document
TRANSCRIPT

articles
Centrosome duplication in mammalian somatic cells requires E2F and Cdk2–Cyclin A
Patrick Meraldi*, Jiri Lukas†, Andrew M. Fry*‡, Jiri Bartek† and Erich A. Nigg*§*Department of Molecular Biology, University of Geneva, 30 quai Ernest-Ansermet, CH-1211 Geneva 4, Switzerland
†Danish Cancer Society, Institute of Cancer Biology, Department of Cell Cycle and Cancer, Strandboulevarden 49, DK-2100 Copenhagen Ø, Denmark‡Department of Biochemistry, Adrian Building, University of Leicester, University Road, Leicester LE1 7RH, UK
§e-mail: [email protected]
Centrosome duplication is a key requirement for bipolar spindle formation and correct segregation of chromosomes during cell division. In a manner highly reminiscent of DNA replication, the centrosome must be duplicated once, and only once, in each cell cycle. How centrosome duplication is regulated and coordinated with other cell-cycle functions remains poorly understood. Here, we have established a centrosome duplication assay using mammalian somatic cells. We show that centrosome duplication requires the activation of E2F transcription factors and Cdk2–cyclin A activity.
he major microtubule-organizing centres (MTOCs) areknown as centrosomes in animal cells and spindle-pole bod-ies (SPBs) in yeast1–4. The mammalian centrosome consists of
two barrel-shaped centrioles surrounded by pericentriolar mate-rial (PCM). In the course of each cell cycle, this organelle needs tobe duplicated once, and only once. During cell division the dupli-cated centrosomes associate with the poles of the mitotic spindleand thereby determine spindle bipolarity5. If proliferating cells failto coordinate centrosome duplication with DNA replication, thiswill inevitably lead to a change in ploidy, and the formation ofmonopolar or multipolar spindles will generally provoke abnormalsegregation of chromosomes. Indeed, it has long been recognizedthat errors in the centrosome duplication cycle may be an impor-tant cause of aneuploidy and thus contribute to cancer formation6.This view has recently received fresh impetus with the descriptionof supernumerary centrosomes in many human tumours7,8.
The regulation of centrosome duplication during the cell cycle ispoorly understood. Electron microscopic studies revealed that inmammalian cells small procentrioles emerge adjacent to each cen-triole during S phase9–12. Furthermore, centrosome duplication wasfound to be independent of DNA synthesis13, but to require anintact nucleus and protein synthesis9,14. The problem has also beenstudied extensively in eggs and early embryos of sea urchin, Xeno-pus and Drosophila15–19, and egg extracts have been used successfullyfor developing cell-free centrosome duplication assays20–22. Inembryonic systems, centrosome duplication is independent ofDNA or protein synthesis, and multiple rounds of centrosomeduplication occur even in the absence of a nucleus. This probablyreflects the fact that early embryos can rely on maternal stores ofcentrosomal components, whereas somatic cells need to synthesizecritical structural or regulatory components in each cell cycle.
Knowledge of the pathways that control MTOC duplication isfragmentary. In Saccharomyces cerevisiae, genetic analyses haveimplicated the centrin-related protein Cdc31 and the protein kinaseMps1 in SPB duplication2, but the precise functions of these pro-teins are unknown. Prominent among the proteins implicated inthe regulation of centrosome duplication in mammalian cells arep53 (ref. 23) and STK15/BTAK, a member of the Aurora family ofprotein kinases24, but the mechanisms of their action remainunclear. Most recently, elegant studies with extracts prepared fromXenopus eggs revealed a requirement for cyclin-dependent kinase 2(Cdk2)22,25. As we show here, a role for Cdk2 in centrosome dupli-cation can also be demonstrated in mammalian somatic cells.
To prevent genomic instability during cell proliferation, centro-some duplication and DNA replication must be coordinated. As afirst step towards understanding how coordination might beachieved in somatic mammalian cells, we have designed an experi-mental approach aimed at determining which, if any, of severalprominent cell-cycle regulators might be important in controllingcentrosome duplication. We find that centrosome duplication insomatic cells is strictly dependent on phosphorylation of the retino-blastoma (Rb) protein and requires both E2F transcription factorsand the activity of Cdk2, the latter most probably in association withcyclin A. Our results indicate that the commonly observed abroga-tion of the Rb pathway in human tumours will not only facilitateprogression towards DNA replication, but may also deregulate thecentrosome duplication cycle.
