7 | carbohydrates and glycobiology - web publishing€¢ two sugar molecules can be joined via a...
TRANSCRIPT

7 | Carbohydrates and Glycobiology
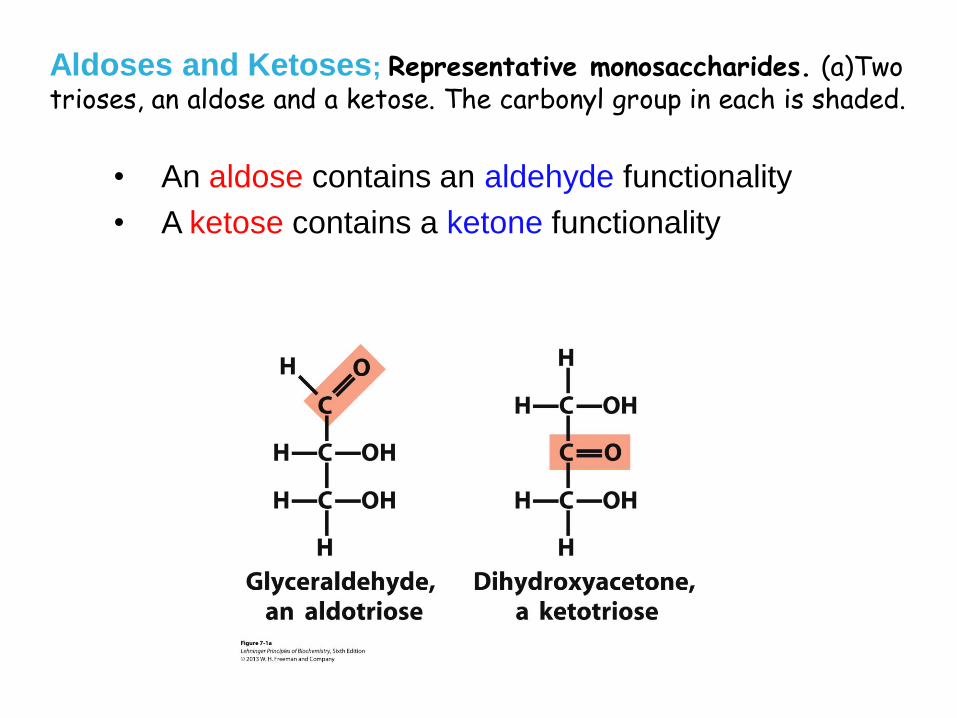
Aldoses and Ketoses; Representative monosaccharides. (a)Two trioses, an aldose and a ketose. The carbonyl group in each is shaded.
• An aldose contains an aldehyde functionality
• A ketose contains a ketone functionality

The Glycosidic Bond
• Two sugar molecules can be joined via a glycosidic bond
between an anomeric carbon and a hydroxyl carbon
• The glycosidic bond (an acetal) between monomers is less
reactive than the hemiacetal at the second monomer
– Second monomer, with the hemiacetal, is reducing
– Anomeric carbon involved in the glycosidic linkage is
nonreducing
• The disaccharide formed upon condensation of two glucose
molecules via 1 4 bond is called maltose

• Formation of maltose. A disaccharide
is formed from two monosaccharides
(here, two molecules of D-glucose)
when an —OH (alcohol) of one glucose
molecule (right) condenses with the
intramolecular hemiacetal of the other
glucose molecule (left), with elimination
of H2O and formation of a glycosidic
bond. The reversal of this reaction is
hydrolysis—attack by H2O on the
glycosidic bond. The maltose molecule,
shown here as an illustration, retains a
reducing hemiacetal at the C-1 not
involved in the glycosidic bond.
Because mutarotation interconverts the
α and β forms of the hemiacetal, the
bonds at this position are sometimes
depicted with wavy lines, as shown
here, to indicate that the structure may
be either α or β.

Nonreducing Disaccharides
• Two sugar molecules can be also joined via a glycosidic
bond between two anomeric carbons
• The product has two acetal groups and no hemiacetals
• There are no reducing ends, this is a nonreducing sugar
• Trehalose is a constituent of hemolymph of insects

• Two common disaccharides.
Like maltose in these are
shown as Haworth
perspectives. The
common name, full
systematic name, and
abbreviation are given for
each disaccharide. Formal
nomenclature for sucrose
names glucose as the
parent glycoside, although
it is typically depicted as
shown, with glucose on
the left.

