a comparative study on the production of pha by three … r.sav, et al.pdf · · 2017-07-18a...
TRANSCRIPT

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
940
Original Research Article
A Comparative study on the production of PHA by three different Pseudomonas sp.
Ajay R.Sav, Amit K. Mittal, Alpana A. Thorat, Sachin Dubey and U.C.Banerjee*
Department of Pharmaceutical Technology (Biotechnology); National Institute of Pharmaceutical Education and Research, Sector 67, S.A.S. Nagar- 160062, Punjab, India.
*Corresponding author
A B S T R A C T
Introduction
Polymers have become indispensible part of our life. Every year close to 150 million tones of plastic is being used and the demand is increasing further more (Crank et al., 2004). Petroleum derived polymers are non-degradable and there is a rising burden of their safe disposal. Biologically produced and easily degradable polymers are often considered as practically feasible
alternatives; these are mostly produced from renewable energy sources unlike to non-renewable petroleum based polymers (Flieger et al., 2003; Nath et al., 2008). Polyhydroxyalkanoates (PHA) are among degradable polymers obtained through microbial fermentation (Raj et al., 2014; El-Shanshoury et al., 2013). These biopolymers belong to polyester class and offers
ISSN: 2319-7706 Volume 3 Number 10 (2014) pp. 940-954 http://www.ijcmas.com
K e y w o r d s
Polyhydroxyalkanoates (PHA), biopolymer, Pseudomonas sp., optimization, bioreactor, biodegradable
Polyhydroxyalkanoates (PHA) are biodegradable polymers with widespread applications, produced intracellularly by microbes in nutrient limiting conditions. PHA s have great potential of replacing conventional non-degradable polymers for various applications. This study was designed with an aim of screening new PHA producing pseudomonas strains. Three strains of Pseudomonas sp., (Pseudomonas MTCC 2496, Pseudomonas putida MTCC 102 and Pseudomonas putida MTCC 2445) were used for this purpose. PHA's obtained from all the three strains were characterized using gas chromatography (GC), Fourier transform infrared spectroscopy (FTIR) and nuclear magnetic resonance (NMR H1 and C13) techniques. The produced PHA's when compared with the marketed PHA, showed the presence of a mixture of different chain lengths. From the characterization point of view, presence of intense ester group was found with the PHA produced by Pseudomonas sp. MTCC 2496. The results suggested that Pseudomonas sp. MTCC 2496 was the most prominent among the screened microbes; it has concentrated up to 39% of PHA per liter of broth. Production of PHA was further optimized in a 7 l bioreactor. With the final optimized conditions (4% glucose; medium pH 7; 10 µg/ml inoculum size; 12 h inoculum age, 30 °C incubation temperature, 250 rpm agitation, and 0.5 vvm aeration rates) the yield was increased upto 57 %.

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
941
biocompatibility and biodegradability and can be produced from renewable source (Reddy et al., 2003). It has comparable physical characteristics (including molecular mass, brittleness, melting point and glass transition temperature) to that of synthetic petrochemical derived polymer such as polypropylene (Crank et al., 2004). Furthermore PHA s found a series of applications ranging from bioplastic, medical implants, drug delivery carriers, printing, photographic materials, synthesis of certain antibiotics, biofuels production etc (Lee et al., 2011).
