8 physiology and testing of respiratory musclespneumonologia.gr/articlefiles/1 respiratory muscles...
TRANSCRIPT

SECTION II
PHYSIOLOGY
810008 Physiology and Testingof Respiratory Muscles
THEODOROS VASSILAKOPOULOS � CHARIS ROUSSOS
s0010 INTRODUCTION
p0010 The respiratory muscles are the only muscles, along with theheart, that have to work continuously, though intermittently,to sustain life. They have to repetitively move a rather com-plex elastic structure, the thorax, to achieve the entry of airinto the lungs and thence effect gas exchange. Their greatnumber mandates that they should interact properly to per-form their task despite their different anatomic location, geo-metric orientation, and motor innervation. They should alsobe able to adapt to a variety of working conditions and respondto many different chemical and neural stimuli. This chapterdescribes some aspects of the respiratory muscles’ functionthat are relevant to the understanding of the way the respira-tory muscles accomplish the action of breathing and how theirfunction can be tested in the laboratory.
s0020 FUNCTIONAL ANATOMY
s0030 The Intercostal Musclesp0020 The intercostal muscles are two thin layers of muscle fibers
occupying each of the intercostal spaces. They are termedexternal and internal because of their surface relations, theexternal being superficial to the internal. The muscle fibersof the two layers run approximately at right angles to eachother: The external intercostals extend from the tubercles ofthe ribs dorsally to the costochondral junctions ventrally, andtheir fibers are oriented obliquely, downward, and forward,from the rib above to the rib below. The internal intercostalsbegin posteriorly, because the posterior intercostal membraneon the inner aspect of the external intercostal muscles. Fromapproximately the angle of the rib, the internal intercostalmuscles run obliquely, upward, and forward from the superiorborder of the rib and costal cartilage below to the floor of thesubcostal groove of the rib and the edge of the costal cartilageabove, ending at the sternocostal junctions. All the intercostalmuscles are innervated by the intercostal nerves.
p0030 The external intercostal muscles have an inspiratory action onthe rib cage, whereas the internal intercostal muscles are expira-tory. An illustrative clinical example of the “isolated” inspiratoryaction of the intercostal muscles is offered by patients who havebilateral diaphragmatic paralysis. In these patients, inspiration isaccomplished solely by the rib cage muscles. As a result, the ribcage expands during inspiration, and the pleural pressure falls.Because the diaphragm is flaccid and no transdiaphragmatic pres-sure can be developed, the fall in pleural pressure is transmitted tothe abdomen, causing an equal fall in the abdominal pressure.Hence the abdomen moves paradoxically inward during
inspiration, opposing the inflation of the lung (Figure 8-1). Thisparadoxic motion is the cardinal sign of diaphragmatic paralysison clinical examination and is invariably present in the supineposture, during which the abdominal muscles usually remainrelaxed during the entire respiratory cycle. However, this signmay be absent in the erect posture.
s0040The Diaphragmp0040The floor of the thoracic cavity is closed by a thin musculoten-
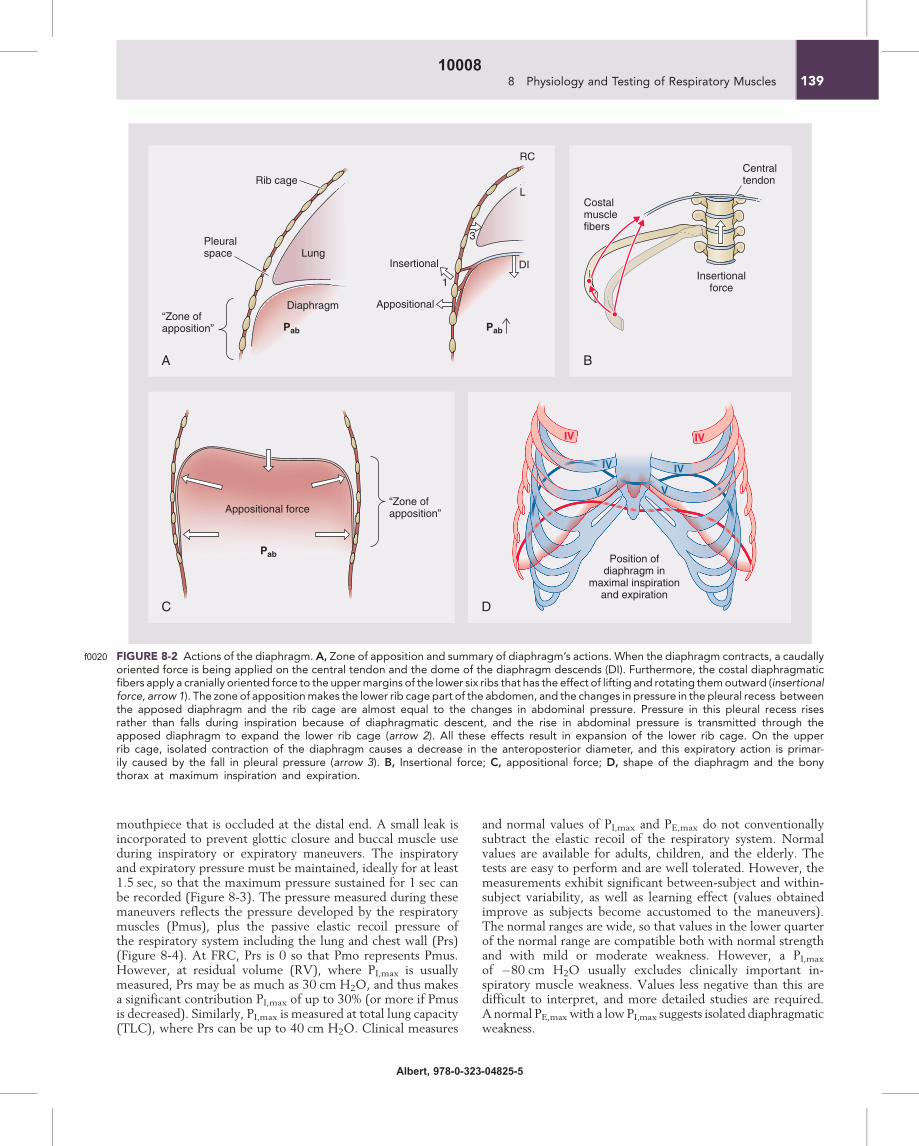
dinous sheet, the diaphragm, the most important inspiratorymuscle, accounting for approximately 70% of minute ventila-tion in normal subjects. The diaphragm is anatomically uniqueamong the skeletal muscles in that its fibers radiate from a cen-tral tendinous structure (the central tendon) to insert periph-erally into skeletal structures. The muscle of the diaphragmfalls into two main components on the basis of its point of ori-gin: the crural (vertebral) part and the costal (sternocostal)part. The crural part arises from the crura (strong, taperingtendons attached vertically to the anterolateral aspects of thebodies and intervertebral disks of the first three lumbar verteb-rae on the right and two on the left) and the three aponeuroticarcuate ligaments. The costal part of the diaphragm arises fromthe xiphoid process and the lower end of the sternum and thecostal cartilages of the lower six ribs. These costal fibers runcranially so that they are directly apposed to the inner aspectof lower rib cage, creating a zone of apposition.
p0050The shape of the relaxed diaphragm at the end of a normalexpiration (functional residual capacity, FRC) is that of twodomes joined by a saddle that runs from the sternum to theanterior surface of the spinal column (Figure 8-2). The motorinnervation of the diaphragm is from the phrenic nerves,which also provide a proprioceptive supply to the muscle.When tension develops within the diaphragmatic musclefibers, a caudally oriented force is applied on the central ten-don and the dome of the diaphragm descends; this descenthas two effects. First, it expands the thoracic cavity along itscraniocaudal axis and consequently the pleural pressure falls.Second, it produces a caudal displacement of the abdominalvisceral contents and an increase in the abdominal pressurethat in turn results in an outward motion of the ventralabdominal wall. Thus, when the diaphragm contracts, a crani-ally oriented force is being applied by the costal diaphragmaticfibers to the upper margins of the lower six ribs that has theeffect of lifting and rotating them outward (insertional force;see Figure 8-2). The actions mediated by the changes in pleuraland abdominal pressures are more complex: if one assumesthat the diaphragm is the only muscle acting on the rib cage,it seems that it has two opposing effects when it contracts.