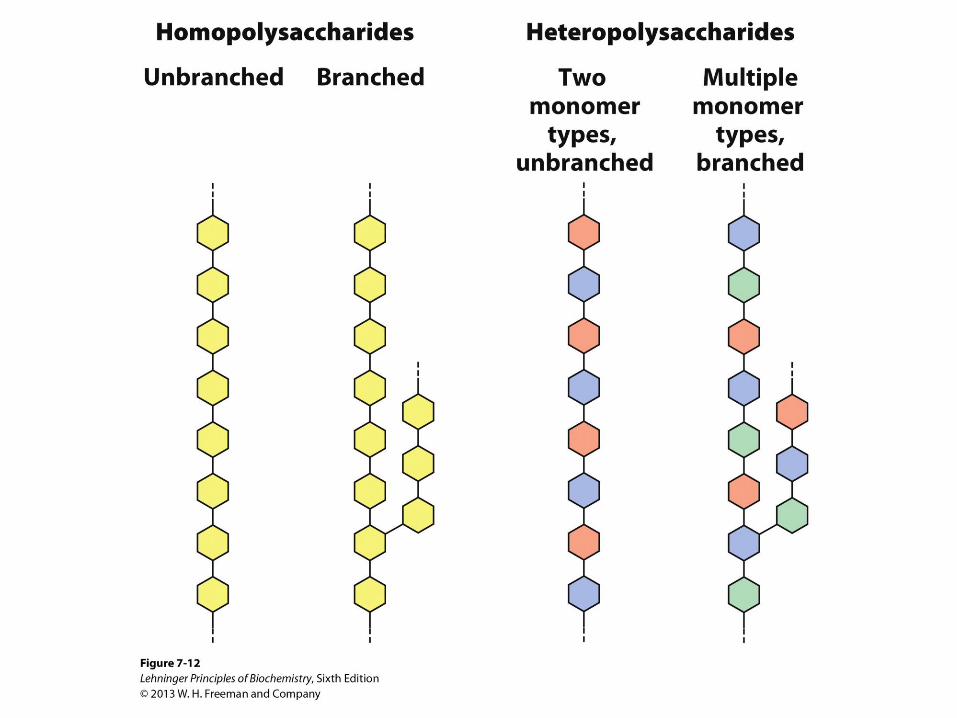
Polysaccharides
• Natural carbohydrates are usually found as
polymers
• These polysaccharides can be
– homopolysaccharides
– heteropolysaccharides
– linear
– branched
• Polysaccharides do not have a defined molecular
weight.
– This is in contrast to proteins because unlike
proteins, no template is used to make polysaccharides

Glycogen
• Glycogen is a branched homopolysaccharide of
glucose
– Glucose monomers form (1 4) linked chains
– Branch-points with (1 6) linkers every 8–12
residues
– Molecular weight reaches several millions
– Functions as the main storage polysaccharide in
animals

Starch
• Starch is a mixture of two homopolysaccharides of
glucose
• Amylose is an unbranched polymer of (1 4)
linked residues
• Amylopectin is branched like glycogen but the
branch-points with (1 6) linkers occur every 24–
30 residues
• Molecular weight of amylopectin is up to 200 million
• Starch is the main storage polysaccharide in plants

Glycosidic Linkages in Glycogen and Starch
• Glycogen and starch. (a) A short segment of amylose, a linear polymer of D-glucose residues in (α1→4) linkage. A single chain can contain several thousand glucose residues. Amylopectin has stretches of similarly linked residues between branch points. Glycogen has the same basic structure, but has more branching than amylopectin. (b) An (α1→6) branch point of glycogen or amylopectin.

Mixture of Amylose and Amylopectin in Starch
• Glycogen and starch. (c) A cluster of amylose and amylopectin like that believed to occur in starch granules. Strands of amylopectin (black) form double-helical structures with each other or with amylose strands (blue). Amylopectin has frequent (α16) branch points (red). Glucose residues at the nonreducing ends of the outer branches are removed enzymatically during the mobilization of starch for energy production. Glycogen has a similar structure but is more highly branched and more compact.

Metabolism of Glycogen and Starch
• Glycogen and starch often form granules in cells
• Granules contain enzymes that synthesize and degrade
these polymers
• Glycogen and amylopectin have one reducing end but
many nonreducing ends
• Enzymatic processing occurs simultaneously in many
nonreducing ends

Cellulose
• Cellulose is a branched homopolysaccharide of
glucose
– Glucose monomers form (1 4) linked chains
– Hydrogen bonds form between adjacent
monomers
– Additional H-bonds between chains
– Structure is now tough and water-insoluble
– Most abundant polysaccharide in nature
– Cotton is nearly pure fibrous cellulose

Hydrogen Bonding in Cellulose

Cellulose Metabolism
• The fibrous structure and water-insolubility make cellulose a
difficult substrate to act on
• Fungi, bacteria, and protozoa secrete cellulase, which allows
them to use wood as source of glucose
• Most animals cannot use cellulose as a fuel source because
they lack the enzyme to hydrolyze (1 4) linkages
• Ruminants and termites live symbiotically with microorganisms
that produces cellulase
• Cellulases hold promise in the fermentation of biomass into
biofuels

Chitin • Chitin is a linear homopolysaccharide of N-
acetylglucosamine
– N-acetylglucosamine monomers form (1 4)-linked
chains
– Forms extended fibers that are similar to those of
cellulose
– Hard, insoluble, cannot be digested by vertebrates
– Structure is tough but flexible, and water-insoluble
– Found in cell walls in mushrooms, and in exoskeletons
of insects, spiders, crabs, and other arthropods

Chitin, a short segment of chitin, a homopolymer of N-acetyl-D-glucosamine units in (β1→4) linkage.