PHA s can be accumulated as discrete granules by various microorganisms as a means of energy storage (Shah, 2014). A lists of PHA s producing microorganism is shown in Table 1. It has been reported that by varying the composition of carbon sources, the chemical and physical properties of PHA can be altered (Nonato et al., 2001). Recently Passanha et al. in 2014 studied the effect of NaCl on the production of PHA by Cupriavidus necator (Passanha et al., 2014). Addition of 0.09 % NaCl in the medium increased the production of PHA upto 30 % (Passanha et al., 2014). Arumugam et al. in 2014 reported the polyhydroxyalkanoate co-production with biohydrogen by Enterobacter aerogenes and Rhodobacter sphaeroides from Calophyllum inophyllum oil cake as carbon source (Arumugam et al., 2014). A polyhydroxyalkanoate (PHA) producing photosynthetic mixed culture under dark/light periods without aeration was also reported, where 30 % PHA content was obtained (Albuquerque et al., 2011). Chen et al. in 2013 reported PHA biosynthesis using seed sludge at pH 9.0 with continuous pulsed feeding (Chen et al., 2013). Similarly Venkateswar Reddy and Venkata Mohan in 2012 reported substrates for PHA production under aerobic and anoxic condition using food waste and fermented
effluent (Venkateswar Reddy and Venkata Mohan, 2012). The results showed that the higher PHA production was obtained at anoxic condition rather than aerobic. Albuquerque et al. in 2011 reported the production of PHA from volatile fatty acids in fermented molasses (Albuquerque et al., 2010). The effect of substrate composition and feeding regime were also reported (Albuquerque et al., 2010).
The present study was conducted to screen the best strains of Pseudomonas sp. for the production of PHA. PHA s produced by various strains were isolated and characterized. The various physico-chemical conditions for the growth and production of PHA by the select strain of Pseudomonas sp. were reported. Finally a laboratory scale bioreactor was used for the detailed study.
Materials and Methods
Chemicals and reagents
Yeast extract, beef extract and peptone were purchased from HI-MEDIA® Chemicals, Mumbai, India. Standard PHA (Poly [(R)-3-hydroxybutyric acid]) was obtained from Sigma-Aldrich Chemicals Pvt. Ltd. (Bangalore, India), Dinitrosalicylic acid was procured from S.D. Fine Chemicals Ltd., Mumbai, India. All other chemicals used in the study were of analytical grade. All the microorganisms used in this study were procured from Microbial Type of Culture Collection, Institute of Microbial Technology, Chandigarh, India.
Cultivations of microorganisms
For the screening of microorganisms and inoculum preparation same medium was used. The medium consisted of (per liter of deionized water) 2 g yeast extract, 1 g beef

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
942
extract, 5 g peptone, 5 g NaCl and the 10 ml trace element solution. The composition of trace element was 10 g FeSO4, 2.25 g ZnSO4.7H2O, 2.25 g CuSO4.5H2O, 0.5 g MnSO4. 5 H2O, 2 g CaCl2.2H2O, 0.1 g (NH4)6 Mo7O24 in a liter of water. Media for the batch fermentation consisted of (per liter of deionized water) 40 g glucose, 30 g (NH4)2SO4, 1.2 g MgSO4, 1.7 g citric acid, 13.3 g KH2PO4 and 10 ml of trace elements as described above. The pH was adjusted to 7 before sterilization. Inoculum (10, % v/v) was generated by growing the various Pseudomonas strains in shake flask 30 °C.
PHA extraction
PHA was extracted following a previously reported method (Valentin and Dennis, 1997; Mohapatra et al., 2014). Briefly the cells were harvested by centrifuging of culture broth at 7000 g for 20 min followed by washing with 0.05N sodium phosphate buffer (pH 7.0). The cell pellets were mixed with chloroform and resultant suspension was shaken (200 rpm) at 37 ºC for 24 h. Bacterial debris was removed by filtering the suspension through Whatman filter paper (0.45 µm). The polymer was precipitated from the filtered solution using 10 volumes of ethanol. The precipitated polymer was re-dissolved in chloroform and re-precipitated with 10 volumes of ethanol. Finally filtered through the Whatman filter paper and allowed to dry.
Characterization of PHA
Determination of PHAs by gas chromatography PHA was quantitatively analyzed by gas chromatography (5765, Nucon Engineers, New Delhi, India). Propanolyzed sample (1.0 l) was injected into Reoplex packed column (2m x OD 3.2 mm x ID 2 mm) and the flame ionization detector (FID) was used for the analysis of PHA. Nitrogen gas was used as the carrier at
a flow rate of 30 ml/min. For analysis, oven temperature was maintained at 180 °C, injector at 230 °C and detector temperature was at 270 °C (Riis and Mai, 1988).
