a comparison of the clavam biosynthetic gene clusters in streptomyces antibioticus ...
TRANSCRIPT

A comparison of the clavam biosynthetic geneclusters in Streptomyces antibioticus Tü1718 andStreptomyces clavuligerus
Sarah Goomeshi Nobary and Susan E. Jensen
Abstract: The production of clavam metabolites has been studied previously in Streptomyces clavuligerus, a species thatproduces clavulanic acid as well as 4 other clavam compounds, but the late steps of the pathway leading to the specific endproducts are unclear. The present study compared the clavam biosynthetic gene cluster in Streptomyces antibioticus, chosenbecause it produces only 2 clavam metabolites and no clavulanic acid, with that of S. clavuligerus. A cosmid library ofS. antibioticus genomic DNA was screened with a clavaminate synthase-specific probe based on the corresponding genesfrom S. clavuligerus, and 1 of the hybridizing cosmids was sequenced in full. A clavam gene cluster was identified thatshows similarities to that of S. clavuligerus but also contains a number of novel genes. Knock-out mutation of the clavami-nate synthase gene abolished clavam production in S. antibioticus, confirming the identity of the gene cluster. Knock-outmutation of a novel gene encoding an apparent oxidoreductase also abolished clavam production. A potential clavam biosyn-thetic pathway consistent with the genes in the cluster and the metabolites produced by S. antibioticus, and correspondinglydifferent from that of S. clavuligerus, is proposed.
Key words: b-lactam, clavulanic acid, valclavam, alanylclavam, clavamycin.
Résumé : La production de métabolites des clavames a déjà été étudiée chez Streptomyces clavuligerus, une espèce qui pro-duit de l’acide clavulanique ainsi que 4 autres clavames, mais les dernières étapes de la voie qui génère les produits finauxspécifiques ne sont pas claires. La présente étude compare la grappe de gènes de biosynthèse des clavames chez Streptomy-ces antibioticus, choisi car il produit seulement 2 métabolites de clavames et pas d’acide clavulanique, avec celle de S. cla-vuligerus. Une banque de cosmides d’ADN génomique de S. antibioticus a été criblée avec une sonde spécifique à laclavaminate synthase conçue à partir des gènes correspondants chez S. clavuligerus, et un des cosmides qui s’hybridait àcette sonde a été complètement séquencé. Une grappe de gènes de la voie des clavames qui présentait des similarités aveccelle de S. clavurigerus a été identifiée, mais elle comportait aussi un certain nombre de gènes nouveaux. Une mutationinactivant le gène de la clavaminate synthase abolissait la production de clavames chez S. antibioticus, confirmant l’identitéde la grappe de gènes. La mutation nulle d’un gène nouveau codant une oxydoréductase présumée abolissait aussi la pro-duction de clavames. Une voie potentielle de biosynthèse des clavames, cohérente avec les gènes de la grappe et les métabo-lites produits par S. antibioticus et par conséquent différente de celle de S. clavuligerus, est proposée.
Mots‐clés : b-lactame, acide clavulanique, valclavame, alanylclavame, clavamycine.
[Traduit par la Rédaction]
Introduction
Clavulanic acid is a potent b-lactamase inhibitor used clini-cally in combination with conventional b-lactam antibiotics tocombat infections caused by b-lactamase-producing organisms,and is the best known member of the clavam family of antibi-otics. It differs structurally from other clavam metabolites inseveral regards but most notably in the stereochemistry of itsbicyclic oxazolidine ring system (Fig. 1). Clavulanic acidshows 5R stereochemistry, which is associated with its b-lacta-mase inhibitory activity, whereas all other clavam metabolitesstudied to date have 5S stereochemistry and lack b-lactamaseinhibitory activity (Baggaley et al. 1997). These other clavamcompounds are referred to collectively as 5S clavams.
The biosynthesis of clavulanic acid and the 5S clavams hasbeen studied in Streptomyces clavuligerus, the sole speciesused for industrial production of clavulanic acid. In additionto clavulanic acid, S. clavuligerus produces the 5S clavamcompounds 2-hydroxymethylclavam, 2-formyloxymethylcla-vam, clavam-2-carboxylic acid, and alanylclavam (Fig. 1).The biosynthetic pathway leading to clavulanic acid and the5S clavams is shared in its early stages (Fig. 2), up to theproduction of clavaminic acid (Egan et al. 1997), and the en-zymes involved in these early steps have been well character-ized (Zhang et al. 2000; Miller et al. 2001; Elkins et al. 2002;Caines et al. 2004). The genes responsible for clavam biosyn-thesis in S. clavuligerus are unusual in several respects, in-cluding the finding that the genes encoding these early
Received 19 November 2011. Accepted 15 December 2011. Published at www.nrcresearchpress.com/cjm on 21 March 2012.