Agar and Agarose
• Agar is a complex mixture of hetereopolysaccharides containing modified galactose units
• Agar serves as a component of cell wall in some seaweeds
• Agarose is one component of agar
• Agar solutions form gels that are commonly used in the laboratory as a surface for growing bacteria
• Agarose solutions form gels that are commonly used in the laboratory for separation DNA by electrophoresis
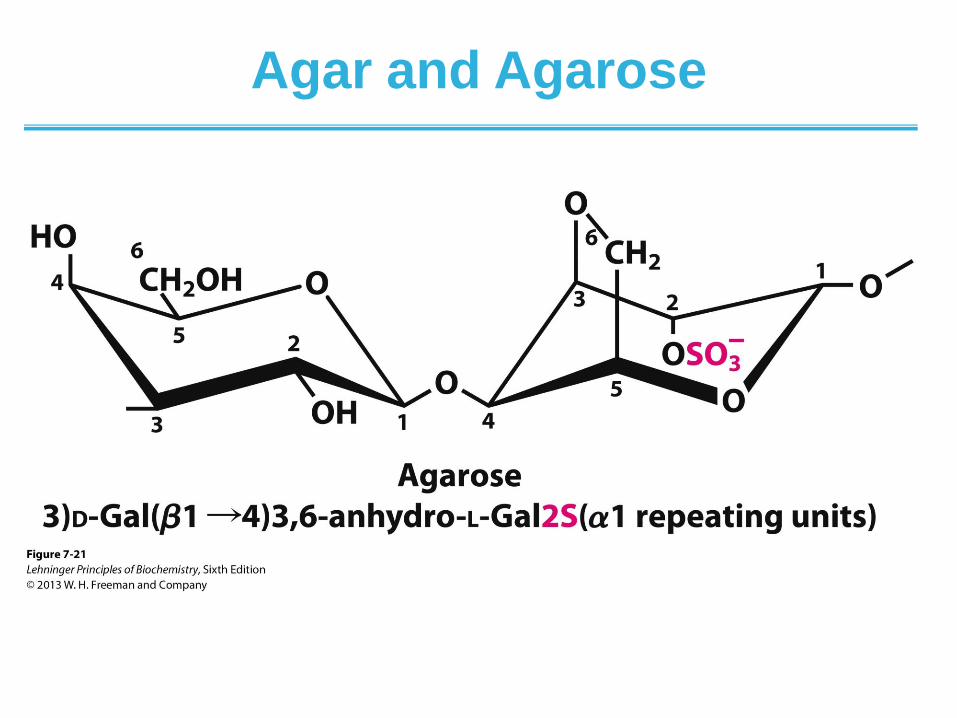
Agar and Agarose



Glycosaminoglycans
• Linear polymers of repeating disaccharide units
• One monomer is either
– N-acetyl-glucosamine or
– N-acetyl-galactosamine
• Negatively charged
– Uronic acids (C6 oxidation)
– Sulfate esters
• Extended hydrated molecule
– Minimizes charge repulsion
• Forms meshwork with fibrous proteins to form extracellular matrix
– Connective tissue
– Lubrication of joints

Repeating units of some common glycosaminoglycans
of extracellular matrix.

• The molecules are copolymers of alternating uronic acid and amino
sugar residues (keratan sulfate is the exception), with sulfate esters in
any of several positions, except in hyaluronan. The ionized
carboxylate and sulfate groups (red in the perspective formulas) give
these polymers their characteristic high negative charge. Therapeutic
heparin contains primarily iduronic acid (IdoA) and a smaller
proportion of glucuronic acid (GlcA, not shown), and is generally highly
sulfated and heterogeneous in length. The space-filling model shows a
heparin segment as its solution structure, as determined by NMR
spectroscopy. The carbons in the iduronic acid sulfate are colored
blue; those in glucosamine sulfate are green. Oxygen and sulfur
atoms are shown in their standard colors of red and yellow,
respectively. The hydrogen atoms are not shown. Heparan sulfate is
similar to heparin but has a higher proportion of GlcA and fewer
sulfate groups, arranged in a less regular pattern.