ResultsA centrosome duplication assay for somatic cells. In both somaticand embryonic systems, centrosome duplication can be experimen-tally uncoupled from DNA replication by the use of DNA synthesisinhibitors9,15,16,26. In particular, prolonged treatment of Chinese ham-ster ovary (CHO) cells with agents such as hydroxyurea (HU) allowsmultiple rounds of centrosome duplication to occur in the absence ofDNA replication or cytokinesis26. Taking advantage of this, we haveestablished a centrosome duplication assay in CHO cells, using bothimmunofluorescent staining with anti-γ-tubulin antibodies and elec-tron microscopy to monitor centrosome multiplication (Fig. 1A). Inthe absence of HU treatment, the vast majority of CHO cells con-tained two closely spaced, γ-tubulin-positive dots (Fig. 1Aa–e) andonly two centrioles (Fig. 1Ak). In contrast, most HU-treated CHOcells contained multiple γ-tubulin-positive centrosomes (Fig. 1Af–j)and multiple centrioles ( Fig. 1Al), in full agreement with a previousreport26. To determine which gene products might be involved in reg-ulating centrosome duplication, we transfected complementaryDNAs encoding known cell-cycle regulators into CHO cells, and usedanti-γ-tubulin staining after HU treatment as a convenient assay tovisualize ongoing centrosome duplication (Fig. 1B).Centrosome duplication is regulated through the Rb/E2F pathway.Progression towards S phase is controlled by the phosphorylationstate of the Rb protein27,28. This protein is phosphorylated primarilyby Cdk4/6-cyclin D, and these kinases are controlled in turn by theCdk4/6-specific inhibitor p16INK4. We thus asked whether the over-expression of p16INK4 could block centromsome duplication in HU-
T
88 NATURE CELL BIOLOGY | VOL 1 | JUNE 1999 | cellbio.nature.com© 1999 Macmillan Magazines Ltd

articles
treated CHO cells. When compared with untransfected cells or withcells transfected with a control plasmid encoding green fluorescentprotein (GFP), centrosome duplication was markedly inhibited incells expressing p16INK4 (Fig. 2). However, no such inhibition wasobserved when p16INK4 was overexpressed after the application of theHU block (Fig. 2b, + HU + p16 after). This suggests that the p16INK4
expressed at an earlier stage acted by imposing a G1 arrest rather
than by interfering with a direct role of Cdk4/6 in the duplicationprocess. To extend and confirm this finding, we next expressed anRb mutant (Rb∆Cdk) which lacks sites at which it can be phosphor-ylated by CDKs and is known to block cell-cycle progression down-stream of Cd4/6 (ref. 29). Expression of RB∆Cdk inhibitedcentrosome duplication to a similar extent as p16INK4 (Fig. 3a), indi-cating that Rb phosphorylation is strictly required.
Figure 1 Centrosome duplication assay. A, Detection of multiple rounds of centrosome duplication in hydroxyurea (HU)-treated cells. CHO cells were incubated for 40 h with HU (f–j, l) or without HU (a–e, k). They were then either stained with an antibody against γ-tubulin and analysed by immunofluorescence microscopy (a–j), or embedded, sectioned and analysed by electron microscopy (k, l). a, f, Whole cells; b–e, g–j, regions where centrosomes could be located by immunofluorescence microscopy. Electron microscopy confirms the presence of multiple centrioles in HU-
treated cells (l, arrowheads). Scale bars in a, f, 10 µm. Scale bar in k, l, 500 nm. B, Schematic representation of centrosome duplication assay. CHO cells were transfected for 6 h with cDNAs encoding different cell-cycle regulators. Two hours later, HU was added and cells were incubated for 40 h before immunofluorescence staining with anti-γ-tubulin antibodies. Centrosomes were counted in transfected cells as well as in surrounding untransfected cells.
Centrosome duplication(40 h in hydroxyurea)
Expression ofcell-cycle regulators
?
A
b c d e g h i j
a fHU
HU
+HU
+HU
B
k l
Figure 2 Overexpression of p16INK4 blocks centrosome duplication. CHO cells were transfected with plasmids encoding either Myc-tagged p16INK4 (a, b) or GFP (control) (c, d), treated with HU, and centrosome duplication was then analysed quantitatively. Transfected cells were identified by staining with anti-Myc antibodies and GPF fluorescence respectively (a, c), and centrosomes visualized by staining with anti-γ-tubulin antibodies (b, d). Centrosome numbers were determined for all transfected cells on every coverslip, using a Zeiss Axioplan II microscope equipped with a ×63 oil immersion objective. Scale bar 10 µm. e, Histograms indicate the percentages of cells with more than two centrosomes, indicative of ongoing
centrosome duplication. In untreated cultures, only about 15% of cells showed multiple (more than two) centrosomes (lower dotted line), whereas more than 50% of cells in HU-treated cultures had clearly undergone several rounds of centrosome duplication. Expression of GFP only marginally reduced the percentage of cells with multiple centrosomes (upper dotted line), whereas overexpression of p16INK4 lowered this value substantially, provided that expression of this inhibitor was induced before HU treatment. Error bars indicate standard deviations for data collected from five independent experiments.