S. Goomeshi Nobary and S.E. Jensen. Department of Biological Sciences, CW-405 Biological Sciences Building, University of Alberta,Edmonton, AB T6G 2E9, Canada.
Corresponding author: Susan E. Jensen (e-mail: [email protected]).
413
Can. J. Microbiol. 58: 413–425 (2012) doi:10.1139/W2012-012 Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.
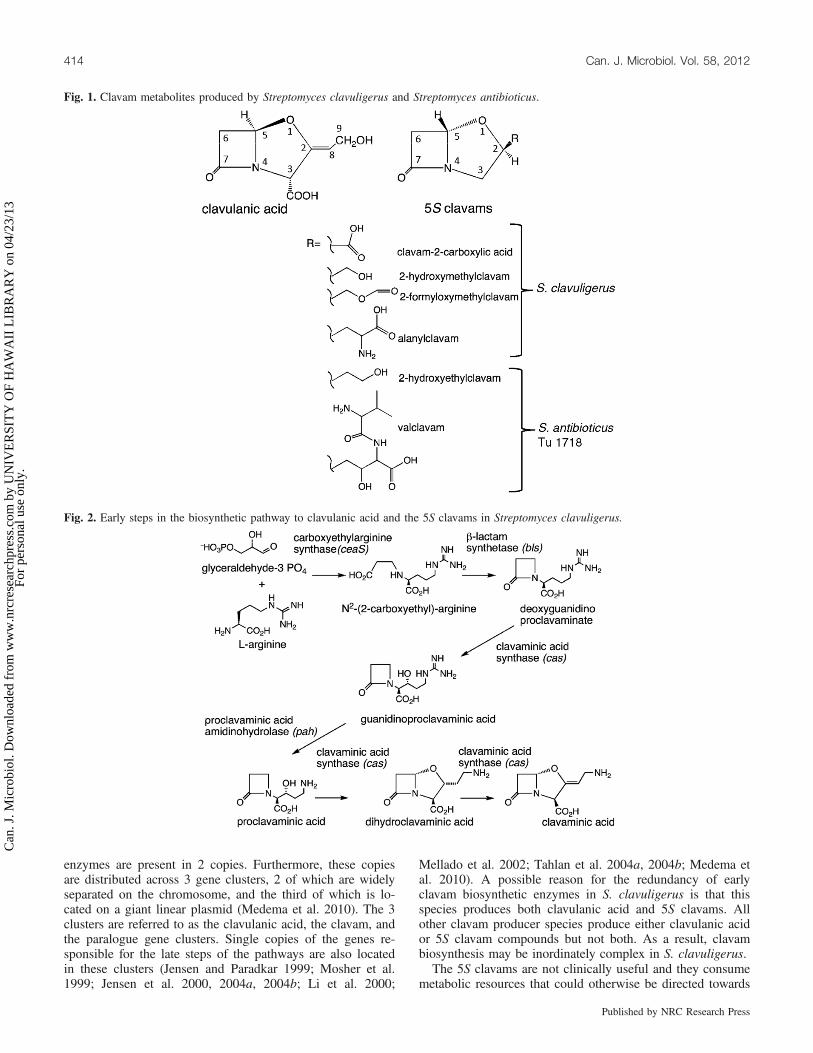
enzymes are present in 2 copies. Furthermore, these copiesare distributed across 3 gene clusters, 2 of which are widelyseparated on the chromosome, and the third of which is lo-cated on a giant linear plasmid (Medema et al. 2010). The 3clusters are referred to as the clavulanic acid, the clavam, andthe paralogue gene clusters. Single copies of the genes re-sponsible for the late steps of the pathways are also locatedin these clusters (Jensen and Paradkar 1999; Mosher et al.1999; Jensen et al. 2000, 2004a, 2004b; Li et al. 2000;
Mellado et al. 2002; Tahlan et al. 2004a, 2004b; Medema etal. 2010). A possible reason for the redundancy of earlyclavam biosynthetic enzymes in S. clavuligerus is that thisspecies produces both clavulanic acid and 5S clavams. Allother clavam producer species produce either clavulanic acidor 5S clavam compounds but not both. As a result, clavambiosynthesis may be inordinately complex in S. clavuligerus.The 5S clavams are not clinically useful and they consume
metabolic resources that could otherwise be directed towards
Fig. 1. Clavam metabolites produced by Streptomyces clavuligerus and Streptomyces antibioticus.