Heparin and Heparan Sulfate
• Heparin is linear polymer, 3–40 kDa • Heparan sulfate is heparin-like
polysaccharide but attached to proteins • Highest negative charge density biomolecules • Prevent blood clotting by activating protease
inhibitor antithrombin • Binding to various cells regulates development
and formation of blood vessels • Can also bind to viruses and bacteria and
decrease their virulence

The ionized carboxylate and sulfate groups (red in the perspective formulas) give these polymers their characteristic high negative charge. Therapeutic heparin contains primarily iduronic acid (IdoA) and a smaller proportion of glucuronic acid (GlcA, not shown), and is generally highly sulfated and heterogeneous in length. The space-filling model shows a heparin segment as its solution structure, as determined by NMR spectroscopy (PDB ID 1HPN). The carbons in the iduronic acid sulfate are colored blue; those in glucosamine sulfate are green. Oxygen and sulfur atoms are shown in their standard colors of red and yellow, respectively. The hydrogen atoms are not shown (for clarity). Heparan sulfate (not shown) is similar to heparin but has a higher proportion of GlcA and fewer sulfate groups, arranged in a less regular pattern.

• Interaction between a glycosaminoglycan and its binding protein. Fibroblast growth factor 1 (FGF1), its cell surface receptor (FGFR), and a short segment of a glycosaminoglycan (heparin) were co-crystallized to yield the structure shown here. The proteins are represented as surface contour images, with color to represent surface electrostatic potential: red, predominantly negative charge; blue, predominantly positive charge. Heparin is shown in a ball-and-stick representation, with the negative charges (—SO3
- and —COO-) attracted to the positive (blue) surface of the FGF1 protein. Heparin was used in this experiment, but the glycosaminoglycan that binds FGF1 in vivo is heparan sulfate on the cell surface.

Glycoconjugates: Glycoprotein
• A protein with small oligosaccharides attached
– Carbohydrate attached via its anomeric carbon
– About half of mammalian proteins are glycoproteins
– Carbohydrates play role in protein-protein recognition
– Only some bacteria glycosylate few of their proteins
– Viral proteins heavily glycosylated; helps evade the
immune system

Glycoconjugates: Proteoglycans
• Sulfated glucoseaminoglycans attached to a large rod-shaped protein in cell membrane
– Syndecans: protein has a single transmembrane domain
– Glypicans: protein is anchored to a lipid membrane
– Interact with a variety of receptors from neighboring cells and regulate cell growth

• Oligosaccharide linkages in glycoproteins. (a) O-linked oligosaccharides have a glycosidic bond to the hydroxyl group of Ser or Thr residues (light red), illustrated here with GalNAc as the sugar at the reducing end of the oligosaccharide. One simple chain and one complex chain are shown. (b) N-linked oligosaccharides have an N-glycosyl bond to the amide nitrogen of an Asn residue (green), illustrated here with GlcNAc as the terminal sugar. Three common types of oligosaccharide chains that are N-linked in glycoproteins are shown. A complete description of oligosaccharide structure requires specification of the position and stereochemistry (α or β) of each glycosidic linkage.

Proteoglycans
• Different glycosaminoglycans are linked to the core
protein
• Linkage from anomeric carbon of xylose to serine
hydroxyl
• Our tissues have many different core proteins;
aggrecan is the best studied

This is aggrecan a major proetoglycan (GAG) of cartilage.
• Proteoglycans are proteins that are heavily glycosylated. The basic proteoglycan unit consists of a core protein with one or more covalently attached GAG chains. The point of attachment is a Ser residue to which the glycosaminoglycan is joined through a tetrasaccharide bridge (e.g. chondroitin sulfate-GlcA-Gal-Gal-Xyl-PROTEIN). The Ser residue is generally in the sequence -Ser-Gly-X-Gly- (where X can be any amino acid residue, but Proline). he protein component of proteoglycans is synthesized by translocated into the lumen of the RER. Glycosylation of the proteoglycan occurs in the Golgi in multiple enzymatic steps. First a special link tetrasaccharide is attached to a serine side chain on the core protein to serve as a primer for polysaccharide growth. Then sugars are added one at a time by glycosyl transferase. The completed proteoglycan is then exported in secretory vesicles to the extracellular matrix.
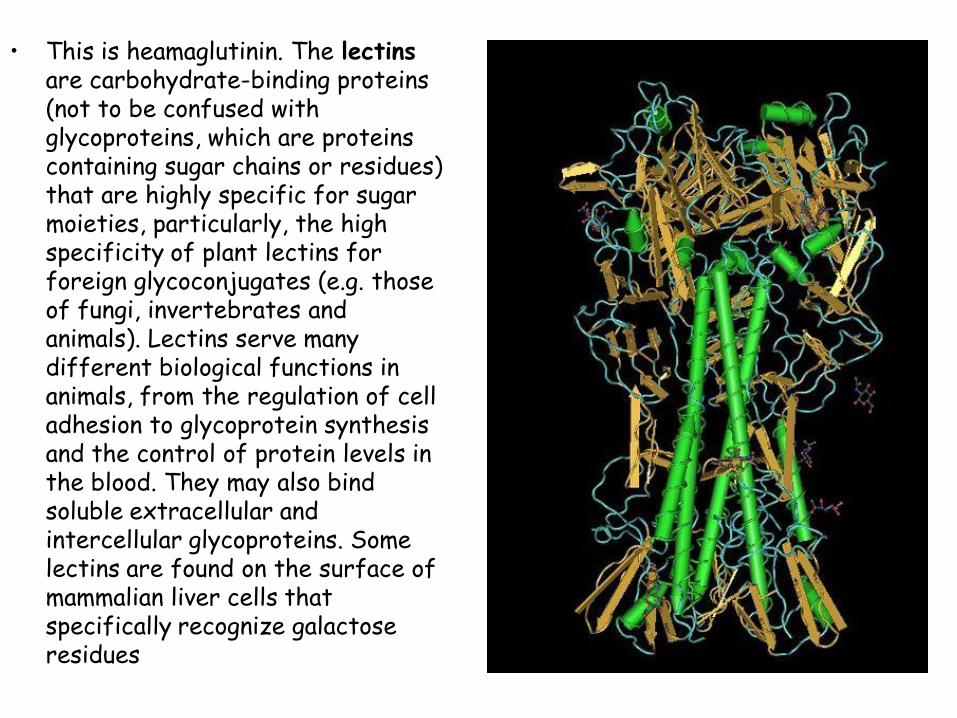
• This is heamaglutinin. The lectins are carbohydrate-binding proteins (not to be confused with glycoproteins, which are proteins containing sugar chains or residues) that are highly specific for sugar moieties, particularly, the high specificity of plant lectins for foreign glycoconjugates (e.g. those of fungi, invertebrates and animals). Lectins serve many different biological functions in animals, from the regulation of cell adhesion to glycoprotein synthesis and the control of protein levels in the blood. They may also bind soluble extracellular and intercellular glycoproteins. Some lectins are found on the surface of mammalian liver cells that specifically recognize galactose residues