Nuclear magnetic resonance (NMR) spectroscopy
For NMR analysis polymer was extracted from 1 g dry cell mass of each bacterial strains of Pseudomonas sp. MTCC 2496, Pseudomonas putida MTCC 102, Pseudomonas putida MTCC 2445 and subjected to both 1H and 13C NMR spectroscopy. Analysis was carried out using 1H /13C dual probe and deuterated chloroform was used as solvent. For C13
NMR, samples were prepared by mixing 1g cellmass of individual cultures with 50 ml chloroform and incubated at 37 °C (200 rpm) for overnight, filtered through Whatman filter paper (0.45µm) and precipitated by 10 volumes of ethanol. Again polymer was dissolved in chloroform and re-precipitated using 10 volume of ethanol and then dissolved in deuterated chloroform.
FTIR analysis
The FTIR analysis of polyhydroxy-alkanoates (PHA) extracted from all the bacterial strains was performed to find out the presence of ester group. PHA produced from all the three bacterial strains were compared with marketed PHA. Samples were prepared by mixing 40 mg pellet of bacterial culture with 50 ml chloroform and incubated at 37 °C (200 rpm) for overnight, filtered through Whatman filter paper (0.45µm) and precipitated by 10 volumes of ethanol. Again the polymer was dissolved in chloroform and re-precipitated using 10 volume of ethanol and then used for FTIR analysis.

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
943
Optimization of studies in 7 l bioreactor
Production of PHA was maximized by optimizing each parameter one at a time. These studies were performed in 7 l bioreactor (BioFlow 410, New Brunswick Scientific, New Jersey, U.S.A.). Glucose concentration was varied from 1 to 8 % (w/v) in the reactor medium. Effect of initial pH was studied by starting the fermentation run at initial medium pH of 6, 7 and 8. Effect of temperature on the production of PHA and growth of Pseudomonas sp. was observed at various temperatures (25, 30 and 35 °C). Effect of agitation and aeration on the growth and production of PHA s by Pseudomonas sp. was studied at various agitation (150, 200 and 250 rpm) and aeration rates (0.5 and 1 vvm).
Determination of cell growth
Cell growth was determined by measuring the absorbance of the samples collected during the course of fermentation at various time interval. The absorbance was measured at 600 nm by dispersing the centrifuged call mass in sodium phosphate buffer using buffer as blank. Apart from measuring absorbance at 600 nm, cell dry weight was also measured. It was determined by centrifuging 100 ml culture broth at 7000 g for 20 min followed by through washing in phosphate buffer and drying the cellmass to a constant weight at 90oC.
Determination of residual glucose
Residual glucose was determined by the DNS (dinitrosalicylic acid) method (Miller, 1972). Briefly 1 ml supernatant (after centrifugation) was taken in a test tube. Distilled water (1 ml) and 3 ml DNS reagent was added and the mixture was boiled at 100 °C for 10 min. The mixture was then allowed to cool and finally 5 ml of distilled water was added before measuring the
absorbance at 540 nm using Perkin Elmer UV-Vis spectrophotometer.
Results and Discussion
Characterization of PHA
All the three Pseudomonas sp. were cultivated for 48 h and each dry cellmass was processed as described in PHA extraction section. The PHA extracted was analyzed and characterized by various techniques.
GC characterization
Figure S1 in the Electronic Supplementary Material shows the GC spectra obtained from the standard PHA and PHA s isolated from the Pseudomonas sp. MTCC 2496, Pseudomonas sp. MTCC 102 and Pseudomonas sp. MTCC 2445. PHA peak was observed at 0.6-0.7/min in all the cases.