Fig. 2. Early steps in the biosynthetic pathway to clavulanic acid and the 5S clavams in Streptomyces clavuligerus.
414 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.

clavulanic acid production. They also complicate downstreamprocessing during the industrial production of clavulanicacid. Genetic manipulation to eliminate the 5S clavams selec-tively can improve clavulanic acid productivity in S. clavuli-gerus (Paradkar et al. 2001). On the other hand, the 5Sclavams differ from clavulanic acid in that they carry a vari-ety of side chains at C-2, which affects their biological prop-erties. A full understanding of 5S clavam biosynthesis couldprovide biological routes for production of clavulanic acidvariants with alternative side chains.Streptomyces antibioticus Tü1718 produces valclavam and
2-hydroxyethylclavam (Fig. 1) (Röhl et al. 1987) but no clav-ulanic acid. As such, clavam biosynthesis in S. antibioticusmay be simpler than in S. clavuligerus, and a comparison ofthe genetic elements involved in 5S clavam production be-tween the 2 species could shed light on the overall process.Clavaminate synthase (Cas) is known to catalyze 3 of theearly biosynthetic steps (Fig. 2) leading to clavaminic acid inS. clavuligerus. The presence of Cas activity has been dem-onstrated for the enzyme isolated from S. antibioticus (Jancet al. 1995), and similarly, proclavaminic acid amidinohydro-lase (Pah) activity, the subsequent enzyme in the pathway,has been detected in S. antibioticus by Baldwin et al.(1994), but otherwise, no studies on clavam biosynthesishave been conducted in that or any species other than S. clav-uligerus. We located and sequenced the clavam biosyntheticgenes from S. antibioticus and compared them to the corre-sponding genes from S. clavuligerus to gain a fuller under-standing of the production of clavam metabolites in thesespecies.
Materials and methods
Bacterial strains and culture conditionsStreptomyces antibioticus Tü1718 was kindly provided by
C.A. Townsend (Department of Chemistry, Johns HopkinsUniversity). Wild-type S. clavuligerus NRRL 3585 was ob-tained from the Northern Regional Research Laboratory, Pe-oria, Illinois, USA. Spores of S. antibioticus were collectedfrom ISP2 (Becton Dickinson, Sparks, Maryland) agar platesafter incubation at 28 °C for 7 days.To assess clavam metabolite production, seed medium
(0.4 g L-methionine, 4.0 g glucose, 4.0 g yeast extract, 10 gmalt extract, 1000 mL H2O, pH 7.3) (Janc et al. 1995) wasinoculated with 108 spores of S. antibioticus and incubatedfor 20 h at 28 °C and 250 r/min. Fermentation medium(30 g mannitol, 30 g soy flour, 1000 mL H2O, pH 7.2) (Jancet al. 1995) was inoculated to 2% (v/v) with seed culture andincubated at 28 °C and 250 r/min for up to 72 h. Trypticasesoy broth (Becton Dickinson) supplemented with 1% (m/v)soluble starch was used to grow Streptomyces spp. to extractgenomic DNA (gDNA).Lennox broth (LB; 10 g tryptone, 5 g yeast extract, 5 g
NaCl, 1000 mL H2O, solidified with Bacto-agar at 15 g/Lwhen solid medium was required) was used for routine culti-vation of Escherichia coli strains. Cultures were incubated at37 °C for 20 h. Antibiotics used as selective agents in E. coliincluded ampicillin (100 µg/mL), chloramphenicol (25 µg/mL), apramycin (50 µg/mL), and kanamycin (50 µg/mL). To
select S. antibioticus mutants containing antibiotic resistancegenes, ISP2 agar plates were supplemented with apramycin(25 µg/mL).