• Fibronectin is a high-MW (~440kDa) glycoprotein of the extracellular matrix that binds to membrane-spanning receptor proteins called integrins. Similar to integrins, fibronectin binds extracellular matrix components such as collagen, fibrin, and heparan sulfate proteoglycans (e.g. syndecans).
• Fibronectin exists as a protein dimer, consisting of two nearly identical monomers linked by a pair of disulfide bonds. The fibronectin protein is produced from a single gene, but alternative splicing of its pre-mRNA leads to the creation of several isoforms. Two types of fibronectin are present soluble plasma fibronectin (formerly called "cold-insoluble globulin", or CIg) is a major protein component of blood plasma (300 μg/ml) and is produced in the liver. Insoluble cellular fibronectin is a major component of the extracellular matrix. It is secreted by various cells, primarily fibroblasts, as a soluble protein dimer and is then assembled into an insoluble matrix in a complex cell-mediated process.

Integrins; are transmembrane receptors that mediate the attachment between a cell and its surroundings, such as other cells or the extracellular matrix (ECM). Integrins couple the ECM outside a cell to the cytoskeleton (in particular the microfilaments) inside the cell. Which ligand in the ECM the integrin can bind to is defined by which α and β subunits the integrin is made of. Among the ligands of integrins are fibronectin, vitronectin, collagen, and laminin. The connection between the cell and the ECM may help the cell to endure pulling forces without being ripped out of the ECM.

• In signal transduction, integrins pass information about the chemical composition and mechanical status of the ECM into the cell. Therefore, in addition to transmitting mechanical forces across otherwise vulnerable membranes, they are involved in cell signaling and the regulation of cell cycle, shape, and motility. Integrins work alongside other proteins such as cadherins, immunoglobulin superfamily cell adhesion molecules, selectins and syndecans to mediate cell–cell and cell–matrix interaction and communication. Integrins bind cell surface and ECM components such as fibronectin, vitronectin, collagen, and laminin. Integrin subunits span the plasma membrane and in general have short cytoplasmic domains of about 40–70 amino acids. The exception is the beta-4 subunit, which has a cytoplasmic domain of 1088 amino acids, one of the largest known cytoplasmic domains of any membrane protein. Outside the cell plasma membrane, the α and β chains lie close together along a length of about 23 nm; the final 5 nm N-termini of each chain forms a ligand-binding region for the ECM. The molecular mass of the integrin subunits can vary from 90 kD to 160 kDa. Beta subunits have four cysteine-rich repeated sequences. Both α and β subunits bind several divalent cations. The role of divalent cations in the α subunit is unknown, but may stabilize the folds of the protein. The cations in the β subunits are more interesting: they are directly involved in coordinating at least some of the ligands that integrins bind.

