414 jacc vol. 20, no. ” august 1992414-9 · 416 prager et al. isoforms of mb creatine kinase jacc...
TRANSCRIPT

414 JACC Vol. 20, No. ”
August 1992414-9
NELSON A. PRAYER, MD, TAD.40 SUZUKI, PHD,* ALLAN S. JAFFE,
BURTON E. SOBEL, MD, FACC, DANA IQ. ABENDSCHEIN, PHD
Saint Louis, Missotrri md Kyoto, Japan
Obje&ves. This study was designed to characterize the nature and time course of carboxpterminal lysine cleavages from the
creatine kinase (CK) in viva. Background. Rapid conversion of the tissue iSOfOrm Of MM CK
to two additional circulating isoforms with one or both carboxy- terminal lysines cleaved facilitates early detection of new tissue isofurm release after acute myocardiai infarction and coronary recanalization. Characterization of changes in plasma MS CK isoform profiles, potentia~Iy enhancing specificity even further, has been hindered by difffcuities in separating the isoform prod. ucts and elucidation of carboxy.terminal lysine cleavages under- lying their formation.
Metllods. Isoform species with carboxy-terminal lysine present on B.monomers were separated from those from which lysine had been cleaved by anion exchange chromatography. Carboxy terminal lysine on M.monomers was assayed with the use of a monospecific antibody.
Results. MB CK in four pooled plasma samples from among 77 normal subjects exhibited carbox~termi~al lysine on 48 f 211% (mean * SEM) af &monomers and 82 f 12% of
Further progress with interventions designed to restore coronary blood flow early in the course of acute myocardial infarction would be enhanced by methods for the rapid diagnosis of infarction and for detection of the occurrence and timing of coronary recanalization. Appearance ot the MB isoenzyme of creatine kinase (CK, EC 2.7.3.2) in plasma is a specific marker of acute myocardiai infarction (I). However, MB CK constitutes only 15% of the total CK activity in myocardiurr. (2) (the other 85% being MM CK),
From the Cardiovascular Division, Washington University, Saint Louis, Missouri and the *Biochemistry Department, Research and Development Center, Unitika, Ltd.. Kyoto, Japan. This study was supported in part by Specialized Center of Research in Coronary and Vascular Diseases Grant HLl%i6 and Grunt ROI HL36274 to Dr. Abendschein from the National Institutes of Health. Bethesda. Maryland and by Unitika. Ltd., which pmvided CKM-GO1 antibody. It was presented in part at the 40th Annual Scientific SeSSion, American College of Cardiology. Atlanta, Georgia. March 191.
Manuscript received July 9. 1991: revised manuscript received March 2, 1992, accepted March 25. 1992.
University Sch; Of Medicine, Cardiovascular Division, 6ijO ‘South Euclid _AddrtSs for e: Dana R. Abendschem, PhD Washington
Avenue. Box 8086, Saint Louis, Missouri 631 IO.
81992 by the American College of Cardiology
30 h, 43 f 9% (three ~00~s fro 95 f 10% of ~.moaolners terminus. After 40 to 50 h, I3 f of B-manamers and 46 9, 19% of M.mo~omers sti!l retained carbox~terminat lysine.
~o~~~us~o~s. In contrast to CF.<, the tissue ~sofQr CK undergoes slow cleavage sine from both monomers m vivo. Sequential cleavage of lysi of ~-monomers and subsequen with generation of at least two a of assays capable of resolving all of the isofor cam occur in vivo might increase sensitivity for early detention of new tissue isoform release associated with acute myoeardial inIarctio~ and coronary reca~alizatio~ compared with cu
and release of CK from myocardium undergoing infarction is slow initially. Thus, plasma MB CK activity remains within the normal range for several hours after the onset of infarc- tion. Even after coronary recanaiization, plasma MB CK activity may increase too si~wiy to permit its rapid recogni- tion.