DNA techniquesThe alkaline lysis method was used to isolate plasmid and
cosmid DNA (Birnboim and Doly 1979). DNA fragmentsgenerated by restriction endonuclease digestion were sepa-rated by electrophoresis using 0.8% agarose gels and purifiedwith QIAquick Gel Extraction kits (Qiagen Inc., Valencia,California).Double-stranded DNA probes were labeled using a nonra-
dioactive protocol, as instructed by the manufacturer (DIGHigh Prime DNA Labeling and Detection Starter kit I,Roche, Laval, Quebec) for use in Southern analyses(Sambrook et al. 1989).DNA sequence analysis in preliminary studies was carried
out by cycle sequencing using BigDye Terminator version3.1 Cycle Sequencing kits (Applied Biosystems, Carlsbad,California). Subsequently, the complete sequence of cosmid6J11 was determined by pyrosequencing, carried out by Seq-Wright DNA Technology Services (Houston, Texas).Plasmids were introduced into S. antibioticus by interspe-
cies conjugation from E. coli ET12567/pUZ8002 as de-scribed by Kieser et al. (2000).
Polymerase chain reactionPolymerase chain reaction (PCR) mixtures contained
50 pmol of each oligonucleotide primer, 50 mmol/L Tris–HCl (pH 9.0), 1.5 mmol/L MgCl2, 0.4 mmol/L b-mercapto-ethanol, 0.1 mg/mL purified bovine serum albumin,10 mmol/L (NH4)2SO4, 0.2 mmol/L (each) dNTP, and 5%(v/v) DMSO in a total volume of 50 µL, with 5–10 ng ofplasmid or cosmid DNA, or 500 ng of gDNA as template,and 0.5 units Taq polymerase (Fermentas-Thermo Fisher,Burlington, Ontario). The cycling conditions were as follows:2 min of initial denaturation at 96 °C, and 30 cycles of dena-turation at 96 °C for 30–60 s and annealing/elongation at 55–60 °C for 30 s, followed by a final elongation at 72 °C for5 min unless otherwise noted. Primers used in the study arelisted in Table S11.To amplify regions containing secondary structures, 7-
deaza-dGTP (New England Biolabs) was used in place ofdGTP, at final concentrations of 0.2, 0.4, 0.6, and 0.8 mmol/L. To amplify the apramycin resistance (Aprr) cassette forPCR-targeted mutagenesis, PCR conditions were as follows:2 min of initial denaturation at 94 °C, 10 cycles of denatura-tion at 94 °C for 45 s, annealing at 50 °C for 45 s, and elon-gation at 72 °C for 90 s; 15 cycles of denaturation at 94 °Cfor 45 s, annealing at 55 °C for 45 s, and elongation at 72 °Cfor 90 s; and a final elongation for 5 min at 72 °C.
Streptomyces antibioticus genomic DNA libraryconstructionGenomic DNA was prepared by a neutral lysis procedure
taking care to prevent shearing (Kieser et al. 2000). gDNA(100 µg) was then digested with 0.08 units of Sau3A for5 min at 37 °C. DNA fragments were separated by centrifu-gation through a 10%–35% sucrose density gradient for 20 h
1Supplementary data are available with the article through the journal Web site at http://nrcresearchpress.com/doi/suppl/10.1139/w2012-012.
Goomeshi Nobary and Jensen 415
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.

at 26 000 r/min (average centrifugal force, 85 175g) (Beck-man SW40Ti rotor), and the separated DNA fragments wererecovered by repeatedly removing 1 mL fractions from thetop of the gradient. Fractions containing DNA fragments ap-proximating the size of l DNA (48 kb) were pooled andtreated with shrimp alkaline phosphatase. Cosmid pOJ436(Bierman et al. 1992) was made linear by digestion withHpaI, treated with shrimp alkaline phosphatase, and then di-gested with BamHI to yield 2 cosmid vector arms, whichwere combined with the size-fractionated gDNA fragmentsand with packaging extract (Gigapack gold III kit, Stratagene,La Jolla, California) according to the manufacturer’s instruc-tions. Appropriate dilutions of the packaged mixture wereadded to E. coli XL-Blue MR cells and spread on 22 cm ×22 cm plates containing 200 mL of LB agar supplementedwith apramycin. After incubation for 16 h at 37 °C,apramycin-resistant colonies were picked using a QPix robot(Genetix, Boston, Massachusetts) and transferred to 384-wellmicrotiter dishes for storage at –80 °C.