• Two families of membrane proteoglycans. (a) Schematic diagrams of a syndecan and a glypican in the plasma membrane. Syndecans are held in the membrane by hydrophobic interactions between a sequence of nonpolar amino acid residues and plasma membrane lipids; they can be released by a single proteolytic cut near the membrane surface. In a typical syndecan, the extracellular amino-terminal domain is covalently attached to three heparan sulfate chains and two chondroitin sulfate chains. Glypicans are held in the membrane by a covalently attached membrane lipid (GPI anchor), but are shed if the bond between the lipid portion of the GPI anchor and the oligosaccharide linked to the protein is cleaved by a phospholipase. All glypicans have 14 conserved Cys residues, which form disulfide bonds to stabilize the protein moiety, and either two or three glycosaminoglycan chains attached near the carboxyl terminus, close to the membrane surface.

Two families of membrane proteoglycans. (b) Along a heparan sulfate chain, regions rich in sulfated sugars, the NS domains (green), alternate with regions with chiefly unmodified residues of GlcNAc and GlcA, the NA domains (gray). One of the NS domains is shown in more detail, revealing a high density of modified residues: GlcNS (N-sulfoglucosamine), with a sulfate ester at C-6; and both GlcA and IdoA, with a sulfate ester at C-2. The exact pattern of sulfation in the NS domain differs among proteoglycans.
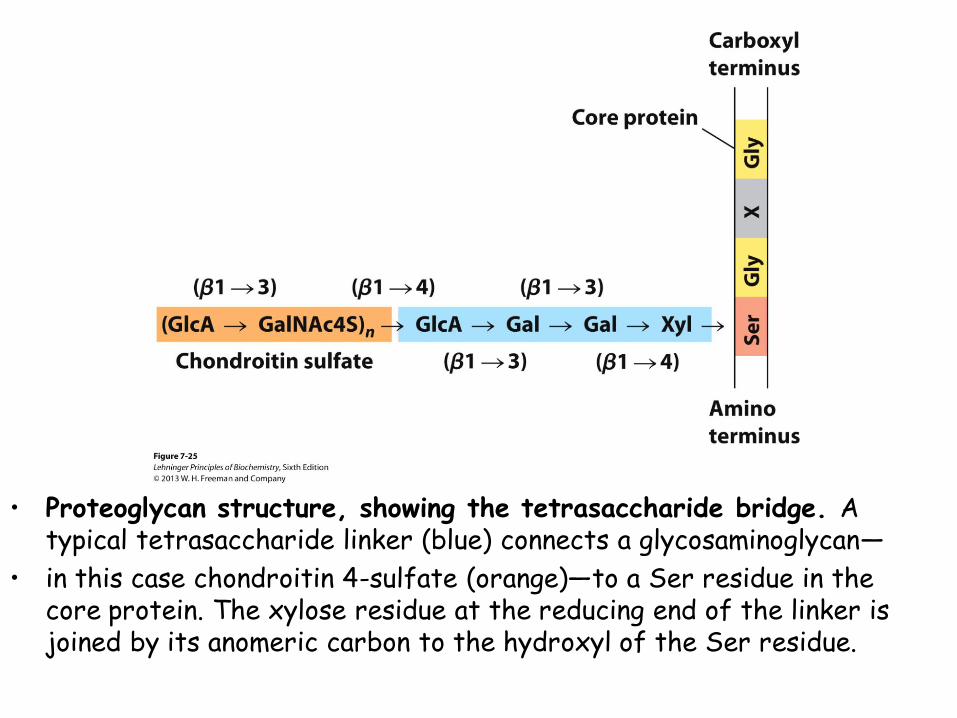
• Proteoglycan structure, showing the tetrasaccharide bridge. A typical tetrasaccharide linker (blue) connects a glycosaminoglycan—
• in this case chondroitin 4-sulfate (orange)—to a Ser residue in the core protein. The xylose residue at the reducing end of the linker is joined by its anomeric carbon to the hydroxyl of the Ser residue.

Proteoglycan Aggregates
• Hyaluronan and aggrecan form huge (Mr > 2•108) noncovalent aggregates
• Hold lots of water (1000 its weight); provides lubrication
• Very low friction material
• Covers joint surfaces: articular cartilage
– Reduced friction
– Load balancing

• Proteoglycan aggregate of the extracellular matrix. Schematic drawing of a proteoglycan with many aggrecan molecules. One very long molecule of hyaluronan is associated noncovalently with about 100 molecules of the core protein aggrecan. Each aggrecan molecule contains many covalently bound chondroitin sulfate and keratan sulfate chains. Link proteins at the junction between each core protein and the hyaluronan backbone mediate the core protein–hyaluronan interaction. The micrograph shows a single molecule of aggrecan, viewed with the atomic force microscope.