p16
GFP
CentrosomesOverexpressed protein
+HU
Cel
ls w
ith m
ore
than
two
cent
roso
mes
(%
)
+HUHU +HU
20
30
40
50
+HU+GFP +p16 +p16
after
(control)
ea b
c d
_
NATURE CELL BIOLOGY | VOL 1 | JUNE 1999 | cellbio.nature.com 89© 1999 Macmillan Magazines Ltd
articles
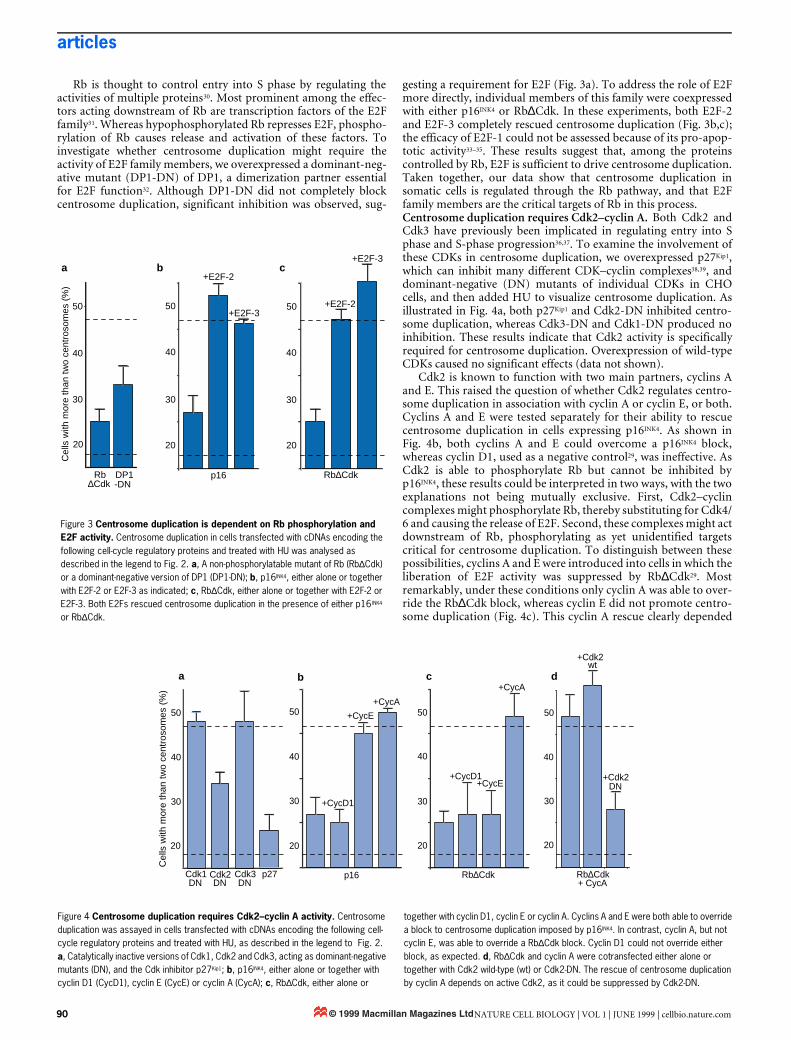
Rb is thought to control entry into S phase by regulating theactivities of multiple proteins30. Most prominent among the effec-tors acting downstream of Rb are transcription factors of the E2Ffamily31. Whereas hypophosphorylated Rb represses E2F, phospho-rylation of Rb causes release and activation of these factors. Toinvestigate whether centrosome duplication might require theactivity of E2F family members, we overexpressed a dominant-neg-ative mutant (DP1-DN) of DP1, a dimerization partner essentialfor E2F function32. Although DP1-DN did not completely blockcentrosome duplication, significant inhibition was observed, sug-
gesting a requirement for E2F (Fig. 3a). To address the role of E2Fmore directly, individual members of this family were coexpressedwith either p16INK4 or Rb∆Cdk. In these experiments, both E2F-2and E2F-3 completely rescued centrosome duplication (Fig. 3b,c);the efficacy of E2F-1 could not be assessed because of its pro-apop-totic activity33–35. These results suggest that, among the proteinscontrolled by Rb, E2F is sufficient to drive centrosome duplication.Taken together, our data show that centrosome duplication insomatic cells is regulated through the Rb pathway, and that E2Ffamily members are the critical targets of Rb in this process.Centrosome duplication requires Cdk2–cyclin A. Both Cdk2 andCdk3 have previously been implicated in regulating entry into Sphase and S-phase progression36,37. To examine the involvement ofthese CDKs in centrosome duplication, we overexpressed p27Kip1,which can inhibit many different CDK–cyclin complexes38,39, anddominant-negative (DN) mutants of individual CDKs in CHOcells, and then added HU to visualize centrosome duplication. Asillustrated in Fig. 4a, both p27Kip1 and Cdk2-DN inhibited centro-some duplication, whereas Cdk3-DN and Cdk1-DN produced noinhibition. These results indicate that Cdk2 activity is specificallyrequired for centrosome duplication. Overexpression of wild-typeCDKs caused no significant effects (data not shown).
Cdk2 is known to function with two main partners, cyclins Aand E. This raised the question of whether Cdk2 regulates centro-some duplication in association with cyclin A or cyclin E, or both.Cyclins A and E were tested separately for their ability to rescuecentrosome duplication in cells expressing p16INK4. As shown inFig. 4b, both cyclins A and E could overcome a p16INK4 block,whereas cyclin D1, used as a negative control29, was ineffective. AsCdk2 is able to phosphorylate Rb but cannot be inhibited byp16INK4, these results could be interpreted in two ways, with the twoexplanations not being mutually exclusive. First, Cdk2–cyclincomplexes might phosphorylate Rb, thereby substituting for Cdk4/6 and causing the release of E2F. Second, these complexes might actdownstream of Rb, phosphorylating as yet unidentified targetscritical for centrosome duplication. To distinguish between thesepossibilities, cyclins A and E were introduced into cells in which theliberation of E2F activity was suppressed by Rb∆Cdk29. Mostremarkably, under these conditions only cyclin A was able to over-ride the Rb∆Cdk block, whereas cyclin E did not promote centro-some duplication (Fig. 4c). This cyclin A rescue clearly depended
Figure 3 Centrosome duplication is dependent on Rb phosphorylation and E2F activity. Centrosome duplication in cells transfected with cDNAs encoding the following cell-cycle regulatory proteins and treated with HU was analysed as described in the legend to Fig. 2. a, A non-phosphorylatable mutant of Rb (Rb∆Cdk) or a dominant-negative version of DP1 (DP1-DN); b, p16INK4, either alone or together with E2F-2 or E2F-3 as indicated; c, Rb∆Cdk, either alone or together with E2F-2 or E2F-3. Both E2Fs rescued centrosome duplication in the presence of either p16INK4 or Rb∆Cdk.