FTIR characterization
To further confirm the presence of functional groups, FTIR spectra s were taken for the PHA s isolated from the three Pseudomonas sp. (Pseudomonas sp. MTCC 2496, Pseudomonas sp. MTCC 102 and Pseudomonas sp. MTCC 2445). The spectra were compared with the standard PHA (Figure S2 in the Electronic Supplementary Material). The FTIR analysis of polyhydroxyalkanoates extracted from all the bacterial strains showed the presence of ester group (Figure S2). The spectra of PHA s from all the bacterial strains were characterized by the presence of following groups: 3370 to 3350cm-1(-OH); intense 2925.0 to 2925.9 cm-1, 2853.8 to 2854.5 cm-
1 (-CH3) large bands to aliphatic C-H.
Characteristic signals from PHA were seen at 1723.7 to 1742.4 cm-1 corresponding to

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
944
the valence carbonyl vibration (C=O of ester group); 1450.6 to 1459.3 cm-1
( as CH3/
CH2); 1377.2 to 1378.0 cm-1 ( s CH3); 1162.9 to 1183.6 cm-1 (C-O). Between 1280 and 1050 cm-1 signals are due to valence antisymmetric and symmetric vibration of C-O-C. Medium signals could be seen at 1000 to 1500 cm-1 range due to bending of CH2 and CH3, -C-O- and C-C- groups. Signals were intense in standard PHA and PHA from Pseudomonas sp. MTCC 2496 (Figure S2), while absorption bands of medium intensity were observed with the PHA s from Pseudomonas putida MTCC 102 and Pseudomonas putida MTCC 2445 (Figure S2). All absorption peaks for the functional groups of PHA found in all the three Pseudomonas sp. were in agreement with the functional groups observed in the standard PHA spectra.
H1 and C13 NMR characterization
The H1 and C13 NMR were used to characterize the polymer, PHA. Figure S3 in the Electronic Supplementary Material shows the H1 NMR spectra of standard PHA (Figure S3a) and PHA s obtained from the different Pseudomonas sps. (Figures S3b, S3c and S3d represent Pseudomonas sp. MTCC 2496, Pseudomonas sp. MTCC 102 and Pseudomonas sp. MTCC 2445, respectively). Similarly Figure S4 in the Electronic Supplementary Material shows the C13 NMR spectra of standard PHA (Figure S4a) and PHA s obtained from the different Pseudomonas sps. (Figures S4b, S4c and S4d represent Pseudomonas sp. MTCC 2496, Pseudomonas sp. MTCC 102 and Pseudomonas sp. MTCC 2445 respectively). Identical signal patterns in both the NMR for all the PHA s confirmed the identity and quality of PHA produced from different Pseudomonas sps. Signal at 5.22-5.28 in 1H spectra and corresponding signal at 68.16 in 13C spectra has confirmed the presence of -CH- group in PHA
extracted from different Pseudomonas sp. Signal at 0.88 in 1H spectra and corresponding signal at 169 in 13C spectra confirmed the carbonyl group in PHA.