After the initial report by Wevers et al. (3), it was discovered that once CK isoenzymes are released into the circulation, they undergo systematic biochemical modifica- tions that favor the early detection of infarction and coro- nary recanalization (4-6). The nature of these modifications has beeE defined for the MM isoenzyme and entails sequen- tial, irreversible cleavage of a single iysine residue from the carboxyl terminus of each M-monomer, which is mediated by circulating carboxypeptidase N (7-11). This results in time-dependent conversion of the tissue iseform (MM,) to two additional enzymatically active isoforms with one or both lysine residues removed (MM, and MMI) and exhibit- ing a stepwise increase in negative charge, facilitating their separation by electrophoresis and chromatofocusing. Be- cause conversion to other isoforms is rapid, MM, constitutes <20% of MM CK in plasma under physiologic conditions
0735-1097/92/$5.00

JACC Vol. 20. No. 2 PR.‘GER ET AL. August 1992:4!4-9 ISOFORMS OF MB CRE.%TINF MIWASE
has been consid-
mers (19). This observation is consi
isoforms analogous to those seen three additional isoforms: two
vivo and to define the time course of generation of isoform products in vivo. Assay techniques that d~stiogoish B-monomers on the basis of presence or absence of carboxy- terminal lysine and that detect -monomers with an intact carboxy terminal lysine permitted di re~tiatio~ of individ- real isoform species (19-2ii).
col was approved by the uman Studies Committee at Washington University. All subjects gave written informed consent. Blood (60 to 120 ml) was withdrawn fro the antecubital vein of 77 healthy volunteers (one sample/ volunteer) into heparinized syringes (final concentration of 20 U/ml), centrifuged at 9,000 rpm for 15 min and the plasma removed. Plasma MB CK mass in individual subjects was verified to be in the normal range (17 PgIliter) and plasma from four groups of subjects was pooled (total volumes of 900, I ,500, 890 and 1,070 ml). Each pool was assayed for isoforms of MB CK.
Serial plasma samples were obtained from patients with acute myocardial infarction who bad neither received tbroor- bolytic drugs nor undergone any other revascularization procedure. Samples (3 to 6 ml) obtained from 6 to 10 patients at each of three intervals after the onset of symptoms (510,
tive solid-phase radioi ues was asses
eluted from the ”
enzymatic activity of was remove
tbe presence or absence of
indicative of isoforms with and without ~arboxy-terminal lysine on the B-monomer, the proportions of the isoforms were expressed as percentages of the total area under tbe absorbance profile. ln some samples with high ties, the peaks were separated by preparative phy of the remaining with monitoring o (A,,,) in the column e t and collection of the individual protein peaks. These dialyzed and concentrated.
To detect the presence of carboxy-terminal lysine on the -monomer, isoforms separated by actor-exchange chro-
matography or the remaining sample not applied to the anion-exchange column were mixed with a rno~ospe~i~~ antibody that inhibits enzymatic activity of ~-monomers
maximal i~~b~bitio~~ of CK activity. The of the antibody was assessed with MB C prepared as described in the following section. The CK-antibody mixture was mixed gently over~~~bt at 4°C. Inhibition of CK activity was expressed as a percentage of the activity in paired control samples containing an equiva-
416 PRAGER ET AL. ISOFORMS OF MB CREATINE KINASE
JACC Vol. 20, No. 2 August lYY2:414-9
lent volume of buffer (IO mmol/liter Tris, I mm@llliter 2-mercaptoethanol, 0.2 mg/ml bovine serum albumin, pN 7.4).
Purification of individual isoenzymes from tissue.
Both MM and MB CK were p ed from the native hearts excised from patients who were recipients of donor cardiac dlografts; BB CK was purified from human umbilical cord. Tissues were rinsed with ice-cold saline solution and frozen at -20°C before processing. Tissue MB CK was extracted from myocardium, as described previously (231, except that precipitation of nor&K protein in the homogenate was done with 30% followed by 70% (weight/volume) ammonium sulfate, and the extract was applied directly to the “Conan- MB”-Scpharosc column. The etllueut was rerun through the affinity column to optimize binding of MB isoform to the antibody. The second cfflucnt, containing MM CK, was dialyzed against 50 mmollliter Tris, (pH 8.5) ovcraight at 4°C before further purification of the MM CK, as described previously (23).
Tissue BB CK was purified as described previously (23) except that after the second ethanol precipitation (70%) [he crude extract was fractionated by a single anion-exchange chromatographic step on a column of Q-Sepharose (PharmaciaILKB). The column was eluted with a linear gradient of sodium chloride (0 to 500 mmol) and fractions eluting at 220 to 250 mmol sodium chloride containing BB CK activity were pooled, dialyzed overnight (4°C) and concentrated before freezing at -70°C.
