliPhi Biosciences AU, Sydney, Australia, 5The
Evergreen State College, Olympia, USA
*E-mail: [email protected]
Objectives: To evaluate the efficacy of a dry powder formulation of Pseudomonas aeruginosa
phage PEV20 in a murine lung model of infection
Methods: Highly purified phage PEV20 with high titre was obtained from AmpliPhi Biosciences
AU. Phage PEV20 was co-spray dried with 17 mg/mL of lactose and 8 mg/mL of leucine in sterile
ultra-pure water (pH 7.4) using Büchi 290 spray dryer coupled with a conventional two-fluid
nozzle for atomization. Stability of phages in spray dried formulation was tested using the standard
double layer plaque assay. Immunocompromised Swiss mice (4 mice per group) were challenged
with 2 x 105 colony forming units (CFU) P. aeruginosa PSA ADL 01 strain using intratracheal
instillation. After 2h post-bacterial administration, 1mg of phage powder was intratracheally
delivered using dry powder insufflator; no treatment was given to the control group. At 4h and 24h
post-phage treatment, the lung and plasma were collected for bacterial and phage counting.
Results: Spray dried phage PEV20 powder was produced at 2 x 107 plaque forming units
(PFU)/mg. Phage-treated group showed 5-log10 reduction in bacterial load (105 CFU) in the lung
after 24h compared to untreated (1010 CFU). All the phages administered were recovered in the
lung (107 – 108 PFU) at 4 h with a 1-log10 titre increase after 24h (108 – 109 PFU) compared with
the actual phage dose given. The concentration of phage PEV20 in plasma was higher at 24h than
at 4h.
Conclusions: Stable PEV20 spray dried formulation was generated at a therapeutic dose for
murine lung infection model. A significant killing effect was observed against a clinical P.
aerugonisa strain in vivo. Both phage and bacterial load stayed consistent after 2h post phage-
administration. After 24h, significant reduction in the pathogen number conveyed phage replication
and bacteriolysis in the lung, which was verified by the increase in phage titre. This is a proof-of-
concept study demonstrating the potential use of phage PEV20 dry powder formulation for
treatment of P. aeruginosa lung infection. Powder formulations provide easy storage, transport and
administration over liquid formulations.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
76 Section VIII: Open Section
CRISPR-Cas and phage-host interactions in Streptococcus mutans
Cas Mosterd and Sylvain Moineau*
Département de biochimie, microbiologie et bio-informatique, Faculté des sciences et de génie,
Groupe de recherche en écologie buccale, Faculté de médecine dentaire,
Université Laval, Québec, Canada, G1V 0A6
*E-mail: [email protected]
Streptococcus mutans is a member of the natural oral microbiota, where it is the main cause of
dental caries. Phages are also abundant in the oral cavity. To evade viral predation in this
environment S. mutans likely encodes several anti-viral mechanisms. One of these mechanisms
may be the CRISPR-Cas system. CRISPR is an acronym for clustered regularly interspaced short
palindromic repeats and this locus is often located close to CRISPR-associated (cas) genes. Each
CRISPR locus consists of short repeat nucleotide sequences interspaced by variable nucleotide
sequences (spacer). Upon the discovery that spacer sequences often matched plasmid and phage
sequences, it was experimentally confirmed to work as a defence mechanism, specifically as an
adaptive microbial immune system. The spacer sequences provide specificity to the system by
targeting foreign DNA sequences (proto-spacers), while the cas genes were found to encode a
variety of proteins that each have a role within the system. In fact, CRISPR-Cas systems show
remarkable diversity of Cas proteins, effector complex structure, locus architecture and
mechanisms of adaptation, crRNA processing and DNA/RNA interference. CRISPR-Cas systems
belong to two classes, with multi-subunit effector complexes in Class 1 and a single-protein
effector in Class 2. Within each class, a number of types and subtypes can also be found.
In this study it is demonstrated that S. mutans strain SMQ-1336 possesses a Type II-A CRISPR-
Cas system. When the wild-type strain SMQ-1336 is challenged with a virulent phage,
bacteriophage-insensitive mutants (BIMs) can be readily obtained. Analysis of the CRISPR array
of these BIMs revealed the acquisition of new spacers. Acquisition of multiple spacers was
observed in several BIMs. Sequence analysis of the new spacers indicated that they were derived
from the infecting phage genome. Surprisingly, all the BIMs were shown to also prevent phage
adsorption, indicting a probable mutation in the unknown phage receptors. Experiments in which a
repeat-spacer-repeat unit targeting the genome of the same virulent phage was introduced into S.
mutans SMQ-1336 on a plasmid did not result in phage resistance. However, plasmid constructs
harbouring a sequence matching one of the spacers present in the CRISPR array of S. mutans
SMQ-1336 could not be transformed in the wild-type strain, confirming the interference activity of
the CRISPR-Cas system.
Taken altogether, the CRISPR-Cas system of S. mutans SMQ-1336 does not seem to protect
against phage infection but prevent plasmid transformation. The exact role of the CRISPR-Cas
system in S. mutans and the other mechanism(s) involved in phage resistance are under
investigation.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VIII: Open Section 77
Mucosal Vaccine Delivery for Peptide Antigens via Lambda Display Phage
Patricia González Cano1, Lakshman N., A. Gammage1,2, Connie Hayes3, Karthic Rajamanickam3
Scott Napper1,4, Sidney Hayes3* and Philip J. Griebel1,5
1VIDO-InterVac, University of Saskatchewan, Saskatoon, SK, Canada S7N 5E3; 2 Current
Address: CFIA, 534-East Cordova St, Vancouver, BC, Canada. V6A 1L7; 3Department of
Microbiology and Immunology, College of Medicine, University of Saskatchewan, Saskatoon, SK,
Canada S7N 5E5; 4Department of Biochemistry, University of Saskatchewan, Saskatoon,
Saskatchewan, Canada S7N 5E5; 5School of Public Health, University of Saskatchewan,
Saskatoon, SK, Canada S7N 2Z4
*E-mail: [email protected]
There is increasing interest in the use of bacteriophage as protein/peptide and DNA vaccine
delivery vehicles. The safety, stability, ease of production, and immunogenicity of bacteriophages
have been characterized but little is known about their capacity to induce mucosal immune
responses in the small intestine. Whole body imaging confirmed that within 24 hours orally
delivered lambda bacteriophage (LP) was distributed through the gastro-intestinal tract of mice.
Targeted delivery of LP to specific sites in the small intestine of newborn calves confirmed LP
were immunogenic in a dose-dependent manner. LP were taken up by Peyer’s patches (PPs) and
induced LP-specific IgA and T cell responses within the mucosa-associated lymphoid tissue. A
lambda display phage (LDP) was constructed to present three immunogenic peptide epitopes
(YYR, YML, RL) fused to lambda coat protein D to display the disease specific epitopes (LDP-
DSE). Delivery of purified LDP-DSE to surgically constructed intestinal segments induced IgA
antibody-secreting cell (ASC) responses to the three displayed peptide epitopes within the targeted
PPs. Further, delivery of bacteria expressing D-DSE or bacteria containing assembled LDP-DSE
also induced epitope-specific IgA-ASC responses PP. These are the first studies to report the use of
LDP to induce mucosal immune responses in the small intestine and identify Peyer’s patches as an
important site for uptake of LP. Furthermore, these responses were observed in the absence of
exogenous immune stimulating compounds or adjuvant. Therefore, we conclude that LDP have
substantial potential to be used as an oral vaccine delivery vehicle for immunogenic protein/peptide
vaccines.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
78 Section VIII: Open Section
Immunophage synergy is essential for eradicating pathogens that provoke
acute respiratory infections.
Dwayne Roach1, Chung Leung2,3, Marine Henry1, Eric Morello1, Devika Singh2, James Di
Santo4,5, Joshua Weitz2,3, and Laurent Debarbieux1*
1Department of Microbiology, Institut Pasteur, Paris 75015, FR 2School of Biology, Georgia Institute of Technology, Atlanta, Georgia 30332, USA 3School of Physics, Georgia Institute of Technology, Atlanta, Georgia 30332, USA
4Innate Immunity Unit, Department of Immunology, Institut Pasteur, Paris 75015, FR 5Inserm U1223, Paris 75015, FR
*E-mail: Laurent. [email protected]
Pseudomonas aeruginosa is an important cause of life-threatening nosocomial pneumonia and
prone to multidrug resistance. Especially at risk are patients with weakened immune systems and
chronic respiratory disorders. Considering the scarcity of new antibiotics, the use of bacteriophage
(phages) as an alternative therapy has re-emerged. While several recent animal studies have
demonstrated the therapeutic potential of phages for treating pseudomonal lung infection in healthy
immunocompetent animals, it remains unknown whether phages are effective in the setting of
immunodeficiency. Here, we provide a systematic in vivo and mathematical investigation of host
innate immune status on the efficacy of monophage curative and prophylactic therapies for
pneumonia caused by P. aeruginosa in mouse. We show that the ability of inhaled phage therapy to
cure acute pneumonia and provide prophylaxis was host neutrophil dependent. In contrast, phage
therapy efficacy was innate and adaptive lymphocyte-independent in this setting. Mathematical
modelling the dynamics between host innate immunity, bacterial infection and phage replication,
we show that both phage lysis and neutrophil activity work in concert to reduce both phage-
sensitive and phage-resistant bacteria in the lungs. This ‘immunophage synergy’ contrasts with the
predominant view that phage therapy efficacy relies largely on the bacterial permissiveness to phage
lysis. Mathematical simulations also imply heterogeneous mixing of phage and bacterial
populations and phage saturation have important consequences on treatment efficacy. Nevertheless,
we show that phage therapy can still be efficacious in hosts with weakened innate immunity that
manifest emergence of phage resistance. In addition, there were no untoward immune effects from
high dose of phages alone in lung tissues. This study supports that phage therapy can be a safe and
effective antibacterial strategy but success is defined by immune status of the patient.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VIII: Open Section 79
Use of bacteriophages for construction and tests of biosensors.
Marcin Łoś
Phage Consultants, Partyzantow 10/18, 80-254 Gdansk, Poland
Department of Bacterial Molecular Genetics, University of Gdansk, Wita Stwosza 59, 80-309
Gdansk, Poland
E-mail: [email protected]
Pathogenic viruses and bacteria are very often a natural target for development of detection and
rapid diagnostics methods. The need for rapid detection and identification of pathogens spans
widely from the healthcare systems through veterinary to military use in e.g. battlefield diagnostics.
Increase in the threats of use of biological warfare against civilians were for a long time a driving
force in development of automated rapid detection devices, which could minimize a risk for
population. The aim is to build a test, which is easy, cheap, sensitive, fast and reliable. Usually it is
impossible to meet all these requirements at the same time. However, the constant increase in our
abilities to miniaturize and to build more and more complex electronics allow us to build more and
more sophisticated sensing elements able to detect even very small changes in the environment.
Despite this, in the construction of the biosensors also biological part plays very important role. My
favorite group of viruses - bacteriophages, appeared to be an ideal to play a role as a detection
target, but also as a biological part of a sensing elements. They posses many desirable
characteristics common with pathogenic viruses, but they are also very efficient in interactions with
bacterial cells. Because of that their use may in many cases appear to be superior to the use of more
traditional ways of detection employing e.g. antibodies in interactions with bacterial pathogens.
During this presentation I will summarize over 15 years of a struggle to use phages in both of those
roles - as a model and as a part of a sensing element and our approach to integrate them with non-
biological part of biosensors.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
80 Section VIII: Open Section
Getting the enemy to help you out: Elucidating the specificity determinants of
anti-phage genomic island excision in Vibrio cholera
Amelia McKitterick, Kimberley Seed*
University of California, Berkeley, Berkeley, California.
E-mail: [email protected]
Vibrio cholerae is the causative agent of cholera and thrives in both aquatic environments and the
human intestinal tract. In both of these environments V. cholerae is susceptible to predation by
bacteriophages, the most prevalent of which is ICP1. In order to combat ICP1 predation, V.
cholerae has evolved a unique defense system referred to as the PLE (Phage-inducible
chromosomal island Like Element), a genomic island that is found integrated into the bacterial
chromosome. Five PLE variants have been identified in clinical isolates of V. cholerae and are
characterized by the ability to specifically block ICP1 production, although the mechanism(s)
underpinning this activity are not understood. PLEs are activated and respond specifically to ICP1
infection by excising from the chromosome and circularizing. PLE 1 encodes Int, a putative large
serine recombinase that drives recombination between attachment sites. Integration is catalyzed
solely by Int, however, excision requires a recombination directionality factor (RDF) that directs
Int-mediated recombination between the integrated attachment sites allowing for excision. As PLE
excision and circularization are observed only after ICP1 infection, we hypothesize that a phage
encoded RDF is necessary for directing PLE excision. We show that Int is necessary for PLE 1
excision, and that Int is the only PLE-encoded factor required for the recombination event in
response to ICP1. Additionally, we identified an ICP1-encoded gene product (Ind1), that is
necessary and sufficient for Int-mediated PLE excision, indicating that it is the RDF. As such, the
integrase and Ind1 required for PLE excision are encoded by separate entities, which helps to
define the molecular specificity of PLE activity. Variability in int is seen in between PLEs,
indicating that PLEs have evolved in response to changing ICP1 gene products, furthering our
hypothesis that PLEs are engaged in a molecular arms race with ICP1.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section VIII: Open Section 81
Is more better? Use of multiple hosts in phage isolation
Paul Hyman
Ashland University, Ashland, Ohio, USA
E-mail: [email protected]
A bacteriophage’s host range is an important characteristic with implications for phage ecology as
much as applications of phages including phage therapy. In our work isolating bacteriophages, and
in the literature, there is indication that isolation methods may bias the host range of novel isolated
bacteriophages. Specifically, it appears that isolating bacteriophages with a mixture of host strains
favors broader host range phages. We have previously applied this in isolating Enterococcus
faecalis phages. Independently isolating two phages using two strains of E. faecalis together did
produce phages with broader host range than two phages isolated on only one of the strains. Oddly
though, the broader host range phages could only infect one of the two isolation strains.
Very recently we have begun isolation of phages from soil with a single host and several
combinations of multiple hosts for the enrichment culture. Our preliminary results are that more
phages (or higher titers of phages) are obtained in the single host cultures than any of the multiple
host cultures. Spot testing shows that these phages also only infect one of the isolating hosts.
These results suggest there may be negative as well as positive effects of the use of multiple hosts
in phage isolation. They also suggest that not all strains of bacteria will be equally useful in phage
isolation.
.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
82 General
POSTER ABSTRACTS
Abstracts are arranged in thematic sections. Within each thematic section, posters are
ordered alphabetically by the last name of the presenting author.
Section I: Agriculture and Food Safety
Section II: Phage-Based Biotechnology
Section III: Ecology & Evolution
Section IV: Genomics & Proteomics
Section V: Structure and Molecular Mechanisms
Section VI: Temperate Phages
Section VII: Phage Therapy
The presenting author’s name appears in bold text. The names of other authors who are
attending the meeting have been underlined.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section I: Agriculture and Food Safety 83
Enhanced inhibition of Salmonella Typhimurium using a phage cocktail
targeting three different host receptors
Jaewoo Bai, and Sangryeol Ryu*
Department of Food and Animal Biotechnology, Department of Agricultural Biotechnology,
Research Institute for Agriculture and Life Sciences, and Center for Food and Bioconvergence,
Seoul National University, Seoul, Korea.
*E-mail: [email protected]
Objectives: With the increasing food safety concerns about fresh produce and limitations of
current disinfectant treatments, the development of novel and safe methods is urgently required to
control the contamination of fresh produce. Bacteriophages are considered as a natural antibacterial
agent for the control of foodborne pathogens. However, the rapid development of bacterial
resistance to phage infection is the most significant barrier to practical phage application. To
overcome this problem, we developed a novel phage cocktail consisting of three virulent phages
(BSPM4, BSP101 and BSP22A) that target different host receptors, including flagella, O-antigen
and BtuB, respectively. This study provides a receptor-based strategy to develop a novel biocontrol
agent against S. Typhimurium in fresh produce.
Methods: Bacteriophages infecting S. Typhimurium LT2C were isolated from various
environmental samples. To confirm the phage receptor, complement strains for rfbP, btuB, and
flgK genes were further subjected to spotting assay. For genomic analysis, Genome Sequencer FLX
(GS-FLX) instrument was used and then assembled with Newbler v2.9. Open reading frames
(ORFs) were predicted using the Glimmer v3.02, GeneMarkS and FgenesB software programs.
The annotation and functional analysis of the predicted ORFs was conducted using the BLASTP
and InterProScan databases. The bacterial growth inhibition abilities of single phage and their
cocktails were tested using S. Typhimurium LT2C strain with an MOI of 1. Fresh iceberg lettuce
and cucumber were used for food application for the phage cocktail. Each sample was cut and
sliced into the size approximately 10 cm2 and then artificially contaminated with Salmonella (~ 1 x
105 CFU). Each sample was treated with phage cocktail at different MOI values (103 to 104) and
incubated. At the indicated time point, each sample was homogenized and spread-plated for the
enumeration of S. Typhimurium.
Results and conclusions: This study was carried out to construct a novel phage cocktail using
agent against Salmonella. For this purpose, three novel phages targeting the different receptors
(i.e., flagella, O-antigen and BtuB) of S. Typhimurium were isolated and characterized. In addition,
genomic study of three phages revealed that all three phages are strictly virulent without no toxin
or virulence-related genes in their genomes, suggesting they are safe to be applied on foods. In-
vitro challenge assay and BIMs frequency analysis revealed the enhanced effect of three phages
cocktail to inhibit the host growth and to reduce the development of resistant strains, indicating the
preparation of cocktail with phages utilizing different host targets is a powerful strategy to control
S. Typhimurium. Furthermore, the effective inhibition of S. Typhimurium growth by the phage
cocktail in food systems showed that the phage cocktail has great potential of being developed as a
biocontrol agent against S. Typhimurium in fresh produces.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
84 Section I: Agriculture and Food Safety
Isolation and characterization of broad host range virulent phages for control
of Salmonella on plants
Sudhakar Bhandare, Brigitte Cadieux, Anna Colavecchio and Lawrence Goodridge*
Department of Food Science and Agricultural Chemistry, McGill University, 21111 Lakeshore
Road, Ste Anne de Bellevue, QC, H9X 3V9.
*E-mail: [email protected]
Objectives: Salmonella is an important cause of foodborne illness in Canada and worldwide. Each
year, approximately 88,000 people become sick in Canada from consuming food that is
contaminated with Salmonella, while the global burden of Salmonellosis is also substantial.
Recently, fresh fruits and vegetables have emerged as important sources of Salmonella apart from
traditional animal origin sources. This is because fruits and vegetables are grown in soil, where
contamination due to animals and non-potable water can occur. Control of Salmonella on fresh
produce is difficult and biocontrol of Salmonella using bacteriophages is a natural and efficient
way to reduce the load of Salmonella on fruits and vegetables.
The objective of this work was the isolation and initial characterization of bacteriophages capable
of infecting a wide range of Salmonella enterica serotypes, and which could be employed in
biological control of Salmonella that contaminate plants. Isolated bacteriophages were
characterized based on host range, their morphology and their genome sizes.
Methods: Bacteriophage isolation was carried out using sewage samples (1L) collected from the
Jean-R. Marcotte Wastewater treatment plant in Montreal by adopting standard enrichment and
agar overlay techniques (Van Twest and Kropinski 2009). Thirteen target Salmonella isolates,
originally isolated for food plants, and representing thirteen serotypes were used for isolation.
Isolated phages were characterized biologically by host range profile using 43 different Salmonella
isolates from 30 serotypes and broad host range phages were also screened for their lytic spectra on
40 other Enterobacteriaceae strains. Broad host range phages were further characterised
morphologically by electron microscopy and their genomes sizes were estimated using Pulsed
Field Gel Electrophoresis (PFGE).
Results: Thirteen phages were successfully isolated for each of the 13 target strains. Phages Φ3
and Φ6 could lyse 88.3%; while phage Φ9 lysed 86% and phage Φ13 lysed 83.7% of the 43 host
strains used for lytic spectra. These four broad host range phages could lyse some of the 40 other
Enterobacteriaceae strains. Φ3 and Φ6 could lyse 4 strains each while Φ9 could lyse 8 and the Φ13
could lyse 14 Enterobacteriaceae strains. Electron Microscopy revealed that phage Φ3 belongs to
the Myoviridae family while phages Φ6, Φ9 and Φ13 belonged to Siphoviridae family. Estimated
genome sizes of these phages were ~ 12.5 kb for Φ3 and ~ 30 kb for Φ6; while Φ9 and Φ13 were ~
48.5 kb in size.
Conclusions: The present study revealed that Φ3, Φ6, Φ9 and Φ13 have broad lytic ability for
Salmonella strains and could also lyse some of the other Enterobacteriaceae strains. After complete
characterisation of these four phages, they can be utilized for phage biocontrol of Salmonella on
plants.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section I: Agriculture and Food Safety 85
PD_RS08885 is required for phage infection and pathogenicity of Xylella
fastidiosa
Tushar Suvra Bhowmick1, 2, Mayukh Das1, 2, Tram T Le1, 2, Ry Young1, 3, Carlos F. Gonzalez1, 2
1Center for Phage Technology, 2Department of Plant Pathology and Microbiology, 3Department of
Biology and Biophysics, Texas A&M University, College Station, Texas.
Xylella fastidiosa (Xf) is the causal agent of Pierce’s Disease (PD) of grapevines. Our laboratory
has isolated and propagated phages active against Xf, and confirmed that Type IV pili (T4P) are the
primary receptor for Xf virulent phages. We have previously reported seven in vitro selected Xf
phage resistant mutants that are non-pathogenic, retained twitching motility and exhibited an
absolute adsorption defect for cocktail phages. Genome analysis of two in vitro selected Xf phage
resistant mutants identified a frameshift in PD_RS08885 (accession number WP_004089904.1).
The PD_RS08885 product is a membrane protein with six predicted transmembrane domain
helices. The selected phage resistant frameshift mutant exhibited a truncated protein with only two
predicted transmembrane domain helices. To confirm the observed results, an in-frame deletion
(XF-15 ΔRS08885) was constructed by replacing the PD_RS08885 open reading frame (ORF)
from nucleotides 1964420 to 1965178 (Xf Temecula 1 [accession number NC_004556.1]) with the
kanamycin resistance cassette using site-directed gene disruption. Complementation of XF-15
ΔRS08885 was accomplished by introducing a wild-type copy of PD_RS08885 using a
chromosome-based complementation system. The XF-15 ΔRS08885 was not sensitive to phages
Sano, Salvo, Prado or Paz, whereas the complemented derivative exhibited sensitivity to all four
phages. Light microscopy studies confirmed twitching motility in both the deletion mutant and
complement. Both the deletion mutant and complement formed biofilms in vitro. Furthermore, XF-
15 ΔRS08885 is non-pathogenic in grapevines but the complement caused development of typical
PD symptoms, whereas both have the ability to colonize and move in grapevines. Bioinformatics
analysis showed PD_RS08885 encoded a transmembrane protein, which has a PAP2 superfamily
domain. Our results indicate that a PAP2 superfamily domain containing PD_RS08885 is required
for Xf phages infection and may be involved in the PD pathogenic process in grapevines.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
86 Section I: Agriculture and Food Safety
Staphylococcus aureus Biocontrol Strategy by Yeast Engineering
- Displaying Phage Endolysin on Yeast
Jihwan Chun, Jaewoo Bai, Sangryeol Ryu*
Department of Food and Animal Biotechnology, Department of Agricultural Biotechnology,
Research Institute for Agriculture and Life Sciences, and Center for Food and Bioconvergence,
Seoul National University, Seoul, Korea.
*E-mail: [email protected]
Objectives: Pathogenic bacteria has long been a threat to human civilization. With the emergence
of multi-resistant bacteria, the development of novel approaches for eradication without the use of
conventional antibiotic agents has become very urgent. Bacteriophages have been explored for a
long time to resolve this issue, even though its application in food industry is still very limited. In
current study, yeast surface display system was selected as a novel endolysin application platform
to develop safe and promising biocontrol mechanism against pathogenic bacteria.
Methods: Staphylococcus aureus ATCC 13301 strain was selected as a target host and S. aureus
phage SA11 endolysin, LysSA11, was examined as a counterpart. Shuttle vector pCTCON, which
carries Aga2 anchor protein involved in yeast surface display system, was ligated to construct
pCTCON::LysSA11. The vector construct was cloned into replication host, Escherichia coli DH5α,
and furthermore, to the expression host, Saccharomyces cerevisiae EBY100 strain. Each
transformation product was selected on antibiotic supplemented media and confirmed by PCR and
sequencing. Cultivation and expression induction of yeast were conducted on synthetic media with
2% of glucose and galactose, respectively (SD-CAA, SG-CAA). Yeasts displaying LysSA11 (A:
8.0x108 CFU/mL, B: 1.0x109 CFU/mL) were applied to S. aureus in early exponential growth (A:
3.4x103 CFU/mL, B: 9.9x103 CFU/mL) at RT. (Group A: 16-hour induction / Group B: 21-hour
induction) Then, the mixtures were cultivated on Baird-Parker media for S. aureus enumeration
and assessment of inhibitory effect.
Results: Phage endolysin LysSA11 was successfully incorporated to pCTCON vector, which was
successfully transferred to E. coli DH5α and S. cerevisiae EBY100. Transformed yeast cells were
cultivated, induced, and LysSA11-displaying yeast cells were treated to S. aureus ATCC 13301
cells. In group A (16 h induction), 1.19 log CFU of S. aureus cells were reduced in 2 hours, and the
viable count was reduced by 3.53 log unit, which means complete inhibition, after 4 hours.
Meanwhile, in group B (21 h induction), S. aureus cells were inhibited by 0.75 log unit in 1 hour,
and 2.90 log unit in 2 hours, implying enhanced lytic activity with elongated induction.
Conclusions: We have constructed a novel yeast surface display system facilitated with
bacteriophage endolysin to successfully regulate pathogenic bacteria, S. aureus. The results suggest
that introduction of phage endolysin in eukaryotic system could express active form of proteins for
inhibition of bacterial pathogen.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section I: Agriculture and Food Safety 87
Storage of Staphylococcus aureus phages intended for food safety
applications
Eva González-Menéndez1, Lucía Fernández1, Diana Gutiérrez1, Ana Rodríguez1, Beatriz Martínez1
and Pilar García1*
1 Dairy Research Institute of Asturias (IPLA-CSIC). Paseo Río Linares s/n. 33300- Villaviciosa,
*E-mail: [email protected]
Bacteriophages have been proven as effective biocontrol agents for the food industry. However,
their extensive use requires the development of improved methodologies for the storage of
bacteriophages, and formulations compatible with their final application as biopreservatives. To
this aim, we determined the stability of four Staphylococcus aureus phages by using different
sample preparation techniques and storage conditions. First, we tested stability during medium- and
long-term storage at different temperatures (4°C, -20°C, -80°C, -196°C) and during lyophilization
(freeze drying) using different stabilizing additives (disaccharides, glycerol, sorbitol and skim
milk). Interestingly, phages belonging to the Siphoviridae family (phiIPLA88 and phiIPLA35)
turned out to be more stable at 4°C and -20°C than Myoviridae phages (phiIPLA-RODI and
phiIPLA-C1C). At lower temperatures (-80°C and -196°C), both siphophages and myophages
showed good infectivity after 24 months regardless of the stabilizer. Differences between families
were also observed after lyophilization. For instance, the addition of skim milk yielded a more
stable dry powder containing the phages, while sorbitol generally caused the highest reduction in
Siphoviridae phage titer after 24 months, in contrast to Myoviridae phages with similar stability in
both additives. Remarkably, storage of phages as lyophilized infected cells proved to be a suitable
option for unstable phages such as phiIPLA-C1C. As an alternative to facilitate storage and
transportation, we also explored phage encapsulation. Phage phiIPLA-RODI encapsulated in
alginate capsules retained high viability when stored at 4°C for 6 months and at room temperature
for 4 weeks. Considering the future application of phages as food biopreservatives, we also
explored the use of the spray-dryer technique to obtain encapsulated phages. Phage phiIPLA-RODI
retained their viability (<5 log units reduction) for 6 months at room temperature and phiIPLA88
up to 12 months in the presence of trehalose (<3 log units reduction). These results suggest that
dried powders might be a suitable way to deliver phages to food.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
88 Section I: Agriculture and Food Safety
Impact of the receptor preference on host range distribution of Salmonella-
infecting bacteriophages
Y. Emre Gencay*, Michela Gambino, Tessa From Prüssing, Lone Brøndsted
Department of Veterinary and Animal Sciences, University of Copenhagen, Stigbøjlen 4, 1870,
Frederiksberg C, Denmark
*E-mail: [email protected]
Salmonella enterica consist of more than 2400 serotypes, yet major foodborne infections in Europe
are often associated serotypes Typhimurium and the monophasic variant (4,[5],12:i:-), Enteritidis,
Infantis and Derby. One of the major contributor of this diversity is the O-polysaccharide (O-PS)
units attached to conserved core sugars of the lipopolysaccharide. Salmonella phages are known to
target either conserved or serotype specific receptors such as repeating units of O-PS, conserved
core sugars, or outer membrane proteins. Except certain model phages, the extent of the host range
of phages that can infect wild type food-associated Salmonella has been poorly described so far. In
this work, we have isolated, sequenced and determined the target receptor and the host range of a
large number of phages in order to associate phage families and the target receptors with the host
range potential of the Salmonella-infecting phages isolated from the environment.
With an attempt to isolate diverse phages we have used 10 different isolation hosts (7 serotypes)
and isolated 50 phages from samples either by direct spotting or after selective enrichment with the
isolation hosts. Plaque formations were determined in a large host range analysis using 71 Danish
Salmonella pork isolates of different serotypes often implicated in foodborne illness in Europe to
reflect a snapshot of the wild isolates. Using defined prophage cured S. Typhimurium LT2 mutants
we found that 9 phages were dependent on vitamin B membrane transporter (BtuB), whereas the
majority (n: 31) was targeting O-PS. While the receptor of three phages were found to be either
partially or totally masked by O-PS, the receptors of the remaining 7 phages could not be
determined.
Subsequently, 40 of these phages were sequenced using MiSeq platform, allowing us to group 31
distinct phages into phage families and associate these with host ranges. O-PS-dependent phages
showed three distinct host range pattern in themselves. Members of Jerseyvirus infects only up to
12 strains, whereas four temperate phages can infect slightly more strains. Among the O-PS-
dependent phages, three phages of Vi1virus infected the most strains (up to 20). Interestingly, none
of the O-PS-dependent phages could infect all strains of a certain serotype, indicating the absence
of serotype specificity. The BtuB-dependent phages were identified as T5virus had the broadest
host range, infecting up to 37 different strains, whereas a few T5virus dependent on an unknown O-
PS-masked receptor showed a more limited host range.
Statistical analyses of the host range showed that the target receptor is the foremost influencer of
the host range compared to isolation host or sample or assigned family. BtuB-dependent phages
belonging to T5virus genus have higher potential to cover the diversity of food-associated
Salmonella strains, making them better candidates for biocontrol purposes.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section I: Agriculture and Food Safety 89
Comparative study of phage antimicrobial proteins against Staphylococcus
aureus
Diana Gutiérrez1*, Silvia Portilla1, Lucía Fernández1, Beatriz Martínez1, Pilar García1 and Ana
Rodríguez1
1 Dairy Research Institute of Asturias (IPLA-CSIC). Paseo Río Linares s/n. 33300- Villaviciosa,
Asturias, Spain.
* E-mail: [email protected]
The rapid rise and dissemination of multi-drug resistant (MDR) pathogens represents a major
global long-term threat to human health. Recently, the World Health Organization (WHO)
published a list of antibiotic-resistant bacteria for which new antibiotics are urgently needed, where
Staphylococcus aureus, was classified as “Priority 2: High”, being of special interest S. aureus
resistant to methicillin (MRSA). Moreover, this bacterium is one of the major bacterial agents
causing foodborne diseases in humans due to their ability to produce enterotoxins. Likewise, the
complete elimination of this bacterium is a difficult task, in part because it can form biofilms on
biotic and abiotic surfaces. In this regard, the use of phage lytic proteins: endolysins and virion-
associated peptidoglycan hydrolases (VAPGHs) has shown a potent antimicrobial capacity as
therapeutic agents, biopreservatives against a range of pathogens and as antibiofilm agents. Herein,
the potential of two novel S. aureus endolysins, LysRODI and LysA72, derived from the
Myoviridae phiIPLA-RODI and the Siphoviridae phage phiA72, respectively was evaluated. In
addition, the lytic activity of the chimeric protein CHAP-SH3b, derived from the fusion of the
catalytic domain (CHAP) from HydH5 (VAPGH from phage phiH5) and the cell wall binding
domain of lysostaphin, was also compared. To do this, the three proteins were cloned into the
pET21vector, expressed in E. coli BL21 and purified by Nickel Affinity Chromatography. The
lytic activity of the proteins against S. aureus was compared by death kinetics measured by
turbidity assays and MIC. LysRODI showed the highest specific activity followed by CHAP-SH3b
and LysA72, meanwhile the lowest MIC was observed for CHAP-SH3b (16±0.2 µg/ml) and the
highest for LysA72 (79 ± 0.5µg/ml). The results obtained, allowed to stablish a protein
concentration to test their sensitivity to different environmental parameters such as pH, temperature
and salt concentration. Moreover, the effect of the presence of certain ions over the activity of the
proteins was tested, revealing that the Zn++ and Mg++ cations might negatively affect their activity.
The lytic spectrum of the proteins was tested against a collection of S. aureus strains, including
MRSA. Besides, the antibiofilm potential of the proteins was screened using the novel technology
‘xCelligence real-time cell analyzer’ (RTCA)’ and parameters such as MBEC50 (protein
concentration that removes 50% of the biofilm), LOABE (lowest protein concentration to observe
an antibiofilm effect) and specific antibiofilm activity were calculated. The results obtained so far
will be used to study the antimicrobial activity of these proteins in vivo using a zebrafish embryo
infection model.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
90 Section I: Agriculture and Food Safety
Viral diversity in rumen microbial communities of Colombian cattle and its
variation under dietary interventions.
Hernández, R1,2., Jiménez, H2., Caro-Quintero, A2., Reyes, A1*
1 Laboratory of Computacional Biology and Microbial Ecology BCEM, Universidad de los Andes.
2 Corpoica, Sede Tibaitatá.
*E-mail: [email protected]
Bovine animals are ruminants that consume fiber-rich diets based on grasses and small shrubs,