Cel
ls w
ith m
ore
than
two
cent
roso
mes
(%
)
Rb DP1
20
30
40
50
Rb∆Cdkp16
+E2F-3
+E2F-2
20
30
40
50
+E2F-2
+E2F-3
20
30
40
50
a b c
∆Cdk -DN
Figure 4 Centrosome duplication requires Cdk2–cyclin A activity. Centrosome duplication was assayed in cells transfected with cDNAs encoding the following cell-cycle regulatory proteins and treated with HU, as described in the legend to Fig. 2. a, Catalytically inactive versions of Cdk1, Cdk2 and Cdk3, acting as dominant-negative mutants (DN), and the Cdk inhibitor p27Kip1; b, p16INK4, either alone or together with cyclin D1 (CycD1), cyclin E (CycE) or cyclin A (CycA); c, Rb∆Cdk, either alone or
together with cyclin D1, cyclin E or cyclin A. Cyclins A and E were both able to override a block to centrosome duplication imposed by p16INK4. In contrast, cyclin A, but not cyclin E, was able to override a Rb∆Cdk block. Cyclin D1 could not override either block, as expected. d, Rb∆Cdk and cyclin A were cotransfected either alone or together with Cdk2 wild-type (wt) or Cdk2-DN. The rescue of centrosome duplication by cyclin A depends on active Cdk2, as it could be suppressed by Cdk2-DN.
p16 Rb∆Cdk
+CycD1
+CycE
+CycA
+CycD1+CycE
+CycA
20
30
40
50
20
30
40
50
20
40
50
Rb∆Cdk
30
+Cdk2
+Cdk2
Cdk1 Cdk2 Cdk3
20
30
40
50
Cel
ls w
ith m
ore
than
two
cent
roso
mes
(%
)
a b c d
p27DN DN DN
wt
DN
+ CycA
90 NATURE CELL BIOLOGY | VOL 1 | JUNE 1999 | cellbio.nature.com© 1999 Macmillan Magazines Ltd

articles
on Cdk2 activity, as it could be abolished by coexpression of aCdk2-DN mutant (Fig. 4d). One might argue that the striking dif-ference in the abilities of cyclins A and E to rescue centrosomeduplication in Rb∆Cdk-expressing cells could be due to differencesin expression levels. This seems unlikely, however, as cyclin E wasas effective as cyclin A in overriding a p16INK4 block (Fig. 4b).Because of low transfection efficiencies, we were unable to measurecyclin A and cyclin E levels directly in Rb∆Cdk-expressing CHOcells, but a recent study on Rb∆Cdk-arrested Rat-1 cells reportsthat levels of cyclin E and cyclinE–Cdk2-activity were normal,whereas expression of cyclin A was suppressed40. Furthermore,even in the presence of Rb∆Cdk, cyclin E was more effective thancyclin A in promoting DNA replication (Fig. 5), in agreement withprevious studies indicating that cyclin E is more potent than cyclinA in promoting DNA replication under a variety of experimentalconditions (ref. 41; J. L. and J. B., unpublished). These results by nomeans exclude a role for cyclin E in centrosome duplication, asreported recently for Xenopus egg extracts22–25. However, if cyclin Edoes perform such a role in somatic cells, our data suggest that itneeds to cooperate with one or several E2F-regulated gene prod-ucts. In contrast, cyclin A is able to promote multiple rounds ofcentrosome duplication even in cells in which activation of E2F hasbeen inhibited by Rb∆Cdk.
To confirm a role for cyclin A in centrosome duplication insomatic cells, we asked whether HU-treated CHO cells could sup-port multiple rounds of centrosome duplication even when HUwas added to cells synchronized in mid-to-late S phase, that is, at atime when cyclin A levels are high but those of cyclin E are low42,43.To this end, CHO cells were synchronized by replating them aftershake-off in M phase, and progression towards S phase was moni-tored by measuring the incorporation of bromodeoxyuridine(BrdU). We found that 53% of the cells were able to undergo mul-tiple rounds of centrosome duplication when a 40-h HU treatmentwas applied 2 h after replating of M-phase cells, when only 9% ofcells were BrdU positive, whereas 83% of the cells showed multiplecentrosomes when HU was added 12 h after replating, when 78%of cells were BrdU positive (data not shown). This findingstrengthens the conclusion that cyclin A is the most likely partnerof Cdk2 in somatic cell centrosome duplication, at least under ourexperimental conditions.
DiscussionWe have established a novel centrosome duplication assay insomatic mammalian cells. Using this assay we asked how the regu-lation of centrosome duplication can be integrated into our currentunderstanding of cell-cycle regulation. We found that centrosomeduplication is dependent on the phosphorylation of Rb andrequires both E2F transcription factors and Cdk2–cyclin A activity.