eparation of individual isoforms of M with lysine selectively removed from the carboxyl terminus d: either the M-monomer or the B-monomer were prepared as described previously (19). Briefly, MM CK and BB CK purified from tissue were thawed rapidly, equilibrated at room temperature for 5 mia and incubated with pancreatic cnrboxypeptidase !3 (Behring Diagnostics) in a mass ratio of I:25 (carboxypeptidase to CK) for 5 min at 22°C and 25 min at 4°C. The reaction was stopped by placing the mixture on ice. The resulting isoenzyme products were separated from carboxypeptidase B by anion-exchange chromatogra- phy (Q-Sepharosek with MM CK eluting from the column at 60 mmol sodium chloride, carboxypeptidase B at 100 mmol sodium chloride and BB CK at 240 mmol sodium chloride. Fractions containing CK activity were verified as devoid of
residual carboxypeptidase activity with use of a spectropho- tometric assay (9).
After dialysis and concentration, each repurified, ::arboxypeptiaase-treated isoenzyme was mixed with an equivalent mass of the tissue isoenzyme of the opposite monomer (for example, MLys(_, MLysq_) with BLys(+) B tYs(+J. The mixtures were made 4 molar with guanidine hydrochloride, incubated for IO min at 22°C. and dialyzed against 50 mmoNiter Trk (pH 7) containing IO mmofliter of 2-mercaptoethanol for 72 h at 4°C. The hybrid isoforms were separated from renatured MM and BB CK by preparative anion-exchange chromatography (Q-Sepharose).
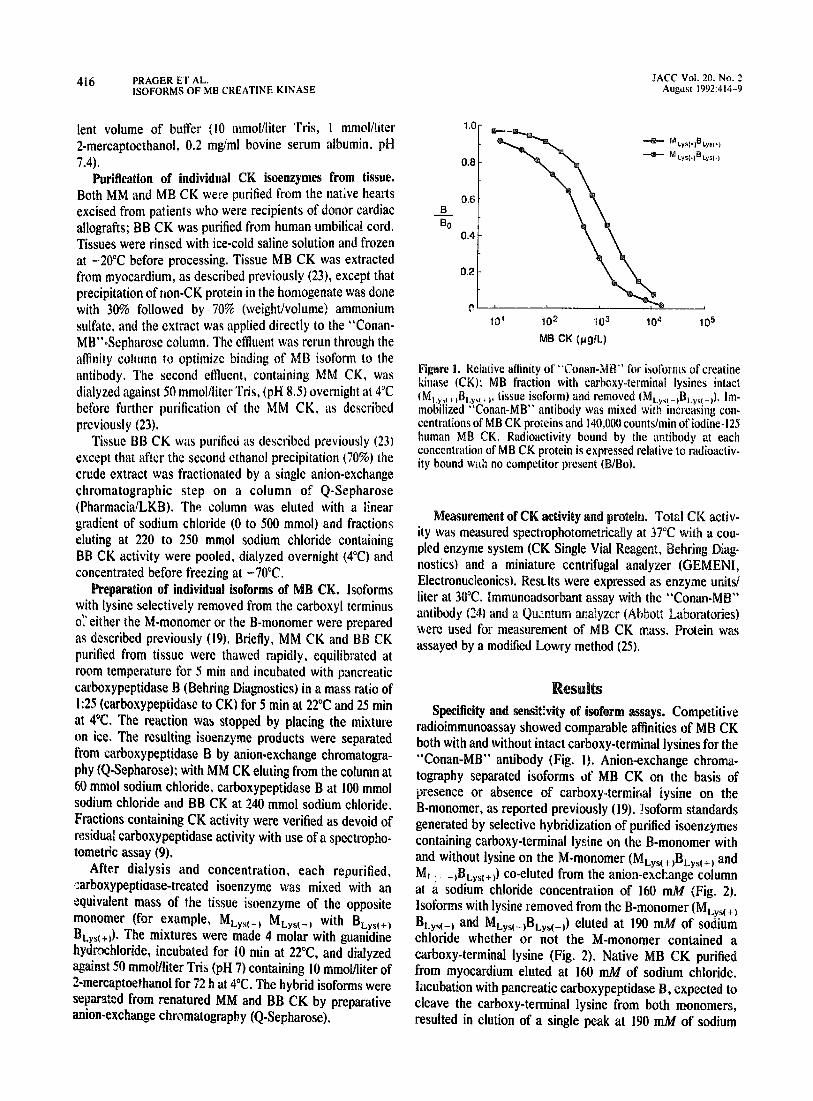
Figure 1. K&live affinity of “Conan-M ” for isoforms of creatine kinase (CK): MB fraction with ~arb~xy-germinal lysines intact (M ,+, , ,B,.y,c + ,, tissue isoform) and removed (MLy,(_,BLys,_,). lm- mobilized “Conan-MB” antibody was mixed with increasing con- centrations of MB CK proteins and 140,000 countslmin of iodine-125 human MB CK. Radioactivity bound by the antibody at each concentration of MB CK protein is expressed relative to radioactiv- ity bound with no competitor present (BIBo).