137
10008
Albert, 978-0-323-04825-5

On the upper rib cage, it causes a decrease in the anteroposter-ior diameter, and this expiratory action is primarily because ofthe fall in pleural pressure (see Figure 8-2). On the lower ribcage, it causes an expansion. In fact this is the pattern of chestwall motion observed in tetraplegic patients with transectioninjury at the fifth cervical segment of the spinal cord or below,who have complete paralysis of the inspiratory muscles exceptfor the diaphragm. This inspiratory action on the lower ribcage is caused by the concomitant action of two differentforces, the “insertional” force already described and the “appo-sitional” force c. The Neck Muscles.Au1
s0050 The Sternocleidomastoidsp0060 The sternocleidomastoids arise from the mastoid process and
descend to the ventral surface of the manubrium sterni andthe medial third of the clavicle. Their neural supply is fromthe accessory nerve. The action of the sternocleidomastoids isto displace the sternum cranially during inspiration, to expandthe upper rib cage more in its anteroposterior diameter than inits transverse one, and to decrease the transverse diameter ofthe lower rib cage. In normal subjects breathing at rest, how-ever, the sternocleidomastoids are inactive, being recruitedonly when the inspiratory muscle pump is abnormally loadedor when ventilation increases substantially. Therefore, theyshould be considered as accessory muscles of inspiration.
s0060 The Scalenesp0070 The scalenes are composed of three muscle bundles that run
from the transverse processes of the lower five cervical verteb-rae to the upper surface of the first two ribs. They receive theirneural supply mainly from the lower five cervical segments.Their action is to increase (slightly) the anteroposterior diam-eter of the upper rib cage. Although initially considered asaccessory muscles of inspiration, they are invariably active dur-ing inspiration. In fact, seated normal subjects cannot breathewithout contracting the scalenes even when they reduce therequired inspiratory effort by reducing tidal volume. There-fore, scalenes in humans are primary muscles of inspiration,and their contraction is an important determinant of theexpansion of the upper rib cage during breathing.
s0070The Abdominal Musclesp0080The abdominal muscles with respiratory activity are those con-
stituting the ventrolateral wall of the abdomen (i.e., the rectusabdominis ventrally and the external oblique, internal oblique,and transverses abdominis laterally). They are innervated bythe lower six thoracic nerves and the first lumbar nerve. As theycontract, they pull the abdominal wall inward, thus increasingthe intraabdominal pressure. This causes the diaphragm tomovecranially into the thoracic cavity, increasing the pleural pressureand decreasing lung volume. Thus, their action is expiratory.Expiration is usually a passive process but can become activewhen minute ventilation has to be increased (e.g., during exer-cise) or during respiratory distress. Expiratory muscle action isalso essential during cough.
s0080TESTING RESPIRATORY MUSCLE FUNCTION
p0090Muscles have two functions: to develop force and to shorten.In the respiratory system, force is usually estimated as pressureand shortening as lung volume change. Thus, quantitativecharacterization of the respiratory muscles usually relies onmeasurements of volumes and pressures.
s0090Vital Capacityp0100Vital capacity (VC) is an easily obtained measurement with
spirometry, which, when decreased, points to respiratory mus-cle weakness. The VC averages approximately 50 mL/kg innormal adults. However, VC is not specific and may bedecreased because of both inspiratory and expiratory muscleweakness and restrictive lung and chest wall diseases. Amarked fall (>30%) in VC in the supine compared with theerect posture (which in the normal individual is 5–10%) isassociated with severe bilateral diaphragmatic weakness.
s0100Maximal Static Mouth Pressuresp0110Measurement of the maximum static inspiratory (PI,max) or
expiratory (PE,max) pressure that a subject can generate at themouth is a simple way to estimate inspiratory and expiratorymuscle strength. These are measured at the side port of a
Normal subject
Chest Abdomen
Patient with diaphragmatic paralysis
Chest Abdomen
FIGURE 8-1f0010 Schematic demonstration of normal abdominal and rib cage movement (left panel) and the paradoxical abdominal motion ofisolated diaphragmatic paralysis (right panel). The diaphragm at resting end-expiration is shown as a solid line and after inspiration as adashed line. In the normal subject (left), the diaphragm moves caudally, and in the patient with diaphragmatic paralysis (right), the diaphragmmoves in a cephalad direction. The anterior abdominal wall moves inward instead of outward.
138 S E C T I ON I I Physiology10008
Albert, 978-0-323-04825-5
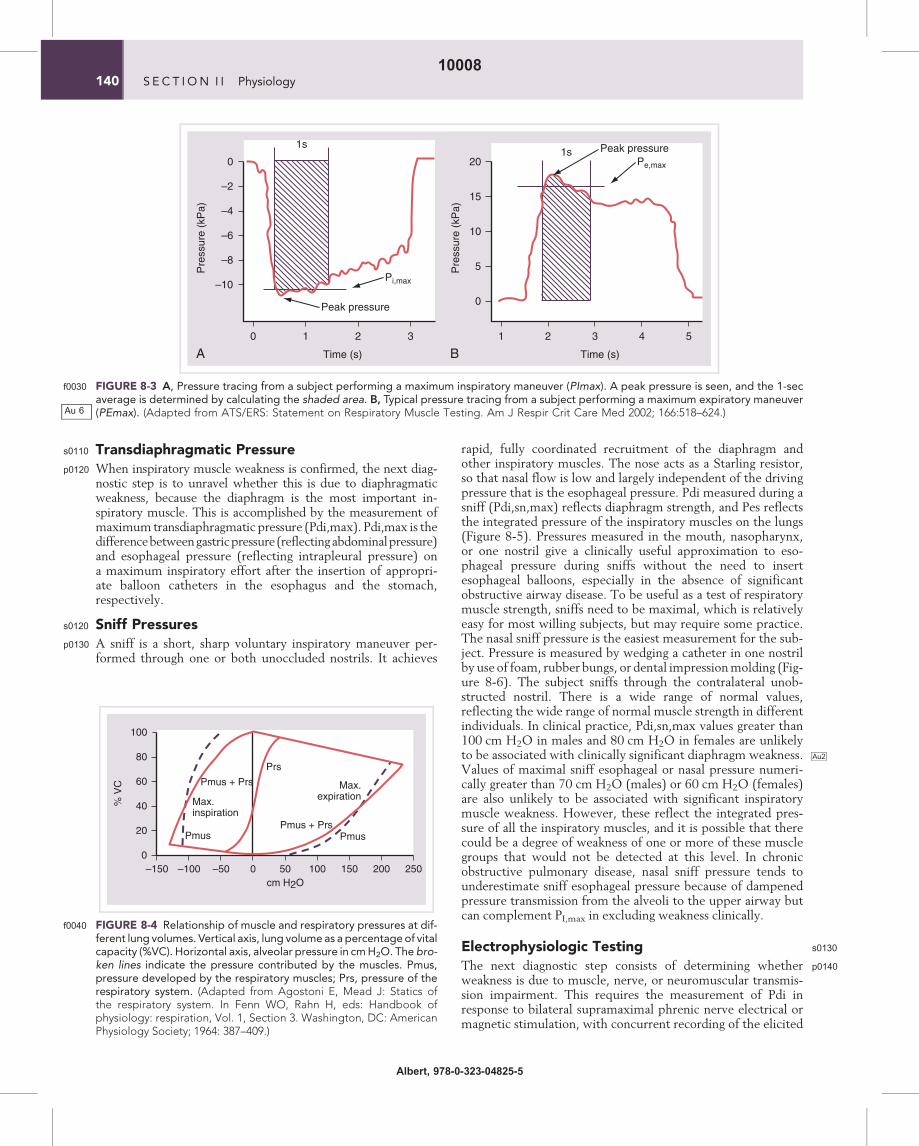
mouthpiece that is occluded at the distal end. A small leak isincorporated to prevent glottic closure and buccal muscle useduring inspiratory or expiratory maneuvers. The inspiratoryand expiratory pressure must be maintained, ideally for at least1.5 sec, so that the maximum pressure sustained for 1 sec canbe recorded (Figure 8-3). The pressure measured during thesemaneuvers reflects the pressure developed by the respiratorymuscles (Pmus), plus the passive elastic recoil pressure ofthe respiratory system including the lung and chest wall (Prs)(Figure 8-4). At FRC, Prs is 0 so that Pmo represents Pmus.However, at residual volume (RV), where PI,max is usuallymeasured, Prs may be as much as 30 cm H2O, and thus makesa significant contribution PI,max of up to 30% (or more if Pmusis decreased). Similarly, PI,max is measured at total lung capacity(TLC), where Prs can be up to 40 cm H2O. Clinical measures
and normal values of PI,max and PE,max do not conventionallysubtract the elastic recoil of the respiratory system. Normalvalues are available for adults, children, and the elderly. Thetests are easy to perform and are well tolerated. However, themeasurements exhibit significant between-subject and within-subject variability, as well as learning effect (values obtainedimprove as subjects become accustomed to the maneuvers).The normal ranges are wide, so that values in the lower quarterof the normal range are compatible both with normal strengthand with mild or moderate weakness. However, a PI,max
of �80 cm H2O usually excludes clinically important in-spiratory muscle weakness. Values less negative than this aredifficult to interpret, and more detailed studies are required.A normal PE,max with a low PI,max suggests isolated diaphragmaticweakness.