Shake flask studies
Optimization of inoculum age and size
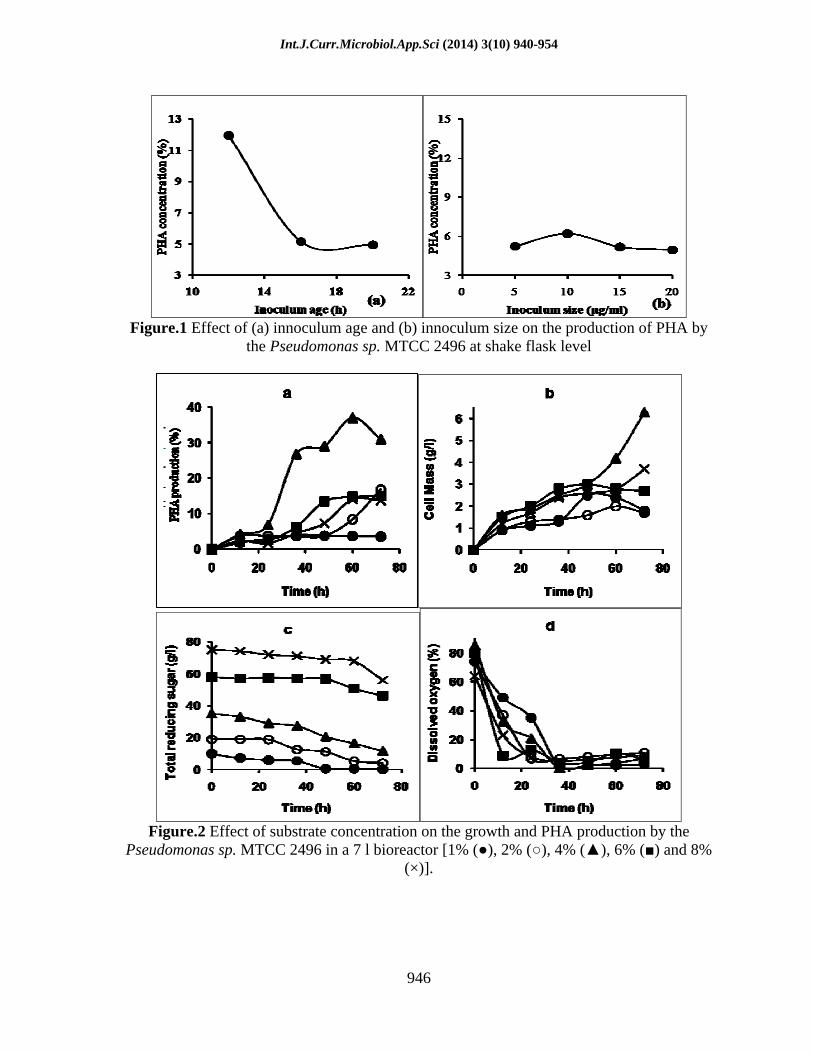
The effect of inoculum age and size on the accumulation of polyhydroxyalkanoates (PHAs) by Pseudomonas sp. MTTC 2496 was studied. Figure 1 summarizes the PHA producing potential of the Pseudomonas sp. as a function of inoculum age and size. Pseudomonas sp. showed the production of high quality PHA as evident by FTIR and NMR spectra s analysis. The optimum innoculum size showing maximum accumulation of PHA was 10 µg/ml and the optimum inoculum age was found to be 12 h.
Optimization of fermentation parameters
Effect of substrate concentration
Carbon source is important for the growth and production of PHA by the microorganism. Glucose was used as the primary carbon source for the synthesis of PHA. Its concentration was varied from 1 to 8% in the bioreactor. Cell mass, residual glucose and PHA concentrations were measured with time. It is evident from Figure 2 that when initial glucose concentration in the reactor is 3%, amount of PHA produced was also very less and there was no residual glucose in the reactor. It is known that higher glucose level in the medium creates stress condition which leads to the accumulation of PHA (Suriyamongkol et al., 2007). The amount of PHA produced with 4% glucose in reactor was 38% of cell dry weight and residual glucose concentration at the end of 72 h was 25% of the initial concentration (Figure 2). Data suggested that further increase of glucose level in the medium was neither necessary

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
945
for PHA accumulation nor for the growth, hence 4% glucose was found to be optimum for PHA production. The decrease of dissolved oxygen concentration followed more or less the same pattern during the course of growth and PHA synthesis by Pseudomonas sp. MTTC 2496. Maximum cellmass growth was observed with 40 g/l glucose in the bioreactor. It was found that higher glucose level had decreased both the cellmass growth as well as PHA synthesis. It might be due to the accumulation of acids (lowering of pH) at higher glucose concentration and subsequent deactivation of enzymes (Ryu et al., 2008; Sangkharak and Prasertsan, 2013).