pled enzyme system (CK Single Vial nostics) and a miniature ccntrifug Electronucleonicsl. ResLIts were ex liter at 30°C. llmmunoadsorbant ass antibody (24’1 and a mere used for measurement of MB CK mass. Protein was assayed by a modified Lowry method (25).
Specificity and sensithity of isoform assays. Competitive radioimmunoassay showed comparable affinities of MB CK both with and without intact carboxy-terminal lysines for the “Conan-MB” antibody (Fig. I). Anion-exchange chroma- tography separated isoforms of MB CK on the basis of presence or absence of carboxy-termirdal lysine on the B-monomer, as reported previously (19). !soform standards generated by selective hybridization of purified isoenzymes containing carboxy-terminal lysine on the B-monomer with and without lysine on the M-monomer (MLys(+)BLys(+) and MI . _,BLys(+J co-eluted from the anion-exchange column at 4 sodium chloride concentration of 160 mA4 ( Isoforms with lysine removed from the B-monomer ( BLys(_) and MLys(-jBLys(-k) eluted at 190 mM of soLdyis:Fr: chloride whether or not the M-monomer contained a carboxy-terminal lysine (Fig. 2). Native MB CK purified from myocardium eluted at 160 m&f of sodium chloride. Incubation with pancreatic carboxypeptidase B, expected to cleave the carboxy-terminal lysine from both monomers, resulted in elution of a single peak at 190 mM of sodium
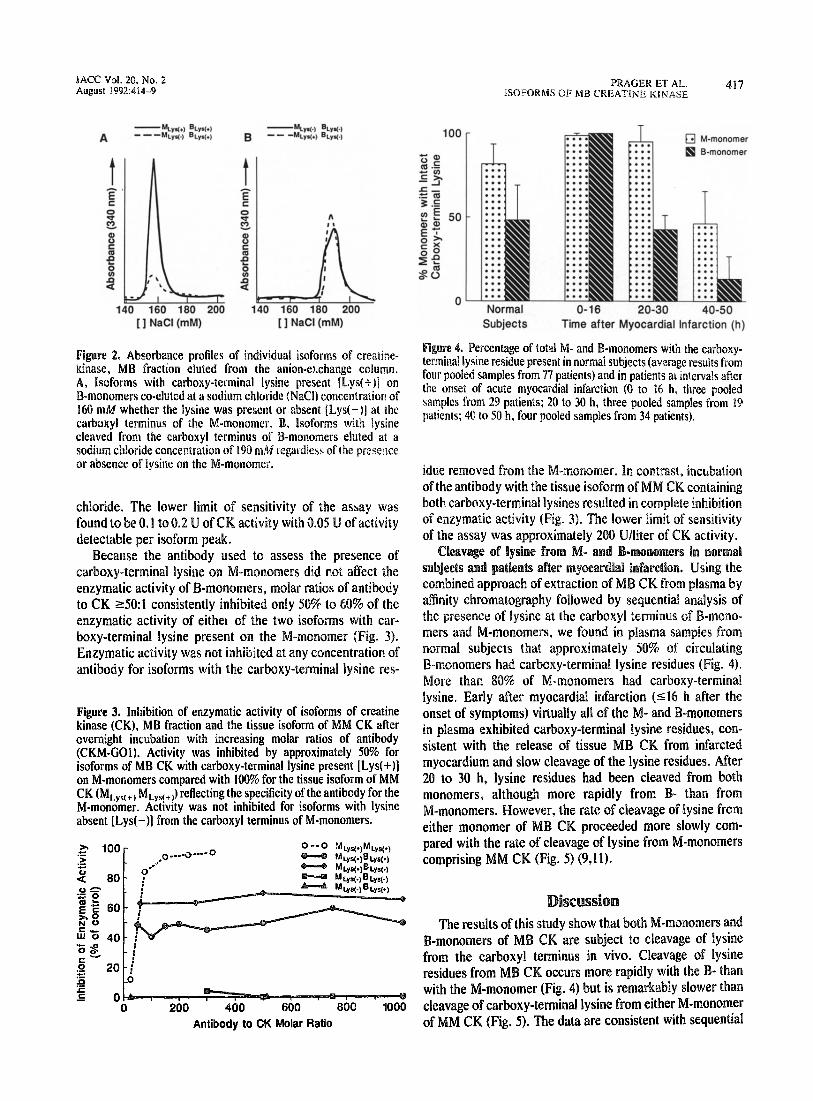
MC@ Vol. 20, No. 2 August ii?92:414-9
Absorbance profiles of individual isofor fraction eluted from the anion-exe
, Isoforms with carboxy-terminal lysiae present [Lys(t)j on -monomers co-eluted at a sodium chloride (WaCl) concentratiol~ of 0 miM whether the lysin absent [Lys(-)I at the
carboxyl terminus of the , lsoforms with lysine cleaved from the carboxyl monomers eluted al a sodium chloride concentrati gardless of tbe presence or absencr of lysine on the
chloride. The lower limit of sensitivity of the assay was found to be 0.1 to 0.2 U of C activity with 0.05 U of activity detectable per isoform peak.