however with the aim to increase the production of milk and meat, diets rich in carbohydrates have
been incorporated, causing changes in the regular metabolism of food digestion and even some
diseases such as acidosis. The aim of this project is to describe the diversity of bacteriophages in
the rumen of Colombian cows that feed on tropical grasses, and how this diversity changes when a
carbohydrate rich food source like corn is introduced in the diet. High concentration of
carbohydrates from corn leads to a change in the pH values inside rumen, which in turns leads to
changes in the microbial community and potentially their phages. The ruminal fluid of five female
cows with similar characteristics (weight, age, etc ...) will be sampled with a cannula. Phage
diversity will be evaluated at different times. The first time will occur when cows feed on grass, a
second time at the moment of introduction of a change in the diet with corn and lastly, when the
original diet is re-established. Viral particles will be concentrated using ultracentrifugation or by
precipitation with polyethyleneglycol. DNA will be extracted using phenol-chloroform method. If
the concentration of DNA obtained is very low, random amplification of viral nucleic acids will be
performed. Shotgun sequencing approach will be used to characterize the viral-derived reads. The
diversity of phage before and after the change in diet will be established by bioinformatic analysis.
We hope to find interactions between the phage and rumen bacteria as they change in response to
the diet switch.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section I: Agriculture and Food Safety 91
Efficacy and safety of a Staphylococcus aureus phage cocktail in a murine
model of bovine mastitis
Koen Breyne1, Ryan W. Honaker2, Zack Hobbs2, Manuela Richter2, Maciej Zaczek2, Rebecca Lu2,
Anika Kinkhabwala2, Bruno Marchon2, Evelyne Meyer1, Lucia Mokres2
1Ghent University, Department of Pharmacology, Toxicology, and Biochemistry, Merelbeke,
Belgium 2EpiBiome, Inc. South San Francisco, CA
The aim of this study was to evaluate the efficacy of a purified phage cocktail for treatment of
bovine Staphylococcus aureus mastitis in a well-defined mouse model. Candidate phages were
selected based on their performance in a series of in vitro assays designed to test various phage
characteristics. Phages were tested individually and as cocktails to determine the most optimal
cocktail composition, a process referred to herein as phage scoring. The phage score incorporates
quantitative measures of killing efficiency in broth, time to resistance, host range, and other factors
predictive of a successful cocktail. The highest scoring phages were further tested for efficacy and
resistance suppression in broth and raw milk with and without supplemental IgG, and significant
decreases in CFU were observed. Based on these in vitro results the cocktail was then purified for
testing in the in vivo model.
Mammary glands were experimentally infected with S. aureus N305 (ATCC 29740), a clinical
bovine mastitis isolate that has been widely used for experimental infection of dairy cows and
mice. Lactating mice mammary glands were inoculated by intramammary infusion with S. aureus
and the phage cocktail treatment was applied via the same route four hours post infection. Treated
mammary glands were graded for gross pathological appearance and excised for bacterial load
(CFU) and phage load (PFU) quantification, and were compared to both sham- or antibiotic-treated
(the current standard of care) infected glands and non-infected controls.
Observation of clinical and gross macroscopic changes as well as quantification of CFU
demonstrated that the phage cocktail treatment significantly improved gross macroscopic clinical
observations, and significantly decreased intramammary bacterial loads. PFU indicated that the
tested phage cocktail treatment was able to maintain high local phage titers in the mammary gland
without spreading systemically. Further analyses (such as inflammatory cytokine profiling and
histopathology) will document the safety and efficacy of this candidate novel curative phage
cocktail mastitis treatment in the elegant preclinical model. These results present promising data
for phage therapy as an alternative to antibiotics for the treatment of bovine mastitis.
The first two and the last two authors contributed equally to this work.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
92 Section I: Agriculture and Food Safety
Search of lytic phages, an alternate for control multi drag resistant Pseudomonas
infections in trout
K. Porchkhidze, T. Khukhunashvili, E.Jaiani, G. Tsersvadze, M. Tediashvili, N. Janelidze
George Eliava Institute of Bacteriophages, Microbiology and Virology, Tbilisi, Georgia
Cold water fish breeding is one of the most rapidly developing sectors of Georgian
Aquaculture, however the presence of bacterial pathogens leads to fish death or damage of aquaculture
products and finally the huge economic losses. Some species, most commonly P. fluorescens and P.
putida, have been reported as fish primary pathogens in the high intensity fish aquaculture systems.
Emergence of drug resistant bacteria in aquaculture industry due to unrestricted use of antibiotics
clearly indicates the need in more sustainable and environmental friendly strategies for controlling fish
infections, such as bacteriophages.
The main goal of present work was to study the antibiotic susceptibility of Pseudomonas
strains isolated from trout farms and to isolate specific bacteriophages against target pathogens. More
than 200 strains of presumptive Pseudomonas spp. were collected from water and sick fish samples.
Based on the results of conventional phenotypic identification in combination with API 20NE
biochemical systems (Biomerieux, France) and genetic identification by Species Specific PCR 28
strains of P. fluorescens and 31 – of P. putida have been revealed. The susceptibility of identified
isolates of both Pseudomonas species to 18 antibiotics most commonly used in Georgian aquaculture
was studied by Kirby-Bauer disc diffusion susceptibility test.
The results of antibiotic susceptibility testing demonstrated the highest sensitivity (94-90%) of
P. putida isolates to Moxifloxacin, Amikacin and Kanamycin and considerably less to Tobramycin
(87%), Cefepime (84%), Colistin sulphate (84%) and Gentamicin (84%). P. fluorescens strains
showed similar effectiveness to same antibiotics with relatively higher sensitivity to Ciprofloxacin
(86%). All Pseudomonas strains demonstrated complete resistance to β-lactams, Furazolidon, Tylosin
and Metronidazol. Obtained data correlated with results of our studies on presence of Antibiotic
Resistance Genes (ARG). Namely, 41 % strains were found to be positive for the blaTEM-1 gene and
31.8% to sul1gene; none of them were found to possess qnrS and TetM genes. The multidrug
resistance and presence of ARG in tested Pseudomonas strains shows the pathogenic potential of these
isolates.
Four bacteriophages isolated from water environments of Georgia were selected out of 20
primary phage isolates. By virion morphology the newly isolated phages were attributed to Myoviridae
and Siphoviridae families and showed characteristics of lytic phages, particularly short latent period
and high burst size, also high survival rate in fresh water microcosms. Broad host range was one of the
main criteria for selection of candidate phages. The phage PpML11 lytic to fish isolate P. putida 1-51
expressed activity to 90% of tested P. putida strains, also in the spot test lytic zones were registered
on the lawns of 85% of P. fluorescens isolates.
Work on selection of therapeutic phage candidates for aquaculture and their detailed
characterization is on-going. For evaluation of antibacterial efficacy of selected bacteriophages small
scale in vivo experiments will be conducted on experimental fish Danio rerio.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section I: Agriculture and Food Safety 93
Chitosan Nanoparticles Loaded Bacteriophage Reduces Bacterial
Colonization and
Shedding in the Faeces of COBB Broiler Chicks
A. A. Kaikabo1,2*, M. S. Abdulkarim1 and F. Abas1
1Faculty of Food Science and Technology, University Putra Malaysia, 43300 UPM Serdang,
Selangor,
Malaysia 2Bacterial Research Department, National Veterinary Research Institute, PMB 01 Vom, 931001,
Plateau State, Nigeria
*E-mail: [email protected]
Avian pathogenic Escherichia coli O1:K1:H7 (APEC) causes infection in poultry leading to great
economic losses to poultry production globally. APEC contaminates finished poultry products thus
a problem to food safety, security and public health. The pathogen has been associated with urinary
tract infection in human and neonatal meningitis in children. It resists antibiotic therapy.
Alternative biological control measures using Bacteriophage has been explored in this work.
Bacteriophage(s) were isolated and characterized and a candidate phage has been selected based on
its characteristics and used in the biological control experiment against avian pathogenic
Escherichia coli O1:K1:H7. Oral application of phage therapy is affected by numerous factors such
as inactivation by gastric acids, enzymes and bile. We have synthesized and characterized chitosan
nanoparticles and loaded candidate phage (C-Φ NPs) to be protected and delivered. The C-Φ NPs
were applied in the in vivo experiments to assessed role in bacterial colonization in gastrointestinal
tract and shedding in the faeces of challenged birds. Microbiological culture of faeces from the
infected C-Φ NPs treated showed a decreased of viable bacterial counts from 1 to 3 weeks post
challenge compared with untreated groups. Similarly, at week 4 post challenge when the
experimental birds were euthanized culture of the intestinal scrub showed a decreased in viable
bacterial cell counts in C- Φ NPs treated group compared with untreated control respectively (p ≤
0.05). These results showed that C- Φ NPs is beneficial in reducing bacterial colonization and
shedding of APEC in the faeces of the infected birds. Thus, useful to public health, food safety and
security.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
94 Section I: Agriculture and Food Safety
Isolation of Vibrio phages from the environmental waters in Kenya and their
application as biocontrol agents against Vibrio cholerae
Alice Nyambura Maina1,2, Francis Mwaura1, Mirriam Jumba1, Samuel Kariuki3
1University of Nairobi, Kenya, 2 Technical University of Kenya, 3Kenya Medical Research Institute
*E-mail: [email protected]
Background: To date Kenyans’ continue to experience Epidemics of Cholera more frequently than
before. In 2014 Cholera outbreak was reported in Kenya and out of the 47 Counties it spread to 30
of them. 15000 cases were reported and about 250 deaths. In May 2015 a total of 3301 cases had
been reported and 65 deaths occurred with case Fatality rates of 2%. In March 2016, 216 deaths
took place and 13 000 admitted to Hospital following Cholera outbreak that affected many in the
country. Since the December 2014 outbreak Cholera has been recurring in Kenya. In May this year
(2017) three people died of Cholera after eating food contaminated with Vibrio cholerae in a
wedding ceremony and seven admitted in the hospital. Cholera outbreaks have also been reported
in refugee and internally displaced people camps. As a developing country, bacteriophages lytic to
Vibrio cholerae can be a cheaper and a novel approach to control cholera outbreaks in Kenya.
Objective: of this research is to isolate bacteriophages lytic to Vibrio cholerae from the
environmental waters of Kenya and apply them as biocontrol agents against the pathogen. The
phages will be disseminated in the water bodies contaminated with the pathogen in regions that
experience cholera epidemics every year.
Methods: A total of one hundred and twenty two environmental water samples used for domestic
purposes were collected from two regions: Lake Victoria and Coast between March 2015 and
October 2016. Water samples were collected from ponds, lakes, rivers, wells and boreholes. Vibrio
cholerae and other bacteria were isolated from these environmental waters. Plaque assay technique
was used to isolate the phages that were purified and amplified. The host range of the Vibrio
phages was tested using bacteria isolated from the same environment.
Results: Pathogenic Vibrio cholerae O1 Eltor was isolated from the environmental waters of the
two regions. The Vibrio phage isolated was lytic against Vibrio cholerae and two more bacteria
E.coli 083 and Providentia sneebia.
Conclusions: The environmental waters of Lake Victoria and Coast regions were found to be
contaminated with pathogenic Vibrio cholerae O1 biovar Eltor. The Vibrio phage was lytic against
Vibrio cholerae, E.coli 083 and Providentia sneebia. The Vibrio phage had broad spectrum and
hence it can be used as a cocktail to control these bacteria. More work is ongoing to extract phage
DNA, determine the genome size of the vibrio phage, TEM and concentrate them for use in water
decontamination.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section I: Agriculture and Food Safety 95
Isolation, characterisation and delivery of giant bacteriophages infecting the
pig enteric pathogen Salmonella enterica
Anisha Thanki1*, Nathan Brown1, Andrew Millard2, Gurinder Kaur Vinner3, Danish Malik3,
Charlotte Evans4 and Martha Clokie1.
1Department of Infection, Immunity and Inflammation, University of Leicester, Leicester, England; 2Unit of Microbiology and Infection, Warwick Medical School, University of Warwick, Coventry,
England; 3Department of Chemical Engineering, Loughborough University, Loughborough,
England; 4AHDB Pork, Stoneleigh Park, Warwickshire, England.
*Email: [email protected]
Salmonella can infect and cause diarrhoea in pigs at any stage of their growth, from newborns to
finishing pigs. The diarrhoea in turn then accelerates transmission in farms, and can ultimately lead
to Salmonella entering the human food chain and causing food poisoning. It is estimated that pigs
cause 11.7% of salmonellosis each year. Antibiotics are used to treat these infections and they are
also used as general growth promoters in some countries. This has led to a surge in antibiotic
resistant Salmonella strains responsible for numerous outbreaks on pig farms. As a consequence,
regulatory incentives worldwide have been introduced in an attempt to reduce antibiotics in the
food chain.
Bacteriophages are a promising alternative to treat Salmonella infections and their ‘Generally
regarded as safe’ (GRaS) status granted by the FDA means that currently their use as a food
additive is compliant with current regulation in the USA. The aim of our research was to isolate a
panel of bacteriophages that could be developed therapeutically to treat UK pigs and data will be
presented on the progress we have made to date.
We liased with farmers to collect pig faecal, boar faecal, and slurry samples, from which 22
myoviruses were isolated. All phages could infect multiple antibiotic resistant strains from the most
prevalent Salmonella serotypes associated with pigs, with significantly similar efficiency of
plating. Further genetic analysis revealed that all of the phages had what is referred to as ‘giant’
genomes at approximately 240 kbp. According to predicted protein similarity they cluster together
with two other genomes in the NCBI database from Salmonella phages previously isolated in
Korea. No phages contained predicted integrases or other genes that would obviously prevent their
being used to remove Salmonella from pigs.
To investigate which phages would be optimal for therapeutic use, in vitro ‘killing assays’ were
conducted with different phage combinations. The most effective formulations were two phage
combinations, which could reduce Salmonella by 4-logs in 2 hours when they were applied at an
MOI of 100. These optimal phage combinations were characterised in terms of stability and most
phages were stable at temperatures up to 60°C for one hour, whilst one phage was stable at 90°C.
Therefore phages remained viable after the spray drying process (converting phage lysate to
powder) and powdered phage preparations will increase the shelf-life and versatility of application
of phages in the pig industry. We are currently collaborating with farmers, vets and end users to
determine the optimum intervention point at which phages could provide maximum benefit in
reducing Salmonella in pigs.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
96 Section I: Agriculture and Food Safety
Optimisation of phage therapy against leek blight
Wagemans, J., van Charante, F., Rombouts, S., Volckaert, A., Pollet, S., Venneman, S., Maes, M.,
Van Vaerenbergh, J. and Lavigne R.
Every year, agriculture suffers from huge economic losses due to plant infectious diseases like
bacterial blight in leek. This bacterial infection is caused by Pseudomonas syringae pv. porri
(Pspo). Until recently, infected crops were still treated with copper or antibiotics like streptomycin.
However, since bacteria are becoming more and more resistant to these chemical treatments and
because of their risk for nature, alternatives are necessary. One possibility would be phage therapy.
Therefore, we first isolated five phages KIL1 to KIL5. These phages were characterized both
genomically and microbiologically (Rombouts et al., 2016). Moreover, one host range mutant
KIL3b was created, that infects 36 different isolates of Pspo. All phages were subsequently used in
preliminary field phage therapy trials in leek. A significant reduction (e.g. from 63% to 38.5%
symptomatic leek plants) of the disease symptoms was observed after application of the cocktail,
which is promising.
However, to be more effective, further research is needed. First of all, the application method has
to be optimized (so far, only one method was tested) and the composition of the phage cocktail has
to be further improved. One other important shortcoming that makes phage therapy commercially
less attractive is the fact that the identified phage cocktail often cannot be produced in an
industrially applicable way. Therefore, we optimized the production of the phage cocktail by using
an automated anion-exchange chromatography to purify the phages on CIM disks (BIA
Separations). This resulted in a fast method for the purification which is also suitable for large scale
production. Up to 80% recovery of the phages could be observed.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section II: Phage-Based Biotechnology 97
Bacteriophages as Tools for Precision Microbiome Therapeutics
Robert J. Citorik1*, Timothy K. Lu1,2
1Microbiology Program, 2EECS, MIT, Cambridge, MA
* E-mail: [email protected]
The various microbial populations residing on and within us are known to have intricate ties to
human health. Metagenomic and metabolomic analyses, in addition to laboratory research, are
revealing potential keystone members, groups, or pathways implicated in preserving or damaging
the health of the human host. Future translation will require the ability to validate and to effect
changes in microbiota based on knowledge gained from these studies, necessitating the creation of
next generation therapeutics with programmable specificity. Additionally, these antimicrobials
could prove pivotal in the battle against increasingly drug-resistant pathogens, which can also
constitute members of the microbiome.
Previously, we have shown the pairing of CRISPR-Cas9 with bacteriophages or conjugative donor
bacteria to create sequence-specific antimicrobials. These programmed phages selectively killed E.
coli harboring resistance or virulence determinants, while leaving “unarmed” E. coli unharmed.
Antimicrobial activity is based on genetic signatures, enabling other potential targets such as
metabolic pathways implicated in disease or therapeutic outcomes.
In addition to rational payload design, work has continued on phage scaffolds, which determine the
diversity of targetable bacteria. The opportunistic pathogen Klebsiella pneumoniae is a major
health concern and can be armed with extensive arrays of resistance genes. This microbe is a prime
candidate for phage therapy to treat infection, given its resistance potential, as well as prophylactic
therapy, since it can preexist in a healthy gut before spreading within or beyond the carrier to a
susceptible site or patient. Using recently developed phage engineering strategies, we are
characterizing a pipeline of bacteria-derived or newly isolated temperate and lytic phages against a
clinical panel of carbapenem-resistant K. pneumoniae for testing in a murine gut decolonization
model. Of particular importance for this group of organisms are varieties of phage enzymes aimed
at recognition and penetration of the extensive and variable capsules that typically protect the
bacteria from environmental and immune insults. The arsenal of capsule types that may be
employed by bacteria like K. pneumoniae presents a challenge in the development of targeted
phage-based therapies, which we seek to address through both analysis of our new isolates and
engineering efforts. Ultimately, we hope these efforts will not only yield phage(s) or scaffolds
optimized for eliminating the pathogen, but will also inform design rules for the rational creation or
modification of novel engineered derivatives.
Given appropriate knowledge and engineering technologies, including genome modification,
rational payload programming, and phage scaffold tuning, bacteriophages could prove pivotal in
the next generation of antimicrobials and precision microbiome therapeutics.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
98 Section II: Phage-Based Biotechnology
Phage endolysins: putative Clostridium perfringens therapeutic
Stephen M. Swift1, D. Treva Rowley1, Jerel Waters1, Brian Oakley2, David M. Donovan1*
1ABBL, ARS, USDA, Beltsville, MD.
2College of Veterinary Medicine Western University of Health Sciences, Pomona, CA
* E-mail: [email protected]
Clostridium perfringens is a major necrotic enteritis causing bacterial pathogen in poultry, and a
source of food poisoning and gas gangrene in people. C. perfringens can also cause mild to severe
enteritis in pigs. In the EU, the occurrence of C. perfringens associated necrotic enteritis in poultry
has increased as antibiotic use has decreased. As the US moves away from use of antibiotics in
animal feed, we can expect an increase in necrotic enteritis with subsequent losses from morbidity
and, in subclinical cases, losses from decreased chicken weights. Alternatives to antibiotics in
animal feed will be needed in the near future. The genomes of 43 unique C. perfringens isolates
from chicken were sequenced, examined for peptidoglycan hydrolase enzymes by homology to
known enzymes. There were more than 450 putative peptidoglycan hydrolases identified that
clustered into 15 groups according to homology [less than 50% amino acid identity between groups
and more than 90% amino acid identity within group]. Of 17 representative lysins cloned, four
lysins were shown to have high lytic activity against all 43 of the initial poultry isolates as well as 9
porcine (all tested) in plate lysis assays but not against other Gram-positive or Gram-negative
species tested. Activity was also demonstrated in zymogram and turbidity reduction assays. The
domain architecture and relative activity of the four lysins has been determined.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section II: Phage-Based Biotechnology 99
Discovery and Biochemical Characterization of PlyA54, PlyN74, and
PlyTB40 - Bacillus Specific Endolysins
Irina Etobayeva1,2, Sara Linden1, Farhang Alem3, Philip D. Mosier4, Allison A. Johnson5, Louise
Temple-Rosebrook6, Ramin H. Hakami3, and Daniel C. Nelson1,2,*
1Institute for Bioscience and Biotechnology Research, Rockville, MD
2Department of Veterinary Medicine, University of Maryland, College Park, MD 3National Center for Biodefense & Infectious Diseases, Biomedical Research Laboratory, George
Mason University, Manassas, VA 4Department of Medicinal Chemistry, Virginia Commonwealth University, Richmond, VA
5Center for the Study of Biological Complexity, Virginia Commonwealth University, Richmond,
VA 6Department of Integrated Science & Technology, James Madison University, Harrisonburg, VA
*E-mail: [email protected]
Three Bacillus spp. specific endolysins, designated PlyA54 (ORF 54 of phage Anthos), PlyN74
(ORF 74 of phage Nigalana), and PlyTB40 (ORF 40 of phage Tsar Bomba), were identified,
cloned, purified, and characterized for their antimicrobial properties. Bioinformatic analysis
revealed that all three endolysins have very different N-terminal enzymatic activity domains
(EADs) but similar C-terminal cell wall binding domains (CBDs). PlyA54 has a Peptidase-M15-4
EAD with a conserved SxHxxGxAxD metal binding motif, PlyN74 contains an Amidase-2 EAD,
and PlyTB40 possesses a MurNAc-LAA EAD. Both, PlyA54 and PlyN74, share 94% homology in
their CBD sequences, identified as a SH3b domain. In contrast, the PlyTB40 CBD only shared
~50% homology to the PlyA54 and PlyN74 CBDs, although it also displayed specificity toward the
Bacillus peptidoglycan. All three endolysins showed relatively broad antimicrobial activity against
Bacillus species with the highest lytic activity against B. cereus, with moderate but less effective
lysis noted for B. megaterium, B. pumilus, B. thuringiensis, and B. amyloliquefaciens. Surprisingly,
none of the enzymes demonstrated any measurable activity against several B. anthracis strains
despite high activity against closely related B. cereus species. Lytic activity of the EADs alone was
significantly diminished compared to that of the full-length endolysins, suggesting a necessary role
for the CBDs. Biochemical characterization showed that optimal lytic activity for all three
endolysins is in the physiological pH range (pH 7.0-8.0), over a broad temperature range (4ºC-
55ºC), and in the absence or low concentrations of NaCl (<50 mM). Out of all three endolysins,
only PlyA54 showed sensitivity to inhibition by EDTA. Dialysis of inhibited PlyA54 in buffer
supplemented with an excess of divalent metal ions (Ca2+, Mg2+) restored its lytic activity to ~80%
of the pre-inhibited level. PlyA54 showed much stronger lytic activity than PlyN74 and PlyTB40
on identical strains. These properties make PlyA54 a good candidate for further antimicrobial
development as well as bioengineering studies.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
100 Section II: Phage-Based Biotechnology
Phage display as a powerful tool to catch specific antibodies against ion
channels
Angela Holguin1, Wei Ye1, Fred Fellouse1
Department of Molecular Genetics, University of Toronto
The aim of this poster is to present a pipeline for isolation of new antibodies using synthetic
libraries against complex antigens as ion channels related to pain. There is a high interest to
develop a non-addictive non-opioid treatment for chronic pain as cancer pain, and different
antigens have been described as optimal targets. These targets could be used to fish antibodies with
phage display. This selection technique is a key tool in health science for the isolation of new
specific antibodies that can be used as treatment. There are critical factors that interfere in the
successful of the selection; for instance, the presentation of antigen. Many antigens as ion channels
are problematic at the presentation step due the difficulty to simulate the desire structural
conformation as in nature the are several changes of the protein. Ion channels, change the structural
conformation depend on the state of the channel, active state or inactive state. Here, we discuss the
phage display pipeline, problems along the process, and troubleshooting when working with
complex antigens as ion channels.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section II: Phage-Based Biotechnology 101
Vibriophage endolysins as a new antibacterial control strategy
Lina A. Zermeño-Cervantes, Sergio F. Martínez-Díaz, César S. Cardona-Félix, Carlos O. Lomelí-
Ortega*
Instituto Politécnico Nacional (IPN), Av. IPN s/n, Col. Playa Palo de Sta. Rita, La Paz, B.C.S.,
México. C.P. 23096. Phone 6121234658 ext. 81565
* E-mail: [email protected]
Vibriosis is a set of diseases of high incidence in the aquaculture sector, responsible for massive
mortalities and economic losses worldwide. Recently, multiple Vibrio parahaemolyticus strains
carrying pVA1 plasmid that encodes a binary toxin called PirAB were identified as the causal agent
of acute hepatopancreatic necrosis disease also known as early mortality syndrome
(AHPND/EMS). AHPND has generated significant economic losses in the major shrimp producing
countries since 2009, spreading rapidly to Asia and Mexico. However, the latest reports about the
presence of pirA and pirB genes in other Vibrio strains apart from V. parahaemolyticus strains
(VPAHPND) are concerning, since the problem is more serious and complex than estimated.
Attributed mainly to gene transfer versatility, the problem is exacerbated by the high degree of
bacterial resistance recorded in VPAHPND strains and the lack of new control strategies available to
address this aquaculture health problem. In this context, bacteriophage lytic enzymes (endolysins)
could be a promising antibacterial alternative. In this study, the recombinant endolysin of a marine
vibriophage called LysVPMS1 was obtained and its lytic activity against different VPAHPND strains
and other Vibrio species was evaluated. Interestingly, LysVPMS1 does not resemble any reported
endolysin. Furthermore, LysVPMS1 has a lytic effect against virulent strains of VPAHPND, V.
alginolyticus, V. harveyi, V. campbelli, and other non-Vibrio Gram-negative bacteria. Its optimum
lytic activity is around pH 6, and increases with Ca2+ ions. The enzyme retains its lytic activity after
being exposed at 50°C for 30 min. These findings show that it would be feasible to use it in
aquaculture due its broad-spectrum activity against various Vibrio strains. The next step will be to
evaluate their ability to prevent or control vibriosis under in vivo conditions. Currently there are
severe restrictions on the use of antibiotics, thus lytic enzymes encoded by bacteriophages could be
a novel promising and safe class of antibacterial alternative to reduce the risks in this industry.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
102 Section II: Phage-Based Biotechnology
Phage transduction for improvement of bacterial strains
Barbara Marcelli*, Jan Kok, Oscar P. Kuipers
Department of Molecular Genetics, Groningen Biomolecular Sciences and Biotechnology Institute,
University of Groningen
*E-mail: [email protected]
L. lactis is one of the most studied species of the Lactic Acid Bacteria (LAB); it is widely used in
the food industry in starter cultures for the production of a wide variety of cheeses and butter milk.
The consumers’ demand for improved products constantly pushes industries to develop even better
products with enhanced characteristics in terms of, for example, quality, taste or shelf life.
Bacteriophage transduction can be used as a naturally occurring tool to transfer genetic material
from one L. lactis strain to another, or in some cases even between LAB species, in order to
introduce desirable traits into industrially relevant strains.
A large number of bacteriophages has been isolated from the dairy environment, as the starting
material for this project: their genomes have been sequenced and annotated as a first step. A
subgroup of the phageshas been further characterized to get more insights into their nature and life
cycle. Finally, a number of the bacteriophages have been tested for their ability to transfer either
plasmid or chromosomal DNA among different LAB strains. The frequency of transduction, the
amount and the nature of bacterial DNA transferred have been examined to evaluate for the
applicability, in the industry, of phage-mediated horizontal gene transfer.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section II: Phage-Based Biotechnology 103
Anti-Biofilm Activity of the Endolysin PlyGRCS against Multi-Drug
Resistant Staphylococcus aureus
Sara B. Linden1 and Daniel C. Nelson1,2,*
1Institute for Bioscience and Biotechnology Research, Rockville, MD
2Department of Veterinary Medicine, University of Maryland, College Park, MD
*E-mail: [email protected]
The ability of Staphylococcus aureus to form biofilms represents a major virulence factor and
contributes to its ability to cause chronic infections. Furthermore, S. aureus can form biofilms on
medical devices, leading to colonization of areas of the body where it would not normally be able
to persist. Biofilms are notoriously difficult to eradicate, as they are resistant to antibiotics and are
resilient against the host immune system. Currently, the treatment for S. aureus biofilm infections
is debridement or removal of the affected implant, which is not a desirable outcome, and as such,
alternative therapies are needed. PlyGRCS, an endolysin which has shown antimicrobial activity
against planktonic S. aureus, represents such an alternative option. In this study, we investigate the
use of PlyGRCS against staphylococcal biofilms. PlyGRCS displays the ability to remove biofilms
from abiotic surfaces at concentrations much lower than its minimum inhibitory concentration. In
addition to its ability to disperse biofilms, as shown by crystal violet staining, in vivo imaging, and
confocal microscopy, PlyGRCS also kills bacteria within biofilms. PlyGRCS removes S. aureus
biofilms formed under dynamic conditions in medical-grade catheters, mimicking an in vivo
infection. Finally, PlyGRCS possesses the ability to kill bacteria within biofilms grown on
epithelium, without harming eukaryotic cells. In summary, we show that PlyGRCS has a potential
application an anti-biofilm therapeutic.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
104 Section II: Phage-Based Biotechnology
Structure-Guided Mutagenesis of the Bacteriophage Endolysin PlyC Cell
Binding Domain: From Octamer to Monomer
Xiaoran Shang1 and Daniel Nelson1,2,*
1Institute for Bioscience and Biotechnology Research, Rockville, MD 2Department of Veterinary Medicine, University of Maryland, College Park, MD
*E-mail: [email protected]
Objectives: The bacteriophage endolysin PlyC possesses potent lytic activity towards specific
streptococcal hosts. Molecular characterization of the plyC operon reveals that PlyC is composed
of two separate gene products, PlyCA, which contains the catalytic domains and PlyCB, which
serves as the cell binding domain. Additionally, the X-ray crystal structure indicates PlyC is a
holoenzyme consisting of a single PlyCA polypeptide tethered to a ring-shaped assembly of eight
PlyCB subunits. Our aims were study the binding mechanism and create new chimeric endolysins
through engineering the PlyCB monomer.
Methods: The crystal structure of PlyCB octamer was analyzed for residues that are involved in
intra-subunit interactions via hydrogen bonding and salt bridges. Systematic mutations to these
residues were generated through use of the Change-IT mutagenesis kit, and the structure (i.e.
octamer or monomer) of each mutant was analyzed by analytical gel filtration. Fluorescence
microscopy was used to assess binding of PlyCB and its mutants to streptococci.
Results: Mutations to Lys40, Asp41, and Glu43 were chosen for construction in seven
combinations. All of the PlyCB mutants were successfully expressed and purified. However, only
one combination, Lys40 and Glu43, indicated a monomer of PlyCB via gel filtration. Furthermore,
this mutant retained the ability to bind streptococci.
Conclusions: Structure-guided mutagenesis of PlyCB was successfully used to create a PlyCB
monomer. This PlyCB-derived cell binding domain retained its binding specificity for streptococci,
which will allow for additional bioengineering studies.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section II: Phage-Based Biotechnology 105
Evaluation of the tumor-limiting capacity of a phage-display selected peptide
motif in a human papillomavirus tumor model
Lanre Precieux Sulaiman1*, Ana Paula Lepique2 and Luisa Lina Villa1.
1. Faculdade de Medicina, Universidade de São Paulo, São Paulo, Brazil.
2. Instituto de Cinecias Biomedicas, Universidade de São Paulo, São Paulo, Brazil.
*E-mail: [email protected]
Objective: Women infected with high-risk human papillomaviruses (HPV) have an increased risk
for developing cervical neoplasia. To better understand the role of immune responses to these
viruses and disease development, we used phage display-based combinatorial selection to isolate
peptide ligands to antibodies present in the sera of HPV-positive women. Some peptides were
significantly enriched from a phage library displaying between 108 and 109 unique random
sequences. The most enriched motifs show different levels of homology to the L1 protein of both
low and high-risk HPV types. We want to see if the selected peptides had the capacity to limit
tumor growth in an HPV-model and if so, try to understand the mechanism of action.
Methods: We tested recombinant M13 phages containing an insert of the selected peptide - in both
prophylactic and therapeutic manners – against TC-1 tumor graft (an HPV model) in C57Black
immune-competent and knock-out mice. Mice were grafted with the tumor and then treated with
the phage injections (with or without adjuvants), or they were first immunized with the phage
injections and then grafted with the tumor. Thereafter, we analyzed tumor growth and kinetics by
pachymeter measurement, cellular infiltrate response by flow-cytometry and humoral response by
both phage-ELISA and peptide-specific ELISA.
Results: The mice immunized with the recombinant phage had reduced tumor growth when
compared with the controls and the mice that were treated at certain time-points after tumor
grafting also had varying degrees of tumor growth reduction. In all the cases, there seems to be
peptide specific stimulation of cytotoxic CD8 cells while also showing detectable peptide-specific
antibody production. This was further confirmed with the use of CD8 knock-out mice where
immunized mice lost the protection observed in the wild-type immune-competent specimens. There
thus seems to be a TH1 type response to the selected peptide that helps to reduce the rate of tumor
growth.
Conclusion: Phage-display is here proven again as a useful tool in selecting possible antigenic
epitopes in cancer and may offer complementary treatment options in HPV-positive cancers. While
our peptide may not offer full protection against the tumor, there may be a possibility for its
usefulness in a combination therapy. Further work is necessary to completely understand the