Effect of pH and temperature
To study the effect of initial pH on the growth and PHA production by Pseudomonas sp., the organism was grown at different initial pH s in the range of 6 to 8 in 5 L bioreactor. The organism grew well in all the three pH s but maximum celmass (3.1 g/L) and higher PHA content (40% w/w) were obtained when the initial pH of the medium was adjusted to pH 7 (Table 2). Deviation on the either of side of the initial pH value (pH 7) led to fall in PHA production. The glucose uptake rate was more or less same in all the three cases and maximum glucose consumption took place up to 60 h of fermentation where residual glucose concentration was around 2-2.2 g/L in the case of bioreactor run with initial pH 6 and 8 (data not shown). The residual glucose concentration was much less (1.5 g/L) when the organism was cultivated in the medium of initial pH 7. The rate of production of PHA was less at initial medium pH 8 while less cell growth was observed at initial medium pH 6. Temperature conditions during PHA production by Pseudomonas sp. have also been studied. Results of these studies have been summarized in Table 2. Yield of PHA
remained same at all the three temperature conditions, thus varying temperature did not create stress required for enhancing overall PHA production. Studies indicated that pH and temperature did not have any significant effect on the specific growth rate of microorganism (µ).The maximum specific product formation rate (qp) and specific product yield [Y(P/X)] was observed at pH 7 and 30 °C. It was observed that pH played an important role for the PHA production while temperature did not make any difference. Effect of agitation
Agitation is important for mixing, mass transfer and heat transfer. Investigations were carried out at different agitation speeds of 150, 200 and 250 rpm for the production of PHA by Pseudomonas sp. MTCC 2496 (Figure 3). It was observed that with the increase of agitation, growth rate remained almost same. On the other hand maximum PHA production was obtained at an agitation rate of 250 rpm. Dissolved oxygen and pH followed a similar profile in all the three cases. However, at the end of fermentation, comparatively higher amount of residual sugar was observed in higher PHA production medium.
Effect of aeration
Aeration is another essential parameter for the metabolic activities of aerobic organisms. Aeration at the same time also promotes mixing in the bioreactor thereby increases the efficiency of the transport. The effect of increasing the aeration rate was studied by sparging the bioreactor with air at different aeration rates (0.5 and 1 vvm). At 0.5 vvm, PHA production was found to be more, the possible reason might be the availability of less oxygen created stress condition and increased the PHA production (Figure 4). On the other hand at the same time there was a lower growth and PHA production at higher aeration rates.
Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
946
Figure.1 Effect of (a) innoculum age and (b) innoculum size on the production of PHA by the Pseudomonas sp. MTCC 2496 at shake flask level
Figure.2 Effect of substrate concentration on the growth and PHA production by the Pseudomonas sp. MTCC 2496 in a 7 l bioreactor [1% ( ), 2% ( ), 4% ( ), 6% ( ) and 8%
(×)].