One of the main reasons why centrosome duplication is less wellunderstood than other cell-cycle events is the shortage of suitableassay systems. This in turn is largely due to the fact that centrosomeduplication is difficult to score reliably, and that only electronmicroscopy provides the necessary resolution for detecting centri-oles unambiguously. However, upon addition of HU to certainsomatic cells, such as CHO, it is possible to observe multiple roundsof centrosome duplication26. These cells can conveniently be stud-ied by immunofluorescence microscopy, but we emphasize thatthey support bona fide duplication of centrioles rather than frag-mentation of PCM, as we demonstrate here by electron microscopy(see also ref. 26). We have combined the use of such cells with trans-fection experiments to study the involvement of individual cell-cycle regulators in centrosome duplication.
Our results clearly show that centrosome duplication in somaticmammalian cells requires the phosphorylation of Rb. This impliesthat both DNA replication and centrosome duplication are control-led through the same pathway. In normal proliferating cells, thiswould seem to provide an efficient means of ensuring the coordi-nate execution of these two key events. It also follows, however, thatthe loss of a functional Rb pathway, either through mutation or theaction of viral oncogenes, might jeopardize the coordinationbetween DNA replication and centrosome duplication and lead togenomic instability.
We further show that centrosome duplication requires theactivity of E2F transcription factors. Rb has been reported to reg-ulate the activities of several potential effector proteins30, but wefound that overexpression of E2F is sufficient to induce centro-some duplication in cells expressing a non-phosphorylatable Rbmutant. This indicates that, among the Rb-binding proteins, E2Fis the major downstream effector regulating centrosome duplica-tion. Furthermore, the dependence of somatic cells on the tran-scriptional activity of E2F may explain why such cells need an
Figure 5 Rescue of DNA replication in Rb∆∆∆∆Cdk-expressing cells by Cdk2–cyclin E. Cells were transfected with the cDNAs indicated and labelled for 3 h with BrdU. DNA replication was then analysed by immunofluorescent staining with anti-BrdU antibodies. The percentages of BrdU-positive cells were determined for untransfected cells (–), cells transfected with GFP alone (GFP), Rb∆Cdk alone, and Rb∆Cdk in combination with either cyclin E or cyclin A. When coexpressed with Rb∆Cdk, cyclin E was more efficient than cyclin A in promoting BrdU incorporation.
Brd
U-p
ositi
ve c
ells
(af
ter
3 h)
(%
)
GFP10
30
50
70
Rb∆CdkRb∆Cdk
+CycE
+CycA
Figure 6 Regulation of centrosome duplication in somatic cells. The model proposes that both centrosome duplication and DNA replication are regulated through the Rb pathway. Of the different proteins proposed to act downstream of Rb, E2F transcription factors are required for both processes. This model predicts, therefore, that some of the E2F target genes will be essential for centrosome duplication. In addition to E2F, we have identified a requirement for Cdk2–cyclin A in centrosome duplication. See text for details.
DNA replication
Centrosome duplication
RbpRb
P PCycACdk2E2F
CycD1Cdk4/6
p16
CycECdk2
NATURE CELL BIOLOGY | VOL 1 | JUNE 1999 | cellbio.nature.com 91© 1999 Macmillan Magazines Ltd

articles
intact nucleus and protein synthesis for centrosome duplication9,14,whereas embryonic systems only require cytoplasmiccomponents15. In the case of DNA synthesis, E2F activity isrequired to synthesize several essential gene products44, includingkey regulators of DNA replication such as Cdc6 (refs 45,46). Byanalogy, it seems plausible that E2F may activate the synthesis ofgene products that are critical for centrosome duplication; thesemay include regulatory proteins as well as bona fide centrosomalcomponents. In view of our results it is particularly intriguing thatboth the cyclin A and E genes are among the known target genesfor E2F47,48.
We show here that centrosome duplication in somatic cellsrequires Cdk2 activity in addition to E2F. In contrast, no evidencecould be obtained for involvement of Cdk1 or Cdk3, indicating thatthe Cdk2 requirement is specific. This conclusion is in excellentagreement with the results of two recent independent studies oncentrosome duplication in cell-free systems based on Xenopus eggextracts22,25. However, in Xenopus extracts cyclin E was identified asthe primary partner of Cdk2, whereas our studies on somatic cellslead us to emphasize the role of cyclin A. We cannot explain thisdiscrepancy but believe that it reflects a genuine difference in theregulation of centrosome duplication in the two systems. In fact,somatic cell cycles differ in several aspects from those of earlyembryonic cells. First, they are characterized by the presence of pro-longed G1 and G2 phases and they exhibit regulation at the tran-scriptional level, neither of which are seen in rapidly dividingembryonic cells. Second, the two systems differ markedly in the reg-ulation of cyclins A and E; in somatic mammalian cells cyclin E lev-els peak sharply around G1/S, whereas they are almost constantduring the cell cycles of early Xenopus embryos. Conversely, cyclinA is a prominent S-phase partner of Cdk2 in somatic mammaliancells, whereas in Xenopus embryos A-type cyclins do not associatewith Cdk2 before the mid-blastula transition49. In view of this, it ispossible that cyclin E has a prominent role in centrosome duplica-tion in early Xenopus embryos22,25 but that in somatic mammaliancells this function is performed primarily by cyclin A.