mechanism of action of the selected peptide.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
106 Section II: Phage-Based Biotechnology
Standardization of protocols to isolate phages against petroleum-souring,
sulfate-reducing bacteria
Santiago Hernández Villamizar, Martha Vives F.*
Universidad de los Andes, Bogotá D.C., Colombia
*E-mail: [email protected] , [email protected]
The oil industry is one of the largest income source in Colombia and is pillar of the world
economy. The souring is the increase of sulphide concentrations in the oil, consequence of the
reducing metabolism of anaerobic bacteria classified as sulfate reducing bacteria (SRB). The
souring affects the quality of the oil and the downstream process are more expensive. It also affects
the infrastructure of the industry, since the biocorrosion process deteriorate the material of the
pipelines and storage tanks. Therefore, the souring of oil and the biocorrosion generate economic
losses in this industry. At present these issues are controlled with biocides but it is necessary to find
other solutions because of its environmental impact, and the bacterial resistance to biocides.
The use of bacteriophages to control the populations of SRB is an alternative. This work seeks to
standardize the techniques to isolate effective bacteriophages against SRB since anaerobic bacteria
requires special conditions to thrive. An initial approximation was tested with Desulfovibrio
vulgaris as a host. Four treatments were carried out to assess the optimal growth of anaerobic
bacteria in solid medium. The best treatment was the traditional double layer method using 6 mL of
agar soft in the second layer. Moreover, waste samples and injection water used in oil recovery
were employed to isolate bacteriophages using direct and enrichment methods in anaerobic
conditions. Through the enrichment method was possible to visualize clear halos, which could
indicate the presence of bacteriophages in the samples. However, these results could not have been
replicated over time. The few Desulfovibrio sp. bacteriophages known in the literature are
temperate, which could be the reason for the no-replication of the said halos.
To test the reproducibility of the methodology of isolation of phages against SRB described in the
current work, new experiments with several SRB will be run. Besides, additional samples will be
tested in order to isolate lytic bacteriophages. The importance of the present work is based in the
fact of the little research made in the SRB bacteriophages; probably, for the demanding conditions
that require the anaerobic bacteria to thrive. Consequently, the result of this work will contribute to
the understanding of the anoxic bacteriophages and will provide clues to use bacteriophages in the
anoxic world.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section III: Ecology & Evolution 107
My Sabbatical (Spring, 2017)
Stephen T. Abedon
Dept. Microbiology, the Ohio State University, phage.org, phage-therapy.org, archaealviruses.org
E-mail: [email protected]
Some things new, some things old. I’ve spent 6 months checking things off my to-do list…
Abedon, S. T. 2017. Multiplicity of infection, Reference Module in Life Sciences. Elsevier.
Abedon, S. T. . 2017. Plaques, Reference Module in Life Sciences. Elsevier.
Blasdel, B. G., Abedon, S. T. 2017. Superinfection immunity, Ref. Mod. Life Sci. Elsevier.
Abedon, S. T. 2017. Phage "delay" towards enhancing bacterial escape from biofilms: a more
comprehensive way of viewing resistance to bacteriophages. AIMS Microbiology 3:186-226. (from
my talks in Liverpool and “Bacteriophage 2017”, both as available online; open access)
Abedon, S. T. 2017. Information phage therapy research should report. Pharmaceuticals 10:43.
(from my talk given in Stratford-Upon-Avon, as available online; open access)
Abedon, S. T. 2017. Commentary: Communication between viruses guides lysis-lysogeny
decisions. Frontiers in Microbiology 8:983. (based on more or less my entire career; open access)
Abedon, S. T., García, P., Mullany, P., Aminov, R. 2017. Editorial: Phage therapy: past, present
and future. Front. Microbiol. 8:981. (phage therapy review for the following; open acc.)
Aminov, R., Abedon, S. T., García, P., Mullany, P. 2017. Phage Therapy: Past, Present and
Future. See: journal.frontiersin.org/researchtopic/4111/phage-therapy-past-present-and-future
Howard-Varona, C., Hargreaves, K. R., Abedon, S. T., Sullivan, M. B. 2017. Lysogeny in
nature: mechanisms, impact and ecology of temperate phages. ISME Journal 11:1511-1520.
Alves, D. R., Abedon, S. T. 2017. An online phage therapy bibliography: separating under-
indexed wheat from overly indexed chaff. AIMS Microbiology 3:525-528. (open access)
Abedon, S. T. Post-proofs. Active bacteriophage biocontrol and therapy towards removal of
unwanted bacteria on sub-millimeter scales: from foods to microbiomes. AIMS Microbiology.
(from my Zurich talk, available online—finally got back to working on these… open access)
Abedon, S. T. Post-proofs. Bacteriophage clinical use as antibacterial "drugs": utility precedent.
Microbiology Spectrum and also in Cani, P., Britton, R. (eds.), Bugs as Drugs. ASM Press.
Abedon, S. T. Pre-proofs. Phage therapy: various perspectives on how to improve the art. Meth.
Mol. Biol. (also from my talk given in Stratford-Upon-Avon, as available online)
Abedon, S. T., Katsaounis, T. I. Pre-proofs. Basic phage mathematics. Meth. Mol. Biol.
Abedon, S. T. Pre-proofs. Why archaea are limited in their exploitation of other, living organisms.
Witzany, G. (ed.), Biocommunication of Archaea. Springer.
Abedon, S. T. Pre-proofs. Viruses of microorganisms: what are they and why should we care?
Hyman, P., Abedon, S. T. (eds.), Viruses of Microorganisms: Diversity and Applications. Caister
Academic Press. (we are desperately working on finalizing this multi-authored monograph…)
Abedon, S. T. Pre-proofs. Archaeal viruses. Ditto.
Abedon, S. T. Pre-proofs. Phage plaques. Harper, D., Abedon, S. T., Burrows, B. H., McConville,
M. (eds.), Bacteriophages: Biology, Technology, Therapy. Springer.
Abedon, S. T., Katsaounis, T. I. Pre-proofs. Phage enumeration via plaque counts. Ditto.
Dąbrowska, K., Górski, A., Abedon, S. T. Pre-proofs. Bacteriophage pharmacology and
immunology. Ditto.
Plus a whole lot of new web pages/postings, and still in preps. See abedon.phage.org for more.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
108 Section III: Ecology & Evolution
Surveillance of Shigella specific phages:
Potential use in epidemiology and diagnosis of Shigellosis
Mahmuda Akter*, Mahmuda Yeasmin, Sharmin Luna, Shahin Mahmud, Arafat Oany, Rubhana
Raqib and Kaisar Talukder
International Centre for Diarrheal Diseases Research, Bangladesh, GPO Box 128, Dhaka- 1000,
Bangladesh
*E-mail: [email protected]
Objectives: Global burden of shigellosis caused by Shigella species is >160 million cases yearly,
mostly affecting under-5 children in resource poor countries. The aim of the study was to monitor
the prevalence of Shigella specific phages in the environment and to utilize this in development of
an accurate diagnostic tool to accurately estimate actual disease burden.
Methods: A total of 180 water samples have been collected from rivers, lakes, canals and ponds
at 16 different sites in and around Dhaka, Bangladesh, for isolation of Shigella-specific phages.
The collection was carried out in two phases—during February, 2006, to January, 2007, and July,
2014, to December, 2015. Standard microbiological and biochemical methods were used for
isolation of phages. Among 142 isolated phages, 85 were characterized phenotypically using 57
serotypes of Shigella including atypical and new variants (n=200), 69 different serotypes of E.
coli (n=132), Salmonella (n=12), Vibrio cholerae (n=2) and Yersinia (n=1) used for testing
susceptibility to phages. PCR and RFLP were done for extensive characterization.
Results: The highest number of serotype diversity of Shigella specific phages was found in
canals, followed by rivers, lakes and ponds. A majority of phages isolated during 2006-2007 lysed
most of the serotypes of all Shigella species although very few phages were S. flexneri specific.
However, in 2014-2015 the pattern changed; very few phages lysed all Shigella species while
maximum number of phages lysed S. sonnei and S. flexneri, always including predominant
serotypes (2a, 2b, 3a, 1c, 6). icddr,b hospital data showed that isolation rate of S. boydi and S.
dysenteriae from patients was lower in 2014-2015 compared to 2006 while isolation rate of S.
sonnei gradually increased and S. flexneri remained broadly unchanged.
Among 85 phages, 4 phages were each specific for S. dysenteriae 3, S. dysenteriae 6, S. boydii 1
and S. sonnei. Again, one phage lysed all S. flexneri serotypes, S. dysenteriae 1 and S. sonnei and
was positive for capsid gene belonging to T4 type phage. Nine phages lysed S. flexneri serotypes
only, while five phages lysed combination of S. flexneri and S. sonnei.
A mixture of 9 phages lysed all serotypes of Shigella including atypical and new variants, and a
mixture of two phages lysed predominant serotypes of Shigella. These phages did not lyse other
enteric bacteria tested. A majority of the phages were different from each other as determined by
restriction pattern analysis.
Conclusions: Detection of Shigella specific phages may be used as a reliable indicator for
monitoring prevalence of Shigella in the environment and provide new insights into epidemiology
of Shigella infection. A single drop of cocktail-phage preparation may be used to diagnose any
Shigella. When applying commercially available polyvalent antisera, however, up to 16 antisera
may be needed to confirm. The isolated phages may also have therapeutic potential.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section III: Ecology & Evolution 109
The Human Gut Virome in Lean and Obese individuals
Laura Avellaneda-Franco1,6, Jaime L Moreno1, Lance Boiling2, Katrine Whiteson3, Andrew
Heath4, Forest Rohwer2, Jeffrey I. Gordon5, Alejandro Reyes1,5,6*
1Departamento de Ciencias Biológicas, Universidad de los Andes, Bogotá, Colombia; 2Department
of Biology, San Diego State University, San Diego, CA 92182; 3Departament of Biology,
University of California, Irvine, CA 92697; 4Department of Psychiatry, and 5Center for Genome
Sciences Washington University School of Medicine, St. Louis, MO 63132; 6 Max Planck Tandem
Group in Computational Biology, Universidad de los Andes, Bogotá, Colombia; 3Department of
Biology, San Diego State University, San Diego, CA 92182.
*E-mail: [email protected]
Objective: Viruses have been cataloged as the “dark matter” of biological sciences because of the
narrow knowledge that we have about them. The aim of this study was to characterize the gut
virome of lean and obese individuals and to assess their correlation with the BMI.
Methods: Fecal samples of 59 individuals with one to three sampling points (0, 3, or 12 months
after the first sampling) were obtained in part from the Missouri Adolescent Female Twin Study,
which consists of samples from female twin pairs and their mothers. Isolation of Viruses Like
Particles was performed and its DNA was extracted and amplified using MDA and sequenced
using 454 GS FLX pyrosequencing. The obtained reads were cleaned using Trimmomatic. Cleaned
reads from each individual were assembled using Newbler. Obtained contigs of all samples were
de-replicated at 90% of identity. Open reading frames (ORF) were predicted using Glimmer. Next,
the reads of each individual were mapped to a) the dereplicated contigs and b) the predicted ORF to
construct two abundance matrices, which were normalized using RPKM counts. De-replicated
contigs were annotated by comparing both the nucleotide and aminoacid sequences against the
NCBI non-redundant database. Also, α and β-diversity were estimated using QIIME from the
constructed matrices as a proxy to OTU tables. Finally, the relationship between specificity and
precision, and mutual information was used to select viral contigs that are specific markers to a
given individual, family, and/or BMI.
Results: We obtained in average 46,000 reads per sample. After the assembly, which incorporated
over 90% of the reads per sample, and de-replication, we obtained 16,223 contigs with at least 500
bp in length, we predicted that 3099 of these contigs were circular. Also, we predicted 44,885
ORFs. We found that the difference among the individuals virome can not be explained by either
the BMI or familial relationship, however, beta diversity analysis suggest that the virome within an
individual is stable over time. We identified discriminatory contigs that were characteristic at the
individual, family, and in certain BMI ranges.
Conclusion: The virome in adults is highly stable over time and its overall composition is a
signature of the individual and not their family or other physiological factor such as BMI.
However, a small set of viral contigs were identified as specific for individual families or BMIs.
Furthermore, we were able to identify viral contigs closely related to prophages contained within
sequenced bacterial genomes. In addition, certain viral contigs were identified with high abundance
and incidence among the sampled population, some of those with high identity to the crAssphage.
In summary, the human virome still poses challenges for our understanding and factors such as
BMI or family relationship do not seem to be major shapers of its diversity.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
110 Section III: Ecology & Evolution
Testing the Ferrojan Horse Hypothesis: Iron-Virus Interactions in the Oceans
C. Bonnain, S. Caprara, K.N. Buck, M. Breitbart*
University of South Florida, Saint Petersburg, Florida, USA
E-mail: [email protected]
Iron is an essential nutrient in the oceans, with the sub-nanomolar concentrations found in open
ocean surface waters often insufficient for supporting biological activity. More than 99.9% of
dissolved iron is bound to organic ligands, yet identifying the sources of these ligands in seawater
remains a major challenge. A significant portion of iron-binding ligands fall into the colloidal
fraction, which is operationally defined as the fraction collected between a 0.02 µm and a 0.45 µm
filter. Among the organic ligands in this fraction persists an extremely abundant biological
candidate: viruses. On average there are 107 viruses per milliliter of seawater, most of which are
phages (viruses that infect bacteria). The impact of viruses on ocean biogeochemistry is often
evoked purely through the act of lysing hosts and very few studies have considered the
geochemical potential of the viral particles themselves. Recent work in non-marine model systems
has revealed the presence of iron atoms within the structure of diverse phages infecting Escherichia
coli. Combined with the small size and sheer abundance of phages in the oceans, the inclusion of
iron in phage structures would translate into a major factor for cycling of this important trace metal.
In addition, iron is so critical for growth that bacteria have evolved multiple uptake systems for
assimilating iron, such as siderophores. Certain outer membrane proteins serve a dual function in
siderophore uptake and as a phage receptor, suggesting that some of the strategies utilized for iron
acquisition make bacteria vulnerable to phage infection. Given the constant arms race between
bacteria and phages to develop resistance and counter-resistance, respectively, it is not surprising
that phage would have evolved to utilize critical regions of surface-exposed proteins which are
indispensable for bacterial growth as receptors. The research presented here explores the potential
of marine phages to serve as iron-binding ligands and discusses the implications for both trace
metal biogeochemistry and marine phage-host interactions.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section III: Ecology & Evolution 111
How diverse are coliphages in the environment?
Stephen Ahern, Michela Gambino, Cæcilie Ingerslev Staffe, and Lone Brøndsted*
Department of Veterinary and Animal Sciences, University of Copenhagen, Stigbøjlen 4, 1870,
Frederiksberg C, Denmark
*E-mail: [email protected]
Escherichia coli is a highly versatile and adaptable species, exhibiting high degree of diversity and
consisting of both harmless commensals and pathogenic variants. In fact, it has been shown that
only 20% of the genes of the E. coli genome are shared among all strains, and there are over 200
serotypes based on somatic, capsular, fimbrial and flagella antigens. The E. coli surface diversity
has fostered highly diverse phages using many different host receptors and infection strategies,
with most phages only infecting a subset of E. coli. The E. coli Reference Collection (ECOR) is a
set of 72 reference strains isolated from a variety of hosts and geographical locations, which have
been shown to represent the genetic diversity of E. coli. The diversity within the E. coli species
makes designing phage biocontrol strategies difficult.
Here we aim to identify phages able to infect diverse E. coli. Using the ECOR library as hosts, we
isolated 128 phages from a variety of environmental samples, infecting 56% of ECOR strains.
Sixty-eight phages were selected for whole genome sequencing using MiSeq. Based on sequence
homology and genome organization, 20 phages could be grouped into 8 different and unrelated
phage types, most of which have not been characterized previously. Within phage types, related
phages were isolated from multiple environments and locations. We set out to identify phage
receptors and other host genes required for infection by using positive phage selection on a library
of Tn5 transposons of phage sensitive ECOR strains. Of the phages in our collection, we identified
phage receptors including outer membrane proteins such as OmpC and FhuA, as well as several
LPS moieties. These findings are currently being confirmed using clean deletion mutants. Our
work highlights the difficulty of isolating phages able to cover a large host range, even within one
species. It also demonstrates that E. coli phages are highly diverse and that seemingly unrelated
phages have adopted similar host recognition and adsorption strategies.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
112 Section III: Ecology & Evolution
Impact of a dominant temperature-dependent phage on the seasonal
population dynamics of a tropical bacterial pathogen assessed by
mathematical modelling.
Andrew Morozov, Halil Egilmez, Martha Clokie and Ed Galyov*
University of Leicester, UK
*E-mail: [email protected]
Melioidosis, a serious environmentally-acquired bacterial infection, is often characterised by a
noticeable seasonality. Burkholderia pseudomallei, the causative agent of melioidosis, is
commonly found in tropical soil and stagnant waters that represent environmental reservoirs of the
disease. Environmental populations of B. pseudomallei are known to be regulated by a variety of
biotic and abiotic factors, but the contribution of different factors to the seasonal dynamics of B.
pseudomallei population is poorly understood. Mathematical modelling has long been recognised
as a powerful, efficient and reliable research tool to predict population dynamics. Here we build
and explore a novel mathematical model to describe and predict both the daily and seasonal
dynamics of the B. pseudomallei population and associated phages. The model focuses on the size
and composition of B. pseudomallei populations that are controlled by phages in the surface water.
The model is centred on our recent findings that a particular clade of phages, which is highly
abundant in the endemic area of Thailand, differently infects B. pseudomallei depending on the
temperature conditions: phages are predominantly lytic at a higher temperature of 37oC and mainly
lysogenic at 25oC. Lysogens survive in the environment at lower temperatures, but are killed and
release their viruses when they are then put into elevated temperatures of 37 oC.
Our model explores the effects of daily and seasonal variations in the environmental temperature
that result in frequent temperature-dependent switching between the types of phage-bacteria
interactions that affect the B. pseudomallei population. The model is based on a system of
differential equations that describe the densities of bacteria in the phage-free, lysogenic and lytic
states as well as the number of free phages. The parameters of the model are informed by our
research, other scientific publications and publicly available records. Various scenarios of the
seasonal progression have been investigated. Based on our extensive computer simulations, we
explored the sensitivity of model predictions on a few key parameters. Our simulations predict the
existence of three main types of fluctuations of the size and composition of bacterial population in
the field: short-term (daily), intermediate (from 1 week to a month) and longer-term (seasonal)
fluctuations. Our striking conclusion from this work is that most of the modelling outcomes appear
to correlate with the apparent higher risk of melioidosis acquisition during the “warm and wet”
season where the phages are predominantly undergoing replication in the lytic cycle and the
bacterial populations are dominated by phage-free B. pseudomallei. The model predicts annual
blooms of phage-free bacteria, and during such periods B. pseudomallei are more likely to establish
an infection. Our data suggest that seasonality of melioidosis and potentially other environmentally
acquired bacterial infectious diseases could be explained by the action of dominant condition-
dependent phages.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section III: Ecology & Evolution 113
New insights into crAssphage
Emma Guerin1,2*, Andrey Shoporov1 Angela McCann1 , Paul Ross1,2 , Colin Hill1,2
1 APC Microbiome Institute, University College Cork, Ireland; 2 Department of Microbiology,
University College Cork
*E-mail: [email protected]
In 2014, Bas E. Dutilh et al. discovered a previously unidentified phage with a circular genome of
~97kb with 80 predicted open reading frames. This phage was named crAssphage based on the
cross-assembly programme used for its assembly. It was found to be highly prevalent in the human
gut, making up as much as 90% of the sequencing reads in faecal viral metagenomes. Through co-
abundance analysis it was predicted that a member of the Bacteroides genus is likely to be the main
host for this phage. However, wet lab work has yet to prove these in silico predictions.
Since the publication of the crAssphage genome there is little knowledge on its significance. Much
of the knowledge generated thus far is in silico based, but given its prevalence it is likely to have a
major role in influencing the bacterial population of the gut. To further investigate in silico
predictions, wet lab experiments are key. Of particular interest is confirming whether Bacteroides
is in fact the host for crAssphage. Gaining insight into phage morphology as well as how it
interacts with and replicates itself within the host may help to shine light on how it helps to shape
the gut microbiota.
The aim of this work is to reinforce bioinformatic predictions by confirming the host for
crAssphage and generating information on its role in the gut environment, using faecal samples
known to be crAssphage rich, based on bioinformatics data. The experimental pipeline developed
for this analysis may prove useful for other applications such as the examination of phage-host
interactions and the role of phage in influencing disease states in the gut.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
114 Section III: Ecology & Evolution
Monitoring Pseudomonas aeruginosa bacteriophages in the environment
Ermir Kadija
”Luigj Gurakuqi” University of Shkodra, Albania
E-mail: [email protected]
Obligately lytic bacteriophage isolation is crucial for phage therapy. Isolation from environmental
samples is the simplest method and often times easy. However, isolation of certain phages, such as
S. aureus phages, remains difficult and challenging. For this reason we have monitored
Pseudomonas phages in environment from May - October 2012. Summer months were chosen for
being the hottest months of the year with temperature reaching 40˚C. Samples from a water
discharge that run on the river Buna were used for phage monitoring and water temperatures were
also monitored.
Environmental samples were collected monthly and delayed two days later if sites experienced
rain. They were centrifuged, filtered, and plated using a double agar layer protocol on a
Pseudomonas aeruginosa strain. The following day, plates were checked for phage plaques.
Results show a correlation between phage titer and temperature with August having 172 PFU mL-1
and the lowest phage titers were observed in May and October with 4 PFU mL-1. These results
correspond with temperature results since on August water temperature was 23˚C.
Repeating the same experiment with S. aureus we weren't able to isolate phages but a specific
phage for S. aureus was possible to isolate using an enrichment protocol with a 50 mL
environmental sample. This data corresponds with previous data with the number of phage
particles increasing proportionally with availability and density of bacteria (temperature).
However, the diversity of phages for P. aeruginosa was higher than that of S. aureus since 3
different phages for Pseudomonas were isolated with direct plating.
As the results shows that temperature is a factor of successful phage isolation, more research is
necessary in this subject since more factors must influence phage isolation like sewage chemical
composition, phage stability, burst size, bacterial growth requirements, bacterial generation time,
etc.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section III: Ecology & Evolution 115
Isolation and characterization of two novel cyanophages from a subarctic
Canadian lake
Alice Lévesque1,2,4,5*, Antony T. Vincent2,5, Simon J. Labrie2,4, Sylvain Moineau2,4,
Warwick F. Vincent1,3 & Alexander I. Culley1,2,4,5
1 Centre d’Études Nordiques (CEN), Université Laval, Québec, Canada; 2 Dept. of Biochemistry,
Microbiology and Bioinformatics, Université Laval, Québec, Canada; 3 Dept. of Biology,
Université Laval, Québec, Canada; 4Groupe de recherche en écologie buccale (GREB), Université
Laval, Québec, Canada; 5Institut de biologie intégrative et des systèmes (IBIS), Université Laval,
Québec, Canada
*E-mail: [email protected]
Cyanobacteria are key components of arctic aquatic ecosystems, as both essential primary
producers, and as significant contributors to the biogeochemical cycling of essential nutrients such
as carbon and nitrogen. The two primary mechanisms of top-down control of cyanobacterial
populations are grazing and viral lysis. Viruses are the most diverse and abundant biological
entities on Earth with estimates of 1031 viral particles globally and 104 virotypes per mL of water. It
is now widely recognized that viruses are important drivers of host evolution, and influence
nutrients cycles and microbial dynamics through viral lysis. Cyanophages, viruses that infect
cyanobacteria, infect keystone primary producers in marine and freshwater environments, and have
been shown to harbour host-like genes that are involved in essential host metabolic processes. In
order to better understand cyanophage dynamics, genomics and ecology, we focused our research
on the isolation and characterization of cyanophages that infect polar cyanobacteria.
Water from a Canadian subarctic lake located in the vicinity of Whapmagoostui-Kuujjuarapik
(Nunavik, Canada) was sampled in August, 2015, and was filtered through 0.45µm and 0.22µm
pore size filters to remove large particles (cells, clay, etc.). The filtrate containing viruses was used
to inoculate different cyanobacterial cultures for viral amplification. Cultures presenting signs of
lysis relative to an untreated control were serially diluted to isolate a single virus. Clonal isolates
were subsequently sequenced on a NGS sequencing platform (MiSeq, Illumina).
We succeeded in isolating two novel cyanophages (B3 and B23). Electron micrographs revealed
that both isolates possessed icosahedral capsids and long contractile tails, typical features of viruses
in the order Caudovirales, family Myoviridae. B3 and B23 genome lengths were 244,930 bp and
243,633 bp, respectively. Genomic analysis and ORF annotation indicated the presence of two
host-related metabolic genes (hli and phoH) in both genomes. However, the function of only 25%
of the putative ORFs was identified using RAST based on homology with sequences in the SEED
database. Phylogenetic analysis based on alignments of the B3 and B23 primase genes and those
from phages from a representative number of taxa showed that both viruses clustered with other
cyanophages. These results suggest that B3 and B23 are highly divergent from previously
described cyanophages and that this study will result in new insight into the world of arctic
cyanophages, of which yet little is known.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
116 Section III: Ecology & Evolution
Which are the host factors influencing the replication of virulent
bacteriophages in the gastrointestinal tract?
Marta Lourenço1, Matthieu Galtier1, Damien Maura1, and Laurent Debarbieux1*
1 Group Interactions Bacteriophages Bacteria in Animals, BMGE Unit, Microbiology Department,
Institut Pasteur
*E-mail: [email protected]
Virulent bacteriophages have been frequently proposed as alternative antibacterials to target
bacterial pathogens resistant to multiple antibiotics. Often, some of these pathogens are
asymptomatically carried in the gastrointestinal tract (GIT) of mammals. However, despite several
attempts using different animal models, the use of virulent bacteriophages to efficiently reduce
carriage levels have often been disappointing, achieving at best a moderate reduction. We believe
the lack of knowledge on the factors that influence the replication of bacteriophage in the GIT is
impeding the development of strategies to improve overall bacteriophage efficacy.
To address this question, we characterized nine bacteriophages isolated using three different strains
of Escherichia coli: the enteroaggregative strain 55989, the adherent invasive strain LF82 and the
uropathogenic strain AL505. Each of these E. coli strains was used to colonize the GIT of mice.
Using homogenized GIT samples from these mice, we assessed the ex-vivo replication of these
bacteriophages in different sections of the GIT. We found that some bacteriophages replicate
efficiently regardless of the GIT sections while others display variable replication efficiency.
Therefore, these data support our hypothesis that the local GIT environment is influencing the
infectivity of bacteriophages, raising questions regarding the identification of the actors involved
and how, in the future, we could design ways to improve overall efficacy of phage therapy
treatments.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section III: Ecology & Evolution 117
Metagenomic time series reveal temporal variations within the viral
community in the Baltic Sea
Emelie Nilsson*, Daniel Lundin, Stina Israelsson, Jarone Pinhassi, Karin Holmfeldt
Linnaeus University, Faculty of Health and Life Sciences, Department of Biology and
Environmental Science, Centre for Ecology and Evolution in Microbial Model Systems (EEMiS),
391 82 Kalmar, Sweden.
*E-mail: [email protected]
Viruses infecting microbes are numerous and play an important role in aquatic ecosystems,
influencing biogeochemical cycles, such as cycling of carbon, nutrients and energy. To increase
our knowledge regarding viruses, metagenomic studies, such as Tara Oceans and the Pacific Ocean
Virome, are important. Such studies have improved our knowledge regarding the great diversity of
aquatic viruses as well as their spatial distribution and variation. However, temporal dynamics of
viral communities is a yet unexplored area within the field. Here, we seek to address this
shortcoming by presenting data from 25 viromes taken across all seasons from 2012 to 2015 at the
Linnaeus Microbial Observatory in the Baltic Sea.
Each time-point was assembled into contigs and annotated with VirSorter and DIAMOND against
a database of all viral genomes in NCBI. A majority of the contigs had little resemblance with
known viruses, but several were large enough to indicate that they are complete or nearly complete
viral genomes. In this study, we investigated large contigs (>20 genes, 290-2449 contigs per
sample). These contigs were diverse and showed similarities to globally abundant T4-like
cyanophages and pelagiphages, but also to more rare Cellulophaga phages, isolated from the Baltic
Sea, and phycodnaviruses. By recruiting reads from each time point back to the contigs from all
time points, we were able to investigate temporal patterns of the large contigs. These showed most
similarity to Myoviridae, Podoviridae, Siphoviridae and Phycodnaviridae, but the majority of
ORFs did not show any resemblance to known viruses. To understand why the relative abundance
of the families varied over time, we selected the contigs that mainly contributed to the increases in
relative abundance and studied the temporal abundance in more detail. The contigs similar to T4-
like cyanophages and pelagiphages were present and had a relatively high abundance in all
samples. Contigs similar to Cellulophaga phage phi38:1 were also present, but the relative
abundance was low except for two samples in spring 2012, in which it rivalled the dominating
pelagiphage. An increase in relative abundance during late summer and autumn could be seen for
the Synechococcus phage S-CBS4, while other Synechococcus phages within Siphoviridae showed
no such seasonal trend.
Overall, this study shows novel temporal patterns within viral communities that indicate seasonal
variability in diversity and the next step is to discover the relationship between viral populations
and microbial succession.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
118 Section III: Ecology & Evolution
Coevolution of human gut Enterococcus faecium with lytic phage V12
Andrew Oliver1, Stephen Wandro1, Tara Gallagher1, Claudia Weihe2, Whitney England1, Jennifer
B.H. Martiny2, Katrine Whiteson1*
1 Department of Molecular Biology and Biochemistry, University of California, Irvine; 2
Department of Ecology and Evolutionary Biology, University of California, Irvine
*E-mail: [email protected]
Bacteriophages are highly abundant in the human microbiota, but their effect on the composition
and diversity of resident microbes is largely uncharacterized. Phage predation can drive the
evolution of bacterial resistance, which can then drive reciprocal evolution in the phage. These
coevolutionary dynamics have been extensively studied in few ecological systems, but studies
relating to the human microbiome are lacking. We performed experimental evolution and
coevolution of isolates of Enterococcus faecium from healthy human stool with or without a single
infecting T4-like bacteriophage, V12. Quadruplicates of E. faecium and phage were grown together
in semi-continuous culture for eight days, diluting in fresh BHI media every twelve hours. In the
evolution scenario, bacteria were either grown alone or naively reintroduced every twelve hours
with an evolving phage, whereas in the coevolution scenario, both bacteria and phage were grown
in the same microcosm. At several time points during the experiment, bacterial and phage DNA
from each replicate was extracted and sequenced. Unique to the coevolution scenario, E. faecium
obtained small nucleotide polymorphisms (SNPs) in a gene related to exopolysaccharide biogenesis
and export. Phage V12 evolved SNPs in a capsid protein and obtained varying amounts of 1.8kb
duplications in the tail fiber gene when coevolving with E. faecium. Host range assays confirmed
that coevolution was directional where phage resistance and host infectivity increased over time.
This study provides insight into genes that may be involved in coevolution in this novel bacteria-
phage system.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section III: Ecology & Evolution 119
Identifying gene signals for phage host-range prediction
Jason Shapiro1, Siobhan C. Watkins2, Catherine Putonti1*
1Loyola University Chicago, Chicago, IL; 2New Mexico Tech, Socorro, NM
*E-mail: [email protected]
Objectives: As more novel, uncultured phage genomes are published, new tools are needed for
placing these genomes in an ecological and evolutionary context. There are no universal genes
shared by all phages, and horizontal gene transfer between viruses is common. In essence, every
phage genome is a mosaic that reflects the often disparate evolutionary histories of its genes. Host
range, in particular, constrains viral ecology and evolution, and predicting a virus’ host is a key
challenge when characterizing novel, uncultured genomes. Taking a computational approach, our
work is aimed at improving methods for phage-host prediction.
Methods: We have built gene-level networks representing the co-occurrence of genes across phage
genomes. In these networks, nodes represent a set of homologous genes (across phage genome
sequences) and two nodes are connected by an edge if the genes co-occur within the same genome.
This is a departure from genome-based networks, which have previously been used for recovering
taxonomic groups of phages, in which nodes represent genomes and edges signify the presence of
homologous genes. Taking this gene-centric approach, we sought to quantify the reliability of
observed clusters within the network and to ask if subsets of genes could be used to predict a
phage’s host. We developed an evolutionary algorithm to identify the subset of genes that
maximizes the mutual information of clusters and hosts.
Results: Here, we first show that clusters of genes within this network provide a robust view of
virus genetic diversity. Gene-level networks provide a higher resolution view of phage genetic
diversity and offer a novel perspective on virus ecology – in particular viral host range. We can
predict virus host range at the genus level with over 80% accuracy for many host genera. This
exceeds the accuracy of current bioinformatic approaches for host prediction.
Conclusions: Gene-level networks can provide a high-resolution view of viral genetic diversity
and have the potential to provide meaningful insight into predicting virus-host interactions. We
expect similar methods can be applied to other aspects of viral ecology, including isolation source
(e.g. freshwater, marine, soil, leaf, gut, hospital, etc.) and abiotic or biotic factors that might vary
across these locations (e.g. temperature, pH, O2, nutrient concentrations, and available host
diversity).