A
C D
B
Diaphragm
Lung
3
1
Insertional
Position ofdiaphragm in
maximal inspirationand expiration
Insertionalforce
IV
VV
IV
Centraltendon
Costalmusclefibers
Appositional
Rib cage
“Zone ofapposition”
“Zone ofapposition”Appositional force
Pab
Pab
Pab
DI
L
RC
Pleuralspace
IVIV
FIGURE 8-2f0020 Actions of the diaphragm. A, Zone of apposition and summary of diaphragm’s actions. When the diaphragm contracts, a caudallyoriented force is being applied on the central tendon and the dome of the diaphragm descends (DI). Furthermore, the costal diaphragmaticfibers apply a cranially oriented force to the uppermargins of the lower six ribs that has the effect of lifting and rotating themoutward (insertionalforce, arrow 1). The zone of appositionmakes the lower rib cage part of the abdomen, and the changes in pressure in the pleural recess betweenthe apposed diaphragm and the rib cage are almost equal to the changes in abdominal pressure. Pressure in this pleural recess risesrather than falls during inspiration because of diaphragmatic descent, and the rise in abdominal pressure is transmitted through theapposed diaphragm to expand the lower rib cage (arrow 2). All these effects result in expansion of the lower rib cage. On the upperrib cage, isolated contraction of the diaphragm causes a decrease in the anteroposterior diameter, and this expiratory action is primar-ily caused by the fall in pleural pressure (arrow 3). B, Insertional force; C, appositional force; D, shape of the diaphragm and the bonythorax at maximum inspiration and expiration.
1398 Physiology and Testing of Respiratory Muscles10008
Albert, 978-0-323-04825-5
s0110 Transdiaphragmatic Pressurep0120 When inspiratory muscle weakness is confirmed, the next diag-
nostic step is to unravel whether this is due to diaphragmaticweakness, because the diaphragm is the most important in-spiratory muscle. This is accomplished by the measurement ofmaximumtransdiaphragmatic pressure (Pdi,max). Pdi,max is thedifferencebetweengastricpressure(reflectingabdominalpressure)and esophageal pressure (reflecting intrapleural pressure) ona maximum inspiratory effort after the insertion of appropri-ate balloon catheters in the esophagus and the stomach,respectively.
s0120 Sniff Pressuresp0130 A sniff is a short, sharp voluntary inspiratory maneuver per-
formed through one or both unoccluded nostrils. It achieves
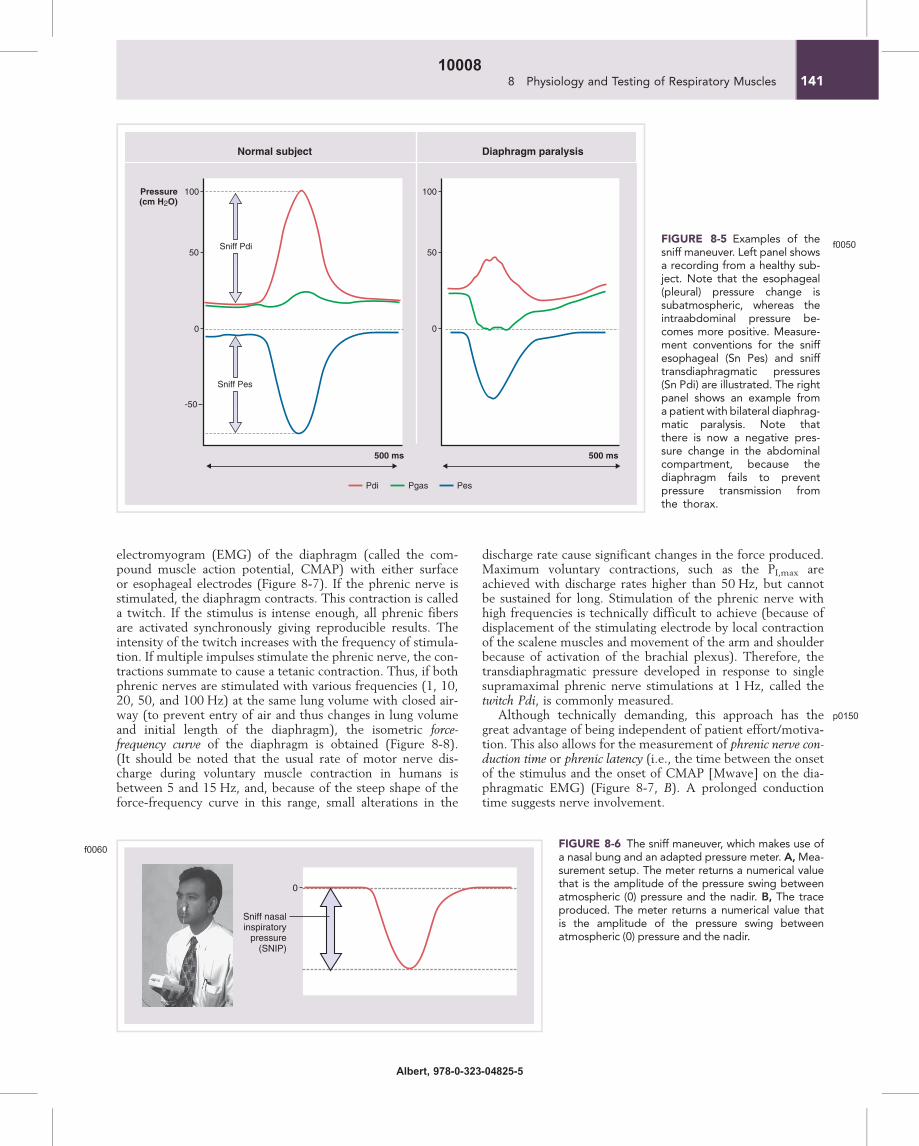
rapid, fully coordinated recruitment of the diaphragm andother inspiratory muscles. The nose acts as a Starling resistor,so that nasal flow is low and largely independent of the drivingpressure that is the esophageal pressure. Pdi measured during asniff (Pdi,sn,max) reflects diaphragm strength, and Pes reflectsthe integrated pressure of the inspiratory muscles on the lungs(Figure 8-5). Pressures measured in the mouth, nasopharynx,or one nostril give a clinically useful approximation to eso-phageal pressure during sniffs without the need to insertesophageal balloons, especially in the absence of significantobstructive airway disease. To be useful as a test of respiratorymuscle strength, sniffs need to be maximal, which is relativelyeasy for most willing subjects, but may require some practice.The nasal sniff pressure is the easiest measurement for the sub-ject. Pressure is measured by wedging a catheter in one nostrilby use of foam, rubber bungs, or dental impressionmolding (Fig-ure 8-6). The subject sniffs through the contralateral unob-structed nostril. There is a wide range of normal values,reflecting the wide range of normal muscle strength in differentindividuals. In clinical practice, Pdi,sn,max values greater than100 cm H2O in males and 80 cm H2O in females are unlikelyto be associated with clinically significant diaphragm weakness Au2.Values of maximal sniff esophageal or nasal pressure numeri-cally greater than 70 cm H2O (males) or 60 cm H2O (females)are also unlikely to be associated with significant inspiratorymuscle weakness. However, these reflect the integrated pres-sure of all the inspiratory muscles, and it is possible that therecould be a degree of weakness of one or more of these musclegroups that would not be detected at this level. In chronicobstructive pulmonary disease, nasal sniff pressure tends tounderestimate sniff esophageal pressure because of dampenedpressure transmission from the alveoli to the upper airway butcan complement PI,max in excluding weakness clinically.