Our experiments have revealed an unexpected but intriguingdifference in the abilities of cyclins A and E to support Cdk2 activitywith respect to either centrosome duplication or DNA replication.In particular, in cells in which E2F activation was blocked by theexpression of a non-phosphorylatable Rb mutant, only cyclin A wasable to promote centrosome duplication; cyclin E was ineffective.Conversely, under very similar experimental conditions, cyclin Ewas more effective than cyclin A in inducing DNA replication (seealso J. L. and J.B., unpublished). In somatic cells therefore, cyclin Amay be the preferred partner of Cdk2 for centrosome duplication,whereas cyclin E may be primarily responsible for promoting theG1/S transition and initiating DNA replication. The temporal coin-cidence between centrosome duplication and cyclin A expression isconsistent with such a model, but more work is required to substan-tiate or refute this hypothesis.
In conclusion, we propose that in somatic mammalian cells cen-trosome duplication and DNA replication are regulated coordinately,as depicted schematically in Fig. 6. According to this model, bothprocesses are controlled by the Rb pathway, both processes depend onthe release of E2F, and both require Cdk2 activity. However, whereassubstrates involved in DNA replication are targeted primarily byCdk2–cyclin E, those involved in centrosome duplication are targetedprimarily by Cdk2–cyclin A, at least in mammalian somatic cells. Val-idation of this model will require the identification of the critical E2Ftarget genes and the physiologically relevant Cdk2 substrates. h
MethodsCell culture.CHO cells were grown at 37 °C in a 5% CO2 atmosphere in Ham’s F-12 medium supplemented with 10%
heat-inactivated fetal calf-serum (FCS) and penicillin-streptomycin (100 IU ml-1 and 100 µg ml-1,
respectively). Before transfection, cells were transferred into Dulbecco’s modified Eagle’s medium
(DMEM) supplemented with 10% heat-inactivated FCS and penicillin-streptomycin.
Centrosome duplication assay.CHO cells were seeded onto HCl-treated glass coverslips, at a density of 3 × 105 cells per 60-mm dish, in
Ham’s F-12 medium and transferred into 5 ml DMEM 2–3 h before transfection. They were transfected
with 20 µg plasmid DNA using calcium phosphate precipitates. For this purpose, DNA was incubated for
5 min in 240 µl CaCl2 (0.25 M), then mixed with 240 µl HBS solution (50 mM HEPES, 1.5 mM
Na2HPO4, 0.28 M NaCl) and added dropwise to the cells. After 6 h the medium was removed and the
cells treated for 3 min with 10% glycerol in phosphate-buffered saline (PBS). The cells were then washed
three times with PBS and incubated in Ham’s F-12. After an additional 2 h, HU was added to a final
concentration of 4 mM. Cells were fixed with cold methanol 40 h after the addition of HU and analysed
by immunofluorescence microscopy as described50. This experimental protocol was held constant for all
transfection experiments described, except for one experiment shown in Fig. 2 (+ HU + p16 after), in
which a 6-h transfection of p16INK4 was performed 12 h after the onset of the 40-h HU treatment.
Cells were stained for centrosomes with anti-γ-tubulin (purified IgG at 3.6 µg ml-1) and for the
overexpressed protein with either 9E10 anti-Myc monoclonal antibody or 12CA5 anti-haemagglutinin
(HA) monoclonal antibody, depending on the tag. Myc tags were used for detection of p16INK4, p27Kip1
and cyclins A, E and D1, whereas HA tags were used for detection of CDKs, Rb and E2Fs. Quantitative
analyses were performed on 200–600 transfected cells, depending on the protein combinations being
expressed, and at least three separate experiments were performed for each transfection.
Electron microscopy.Cells grown on coverslips were fixed with 2.5% glutaraldehyde (Sigma) (5 min, 37 °C, 55 min on ice),
washed with PBS, post-fixed with 1% osmium tetroxide, 1% potassium ferricyanide in PBS for 60 min
on ice, washed with distilled water, and treated with 1% aqueous uranyl acetate. Thereafter, cells were
dehydrated in ethanol, and embedded in epoxy resin (Fluka). After polymerization, the resin containing
the cells was separated from the coverslip by dipping into liquid nitrogen. Ultrathin sections were cut in
parallel to the monolayer and stained with aqueous uranyl acetate and lead citrate.
DNA replication assay.CHO cells seeded onto HCl-treated glass coverslips were transfected for 6 h as described above. To identify
transfected cells, a plasmid encoding GFP was added to each transfected DNA at a ratio of 1:10. Twenty
hours after the 10% glycerol treatment, BrdU (bromodeoxyuridine/fluorodeoxyuridine; Amersham) was
added to the medium for 3 h. Cells were fixed with 3% paraformaldehyde/2% sucrose for 10 min and
permeabilized with 0.5% Triton X-100 in PBS for 5 min. They were then treated for 10 min with 1.5 M
HCl, followed by extensive washing with PBS. BrdU incorporation was revealed using an anti-BrdU mouse
monoclonal antibody (Amersham), followed by Cy3-conjugated anti-mouse IgG antibodies (Jackson).
RECEIVED 16 MARCH 1999; REVISED 16 APRIL 1999; ACCEPTED 19 APRIL 1999; PUBLISHED 15 MAY 1999.
1. Stearns, T. & Winey, M. The cell center at 100. Cell 91, 303–309 (1997).
2. Winey, M. & Byers, B. Assembly and functions of the spindle pole body in budding yeast. Trends
Genet. 9, 300–304 (1993).