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
120 Section III: Ecology & Evolution
Broad host range – lessons from the literature
Alexa Ross and Paul Hyman*
Ashland University, Ashland, Ohio, USA
*E-mail: [email protected]
A bacteriophage’s host range is an important characteristic. While conceptually simple, host range
determination has several technical challenges. First, it can only be tested on the species and
strains available to the tester, which is by necessity a subset of potential hosts found outside the
laboratory. As well, there are few standard sets of strains for any particular bacterial species.
Second, host range varies depending on the specific testing method used. Different methods will
show varying host ranges even when done on the same hosts by the same researchers. It is quite
common, for example, for a bacteriophage to show a different host range on a single set of hosts
when tested by spot testing versus by plaque formation. Third, the terminology of “narrow” and
“broad” host range is inconsistently used.
In our work isolating bacteriophages, and in the literature, there is indication that isolation methods
may bias the host range of novel isolated bacteriophages. Specifically, it appears that isolating
bacteriophages with a mixture of host strains favors broader host range phages. In order to test this
contention, we have surveyed the literature on newly isolated bacteriophages published over the
last 70 years on a wide variety of bacterial types. Not surprisingly, the majority of phages were
isolated on a single host, but there are a few examples of multi-host isolation. While there are not
enough examples to make definitive statements, there is certainly enough evidence to support
further exploration of multi-host isolation.
We also note that when host range is tested, even on single-host isolated phages, host range appears
to be a trait with significant diversity between bacteriophage. Both narrow and broad host range
phages are often isolated at the same time on the same host strain suggesting that both are
reasonably common. In the few cases where hosts of multiple genera are tested, as opposed to just
strains of the same species, both single-species and multiple genera (polyvalent) host-range phages
can be isolated.
Our literature review suggests that the belief that bacteriophages are generally species specific may
be less true among phages as a group, perhaps due to a narrower host-range bias resulting from
single-host isolation procedures. Already isolated phage species may also harbor greater host
range diversity that has not been identified because generally the complete potential breadth of
phage host range is never fully tested. This has implications for the evolution and ecology of
bacteriophages as well as for phage therapy applications, for which polyvalency is not adequately
considered even as broad host range on the target host species is explored.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section III: Ecology & Evolution 121
Genome-derived insights into interactions between cyanophage Vb-AphaS-
CL131 and filamentous cyanobacteria Aphanizomenon flos-aquae
Sigitas Šulčius1,3*, Gediminas Alzbutas1, Eugenijus Šimoliūnas2, Emelie Nilsson3, Giedrius
Gasiūnas2, Inga Songailienė2, Vykintas Jauniškis2, Jolita Kemeraitė1 & Karin Holmfeldt3
1 Nature Research Centre, Vilnius, Lithuania; 2 Vilnius University, Vilnius, Lithuania; 3 Linnaeus
University, Kalmar, Sweden
*E-mail: [email protected]
Filamentous diazotrophic cyanobacteria Aphanizomenon
flos-aquae contribute significantly to community
composition and biogeochemical processes in many
temperate lakes and brackish water ecosystems, among
other locations in the Baltic Sea. Recent experimental
studies demonstrated an important role of cyanophage
infections in A. flos-aquae population dynamics, with
implications for food web structure and functioning. In
addition, metagenomic and transcriptomic surveys have
indicated that cyanophages are common and active in the
Baltic Sea. However, our understanding of the genomic
basis of phage-host interactions in these ecologically relevant cyanobacteria and their viruses
remains elusive. Therefore, in this study we performed genome sequence analysis of a cyanophage
Vb-AphaS-CL131 (CL131) and A. flos-aquae host strain 2012/KM1/D3 (D3), to better understand
the complexity of their reciprocal interactions at the genetic level.
Cyanophage CL131 has a linear dsDNA genome 112,793 bp in length. The genome contains 147
predicted protein coding genes and 2 tRNA genes. The majority of the predicted genes have no
detectable homologues in the present databases and the genome showed no co-linearity with
previously sequenced genomes of other cyanophages. The recruitments of CL131 reads from
available and local viral metagenomes are currently being performed to represent spatial and
temporal distribution of CL131 cyanophage. Neither integrase genes were found in CL131 genome
nor prophage sequences in the D3 genome, suggesting that CL131 does not enter into lysogenic
interactions with their hosts. No genes related to photosynthesis and carbon metabolism were
found, implying that CL131 replication and progeny formation strongly relies on host metabolism
and its intracellular resources. Cyanophages encode putative Class 2 type V-U4 CRISPR-Cas
system; those complexes are composed of single Cas protein and crRNA. Northern blot analysis
confirmed transcription of cyanophage CRISPR region during cyanophage infection. On the other
hand, host strain D3 encodes an I-D type CRISPR system that has a spacer that is complementary
to CL131 cyanophage genome, implying that I-D type CRISPR system may provide resistance to
the host against CL131 infection. The simultaneous genomic characterization of both cyanophage
and its host should enable better insights into possible virus-host interactions of bloom forming
filamentous cyanobacteria.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
122 Section III: Ecology & Evolution
Identification and characterization of bacteriophages in global sewage
samples
Henrike Zschach1*, Barbara Lindhard1, Vanessa Jurtz1, Frank M. Aarestrup2, Morten Nielsen1,
Ole Lund1
1 Department of Bio and Health Informatics, Technical University of Denmark (DTU), 2800 Kgs.
Lyngby, Denmark; 2 National Food Institute, Technical University of Denmark (DTU), 2800 Kgs.
Lyngby, Denmark
*E-mail: [email protected]
Sewage is a major source of both human pathogens and their associated bacteriophages. Phages
control bacterial populations by predation and can act as natural reservoirs for accessory genes
such as antimicrobial resistance genes and virulence factors. However, currently limited knowledge
is available about the sequence and functional diversity of such sewage phage communities. We
here present a study of the phage communities of 81 sewage samples from 62 different countries
around the world.
The samples consist of metagenomic assemblies in which we identified phage contigs by using the
MetaPhinder tool. These contigs were subsequently screened for the presence of known virulence
and resistance genes with the VirFinder and ResFinder tools. Additionally, we performed host
prediction with HostPhinder and taxonomic classification.
Antimicrobial resistance genes were found in the phage population of 52 out of 80 samples and
virulence factors in 18 of the samples. Potential hosts were predicted for 12.7 ± 3 % of phage
contigs. Among the most common host genera were Escherichia, Caulobacter and Bacillus.
Taxonomic classifications were assigned to 0.5% of the phage contigs on average. Among the
most common taxonomic assignments is crAssphage which was identified in 74 of the samples.
In conclusion, we found that the phage communities in sewage are extremely diverse and contain
many novel sequences.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section IV: Genomics & Proteomics 123
Comparative genomics of CEV2, a T5-family bacteriophage infecting
Escherichia coli O157:H7
María Cecilia Aristimuño Ficoseco1*, María Jose Olaya Passarell1, Elvira Hebert1, Elizabeth
Kutter2* and Raul Raya1
1CERELA (Centro de Referencia para Lactobacilos)-CONICET. Tucumán, Argentina.
2Evergreen State College Olympia, WA, USA.
*E-mail: [email protected]; [email protected]
Escherichia coli phage CEV2, a siphovirus (phages with long non-contractile tails) of the T5-like
family, possesses a 118,347 bp genome (39.3% GC) which encodes 160 ORFs and 22 tRNAs.
Phages CEV2 and T5 showed similar host ranges, but only CEV2 infects virtually all tested E. coli
O157: H7 strains. In their irreversible final step of phage binding to their hosts, both phages
recognize, through their PB5 proteins (oad gene; 99% identity), the outer cell membrane FhuA
protein as receptor. However, their long tail fiber (LTF) proteins, responsible for the reversible
interaction that occurs between the phage and the LPS (antigen O) of the bacterium, presented only
46% identity. These results could explain the difference in host range observed between these
phages. A comparative genomic study of eight T5-like phages [T5 (AY543070.1 and
NC_005859.1); CEV2; SPC35 (HQ406778.1); DT571/2 (KM979355.1); DT57C (KM979354.1);
FFH1 (KJ190157.1); AKFV33 (HQ665011.1); and APCEc03 (KR422353.1)], conducted primarily
with the programs BlastP and BlastX; Clustal Omega; Easyfig 2.1; ARAGORN and tRNAscan,
revealed a collinear organization of their genes, with identities over 80% throughout their
sequences, with phages CEV2 and FFH1 being most closely related, except in two regions: the
“LTF” and the “tRNAs” regions located, respectively, at 80-85- and 28-37-kb of their genomes.
The LFT protein of CEV2 presented a 71% identity with that of phages FFH1 and APCEc03. The
number of tRNA genes in these phages ranged from 9 to 23. CEV2 encodes 22 tRNAs, although
Gly- and Tyr-tRNA isoaccepting species were not detected. On the other hand, T5 encodes 23
tRNAs and lacks tRNAs genes for glycine (Gly) and tryptophan (Trp). The greater diversity
observed among the LFT proteins of these phages indicates their ability to adapt to different
scenarios, in particular to the different types of specific receptors located in the LPS of their host
cells. In turn, the presence of a variable number and specific tRNAs may contribute to the overall
rate of protein synthesis during phage infection.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
124 Section IV: Genomics & Proteomics
Making publicly available viral sequence data easier to use
J. Rodney Brister*, Olga Blinkova, Eneida L. Hatcher, Yuri Ostapchuck, Igor Tolstoy
National Center for Biotechnology Information, National Library of Medicine, National Institutes
of Health, Bethesda, MD 20894, USA
*E-mail: [email protected]
What’s in my tube? In the age of next generation sequencing, this is an increasingly important
question in the realm of biology. Perhaps the easiest way of answering this question is to find
similar sequences, but even when a match can be found, the effectiveness of this approach is
dependent on the availability and quality of sequences used for comparison. While GenBank and
other International Sequence Database Consortium members provide public platforms for the
storage and dissemination of data, the sheer volume of currently available data makes it
increasingly difficult to use. Moreover, the archival nature of these databases all but guarantees that
some aspects of the data will become outdated. Our group is focused on understanding the ever
expanding viral sequence space and making this data easier to use. Here I will describe the various
approaches we are implementing to build a validated layer upon viral GenBank and Short Read
Archive (SRA) sequence data. These approaches include working with other groups on taxonomy
issues, defining approaches to provide better nucleotide and protein reference space, developing
processing approaches that increase sequence data value, and building easy to use interfaces that
allow people to find the sequences they want.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section IV: Genomics & Proteomics 125
Hairy jumbo phages of Enterobacteriaceae
Colin Buttimer1, Yannick Borne2, Hanne Hendrix3, Hugo Oliveria4, Aidan Casey5, Horst Neve6,
Olivia McAuliffe5, R. Paul Ross5, Colin Hill7, Jean-Paul Noben8, Jim O’Mahony1, Lars Fieseler2,
Rob Lavigne3, Aidan Coffey1*
1Department of Biological Sciences, Cork Institute of Technology, Bishopstown, Cork, Ireland; 2ZHAW – Zürcher Hochschule für Angewandte Wissenschaften, Institut für Lebensmittel- und
Getränkeinnovation, Fachstelle Mikrobiologie, Switzerland; 3Laboratory of Gene Technology, KU
Leuven, Leuven, Belgium; 4CEB – Centre of Biological Engineering, LIBRO – Laboratório de
Investigação em Biofilmes Rosário Oliveira, University of Minho, 4710-057, Braga, Portugal; 5Teagasc Food Research Centre, Moorepark, Fermoy, Co. Cork, Ireland; 6Department of
Microbiology and Biotechnology, Max Rubner-Institut, Kiel, Germany; 6APC Microbiome
Institute and School of Microbiology, University College, Cork, Ireland; 8Biomedical Research
Institute and Transnational University Limburg, Hasselt University, Diepenbeek 3950, Belgium.
*E-mail: [email protected]
Myoviridae have the most sophisticated virion design, possessing a tail capable of contracting on
infection and generally having the largest genomes when compared to the other families. However,
only a small number of the known Myoviridae phages have genomes greater than 200 kbp; and
these are often referred to as “giant” or “jumbo” phages. These are not often isolated by
conventional methods, and this is believed to be due to their larger virion structures. As a result,
they have received less attention than the smaller members of the family. Examples of the jumbo
phages are Enterobacteria phage vB_PcaM_CBB (CBB) and Erwinia amylovora phage Y3; and
both are unusual in that they possesses atypical whisker-like structures along the length of their
contractile tails. With a genome of 355,922 bp, excluding a predicted terminal repeat of 22,456 bp,
phage CBB is the third largest phage sequenced to date, sharing homology with the Rak2-like
phages. Phage Y3 possesses a much smaller genome of 261,365, sharing homology with
Pseudomonas phages Lu11 and PaBG as well as Ralstonia phage RSL1. This work presents the
insights gained from the genomic and proteomic analysis of these atypical jumbo myoviridae
phages.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
126 Section IV: Genomics & Proteomics
Host-phage interactions between novel jumbo phage PA5oct and
Pseudomonas aeruginosa
Katarzyna Danis-Wlodarczyk1*, Ho Bin Jang1, Bob Blasdel1, Yves Briers2, Jean-Paul Noben3,
Zuzanna Drulis-Kawa4, Rob Lavigne1
1 Laboratory of Gene Technology, KU Leuven, Kasteelpark Arenberg 21, 3001 Leuven, Belgium 2 Laboratory of Applied Biotechnology, Ghent University, Valentin Vaerwyckweg 1, 9000 Gent,
Belgium 3 Biomedical Research Institute and Transnational University Limburg, Hasselt University, 3950
Diepenbeek, Belgium 4 Department of Pathogen Biology and Immunology, Institute of Genetics and Microbiology
Institute of Genetics and Microbiology, University of Wroclaw, Przybyszewskiego 63/77,
51-148 Wroclaw, Poland
*E-mail: [email protected]
The PA5oct, a novel jumbo myovirus, has a giant linear A+T-rich double-stranded DNA genome
(287,182 bp), making it the third largest sequenced Pseudomonas phage genome. Based on DNA &
RNA sequencing, 462 genes could be confirmed, of which only a small number could be
functionally assigned. The number of genes was further supported by structural mass spectrometry
analysis. The genetic relationships between PA5oct and other jumbo bacteriophages and other
Caudovirales viruses, was constructed using a protein-sharing network. The PA5oct, Escherichia
phages 121Q/PBECO4, Klebsiella phage vB_KleM-RaK2, Klebsiella phage K64-1, and
Cronobacter phage vB_CsaM_GAP32 presented closer relationships based on shared conserved
core genes. Furthermore, PA5oct, PBECO4, GAP32, T5, FelixO1 appear to be distantly diverged
members of Tevenvirinae.
To examine phage transcription across the temporal phases of infection cycle at single-nucleotide
resolution as well as the phage- host interactions, RNA-seq was performed based on three selected
infection time points (5, 15 and 25 min). Reads originating from the phage and the host at each
stage of infection were mapped to the phage and host genomes respectively, revealing that PA5oct
progressively dominates host transcription. Indeed, PA5oct transcripts represent 21% of total non-
rRNA transcripts within 5 minutes and eventually proceeds to 69% then 92% by middle and late
infection, respectively. The PAO1 host-mediated transcriptional response to PA5oct infection is
similar to 14-1, PEV2, YuA, and LUZ19 infection response. Interestingly, PA5oct progressively
dominates the mRNA and sRNA environment of the cell with strong transcription, as well as the
expression of an RNase-H-like protein putatively involved in degrading host transcripts. This shift
from host transcripts to phage transcripts reduces all host transcripts relative to the total in the cell.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section IV: Genomics & Proteomics 127
Comparative genomics of rhizobiophages from western Canada
K.M. Damitha Gunathilake1*, Anupama P. Halmillawewa1, Marcela Restrepo-Córdoba1,
Benjamin J. Perry2, Christopher K. Yost2, Michael F. Hynes1
1University of Calgary, Calgary AB, Canada, 2University of Regina, Regina, SK Canada,
*E-mail: [email protected]
Bacteriophages that infect the rhizobia, bacteria that form nitrogen-fixing nodules on the roots of
legumes, have been studied for decades, and have been suggested as a means of manipulating
rhizosphere rhizobial populations. Genomic studies on rhizobiophages are relatively recent, and
only a few complete genome sequences are available in the databases. We have isolated a large
collection of phages that infect Rhizobium leguminosarum, Mesorhizobium loti, and R. gallicum
from rhizosphere soils from a variety of legumes in western Canada, and characterized them by
examining host range, lysogeny, morphology, and genome composition. Complete genome
sequences have been produced for six of them: vB_RleS_L338C genome: accession no:
KF614509, vB_RglS_P106B (KF977490), vB_RleM_PPF1 (KJ746502), vB_RleM_P10VF
(KM199770) and vB_MloP_Lo5R7ANS (KM199771). The completed genome of M. loti phage
Cp1R7ANS-C2 is also available but has not been submitted to a public database. Draft genomes
for a further five R. leguminosarum phages P9VFCI, V1VFA, AF3, P11VFA, L338G and M. loti
phage Cp1R7A-A1 have also been produced. Phages isolated using R. leguminosarum as host all
belonged to the Myoviridae or the Siphoviridae, whereas phages that infected M. loti were mostly
members of the Podoviridae, with one example of a probable Siphovirus. The majority of the
phages had genomes that were modified and resistant to restriction digestion with many common
enzymes. On the phages with larger genomes, such as P10VF, L338G, AF3, and P9VFC1, many
genes encoded predicted proteins with no significant homology to anything in the databases. We
have performed genome comparisons between our isolates, and comparisons and phylogenetic
analysis in the context of all rhizobiophages sequenced so far. Rhizobium phage P10VF shows no
significant similarities to 16 other whole genome sequences of Rhizobium, Mesorhizobium and
Sinorhizobium phages available in the NCBI database. However, it shows significant similarities
and phylogenetic relationship to Sinorhizobium meliloti Phage ΦM9 for which the complete
genome is now available. Furthermore, the draft genomes of Rhizobium phages P9VFCI and AF3
show similarities to P10VF, although both have genomes larger than P10VF. Another Rhizobium
phage, V1VFA, has a part of its genome identical to P10VF. In spite of being isolated from
different geographic locations, from soils planted to different legumes, and being isolated using
different strains, all three of phages P9VFCI, AF3 and V1VFA are related to P10VF as indicated
by the phylogenetic analysis with amino acid sequences of putative terminases and capsid proteins.
Siphophage P11VFA shows significant similarity to L338C, with identical terminase and capsid
proteins, but also contains large sections of DNA similar to a Sinorhizobium plasmid.
Mesorhizobium phages Cp1R7AS-C2 and Lo5R7ANS (Podoviridae) have similar genome
arrangements and sizes, and share some properties with T7-like phages. Mesorhizobium phage
Cp1R7A-A1 has a much larger genome with no strong similarities to anything so far sequenced.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
128 Section IV: Genomics & Proteomics
Lytic bacteriophages present in the lower female genital tract targeting E. coli
Anna-Ursula Happel1, Shameem Z. Jaumdally1,2, Hoyam Gamieldien1, Heather B. Jaspan1,3,
Smritee Dabee,1 Shaun Barnabas,1,4 Linda-Gail Bekker MD PhD,1,4 Jo-Ann S. Passmore1,2,5, Remy
Froissart6*
1Institute of Infectious Disease and Molecular Medicine (IDM), University of Cape Town, South
Africa; 2DST-NRF Centre of Excellence in HIV Prevention, 3Seattle Children’s Research Institute,
University of Washington, Seattle, Washington, USA, 4Desmond Tutu HIV Centre, University of
Cape Town, South Africa, 5National Health Laboratory Service, Groote Schuur Hospital,
Observatory, Cape Town, South Africa, 6UMR 5290 MIVEGEC, CNRS-IRD-University of
Montpellier, Montpellier, France Desmond Tutu HIV Centre, University of Cape Town, South
Africa
*E-mail: [email protected]
Background: As in other human mucosal niches, bacteriophages are present in the lower female
genital tract (FGT). Little is known about the origin and transmission of bacteriophages in the FGT,
their bacterial targets and their effect on commensal bacterial communities in this niche over time.
Understanding the pathogenic interaction between bacteriophages and their specific bacterial hosts
in important since they may be a cause of dysbiosis in the FGT, typified by commonly occurring
bacterial vaginosis (BV). Several lines of evidence suggest the presence of bacteriophages in the
lower FGT viral sequences related to Caudovirales have been found in the lower FGT, and
temperate bacteriophages have been demonstrated within commensal or anaerobic pathogenic
bacteria from the FGT; in addition to these genital bacterial isolates having CRISPR loci and Cas-
genes. To our knowledge, no lytic phages have ever been isolated from human FGT and
characterized phenotypically. The aim of this study was to screen for and isolate lytic
bacteriophages from the lower FGT and test experimentally how these are involved in the etiology
of vaginal dysbiosis within human FGTs.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section IV: Genomics & Proteomics 129
Isolation and Analysis of Cluster Q Phage Ein37 and F1 Phage Plumbus
Daniel Devonshire, Sophie Harrison*, Cody Allison, Samantha Altheide, Brianna Bordwell,
Mandy Chen, Adrienne Chitwood, Tokala Christensen, Carollyne Coulson, Nichole Criss, Melinda
Dauley, Christopher Davis, Kyle Dobbs, Jacob Durham, Michael Espy, Jarred Figlar-Barnes, Parke
Funderburk, Evan Furrer, Brianna Gage, Daniel Gilberts, Andrew Glassbrook, Kyle Grosten, Jenny
Green, Annelise Hooper, Casey Irwin, Coleman Kaestner, Katherine Kallus, Nolan Knaus, Tamara
Koledin, Randolph Kraus, Stacia Krause Ledbetter, Tessa LaLonde, Madesyn Lockwood, Anthony
Lopez, Kody Ludwig, Nacia Magass, Trevor McCliment, Bryanne McNamara, Robert Munoz,
Micah Needham, Melyssa Nocis, Taylor Nowotny, Luke Ogden, David Parker, Habib Quaraishi,
Olivia Parks, Kaden Phillips, Rian Plastow, Justin Roberts, Shayna Rossiter, Breck Roundtree,
Candace Saunders, Marissa Scoville, Alexea Seil, Petal Shohadai, Keelan Smith, Sean Staub,
Charles Thompson, Steven Vananzo, Amber Vidal, Jenny von Henkelman, Crystal Whitford,
Chandler Williams, Clarissa Dirks, Alberto Napuli, James Neitzel
*E-mail: [email protected] (The Evergreen State College)
The student participants in our third cycle of the SEA-PHAGE project were again members of the
interdisciplinary program Introduction to Natural Sciences, a year-long, full-time learning
community with integrated instruction in biology, chemistry, and science process skills. During the
fall quarter students collected and purified phages using enrichment cultures of Mycobacterium
smegmatis mc² 155 at 37 C. This year 23 phages were isolated from local soils, purified, and
entered into the PhagesDB collection. Compared to prior years, we had a lower percentage of
successful isolations. This set of phages had their DNA purified and analyzed by restriction
enzyme digestion and gel electrophoresis. Successful DNA extractions were completed for most of
the phages, although some difficulty in dealing with DNA stability was encountered. DNA quality
and restriction enzyme experiments were used to select genomes for sequence analysis. Phages
were also analyzed by transmission electron microscopy after negative staining with uranyl acetate.
This resulted in clear images of all of the isolated phages, which appeared to be Siphoviridae. The
phages sequenced and analyzed were Ein37 and Plumbus. DNA from these phages was sequenced
using the Illumina process at the Pittsburg Bacteriophage Institute. The sequence of Ein37 revealed
a 53,748 bp linear double stranded DNA genome with a sticky fourteen bp 3’ overhang and with a
GC content of 67.4 %. Analysis of the sequence of this phage confirmed that it was a Siphoviridae
in the Q cluster. BLASTn results showed that Ein37 had a 99% sequence identity with the other 7
members of cluster Q, indicating a high degree of similarity in this group, with almost half of the
predicted genes being Phams unique to this cluster.The genome of Plumbus is a 54,468 bp dsDNA
with a sticky ten bp 3’ overhang and a GC content of 61.1%. BLASTn results indicated its closest
relative was Cluster F1 phage Kimberlium. Both genomes were analyzed for potential protein
coding open‐reading frames using Glimmer and GeneMark, and protein functions were predicted
by BLASTp and HHPred, as well as examining synteny with related phages. Preliminary results
suggest the presence of 86 protein coding genes in Ein39 and 106 in Plumbus. We identified no
tRNA or tmRNA genes in either phage. Further work is being conducted to identify and confirm all
protein coding regions and to identify functions for predicted protein products.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
130 Section IV: Genomics & Proteomics
Klebsiella podovirus KP32 produces two polysaccharide depolymerases,
promising anti-virulence tools
Agnieszka Latka1,2, Grazyna Majkowska-Skrobek1, Flavia Squeglia3, Rita Berisio3, Yves Briers2
and Zuzanna Drulis-Kawa1*
1Department of Pathogen Biology and Immunology, Institute of Genetics and Microbiology,
University of Wroclaw, Przybyszewskiego 63/77, 51-148 Wroclaw, Poland 2Laboratory of Applied Biotechnology, Department of Applied Biosciences, Ghent University,
Valentin Vaerwyckweg 1, 9000 Gent, Belgium 3Institute of Biostructures and Bioimaging, National Research Council, Via Mezzocannone 16,
I-80134 Naples, Italy
*E-mail: [email protected]
Bacteriophages (phages) as natural enemies of bacteria have a huge ecological impact on the
dynamics of bacterial populations. To overcome the capsule barrier, some phages are equipped
with a virion-associated depolymerase – highly specific enzymes depolymerizing capsular
polysaccharides (CPS), thus also responsible for host cell recognition and receptor binding.
Bacteriophages possessing proteins with such activity are also able to better diffuse in the biofilm,
gaining access to the present microcolonies, and infecting embedded bacteria. Depolymerization of
CPS results in the increasing susceptibility to chemical and physical agents as well as to host
defences such as phagocytosis or complement-mediated killing. Although depolymerases do not
kill the bacterium, they have special interest as anti-virulence compounds that disarm the
bacterium, reducing or even avoiding the infection process.
In the presented study two genes encoding putative depolymerases have been identified in the
genome of K. pneumoniae specific KP32 podovirus. According to in silico prediction (PHYRE2),
the β-helical structure is highly probable for both proteins. Selected gene products have been
produced as recombinant proteins. Zymographic analysis using the crude exopolysaccharides
extracted from bacterial strains, confirmed their enzymatic activity. Both depolymerases, one of
which has been annotated as a tail fiber with similarity to T7 tail fiber region, while the other as
hypothetical protein, recognize and cleave CPS of different Klebsiella serotypes. They possess
mainly β-conformation, according to circular dichroism (CD) spectroscopy, what explains their
stability at high temperature and wide range of pH. The oligomeric state of all depolymerases is
trimeric, according to multi-angle static light scattering (MALS) spectroscopy. Microbiological
results showed that these depolymerases by modifying the phenotype of the bacterial cells without
affecting their viability, make the bacteria easier to eradicate by immune response mechanisms.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section IV: Genomics & Proteomics 131
Characterization of Temperature Sensitive Mutants of Bacteriophage K
Justin Leavitt* and Jason Gill
Department of Animal Science, Texas A&M University, College Station, TX 77843, USA
Center for Phage Technology, Texas A&M University, College Station, TX 77843, USA.
*E-mail: [email protected]
Bacteriophage K is a Twort-like virus that infects many clinically important strains of
Staphylococcus aureus. Many of the genes in the phage K genome have unknown functions, and
many details about the phage K lifecycle require further study. Because Bacteriophage K has
therapeutic potential in treating staphylococcal infections, there is added incentive to learn more
about its biology. Using a classical genetic screen, a large number of temperature sensitive mutants
were isolated. These mutants were sequenced and further characterized by their ability to lyse the
host cell, make complete phage particles, and infect other host cells at the non-permissive
temperature. The phenotypes of these mutants and corresponding genetic changes will be
discussed.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
132 Section IV: Genomics & Proteomics
Characterization of Bdellovibrio bacteriovorus phage
Kema Malki1*, Mark Martin2, Bentley Fane3, Mya Breitbart1
1College of Marine Sciences, University of South Florida, Saint Petersburg, Florida, USA
2Biology Department, University of Puget Sound, Tacoma, Washington, USA 3Department of Immunobiology, School of Plant Sciences, University of Arizona, Tucson, Arizona,
USA
*E-mail: [email protected]
Bdellovibrio are small, parasitic bacteria that are best known for their ability to attack and lyse
gram-negative bacterial cells. These δ-Proteobacteria were originally isolated from soil in the
1960s during a search for bacteriophage (phage). Since then, different strains of Bdellovibrio with
varying host ranges have been discovered from diverse environments including freshwater,
seawater, soil, and sewage. Though originally thought to be obligate parasites, strains of host-
independent Bdellovibrio have also been isolated and cultured in laboratory settings. Due to their
capability to lyse bacteria, Bdellovibrio have important ecological implications and have even been
proposed as potential antimicrobial agents for human health applications.
The purpose of this study is to expand our knowledge of phage capable of infecting Bdellovibrio
bacteriovorus. Several studies in the early 1970s isolated Bdellovibrio phage and described their
morphology and infection parameters. However, to date, genomic information is only available for
two Bdellovibrio phage, the small ssDNA phage ΦMH2K and the dwarf myovirus Φ1402, which is
the smallest autonomous myovirus. These phage genomes suggest that Bdellovibrio phage may
represent a large reservoir of unexplored diversity, particularly regarding phage with reduced
genome sizes. In this study, phage capable of creating plaques on the host-independent,
Bdellovibrio bacteriovorus strain HI 109 were isolated from sewage samples, plaque purified, and
are currently being characterized based on their morphology and genome sequences. The
availability of well-characterized Bdellovibrio phage systems in the lab represents a critical step
towards future work examining the net ecological impact of Bdellovibrio phage on environmental
bacterial communities.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section IV: Genomics & Proteomics 133
Genomic Changes in a Host Range Mutant of Staphylococcus aureus
Bacteriophage Sb-1
K.V. Sergueev1, A.A. Filippov1, J. Farlow1, A.D. Reddy1, L. Kvachadze2, N. Balarjishvili2, M.
Kutateladze2, M.P. Nikolich1*
1 Walter Reed Army Institute of Research, Silver Spring, MD, USA; 2 G. Eliava Institute of
Bacteriophages, Microbiology and Virology, Tbilisi, Georgia.
*E-mail: [email protected]
Infections with methicillin-resistant Staphylococcus aureus (MRSA) is a major and growing
multidrug resistance problem in global medicine. Bacteriophage (phage) therapy is one of
promising approaches for treatment of MRSA infections in concert with antibiotics. S. aureus-
specific bacteriophage Sb-1 was isolated in the Georgian Republic (USSR) in 1977 and has been
widely used for treatment of various human S. aureus infections. Sb-1 has a very broad host range
within S. aureus that includes MRSA strains, and its host range can be further expanded. We
tested the activity of Sb-1 phage manufactured at the Eliava Institute against a panel of 25 diverse
US Military MRSA isolates and showed that it was able to lyse 23 of them. A host range mutant of
Sb-1 designated Sb-1M was then isolated to address the two additional strains in the military
MRSA panel, and then it was tested in comparison with the parental phage. Sb-1 was active
against 79 out of 94 (84%) diverse global S. aureus isolates, while eight additional strains (87/94 =
92.6%) were susceptible to Sb-1M. In an effort to better understand the genetic basis for this host
range expansion, we sequenced the complete genomes of Sb-1 and Sb-1M. Comparative genomic
analysis revealed a complex hypervariable repeat structure in the Sb-1 genome in which a distinct
allele correlated with the expansion of host range. This hypervariable region appears to be a novel
host range determinant that had not previously been characterized in Twort-like phages.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
134 Section IV: Genomics & Proteomics
Comparative genomics of fully sequenced Cp8viruses, with reference to the
Irish Campylobacter phage, vB_CjeM_Los1
L. O’Sullivan1,3, H. Neve2, D. Bolton3, O. McAuliffe4, and A. Coffey1
1 Department of Biological Sciences, Cork Institute of Technology, Co. Cork, Ireland; 2 Max
Rubner-Institute, Federal Research Institute of Nutrition, Hermann-Weigmann-Strasse 1, Kiel,
Germany; 3 Food Research Centre, Teagasc, Ashtown, Co. Dublin, Ireland; 4 Biotechnology
Department, Teagasc, Moorepark Food Research Centre, Fermoy, Co. Cork, Ireland.
*E-mail: [email protected]
Campylobacter jejuni is a commensal of up to 83.1% of Irish broiler chickens
and it has the ability
to cause severe gastrointestinal infection in humans. Phages against C. jejuni have been previously
isolated and proven effective in reducing C. jejuni numbers in live poultry and contaminated
carcass skin. The lytic C. jejuni Myovirus vB_CjeM_Los1 (hereafter Los1) was isolated from a
poultry faecal sample in a Cork slaughterhouse and it has proven the ability to lyse C. jejuni strains
isolated from both poultry and clinical origin. Well-studies Campylobacter phages are
representative of two genera, Cp220viruses and Cp8viruses as proposed by the ICTV. To date, 8
Campylobacter phages have been sequenced all with similarity to Cp8 (regarding size and whole-
genome sequence), including Los1 and this poster details some of the comparative analyses
performed using the eight genomes, whose phages have a diverse range of origins.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section IV: Genomics & Proteomics 135
Broad host-range T5-like Yersinia enterocolitica phage R2-01
Maria I. Pajunen1*, Lotta J. Happonen2, Jin Woo Jun1, Ayesha Nawaz1, Laura Mattinen1 and
Mikael Skurnik1*
1Department of Bacteriology and Immunology, Medicum, Research Programs Unit,
Immunobiology, University of Helsinki, Helsinki, Finland. 2Lund University, Department of Clinical Sciences Lund, Infection Medicine, Lund, Sweden.
*E-mail: [email protected], [email protected]
Here we report the complete genome sequence and morphological characterization of the T5-like
siphovirus vB_YenP_R2-01 (in short R2-01) infecting Yersinia enterocolitica, a zoonotic food-
borne pathogen causing yersiniosis in humans and animals. Bacteriophage R2-01 encodes for 152
open reading frames (ORFs) and 19 tRNA-molecules on its 122,696 bp-long double-stranded DNA
genome including 9,901 bp terminal repeats. A total of 115 of the ORFs are similar to genes of the
well-characterized bacteriophage T5, including all genes involved in phage morphogenesis. The
genome of R2-01 is mostly syntenic to that of T5, and the major differences between the genomes
reside in areas with genes encoding for hypothetical R2-01 proteins. Other phages similar to R2-
01 are the Salmonella enterica serovar Typhimurium phage Stitch, the Escherichia phage EPS7,
the Salmonella phage Shivani, the Escherichia phage vB_EcoS_FFH1, the Enterobacteria phages
DT57C and DT571/2, the Escherichia phage AKFV33, and the Salmonella phage SPC35. The host
range of the phage is broad among Y. enterocolitica and it infects strains of both pathogenic and
non-pathogenic serotypes. It does not infect strains of Y. pseudotuberculosis, Y. nurmii, Y.
pekkanenii, Y. mollaretii, Y. frederiksenii, Y. intermedia, Y. ruckeri, Y. bercovieri, Y. kristenseni
(except one strain), and Y. aleksiciae. The phage also failed to infect Escherichia coli and Shigella
strains. Isolation of phage-resistant mutants from a transposon-insertion library revealed that the
phage receptor is the BtuB protein.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
136 Section IV: Genomics & Proteomics
Development of a random mutagenesis protocol for marine cyanophage
Tamsin Redgwell1*, Andrew Millard 2, Dave Scanlan1
1 School of Life Sciences, University of Warwick, Coventry, CV4 7AL, UK. 2 Warwick Medical School, University of Warwick, Coventry, CV4 7AL, UK.
*E-mail: [email protected] Objectives:
Viruses are the most abundant biological entities on the planet, with viruses specifically infecting
cyanobacteria termed cyanophage. Marine cyanobacteria of the genera Prochlorococcus and
Synechococcus are key oceanic primary producers, and through their lysis following cyanophage
infection large amounts of dissolved organic matter are released into the environment directly
affecting marine carbon cycling. The discovery of auxiliary metabolic genes (AMGs) in
cyanophage genomes has altered our understanding of the role viruses play in marine ecosystems.
These AMGs are widespread in nature and whilst their specific roles are poorly understood it is
thought they play a key role subverting host metabolism to increase infection success. The
development of a random mutagenesis protocol for cyanophage would allow us to assign function
to these phage AMGs, and aid in the discovery of more. Through development of a chemical
mutagenesis system I hope to develop a protocol that not only is applicable to marine cyanophage
but can also be applied to freshwater cyanophage systems.
Methods:
A 24-hour period of hydroxylamine mutagenesis was conducted on both a coliphage (control) and
cyanophage S-PM2d, to create single nucleotide polymorphisms (SNPs). Time points were taken
every 6 hrs and dialysed to remove the mutagen. The time point at which there was a 3-log drop in
phage titre (99.9% killing) was used to identify the length of incubation required to create 1 SNP
per genome. To confirm this 15 individual plaques per time point were picked, and DNA extracted
and sequenced on an Illumina MiSeq using the Nextera XT library prep kit, for each phage. Reads
were trimmed with Sickle, assembled with SPAdes, and SNPs were called using VarScan.
Results:
In the coliphage control 99.9% killing did correspond to 1 SNP per genome. However, this was not
the case for cyanophage S-PM2d, where the number of SNPs per genome does not appear to be
related to the length of incubation with mutagen. Here, the number of SNPs is initially and
consistently higher than in the control. The types of mutation caused are also not limited to the
transition changes known to be generally attributed to hydroxylamine mutagenesis.
Conclusions:
It is evident that cyanophage possess auxiliary metabolic genes that likely affect the behaviour of
the host during infection. Elucidation of the function of these genes has been limited by the lack of
a cyanophage genetic system. Initial results show that hydroxylamine is capable of creating SNPs
in cyanophage genomes, but at a rate much higher than in a coliphage control. Further work will
aim to determine whether this behaviour is specific to this particular cyanophage, as well as
characterise mutants of interest.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section IV: Genomics & Proteomics 137
Digging in the dirt for diversity - Genomic and molecular analysis of a
diverse set of E.coli phages
Imke Schmidt1, Christine Rohde1, Boyke Bunk1, Cathrin Spröer1, Manfred Rohde2 and Johannes
Wittmann1*
1Leibniz Institute DSMZ - German Collection of Microorganisms and Cell Cultures,
Braunschweig, Germany; 2Helmholtz Centre for Infection Research, Braunschweig, Germany
*E-mail: [email protected]
Escherichia coli is among the most common pathogens in poultry and the causative agent of
colibacillosis. This frequent disease creates significant economic losses and is commonly treated by
antibiotics, which in turn promotes the selection of multidrug resistant (MDR) bacteria. A high
incidence of MDR bacteria is problematic not only for animal health but also because of the
potential for zoonotic transfer to humans through contaminated food. Therefore, there is a high
demand to develop new strategies as alternatives to antibiotic therapies. In accordance with the
One-Health concept this project aims to isolate and characterize phages to fight E. coli infections in
broilers.
Apart from generating a suitable phage mixture for a field trial with broilers, one further aim of this
study is to get further insight into the diversity of E. coli phages followed by enhanced work on
taxonomic issues in that field.
Thus far, we isolated phages from sewage (21), surface water (4), manure (34), poultry (18)
respectively horse dung (1), and hospital wastewater (5). Morphological characterization revealed a
diversity of morphotypes (77% Myo-, 17% Sipho-, and 6% Podoviridae), genome sequencing
resulted in genomes sizes from 44 up to 370 kb. Annotation and comparison with databases
showed similarities in particular to T4- and T5-like phages. Host range analyses were performed by
spot tests on 72 E. coli strains with different resistance patterns.
We identified E28 as suitable strain for in vivo experiments, 11 of the isolated phages infect E28
with varying efficiency. In order to avoid the development of phage resistance we composed a
mixture of virulent phages that differ in host coverage, genotype, growth parameters, and inhibit
the growth of E28 in vitro.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
138 Section V: Structure and Molecular Mechanisms
Investigating immunity mechanisms of an antiphage island in Vibrio cholera
Zachary Barth and Kimberley Seed*
University of California, Berkeley
*E-mail: [email protected]
The lytic myovirus ICP1 is engaged in an evolutionary arms race with its host, pandemic Vibrio
cholerae. V. cholerae strains possessing a phage inducible chromosomal island-like element (PLE),
are able to resist infection by ICP1. The V. cholerae PLE shares several functional similarities with
phage inducible chromosomal islands, a class of satellite phages found in Gram positive bacteria.
These functional similarities include excision of the island from the host chromosome upon phage
infection, followed by replication of the island, as well as mobilization and integration into other V.
cholerae cells. While the PLE does not prevent death of infected cells, it is able to completely
block production of ICP1 from infected cells, protecting V. cholerae on a population level. A
precise mechanism through which the PLE blocks ICP1 production remains unknown. Previously,
the PLE was found to decrease ICP1 DNA replication during infection, and to alter cell lysis
kinetics. Here, we further investigated the PLE’s impact on processes necessary for ICP1
replication in order to understand the mechanism(s) of PLE conferred immunity. We investigated
PLE effects on ICP1 gene expression, and tested for potential links between PLE replication and
impairment of ICP1 replication. Our efforts to elucidate the mechanisms underpinning PLE
mediated immunity will provide an improved understanding of phage-host interactions.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section V: Structure and Molecular Mechanisms 139
Spacer capture and integration by a type I-F Cas1:Cas2-3 CRISPR adaptation
complex
Robert D. Fagerlund1, Max E. Wilkinson1,2, Oleg Klykov3, Arjan Barendregt3, F. Grant Pearce4,
Sebastian N. Kieper1, Howard W.R. Maxwell1, Angela Capolupo3, Albert J.R. Heck3, Kurt L.
Krause2, 1,5, Richard A. Scheltema3, Raymond H.J. Staals1 and Peter C. Fineran1*
1Department of Microbiology and Immunology and 2Department of Biochemistry, University of
Otago, PO Box 56, Dunedin 9054, NZ; 3Biomolecular Mass Spectrometry and Proteomics, Bijvoet
Center for Biomolecular Research and Utrecht Institute of Pharmaceutical Sciences, and
Netherlands Proteomics Center, Utrecht University, 3584 CH Utrecht, NL; 4Biomolecular
Interactions Centre and School of Biological Sciences, University of Canterbury, Private Bag 4800,
Christchurch 8020, NZ; 5Otago Centre for Electron Microscopy, University of Otago, PO Box 56,
Dunedin 9054, NZ
*E-mail: [email protected]
CRISPR-Cas adaptive immune systems capture DNA fragments from invading bacteriophages and
plasmids and integrate them as spacers into CRISPR arrays. In most CRISPR-Cas systems,
adaptation is driven by Cas1:Cas2 complexes. Type I-F systems however, contain a unique fusion
of Cas2 with the type I effector helicase and nuclease for invader destruction, Cas3. Here, we
present a structural model of the 400 kDa Cas14:Cas2-32 complex from Pectobacterium
atrosepticum bound to protospacer substrate DNA. The two Cas3 domains flank Cas2 of the
Cas1:Cas2 core and form a groove where the protospacer binds. We developed a sensitive in vitro
integration assay and demonstrated that Cas1:Cas2-3 catalyzed spacer integration into CRISPR
arrays via the integrase domain of Cas1 and this was independent of Cas3 activities. Integration
required at least partially duplex protospacers with free 3’-OH groups and leader-proximal
integration was stimulated by integration host factor (IHF). In a coupled capture and integration
assay, Cas1:Cas2-3 processed and integrated protospacers independently of Cas3 activity. These
results provide insight into the structure of protospacer-bound type I Cas1:Cas2-3 adaptation
complexes and their integration mechanism.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
140 Section V: Structure and Molecular Mechanisms
Imprecision during spacer acquisition by type I CRISPR-Cas systems
increases CRISPR diversity in bacterial populations
Simon A. Jackson1, Nils Birkholz1, Corinda Taylor1 and Peter C. Fineran1, 2
1Department of Microbiology and Immunology, University of Otago, PO Box 56, Dunedin 9054,
New Zealand. 2Bio-Protection Research Centre, University of Otago, PO Box 56, Dunedin 9054,
New Zealand.
*E-mail: [email protected]
Type I CRISPR-Cas systems use protospacer adjacent motifs (PAMs) to discriminate between
target and non-target sequences. PAMs are also cues for the Cas1-Cas2 complex that trims pre-
spacer substrates prior to their integration into CRISPR arrays. The Cas1 protein possesses a PAM-
sensing domain and cleaves pre-spacer substrates adjacent to PAMs (or within PAMs for type I-E).
This ensures crRNAs produced from spacers are compatible with the PAM-sensing domains of
interference complexes. The Pectobacterium atrosepticum type I-F CRISPR-Cas system has a
PAM-proximal spacer acquisition fidelity of greater than 93% in vivo. Most of the remaining
spacers result from cleavage within one nucleotide of the canonical position. This imprecision is
termed ‘slipping’. Here, we show that slipping typically results in spacers that are not functional for
interference. Instead, slipped spacers strongly stimulate primed CRISPR adaptation (priming),
resulting in increased CRISPR diversity within bacterial populations. We propose that slipping-
induced priming pre-empts escape mutations in phage and mobile genetic elements. Thus, the
diversity-generating characteristics of slipping might account for why higher fidelity Cas1-Cas2
activity has not arisen.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section V: Structure and Molecular Mechanisms 141
Quorum sensing controls adaptive immunity through multiple CRISPR-Cas
systems
Adrian G. Patterson1, Simon A. Jackson1, Corinda Taylor1, Gary B. Evans2, George P.C. Salmond3,
Rita Przybilski1, Raymond, H.J. Staals1 and Peter C. Fineran1,4
1Department of Microbiology and Immunology, University of Otago, PO Box 56, Dunedin 9054,
New Zealand; 2Ferrier Research Institute, Victoria University of Wellington, 69 Gracefield Rd,
Lower Hutt 5010, New Zealand; 3Department of Biochemistry, University of Cambridge, Tennis
Court Road, Cambridge CB2 1QW, UK; 4Bio-Protection Research Centre, University of Otago, PO
Box 56, Dunedin 9054, New Zealand.
E-mail: [email protected]
Bacteria commonly exist in populations at high cell density, making them prone to viral predation
and horizontal gene transfer (HGT). To combat these invading elements, bacteria possess defence
strategies, such as the CRISPR-Cas adaptive immunity. These systems capture short sequences of
genetic material from invading nucleic acids (termed ‘spacers’) which are incorporated into
genomic CRISPR arrays, forming a genetic ‘memory bank’ of previous exposure. When expressed,
these provide RNA-guides for Cas protein complexes to identify specific cognate sequences and
mediate their destruction. Since their discovery, a plethora of research has been directed at
understanding CRISPR-Cas function, while comparatively little is known about their regulation
within natural hosts. Many bacterial populations coordinate their behaviour as cell density
increases, using a signalling mechanism called quorum sensing (QS). Here we show that QS
upregulates CRISPR-Cas activity to protect bacterial populations when they would be most
vulnerable to HGT and the spread of viruses. Strains unable to communicate were less effective at
defending against invading elements targeted by the CRISPR-Cas systems. Additionally, the
acquisition of new spacer sequences, required for subsequent identification of foreign targets, was
impaired in the absence of QS signalling. These results demonstrate that bacterial populations use
QS-mediated communication to regulate CRISPR-Cas activity, thereby modulating the efficacy of
community level defence.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
142 Section V: Structure and Molecular Mechanisms
Characterization of bacteriophage CPS1 infecting
Clostridium perfringens
Eunsu Ha1 and Sangryeol Ryu1,2
1Department of Food and Animal Biotechnology, Department of Agricultural Biotechnology, and
Research Institute of Agriculture and Life Sciences, Seoul National University, Seoul 08826, Korea 2Center for Food and Bioconvergence, Seoul National University, Seoul 08826, Korea
Objectives: Clostridium perfringens is a Gram-positive, anaerobic, and spore forming bacterium
widely distributed in the environment. It is one of the most common causes of foodborne illnesses.
Bacteriophages are regarded as one of the alternatives to control pathogenic bacteria without
causing antibiotics resistance problem. However, the number of reported C. perfringens phages are
small and nothing much is known about their receptors. In this study, we isolated C. perfringens
virulent bacteriophage CPS1 and identified its receptor.
Methods: CPS1 was isolated from animal feces. CPS1 morphology was examined by transmission
electron microscopy. Phylogenetic tree was constructed with MEGA7 by the neighbor-joining
method. C. perfringens ATCC 13124 random transposon mutant library was constructed using Ez-
Tn5 (Epicentre) and screened for CPS1 resistant mutant. The gene disrupted by Tn5 insertion was
cloned in pET28a plasmid and expressed in E. coli BL21 (DE3). The protein was purified by Ni-
NTA chromatography and the histidine tag of the protein was removed by thrombin kit (Novagen).
Glucose oxidation assay and Morgan-Elson reaction were done with the purified protein to
elucidate the activity of the protein. Cell wall associated polysaccharides were quantified by
phenol-sulfuric acid analysis. Capsule staining was done with Anthony’s method.
Results: Analysis of full genome and morphology of CPS1 showed that CPS1 belongs to
Picovirinae subfamily. Screening of Ez-Tn5 random transposon mutant library of C. perfringens
ATCC 13124 revealed that C. perfringens ATCC 13124 CPF_0486::Tn5 had CPS1 resistance. The
gene was initially annotated as UDP-glucose 4-epimerase (galE). However, glucose oxidase assay
and Morgan-Elson reaction showed that CPF_0486 encoded protein had both UDP-glucose 4-
epimerase and UDP-N-acetyl glucosamine 4-epimerase activities, suggesting that the gene is gne.
According to the previous reports (Bernatchez et al., 2005; Valiente et al., 2008), gne but not galE
is required for synthesis of capsular polysaccharides (CPs). These results indicate that CPs are
possible receptors of the phage. Comparison of the amount of CPs produced in the wild type C.
perfringens and the mutant by phenol-sulfuric acid analysis revealed that the amount of CPs was
lower in the mutant. Capsule staining also showed that the wild type produced larger amount of
CPs than the mutant. These results support that CPS1 phage receptor is CPs.
Conclusions: C. perfringens bacteriophage CPS1 was isolated from environmental samples.
Analysis of morphology and genome of CPS1 revealed that CPS1 belonged to Picovirinae
subfamily. The CPS1-resistant mutant of C. perfringens was found to have mutation in the
CPF_0486 gene associated with biosynthesis of CPs, suggesting that CPs are the receptor of CPS1
phage.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section V: Structure and Molecular Mechanisms 143
Characterization of a potential toxin/anti-toxin system within Vibrio
cholerae’s anti-phage immunity island
Kristen N LeGault*, Kimberley D Seed
University of California, Berkeley, Berkeley, CA
*E-mail: [email protected]
Bacteriophage predation has driven the evolution of several anti-phage immunity mechanisms in
their bacterial hosts. One such system is the phage-inducible chromosomal-like element (PLE) of
Vibrio cholerae, recently shown to block infection by specific lytic phage (O’Hara et al. 2017).
The mechanism of phage-immunity conferred by the PLE is not yet known, as the PLE
encompasses a number of open reading frames (ORFs) of unknown function. Two of these ORFs
possess features of a toxin/anti-toxin system. Here we show that overexpression of the putative
toxin causes cell filamentation and a loss of viability, while co-expression of the putative anti-toxin
restores normal growth. However, bacterial adenylate cyclase two-hybrid (BACTH) assay suggests
that these genes do not interact with each other at the protein level. Two determine what these
proteins are interacting with, co-immunoprecipitation with the tagged toxin and antitoxin will be
down during phage infection. The role of the toxin/anti-toxin system within the PLE is not known,
yet we hypothesize that the toxin/anti-toxin may help maintain the PLE within the superintegron.
To investigate this hypothesis, the toxin and anti-toxin were expressed on plasmids in a PLE minus
background, either by themselves or together. Plasmid loss was lower when the toxin and anti-
toxin were co-expressed compared to when the anti-toxin was expressed on its own. Preliminary
results from a subsequent experiment suggest that PLE loss is higher when the toxin has been
deleted. Further molecular studies will improve our understanding of how the unknown ORFs of
the PLE function to prevent productive phage infections.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
144 Section V: Structure and Molecular Mechanisms
Septimavirus genus bacteriophage vB_PaeS_PS9N – virion DNA structure
and post-infection bacterial phenotypes
Aleksandra Głowacka a, Agnieszka Bednareka, Kamil Dąbrowski a, Jan Gaworb, Wioleta Woźnicaa,
Monika S. Hejnowicza, Beata Weber-Dąbrowskac, Anna Baraniakd , Marek Gniadkowskid, Robert
Gromadkab, Andrzej Górskic, Małgorzata Łobocka a,e,*.
aDepartment of Microbial Biochemistry, bLaboratory of DNA Sequencing and Oligonucleotide
Synthesis, Institute of Biochemistry and Biophysics, Polish Academy of Sciences, Warsaw,
Poland, cLaboratory of Bacteriophages, Ludwik Hirszfeld Institute of Immunology and
Experimental Therapy, Polish Academy of Sciences, Wrocław, Poland, dDepartment of
Epidemiology and Clinical Microbiology, National Medicines Institute, Warsaw, Poland, eAutonomous Department of Microbial Biology, Faculty of Agriculture and Biology, Warsaw
University of Life Sciences, Warsaw, Poland
*E-mail: [email protected]
Pseudomonas aeruginosa bacteriophages are considered to be alternatives to antibiotics in curing
infections by antibiotic resistant P. aeruginosa strains. However, only phages that are obligatorily
lytic and cannot transfer DNA between bacteria by transduction can be considered as possible
candidates for therapeutic applications. Here we characterize a newly isolated, obligatorily lytic P.
aeruginosa siphovirus, vB_PaeS_PS9N (PS9N). Its genomic sequence appears to be highly
homologous to the sequences of Septima3virus genus phages (1). Certain of them in addition to
infecting P. aeruginosa strains, can also infect certain strains of Stenotrophomonas maltophilia (2).
The distribution of homologies between PS9N and its relatives is mosaic, with no single phage
being the most closely related over the entire genome length. However, based on homologies of
PS9N tail structures to the tail structures of selected Septima3virus genus representatives, we
predicted that the specificity of PS9N could also cover strains of two different bacterial families.
Indeed, of 13 clinical S. maltophilia isolates one was sensitive to PS9N. Despite the narrow range
of strain specificity against P. aeruginosa and S. maltophiliaI, PS9N could infect certain
carbapenem sensitive as well as carbapenem resistant P. aeruginosa strains. Surprisingly, the
dramatic lysis of susceptible cells infected with PS9N was followed after a few hours by the
overgrowth of phage resistant cells. Moreover, when the infecting phage was propagated in
meropenem resistant P. aeruginosa cells, cells that could grow on meropenem supplemented
medium appeared among the progeny of phage resistant cells. Whether these cells represent
transductants or spontaneous mutants whose selection is favored in phage resistant cells remains to
be found. We show that PS9N DNA is packed to phage heads by a headful packaging mechanism
and identify the PS9N pac site. Analysis of the PS9N genome indicates the lack of genes encoding
enzymes that degrade DNA of infected bacterium. Headful packaging and the lack of genes
encoding host DNA degrading enzymes are characteristic features of transducing phages. Thus, a
possible transducing potential of PS9N and other phages of Septima3virus genus should be taken
into consideration in the use of this phages for the therapeutic purposes.
1.Krupovic M, et al., 2016. Taxonomy of prokaryotic viruses: update from the ICTV bacterial and archaeal viruses
subcommittee. Arch Virol. 161:1095-1099.
2. Peters DL, et al., 2015. The isolation and characterization of two Stenotrophomonas maltophilia bacteriophages
capable of cross-taxonomic order infectivity. BMC Genomics.16:664.
This work was supported by funds from the Operational Program ‘Innovative Economy, 2007-2013’ Development
Project No. POIG 01.03.01-02-003/08, and from the statutory funds for the Institute of Biochemistry and Biophysics of
the Polish Academy of Sciences