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
947
Table.1 List of the selected PHA producing microorganisms
Organism/ microenvironment
Fermentation method
Substrate PHA content (%, DCW)
References
Pseudomonas aeruginosa
Batch Activated sludge with nonanoic acid
48.6, YP/S 0.94 g/g [13]
Mixed microbial culture
2-stage CSTR Sugar cane molasses
61 [1]
Ralstonia pickettii Shake flask Sugar cane molasses
50 [6]
Pseudomonas fluorescens
SMF, Batch Sugarcane liquor, glutamate and salts
70 [12]
Bacillus cereus M5 SMF, Batch Beet molasses 73.8 [30]
Azotobacter vinelandii UWD
SMF, Fed Batch Beet molasses and valerate
59-71 [19]
Bhurkolderia sp. and Cupriavidus necator (formerly R. eutropha)
SMF, Fed Batch Sucrose from sugarcane
65-70 []18]
Rec. E. coli (C. necator genes)
SMF, Fed Batch Beet molasses + salts + trace metals
80 [14]
Aerobic and anoxic condition
Batch Food waste and acidogenic effluents
40 [29]
Anaerobic condition Continuous, Batch
fermented volatile fatty acids
65 [8]
Cupriavidus necator Fed Batch Peptone, meat extract and NaCl
80 [20]
Table.2 Effect of pH and temperature on PHA production
Condition Cell mass (g/l)
µ (x 10-2)
(h-1)
qp
(x 10-
2) (h-1)
Y(P/X)
PHA content (%,
DCW)
Time to achieve maximum
production (h) 6 2.0 1.46 5.64 3.86 32 60 7 3.1 1.94 5.82 3.0 40 60
pH
8 2.8 1.96 4.62 2.36 33 48 25
2.5 2.03 4.51 2.22 38 36 30
3.1 1.94 5.19 2.67 39 60 Temperature
(°C) 35
2.8 2.19 2.67 2.41 40 60

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
948
Figure.3 Effect of agitaion on the growth and PHA production by the Pseudomonas sp. MTCC 2496 in a 7 l fermentor [150 rpm ( ), 200 rpm ( ) and 250 rpm( )]
Figure.4 Effect of aeration on the growth and PHA production by the Pseudomonas sp. MTCC 2496 in a 7 l fermentor [0.5 and 1vvm]

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
949
Figure.S1. Gas Chromatography spectra of standard and extracted polymers from different
Pseudomonas strains (a) Standard PHA, (b) MTCC 2496, (c) MTCC 102 and (d) MTCC 2445.

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
950
Figure.S2 FTIR spectra of standard and extracted polymers from different Pseudomonas strains (a)
Standard PHA, (b) MTCC 2496, (c) MTCC 102 and (d) MTCC 2445
ab
c d

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
951
Figure.S3 1H NMR spectra of standard and extracted polymers from different Pseudomonas
strains (a) Standard PHA, (b) MTCC 2496, (c) MTCC 102 and (d) MTCC 2445.

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
952
Figure.S4 13C NMR spectra of standard and extracted polymers from different Pseudomonas
strains (a) Standard PHA, (b) MTCC 2496, (c) MTCC 102 and (d) MTCC 2445
a b
c d

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
953
Acknowledgements
All the authors gratefully acknowledge for financial support provided by Department of Biotechnology, New Delhi, India.
References
Albuquerque, M.G.E., Concas, S., Bengtsson, S., Reis, M.A.M. 2010. Mixed culture polyhydroxyalkanoates production from sugar molasses: The use of a 2-stage CSTR system for culture selection. Biores. Technol., 101: 7112 7122.
Albuquerque, M.G.E., Martino, V., Pollet, E., Avérous, L., Reis. M.A.M. 2011. Mixed culture polyhydroxyalkanoate (PHA) production from volatile fatty acid (VFA)-rich streams: effect of substrate composition and feeding regime on PHA productivity, composition and properties. J. Biotechnol., 151: 66 76.
Arumugam, A, Sandhya, M., Ponnusami, V. 2014. Biohydrogen and polyhydroxyalkanoate co-production by Enterobacter aerogenes and Rhodobacter sphaeroides from Calophyllum inophyllum oil cake. Biores. Technol., 164: 170 176.
Chen, H., Meng, H., Nie, Z., Zhang, M. 2013. Polyhydroxyalkanoate production from fermented volatile fatty acids: effect of pH and feeding regimes. Biores. Technol., 128: 533 538.
Crank, M., Patel, M., Marscheider-Weidemann, F., Schleich, J., Hüsing, B., Angerer, G. 2004. Techno-economic Feasibility of Large-scale Production of Bio-based Polymers in Europe (PRO-BIP). Final Report Prepared for the European Commission s Institute for Prospective Technological Studies (IPTS).
El-Shanshoury, A.E.R.R., Kenawy, E.R., Amara, A.A., Salama, A.F., Kishk, S.S.
2013. Optimization of Polyhydroxybutyrate (PHB) production by Azotobacter vinelandii using experimental design. Int. J. Curr. Microbiol. Appl. Sci., 2: 227 241.