3. Paoletti, A. & Bornens, M. Organisation and functional regulation of the centrosome in animal cells.
Prog. Cell Cycle Res. 3, 285–299 (1997).
4. Kellogg, D. R., Moritz, M. & Alberts, B. M. The centrosome and cellular organization. Annu. Rev.
Biochem. 63, 639–674 (1994).
5. Heald, R., Tournebize, R., Habermann, A., Karsenti, E. & Hyman, A. Spindle assembly in Xenopus
egg extracts: respective roles of centrosomes and microtubule self-organization. J. Cell Biol. 138, 615–
628 (1997).
6. Boveri, T. Zur Frage der Entstehung maligner Tumoren (Fisher Verlag, Jena, 1914).
7. Lingle, W. L., Lutz, W. H., Ingle, J. N., Maihle, N. J. & Salisbury, J. L. Centrosome hypertrophy in
human breast tumors: implications for genomic stability and cell polarity. Proc. Natl Acad. Sci. USA
95, 2950–2955 (1998).
8. Pihan, G. A. et al. Centrosome defects and genetic instability in malignant tumors. Cancer Res. 58,
3974–3985 (1998).
9. Kuriyama, R. & Borisy, G. G. Centriole cycle in Chinese hamster ovary cells as determined by whole-
mount electron microscopy. J. Cell Biol. 91, 814–821 (1981).
10. Vorobjev, I. A. & Chentsov Yu, S. Centrioles in the cell cycle. I. Epithelial cells. J. Cell Biol. 93, 938–
949 (1982).
11. Chretien, D., Buendia, B., Fuller, S. D. & Karsenti, E. Reconstruction of the centrosome cycle from
cryoelectron micrographs. J. Struct. Biol. 120, 117–133 (1997).
12. Brinkley, B. R. Microtubule organizing centers. Annu. Rev. Cell Biol. 1, 145–172 (1985).
13. Kuriyama, R. & Borisy, G. G. Microtubule-nucleating activity of centrosomes in Chinese hamster
ovary cells is independent of the centriole cycle but coupled to the mitotic cycle. J. Cell Biol. 91, 822–
826 (1981).
14. Phillips, S. G. & Rattner, J. B. Dependence of centriole formation on protein synthesis. J. Cell Biol. 70,
9–19 (1976).
15. Sluder, G., Miller, F. J. & Rieder, C. L. The reproduction of centrosomes: nuclear versus cytoplasmic
controls. J. Cell Biol. 103, 1873–1881 (1986).
16. Raff, J. W. & Glover, D. M. Nuclear and cytoplasmic mitotic cycles continue in Drosophila embryos
in which DNA synthesis is inhibited with aphidicolin. J. Cell Biol. 107, 2009–2019 (1988).
17. Sluder, G., Miller, F. J., Cole, R. & Rieder, C. L. Protein synthesis and the cell cycle: centrosome
reproduction in sea urchin eggs is not under translational control. J. Cell Biol. 110, 2025–2032 (1990).
18. Gard, D. L., Hafezi, S., Zhang, T. & Doxsey, S. J. Centrosome duplication continues in cycloheximide-
treated Xenopus blastulae in the absence of a detectable cell cycle. J. Cell Biol. 110, 2033–2042 (1990).
19. Hinchcliffe, E. H., Cassels, G. O., Rieder, C. L. & Sluder, G. The coordination of centrosome
reproduction with nuclear events of the cell cycle in the sea urchin zygote. J. Cell Biol. 140, 1417–1426
(1998).
20. Palazzo, R. E., Vaisberg, E., Cole, R. W. & Rieder, C. L. Centriole duplication in lysates of Spisula
solidissima oocytes [erratum Science 256, 1746 (1992)]. Science 256, 219–221 (1992).
21. Tournier, F., Cyrklaff, M., Karsenti, E. & Bornens, M. Centrosomes competent for parthenogenesis
in Xenopus eggs support procentriole budding in cell-free extracts. Proc. Natl Acad. Sci. USA 88,
9929–9933 (1991).
22. Hinchcliffe, E. H., Li, C., Thompson, E. A., Maller, J. L. & Sluder, G. Requirement of Cdk2-cyclin E
activity for repeated centrosome reproduction in xenopus egg extracts. Science 283, 851–854 (1999).
92 NATURE CELL BIOLOGY | VOL 1 | JUNE 1999 | cellbio.nature.com© 1999 Macmillan Magazines Ltd

articles
23. Fukasawa, K., Choi, T., Kuriyama, R., Rulong, S. & Van de Woude, G. F. Abnormal centrosome
amplification in the absence of p53. Science 271, 1744–1747 (1996).
24. Zhou, H. et al. Tumour amplified kinase STK15/BTAK induces centrosome amplification,
aneuploidy and transformation. Nature Genet. 20, 189–193 (1998).
25. Lacey, K. R., Jackson, P. K. & Stearns, T. Cyclin-dependent kinase control of centrosome duplication.
Proc. Natl Acad. Sci. USA 96, 2817–2822 (1999).
26. Balczon, R. et al. Dissociation of centrosome replication events from cycles of DNA synthesis and
mitotic division in hydroxyurea-arrested Chinese hamster ovary cells. J. Cell Biol. 130, 105–115
(1995).