Extracellular Matrix (ECM)
• Material outside the cell
• Strength, elasticity, and physical barrier in tissues
• Main components – Proteoglycan aggregates
– Collagen fibers
– Elastin (a fibrous protein)
• ECM is a barrier for tumor cells seeking to invade new tissues – Some tumor cells secrete heparinase that degrades
ECM

Interaction of the Cells with ECM
• Some integral membrane proteins are proteoglycans – Syndecans
• Other integral membrane proteins are receptors for extracellular proteoglycans – Integrins
• These proteins link cellular cytoskeleton to the ECM and transmit signals into the cell to regulate: – cell growth – cell mobility – apoptosis – wound healing
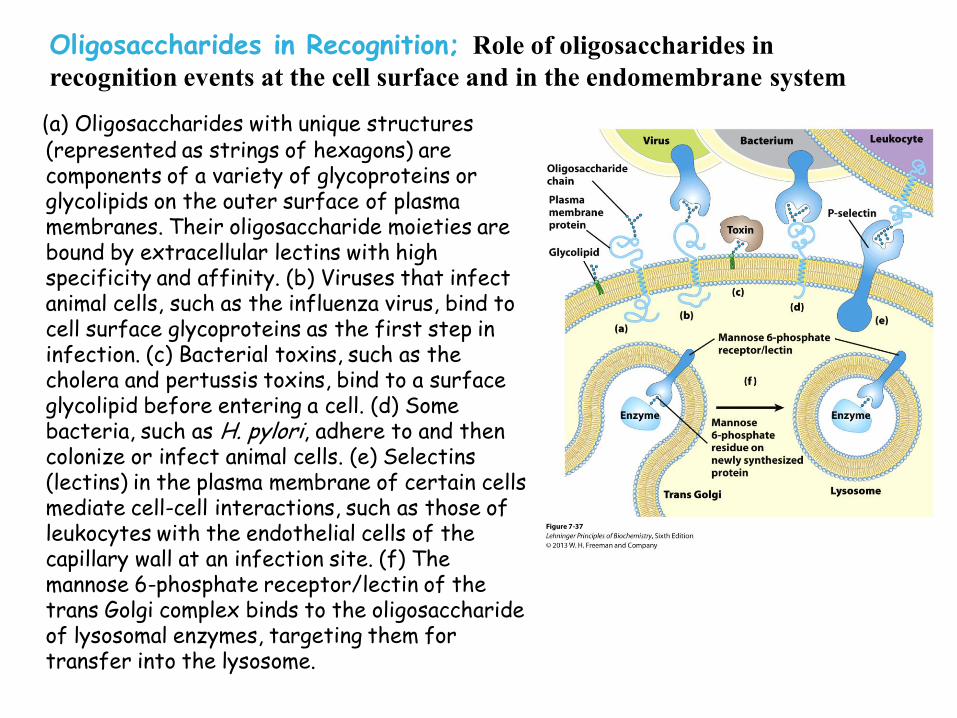
Oligosaccharides in Recognition; Role of oligosaccharides in
recognition events at the cell surface and in the endomembrane system
(a) Oligosaccharides with unique structures (represented as strings of hexagons) are components of a variety of glycoproteins or glycolipids on the outer surface of plasma membranes. Their oligosaccharide moieties are bound by extracellular lectins with high specificity and affinity. (b) Viruses that infect animal cells, such as the influenza virus, bind to cell surface glycoproteins as the first step in infection. (c) Bacterial toxins, such as the cholera and pertussis toxins, bind to a surface glycolipid before entering a cell. (d) Some bacteria, such as H. pylori, adhere to and then colonize or infect animal cells. (e) Selectins (lectins) in the plasma membrane of certain cells mediate cell-cell interactions, such as those of leukocytes with the endothelial cells of the capillary wall at an infection site. (f) The mannose 6-phosphate receptor/lectin of the trans Golgi complex binds to the oligosaccharide of lysosomal enzymes, targeting them for transfer into the lysosome.
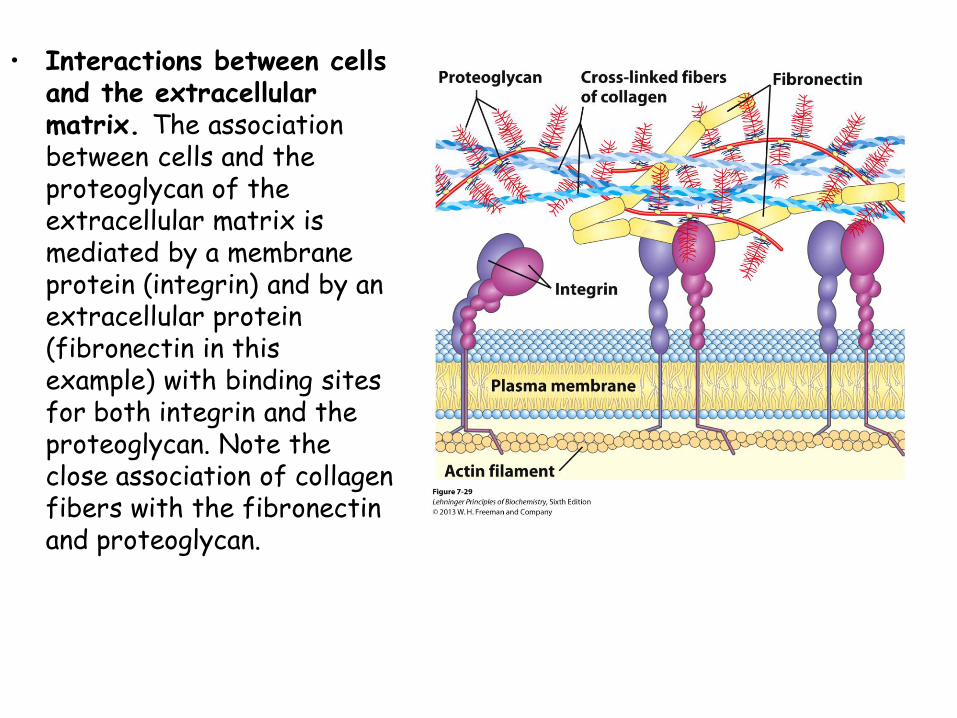
• Interactions between cells and the extracellular matrix. The association between cells and the proteoglycan of the extracellular matrix is mediated by a membrane protein (integrin) and by an extracellular protein (fibronectin in this example) with binding sites for both integrin and the proteoglycan. Note the close association of collagen fibers with the fibronectin and proteoglycan.