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section V: Structure and Molecular Mechanisms 145
Capsid localization of the AFN38122.1 (A2), AFN38181.1 (A4),
AFN38152.1 (A8) proteins on staphylococcal phages 676Z and A3R.
Zuzanna Kaźmierczak1, Katarzyna Hodyra-Stefaniak1, Joanna Majewska1, Sylwia Nowak2,
Marek Harhala1, Paulina Miernikiewicz1, Katarzyna Zielonka1, Dorota Lecion1, Andrzej Górski1,
Krystyna Dąbrowska1, Marta Mazurkiewicz-Kania2, Andrzej Górski1, Barbara Owczarek1, Anna
Kłopot1, Karolina Wojtyna1
1Institute of Immunology and Experimental Therapy, Polish Academy of Sciences, R. Weigla 12,
53-114 Wroclaw, Poland, 2Institute of Experimental Biology, University of Wrocław, Sienkiewicza
21, 50-335 Wrocław, Poland
Objectives: Bacteriophages A3R and 676Z infect Staphylococcus aureus strains pathogenic to
humans. Recently both phages were a subject of an extensive studies. Genomes of A3R and 676Z
were sequenced in 2014. The size of genomes are 141018 bp and 148564 bp, respectively. Both
phages are used in the Phage Therapy Unit in Wrocław, Poland. Phages were morphologically
described and sequenced. The protein structure of phages is not known.
Interests: In these studies we analyzed genomes of A3R and 676Z and annotations of hypothetical
capsid proteins. We have selected several genes encoding hypothetical capsid proteins. In further
research we cloned and expressed genes to obtain selected proteins in order to investigate whether
they are present on phages capsids.
Methods: C57Bl6/J mice were given purified phage proteins (A2 or A4 or A8) in PBS,
subcutaneously in day 0, 20, 40. Control group was given PBS s.c. After 40 days blood samples
were collected and evaluated for the presence of anti-protein IgG antibodies. Antibody levels were
measured by ELISA immunoassay. Phages were purified and concentrated. Next, phages were
incubated with an appropriate antibody conjugated with gold nanoparticles (5nm in diameter),
deposited on a girds, fixed and stained. Phages were examined by TEM.
Conclusions: These data show that proteins A2, A4, A8 are present on capsids of both
staphylococcal bacteriophages A3R and 676Z. A2 which was annotates as hypothetical major
capsid protein, occurs on both heads of staphylococcal phages. Protein A4 is helically present on
the tail of both phages. The base plate of A3R and 676Z phages is co-built by A8 protein.
Acknowledgements: This work was supported by the National Science Centre in Poland grant
UMO-2012/05/E/NZ6/03314 and by the Wroclaw Centre of Biotechnology, programme The
Leading National Research Centre (KNOW) for years 2014–2018.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
146 Section V: Structure and Molecular Mechanisms
Natural deletion mutant LysF1 of Kayvirus endolysin with a broad range of
lytic activity and characterization of its SH3b cell wall binding domain
Martin Benesik1, Jiri Novacek2, Lubomir Janda2, Radka Dopitova2,3, Marketa Pernisova2,3, Katerina
Melkova2, Jiri Doskar1, Lukas Zidek2,3, Jan Hejatko2,3, Roman Pantucek1*
1 Department of Experimental Biology, Faculty of Science, Masaryk University, Kotlarska 2, CZ-
61137 Brno, Czech Republic, 2CEITEC – Central European Institute of Technology, Masaryk
University, Kamenice 5, CZ-62500 Brno, Czech Republic, 3National Centre for Biomolecular
Research, Faculty of Science, Masaryk University, Kamenice 5, CZ-62500 Brno, Czech
Republic
*E-mail: [email protected]
Bacteriophages with a broad host range are suitable candidates for fighting unwanted bacteria in
medicine, food industry, biotechnology and agriculture. Polyvalent ”Twort-like” bacteriophages of
the family Myoviridae and genus Kayvirus are used for the lysis of the serious bacterial pathogens
of the genus Staphylococcus. Their genomic, proteomic and structural characterization suggests
they are safe for phage therapy.
In the Czech Republic, the spontaneous broad host-range mutants 812F1 and 812K1/420 derived
from polyvalent phage 812, belonging to family Myoviridae and genus Kayvirus, are used for
phage therapy of staphylococcal infections. Their endolysin, designated LysF1, has a deleted
middle amidase (Ami-2) domain compared to wild-type LysK endolysin. This two-domain
enzyme, consisting of an N-terminal cysteine-histidine-dependent aminohydrolase/peptidase
(CHAP) domain and C-terminal SH3b cell wall-binding domain (CBD) has never been tested for
its antimicrobial activity. In this work, LysF1 and both its domains were prepared as recombinant
proteins and their function was analyzed. LysF1 had an antimicrobial effect on 31 Staphylococcus
species of the 43 tested, while phage 812F1 was only able to lyse 14 of them. CBD had a crucial
role in binding the enzyme to the bacterial cell wall, since the lytic activity of the truncated variant
LysF1 containing the CHAP domain alone was decreased.
The results of a newly designed co-sedimentation assay of CBD with peptidoglycan (PG) showed
that it was able to bind to three types of purified staphylococcal PGs 11.2, 11.3 and 11.8, which
differ in their peptide bridge, but also to the PG of Streptococcus uberis (PG group 11.5), and this
capability was verified in vivo using the fusion protein with GFP and fluorescence microscopy.
Using several different approaches, including NMR, we have not confirmed the previously
proposed interaction of the SH3b domain with the pentaglycine bridge in the bacterial cell-wall.
The new naturally raised deletion mutant endolysin LysF1 is smaller than LysK, has a broad lytic
spectrum and therefore is an appropriate enzyme for practical use as an enzybiotic against most
frequent Staphylococcus pathogenic species, such as S. aureus, S. epidermidis, but also against
other significant pathogens such as S intermedius, S. lugdunensis, S. haemolyticus, S.
saprophiticus, and S. warneri. The broad binding spectrum of SH3b domain is a promising feature
for creating new chimeric endolysins by combining it with more effective catalytic domains.
Supported by the Ministry of Agriculture (QJ1510216) and Ministry of Health of the Czech
Republic (16-29916A).