Because antibody u ca~.boxy-tee 1 lysine on enzymatic activity of B-monomers, molar ratios of to CK m5O:l consistently inhibited only 50% to 60% of the enzymatic activity of either of the two isoforms with car- boxy-terminal lysine present on the -momomer (Fig. 3). Enzymatic activity was not inhi’oited at any concentration of antibody for isoforms with the carboxy-terminal lysine res-
Figure 3. Inhibition of enzymatic activity of isoforms of creatine kinase (CK), MB fraction and the tissue isoform of overnighl incubation with increasing molar ratios of antibody (CKM-GOl). Activity was inhibited by approximately 50% for isoforms of MB CK with carboxy-terminal lysine present [Lys(t)] on M-monomers compared with 100% for the tissue isoform of MM
CK (M,.ys(+, MW+I ) reflecting the specificity of the antibody for the M-monomer. Activity was not inhibited for isoforms with lysine absent [Lys(-)I from Ihe carboxyl terminus of M-monomers.
Antibody to GK Molar Ratio
M-monomer
B-monomer
~ocard~a~ Infarction (h)
and B-monomers with the carboxy- terminal lysine residue present in normal subjects (average results from four pooled samples from 99 patients) and in patients at intervals after the onset of acute myocardial infarction (0 to 16 h, three pooled samples from 29 patients: 20 to 30 h, three pooled samples from 19 patients; 40 to 50 h, four pooled samples from 34 patients).
of the assay was approximately
of lysine at the carboxyl -monomers, we found in plasma sa
normal subjects that approximately 50% of circulatang monomer5 bad car y-terminal lysine residues (Fig. 4). ore than 80% of monomers had carboxy-terminal
lysine. Early after myocardial infarction (516 h after the onset of symptoms) virtually all of the in plasma exhibited carboxy-terminal sistent with the release of tissue CK from infarcted myocardium and slow cleavage of the lysine residues. After 20 bo 30 h, lysine residues had been cleaved from both
ough more rapidly from B- than from rate of cleavage of lysine from proceeded more slowly com-
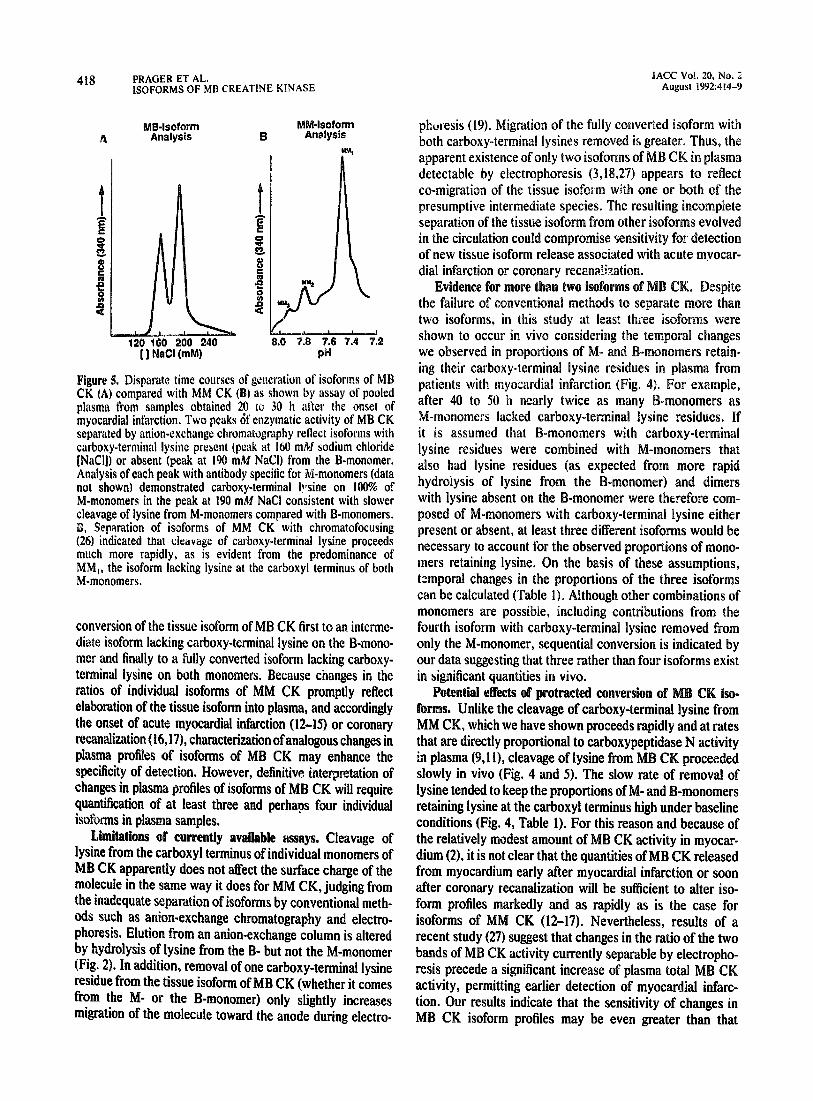
pared with the rate of cleavage of lysine from ~-monomer§ comprising MM CK (Fig. 5) (9,ll).
The results of this study show that both are subject to cleavage of lysine
I lerminus in vivo. Cleavage CK occurs more rapidly with th
-monomer (Fig. 4) but is remarkably slower t cleavage of carboxy-terminal lysine from either M-monomer of MM CK (Fig. 5). The data are consistent with sequential