Flieger, M., Kantorová, M., Prell, A., ezanka, T., Votruba, J. 2003.
Biodegradable plastics from renewable sources. Folia Microbiol., 48: 27 44.
Lee, S.H., Kim, J.H., Mishra, D., Ni, Y.Y., Rhee, Y.H. 2011. Production of medium-chain-length polyhydroxyalkanoates by activated sludge enriched under periodic feeding with nonanoic acid. Biores. Technol., 102: 6159 6166.
Miller, G.L. 1972. Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal. Chem., 31: 426428.
Mohapatra, S., Samantaray, D.P., Samantaray, S.M. 2014. Phylogenetic heterogeneity of the rhizospheric soil bacterial isolates producing PHAs revealed by comparative analysis of 16s-rRNA. Int. J. Curr. Microbiol. Appl. Sci., 3: 680 690.
Nath, A., Dixita, M., Bandiyaa, A., Chavda, S., Desai, A.J. 2008. Enhanced PHB production and scale up studies using cheese whey in fed batch culture of Methylobacterium sp. ZP24. Biores. Technol., 99: 5749 5755.
Nonato, R.V., Mantelatto, P.E., Rossell, C.E.V. 2001. Integrated production of biodegradable plastic, sugar and ethanol. Appl. Microbiol. Biotechnol., 57: 1 5.
Passanha, P., Kedia, G., Dinsdale, R.M., Guwy, A.J., Esteves, S.R. 2014. The use of NaCl addition for the improvement of polyhydroxyalkanoate production by Cupriavidus necator. Biores. Technol., 163: 287294.
Raj, A., Ibrahim, V., Devi, M., Sekar, K.V.,

Int.J.Curr.Microbiol.App.Sci (2014) 3(10) 940-954
954
Yogesh, B.J., Bharathi, S. 2014. Screening, optimization and characterization of poly hydroxy alkanoates (pha) produced from microbial isolates. Int. J. Curr. Microbiol. Appl. Sci., 3: 785 790.
Reddy, C.S.K., Ghai, R., Khan, R., Kalia, V. 2003. Polyhydroxyalkanoates: an overview. Biores. Technol., 87: 137146.
Riis, V., Mai, W. 1988. Gas chromatographic determination of poly-ß-hydroxybutyric acid in microbial biomass after hydrochloric acid propanolysis. J. Chromatogr., 445: 285 289.
Ryu, H.W., Cho, K.S., Goodrich, P.R., Park, C.H. 2008. Production of polyhydroxyalkanoates by Azotobacter vinelandii UWD using swine wastewater: Effect of supplementing glucose, yeast extract, and inorganic salts. Biotechnol. Bioproc. Eng., 13: 651 658.
Sangkharak, K., Prasertsan, P. 2013. The production of polyhydroxyalkanoate by Bacillus licheniformis using sequential mutagenesis and optimization. Biotechnol. Bioproc. Eng., 18: 272 279.
Shah, K.R. 2014. Optimization and production of Polyhydroxybutarate (PHB) by Bacillus subtilis G1S1from soil. Int. J. Curr. Microbiol. Appl. Sci., 3: 377 387.
Suriyamongkol, P., Weselake, R., Narine, S., Moloney, M., Shah, S. 2007. Biotechnological approaches for the production of polyhydroxyalkanoates in microorganisms and plants a review. Biotechnol. Adv., 25: 148 175.
Valentin, H.E., Dennis, D. 1997. Production of poly (3-hydroxybutyrate-co-4-hydroxybutyrate) in recombinant Escherichia coli grown on glucose. J. Biotechnol., 58: 33 38.
Venkateswar Reddy, M., Venkata Mohan, S. 2012. Effect of substrate load and nutrients concentration on the polyhydroxyalkanoates (PHA) production using mixed consortia through wastewater treatment. Biores. Technol., 114: 573 582.