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Section V: Structure and Molecular Mechanisms 147
Campylobacter bacteriophage DNA modifications are conserved in each
phylogenetic group
Jessica C. Sacher1, 2, Yan-Jiun Lee3, James Butcher4, Annika Flint4, Ian F. Connerton5, Alain
Stintzi4, Peter R. Weigele3, and Christine M. Szymanski1,2
1Department of Biological Sciences, University of Alberta, Edmonton, Canada, 2Complex
Carbohydrate Research Center and Department of Microbiology, University of Georgia, Athens,
Georgia, USA, 3New England BioLabs, Ipswich, Massachusetts, USA, 4Department of
Biochemistry, Microbiology and Immunology, University of Ottawa, Ottawa, Ontario, Canada, 5Division of Food Sciences, University of Nottingham, Nottingham, UK
E-mail: [email protected]
The refractory nature of Campylobacter bacteriophage DNA to manipulation has been cited for
several years, leading to the speculation that phages targeting this organism encode heavily
modified DNA. Campylobacter phages fall into three phylogenetic groups, all within the family
Myoviridae: Cp220likeviruses (group II), Cp8unalikeviruses (group III) and the as-yet
understudied group I phages. These groups were originally designated based on capsid and genome
size. However, the genomes of several phages from groups II and III have since been sequenced,
which has provided phylogenetic support for these groupings. It has also recently been
demonstrated that phages of groups II and III separate based on their host recognition properties,
with group II phages requiring motile host flagella and group III phages recognizing capsular
polysaccharide receptors.
To characterize the DNA modifications in Campylobacter phages, we analyzed genomic DNA
from one representative group I phage, two group II phages and two group III phages by liquid
chromatography/electrospray ionization mass spectrometry and restriction endonuclease profiling.
Remarkably, we found that phages from groups II and III display complete substitution of
deoxyguanosine residues for modified bases, while group I phage DNA displays partial base
substitution. We found the modifications to be conserved within, but not between, phylogenetic
groups. To gain insights into the biosynthesis of these non-canonical bases, we analyzed the
genome of phages from each of groups II and III. We found several putative group II modification
biosynthetic genes encoded on the group II phage genome, but a clear pathway for biosynthesis of
the group III-specific modification was not apparent. To identify candidate genes involved in
synthesizing this modified base, we examined total host and phage gene expression during group
III phage infection using RNA-Seq. Interestingly, we observed phage-induced up-regulation of
several C. jejuni purine biosynthesis genes as well as several genes involved in nucleic acid
modification and deoxyribonucleotide synthesis. Further studies are required to elucidate the extent
to which these or other phage-induced host genes contribute to the biosynthesis of modified phage
DNA.
Together our results point toward three separate mechanisms for Campylobacter phage DNA
modification, which may have evolved as a method of protecting DNA from enzymatic digestion
upon host infection. In addition to elucidating the biosynthetic pathways for synthesis and
incorporation of these modifications, our current and future studies are focused on the properties
that allow for their accommodation by phage DNA polymerases as well as the biological
implications of these modifications on phage-host interactions.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
148 Section V: Structure and Molecular Mechanisms
In silico analysis of phage proteins based on machine learning
Patryk Waśko
E-mail: [email protected]
Introduction: Mammalian immune system is designed to protect them against potential pathogens.
For the needs of bioinformatics, these potential pathogens could be simplified to particles that do
not belong to the organism and are therefore recognized as foreign. However, some proteins trigger
more intense immune response than the others and the intensity of immune response can be
determined by a variety of biological and physical factors.
Objectives: Machine learning is a promising approach for an out of the box analysis of complex
sets of data with many dependencies. I applied the principles of machine learning to investigate
phage proteins with respect to their potential interactions with mammalian immune system. In the
analyses, I chose to focus on the induction of immune response based on major histocompatibility
complex epitopes.
Methods: In each iteration I analyzed a model with two sets of proteins: one that contains proteins
with high probability of being immunogenic and the second one with “normal” ones that reflect
standard distribution of proteins. Each protein in both sets was mapped to its epitopes with suitable
score from epitope database. Those were used to train the classifier that will be used in the second
phase of the project to analyze other – in this case phage – proteins.
Discussion: One of the features that comes with machine learning is that the results are self-tested.
When sets of data are used to train the classifier, random part of the set is picked to check the
accuracy of the method and logic. Self-testing also allows for determining which of the data have a
real impact on the investigated attribute.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VI: Temperate Phages 149
Intact, widespread, and mosaic prophages of the prominent gut symbiont
Bacteroides
Danielle Campbell*, Patrick Degnan
Department of Microbiology, University of Illinois at Urbana-Champaign, Urbana, IL
*E-mail: [email protected]
Objectives: The human gut harbors a dense and diverse microbial community that plays a
significant role in human health and development. The bacterial genus Bacteroides is one of the
most abundant bacterial genera in the human gut microbiota, playing important roles in complex
carbohydrate digestion and immune modulation. Despite this, little is known about the phages that
infect it. Characterized here is BVΦ01, a novel temperate phage infecting the Bacteroides.
Methods: Genetic modification of phage and host genomes was performed by allelic exchange
using a counter-selectable vector system. Release of prophages from bacterial hosts was tested by
PCR of cell and phage fractions. In silico comparative analyses of related prophages and genes was
accomplished using a combination of homology, alignment and phylogenetic approaches.
Results: Phage BVΦ01 was first identified as a prophage on the chromosome of the genetically-
tractable host B. vulgatus ATCC 8482. The bacterial attachment site (attB) utilized by phage
BVΦ01 is widely conserved across gut isolates of the family Bacteroidales. This attachment site is
used by several intact BVΦ01-like prophages as well was as several divergent prophages in other
Bacteroidales. Moreover, regions of the BVΦ01 genome have been observed in human gut-
associated viromes.
Prophage BVΦ01 is not inducible with well-known phage-inducing agents that cause DNA stress,
however it is constitutively released from the host at low levels under laboratory conditions. Phage
BVΦ01 harbors three integrase-like genes that are not only highly divergent from each other, but
also from other known phage integrase-like sequences. Here, we’ve demonstrated that one of these
integrases is required for BVΦ01 excision from the host genome. Cell lysis is predicted to be
caused by a conserved, putative holin-lysin-spanin operon in BVΦ01. Heterologous expression of
this operon is sufficient to cause death in both Bacteroides BVΦ01-lysogens and non-lysogens. Co-
culture experiments show that phage BVΦ01 confers context-dependent growth advantages for its
bacterial host.
Conclusions: Phage BVΦ01 is an intact prophage infecting the gut symbiont Bacteroides. We
have validated the activity of 4 highly divergent genes encoding essential functions for BVΦ01
lysis and integration. BVΦ01 has homology to a small cluster of closely related BVΦ01-like
phages and also a broader group of phages utilizing the same attB suggesting that BVΦ01 is a
representative of a broad group of mosaic phages infecting gut symbionts.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
150 Sections VI: Temperate Phages
Insights into phage-mediated mechanisms of Mycobacterial host immunity
Catherine M. Mageeney*1, Marta Dies*2, Javier Buceta2, Vassie C. Ware1
1Department of Biological Sciences, 2Department of Chemical and Biomolecular Engineering
Lehigh University, Bethlehem, PA 18015 (*contributed equally to work)
Corresponding Author: [email protected]
Objectives: Mycobacteriophage Butters (cluster N) has one of the smallest known annotated
mycobacteriophage genomes: 41,491 bp with 66 ORFs and 4 novel genes (gp30, gp31, gp33, gp34)
with no mycobacteriophage homology called orphams, found within the central “variable” region
of the genome where nucleotide and gene content differences are most apparent between N cluster
phages. Work presented here is aimed at characterization of Butters orphams gp30 and gp31. First,
we have explored the cellular localization of both proteins: GP31 is predicted computationally to
be a 4-pass transmembrane protein while GP30 has no predicted membrane domains and is likely
cytoplasmic. Secondly, we have utilized M. smegmatis strains expressing gp30 or gp31 alone or
together to probe immunity mechanisms related to prophage-mediated defense against viral attack.
Methods: Butters GP30 and GP31 were tagged with a tetracysteine tag and expressed in E. coli.
Imaging of fluorescent proteins was completed using a microfluidics fluorescence microscopy
system. Immunity testing was completed using a series of phages from various clusters, excluding
other N cluster phages. Phages were spotted onto lawns of M. smegmatis expressing gp30 or gp31
alone or in tandem. Changes in phage infection for wildtype M. smegmatis, a Butters lysogen, and
strains expressing Butters proteins were monitored over a 48-hour period and efficiency of plating
was recorded.
Results: Mycobacteriophage Butters GP30 is localized in the cytoplasm while GP31 is localized to
the membrane, co-localizing with an E. coli membrane marker. With dual expression of GP30 and
GP31 from the same vector, GP30 localization changes and appears to be sequestered at the
membrane, suggesting a potential interaction between GP30 and GP31. Immunity results differ
depending on which genes are expressed. M. smegmatis strains expressing only gp30 mount a
defense against A3 phage PurpleHaze. Defense against viral attack is not observed when gp31 is
expressed alone but is abrogated when gp30 and gp31 are expressed together.
Conclusions: Taking the imaging and immunity data together, we propose that GP30, in its
cytoplasmic state, confers defense against specific viral attack in the lysogen. Additional factors
contributing to the defense mechanism are unknown, but may include GP30 release from
membrane sequestration (facilitated by GP31). Other Butters variable region genes (expressed in
the lysogen) may influence the putative release of GP30 from membrane association and confer
defense against other phages. A model for Butters defense against viral attack involving GP30 and
GP31 will be presented.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VI: Temperate Phages 151
Lysogenic conversion of Stenotrophomonas maltophilia D1585 by temperate
phage DLP4
Danielle L. Peters*, Paul Stothard and Jonathan J. Dennis
Department of Biological Sciences, University of Alberta, Edmonton, AB T6G 2E9, Canada.
*E-mail: [email protected]
Objectives: The use of bacteriophages to treat antibiotic resistant bacterial infections requires the
characterization of potential therapeutic phages. Lifestyle determination and genome annotation for
each phage must be conducted as temperate phages can produce lysogenic conversion of their
bacterial host. Bacteriophage DLP4 was isolated from asparagus soil collected in Edmonton,
Alberta, Canada using the highly antibiotic resistant bacterium Stenotrophomonas maltophilia
strain D1585. Characterization of DLP4 revealed it is a temperate phage capable of establishing a
lysogenic cycle in D1585. Due to the temperate nature of DLP4, genomic analysis and
identification of important moron genes was imperative to determine how suitable DLP4 is for
inclusion in a therapeutic phage cocktail.
Methods: Morphology was determined with a 4% uranyl acetate stain of DLP4 lysate on a copper
grid and viewed with a Philips/FEI transmission electron microscope. Host range analysis was
conducted with 27 clinical S. maltophilia and 11 Pseudomonas aeruginosa strains. Restriction
fragment length polymorphism analysis was performed using 36 FastDigest enzymes and separated
on a 1% agarose gel. To date, two moron genes of particular interest have been identified and are
being investigated for their functionality using complementation testing and observation of
phenotypic and transcriptomic changes between the DLP4 lysogen and wild type D1585.
Results: DLP4 is a member of the Siphoviridae family. Host range analysis indicated that DLP4
infects 14 of 27 clinical S. maltophilia isolates tested. All 36 restriction enzymes tested failed to
digest the genomic DNA. DLP4 was assembled into a 63,945 bp contig that was confirmed with
PCR and Sanger sequencing. DLP4 is predicted to encode 83 ORFs, one tRNA and an attP site.
Two moron genes currently under investigation are folA (BIT20_024) and ybiA (BIT20_065). The
folA gene encodes dihydrofolate reductase (DHFR), an enzyme responsible for trimethoprim
resistance. I-TASSER analysis of DLP4 DHFR predicts it is structurally similar to Bacillus
anthracis DHFR. Trimethoprim resistance in Escherichia coli (E. coli) DH5α increases 260-fold
with DLP4 folA expressed from the pBBR1-MCS plasmid compared to the empty vector control.
The ybiA gene encodes N-glycosidase YbiA, which is required for the swarming phenotype of E.
coli K-12. Complementation of E. coli BW25113 ybiA- with DLP4 ybiA shows partial
complementation, with swarming ability greatly improved over the ybiA- pBBR1-MCS control.
Conclusions: Although DLP4 is not suitable for therapeutic use due to the presence of moron
genes, these findings are the third documented incidence of lysogenic conversion in S. maltophilia.
More research into S. maltophilia temperate bacteriophages could help elucidate the role of phages
in S. maltophilia infections. Future research using reverse transcription PCR will identify whether
ybiA and folA are expressed in the lysogenic cycle so their potential impact on host cell
pathogenicity and antibiotic resistance can be determined.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
152 Sections VI: Temperate Phages
Activation of a Bacillus subtilis prophage encoded on the pBS32 plasmid
releases defective particles
Jolene Ramsey, Bat-Erdene Myagmarjav, Melissa A. Konkol, Suchetana Mukhopadhyay, Daniel
B. Kearns
Indiana University Bloomington, Bloomington, IN
*E-mail: [email protected]
Objective: The ancestral Bacillus subtilis strain 3610 contains several prophages in the
chromosome, including PBSX and SPβ, that can be induced to excise and generate particles. The
ancestral strain also harbors an 84-kb plasmid called pBS32 that was lost during domestication of
commonly used laboratory derivatives. pBS32 encodes the protein ComI, which is a potent
inhibitor of DNA uptake by natural transformation, and the genes of a putative prophage. We
hypothesized that the coexistence of the prophage genes and competence inhibition were linked to
lysogeny and therefore sought to characterize the phage.
Methods: After induction with the DNA damaging agent mitomycin C, we monitored plasmid-
dependent host growth and lysis. Supernatant from lysed host cells was assayed for phage particles
and the ability to transfer plasmid DNA. Isolation of phage particles for transmission electron
microscopy was attempted using precipitation and various gradients.
Results: When chromosomal prophages were deleted, we observed pBS32-dependent cell killing
upon addition of mitomycin C. The plasmid copy number increased by nearly 100 fold, and was
protected in supernatant from DNase treatment by a heat-labile factor. A complex including the
ZpbH capsid protein from the pBS32 prophage sedimented in CsCl gradients, but particles were
not observed by electron microscopy, nor were naïve B. subtilis killed by addition of these purified
complexes. Killing activity was linked to expression of a plasmid-encoded sigma factor, ZpdN.
Conclusion: The extrachromosomal plasmid pBS32 of B. subtilis encodes a prophage that, when
activated, kills the host. We hypothesize that pBS32-mediated cell death releases a phagelike
particle that is defective and unstable.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VI: Temperate Phages 153
Genomics of Staphylococcus sciuri siphophages
Michal Zeman1*, Ivana Mašlaňová1, Adéla Indráková1, Marta Šiborová2, Pavel Plevka2, Veronika
Vrbovská1, Jiří Doškař1, Roman Pantůček1
1Department of Experimental Biology, Faculty of Science, Masaryk University, Kotlářská 2, 611
37 Brno, Czech Republic. 2Central European Institute of Technology, Masaryk University,
Kamenice 5, 625 00 Brno, Czech Republic.
*E-mail: [email protected]
Objectives: Staphylococcus sciuri is a bacterial pathogen associated with infections in animals and
humans, and represents a reservoir for the mecA gene encoding methicillin-resistance in
staphylococci. Even though there have been 15 genomes of S. sciuri in the public databases, no
S. sciuri siphophages or prophages were known until now.
Methods: Genomes of S. sciuri strains P575 and P879, and two phages induced from these strains
were sequenced by Ion Torrent™ PGM. Phages were subjected to electron microscopy, adsorption
experiments, their genomes were in silico analysed for presence of pathogenicity related genes and
compared to publicly available prophage genomes. Twenty-one S. sciuri prophages were manually
located in S. sciuri genomes obtained from the public database. Phage genomes were compared by
Mauve and ClustalW.
Results: Transmission electron microscopy and cryo-electron microscopy confirmed that phages
φ575 and φ879 belong to Siphoviridae family. Cryo-EM revealed spike at the base plate of the both
phages. Phages φ575 and φ879 were able to adsorb to various hosts even from different
staphylococcal species but they were unable to propagate on non-S. sciuri strains. Reversible
adsorption to Staphylococcus aureus suggested a two-stage adsorption mechanism. Whole genome
sequencing revealed that both phages have genomes with cos ends, and the size and modular
structure of their genomes is similar to other staphylococcal siphoviruses. Phage φ575 encoded
genes for staphylokinase and phospholipase in the genome and phage φ879 was able to package
plasmid mediating resistance to aminoglycoside antibiotics and staphylococcal chromosome
cassette mec from host strain. Two additional prophages were identified in the S. sciuri strain P879
(phage φ879 was induced from this host). Comparative genomic analysis distinguished 10 different
prophages and divided them into 4 distinct clusters. Phage integration sites varied in the host
genome. No known virulence factors were found however metallo-beta-lactamase or
acetyltransferase were present in the genome of some prophages.
Conclusions: The identification of bacterial virulence factors and possible antibiotic resistance
determinants encoded by S. sciuri phages and prophages, their ability to package and transmit
mobile elements and to adsorb onto the cells of other staphylococcal species show that S. sciuri
siphoviruses may contribute to the horizontal gene transfer within the Staphylococcus genus.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
154 Sections VII: Phage Therapy
Biology and genomics of an historic therapeutic Escherichia coli
bacteriophage collection
Abiyad Baig1, Joan Colom1, Paul Barrow1, Catherine Schouler2, Arshnee Moodley2, Rob Lavigne3
and Robert Atterbury1*
1School of Veterinary Medicine and Science, Sutton Bonington Campus, University of
Nottingham, United Kingdom, 2ISP, INRA, Université François Rabelais de Tours, 37380
Nouzilly, France, 3Department of Veterinary and Animal Science, Faculty of Health and Medical
Science, University of Copenhagen, 4Laboratory of Gene Technology, Department of Biosystems,
KU Leuven, Belgium
*E-mail: [email protected]
We have performed microbiological and genomic characterisation of an historic collection of nine
bacteriophages, specifically infecting a K1 E. coli O18:K1:H7 ColV+ strain. These phages were
isolated from sewage and tested for their efficacy in vivo for the treatment of systemic E. coli
infection in a mouse infection model by Smith and Huggins, 1982. Transmission electron
microscopy showed that six of the nine phages were Podoviridae and the remaining three were
Siphoviridae. The four best performing phages in vivo belonged to the Podoviridae family. In vitro,
these phages exhibited very short latent and rise periods in our study.
In agreement with their microbiological profiles, characterisation by genome sequencing showed
that all six podoviruses belong to the Autographivirinae subfamily. Of these, four were isolates of
the same species (99 % identity), whereas two had divergent genomes compared to other
podoviruses. The Siphoviridae phages, which were moderate to poor performers in vivo, exhibited
longer latent and rise periods in vitro. Two of the three siphoviruses were closely related to each
other (99 % identity), but all can be associated with the Guernseyvirinae subfamily. Genome
sequence comparison of both types of phages showed that a gene encoding for DNA-dependent
RNA polymerase was only present in phages with faster replication cycle, which may account for
their better performance in vivo. These data allow a more rational evaluation of the original in vivo
data and pave the way for the selection of phages for future phage therapy trials.
The current alarming rise in levels of antibiotic resistance is an increasing threat to animal and
public health. To address this concern there is an immediate need for new alternative therapeutics.
Phages have been investigated historically and shown to be effective in controlling bacterial
infections. The significance of the present study is that it correlates the microbiological, genomic
and in vivo performance of an historic collection of phages to identify a combination of
characteristics which can be used to make an informed and rational selection of phages for use in
the treatment of systemic E. coli infections.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 155
Isolation of phages targeting different adherent-invasive E. coli strains
Andrei Sorin Bolocan*, Siobhan McDonnell, Ciorsdan Campion, Emma Guerin, Amanda Forde,
Fergus Shanahan, Paul Ross, Colin Hill
APC Microbiome Institute, University College Cork, Cork, Ireland
*E-mail: [email protected]
The threat of an antibiotic resistance crisis has revived interest in diverse biological approaches
against infectious diseases, including the use of phages against bacterial infections. Due to its
malleable genetic character, E. coli has one of the widest spectra of disease of any bacterial species.
E. coli phages are commonly isolated from sewage, hospital waste water, polluted rivers and faecal
samples of humans or animals. The adherent invasive E. coli (AIEC) pathotype has been associated
with intestinal diseases but its prevalence in these diseases remains largely unexplored. The aim of
this study was to isolate broad host range phages from human faeces to target different AIEC E.
coli isolates.
Phages were isolated from faeces using two enrichment steps, whereby the faecal material was
mixed with multiple- or individual- E. coli strains in Luria Bertani broth (LB) supplemented with
CaCl2 and MgSO4, and incubated shaking for 24h at 37°C. Following centrifugation and filtration,
samples were analysed for the presence of the phages using standard spot and double-layer plaque
assay techniques. Of 50 plaques tested we found at least five distinct phage types on the bases of
RAPD-PCR and DNA enzymatic restriction profile analysis. Genome sequencing revealed the
virulent nature of all five phages. Eighteen AIEC strains isolated from faeces and discriminated by
the fingerprint profiles generated by rep-PCR were included for host range assay. For comparison,
15 non-AIEC E. coli strains were also tested, including representatives of the uropathogenic E.
coli (UPEC- UTI), enterotoxigenic, E. coli (ETEC), enteropathogenic E. coli (EPEC)
enterotoxigenic E. coli (ETEC) and enteroaggregative E. coli (EAggEC or EAEC). The five phages
demonstrated broad host range specificity in a spot test, especially against UPEC and AIEC. We
plan to investigate their therapeutic potential for some of the diseases in which these pathotypes are
implicated as causative agents.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
156 Sections VII: Phage Therapy
Phage D29 titer reduction due to shipping, saline preparation, and
aerosolization
Nicholas B. Carrigy1, Rachel Y. Chang2, Sharon S.Y. Leung2, Melissa Harrison3, Zaritza Petrova4,
Welkin H. Pope4, Graham F. Hatfull4, Warwick J. Britton5, Hak-Kim Chan2, Dominic Sauvageau3,
Warren H. Finlay1, and Reinhard Vehring1*
1 Department of Mechanical Engineering, University of Alberta, Edmonton, AB, Canada, 2 Faculty
of Pharmacy, University of Sydney, Sydney, NSW, Australia, 3 Department of Chemical &
Materials Engineering, University of Alberta, Edmonton, AB, Canada, 4 Department of Biological
Sciences, University of Pittsburgh, PA, USA, 5 Centenary Institute of Cancer and Cell Biology, and
Sydney Medical School, Royal Prince Alfred Hospital, and University of Sydney, Sydney, NSW,
Australia
*E-mail: [email protected]
Objective: To determine titer reduction when anti-tuberculosis phage D29 is shipped, diluted into
saline, and aerosolized with clinically-relevant inhalation devices.
Methods: Phage D29 was amplified and shipped from Canada to Australia using cold packs and an
insulated container. At arrival, this lysate was diluted 1:100 in isotonic saline, and the titer
measured after 18 hours of storage at room temperature. The diluted phage D29 preparation was
then input to three inhalation devices: 1) PARI LC Sprint jet nebulizer; 2) Aerogen Solo vibrating
mesh nebulizer; 3) Respimat soft mist inhaler. The titer reduction due to aerosolization was
measured by capturing the emitted aerosol on a filter and performing full-plate plaque assays in
triplicate at multiple dilution levels with Mycobacterium smegmatis. A mathematical model
predicted the number of nebulization cycles the phage underwent with the jet nebulizer, which
corresponds to the number of baffle impactions.
Results: Titer was not significantly different (p > 0.2) after 2-day shipping with cold packs, during
which the temperature increased from 5°C to 20°C, with a mean temperature of 11°C. Neither was
there significant phage deactivation due to dilution into isotonic saline over the tested time period
of 18 hours (p > 0.1). Aerosolization titer reduction was device-dependent, being 3.7 ± 0.1
log(pfu/mL) for the jet nebulizer, 0.4 ± 0.1 log(pfu/mL) for the vibrating mesh nebulizer, and 0.6 ±
0.3 log(pfu/mL) for the soft mist inhaler. On average, phage were predicted to undergo 108
nebulization cycles prior to exiting the mouthpiece of the jet nebulizer.
Conclusions: Phage D29 stable after shipping and dilution in saline at room temperature for at
least 18 hours. It survived aerosolization depending on the inhalation device used. The vibrating
mesh nebulizer may be useful for animal studies requiring high titers of active phage delivered
quickly, and the soft mist inhaler for self-administration phage therapy applications. Repeated
baffle impaction may be the cause of titer reduction with the jet nebulizer, which makes this device
a poor choice for animal studies with D29. Other phage readily survive jet nebulization, however,
showing that robustness of phages is variable and needs to be tested on a case-by-case basis.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 157
Preparation of liposome-encapsulated bacteriophage PEV2 formulation
Sharon S.Y. Leung1, Sandra Morales2, Warwick Britton3, Elizabeth Kutter4, Hak-Kim Chan1*
1 Faculty of Pharmacy, University of Sydney, Sydney, NSW 2006, Australia, 2AmpliPhi
Biosciences AU, 7/27 Dale Street, Brookvale, Sydney, NSW 2100, Australia, 3The Evergreen State
College, Olympia, Washington 98502, USA, 4Centenary Institute of Cancer and Cell Biology, and
Sydney Medical School, Royal Prince Alfred Hospital, and University of Sydney, Sydney, NSW
2006, Australia
*E-mail: [email protected]
Objective: Liposome-encapsulated (LE) phages have been reported to improve phage survival rate
and biodistribution in body. Conventional thin film hydration followed by extrusion was generally
employed to produce LE formulation. This study investigated the feasibility of using a microfluidic
approach to encapsulate a Pseudomonas aeruginosa phage PEV2 into liposomes.
Methods: A microfluidic setup consisted of a stainless steel cross connector (U-430, IDEX Health
& Science, USA) and a fluorinated ethylene propylene tube with an internal diameter of 200 µm
and 100 mm in length were used to encapsulate phage into liposomes. A mixture of soy
phosphatidylcholine and cholesterol at a ratio of 4:1 dissolved in absolute ethanol with a total solid
content of 17.5 mg/mL was injected through the center inlet channel of the cross mixer. Phage
suspension was injected into the cross mixer from the two side channels intersecting with the center
channel. The produced liposome was collected into an Eppendorf tube at the outlet of the mixing
FEP tube. The lipid and phage suspension flow rates were set at 80 and 160 µL/min, respectively.
The size and polydispersity index (pdi) of the produced liposomes were analyzed by dynamic light
scattering using Zetasizer Nano-ZS (Malvern Instruments, Worcestershire, UK). The titer reduction
due to the encapsulation process and encapsulation efficiency was determined using a surface-
droplet (Miles-Misra) technique. The phage encapsulation efficiency (EE) was calculated
according to the equation EE (%)=100−(Cfree/Ctotal) × 100, where Cfree is the titer of unencapsulated
phage, and Ctotal is the total phage titer (unencapsulated and encapsulated phages) obtained by
lysing 0.5 ml of liposome-encapsulated phage formulation with 0.5 ml of bile salts (50 mM). Phage
encapsulation using conventional method at a hydration and extrusion temperature of 40 ºC was
performed to compare the encapsulation efficiency and titer reduction upon processing.
Results: The average size and pdi of liposomes produced by the microfluidic method were 242 ±
2.6 nm and 0.33 ± 0.01, respectively, which were larger than the liposomes generated by the thin
film hydration method (size = 207.2 ± 30.1 nm and pdi = 0.22 ± 0.06). The titer reduction caused
by the encapsulation process was similar for both methods (~0.6 log). The microfluidic approach
had slightly lower encapsulation efficiency (35 ± 3%) than the thin film hydration method (46 ±
18%). The formation of liposome in the microfluidic approach relies on the diffusion of ethanol
and water across the ethanol/water interface through a “self-assembly” mechanism. Therefore,
encapsulation of phage would depend on the availability of phage at the ethanol/water interface. On
the other hand, the thin film hydration method involved agitation between the lipid film and phage
suspension to form large heterogeneous liposome before they were extruded to form homogeneous
liposomes of smaller size. This mixing step would more likely increase the contact between phage
and lipid, hence higher encapsulation efficiency.
Conclusions: This study demonstrated the feasibility of a microfluidic approach to encapsulate
phage into liposomes. The effects of process parameters, total liquid flow rate and flow rate ratio,
on the phage encapsulation efficiency and phage viability will have to be optimized.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
158 Sections VII: Phage Therapy
Nebulisation effects on structural stability of bacteriophage PEV 44
Ariel Astudillo1, Sharon Shui Yee Leung1, Elizabeth Kutter2, Sandra Morales3, Warwick J.
Britton4, Hak-Kim Chan1*
1 Faculty of Pharmacy, University of Sydney, Sydney, NSW 2006, Australia, 2 The Evergreen State
College, Olympia, Washington 98502, USA, 3AmpliPhi Biosciences AU, 7/27 Dale Street,
Brookvale, Sydney, NSW 2100, Australia, 4Centenary Institute of Cancer and Cell Biology, and
Sydney Medical School, Royal Prince Alfred Hospital, and University of Sydney, Sydney, NSW
2006, Australia
*E-mail: [email protected]
Objective: Nebulization being used for inhaled phage therapy can cause phage titer losses which
may be the result of mechanical damage to the phages. This study aimed to determine effects of
nebulization on the structural stability and bioactivity of a Pseudomonas lytic phage PEV44, which
is a myovirus consisting of a hexagonal head and a contractile tail connected by a neck.
Methods: Three nebulizers were investigated: 1) Sidestream® jet nebulizer with Porta-neb®
compressor operated at 7.5 L/min, 2) Aeroneb® Go vibrating mesh nebulizer and 3) Omron® NE22
static mesh nebulizer. A volume of 3 ml PEV44 phage suspension (5.2×1010 pfu/mL) was
aerosolized and the emitted aerosols were collected using a test tube impaction apparatus. The
nebulized aerosol was drawn into an ice-cooled test tube placed inside an aspiration flask through a
Tygon tubing connected with a 2.0 mL plastic pipette at 8 L/min. The morphologies of phages in
the stock/nebulized samples were examined by transmission electron microscopy (TEM, JEOL-
JEM 2100 microscope, Japan) using random sampling to identify the proportion of different phage
morphologies. The titer reduction due to nebulization was determined using a surface-droplet
(Miles-Misra) technique. The viable phage respirable fraction (fraction of phages delivered by
aerosols smaller than 4.5 µm) of the three nebulizers was determined using a multi-stage liquid
impinger (MSLI, Copley, U.K.) operated at 30 L/min.
Results: The morphology of the phage in the nebulized samples was categorized into two groups:
“whole” (the capsid and tail held together) and broken (the capsid separated from the tail) phages.
Within the “whole” phage group, they were divided into three groups: 1) intact, 2) contracted tail
and 3) empty capsid. In the stock phage suspension, considerable portions of broken phages (36%),
“whole” phages with contracted tail (10%) and empty capsid (19%) were detected. The fraction of
broken phages was significantly increased after nebulization, with the jet nebulizer (83%) being
more pronounced than the mesh type nebulizers (50 – 60%). While the amount of intact phage
(from 35% to 2 – 10%) and phage with contracted tail (from 10% to 0.2 – 3%) were significantly
reduced, the proportion of phage with empty capsids were not significantly different after the
nebulization process, possibly due to innate elasticity properties of the phage capsids. The loss
titres obtained by TEM quantification (94%, 75% and 70% for the jet, actively and passively
vibrating-mesh nebulizers, respectively) were comparable with those obtained by the conventional
plaque assay results. Among the three nebulizers, the Omron nebulizer achieved a much higher
viable phage respirable fraction (30%) than the SideStream (3%) and Aeroneb Go (6%).
Conclusions: This study identified various changes on the phage structure and viability, and
respirable phage dose from different types of nebulizers. Understanding these effects and the phage
tolerance to nebulization stress can potentially improve our choice of the delivery method for
inhaled phage therapy.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 159
Bacteriophage therapy treats cholera
Sudhakar Bhandare1^, Joan Colom2^, Abiyad Baig2, Jenny M Ritchie3 Paul Barrow2, Robert
Atterbury2*
1Department of Food Science and Agricultural Chemistry, Faculty of Agricultural and
Environmental Sciences, Macdonald Campus, McGill University, 21,111 Lakeshore Ste Anne de
Bellevue, Quebec, Canada, 2School of Veterinary Medicine and Science, Nottingham University,
Loughborough, Leicestershire, UK, 3Faculty of Health and Medical Sciences, AX Building,
08AX01 University of Surrey, Guildford, Surrey, UK.
^ These authors contributed equally to the study
*E-mail: [email protected]
Cholera is a persistent threat to public health and an endemic disease in many countries. Recent
reports of emergence of antibiotic resistance in Vibrio cholerae show the need to obtain new
effective treatments of cholera. One such treatment could be the use of bacteriophages. This study
evaluates the potential of phage being used as a biocontrol for V. cholerae.
Seven phages against V. cholerae were isolated in China and five were obtained from various
sources. These twelve phages were biologically characterised and their genomes were sequenced.
Amongst the phages studied, the phage vB_VcholP_1 had the widest host range (67 % of 91 V.
cholerae strains) and lacked genes related to lysogeny or virulence (antimicrobial resistance or
toxins). The biological characterization of vB_VcholP_1 showed that this phage had a latent period
of 12 ± 0.0 min and a burst size of 43 ± 5.5 PFU/cell. This phage was selected for use in a
therapeutic trial to treat V. cholerae infections in infant rabbits.
The animals in the control group developed cholera whereas no sign of disease was observed in the
phage-treated animals. V. cholerae reductions in the caecal fluids were 3.5, 4.1 and 3.9 log10 CFU/g
(P < 0.01). Similarly, the reductions in the mid colon were 3.0, 3.1 and 4.2 CFU/g (P < 0.01).
Reductions in the counts of more proximal segments upper, middle and lower small intestine were
comparatively lower than those in the distal regions, but still significant (P < 0.01). Phage
replication was detected in all the sampling sites and the MOI was constant through the intestinal
tract with values between 1.8 and 3.5.
Furthermore, phage vB_VcholP_1 was tested as a prophylactic treatment for V. cholerae infection
in the infant rabbit model. After treatment, the animals looked healthy and significant reductions of
V. cholerae were observed along the intestinal tract. In stools reductions of 3.4, 4.3 and 2.5 log10
CFU/g were observed (P < 0.001, P < 0.01, P < 0.05). Comparable reductions of 3.8, 2.9 and 4.0
log10 CFU/g were obtained in mid colon samples (P < 0.001 and P < 0.01). Finally, in the proximal
intestinal tract, reductions were similar to those obtained in the therapeutic trials (P < 0.001, P <
0.01). In this case, the phage replication was only evident in mid colon and stools with constant
MOI values ranging from 1.6 to 2.3.
No phage resistant V. cholerae colonies were detected in any of the treated animals. In conclusion,
phage vB_VcholP_1 applied as a therapeutic or prophylactic agent prevents the death of infant
rabbits infected by V. cholerae, therefore, it may be suitable for the treatment of humans.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
160 Sections VII: Phage Therapy
Faecal phageome transfer reshapes the murine bacteriome after antibiotic
perturbation
Lorraine A. Draper1,2, Feargal J. Ryan1,2, Marion Dalmasso1,2,†, Pat G. Casey1,2, Angela
McCann1,2, Vimalkumar Velayudhan 1,2, R. Paul Ross1,3, Colin Hill1,2*.
1 APC Microbiome Institute, University College Cork, Cork, Ireland, 2 School of Microbiology,
University College Cork, Cork, Ireland, 3 Teagasc Food Research Centre, Moorepark, Fermoy, Co.
Cork, Ireland, †Present address: Normandie Univ, UNICAEN, ABTE, 14000 Caen, France
*E-mail: [email protected]
Background: It has become increasingly apparent that creating and maintaining a complex and
diverse gut microbiome is fundamental to human health. Thus, there are increasing efforts to
identify methods that can modulate and influence the microbiome, especially in those who due to
disease or circumstance have had their native microbiome disrupted.
Methods: The current study describes how the administration of virus-like particles (VLPs)
influenced and reshaped the murine microbiome of test mice following microbial perturbation with
antibiotics. Subsequent to administration of penicillin and streptomycin, test mice (n=8) received a
bacteria free-VLP enriched faecal transplant(s), while control mice (n=8) received a heat-treated
nuclease treated version of the same.
Results: Mice that received VLPs, in which bacteriophages predominate, had a bacteriome (as
determined via 16S rRNA sequencing) that separated into distinct groups via principle co-ordinate
analysis (PCoA) and contained differentially abundant taxa when compared to the control group.
Additionally through metagenomic sequencing of VLP DNA, we were able to obtain a snapshot of
the putative viruses present in the gut of test and control mice at various time points. It was
observed that viral content appeared to differ in both abundance and diversity.
Conclusions: This study has validated the role of bacteriophage in gut microbiome population
dynamics. There is potential to take advantage of such, especially with the increasing trend toward
performing FMTs. Up until now these have focussed on transferring living bacteria and spores; as
bacteriophage are non-living proteinaceous entities they could form a robust, inexpensive
alternative that is susceptible to standardisation and could be delivered as a freeze dried
formulation.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 161
Isolation of broad host range bacteriophages active against multidrug resistant
Acinetobacter baumannii
Reham Samir, Maha Omran and Marwa ElRakaiby*
Faculty of Pharmacy, Cairo University, Cairo, Egypt
E-mail: [email protected]
The evolution of pathogenic microorganisms and their acquisition of resistance elements against
antimicrobial agents is one of the most threatening global risks to human health. A “red alert”
pathogen, Acinetobacter baumannii, poses a particular challenge to the currently used antimicrobial
agents owing to its remarkable ability to up-regulate existing genes or acquire new resistance
mechanisms. With the emergence of multidrug or even pandrug resistant A. baumannii, new
treatment strategies are required. Phage therapy is a promising treatment option because of its high
specificity and minimal side effects.
Objectives: Here, we aim to preemptively fight multidrug resistant A. baumannii, before their
possible spread, using phages isolated from the Egyptian environment.
Methods: Eighteen Acinetobacter baumannii strains were isolated from patients in Egyptian
hospitals. All strains were molecularly typed by oxa51 gene primers. Bacteriophages were isolated
from sewage samples obtained from Al-Gabal El-Asfar treatment station in Cairo governorate.
Phages were purified and propagated in Tryptic Soya Broth (TSB), according to standard
procedures. The agar overlay plaque assay was used to check the lysate for the presence of phages
and subsequent isolation of pure phage particles. Host range of the isolated bacteriophages was
assessed against the clinical isolates. Phages with broad host ranges were chosen for further
analyses.
Results: Twenty-nine phages were isolated from the sewage samples using combinations of A.
baumannii clinical strains. Five of these phages showed high degree of infectivity against the A.
baumannii isolates.
Conclusion: Bacteriophages isolated from the Egyptian environment are capable of infecting and
lysing resistant strains of A. baumannii. Candidate phages will be characterized and identified then
their therapeutic and prophylactic abilities will be tested in murine animal models.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
162 Sections VII: Phage Therapy
Evaluation of the therapeutic potential of bacteriophage ZCKP1 in the
treatment of Klebsiella pneumonia isolated from Egyptian diabetic foot
patients
Omar A. Taha1, Phillippa Connerton2, Ian Connerton2 and Ayman El-Shibiny1*
1Biomedical Sciences, University of Science and Technology, Zewail City of Science and
Technology, Sheikh Zayed District, 12588, Giza, Egypt, 2Division of Food Sciences, School of
Biosciences, University of Nottingham, Sutton Bonington Campus, Loughborough, Leicestershire
LE12 5RD, United Kingdom
*E-mail: [email protected]
The predicted growth in infection by multidrug resistant (MDR) bacteria necessitates prompt
efforts towards developing alternatives to antibiotics, such as bacteriophage therapies. Immuno-
compromised patients with diabetes mellitus are particularly prone to foot infections by MDR
Klebsiella pneumonia, which may be compounded by chronic osteomyelitis. Bacteriophage
ZCKP1ɸ, isolated from Giza, Egypt, was tested in vitro to evaluate its lytic activity against a MDR
Klebsiella pneumonia KP/01, isolated from the foot of a diabetic patient in Egypt. Characterization
of ZCKP1ɸ indicated that it belonged to the Myoviridae family of bacteriophages with an
approximate genome size of 48 kb. ZCKP1ɸ lysed a range of osteomyelitis pathogenic agents with
activity against different Klebsiella spp. strains but also against Proteus spp. and E. coli isolates.
The bacteriophage reduced viability of its hosts by ≥2 log10 CFU/ml at 25°C and remained
functional at 4°C, permitting treatment over a range of temperatures. The phage demonstrated a
significant reduction in biofilm biomass (more than 50%) and viability (up to 66%) with high MOI
(50 PFU/CFU), achieving the greatest effect. These characteristics of ZCKP1ɸ phage indicate
potential therapeutic value to treat Klebsiella pneumonia infections and warrant further
investigation.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 163
Preliminary pharmacokinetic study of phage therapy for the control of
Salmonella in commercial broilers
Juan C. Farfan-Esquivel, Martha J. Vives Florez1
1 University of Los Andes, Colombia.
E-mail: [email protected], [email protected]
Salmonella is a common foodborne pathogen usually found in broilers and related products. Due to
the increasing acquisition of mechanisms that generates resistance to antibiotics there is a necessity
to find new methods for its control. One of these alternative methods is phage therapy, which has
been tested in animal models and germ-free chickens. Nevertheless, this approach has not been
proven in broilers in a real setting or at a production scale. The aim of this project was to perform a
pharmacokinetic analysis of a selected phage from a phage cocktail effective against Salmonella, in
broiler chickens housed in a production facility.
The first, necessary step of the project was the evaluation of the protective effect of three different
excipients to improve the stability and viability of a phage. The excipients tested were the
following: saccharose 64% (SAC), magnesium hydroxide 8.4% (MGH), calcium carbonate 30%
(CAR). Stability assays with excipients were performed in three temperatures: 4°C, 25°C and
37°C. After one month, all excipients retained similar phage titers (1010 PFU/ml) at 4°C and 25°C.
At 37°C, a decrease in phage titer was measured: SAC 106 PFU/ml, MGH 107 PFU/ml, CAR 109
PFU/ml.
To evaluate acid stability, a model was developed resembling the average pH values and retention
times present in the digestive tract of 22-days broilers. Phage suspensions in excipients were
consecutively transferred in three solutions of hydrochloric acid (HCl) at different pH values: 2 to 5
and 5 to 6. Calcium carbonate and magnesium hydroxide were effective at protecting the phages
from acid with the concentration of phages reduced by three orders of magnitude (1010 to 107
PFU/ml). Magnesium hydroxide the best excipient, while saccharose had no protective effect.
The pharmacokinetic assay was designed with the following conditions: a phage cocktail (with a
titer up to 1010 PFU/ml) was dispensed into the drinking water of 22-day old broilers (n=20). Four
treatments were applied: one negative control group with saline solution without phages (n=5);
another negative control group with the most protective excipient and without phages (n=5);
phages in saline solution with no protective excipient (n=5); and phages with the most protective
excipient (n=5). Blood samples were taken at 0, 2, 4, 8, 24 and 48 hours from the beginning of the
treatment. Fecal samples were taken at 0, 2, 4, 8, 24 and 48 hours after the beginning of the
treatment. Phage titers in these samples were measured by quantitative PCR using a single phage as
a marker for the cocktail. The purpose of this study was to improve our understanding of the
practicalities of phage treatment in a real scenario to advance phage therapy in the industry.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
164 Sections VII: Phage Therapy
Characterization of lytic bacteriophages active against drug resistant clinical
isolates of Pseudomonas aeruginosa
Kirill Sergueev 1, Nathan Brown 2, Jenny He 1, Akhil Reddy 1, Mikeljon Nikolich 1, and Andrey
Filippov 1 *
1 Department of Bacteriophage Therapeutics, Bacterial Diseases Branch, Walter Reed Army
Institute of Research, 503 Robert Grant Ave., Silver Spring, MD, USA; 2 Department of Infection,
Immunity and Inflammation, University of Leicester, Leicester, United Kingdom.
*E-mail: [email protected]
Pseudomonas aeruginosa is a frequent and important cause of multiple infections, with common
multidrug resistance and robust biofilm formation. Expanding antibiotic resistance limits
therapeutic options and necessitates the search for alternative antibacterials including lytic
bacteriophages (phages). Multiple publications have shown high efficiency of phage therapy
against experimental and veterinary P. aeruginosa infections. Since P. aeruginosa phages have
limited host ranges, the recommended approach is to use phage cocktails to cover the diversity of
clinical isolates. Rational formulation will require the use of phages that belong to different
families and bind to different receptors to prevent phage resistance.
The objective of this work was to isolate and characterize a diversity panel of potentially
therapeutic phages active against P. aeruginosa. Phages were isolated from sewage and
environmental waters, using the cultures of several clinical isolates of P. aeruginosa for
enrichment. The phages were first screened based on plaque morphology and host ranges
determined on a small number of strains, and the final screening was done using DNA isolation and
digestion with EcoRV. This allowed for the identification of 30 new different bacteriophages lytic
for P. aeruginosa. Based on full genome sequencing and electron microscopy, the phages belong to
five different phylogenetic groups. The genome sizes varied from 32 to 89 kb. Host range testing
on 52 multidrug resistant strains of P. aeruginosa mostly isolated in military hospitals revealed
lytic spectra ranging from 11% to 71%. Overall, 93% of strains were susceptible to one or more
phages from the panel. Three phages demonstrated a marked lytic activity against P. aeruginosa
PAO1 biofilms. Therefore, several of the 30 diverse lytic P. aeruginosa phages we isolated and