MM-lsoform B Analysis
Pinure 5. Disoarats time courses of generation of isoforms of MB Ck (A) comp&d with MM CK (W-as shown by assay of pooled plasma from samples obtained 20 tu 30 h after the onset of myocardial infarction. Two peaks df enzymatic activity of MB CK separated by anion-exchange chromatography reflect isoforms wirh carboxy-terminal lysinc present (peak at 160 m&I sodium chloride [NaCl]) or absent (peak at 190 mM NaCI) from the B-monomer. Analysis of each peak with antibody specific for M-monomers (data not shown) demonstrated carboxy-terminal bfsine on 100% of M-monomers in the peak at 190 mM NaCl consistent with slower cleavage of iysine from M-monomers compared with B-monomers. S, Separation of isoforms of MM CK with chromatofocusing (26) indicated that cirav~ge of carboxy-terminal lysine proceeds much more rapidly, as is evident from the predominance of MM,, the isoform lacking lysine at the carboxyl terminus of both M-monomers.
PRAGER ET AL. JACC Vol. 20. No. 2
ISOFORMS OF MD CREATINE KINASE Augusl 1!392:414-9
MB-lsoform Analysis
ptoresis (19). Migration of the fully converted &form with both carboxy-termina! lysines removed is greater. Thus, the apparent existence of only two isoforms of detectable by electrophoresis (3,18,27) appears co-migration of the tissue isofoim with one or both of the presumptive intermediate species. The resulting incomplete separation of the tissue isoform from other isofor in the circulation could compromise sensitivity for detection of new tissue isoform release associated with acute m,vocar- dial infarction or coronary reca~a~~ati~~.
11 NaCl (mM)
conversion of the tissue isoform of MB CK first to an interme- diate isoform lacking carboxy-terminal lysine on the B-mono- mer and finally to a fully converted isoform lacking carboxy- terminal lysine on both monomers. Because changes in the ratios of individual isoforms of MM CK promptly reflect elaboration of the tissue isoform into plasma, and accordingly the onset of acute myocardial infarction (12-15) or coronary ~c~~ization (16,17), characterization of analogous changes in plasma profiles of isoforms of MB CK may enhance the specificity of detection. However, definitive interpretation of changes in plasma profiles of isoforms of MB CK will require quantification of at least three and perhaps four individual isofofms in plasma samples.