s0130Electrophysiologic Testingp0140The next diagnostic step consists of determining whether
weakness is due to muscle, nerve, or neuromuscular transmis-sion impairment. This requires the measurement of Pdi inresponse to bilateral supramaximal phrenic nerve electrical ormagnetic stimulation, with concurrent recording of the elicited
0
–2
–4
–6
–8
–10
0 1 2 3
Time (s)
Pre
ssur
e (k
Pa)
1s
Pi,max
Peak pressure
Pre
ssur
e (k
Pa)
1s Peak pressurePe,max
Time (s)
20
15
10
5
0
1 2 3 4 5
BA
FIGURE 8-3f0030 A, Pressure tracing from a subject performing a maximum inspiratory maneuver (PImax). A peak pressure is seen, and the 1-secaverage is determined by calculating the shaded area. B, Typical pressure tracing from a subject performing a maximum expiratory maneuver(PEmax). (Adapted from ATS/ERS: Statement on Respiratory Muscle Testing. Am J Respir Crit Care Med 2002; 166:518–624.)
80
60
40
20
0–150 –100 –50 0 50 100 150 200 250
Pmus + Prs
Max.expirationMax.
inspiration
Pmus
Pmus + Prs
Prs
Pmus
% V
C
cm H2O
100
FIGURE 8-4f0040 Relationship of muscle and respiratory pressures at dif-ferent lung volumes. Vertical axis, lung volume as a percentageof vitalcapacity (%VC). Horizontal axis, alveolar pressure in cmH2O. The bro-ken lines indicate the pressure contributed by the muscles. Pmus,pressure developed by the respiratory muscles; Prs, pressure of therespiratory system. (Adapted from Agostoni E, Mead J: Statics ofthe respiratory system. In Fenn WO, Rahn H, eds: Handbook ofphysiology: respiration, Vol. 1, Section 3. Washington, DC: AmericanPhysiology Society; 1964: 387–409.)
Au 6
140 S E C T I ON I I Physiology10008
Albert, 978-0-323-04825-5
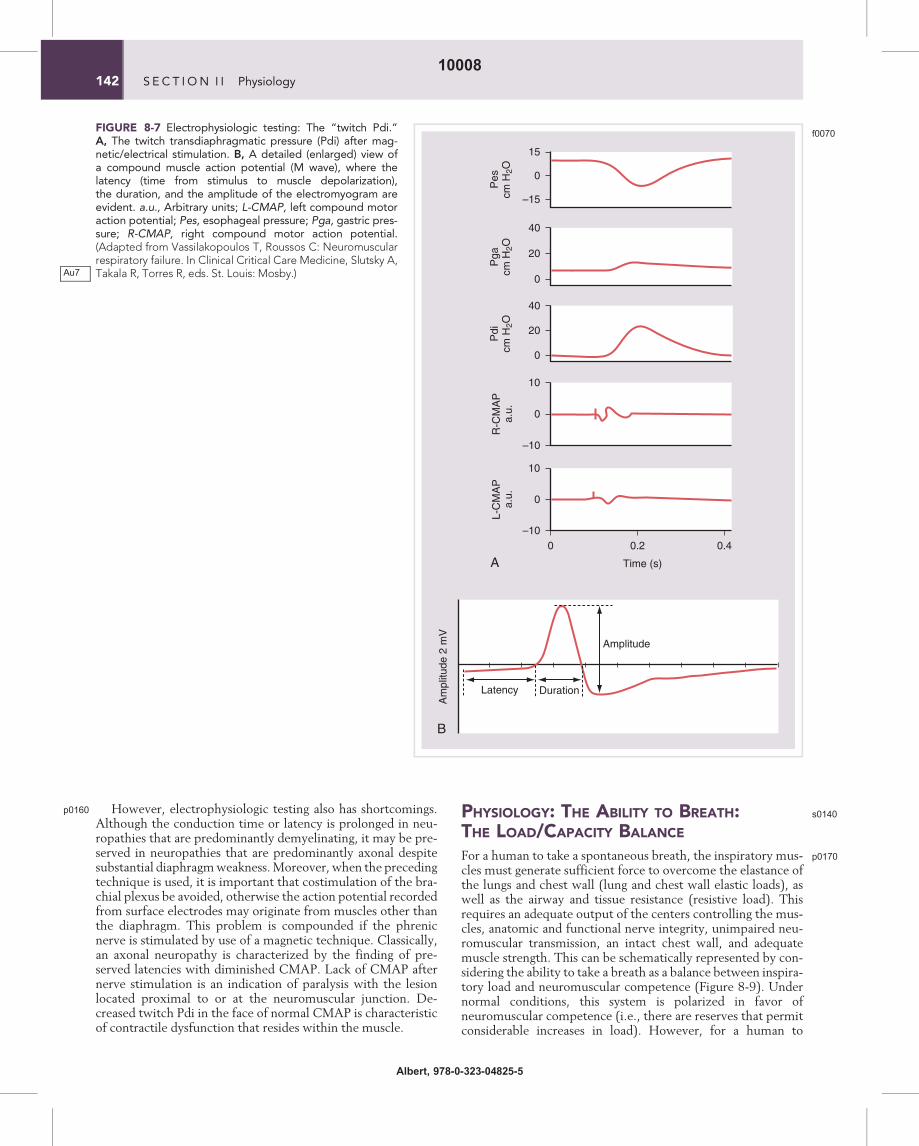
electromyogram (EMG) of the diaphragm (called the com-pound muscle action potential, CMAP) with either surfaceor esophageal electrodes (Figure 8-7). If the phrenic nerve isstimulated, the diaphragm contracts. This contraction is calleda twitch. If the stimulus is intense enough, all phrenic fibersare activated synchronously giving reproducible results. Theintensity of the twitch increases with the frequency of stimula-tion. If multiple impulses stimulate the phrenic nerve, the con-tractions summate to cause a tetanic contraction. Thus, if bothphrenic nerves are stimulated with various frequencies (1, 10,20, 50, and 100 Hz) at the same lung volume with closed air-way (to prevent entry of air and thus changes in lung volumeand initial length of the diaphragm), the isometric force-frequency curve of the diaphragm is obtained (Figure 8-8).(It should be noted that the usual rate of motor nerve dis-charge during voluntary muscle contraction in humans isbetween 5 and 15 Hz, and, because of the steep shape of theforce-frequency curve in this range, small alterations in the
discharge rate cause significant changes in the force produced.Maximum voluntary contractions, such as the PI,max areachieved with discharge rates higher than 50 Hz, but cannotbe sustained for long. Stimulation of the phrenic nerve withhigh frequencies is technically difficult to achieve (because ofdisplacement of the stimulating electrode by local contractionof the scalene muscles and movement of the arm and shoulderbecause of activation of the brachial plexus). Therefore, thetransdiaphragmatic pressure developed in response to singlesupramaximal phrenic nerve stimulations at 1 Hz, called thetwitch Pdi, is commonly measured.
p0150Although technically demanding, this approach has thegreat advantage of being independent of patient effort/motiva-tion. This also allows for the measurement of phrenic nerve con-duction time or phrenic latency (i.e., the time between the onsetof the stimulus and the onset of CMAP [Mwave] on the dia-phragmatic EMG) (Figure 8-7, B). A prolonged conductiontime suggests nerve involvement.