characterized are promising therapeutic candidates.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 165
Bacteriophage engineering for therapeutics
Aurelija Grigonyte, Andrew Millard*, Alfonso Jaramillo
University of Warwick, UK
*E-mail: [email protected]
Bacteriophages are being increasingly explored as an alternative to antibiotics. The ability to
engineer bacteriophages will allow advantageous alterations of their phenotype, e.g., increased host
range, making them more pertinent as therapeutics.
In this study, we show how marker-based (trxA) as well as marker-less (CRISPR-Cas9) selection
can be used for engineering using bacteriophage T7 as a model system. Here we generated a
chimeric phage for which T7 was used as the main scaffold and appended with a tail fiber from
BBP-1 phage.
We have showed that the marker based method is more suitable for the tail fiber engineering. In
developing this system, we also observed co-infection between T7 and engineered phage, where
both wild type and engineered phage are required for a successful infection (figure below). The
idea of co-infection challenges the T7 exclusion principle previously delineated in the literature.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
166 Sections VII: Phage Therapy
Hypersensitivity to phages is a rare phenomenon
Karolina Wojtyna*, Zuzanna Kaźmierczak, Joanna Majewska, Dorota Lecion, Anna Kłopot,
Weronika Kęska, Paulina Miernikiewicz, Barbara Owczarek, Marek Harhala, Andrzej Górski,
Krystyna Dąbrowska
Institute of Immunology and Experimental Therapy, Polish Academy of Sciences, ul. R. Weigla
12, 53-114 Wroclaw, Poland
*E-mail: [email protected]
Phages can induce specific antibody production in mammals. According to the current state of
knowledge there are no reports documenting hypersensitivity to phages, suggesting that it is not
common. However, hypersensitivity to phages, if present, could have a serious impact not only on
phage therapy, but on all other medical applications of phages.
We investigated T4 as a model phage and 676Z, A3R, F8 - therapeutic phages from the Phage
Therapy Unit. First, we examined human sera to determine presence or absence of phage-specific
IgE in population. Sera were obtained from 50 healthy volunteers who had neither been subjected
to phage therapy nor involved in phage work. Second experiment was the evaluation of phage-
specific IgE level in sera of mice exposed to phages. Mice were challenged orally (4 x 109 pfu/ml
in drinking water continuously for 100 days) or intraperitoneally (1010 pfu, injections were repeated
after 20 and 50 days). Sera were collected at following time points: day 1, 10, 50 and 100 after first
immunization.
To analyze hypersensitivity-promoting conditions, we used oral mice allergy model. Mice were
injected subcutaneously with highly purified phage preparation in dose 5 x 109 pfu with an addition
of Al(OH)3 as an adjuvant promoting hypersensitivity reaction. The injection was repeated after 14
days. Seven days after the second sensitization mice were given phage preparations in drinking
water in dose 5 x 109 pfu/ml. IgE levels specific for tested phages in all experiments were evaluated
by ELISA.
We did not observe high level of phage-specific IgE in any of examined samples. In human
population only one sample (a person with other symptoms of general hypersensitivity problems)
had detectable level of phage-specific IgE. No increase of IgE production in mice exposed to
phages in normal conditions was observed. In oral allergy model only one group of animals (676Z)
responded with a low increase of specific IgE level. Other groups did not respond in IgE level,
even though the adjuvant was applied.
These results demonstrate that hypersensitivity to phages is a rare phenomenon. In our study, it
appeared in very specific, allergy-inducing conditions. This reaction also seemed to depend on
individual phage properties. However, the exact mechanism of this reaction still remains unknown.
Project supported by Wroclaw Centre of Biotechnology, programme The Leading National
Research Centre (KNOW) for years 2014-2018.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 167
Real-time biodistribution imaging of intravenous and intraperitoneal
Pseudomonas aeruginosa bacteriophage using molecular labeling
Derek Holman1, Matthew P. Lungren2, Jonathan Hardy3, Chris Contag1,2, 3, Francis Blankenberg2*
1Department of Pediatrics, Division of Medical Genetics and Biochemistry,
Stanford University School of Medicine, Stanford, CA 94305, USA, 2Department of Radiology, Division of Pediatric Radiology and Nuclear
Medicine, Lucile Packard Children’s Hospital, Stanford, CA 94305, USA, 3Department of Microbiology and Immunology, E150 Clark Center MC 5427,
Stanford University School of Medicine, Stanford, CA 94305, USA
*E-mail: [email protected]
Bacteriophages are ubiquitous viruses which have adapted to infect and replicate within target
bacteria, their only known hosts, in a strain specific fashion with little to no cross infectivity. The
recent steep rise in antibiotic resistance throughout the world has renewed interest in adapting
bacteriophages for the imaging and treatment of bacterial infection in humans.
In order to develop a method with which to image, in real-time, the biodistribution of phage
treatments via radiolabeling, we examined the effects of hydrazinonicotinamide (HYNIC)
conjugation and removal of bacterial DNA on the infectivity, biodistribution, and radionuclide
imaging of a Pbunavirus bacteriophage lytic for a clinically relevant strain of Pseudomonas
aeruginosa, a common gram negative bacterial pathogen often resistant to multiple antibiotics.
We found all but the briefest reaction of concentrated bacteriophage with HYNIC (≤ 3 minutes)
resulted in nearly complete loss of infectivity. Furthermore, we determined that digestion and
removal of bacterial DNA was needed to avoid high non-specific uptake of HYNIC-labeled
bacteriophage within the liver and spleen as well as prolonged circulation in the blood. We also
demonstrated the surprisingly wide soft tissue and organ biodistribution and rapid
pharmacokinetics of Tc99m-HYNIC-labeled bacteriophage in normal mice as well as its imaging
characteristics and efficacy in wounded mice infected with bioluminescent Pseudomonas
aeruginosa. In conclusion, the preservation of bacteriophage infectivity and removal of all bacterial
containments including DNA are critical methodologic considerations in the labeling of
bacteriophages for imaging and therapy.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
168 Sections VII: Phage Therapy
Assessment of bacteriophage cocktails against MDR biofilms Anna C. Jacobs1*, James M. Regeimbal2, Luis A. Estrella3, Javier Quinones3, Matthew S. Henry3,
Nathan A. Franklin4, Samandra Demons1, Biswajit Biswas3, Joseph C. Wenke4, Theron C.
Hamilton3, Michael G. Stockelman2, Stuart D. Tyner1.
1Walter Reed Army Institute of Research, Silver Spring, MD; 2Naval Medical Research Center,
Silver Spring, MD; 3Naval Medical Research Center-Frederick, Fort Detrick, MD; 4U.S. Army
Institute of Surgical Research, San Antonio, TX.
*E-mail: [email protected]
Objectives: Some of the most challenging infections among war wounded are those caused by
biofilm-producing, multidrug resistant bacteria (MDR), which may require extended antibiotic
treatment and can result in treatment failure. Our research is focused on developing phage cocktails
to target complex MDR infections, including those associated with implants or orthopedic
hardware. We have developed a pipeline for the isolation, purification, and pre-clinical assessment
of phage cocktails against P. aeruginosa and S. aureus in preclinical trauma models. Our main
objective is to develop therapeutic phage cocktails that can be used in conjunction with antibiotics
to treat complex wound infections.
Methods: Phages were isolated from raw sewage water, and phage cocktails were compounded
using high throughput phage lysis assay. Phage combinations that resulted in synergistic bacterial
killing were further assessed in in vitro assays, including biofilm assays to determine if phages
could penetrate and destroy mature biofilms, and checkerboard assays to evaluate phages in
combination with antibiotics. Phage cocktails were also evaluated in in vivo mouse and rat wound
models. The measured outcomes of these animal models included weight loss post-surgery, clinical
signs of infection, bacterial burden as determined by bioluminescence signal, CFU, wound size,
and healing time.
Results: In vitro biofilm assays of P. aeruginosa or S. aureus showed that the phage cocktails
could reduce established biofilm by at least 50% based on crystal violet staining. In a proof-of-
concept mouse dorsal full-thickness cutaneous wound model, S. aureus-infected, phage-treated
mice had significantly smaller wounds than the control-treated mice (P ≤ 0.01) through day 10
post-infection, with a median area of 44 mm2 compared to 67 mm2. A preliminary study assessing a
S. aureus phage cocktail in a clinically-relevant rat femur defect model resulted in complete
clearance of bacteria from bone and hardware of several of the phage-treated rats, while all control-
treated rats had a bacterial burden of 106 CFU in bone and hardware.
Conclusions: Bacteriophages represent a novel therapeutic option for recalcitrant, MDR bacterial
infections. Our preliminary results suggest that phage cocktails can reduce biofilm and bacterial
burden in vitro and in in vivo animal models. Studies are ongoing to further evaluate both P.
aeruginosa and S. aureus alone and in the presence of antibiotics.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 169
Novel Klebsiella bacteriophages to control the spread of
Klebsiella pneumoniae carbapenemase-producing Klebsiella pneumonia
in hospital setting
Jongsoo Jeon*, Dongeun Yong, Kyungwon Lee
Department of Laboratory Medicine and Research Institute of Bacterial Resistance,
Yonsei University College of Medicine, Seoul, Korea
*E-mail: [email protected]
Objectives: In recent years, antimicrobial resistance has become a major medical threat
worldwide. Among them the rapid increase in Klebsiella pneumoniae carbapenemase (KPC)-
producing Klebsiella pneumoniae is one of the particular global issues challenging the health care
setting. In order to overcome the problems, bacteriophages have been newly reviewed as an
alternative strategy for the control of these pathogens.
Methods: Fifty-eight KPC-K. pneumoniae clinical strains were collected from a university hospital
in South Korea and these strains were used to isolate lytic phages from sewage water samples. Host
ranges of isolated phages were determined by the spot test and these phages were characterized by
transmission electron microscopy, one-step growth curve, adsorption rate, thermal/pH stability
analysis and host cell lysis test.
Results: In this study, we isolated and purified 4 K. pneumoniae phages (Bϕ-U2874, Bϕ-N54, Bϕ-
N98, Bϕ-N137), which cause lysis of KPC-K. pneumoniae strains, from sewage water at a hospital
in South Korea and these phages were characterized in detail. In morphological properties of
phages by TEM, all K. pneumoniae phages belong to the family Siphoviridae with an isometric
head and a long flexible tail. These phages exhibited a very broad spectrum against clinical KPC-K.
pneumoniae isolates used in this study, but they did not lyse other bacterial strains including Gram-
negative or positive strains. These phages showed high absorption rate > 75-99% within 5 min and
bust size of 24-74 PFU/cell. Most of them exhibited stability at 40-50°C and pH 4-10. All phages
showed strong host cell lytic activities at MOI=10.
Conclusions: Overall, several in vitro results revealed that 4 novel K. pneumoniae phages have a
potential as an antimicrobial agent to control the spread of KPC-K. pneumoniae strains in clinical
settings and can be considered as therapeutic agents for infectious diseases caused by KPC-K.
pneumoniae strains. Further studies will be performed on next-generation sequencing (NGS) of
selected phages and in vivo test for efficacy and safety of phages by using mouse infection model.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
170 Sections VII: Phage Therapy
V. cholerae O1 phages: effective subtyping and potential therapeutic mean for
cholera outbreaks
T. Kokashvili1, N. Janelidze1, T. Koberidze1, K. Porchkhidze1, G.Tsertsvadze1, A. Huq2, and M.
Tediashvili1
1 George Eliava Institute of Bacteriophages, Microbiology and Virology, Tbilisi, Georgia; 2 Maryland Pathogen Research Institute, University of Maryland, College Park, MD, USA
The disease cholera, caused by highly variable waterborne bacterium V. cholerae O1, remains a
serious public health concern worldwide, especially in the developing countries with warm climate.
The global climate change increases the possibility of its emergence in the non-endemic regions.
Thus, development of effective means of detection, identification and subtyping of V. cholerae,
also for prevention and treatment of cholera outbreaks, is of high-priority. Specific bacteriophages
can be used for wider phenotypic diversification and tracking of V. cholera isolates in the
environment, and also to prevent and treat infections, caused by antibiotic resistant V. cholerae
strains.
The strain collections of the Eliava Institute, Georgia, and Maryland Pathogen Research Institute,
UMD, USA, were screened to determine the susceptibility to V. cholerae – specific phages,
isolated in Georgia. The clinical and environmental strains of V. cholerae O1 (382 strains in total)
and 16 strains of V. cholerae O139, collected in different geographical regions (Georgia, South
East Asia, South America, Haiti, West Africa, etc,) were used in the study along with the set of 14
Eliava phages specific to V. cholerae O1 and non- O1/non139.
These phages were characterized by virion morphology, serologic properties and DNA restriction
profiles. The phage lytic spectrum was determined by spot test technique followed by estimation of
the phage titer on selected bacterial strains. The strains of V. cholerae O1, obtained from patients,
appeared to be more susceptible to the Eliava phage set: in average 95,2% of strains were lysed by
at least one phage. Some differences in the phage susceptibility pattern was observed for particular
groups of V. cholerae O1 isolates: phages lysed all Haiti strains, 95% of Bangladeshi strains, and
91 % of isolates from different regions.
The Georgian environmental isolates of V. cholerae O1 showed less susceptibility (77 %) to V.
cholerae phages that can be explained by high genetic and phenotypic variability of environmental
V. cholerae strains. The susceptibility patterns to eight phages, selected for the phage-typing set,
allowed to divide clinical V. cholerae O1 isolates into a number of phage groups, with the number
of strains in each group depending on strain collection and geographical origin.
The results obtained have confirmed high distinctive features of specific bacteriophages for
subtyping of V. cholerae isolates of different origin as well as obvious potential of lytic phages for
future therapeutic applications in cholera outbreaks.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 171
Phage Typing, Antibiotic Resistance and Genomic Rep-PCR Fingerprinting
of Clinical Salmonella Isolates from the Caucasus Region
Elene Kakabadze,1* Khatuna Makalatia,1 Maia Merabishvili,1,2 Nino Grdzelishvili,1 Nata
Bakuradze,1 Mkhtar Mkrtchyan,3 Marina Goderdzishvili,1 Jean-Paul Pirnay,2 Nina Chanishvili,1
Anahid Sedrykyan3 1George Eliava Institute of Bacteriophage, Microbiology and Virology, Tbilisi, Georgia; 2Queen
Astrid Military Hospital, Brussels, Belgium; 3Institute of Molecular Biology, Academy of
Sciences, Yerevan, Armenia
*E-mail: [email protected]
Fluoroquinolone-resistant Salmonella infections are emerging to cause considerable
morbidity and mortality worldwide. In reaction to the use of antibiotics in farming and veterinary
practices, MDR phenotypes have emerged. As Salmonella enterica serovars are widely diverse and
spread through the food chain, rapid increase in antibiotic resistance is also observed in clinical
strains. Therefore, rapid identification and treatment solutions are direly needed to treat MDR
infections. The use of lytic phage-based systems offers a triple benefit: rapid and cost effective
typing using therapeutic agents, which can potentially lyse the examined MDR strains.
In this study we have focused on isolates from patients with Salmonellosis, who have not
yet received any antibiotic therapy. In total, 71 non-typhoid Salmonella enterica clinical isolates
from the “Nork” Clinical Hospital of Infectious Diseases (Yerevan, Armenia) and 22 strains from
the Referral Laboratory at the Republican Clinical Hospital of Infectious Diseases (RCHID, Tbilisi,
Georgia) were selected.
Isolates were maintained on Salmonella-Shigella selective media. Serotyping by polyvalent
antisera for flagellar (H) and lipopolysaccharide (O) antigens, and by biochemical analysis, were
performed in the respective clinical settings. MALDI-TOF analysis was performed to confirm the
identification at the genus level. Semi-automated rep-PCR fingerprinting profiles were determined
using the DiversiLab® system and analyzed using Pearson’s correlation analysis. An UPGMA
dendrogram was constructed and strains showing a similarity value above 95% were marked as
linked (cluster). The thus obtained clusters were compared with Salmonella group B, C, D, E, F, G,
K, L, O, R reference libraries for serotype identification. Antibiotic resistance profiles (different
classes of antibiotics) were determined in accordance with the CLSI guidelines for standard disk
diffusion assays. Phage typing was performed using the line spot-test method. For this purpose, 5
multivalent (active against Salmonella and Escherichia coli), 3 phage mixtures and 7 Salmonella
specific phage lines were selected.
Species identification by phage typing confirmed the MALDI-TOF identifications. Forty-
one percent of the tested clinical isolates were shown to be MDR, while none of the strains showed
complete resistance to the tested phages. Despite of their resistance to different classes of
antibiotics, all isolates were susceptible to at least one of the phage lines (clones) used in our typing
system. According to rep-PCR fingerprinting, 93 examined isolates clustered into 24 different
cluster-serotypes. One isolate from RCHID showed 97.3% similarity with a Salmonella Dublin
reference strain, one cluster harbored 14 strains with >95% similarity to a Salmonella typhimurium
reference strain and another cluster consisted of 21 strains identified as Salmonella enteritidis. As
expected, no correlation was observed between strain genotypes (rep-PCR fingerprints) and phage
types. A robust lytic phage-based characterization system can be relevant as a tool for
epidemiological studies and as a rapid screening method for therapeutic or prophylactic phage
applications in food, veterinary or human clinical settings.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
172 Sections VII: Phage Therapy
Drosophila melanogaster as a model to study host-phage infection, immunity
and interactions with the microbiome
Thomas Leech*, Amanda Bretman
University of Leeds, England
*E-mail: [email protected]
Bacteriophage therapy has long promised an enticing alternative to antibiotics in the treatment of
bacterial infections, but over 100 years since the discovery of bacteriophages, they are still yet to
realise their therapeutic potential. A number of aspects of bacteriophage biology have hindered
progress, amongst which the relative dearth of information available regarding phage-host
immunology and the unintended off-target effects of bacteriophage infection on host microbes, are
key.
In order to better understand the complex relationship between bacteriophage infection, the host
immune response and the microbiome, a suitable model is required. The fruit fly Drosophila
melanogaster is a useful and well established model of innate immunity and the microbiome due to
its genetic tractability, ease of housekeeping and the conserved nature of many key genes and
proteins, as well as it’s relatively simple resident bacterial community.
Here, we demonstrate the significant potential that D. melanogaster has in helping to elucidate
specific aspects of phage biology such as immune inactivation of therapeutic phages,
immunological sensitisation by phage infection and perturbation of the host microbiome structure
by potential therapeutic bacteriophages. D. melanogaster may become a powerful model in the
advancement of phage therapy.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 173
Evaluation of phages cocktails for inhibition of V. parahaemolyticus in
aquaculture
Carlos Lomelí-Ortega; Sergio Martínez-Díaz
Microbiology Laboratory, IPN-CICIMAR, La Paz, B.C.S., 23096, México.
E-mail: [email protected]
Vibrio parahaemolitycus is an important cause of disease, mortality, and economical losses in the
shrimp aquaculture industry, and in recent years has been associated to early mortality syndrome
(EMS), which is a new disease that has been detected at shrimp farms in Asia since 2010. New
therapies are necessary for control of this kind of diseases, and phage therapy is an alternative
prophylaxis treatment of bacterial infectious diseases that has gained interest. Phage therapy is the
use of naturally occurring viral predators of bacteria, and are recognized for their ability to reduce
the pathogen populations.
In this study, we evaluated individual effect of 12 phages against V. parahaemolyticus, and select
the best cocktail for biocontrol in artemia culture. A single application of each phage inhibited
growth of V. parahaemolyticus even 24h post application (p<0.05). Dual applications presented
interference between phages, with phages A3S, CVH4, and 4P the worst phages for combination
use.
On the other hand, phages VH5, SH, SP, PL14D, and VPMS1 were the best phages for dual
combination with themselves. A cocktail of VH5, SP, PL14D, and VPMS1 in equal proportion was
the best combination against V. parahaemolyticus, reducing growth in 73 % ± 0.9 during 24h post
application of a single dose. The cocktail was evaluated against four EMS positive strains and was
able to stop growth during 5h post application. A significant difference of 15 % was seen at the end
of the test (p<0.05). The next step will be the evaluation of activity in an in vivo model with
Artemia, to determinate the best conditions for application.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
174 Sections VII: Phage Therapy
Humoral response to staphylococcal bacteriophages applied per os in
a murine model
Joanna Majewska, Zuzanna Kaźmierczak, Karolina Wojtyna, Dorota Lecion, Katarzyna Hodyra-
Stefaniak, Anna Kłopot, Barbara Owczarek, Marek Harhala, Paulina Miernikiewicz, Andrzej
Górski, Krystyna Dąbrowska
Hirszfeld Institute of Immunology and Experimental Therapy, Polish Academy of Sciences,
R. Weigla 12, 53-114 Wrocław, Poland
*E-mail: [email protected]
Objectives: In our previous studies, we showed that a model bacteriophage T4 applied orally over
a long period of time induced production of anti-phage antibodies in mice, both in the gut and in
the blood. Here we investigated a similar model in which two staphylococcal bacteriophages: A3R
and 676Z were applied per os.
Methods: C57BL/6J mice were given purified phage preparations in PBS, mixed with drinking
water (1:1) to a final concentration of 4×109 pfu/ml continuously for 100 days. Control group was
given drinking water with PBS. Phage treatment was followed by 130 days without the phages and
subsequent second continuous treatment with staphylococcal phages until the experiment was
terminated. Throughout the whole experiment blood and fecal samples were collected and
evaluated for the presence of viable phage particles and anti-phage antibodies (IgG, IgA, IgM).
Antibody levels were measured by ELISA immunoassay.
Results: Phages were detectable in the blood in small titers only during the first week of the
treatment. On the contrary, high phage titers (107-6×109 pfu/g) were simultaneously observed in
fecal samples, decreasing gradually to reach an approximate value of 102 and 103 pfu/g (for A3R
and 676Z, respectively) after 5 weeks and eventually decreasing to undetectable levels at the end of
the first phage treatment. Second treatment resulted in viable phage particles being again detectable
in feces. However, phage titers were significantly lower (5,7×102 for A3R; 2,1×103 for 676Z) than
in the initial phase of the experiment and decreased to undetectable levels within 25-38 days.
Decrease of phage titers in faeces correlated with an increase of IgA, but not IgG. Plasma IgA
levels started to increase 4-5 weeks after the initiation of phage treatment, reached its maximum
within 72-85 days and then gradually decreased after phage treatment was terminated. However,
second administration of phage preparations induced IgA production within 10 days. A marked
increase in plasma IgG was observed as soon as two weeks following the initiation of the
treatment. IgG levels then continued to rise until day 36 and – contrary to IgA – remained at high
concentration throughout the whole experiment. IgM peak was observed on day 15.
Conclusions: These data show that staphylococcal bacteriophages applied per os induced humoral
response in vivo, interestingly, with a lower contribution of primary response, but with a significant
level of response in IgG and IgA. Continuous oral treatment with phage preparations and the
induction of humoral responses resulted in a decreased phage passage through the murine
gastrointestinal tract. Increase in IgA levels correlates with a decrease of phage titers in feces.
Acknowledgements: This work was supported by the National Science Centre in Poland grant
UMO-2012/05/E/NZ6/03314 and by the Wroclaw Centre of Biotechnology, programme The
Leading National Research Centre (KNOW) for years 2014–2018.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 175
Formulation and Encapsulation of Bacteriophages for Targeted Controlled
Release
Danish J. Malik1*, Goran T. Vladisavljević1, Martha R. J. Clokie2, Gurinder Vinner1, Salvatore
Cinquerrui1, Francesco Mancuso1
1Chemical Engineering Department, Loughborough University, Loughborough, UK; 2 Department
of Infection Immunity and Inflammation, University of Leicester, Leicester, UK.
*E-mail: [email protected]
Objectives: The aim of the work was to evaluate the use of novel microfluidic and membrane
based techniques for encapsulation of purified (using ultrafiltration and ion exchange)
bacteriophages in stimuli responsive polymers (microparticles) and liposomes (microparticles and
nanoparticles) for treating multi-drug resistant gastrointestinal infections.
Methods: Bacteriophage K (lytic, S. aureus, myovirus) and Felix O1 (lytic, S. enterica, myovirus)
were used in the study. Phages were amplified using a Sarorius Biostat® operated in batch mode.
Phages were purified using centrifugation, ultrafiltration, anion exchange chromatography and size
exclusion chromatography to remove host cell proteins and host cell DNA. Encapsulation of SYBR
green labelled phage in micro- and nanoparticles was shown using confocal microscopy. TEM
imaging of encapsulated phage in liposomes was carried out. Phage stability upon exposure
(exposure time 3hr at pH 2) to simulated gastric fluid and subsequent phage release kinetics was
studied using simulated intestinal fluid (phage titre measured using plaque assay). The effects of
formulation (e.g. polymer or lipid composition), phage titre and experimental setup conditions were
investigated on phage encapsulation, phage viability and storage stability. The phages were also
spray dried and freeze dried using excipients to improve long-term storage stability.
Results: We show that we can achieve significant concentration of highly purified phages (~1012
PFU/ml) using downstream processing operations by removing host cell proteins and host cell
DNA. We have proof-of-principle results demonstrating controlled encapsulation of phages in pH
responsive polymeric microparticles and liposomal nanoparticles using microfluidic and membrane
encapsulation techniques. Our approach allowed precise control over phage loading per particle
(i.e. control over phage dose). We show how scalable production of highly uniform micro- and
nanoparticles may be achieved using microfluidic and membrane emulsification with control over
particle size through manipulation of microfluidic hydrodynamic conditions, membrane shear and
fluid properties (e.g. viscosity). Through changes to the formulation and the particle size, we also
demonstrate control over the release dynamics (burst and sustained release formulations) of the
phages in response to a pH or enzyme trigger. We demonstrate that encapsulated phages may be
stored under refrigerated conditions following drying by addition of disaccharides such as trehalose
as excipients.
Conclusions: We have demonstrated that novel microfluidic and membrane based processes can
be successfully used to encapsulate phages in stimuli responsive micro- and nanoparticles. These
systems would allow delivery of phages to the gastrointestinal tract giving unprecedented control
over the phage dose and the release profile to treat multi-drug resistant infections.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
176 Sections VII: Phage Therapy
Identification of the Type IV Pilus as the Receptor for Bacteriophage DLP1
Adherence to Stenotrophomonas maltophilia and Pseudomonas aeruginosa
Jaclyn G. McCutcheon, Danielle L. Peters and Jonathan J. Dennis*
University of Alberta, Edmonton, Alberta Canada
*E-mail: [email protected]
The opportunistic bacterial pathogen Pseudomonas aeruginosa is a major cause of nosocomial
infections, particularly in immunocompromised individuals, and is the second most prevalent
pathogen found in the lungs of cystic fibrosis patients. The inability to control P. aeruginosa
infections with antibiotics due to its intrinsic and adaptive multi-drug resistance and range of
virulence factors is cause for concern. A possible alternative treatment is phage therapy, the clinical
application of bacteriophages to eradicate target bacteria. While bacteriophages typically have a
very narrow host range, a novel phage DLP1 previously isolated in our lab is capable of infecting
across taxonomic orders, lysing strains of both P. aeruginosa and Stenotrophomonas maltophilia,
another highly antibiotic resistant bacterial pathogen. To explain this unique ability, we have
identified and are characterizing the primary receptor for DLP1 and the mechanism of attachment
to both host species.
Initial screening of a 2,242 member P. aeruginosa PA01 mutant library suggested that DLP1 uses
the type IV pilus, a virulence factor on the surface of the bacterium, as its primary receptor for
adherence. Results show that mutants with transposon insertions in genes encoding pilus structural
and regulatory components are resistant to DLP1 infection. Subsequent complementation of the
disrupted pilin subunit genes in PA01 restored DLP1 adherence and lysing ability. Preliminary data
from a cross-species complementation experiment using appropriate S. maltophilia D1585 pili
genes in a PA01 pili deficient background also restored phage binding and lysis, suggesting that a
similar pili structure serves as the receptor for DLP1 in the main S. maltophilia host strain, D1585.
Visualization using electron microscopy shows that DLP1 interacts with pilus structures on the
surface of PA01 and D1585. When the host bacteria is grown in media containing mucin, a major
component of mucus and a nutrient that facilitates colonization of the lungs by pathogenic bacteria,
the bacteria show increased twitching motility indicative of induced pilin expression. This increase
in pilin expression enhances the association of DLP1 to its host bacteria, visualized by transmission
electron microscopy.
Future research will examine mutagenesis of pilin associated genes in the main S. maltophilia host
strain, D1585, with the prediction that this will knockout DLP1’s ability to adhere to and lyse the
cells. This research will further characterize DLP1 as a candidate for phage therapy in the treatment
of highly antibiotic resistant bacteria from multiple genera.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 177
Determination of safety and therapeutic potential of environmental waste
water lytic bacteriophages against multidrug resistant Staphylococcus aureus
(MDRSA) in BALB/c mice
Oduor Joseph M. Ochieng’1, 2, 3*, Onkoba Nyamongo 4, Maloba Fredrick 5, Arodi Washington
Ouma 6, Atunga Nyachieo 1
1Institute of Primate Research (IPR), Nairobi-Kenya, 2 KAVI-Institute of Clinical Research,
University of Nairobi-Kenya, 3Haartman Institute, University of Helsinki-Finland, 4Technical
University-Kenya, 5 School of Applied Pure Science, Kenyatta University, Nairobi-Kenya, 6 School
of Medicine, Kenyatta University, Nairobi- Kenya
*E-mail: [email protected]
The use of phage therapy as an alternative method of treating infections caused by multidrug
resistant bacteria is gaining ground as most bacteria are becoming resistance against the commonly
available antimicrobials.
Objective: The study sought to determine safety and therapeutic efficacy of environmentally
obtained Staphylococcus aureus lytic phages against multidrug resistant S. aureus (MDRSA) in
mice.
Methodology: Phages and MDRSA were isolated from sewage and waste water collected from
within Nairobi County. The isolated S. aureus bacteria were screened for resistance towards;
ceftazidime, oxacillin, vancomycin, netilmicin, gentamicin and erythromycin, trimethroprim-
sulfamethoxazole and cefuroxime. Thirty BALB/c mice were randomly assigned into three groups;
the MDRSA infection group (n=20), the phage-infection group (n=5) and non-infection group
(n=5). After 24 or 72h post-infection (p.i.) with MDRSA, the infected mice were either treated with
a single dose of clindamycin (8mg/kg/bwt) or 108 PFU/ml of S. aureus phage or a combination
treatment (clindamycin and S. aureus phage). Efficacy and safety of the treatments were
determined by monitoring animal physical health post-infection as well as gross pathology and
histopathology. Bacteremia was determined daily for 10 days and used to establish therapeutic
efficacy of the phage.
Results: Treatment with phage was efficacious (100%) compared to clindamycin (62.25%) at 24h
p.i and 87.5% at 72h p.i., and the combination therapy, 75% at 24h p.i. and 100% at 72hrs p.i.
Efficacy of the treatment regimens was dependent on the time of treatment post infection. The mice
infected with MDRSA and treated with phage had no bacteremia at day 7 post-treatment compared
to those treated with clindamycin and combination therapy (P < 0.001). There were no tissue
abscesses, inflammation in the brain, lungs and liver tissues of phage treated mice compared to
those treated with clindamycin and combination therapy.
Conclusion: Staphylococcus aureus phage obtained from sewage and waste water from within
Nairobi County was safe and possessed therapeutic efficacy against MDRSA bacterium.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
178 Sections VII: Phage Therapy
Characterization of Pseudomonas aeruginosa phage-resistant clones obtained
by controlled phage infection
Tomasz Olszak1, Pawel Markwitz1, Grzegorz Gula1, Miguel Valvano2 and Zuzanna Drulis-
Kawa1*
1Institute of Genetics and Microbiology, University of Wroclaw, Przybyszewskiego 63/77, 51-148
Wrocław, Poland 2Centre of Experimental Medicine, School of Medicine, Dentistry and Biomedical Sciences,
Queen’s University Belfast, 97 Lisburn Road, Belfast BT9 7BL, UK
*E-mail: [email protected]
Phage therapy is a promising strategy to combat bacterial infections, but it has also some
drawbacks. Besides difficulties in the standardization of phage preparations and regulatory issues
related to the use of phage products in humans, a major biological problem is the onset of phage
resistance upon phage treatment. The emergence of the phage-resistant mutants has been
recognized in phage therapy application. One of the simplest, effective, and widely accepted
practice of counteracting the effects of phage resistance is the use of phage cocktails containing
several phages with a similar host spectrum, but different attachment mechanism.
Most microbial surface structures recognized by phages are also important bacterial virulence
factors that strongly stimulate the immune system of infected humans. Therefore, changes in the
expression level or the structure modification of these receptors reduce bacterial virulence. In this
study, we examined the diversity of Pseudomonas aeruginosa clones obtained as a result of
controlled infection by single bacteriophages and cocktails composed of two or three
bacteriophages. Three well-characterized bacteriophages of the Caudovirales order were selected
for these experiments: phiKZ (fimbriae-dependent, giant Myoviridae) LUZ7 (fimbriae-dependent,
Podoviridae) and KTN6 (LPS-dependent, Myoviridae). Isolated bacterial mutants were tested for
twitching, swarming and swimming motility, the intensity of pyoverdine, pyocyanin and biofilm
production, the LPS patterns and the sensitivity to antibiotics, phages, complement system and
phagocytosis.
Our results indicate that the selection pressure caused by phage infection leads to modification or
even loss of important virulence factors (i.e. type IV fimbriae or LPS O-antigen). It was confirmed
that the virulence of isolated phage-resistant clones was reduced. In most cases, observed changes
remain stable, what suggests their mutational character. Nevertheless, bacterial resistance based on
the loss or modification of the particular receptor did not always result in cross-resistance to other
phages recognizing the same receptor.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 179
Characterization of novel broad-spectrum Citrobacter freundii bacteriophage
Graça Pinto, Hugo Oliveira, Ana Oliveira and Joana Azeredo*
CEB – Centre of Biological Engineering, LIBRO – Laboratório de Investigação em Biofilmes
Rosário Oliveira, University of Minho, 4710-057 Braga, Portugal
*E-mail: [email protected]
Citrobacter spp. although frequently ignored, is an emergent nosocomial bacterium able to cause
various superficial and systemic life-threatening infections. Considered to be a hard-to-treat
bacterium, due to the acquisition of resistance to a large range of antibiotics, new measures of early
and efficient therapies are needed. Bacteriophages, the natural predators of bacteria, have elicited
interest due to their high specificity, low inherent toxicity, and ability to kill antibiotic-resistant
bacteria. To assess the potential for phage therapy, we have characterized and sequenced a novel
broad host range T4virus: the myovirus vB_CfrM_CfP1, that specifically infects Citrobacter
freundii (>85% of 21 strains tested). To demonstrate its value as an antimicrobial alternative, CfP1
was fully characterized, shown to be stable at a large range of temperature and pH values, capable
of killing multidrug-resistant clinical isolates (showing resistance up to 12 antibiotics), and with a
burst size of 45 PFU/cell. Genomically, CfP1 presents a dsDNA molecule of 180,219 bp, coding
273 CDSs and sharing 46 to 94% of nucleotide identity to other Citrobacter phages. The absence
of genes with known function encoding for virulence factors, its high killing activity of suspended
cultures and the elevated stability at a wide range of pH suggest that this phage possesses suitable
properties for therapy.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
180 Sections VII: Phage Therapy
Approaches to bacteriophage therapeutics for Staphylococcus aureus
Melissa D. Ray*, Roger D. Plaut, E. Scott Stibitz
US Food and Drug Administration, Division of Bacterial, Parasitic, and Allergenic Products,
Center for Biologics Evaluation and Research, 10903 New Hampshire Ave., Silver Spring,
Maryland
E-mail: [email protected]
Staphylococcus aureus can be a commensal organism as well as a serious pathogen capable of
causing skin abscesses, wound infections, endocarditis, osteomyelitis, pneumonia, and toxic shock
syndrome. The treatment of these infections has become difficult because of the emergence of
strains resistant to multiple antibiotics. Bacteriophage therapy has received more attention recently
as an alternative approach to combat multidrug-resistant bacterial infections.
Our work involves developing a platform to investigate the genetic aspects of, and utility of,
bacteriophage therapy aimed at decolonization of methicillin-resistance S. aureus (MRSA). The
utility of S. aureus bacteriophage K and the Eliava Fersisi bacteriophage cocktail have been
assessed in a mouse model of MRSA nasal and lung carriage. We used a bioluminescent S. aureus
NRS384 derivative, SAP231, to colonize mice. Staphylococcal bacteriophage K and the Eliava
Fersisi cocktail were used for decolonization treatment. Bacteriophage K is a virulent, polyvalent
phage that lyses both coagulase-positive and -negative staphylococci. Bacteriophage K and the
Eliava Fersisi bacteriophage cocktail were shown to comprehensively lyse a panel of twenty-two
MRSA strains. Preliminary results show a significant decrease in MRSA nasal and lung carriage
using the Eliava Fersisi cocktail.
One frequently raised issue is the potential ability of bacteria to develop resistance to phage. This
study also characterizes S. aureus mutants resistant to bacteriophage K in vitro. Using S. aureus
strain NRS384, nine genetically independent spontaneous bacteriophage K resistant mutants were
isolated. Whole genome sequencing identified single-nucleotide polymorphisms (SNPs) in each of
the nine strains. Interestingly, none of SNPs are predicted to be null mutations, but all resulted in
amino acid substitutions in the affected gene products. These mutations provide insight into
potential phage receptors, replicative pathways, and other cellular components that are crucial for
phage growth. The results of these genetic screens lead to a number of testable hypotheses that we
will pursue regarding interactions between phage K and host cells.
Although there have been numerous anecdotal reports describing instances of successful phage
therapy, there is still a need for controlled clinical trials to assess its effectiveness. Overall, this
work should allow us to address important questions regarding phage therapy in a preclinical
platform.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 181
Accounting for transient or phenotypic host resistance in modeling infections
by phage cocktails in complex systems
Juliano Bertozzi-Silva1, Dominic Sauvageau1*
1Chemical and Materials Engineering, University of Alberta, Edmonton, AB, Canada
*E-mail: [email protected]
Objectives: The successful implementation of phage cocktails in applications such as phage
therapy is often impeded by the limited information available for a given system. Mathematical
models can help alleviate these issues by providing predictions for population dynamics. However,
many current models forego the impact of important factors such as the presence of organisms
competing with the host, adsorption efficiency, and, with mounting evidence, transient or
phenotypic host resistance. The aim of this study was to develop and validate experimentally a
model taking these factors into account to provide accurate information on multi-phage infections
in complex systems.
Methods: The system studied consisted of two competing Lactobacillus plantarum phages (phages
B1 and B2), the host L. plantarum, and the yeast Saccharomyces cerevisiae (included as an non-
susceptible organism competing with the host for resources). A kinetic model was developed to
describe growth, interactions and resource allocation of each population present. The model
parameters were determined experimentally through one-step growth (lysis time, lysis rate
constant, latent period, burst size), adsorption (adsorption rate constant, adsorption efficiency), and
microbial growth experiments (maximum specific growth rate, substrate affinity constant, yield).
Transient or phenotypic host resistance was accounted for by the inclusion of two reversible states,
susceptible and non-susceptible hosts, in the bacterial population. The reaction rate constant and
equilibrium constant were determined through model fitting of population dynamics in single
phage infections. Various relationships between susceptible and non-susceptible host states were
tested.
Results: The model was validated using phages B1 and B2, L. plantarum and S. cerevisiae
populations growing concurrently in a bioreactor (closed system). Not only was the model able to
describe the evolution of populations accurately, but it was also predictive. Using the model, it was
possible to predict the behaviour of the various populations based on the initial multiplicity of
infections of each phage, and the initial cell loads of bacteria and yeast. Additionally, the model
provided inferred information which is otherwise not possible to obtain in real-time, such as the
ratio of susceptible and non-susceptible host bacteria, and the individual titers of competing phages
B1 and B2.
Conclusions: The model developed, taking into account adsorption efficiency and transient or
phenotypic host resistance, provided valuable information on population dynamics, even for multi-
phage systems. The model can be applied to a variety of applications and settings where multiple
organisms and phages are present, from phage therapy and biocontrol to ecological studies.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
182 Sections VII: Phage Therapy
Characterization of four virulent bacteriophages isolated from sewage in
Waukesha Wisconsin and their use to combat Pseudomonas aeruginosa
infections
Christine Schneider*, Eric Graham, and Kimani Njoya, William C. Hutchins3
1Department of Life Sciences, Carroll University, Waukesha WI 53186; 2Penn State College of
Medicine, Hershey, PA; 3University of Wisconsin Milwaukee, Milwaukee WI
*E-mail: [email protected]
The global increase in antibiotic resistant bacteria threatens to undermine our ability to perform
many life-saving medical procedures. Pseudomonas aeruginosa is a gram-negative opportunistic
pathogen that is a significant cause of nosocomial infections as well as the most common cause of
lung infections in cystic fibrosis patients. The CDC has categorized P. aeruginosa as a serious
threat due to prevalence of multidrug resistant strains and the clinical significance of this pathogen.
Clearly, alternative strategies to combat P. aeruginosa infections are needed.
Although the interest in using virulent phages to combat bacterial infections dropped out of favor in
the United States after the development of antibiotics, there is now renewed interest in using phage
therapy to combat antibiotic resistant bacteria. Bacteriophage specific to P. aeruginosa would be of
clinical interest as an alternative to antibiotics for the treatment of infections with multidrug
resistant strains or to control biofilm formation in hospital settings to reduce the risk of nosocomial
infections. Toward that aim, we isolated four virulent phages from sewage and here we describe the
characterization of these novel bacteriophages.
To identify novel phage therapy candidates to combat P. aeruginosa infections, we attempted to
isolate bacteriophages from raw, filtered sewage using an enrichment protocol with a cystic fibrosis
clinical mucoid strain of P. aeruginosa (FRD1) and a spontaneous non-mucoid mutant (FRD2). All
four of the virulent phages described here were isolated using FRD2. Unfortunately, we were
unable to isolate phages using the mucoid strain FRD1. Host range analysis revealed that individual
phages lysed 30-40% of the seventeen P. aeruginosa strains examined with more lab strains being
lysed than clinical isolates. Interestingly, the efficiency of plating values (EOP) were typically
higher than 1 for most of the clinical isolates indicating the phage had a higher plaque forming
efficiency on the clinical strain relative to FRD2. In addition, one isolate (BB2) was able to lyse
FRD1, although with reduced efficiency relative to FRD2 (EOP=0.29). Each phage isolate was
able to inhibit the planktonic growth of FRD2 completely for at least 16 hrs, with BB2 completely
blocking growth for 24 hrs. Whole genome sequencing and EM results indicate these phages are
Myoviridae most similar to the ubiquitous and well-conserved family of PB-1 like phages.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
Sections VII: Phage Therapy 183
Phages targeting Streptococci of the oral cavity
Muireann Kate Smith1, Lorraine Draper1, Cillian Hayes1, Olivia Connolly1, Marion Dalmasso1†,
R. Paul Ross1 and Colin Hill1
1APC Microbiome Institute, University College Cork, Cork, Ireland, †Present address: Normandie
Univ, UNICAEN, ABTE, 14000 Caen, France
*E-mail: [email protected]
Streptococci are important pathogens and commensals of the mouth, skin, intestine, and upper
respiratory tract. Infections caused by these bacteria can vary in severity from mild
throat infections and those associated with tooth decay, to those that are systemic and life-
threatening. In many cases such infections are derived from pathogenic species that are the
causative agents of a number of invasive and non-invasive infections and diseases such as
Streptococcal pharyngitis, Scarlett fever, pneumonia, necrotising fasciitis and toxic shock
syndrome. In addition to known pathogens, commensal Streptococci are known to become
opportunistic pathogens should they gain access to the bloodstream from the oral cavity. Examples
of such commensals include: Streptococcus mutans, the leading cause of dental caries that affects
2.4 billion people worldwide; and Streptococcus anginosus that has been implicated in
subcutaneous abscesses, bacteraemia and infections of the upper respiratory tract.
Discovery of new and effective antimicrobial agent(s) in the current battle against antibiotic
resistance is of global concern. Thus, bacteriophages (phages) have been identified as promising
antibacterial agents that could be used to prevent or treat Streptococcal infections. In this study,
four different phages targeting clinically relevant Streptococci were isolated and characterised.
Their respective hosts were identified to be S. mutans, S. anginosus, S. pyogenes and S. infantis.
Following characterisation, two of these phages were determined to be lytic and effectively
diminished S. mutans and S. pyogenes in both planktonic and biofilm forms.
This bactericidal effect was further amplified by combining phages with the food grade bacteroicin
nisin. While both antimicrobials were found to be effective alone, an enhanced bactericidal effect
was witnessed when they were combined. Not only are these antimicrobials highly effective
against Streptococci, they offer a safe and natural means to protect against and treat bacterial
disease. Thus, making the combination of phages and nisin an attractive, practical and attainable
contribution to addressing Streptococcal infections in human health.