Identification of cosmid clones carrying the clavamcluster from S. antibioticusClones from the cosmid library were patched in duplicate
onto PerForma nylon membranes (Genetix) placed on22 cm × 22 cm plates containing LB agar + apramycin.After incubation for 16 h at 37 °C, the colonies were lysedand the DNA was fixed on the membranes by baking in avacuum oven. The membranes were then subjected to South-ern analysis using a clavaminate synthase-specific probe gen-erated by PCR using primers SGN01 and SGN02 withgDNA from S. antibioticus as template.
Generation of cas3 and oxr mutantsMutants were generated using a modification of the REDI-
RECT PCR-targeting protocol described by Gust et al.(2003). pOJ436 cosmid clones bearing the genes to be de-leted could not be used for mutagenesis because of the inte-grative nature of the cosmid, and so fragments of the cosmidDNA insert carrying the genes of interest were subclonedinto pUC119 or pUC120. Aprr cassettes for gene replacementwere amplified by PCR using primer pairs as shown in TableS11. Each of these primers is chimeric, comprising 39 nt ofsequence at the 5′ end to direct the amplified Aprr cassetteto the gene target, fused to 19–20 nt of sequence at the 3′end, which primes the amplification of the cassette. The am-plified Aprr cassettes were then introduced by electroporationinto E. coli BW25113/pIJ790 cells already carrying plasmidscontaining the genes to be deleted. Following electroporation,plasmids conferring Aprr to the E. coli host were extractedand analyzed to confirm replacement of the genes of interestby the Aprr cassette. The mutant plasmids were then intro-duced into E. coli ET12567/pUZ8002 and from there, weretransferred to S. antibioticus through intergeneric conjugationwhere they combined with the chromosome by homologousrecombination. Exconjugants showing Aprr were transferredto ISP2 plates and allowed to sporulate in the absence ofantibiotic selection to encourage the loss of integrated plas-mid. Colonies arising from single spores were checked for re-tention of Aprr.Genomic DNA of apramycin-resistant colonies was then
extracted and subjected to PCR analysis to confirm the re-
placement of the target gene by the Aprr cassette. For cas3mutants, primers Aprr-UP with SGN06 (for the region up-stream of the cassette), and SGN06 with SGN14 (for the en-tire cas3 region) were used. For oxr mutants, primers Aprr-UP with SGN16 (for the region upstream of the cassette),and SGN14 and SGN16 (for the entire oxr region) were used.
Analysis of clavam production via high-performanceliquid chromatography and mass spectrometryThe production of clavams by wild-type and mutant strains
of S. antibioticus was assessed by analyzing samples of fer-mentation broth by high-performance liquid chromatography(HPLC). Fermentation broth was reacted with imidazole toderivatize clavam compounds specifically as described previ-ously (Foulstone and Reading 1982). The derivatized sampleswere then subjected to reversed-phase chromatography on aC18 Bondclone column (Phenomenex, Torrence, California)with a mobile phase consisting of 0.1 mol/L Na2HPO4 ad-justed to pH 3.68 with glacial acetic acid – methanol (94:6),as described previously (Paradkar and Jensen 1995). The pro-duction of valclavam by wild-type S. antibioticus was veri-fied by HPLC with on-line mass spectrometry (MS)detection by analyzing imidazole-derivatized fermentationbroths on a Xterra reversed phase column (Waters Scientific,Milford, Massachusetts) using a volatile mobile phase con-sisting of 0.1% acetic acid adjusted to pH 3.68 with NH4OH –methanol (96:4). Electrospray mass spectra were acquired ona ZMD-2 single quadropole instrument (Waters Scientific), asdescribed previously (Jensen et al. 2004a).
Analysis of clavam production by bioassayMethionine antimetabolite activity of 5S clavam metabo-
lites in fermentation broth was detected by agar diffusion bio-assay using Bacillus sp. strain ATCC 27860 as indicatororganism, as described previously (Pruess and Kellett 1983).