Limitations of currently available wsays. Cleavage of lysine from the carboxyl terminus of individual monomers of MB CK apparently does not affect the surface charge of the molecule in the same way it does for MM CK, judging from the inadequate separation of isoforms by conventional meth- ods such as anion-exchange chromatography and electro- phoresis. Elution from an anion-exchange column is altered by hydrolysis of lysine from the B- but not the M-monomer (Fig. 21. In addition, removal of one carboxy-terminal lysine residue from the tissue isoform of MB CK (whether it comes from the M- or the B-monomer) only slightly increases migration of the molecule toward the anode during electro-
hydrolysis of lysine from the B-rno~omcr~ and dimers absent on the B-monomer were therefore com- -monomers with carboxy-te
present or absent, at least three different i necessary to account for the observed pro mers retaining lysine. On the basis of these assumptions, temporal changes in the proportions of the three isoforms can be calculated (Table I). Although other combinations of monomers are possible, including contributions from the fourth isoform with carboxy-terminal lysine removed from only the M-monomer, sequential conversion is indicated by our data suggesting that three rather than four isoforms exist in significant
Potential e iso- forms. Unlike the cleavage of carboxy-terminal lysine from MM CK, which we have shown proceeds rapidly and at rates that are directly proportional to carboxypeptidase N activity in plasma (9,l I), cleavage of lysine from MB CK proceeded slowly in vivo (Fig. 4 and 5). The slow rate of removal of lysine tended to keep the proportions of M- and B-monomers retaining lysine at the carboxyl terminus high under baseline conditions (Fig. 4, Table 1). For this reason and because of the relatively modest amount of MB CK activity in myocar- dium (2), it is not clear that the quantities of MB CK released from myocardium early after myocardial infarction or soon after coronary recanalization will be sufficient to alter iso- form profiles markedly and as rapidly as is the case for isoforms of MM CK (12-17). Nevertheless, results of a recent study (27) suggest that changes in the ratio of the two bands of MB CK activity currently separable by ele resis precede a significant increase of plasma total activity, permitting earlier detection of myocatdial infarc- tion. Our results indicate that the sensitivity of changes in MB CK isoform profiles may be even greater than that

PRRGER ET AL. ISQFORMS OF MB CREATBFJE KlNASE
419
Normal s;ubjecis
Patients after infarction
Qto 16h
Xl to 30 h
40 to 50 b
*Calculated by assuming that all B-monomers with carboxy-terminal
lysine are combined with M-monomers with carboxy-terminal lysine. tThe
tissue isoform.
enon known to induce a
to be elucidated. The antibodies
playing antibodies specific the tissue isoform or as capable of separation of the tissue isofonn from all other CirC isoform species may enhance greatly the sensitivity and city for detection of acute myocardial infarction and coronary recanalization.
We thank Unitika for providing CKM-601 antibody; Jack H. Ladensen, PhD and Yvonne Land1 for providing “Conan-MB” antibody and for performing competitive radioimmunoassays; Bradley Reznick, Janice Gualdont, Timothy Bateman and James Pippin for technical assistance, and Barbara Donnelly for secretarial assistance.
Lott JA, Stang SM. Serum enzymes and isoenzymts in the diagnosis and differential diagnosis of myocardial ischemia and necrosis. Clin Chem 1980;26:1241-50.
Robeits R, Gowda KA, Ludbrook PA, Sobel BE. Specificity of elevated serum MB creatine phosphokinase activity in the diagnosis of acute myocardial infarction. Am J Cardiol 1975;36:433-7.
Wevers RA, Olthuis HP, Van Niel JCC, Van Wilgenburg MGM, Soons JBJ. A study on the dimeric structure of creatine kinase. Clin Chim Acta 1977;75:377-85.
R. Rapid d&no& 0; rllyucardial mfarction and rep&u- sion by assay of plasma isoforms of creatine kinase isoenzymes. Ciin Biocheal 19%X3:399-407.
5. Panteghim M. Serum isoforms of crectine kinase isoenzymes. Clin Bio 198X:21:21 I-P.
6. WM Creatine kinase isoforms in iscbemic heart disease. Clin Chem 198 -13.
7. George S. lshikawa Y, Perryman MB, Roberts R. Purification and characterization of naturally occurring and in vitro induced multiple forms of MM crea~ine k&se. J Biol C 1984;259:2667-74.