This abstract book is not a publication. Information can not be referenced without explicit
permission of the author(s).
184 Sections VII: Phage Therapy
Preventing Phage Resistance Through Tail Fiber Mutagensis
Kevin Yehl, Sébastien Lemire, Andrew C. Yang, Hiroki Ando, and Timothy K. Lu*
Massachusetts Institute of Technology, Cambridge, MA 02139, USA
*E-mail: [email protected]
The rapid emergence of antibiotic-resistant infections is prompting increased interest in alternative
antimicrobials. Acquisition of resistance against a new antimicrobial is a large issue in their
development. We created a high-throughput method to engineer antimicrobial bacteriophages that
achieve long-term suppression of bacterial growth and prevent resistance appearance. Analogous to
antibody specificity engineering, we direct mutations to small host range determining domains of
the phage’s tail fiber using protein structure as a guide to synthesize large (>107 different members)
“phagebody” libraries. This approach generates high functional diversity while minimizing
disruptions to the overall protein structure. We demonstrate that phagebodies isolated from such
libraries surpass natural phages at suppressing bacterial resistance appearance. We anticipate that
this approach will facilitate the creation of next-generation antimicrobials that defeat resistance,
and that it can be extended to other viral scaffolds for a broad range of applications.

NOTES

NOTES

NOTES

NOTES

NOTES

Author Index (Bold text indicates that the author is attending this meeting)
Aarestrup, Frank ................................. 122
Abas, F. ................................................ 93
Abdulkarim, M. S. ................................ 93
Abedon, Stephen ............................... 107
Adamia, Rezo ....................................... 60
Adhya, Sankar ...................................... 17
Aertsen, Abram .................................... 71
Ahern, Stephen ................................... 111
Ahmed, Rafath ..................................... 28
Akella, Srividya.................................... 33
Akter, Mahmuda .............................. 108
Alem, Farhang ...................................... 99
Alemnah, Yonas ................................... 49
Ali, Monsur .......................................... 30
Allison, Cody ..................................... 129
Altheide, Samantha ............................ 129
Alzbutas, Gediminas .......................... 121
Anany, Hany ....................................... 30
Ando, Hiroki ................................ 34, 184
Arevalo, Alejandra ............................... 29
Astudillo, Ariel ................................... 158
Attai, Hedieh ....................................... 27
Atterbury. Robert .................... 154, 159
Azeredo, Joana ................................... 42
Bai, Jaewoo ................................... 83, 86
Baig, Abiyad ............................. 154, 159
Bailey, Barbara ..................................... 53
Baker, James ........................................ 17
Baker, Kristi ......................................... 53
Bakuradze, Nata ................................. 171
Balamurugan, Sampathkumar .............. 28
Balarjishvili, N ................................... 133
Baquero, Diana ..................................... 29
Baran, Nava .......................................... 16
Baraniak, Anna ................................... 144
Barbu, Magda ..................................... 33
Barendregt, Arjan ............................... 139
Barnabas, Shaun ................................. 128
Barr, Jeremy ....................................... 53
Barrow, Paul ............................... 154, 159
Barth, Zachary ................................. 138
Batumbo, Doudou ................................. 32
Bednarek, Agnieszka .......................... 144
Bekker, Linda-Gail ............................. 128
Benesik, Martin ................................... 146
Berisio, Rita ........................................ 130
Bertozzi-Silva, Juliano ........................ 181
Bhandare, Sudhakar ................... 84, 159
Bhowmick, Tushar ........................ 25, 85
Birkholz, Nils ...................................... 140
Biswas, Biswajit ..................... 49, 62, 168
Blankenberg, Francis .......................... 167
Blasdel, Bob ........................... 60, 64, 126
Blinkova, Olga .................................... 124
Bliskovsky, Valery ................................ 17
Blumberg, Richard ................................ 53
Bolocan, Andrei ................................. 155
Bolton, D ............................................. 134
Bompangue, Didier ............................. 32
Bondy-Denomy, Joseph ...................... 43
Bonnain, Chelsea ............................... 110
Bordwell, Brianna ............................... 129
Borges, Adair ........................................ 43
Borne, Yannick ................................... 125
Breitbart, Mya ..................... 20, 110, 132
Bretman, Amanda ............................... 172
Breyne, Koen ........................................ 91
Briers, Yves................................. 126, 130
Brister, J. Rodney ............................. 124
Britton, Warwick................. 156, 157, 158
Brøndsted, Lone .................... 24, 88, 111
Brouns, Stan ........................................ 44
Brown, Nathan .............................. 95, 164
Brown, Pamela ...................................... 27
Brum, Jennifer .................................... 15
Bryan, Dan ........................................... 60
Buceta, Javier ...................................... 150
Buck, K.N ........................................... 110
Bunk, Boyke ....................................... 137
Butcher, James .................................... 147
Buttimer, Colin .................................. 125
Cadieux, Brigitte ................................... 84

Cambillau, Christian ............................ 69
Campbell, Danielle ........................... 149
Campion, Ciorsdan ............................. 155
Cano, Patricia ....................................... 77
Capolupo, Angela ............................... 139
Caprara, S. .......................................... 110
Cardona-Félix, César.......................... 101
Carlson, Michael .................................. 16
Carlson, Paul ........................................ 54
Caro-Quintero, A.................................. 90
Carrigy, Nicholas ....................... 32, 156
Carstens, Alexander ............................. 46
Casey, Aidan ...................................... 125
Casey, Pat ..................................... 50, 160
Cenens, William ................................... 71
Chan, Benjamin .................................... 32
Chan, Hak-Kim .......... 75, 156, 157, 158
Chang, Rachel ............................. 75, 156
Chanishvili, Nina ............................... 171
Chen, Ke ............................................... 75
Chen, Mandy ...................................... 129
Chitwood, Adrienne ........................... 129
Christensen, Tokala ............................ 129
Chun, Jihwan ...................................... 86
Cinquerrui, Salvatore ......................... 175
Citorik, Robert ................................... 97
Clavijo, Viviana .................................. 29
Clement, Frédéric ................................. 48
Clokie, Martha ....... 32, 52, 95, 112, 175
Coffey, Aidan ............................. 125, 134
Colavecchio, Anna ............................... 84
Colom, Joan .............................. 154, 159
Connerton, Ian ...................... 32, 147, 162
Connerton, Phillippa .......................... 162
Connolly, Olivia ................................. 183
Contag, Chris ..................................... 167
Coulson, Carollyne ............................. 129
Criss, Nichole ..................................... 129
Cucic, Stevan........................................ 30
Culley, Alexander .............................. 115
Cuypers, Maxime ................................. 65
Dabee, Smritee ................................... 128
Dąbrowska, Krystyna 74, 145, 166, 174
Dąbrowski, Kamil .............................. 144
Dalmasso, Marion ................. 50, 160, 183
Danis-Wlodarczyk, Katarzyna ........ 126
Das, Mayukh .................................. 25, 85
Dauley, Melinda .................................. 129
Davis, Christopher .............................. 129
de Crécy-Lagard, Valérie ...................... 46
de la Croix, Nick ................................... 71
De Smet, Jeroen .................................... 64
De Vos, Daniel ...................................... 32
Debarbieux, Laurent ..................... 78, 116
Dedon, Peter .......................................... 46
Degnan, Patrick ................................... 149
Del Rosario, Yvette ............................... 33
Demons, Samandra ....................... 49, 168
Dennis, Jonathan ......................... 151, 176
Devonshire, Daniel ............................. 129
Dewi, Fitriya ......................................... 32
Di Santo, James ..................................... 78
Dies, Marta .......................................... 150
Dirks, Clarissa ..................................... 129
Dishaw, Larry ....................................... 20
Doan, Matthew ...................................... 62
Dobbs, Kyle ........................................ 129
Donado, Pilar ........................................ 29
Donovan, David ................................... 98
Dopitova, Radka ................................. 146
Doskar, Jiri .......................................... 146
Draper, Lorraine ................. 50, 160, 183
Drulis-Kawa, Zuzanna ..... 126, 130, 178
Durham, Jacob .................................... 129
Dutilh, Bas ........................................... 18
Edner, N.M............................................ 44
Egilmez, Halil ..................................... 112
Eldougdoug, Noha ................................ 30
ElRakaiby, Marwa ............................ 161
El-Shibiny, Ayman ...................... 32, 162
England, Whitney ............................... 118
Espy, Michael ..................................... 129
Estrella, Luis ................................. 62, 168
Etobayeva, Irina .................................. 99
Evans, Charlotte .................................... 95
Evans, Gary ......................................... 141
Fagerlund, Robert ............................... 139
Fane, Bentley ...................................... 132

Farfan-Esquivel, Juan ..................... 163
Farlow, J ............................................. 133
Fellouse, Fred.................................... 100
Fernández, Lucía ............................ 87, 89
Ficoseco, María ................................. 123
Fieseler, Lars ...................................... 125
Figlar-Barnes, Jarred .......................... 129
Filip, Carlos .......................................... 30
Filippov, Andrey ........................ 133, 164
Fineran, Peter ............. 21, 139, 140, 141
Finlay, Warren.................................... 156
Fish, Randy ......................................... 60
Flint, Annika ...................................... 147
Forde, Amanda ................................... 155
Fortuna, Wojciech ................................ 57
Franklin, Nathan ................................. 168
Fredrick, Maloba ................................ 177
Froissart, Remy .................................. 128
Funderburk, Parke .............................. 129
Furrer, Evan........................................ 129
Gage, Brianna ..................................... 129
Gallagher, Tara ................................... 118
Galtier, Matthieu ................................ 116
Galyov, Ed ......................................... 112
Gambino, Michela ........................ 88, 111
Gamieldien, Hoyam ........................... 128
Gammage, A. ....................................... 77
Ganichev, D.A. ..................................... 59
García, Pilar.................................. 87, 89
Gasiūnas, Giedrius ............................. 121
Gawor, Jan.......................................... 144
Gencay, Y. Emre .......................... 24, 88
Gilberts, Daniel .................................. 129
Gill, Jason ................................... 47, 131
Glassbrook, Andrew ........................... 129
Głowacka, Aleksandra ....................... 144
Gniadkowski, Marek .......................... 144
Goderdzishvili, Marina ...................... 171
Gogokhia, Lasha .................................. 32
Goldin, Sveta ........................................ 16
Golomidova, Alla ................................. 70
Gonzalez, Carlos ............................ 25, 85
González-Menéndez, Eva .................... 87
Goodridge, Lawrence ........................... 84
Górski, Andrzej ..... 57, 144, 145, 166, 174
Graham, Eric ....................................... 182
Graham, Zachary................................... 20
Grdzelishvili, Nino .............................. 171
Green, Jenny ....................................... 129
Griebel, Philip ....................................... 77
Griffiths, Mansel ................................... 30
Grigonyte, Aurelija ..................... 26, 165
Gromadka, Robert ............................... 144
Grosten, Kyle ...................................... 129
Guerin, Emma ........................... 113, 155
Guerrero-Ferreira, Ricardo ................... 70
Guild, Brandon ...................................... 28
Gula, Grzegorz .................................... 178
Gunathilake, Damitha ...................... 127
Gutiérrez, Diana ............................ 87, 89
Ha, Eunsu........................................... 142
Hakami, Ramin ..................................... 99
Halmillawewa, Anupama .................... 127
Hamilton, Theron .................... 49, 62, 168
Hansen, Lars ......................................... 46
Happel, Anna-Ursula ........................ 128
Happonen, Lotta .................................. 135
Hardy, Jonathan .................................. 167
Harhala, Marek ................. 145, 166, 174
Harrison, Melissa ................................ 156
Harrison, Sophie ............................... 129
Hatcher, Eneida ................................... 124
Hatfull, Graham ............................ 46, 156
Hawsawi, Hajar ..................................... 30
Hayes, Cillian ...................................... 183
Hayes, Connie ....................................... 77
Hayes, Sidney....................................... 77
He, Jenny............................................. 164
Hebert, Elvira ...................................... 123
Heck, Albert ........................................ 139
Hejatko, Jan......................................... 146
Hejnowicz, Monika ............................. 144
Hendrix, Hanne ............................. 64, 125
Henry, Marine ....................................... 78
Henry, Matthew ...................... 49, 62, 168
Hernández, R ....................................... 90
Hill, Colin ..... 50, 113, 125, 155, 160, 183
Hillebrand, Roman ................................ 46

Hinton, Debbie .............................. 65, 66
Hobbs, Zack ........................................ 91
Hodyra-Stefaniak, Katarzyna ..... 145, 174
Holguin, Angela ................................ 100
Holman, Derek ................................. 167
Holmfeldt, Karin ........................ 117, 121
Honaker, Ryan...................................... 91
Hooper, Annelise................................ 129
Hoyle, Naomi ...................................... 12
Hudson, Corey ..................................... 38
Huq, A ................................................ 170
Hurwitz, Bonnie ................................. 19
Hutchins, William .............................. 182
Hutinet, Geoffrey................................ 46
Hyman, Paul ............................... 81, 120
Hynes, Michael .................................. 127
Indráková, Adéla ................................ 153
Irwin, Casey ....................................... 129
Iskandriati, Diah ................................... 32
Israelsson, Stina ................................. 117
Iyer, L. .................................................. 66
Jackson, Simon ........................... 140, 141
Jacobs, Anna ............................... 49, 168
Jaiani, E ................................................ 92
Janda, Lubomir ................................... 146
Janelidze, N ................................. 92, 170
Jang, Ho Bin ....................................... 126
Jaramillo, Alfonso .............................. 165
Jaspan, Heather .................................. 128
Jaumdally, Shameem.......................... 128
Jauniškis, Vykintas ............................. 121
Jeon, Jongsoo .................................... 169
Jiménez, H ............................................ 90
Johnson, Allison ................................... 99
Jumba, Mirriam .................................... 94
Jun, Jin Woo ....................................... 135
Jurtz, Vanessa ..................................... 122
Kadija, Ermir ................................... 114
Kaestner, Coleman ............................. 129
Kaikabo, Adamu .......................... 32, 93
Kakabadze, Elene ............................. 171
Kallus, Katherine................................ 129
Kamwiziku, Guyguy ............................ 32
Kang'ethe, Erastus ................................ 32
Kariuki, Samuel .............................. 32, 94
Kaźmierczak, Zuzanna ........ 145, 166, 174
Kazwala, Rudovick ............................... 32
Kearns, Daniel..................................... 152
Keen, Eric ............................................ 17
Kemeraitė, Jolita ................................. 121
Kęska, Weronika ................................. 166
Khukhunashvili, T................................. 92
Khvichia, G. .......................................... 12
Kieper, Sebastian ................................ 139
Kilcher, Samuel ................................... 39
Kincaid, Randall ................................. 63
Kinkhabwala, Anika ............................. 91
Kirby, Bronwyn .................................... 22
Kirzner, Shay ........................................ 16
Klaus, James ......................................... 17
Kłopot, Anna ....................... 145, 166, 174
Klumpp, Jochen .................................... 39
Klykov, Oleg ....................................... 139
Knaus, Nolan....................................... 129
Knipling, Leslie..................................... 65
Knirel, Yuriy ......................................... 70
Koberidze, ........................................... 170
Kok, Jan .............................................. 102
Kokashvili, T....................................... 170
Koledin, Tamara ................................. 129
Konkol, Melissa .................................. 152
Kot, Witold ........................................... 46
Kozlova, Yu .................................... 55, 59
Kraus, Randolph ................................. 129
Krause, Kurt ........................................ 139
Krishnakumar, Raga ............................. 38
Kuhl, Sarah ........................................... 60
Kuipers, Oscar..................................... 102
Kutateladze, M .............................. 60, 133
Kutter, Betty .... 60, 68, 75, 123, 157, 158
Kvachadze, L ...................................... 133
Labrie, Simon ...................................... 115
LaFave, Matt ......................................... 33
Lakshman, N ......................................... 77
LaLonde, Tessa ................................... 129
Langevin, S. .......................................... 60
Latka, Agnieszka ............................... 130
Lau, Britney .......................................... 38

Lavigne, Rob ...... 40, 48, 64, 71, 96, 125,
126, 154
Lazarou, Michael.................................. 53
Le, Tram ............................................... 85
Leavitt, Justin ................................... 131
Lecion, Dorota.................... 145, 166, 174
Ledbetter, Stacia ................................. 129
Lee, Kyungwon .................................. 169
Lee, Yan-Jiun ..................................... 147
Leech, Thomas .................................. 172
LeGault, Kristen ............................... 143
Leigh, Brittany ................................... 20
Leiman, Petr ......................................... 70
Lemire, Sébastien ....................... 34, 184
Lepique, Ana ...................................... 105
Letarov, Andrey ................................... 70
Letarova, Maria.................................. 23
Letkiewicz, Sławomir .......................... 57
Leung, Chung ....................................... 78
Leung, Sharon .................... 156, 157, 158
Leung, Vince ........................................ 30
Lévesque, Alice ................................. 115
Li, Jian .................................................. 75
Liberti, Assunta .................................... 20
Lim, Loong-Tak ................................... 28
Lindell, Debbie ................................... 16
Linden, Sara ................................. 99, 103
Lindhard, Barbara .............................. 122
Liu, Y. .................................................. 44
Łobocka, Małgorzata ....................... 144
Lockwood, Madesyn .......................... 129
Loessner, Martin .................................. 39
Lomelí-Ortega, Carlos ............. 101, 173
Lopez, Anthony .................................. 129
Łoś, Marcin ......................................... 79
Lourenço, Marta .............................. 116
Lu, Rebecca .......................................... 91
Lu, Timothy ........................... 34, 97, 184
Ludwig, Kody .................................... 129
Luna, Sharmin .................................... 108
Lund, Ole ........................................... 122
Lundin, Daniel.................................... 117
Lungren, Matthew .............................. 167
Luque, Antoni ...................................... 53
Maes, M ................................................ 96
Maestri, Alice ........................................ 32
Magass, Nacia ..................................... 129
Mageeney, Catherine ........................ 150
Mahmud, Shahin ................................. 108
Mahony, Jennifer .................................. 69
Maidanik, Ilia ........................................ 16
Maina, Alice ................................... 32, 94
Majewska, Joanna............. 145, 166, 174
Majkowska-Skrobek, Grazyna ............ 130
Makalatia, Khatuna ............................. 171
Makumi, Angela ...................... 32, 69, 71
Malagon, Francisco ............................... 17
Malik, Danish .............................. 95, 175
Malki, Kema ...................................... 132
Mancuso, Francesco ............................ 175
Marcelli, Barbara .............................. 102
Marchon, Bruno .................................... 91
Markwitz, Pawel ................................. 178
Martin, Mark ....................................... 132
Martínez, Beatriz............................. 87, 89
Martínez-Díaz, Sergio ................. 101, 173
Martiny, Jennifer ................................. 118
Mašlaňová, Ivana ................................ 153
Mateczun, Alfred .................................. 62
Mattinen, Laura ................................... 135
Maura, Damien ................................... 116
Maxwell, Howard ............................... 139
Mazurkiewicz-Kania, Marta ............... 145
McAuliffe, Olivia ........................ 125, 134
McCann, Angela ................... 50, 113, 160
McCliment, Trevor ............................. 129
McCutcheon, Jaclyn ......................... 176
McDonnell, Siobhan ........................... 155
McKitterick, Amelia ........................... 80
McNamara, Bryanne ........................... 129
Melkova, Katerina............................... 146
Melo, Luis ............................................. 42
Merabishvili, Maia ........................ 48, 171
Meyer, Evelyne ..................................... 91
Międzybrodzki, Ryszard .................... 57
Miernikiewicz, Paulina ....... 145, 166, 174
Millard, Andrew ...... 26, 52, 95, 136, 165
Miller, Stefan ....................................... 35

Mkrtchyan, Mkhtar ............................ 171
Moineau, Sylvain ................... 46, 76, 115
Mokres, Lucia ...................................... 91
Moodley, Arshnee .............................. 154
Morales, Sandra ........... 58, 75, 157, 158
Morello, Eric ........................................ 78
Morozov, Andrew .............................. 112
Morozova, V.V. ............................. 55, 59
Mosier, Philip ....................................... 99
Mosterd, Cas ....................................... 76
Motoi, Maria ........................................ 61
Muessner, Christina ............................. 39
Mukhopadhyay, Suchetana ................ 152
Munoz, Robert ................................... 129
Mwaura, Francis ................................... 94
Myagmarjav, Bat-Erdene ................... 152
Nadareishvili, L. ................................... 12
Nagel, Tobi .......................................... 32
Nakavuma, Jesca ................................ 32
Nale, Janet ........................................... 32
Napper, Scott ........................................ 77
Napuli, Alberto ................................... 129
Nasinyama, George ............................ 32
Nawaz, Ayesha ................................... 135
Nazarov, Sergey ................................... 70
Needham, Micah ................................ 129
Neguț, Alina ........................................ 61
Neitzel, James .................................... 129
Nelson, Daniel ............... 36, 99, 103, 104
Nelson, Scott ....................................... 67
Neve, H .............................................. 134
Neve, Horst ........................................ 125
Nguyen, Sophie .................................... 53
Nielsen, Morten .................................. 122
Nikolich, Mikeljon .................... 133, 164
Nilsson, Emelie ......................... 117, 121
Nizharadze, D. ...................................... 12
Njoya, Kimani .................................... 182
Noben, Jean-Paul........................ 125, 126
Nobrega, F.L. ....................................... 44
Nocis, Melyssa ................................... 129
Novacek, Jiri ...................................... 146
Nowak, Sylwia ................................... 145
Nowotny, Taylor ................................ 129
Ntaongo, Julien ..................................... 32
Nyachieo, Atunga .............................. 177
Nyamongo, Onkoba ............................ 177
O’Mahony, Jim ................................... 125
O’Sullivan, Lisa ................................. 134
Oakley, Brian ........................................ 98
Oany, Arafat ........................................ 108
Ochieng, Joseph................................. 177
Odishelidze, N....................................... 12
Ogden, Luke ........................................ 129
Oliver, Andrew .................................. 118
Oliveria, Hugo..................................... 125
Olszak, Tomasz ................................. 178
Omran, Maha ...................................... 161
Ostapchuck, Yuri ................................ 124
Ouma, Arodi ....................................... 177
Owczarek, Barbara .............. 145, 166, 174
Padman, Benjamin ................................ 53
Pajunen, Maria .................................. 135
Pamungkas, Joko................................... 32
Pantůček, Roman ...................... 146, 153
Parker, David ...................................... 129
Parks, Olivia ........................................ 129
Parsley, Todd ....................................... 37
Passarell, María ................................... 123
Passmore, Jo-Ann ............................... 128
Patterson, Adrian................................. 141
Pearce, F. Grant................................... 139
Pekarsky, Irena ...................................... 16
Pernisova, Marketa ............................. 146
Perry, Benjamin .................................. 127
Peters, Danielle .......................... 151, 176
Peterson, Todd ...................................... 33
Petrova, Zaritza ................................... 156
Phillips, Kaden .................................... 129
Phillips, Kenya ...................................... 27
Pinhassi, Jarone ................................... 117
Pipia, L. ................................................. 12
Pires, Priscila......................................... 42
Pirnay, Jean-Paul .................. 11, 32, 171
Piya, Denish ......................................... 47
Plastow, Rian ...................................... 129
Plaut, Roger ....................................... 180
Plevka, Pavel ....................................... 153

Pollet, S ................................................ 96
Pope, Robert ......................................... 62
Pope, Welkin ...................................... 156
Porchkhidze, K ............................. 92, 170
Portilla, Silvia ....................................... 89
Prince, Jeffrey ...................................... 17
Prokhorov, Nikolai ............................. 70
Pruidze, N. ............................................ 12
Prüssing, Tessa ..................................... 88
Przybilski, Rita ................................... 141
Putonti, Catherine ............................ 119
Quaraishi, Habib ................................ 129
Quinones, Javier ........................... 62, 168
Radford, Devon .................................. 28
Rajamanickam, Karthic ........................ 77
Ramsey, Jolene ................................. 152
Raqib, Rubhana .................................. 108
Rauch, Benjamin .................................. 43
Ray, Melissa ...................................... 180
Raya, Raul .......................................... 123
Reddy, Akhil .............................. 133, 164
Redgwell, Tamsin ................. 26, 52, 136
Regeimbal, James ................... 49, 62, 168
Restrepo-Córdoba, Marcela ............... 127
Reyes, Alejandro .................... 13, 29, 90
Riccio, Cristian ..................................... 70
Richardson, Toby ................................. 33
Richter, Manuela .................................. 91
Rihtman, Branko .................................. 26
Rimbey, Jeanette .................................. 27
Ritchie, Jenny ..................................... 159
Roach, Dwayne ................................... 78
Roberts, Justin .................................... 129
Robertson, Rosanna ............................. 65
Rodríguez, Ana .............................. 87, 89
Rogóż, Paweł ....................................... 57
Rohde, Christine ................................. 137
Rohde, Manfred.................................. 137
Rohwer, Forest ..................................... 53
Roman, Jessica ..................................... 49
Rombouts, S. ........................................ 96
Ross, Alexa ........................................ 120
Ross, R. Paul 50, 113, 125, 155, 160, 183
Rossiter, Shayna ................................. 129
Roundtree, Breck ................................ 129
Rouse, Michael .................................... 49
Rowley, D. Treva .................................. 98
Russell, William .................................... 47
Ryan, Feargal ................................ 50, 160
Ryu, Sangryeol ........................ 83, 86, 142
Sacher, Jessica ................................... 147
Salmond, George................................. 141
Samir, Reham ...................................... 161
Săndulescu, Oana .................................. 61
Saunders, Candace .............................. 129
Sauvageau, Dominic ................... 156, 181
Sazinas, Pavelas .................................... 26
Scanlan, Dave ..................................... 136
Scheltema, Richard ............................. 139
Schlosser, Kyle ..................................... 53
Schmidt, Imke ..................................... 137
Schneider, Christine ......................... 182
Schoeniger, Joseph ................................ 38
Schooley, Robert ................................... 62
Schouler, Catherine ............................. 154
Schubert, Alyxandria .......................... 54
Scoville, Marissa ................................. 129
Sedrykyan, Anahid .............................. 171
Seed, Kimberley ............ 41, 80, 138, 143
Seil, Alexea ......................................... 129
Sergueev, Kirill ........................... 133, 164
Shanahan, Fergus ................................ 155
Shang, Xiaoran .................................. 104
Shapiro, Jason ..................................... 119
Shearer, Jonathan .................................. 49
Shohadai, Petal .................................... 129
Shoporov, Andrey ............................... 113
Šiborová, Marta................................... 153
Sillankorva, Sanna ................................ 42
Šimoliūnas, Eugenijus......................... 121
Simons, Mark ........................................ 49
Singhm,Devika...................................... 78
Skurnik, Mikael........................... 56, 135
Smith, George ....................................... 27
Smith, Keelan ...................................... 129
Smith, Muireann ............................... 183
Songailienė, Inga................................. 121
Sørensen, Martine ................................. 24

Speicher, David ................................... 51
Spröer, Cathrin ................................... 137
Squeglia, Flavia .................................. 130
Staals, Raymond ................... 21, 139, 141
Staes, Ines ............................................ 71
Staffe, Cæcilie .................................... 111
Stanbro, Josh ........................................ 49
Staub, Sean ......................................... 129
Stibitz, Scott ................................ 14, 180
Stintzi, Alain ...................................... 147
Stockelman, Michael .................... 49, 168
Stothard, Paul ..................................... 151
Strange, Philip ...................................... 28
Streinu-Cercel, Adrian ......................... 61
Streinu-Cercel, Anca ............................ 61
Studer, Patrick ...................................... 39
Sulaiman, Lanre ............................... 105
Šulčius, Sigitas .................................. 121
Swift, Stephen ...................................... 98
Szymanski, Christine ......................... 147
Taha, Omar ......................................... 162
Talukder, Kaisar ................................. 108
Taylor, Corinda .......................... 140, 141
Tediashvili, M .............................. 92, 170
Tedoradze, I.......................................... 12
Temple-Rosebrook, Louise .................. 99
Thanki, Anisha ..................................... 95
Thompson, Charles ............................ 129
Tikunova, N.V. ............................. 55, 59
Tolstoy, Igor ....................................... 124
Trindade, Marla .................................... 22
Tsersvadze, G ....................................... 92
Tsertsvadze, G .................................... 170
Turner, Paul .......................................... 32
Tyner, Stuart ................................. 49, 168
Valvano, Miguel ................................. 178
Van Belleghem, Jonas ........................ 48
van Charante, F .................................... 96
van Rijswick, Irma ............................... 69
van Sinderen, Douwe ........................... 69
Van Vaerenbergh, J .............................. 96
van Zyl, Lonnie .................................... 22
Vananzo, Steven ................................. 129
Vaneechoutte, Mario ............................ 48
Vara, Leonardo ..................................... 47
Vehring, Reinhard ......................... 32, 156
Velayudhan, Vimalkumar ............. 50, 160
Venneman, S ......................................... 96
Vidal, Amber....................................... 129
Villa, Luisa .......................................... 105
Villamizar, Santiago ......................... 106
Vincent, Antony .................................. 115
Vincent, Warwick ............................... 115
Vinner, Gurinder ........................... 95, 175
Vives, Martha ........................ 29, 106, 163
Vladisavljević, Goran ......................... 175
Vlassov, V.V. ........................................ 59
Volckaert, A .......................................... 96
von Henkelman, Jenny ........................ 129
Vrbovská, Veronika ............................ 153
Waddell, M. Brett ................................. 65
Wagemans, Jeroen .............................. 96
Wagner, Julian ...................................... 38
Wandro, Stephen ................................. 118
Wang, Jiping ......................................... 75
Ware, Vassie ....................................... 150
Waśko, Patryk ................................... 148
Waters, Jerel .......................................... 98
Watkins, Siobhan .............................. 119
Watson, Bridget................................... 21
Watters, Chaselynn ............................... 49
Weber-Dąbrowska, Beata ............. 57, 144
Weigele, Peter ..................................... 147
Weihe, Claudia .................................... 118
Weitz, Joshua ........................................ 78
Wenke, Joseph .................................... 168
West, Jennifer ...................................... 66
Weston, Thomas ................................... 53
Westra, Edze ........................................ 45
Wheat, Gordon ...................................... 60
White, Stephen ...................................... 65
Whiteson, Katrine ............................... 118
Whitford, Crystal ................................ 129
Wilkinson, Max................................... 139
Williams, Chandler ............................. 129
Williams, Kelly .................................... 38
Wittmann, Johannes ......................... 137
Wojtyna, Karolina ............... 145, 166, 174
Wolfe, David ........................................ 62
Wolput, Sanne ...................................... 71
Woolston, Joelle .................................. 31
Woźnica, Wioleta ............................... 144
Yakovets, E.A ...................................... 55
Yang, Andrew .............................. 34, 184
Ye, Wei .............................................. 100
Yeasmin, Mahmuda ........................... 108
Yehl, Kevin ................................. 34, 184
Yong, Dongeun ................................... 169
Yost, Christopher ............................... 127
Young, Ry ...................................... 47, 85
Zablocki, Olivier ................................. 22
Zaczek, Maciej ...................................... 91
Zdorovenko, Evelina ............................. 70
Zeman, Michal................................... 153
Zeng, Lanying ...................................... 72
Zermeño-Cervantes, Lina ................... 101
Zhang, Junjie ....................................... 73
Zhvania, P. ............................................ 12
Zidek, Lukas ....................................... 146
Zielonka, Katarzyna ............................ 145
Zschach, Henrike .............................. 122

Save the dates! ISVM Viruses of Microbes, July 9th – 13th 2018 We are really pleased to invite you to the 2018 Viruses of Microbes meeting: Biodiversity and
future applications that will take place in Wrocław, Poland.
The Wrocław meeting will be the 5th flagship meeting for the International Society of Viruses of
Microbes, following conferences organized in Paris 2010, Brussels 2012, Zurich 2014, and
Liverpool 2016, and bringing together research communities studying viruses infecting bacteria,
archaea, algae, fungi and protozoa. The conference will cover a broad range of topics, from
evolutionary and ecological aspects, through viral molecular structures, to practical applications in
industry and medicine. A special focus will be given to microbial virus encoded enzymes and
phage therapy, as these represent the main Polish experience in the field. Sessions will include:
Viral entry and exit in the variety of microbial hosts
Transcription and replication in viruses of microbes
Host-virus interactions: structure and function, microbial resistance to viral infection
Microbial virus encoded enzymes: fundamentals and application
Biotechnology and molecular engineering of microbial viruses
Phage therapy (I) fundamentals, (II) application, and discussion panel: direction to the future
Diversity and evolution
Ecology and systems biology
Application to market: food production, agriculture, environmental, diagnostics
All of our invited speakers are international experts, many of whom are new to speaking at a VoM
meeting so will bring fresh and innovative perspectives.
Dating back to the 10th century, Wrocław is a beautiful
city and a thriving multicultural centre attracting visitors
from all over the world. Its extensive heritage combines
almost all religions and cultures of Europe! Wrocław is
Poland’s fourth largest city, situated in the south-
western part of the country, on the banks of the Oder
river and its four confluents. With over 100 bridges,
Wrocław is often called the Venice of the North or the
City of Bridges. Besides its fascinating architecture and
being the cultural and academic centre of Lower Silesia, Wrocław is also famous for its dwarfs –
over 300 bronze figurines are scattered and hidden around the city. The city offers many cultural
attractions: 8 theatres, 14 museums, Opera House and National Forum of Music. The conference
venue – a spectacular, modern Conference Centre at the University of Wrocław – is located right in
the heart of the city, in front of the main building of the University and within walking distance of
the vibrant and colorful Market Square (Rynek) and picturesque Ostrów Tumski with a XIII
century gothic cathedral. The opening ceremony and choir concert will be held in the magnificent
Aula Leopoldinum, a representative baroque auditorium of the University of Wrocław. Social
events will also include welcome party in Oratorium Marianum, a get-together barbeque at the
Botanic Garden, and a special evening in the unique africarium of the Wrocław ZOO.
We look forward to seeing you in Wrocław! Please contact Prof. Zuzanna Drulis-Kawa
([email protected]) or Dr Krystyna Dąbrowska ([email protected]) if
interested in sponsorships or with any questions, comments or suggestions. Website with further
information and registration form will be available soon.