Normal subject Diaphragm paralysis
Pressure(cm H2O)
500 ms
100
0
500 ms
50
-50
Pgas PesPdi
Sniff Pdi
Sniff Pes
100
50
0
FIGURE 8-5f0050
Examples of thesniff maneuver. Left panel showsa recording from a healthy sub-ject. Note that the esophageal(pleural) pressure change issubatmospheric, whereas theintraabdominal pressure be-comes more positive. Measure-ment conventions for the sniffesophageal (Sn Pes) and snifftransdiaphragmatic pressures(Sn Pdi) are illustrated. The rightpanel shows an example froma patient with bilateral diaphrag-matic paralysis. Note thatthere is now a negative pres-sure change in the abdominalcompartment, because thediaphragm fails to preventpressure transmission fromthe thorax.
Sniff nasalinspiratory
pressure(SNIP)
0
FIGURE 8-6f0060
The sniff maneuver, which makes use ofa nasal bung and an adapted pressure meter. A,Mea-surement setup. The meter returns a numerical valuethat is the amplitude of the pressure swing betweenatmospheric (0) pressure and the nadir. B, The traceproduced. The meter returns a numerical value thatis the amplitude of the pressure swing betweenatmospheric (0) pressure and the nadir.
1418 Physiology and Testing of Respiratory Muscles10008
Albert, 978-0-323-04825-5
p0160 However, electrophysiologic testing also has shortcomings.Although the conduction time or latency is prolonged in neu-ropathies that are predominantly demyelinating, it may be pre-served in neuropathies that are predominantly axonal despitesubstantial diaphragmweakness.Moreover, when the precedingtechnique is used, it is important that costimulation of the bra-chial plexus be avoided, otherwise the action potential recordedfrom surface electrodes may originate from muscles other thanthe diaphragm. This problem is compounded if the phrenicnerve is stimulated by use of a magnetic technique. Classically,an axonal neuropathy is characterized by the finding of pre-served latencies with diminished CMAP. Lack of CMAP afternerve stimulation is an indication of paralysis with the lesionlocated proximal to or at the neuromuscular junction. De-creased twitch Pdi in the face of normal CMAP is characteristicof contractile dysfunction that resides within the muscle.
s0140PHYSIOLOGY: THE ABILITY TO BREATH:THE LOAD/CAPACITY BALANCE
p0170For a human to take a spontaneous breath, the inspiratory mus-cles must generate sufficient force to overcome the elastance ofthe lungs and chest wall (lung and chest wall elastic loads), aswell as the airway and tissue resistance (resistive load). Thisrequires an adequate output of the centers controlling the mus-cles, anatomic and functional nerve integrity, unimpaired neu-romuscular transmission, an intact chest wall, and adequatemuscle strength. This can be schematically represented by con-sidering the ability to take a breath as a balance between inspira-tory load and neuromuscular competence (Figure 8-9). Undernormal conditions, this system is polarized in favor ofneuromuscular competence (i.e., there are reserves that permitconsiderable increases in load). However, for a human to
Amplitude
DurationLatency
Time (s)
Am
plitu
de 2
mV
B
A0 0.2 0.4
10
0
–10
L-C
MA
Pa.
u.R
-CM
AP
a.u.
10
0
–10
Pdi
cm H
2O
40
20
0
40
20
0
Pga
cm H
2O
15
0
–15
Pes
cm H
2O
FIGURE 8-7f0070
Electrophysiologic testing: The “twitch Pdi.”A, The twitch transdiaphragmatic pressure (Pdi) after mag-netic/electrical stimulation. B, A detailed (enlarged) view ofa compound muscle action potential (M wave), where thelatency (time from stimulus to muscle depolarization),the duration, and the amplitude of the electromyogram areevident. a.u., Arbitrary units; L-CMAP, left compound motoraction potential; Pes, esophageal pressure; Pga, gastric pres-sure; R-CMAP, right compound motor action potential.(Adapted from Vassilakopoulos T, Roussos C: Neuromuscularrespiratory failure. In Clinical Critical Care Medicine, Slutsky A,Takala R, Torres R, eds. St. Louis: Mosby.)Au7
142 S E C T I ON I I Physiology10008
Albert, 978-0-323-04825-5

breathe spontaneously, the inspiratorymuscles should be able tosustain the aforementioned load over time as well as adjust theminute ventilation in such a way that there is adequate gasexchange. The ability of the respiratory muscles to sustain thisload without the appearance of fatigue is called endurance andis determined by the balance between energy supplies andenergy demands (Figure 8-10).
p0180Energy supplies depend on the inspiratory muscle bloodflow, the blood substrate (fuel) concentration and arterial oxy-gen content, the muscle’s ability to extract and use energysources, and the muscle’s energy stores. Under normal circum-stances, energy supplies are adequate to meet the demands,and a large recruitable reserve exists (see Figure 8-10). Energydemands increase proportionally with the mean pressuredeveloped by the inspiratory muscles per breath (PI) expressedas a fraction of maximum pressure that the respiratory musclescan voluntarily develop (PI/PI,max), the minute ventilation(VE), the inspiratory duty cycle (TI/TTOT), and the meaninspiratory flow rate (VT/TI) and are inversely related to theefficiency of the muscles. Fatigue develops when the mean rateof energy demands exceeds the mean rate of energy supply(i.e., when the balance is polarized in favor of demands).
p0190The product of TI/TTOT and the mean transdiaphragmaticpressure expressed as a fraction of maximal (Pdi/Pdi,max)defines a useful “tension-time index” (TTIdi) that is related tothe endurance time (i.e., the time that the diaphragm can
Force(% maximum)
80
60
40
20
100
0
Stimulation frequency20 40 60 80 60
Fatigued muscle Fresh muscle
0
FIGURE 8-8f0080 Force-frequency relationship of in vivo human respira-toryAu4 muscles. The force-frequency curve of the fresh muscle is shownin red, and the relationship after a fatiguing task is shown in blue; adisproportionate force loss at low stimulation frequencies isobserved. (Adapted from Moxham J, Wiles CM, Newham D,Edwards RHT: Ciba Found Symp 1981; 82:197–212.)
Efficiency
(VT/tl)
V’E
(tl/ttot)
PI/PI,max
Blood substrateconcentration
Energy demands
Energy supplies
Arterial oxygencontent
Energy stores/nutrition
Ability to extractenergy sources
Inspiratory muscleblood flow
FIGURE 8-10 f0100Balance between energy supplies and energydemands. Respiratory muscle endurance is determined by the bal-ance between energy supplies and demands. Normally, the suppliesmeet the demands, and a large reserve exists. Whenever this bal-ance weighs in favor of demands, the respiratory muscles ultimatelybecome fatigued, leading to inability to sustain spontaneous Au8
breathing decreased. VT/tI, Mean inspiratory flow (tidal volume/inspiratory time); tI/ttot, duty cycle (fraction of inspiration to totalbreathing cycle duration); PI/PI, max, inspiratory pressure/maximuminspiratory pressure ratio; V 0E, minute ventilation. (Adapted fromVassilakopoulos T, Roussos C: Neuromuscular respiratory failure.In Slutsky A, Takala R, Torres, eds. Clinical critical care medicine.St. Louis: Mosby.)