• Cadherins (calcium-dependent adhesion protien) are a class of type-1 transmembrane proteins. They play important roles in cell adhesion, forming adherens junctions to bind cells within tissues together. They are dependent on Ca2 to function, hence their name.
• The cadherin superfamily includes cadherins, protocadherins, desmogleins, and desmocollins, and more.[ In structure, they share cadherin repeats, which are the extracellular Ca2+-binding domains.


• Nucleotide–sugar transporters such as the Frc/UST74C protein are responsible for importing nucleotide sugars (bottom left) into the endoplasmic reticulum (ER) and Golgi compartments, where sequential enzymatic reactions convert the sugar substrates into complex, structurally diverse carbohydrate polymers attached to newly translated proteins. Proteoglycans, consisting of a protein core decorated with long unbranched carbohydrate chains, are exported through the secretory apparatus and become major constituents of the extracellular matrix (left) and outer surface of the cell (middle). Membrane-anchored proteoglycans on the cell surface are positive regulators of several key developmental signalling mechanisms, including the Wg/Wnt and Hh pathways, in which they are required for the optimal assembly or functioning of signalling complexes. Direct glycosylation of the receptor itself is an important regulatory feature of the Notch signalling pathway (right). During its transit through the trans-Golgi compartment, the Notch precursor protein (bottom right) might be cleaved by furin convertases and modified by an unusual O-linked fucosylation (green circles) performed by the glycosyltransferase Fringe (middle right). In cells expressing Fringe, the resulting Notch heterodimer is exported to the cell surface, where the O-linked carbohydrate modifications alter the responsiveness of the Notch receptor for its ligands Delta and Serrate (top right).
previous slide;

Glycoconjugates: Glycolipids
• A lipid with covalently bound oligosaccharide
– Parts of plant and animal cell membranes
– In vertebrates, ganglioside carbohydrate
composition determines blood groups
– In gram-negative bacteria, lipopolysaccharides
cover the peptidoglycan layer

• Bacterial LPS. Schematic diagram of the
LPS of the outer membrane of Salmonella
typhimurium. Kdo is 3-deoxy-D-manno-
octulosonic acid (previously called
ketodeoxyoctonic acid); Hep is L-glycero-D-
manno-heptose; AbeOAc is abequose (a 3,6-
dideoxyhexose) acetylated on one of its
hydroxyls. There are six fatty acid residues in
the lipid A portion of the molecule. Different
bacterial species have subtly different LPS
structures, but they have in common a lipid
region (lipid A), a core oligosaccharide also
known as endotoxin, and an “O-specific” chain,
which is the principal determinant of the
serotype (immunological reactivity) of the
bacterium. The outer membranes of the gram-
negative bacteria S. typhimurium and E. coli
contain so many LPS molecules that the cell
surface is virtually covered with O-specific
chains.

• LPS is the major component of the outer membrane of Gram-negative bacteria, contributing greatly to the structural integrity of the bacteria, and protecting the membrane from certain kinds of chemical attack. LPS also increases the negative charge of the cell membrane and helps stabilize the overall membrane structure. It is of crucial importance to gram-negative bacteria, whose death results if it is mutated or removed. LPS is an endotoxin, and induces a strong response from normal animal immune systems. It has also been implicated in non-pathogenic aspects of bacterial ecology, including surface adhesion, bacteriophae sensitivity, and interactions with predators such as amoebae.


• Toll-like receptors (TLRs) are a class of proteins that play a key role in the innate immune system as well as the digestive system. They are single, membrane-spanning, non-catalytic receptors usually expressed in sentinel cells such as macrophages and dendritic cells that recognize structurally conserved molecules derived from microbes. Once these microbes have breached physical barriers such as the skin or intestinal tract mucosa, they are recognized by TLRs, which activate immune cell responses.