27. Lukas, J. et al. Retinoblastoma-protein-dependent cell-cycle inhibition by the tumour suppressor
p16. Nature 375, 503–506 (1995).
28. Weinberg, R. A. The retinoblastoma protein and cell cycle control. Cell 81, 323–330 (1995).
29. Lukas, J. et al. Cyclin E-induced S phase without activation of the pRb/E2F pathway. Genes Dev. 11,
1479–1492 (1997).
30. Wang, J. Y. Retinoblastoma protein in growth suppression and death protection. Curr. Opin. Genet.
Dev. 7, 39–45 (1997).
31. Dyson, N. The regulation of E2F by pRB-family proteins. Genes Dev. 12, 2245–2262 (1998).
32. Wu, C. L., Classon, M., Dyson, N. & Harlow, E. Expression of dominant-negative mutant DP-1
blocks cell cycle progression in G1. Mol. Cell. Biol. 16, 3698–3706 (1996).
33. Qin, X. Q., Livingston, D. M., Kaelin, W. G., Jr & Adams, P. D. Deregulated transcription factor E2F-
1 expression leads to S-phase entry and p53-mediated apoptosis. Proc. Natl Acad. Sci. USA 91, 10918–
10922 (1994).
34. Wu, X. & Levine, A. J. p53 and E2F-1 cooperate to mediate apoptosis. Proc. Natl Acad. Sci. USA 91,
3602–3606 (1994).
35. Kowalik, T. F., DeGregori, J., Schwarz, J. K. & Nevins, J. R. E2F1 overexpression in quiescent
fibroblasts leads to induction of cellular DNA synthesis and apoptosis. J. Virol. 69, 2491–2500 (1995).
36. van den Heuvel, S. & Harlow, E. Distinct roles for cyclin-dependent kinases in cell cycle control.
Science 262, 2050–2054 (1993).
37. Pagano, M. et al. Regulation of the cell cycle by the cdk2 protein kinase in cultured human fibroblasts.
J. Cell Biol. 121, 101–111 (1993).
38. Polyak, K. et al. Cloning of p27Kip1, a cyclin-dependent kinase inhibitor and a potential mediator of
extracellular antimitogenic signals. Cell 78, 59–66 (1994).
39. Toyoshima, H. & Hunter, T. p27, a novel inhibitor of G1 cyclin-Cdk protein kinase activity, is related
to p21. Cell 78, 67–74 (1994).
40. Knudsen, E. S., Buckmaster, C., Chen, T. T., Feramisco, J. R. & Wang, J. Y. Inhibition of DNA
synthesis by RB: effects on G1/S transition and S-phase progression. Genes Dev. 12, 2278–2292
(1998).
41. Stoeber, K. et al. Cdc6 protein causes premature entry into S phase in a mammalian cell-free system.
EMBO J. 17, 7219–7229 (1998).
42. Koff, A. et al. Formation and activation of a cyclin E-cdk2 complex during the G1 phase of the human
cell cycle. Science 257, 1689–1694 (1992).
43. Dulic, V., Lees, E. & Reed, S. I. Association of human cyclin E with a periodic G1-S phase protein
kinase. Science 257, 1958–1961 (1992).
44. Helin, K. Regulation of cell proliferation by the E2F transcription factors. Curr. Opin. Genet. Dev. 8,
28–35 (1998).
45. Hateboer, G. et al. Cell cycle-regulated expression of mammalian CDC6 is dependent on E2F. Mol.
Cell. Biol. 18, 6679–6697 (1998).
46. Yan, Z. et al. Cdc6 is regulated by E2F and is essential for DNA replication in mammalian cells. Proc.
Natl Acad. Sci. USA 95, 3603–3608 (1998).
47. Ohtani, K., DeGregori, J. & Nevins, J. R. Regulation of the cyclin E gene by transcription factor E2F1.
Proc. Natl Acad. Sci. USA 92, 12146–12150 (1995).
48. Schulze, A. et al. Cell cycle regulation of the cyclin A gene promoter is mediated by a variant E2F site.
Proc. Natl Acad. Sci. USA 92, 11264–11268 (1995).
49. Strausfeld, U. P. et al. Both cyclin A and cyclin E have S-phase promoting (SPF) activity in Xenopus
egg extracts. J. Cell Sci. 109, 1555–1563 (1996).
50. Fry, A. M., Meraldi, P. & Nigg, E. A. A centrosomal function for the human Nek2 protein kinase, a
member of the NIMA-family of cell cycle regulators. EMBO J. 17, 470–481 (1998).
ACKNOWLEDGEMENTS
We thank Y.-D. Stierhof for the electron microscopic data in Fig. 1, N. Roggli for help with artwork, and
several colleagues for kind gifts of plasmids, particularly E. Harlow for Cdk1, 2 and 3, J. Massagué for
p27Kip1, S. Reed for cyclin E , J. Pines for cyclin A , C.-L. Wu for DP1, and K. Helin for E2F. We also
thank B. Amati as well as all members of the laboratory for stimulating discussions. This work was
supported by grants from the Swiss National Science Foundation, the Swiss Cancer League, the Human
Frontier Science Program and the Canton of Geneva.
Correspondence and requests for materials should be addressed to E.A.N.
NATURE CELL BIOLOGY | VOL 1 | JUNE 1999 | cellbio.nature.com 93© 1999 Macmillan Magazines Ltd