Lung elastic loads
Resistive loads
Chest wallelastic loads
Central drive
Load (per breath)(PI)
Neuromuscularcompetence
(PI, max)
Muscle strength
Neural andneuromusculartransmission
FIGURE 8-9f0090 Balance between inspiratory load and neuromuscularcompetence. The ability to take a spontaneous breath is deter-mined by the balance between the load imposed on the respiratorysystem (pressure developed by the inspiratory muscles; PI) and theneuromuscular competence of the ventilatory pump (maximuminspiratory pressure; PI, max). Normally, this balance weighs in favorof competence, permitting significant increases in load. However, ifthe competence is, for whatever reason, reduced below a criticalpoint (e.g., drug overdose, myasthenia gravis), the balance maythen weigh in favor of load, rendering the ventilatory pump insuffi-cient to inflate the lungs and chest wall. (Adapted from Vassilakopou-los T, Roussos C: Neuromuscular respiratory failure. In Slutsky A,Takala R, Torres, eds. Clinical critical care medicine. St. Louis: Mosby.)
1438 Physiology and Testing of Respiratory Muscles10008
Albert, 978-0-323-04825-5

sustain the load imposed on it). Whenever TTIdi is smallerthan the critical value of 0.15, the load can be sustained indefi-nitely; but when TTIdi exceeds the critical zone of 0.15–0.18,the load can be sustained only for a limited time period, in otherwords, the endurance time. This was found to be inverselyrelated to TTIdi. The TTI concept is assumed to be applicablenot only to the diaphragm but also to the respiratory musclesas a whole:
TTI ¼ PI=PI;maxwTI=TTOT
p0210 Because endurance is determined by the balance betweenenergy supply and demand, TTI of the inspiratory muscleshas to be in accordance with the energy balance view. In fact,as Figure 8-4 demonstrates, PI/PI,max and TI/TTOT, which con-stitute the TTI, are among the determinants of energydemands; an increase in either that will increase the TTI valuewill also increase the demands. But what determines the ratioPI/PI,max? The nominator, the mean inspiratory pressure devel-oped per breath, is determined by the elastic and resistiveloads imposed on the inspiratory muscles. The denominator,the maximum inspiratory pressure, is determined by the neuro-muscular competence (i.e., the maximum inspiratory muscleactivation that can be voluntarily achieved). It follows, then,that the value of PI/PI,max is determined by the balance betweenload and competence (see Figure 8-9). But PI /PI,max is also oneof the determinants of energy demands (see Figure 8-10);therefore, the two balances (i.e., between load and competenceand energy supply and demand) are in essence linked, creating a
system (Figure 8-11). Schematically, when the central hingeof the system moves upward, or is at least at the horizontallevel, spontaneous ventilation can be sustained indefinitely(see Figure 8-11). The ability of a subject to breathe spontane-ously depends on the fine interplay of many different factors.Normally, this interplay moves the central hinge far upwardand creates a great ventilatory reserve for the healthy individ-ual. When the central hinge of the system, for whatever reason,moves downward, spontaneous ventilation cannot be sustained,and ventilatory failure ensues.
s0150Hyperinflation
p0220Hyperinflation (frequently observed in obstructive airway dis-eases) compromises the force-generating capacity of the dia-phragm for a variety of reasons: First, the respiratory muscles,like other skeletal muscles, obey the length-tension relation-ship. At any given level of activation, changes in musclefiber length alter tension development. This is because theforce-tension developed by a muscle depends on the interactionbetween actin and myosin fibrils (i.e., the number of myosinheads attaching and thus pulling the actin fibrils closer withineach sarcomere). The optimal fiber length (Lo) wheretension is maximal is the length at which all myosin headsattach and pull the actin fibrils. Below this length (as withhyperinflation, which shortens the diaphragm), actin-myosininteraction becomes suboptimal, and tension developmentdeclines. Second, as lung volume increases, the zone of apposi-tion of the diaphragm decreases in size, and a larger fraction of
Blood substrateconcentration
Arterial oxygencontent
Energy stores/nutrition
Ability to extractenergy sources
Inspiratory muscleblood flow
Energy supplies
Efficiency↓
(VT, tl)
VE
(tl/tlot)
Pl/Pt, max
Lung elastic loads
Resistive loads
Chest wallelastic loads
Energy demands
Load
Central drive
Muscle strength
Neural and neuromusculartransmission
Competence
FIGURE 8-11f0110 System of two balances: load and competence, energy supplies and demands. The system of two balances, incorporating thevarious determinants of load, competence, energy supplies, and demands is represented schematically. The PI/PI, max, one of the determi-nants of energydemands (see Figure 8-10) is replacedby its equivalent: the balancebetween load andneuromuscular competence (see Figure8-9). In fact, this is the reason the two balances are linked.When the central hinge of the systemmoves upward or is at least at the horizontal level, abalance exists between ventilatory needs and neurorespiratory capacity, and spontaneous ventilation can be sustained. In healthy persons, thehinge moves far upward, creating a large reserve. For abbreviations, see legends to Figures 8-9 and 8-10. (Adapted from Vassilakopoulos T,Roussos C: Neuromuscular respiratory failure. In Slutsky A, Takala R, Torres R, eds. Clinical critical care medicine. St. Louis: Mosby.)
144 S E C T I ON I I Physiology10008
Albert, 978-0-323-04825-5

the rib cage becomes exposed to pleural pressure. Hence, thediaphragm’s inspiratory action on the rib cage diminishes.When lung volume approaches total lung capacity, the zoneof apposition all but disappears (Figure 8-12), and the dia-phragmatic muscle fibers become oriented horizontally inter-nally (Figure 8-12, B). The insertional force of the diaphragmis then expiratory, rather than inspiratory, in direction. Thisexplains the inspiratory decrease in the transverse diameter ofthe lower rib cage in subjects with emphysema and severehyperinflation (Hoover’s sign). Third, the resultant flatteningof the diaphragm increases its radius of curvature (Rdi) (seeFigure 8-12, B) and, according to Laplace’s law, Pdi ¼ 2Tdi/Rdi, diminishes its pressure-generating capacity (Pdi) for thesame tension development (Tdi).
s0160 RESPIRATORY MUSCLE RESPONSES
TO CHANGES IN LOAD
s0170 Acute Responsess0180 Increased Load
s0190
Respiratory Muscle Fatigue.p0230 Fatigue is defined as the loss ofcapacity to develop force and/or velocity in response to a loadthat is reversible by rest. Thus, fatigue may be present beforethe point at which a muscle is unable to continue to performa particular task (task failure). In applying this concept to theinspiratory muscles, one could conclude that they may befatigued before there is hypercapnia because of their inabilityto continue to generate sufficient pressure to maintain alveolarventilation.
p0240 Fatigue should be distinguished from weakness in whichreduced force generation is fixed and not reversed by rest,although the presence of weakness may itself predispose amuscle to fatigue. The site and mechanisms of fatigue remaincontroversial. Theoretically, the site of fatigue may be locatedat any link in the long chain of events involved in voluntarymuscle contraction leading from the brain to the contractilemachinery. A widely used convention is to classify fatigue as
central fatigue, peripheral high-frequency fatigue, or peripherallow-frequency fatigue.
s0200
Central Fatigue. p0250Central fatigue is present when a maximalvoluntary contraction generates less force than does maximalelectrical stimulation. If maximal electrical stimulation super-imposed on a maximal voluntary contraction can potentiatethe force generated by a muscle, a component of centralfatigue exists. This procedure applied to the diaphragm con-sists of the twitch occlusion test that may separate central fromperipheral fatigue. This test examines the transdiaphragmaticpressure (Pdi) response to bilateral phrenic nerve stimulationsuperimposed on graded voluntary contractions of the dia-phragm. Normally, the amplitude of the Pdi twitches inresponse to phrenic nerve stimulation decreases as the volun-tary Pdi increases. During Pdi,max, no superimposed twitchescan be detected. When diaphragmatic fatigue is present, super-imposed twitches can be demonstrated. A number of experi-ments have suggested that a form of central diaphragmatic“fatigue” may develop during respiratory loading so that, atthe limits of diaphragmatic endurance, a significant portionof the reduction in force production is due to failure of thecentral nervous system to completely activate the diaphragm.Central fatigue may be caused by a reduction in the numberof motor units that can be recruited by the motor drive or bya decrease in motor unit discharge rates or both. The observeddecreased central firing rate during fatigue may, in fact, be abeneficial adaptive response preventing the muscle’s self-destruction by excessive activation.
s0210
Peripheral Fatigue. p0260Peripheral fatigue refers to failure at theneuromuscular junction or distal to this structure and is pres-ent when muscle force output falls in response to direct electri-cal stimulation. This type of fatiguemay occur because of failureof impulse propagation across the neuromuscular junction,the muscle surface membrane or the t tubules (transmissionfatigue), impaired excitation—contraction coupling, or failureof the contractile apparatus of the muscle fibers (because ofalterations in metabolism or changes in contractile proteins).