8. BilladelPo ii. Roman DG, Grace Sobel BE. Strauss AW. The nature ot’ post-translational formation M creatine kinase isoforms. J Biol Chem E985;260:14988-92.
R, Serota H, Plummer Jr TN, et al. Conversion of MM creatine kinase isoforms in human plasma by carboxypeptidase N. J Lab
IO.
II.
I?.
13.
14.
15.
16.
17.
18.
19.
Micbelutti L, Falter H. Certossi S, Marcotte B, Mazzuchin A. Isolation and purification of creatine kinase conversion factor from human serum and its identification as carboxypeptidase N. Clin Biochem 1987;20:2i-9.
Hashimoto PI, Grace AM, Billadello JJ, Gross RW, Strauss AW. Sobel E. Norrdenaturing quantification of subforms of canine MM crcatirle aase isoeazymes (isoforms) and their interconversion J Lab Clin Med
1984;103:470-84.
Abendschein U, Seacord LM, Nobara R, Sobel BE. Jaffe AS. Prom@ detection of myocardial injury by assay of creatine kinase isoforms in initial plasma ..an Clm Cdl&i 1989;11:66,1-4.
Jaffe AS, Serota race A, Sobel BE. Diagnostic changes in plasma creatine kinase is ms early after the onset of acute myocardial infarc- tion. Circulation 1986;74:105-9.
B, Gornet TG, Wu Q+., Br0ckie RE, Nishikawa A. Early diagnosis of acute myocardial infarction by rapid analysis of creatine kinase isoenzyme-3 (CK-MM) sub-types. Clin Chem 1987;33:358-62.
Panteghini M. Calarco M. Serum isoforms of creatine kinase MM and MB in myocardial infarction. An appraisal of quantitative, clinical and patho- physiological information. Stand J Clin Lab Invest 1987;47:325-9.
Seacord LM, Abendschein DR, Nohara R. Hartzler 6. Sobel BE, Jaffe AJ. Detection of reperfusion within I hour after coronary recanalisatlon by analysis of isoforms of the MM creatine kinase isoenzyme in plasma. Fibrinolysis 1988;2:151-6. Abendscbein DR. Ellis AK, Eisenberg PR, Klocke FJ, Sobel BE, Jaffe AS. Prompt detection of coronary recanalization by analysis of rates of changes of concentrations of macromolecular markers in plasma. Coro- nary Art Dis 1991;2:201-12. Wevers RA, Wolters RJ, Soons JBJ. isoelectric focusing and hybridisa- tion experiments on creatine kmase. Clin Chim Acta 1977;78:271-6.
Billadello JJ. Fontanet HL, Strauss AW, Abendschein DR. Characteri- zation of MB creatine kinase isoform conversion in vitro and in vivo in does. J Clin Invest 1989:83:1637-43.
20. L&dt Y, Vai&ja HC, Porter SE, Dietzler DN, Ladenson JH. Immunoaf- finity purification of creatine kinase-MB from human, dog, and rabbit heart with ose of a monoclonal antibody specific for CM-MB. Clin Chem 1989;35:985-9.
21. Suzuki T. Shiraishi T, Tomita K, Totani M, Murachi T. Monoclonal antibody inhibiting creatine kinase MM3 but not isoform MM,. Clin Chem 199O;36:153-6.
22. Suzuki T, Tomita K. Murachi T. Creatine kinase-inhibiting monoclonai antibodies: preparation and characterization of porcine MM iscenzyme- specific antibodies. Mol Cell Probes 1988;2:157-67.
23. Grace A, Roberts R. Improved procedures for purification of human and canine creatine kinase isoenzymes. Clin Chim Acta 1982;123:59-71.
24. Eisenberg PR, Shaw D, Schaab C, Jaffe AS. Concordance of creatine kinase-MB activity and mass. Clin Chem 1989;35:440-3.
25. Peterson GL. A simplification of the protein assay method of Lowry et al which is more generally applicable. Anal Biochem 1977;83:346-56.
26. Abendschein DIR, Fontanet HL, Nohara R. Optimized preservation of isoforms of creatine kinase MM isoenzyme in plasma specimens and their rapid quantification by semi-automated chromatofocusing. Clin Chem l%O;36:723-7.
27. Puleo PR. Guadagno PA, Roberts R, et al. Early diagnosis of acute myocardial infarction based on assay for subforms of creatine kinase-MB. Circulation 1990;82r759-64.