Ppl Ppl
Diaphragm(hyperinflated)
A B
Upperrib cage
Ppl
Diaphragm
Zone ofapposition
Lowerrib cage
ThoraxThorax
Decreased zoneof apposition
ShortenedShortenedmuscle fibersmuscle fibersShortenedmuscle fibers
3
1Insertionalforce
Appositionalforce
Decreasedcurvature
Medial orientation ofdiaphragmatic fibers
2DI
Pap
FIGURE 8-12 f0120Consequences of hyperinflation on the diaphragm. A, Normal actions of the diaphragm as in Figure 8-2. B, Deleterious effectsof hyperinflation on the diaphragm.
1458 Physiology and Testing of Respiratory Muscles10008
Albert, 978-0-323-04825-5
Peripheral fatigue can be further classified into high-frequencyand low-frequency fatigue on the basis of the shape of the muscleforce-frequency curve (Figure 8-8). High-frequency fatigueresults in depression of the forces generated by a muscle inresponse to high-frequency electrical stimulation (50–100 Hz),whereas low-frequency fatigue results in depression of forcegeneration in response to low-frequency stimuli (1–20 Hz).Low-frequency fatigue can occur in isolation, but high-frequency fatigue is invariably associated with some alterationsin muscle force generation at lower frequencies.
s0220
High-Frequency Fatigue.p0270 During artificial stimulation of amotor neuron, especially at high frequencies, muscle forcedeclines rapidly in association with the decline in CMAPamplitude. This response, known as “high-frequency fatigue”(see Figure 8-8), is attributed to transmission fatigue. The siteof this type of fatigue may be located postsynaptically (from adecrease in sarcolemmal membrane endplate excitability or areduction in action potential propagation into the t tubularsystem) or presynaptically (probably in fine terminal filamentsof the motor nerve or less frequently from depletion of synap-tic transmitter substance). Teleologically, transmission blockcould be beneficial in some instances by protecting the muscleagainst excessive depletion of its ATP stores, which would leadto rigor mortis. Normal subjects breathing against high-intensityinspiratory resistive loads develop high-frequency fatigue,which resolves very quickly after cessation of the strenuousdiaphragmatic contractions.
s0230
Low-Frequency Fatigue.p0280 All processes that link the electricalactivation of the muscle fiber and the various metabolic andenzymatic processes providing energy to the contractilemachinery are called excitation-contraction coupling pro-cesses. Impaired excitation contraction coupling is thought tobe responsible when the loss of force is not accompanied bya parallel decline in the electrical activity. This type of fatigueis characterized by a selective loss of force at low frequenciesof stimulation (low-frequency fatigue) (see Figure 8-8) despitemaintenance of the force generated at high frequencies ofstimulation, indicating that the contractile proteins continueto generate force provided that sufficient calcium is releasedby the sarcoplasmic reticulum. The mechanism of this typeof fatigue is not understood. It may occur because of a reducedlevel of calcium availability caused by alterations in sarcoplas-mic reticulum function or a reduction in the calcium sensitiv-ity of the myofilaments (troponin binding site) at submaximalcalcium concentrations (because of high hydrogen and phos-phate ion concentration). These defects would reduce theforce developed at low-frequency stimulation. In contrast, athigher stimulation frequencies, a relatively normal force canbe generated when the interior of the fiber is saturated withcalcium.
p0290 This type of fatigue is characteristically long lasting, taking sev-eral hours to recover. Low-frequency fatigue occurs during high-force contractions and is less likely to develop when the forcesgenerated are smaller, even if these aremaintained for longer timeperiods, thereby achieving the same total work. As previouslystated, fatigue develops when the mean rate of energy demandsexceeds the mean rate of energy supply to the muscle (see Fig-ure 8-10), resulting in depletion of muscle energy stores, pHchanges from lactate accumulation, and excessive production ofoxygen-derived free radicals. However, the exact interplay of allthese factors is not yet identified in either the diaphragm or the
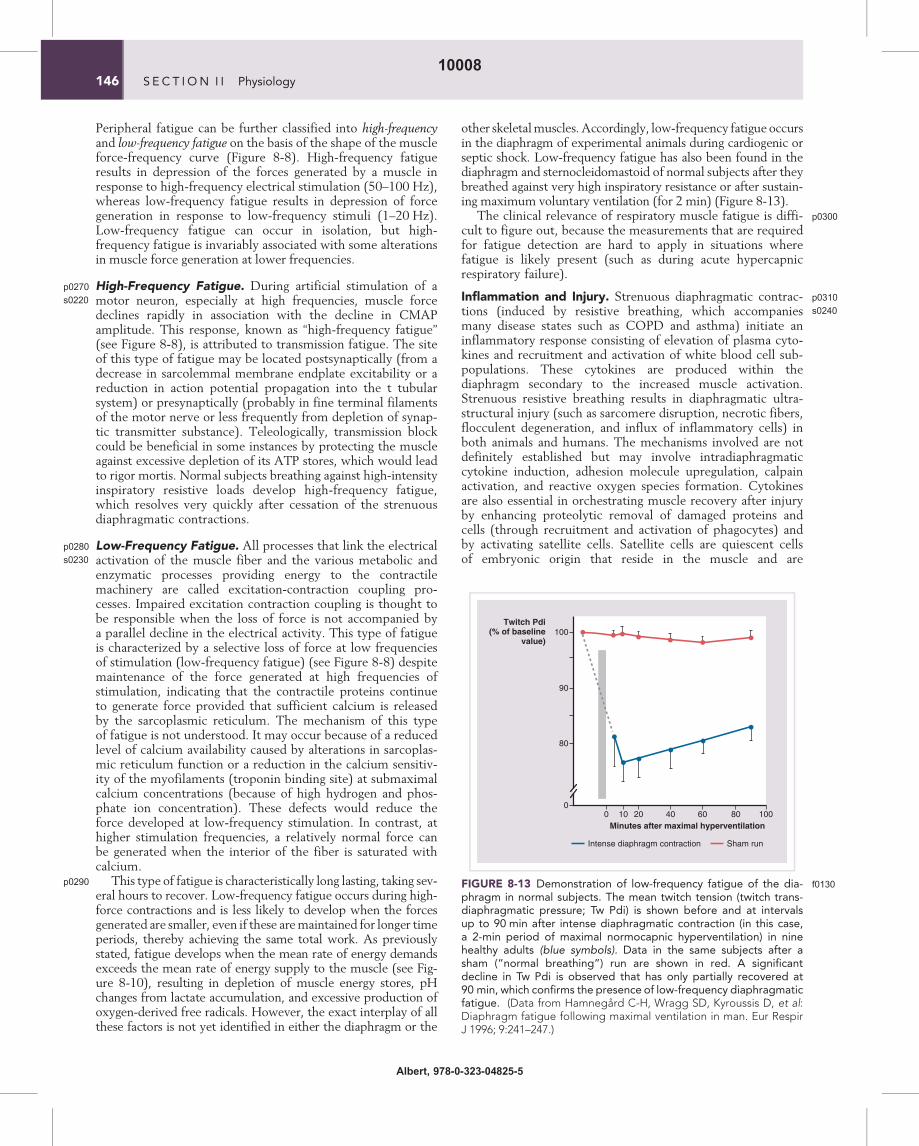
other skeletalmuscles. Accordingly, low-frequency fatigue occursin the diaphragm of experimental animals during cardiogenic orseptic shock. Low-frequency fatigue has also been found in thediaphragm and sternocleidomastoid of normal subjects after theybreathed against very high inspiratory resistance or after sustain-ing maximum voluntary ventilation (for 2 min) (Figure 8-13).
p0300The clinical relevance of respiratory muscle fatigue is diffi-cult to figure out, because the measurements that are requiredfor fatigue detection are hard to apply in situations wherefatigue is likely present (such as during acute hypercapnicrespiratory failure).
s0240
Inflammation and Injury. p0310Strenuous diaphragmatic contrac-tions (induced by resistive breathing, which accompaniesmany disease states such as COPD and asthma) initiate aninflammatory response consisting of elevation of plasma cyto-kines and recruitment and activation of white blood cell sub-populations. These cytokines are produced within thediaphragm secondary to the increased muscle activation.Strenuous resistive breathing results in diaphragmatic ultra-structural injury (such as sarcomere disruption, necrotic fibers,flocculent degeneration, and influx of inflammatory cells) inboth animals and humans. The mechanisms involved are notdefinitely established but may involve intradiaphragmaticcytokine induction, adhesion molecule upregulation, calpainactivation, and reactive oxygen species formation. Cytokinesare also essential in orchestrating muscle recovery after injuryby enhancing proteolytic removal of damaged proteins andcells (through recruitment and activation of phagocytes) andby activating satellite cells. Satellite cells are quiescent cellsof embryonic origin that reside in the muscle and are
Twitch Pdi(% of baseline
value)
Minutes after maximal hyperventilation0 2010 40 60 80 100
90
80
100
Intense diaphragm contraction Sham run
0
FIGURE 8-13 f0130Demonstration of low-frequency fatigue of the dia-phragm in normal subjects. The mean twitch tension (twitch trans-diaphragmatic pressure; Tw Pdi) is shown before and at intervalsup to 90 min after intense diaphragmatic contraction (in this case,a 2-min period of maximal normocapnic hyperventilation) in ninehealthy adults (blue symbols). Data in the same subjects after asham (“normal breathing”) run are shown in red. A significantdecline in Tw Pdi is observed that has only partially recovered at90 min, which confirms the presence of low-frequency diaphragmaticfatigue. (Data from Hamnegard C-H, Wragg SD, Kyroussis D, et al:Diaphragm fatigue following maximal ventilation in man. Eur RespirJ 1996; 9:241–247.)
146 S E C T I ON I I Physiology10008
Albert, 978-0-323-04825-5

transformed into myocytes, when the muscle becomes injured,to replace damaged myocytes.
s0250 Chronic Responsess0260 Increased Load
s0270
Plasticity and Adaptation.p0320 The respiratory muscles are plas-tic organs that respond to chronic changes of the load theyare facing and thus of their activity with structural and func-tional changes/adaptations.
p0330COPD is the paradigm of a disease with chronicallyincreased respiratory muscle load. A major adaptation of therespiratory muscles is fiber type transformation. The myosinheavy chain component of the myosin molecule constitutesthe basis for the classification of muscle fibers as either (type I)or (type II) (Figure 8-14, A). Myosin heavy chain exists invarious isoforms, which in increasing order of maximumshortening velocity are myosin heavy chain (MHC) I, IIa,and IIb, the latter being the faster (Figure 8-14, C ). The
A
B
C
FF
IIb
IIb IIa
I
IIa I
MHC2B MHC2A MHCSlow
FR S
Time
Fo
rce
Time
Fo
rce
Time
Fo
rce
Siz
e
Fatigue resistance
Speed
FIGURE 8-14 f0140Properties of skeletal muscle fiber types. Different fiber types in the diaphragm muscle are distinguished by size, myosin heavychain content, contractile characteristics (force and speed of contraction), and fatigue resistance (type S, slow; type FR, fast-twitch, fatigueresistant; and type FF motor units, fast-twitch, fatigable), as well as myosin heavy chain (MHC) isoform expression (MHCSlow, MHC2A, andMHC2B). A, Size; B, force; C, size, speed of contraction, fatigue resistance. (Adapted from Mantilla CB, Sieck GS: Mechanisms underlyingmotor unit plasticity in the respiratory system. J Appl Physiol 2003; 94:1230–1241; and Jones DA: Skeletal muscle physiology. In Roussos C,ed: The Thorax, 2nd ed. New York: Marcel Dekker; 3–32).
1478 Physiology and Testing of Respiratory Muscles10008
Albert, 978-0-323-04825-5

diaphragm in healthy humans is composed of approximately50% type I fatigue-resistant fibers, 25% type IIa, and 25% typeIIb. There are twoways in whichmuscles canmodify their over-all MHC phenotype: preferential atrophy/hypertrophy of fiberscontaining a specific MHC isoform and actual transformationfrom one fiber type to another. In COPD, there is a transforma-tion of type II to type I fibers, resulting in a great predominanceof type I fatigue-resistant fibers. This increases the resistanceof the diaphragm to fatigue development, but at the same timecompromises the force-generating capacity, because type Ifibers can generate less force than type II fibers.
p0340 Adaptation is not only restricted to fiber type transforma-tion. In an animal model of COPD (emphysematous ham-sters), the number and the length of sarcomeres decrease,resulting in a leftward shift of the length-tension curve, so thatthe muscle adapts to the shorter operating length induced byhyperinflation. These alterations may help restore the mechan-ical advantage of the diaphragm in chronically hyperinflatedstates. In humans, this adaptation seems to occur by sarcomerelength shortening.
s0280 Inactivity-Unloading
p0350 Respiratory muscles do not only adapt when they functionagainst increased load but also when they become inactive, ashappens during denervation or when a mechanical ventilatorundertakes their role as force generator to create the drivingpressure permitting airflow into the lungs. Inactivity andunloading of the diaphragm caused by mechanical ventilationis harmful, resulting in decreased diaphragmatic force-
generating capacity, diaphragmatic atrophy, and diaphragmaticinjury, which are described by the term ventilator-induced dia-phragmatic dysfunction (VIDD). The mechanisms are not fullyexplained, but muscle atrophy, oxidative stress, structuralinjury, and muscle fiber remodeling contribute to variousextents in the development of VIDD.
SUGGESTED READINGS
ATS/ERS: Statement on Respiratory Muscle Testing. Am J Respir CritCare Med 2002; 166:518–624.
Laghi F, Tobin M: Disorders of the respiratory muscles. Am J Respir CritCare Med 2003; 168:10–48.
Mantilla CB, Sieck GS: Invited review: Mechanisms underlying motor unitplasticity in the respiratory system. J Appl Physiol 2003; 94:1230–1241.
Orozco-Levi M: Structure and function of the respiratory muscles inpatients with COPD: impairment or adaptation? Eur Respir J 2003;22:Suppl.46, 41s–51s.
Roussos C, Zakynthinos S: Fatigue of the respiratory muscles. IntensiveCare Med 1996; 22:134–55 Au5.
Vassilakopoulos T: Ventilator-induced diaphragmatic dysfunction: Theclinical relevance of animal models. Intensive Care Med 2007 (in press) Au3.
Vassilakopoulos T, Roussos C: Neuromuscular respiratory failure. InSlutsky A, Takala R, Torres R, eds. Clinical Critical Care Medicine.St. Louis: Mosby: 275–282.
Vassilakopoulos T, Roussos C, Zakynthinos S: The immune response toresistive breathing. Eur Respir J 2004; 24:1033–1043.
Vassilakopoulos T, Zakynthinos S, Roussos C: Respiratory muscles andweaning failure. Eur Respir 1996; J9:2383–2400.
Vassilakopoulos T, Zakynthinos S, Roussos C: Muscle function: Basic con-cepts. In Marini JJ, Slutsky A, eds. Physiologic Basis of VentilatorSupport. New York: Marcel Dekker; 1998: 103–152.
148 S E C T I ON I I Physiology10008
Albert, 978-0-323-04825-5