Results
Locating the clavam gene cluster in S. antibioticusTo locate the clavam gene cluster in S. antibioticus, a
probe specific for the clavaminate synthase-encoding genewas generated by PCR using primers based on conserved re-gions from the cas1 and cas2 genes of S. clavuligerus, withgDNA from S. antibioticus as template. A PCR product ofthe expected size (551 bp) was obtained (Fig. 3A) and aftergel purification was subjected to DNA sequence analysis.The sequence information obtained showed high similarity tocas1 and cas2 from S. clavuligerus (Fig. 3B), confirming thata fragment of the cas gene from S. antibioticus had been am-plified. Since the enzyme previously studied in S. antibioti-cus was designated clavaminate synthase 3 (Janc et al.1995), we named the gene encoding this enzyme cas3. ThePCR product was then labeled using a nonradioactive proto-col for use as a probe.A library of S. antibioticus gDNA fragments was prepared
in the E. coli – Streptomyces shuttle cosmid vector, pOJ436,and transfected into E. coli with selection for Aprr.Apramycin-resistant clones (2304 clones) were patched induplicate onto a nylon membrane and then hybridized withthe labeled cas3 probe. Eight cosmid clones hybridized withthe cas3 probe (Fig. S11).
416 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.
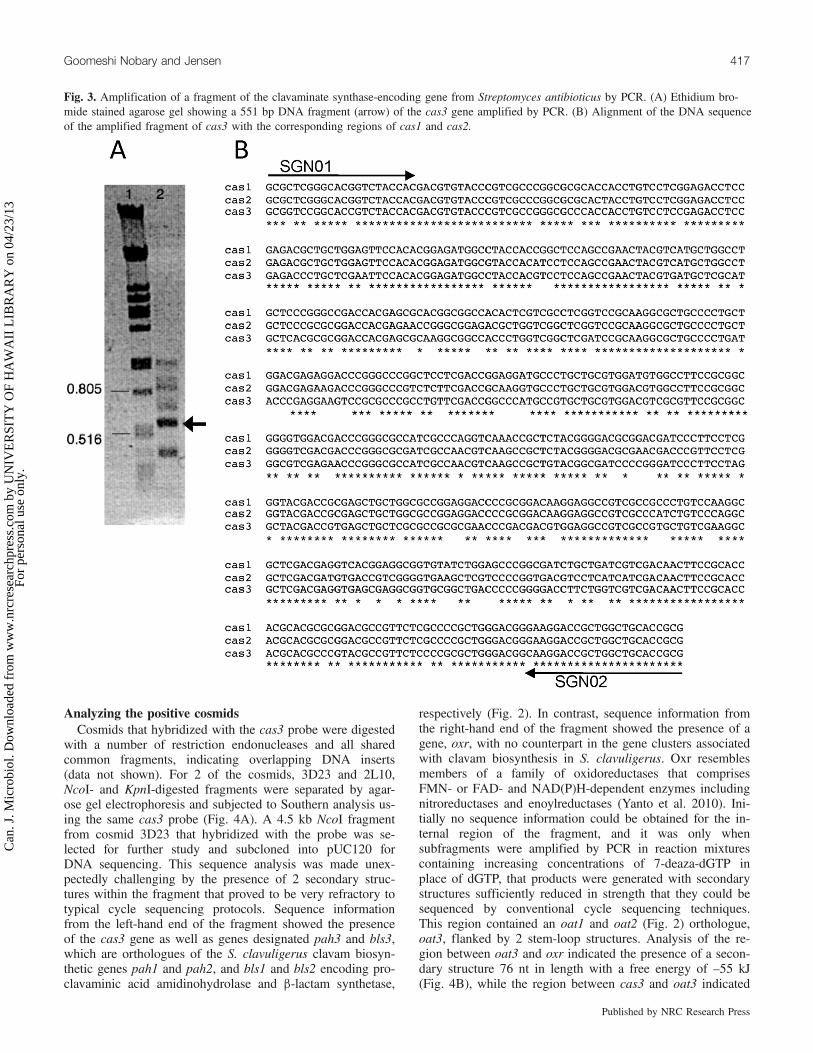
Analyzing the positive cosmidsCosmids that hybridized with the cas3 probe were digested
with a number of restriction endonucleases and all sharedcommon fragments, indicating overlapping DNA inserts(data not shown). For 2 of the cosmids, 3D23 and 2L10,NcoI- and KpnI-digested fragments were separated by agar-ose gel electrophoresis and subjected to Southern analysis us-ing the same cas3 probe (Fig. 4A). A 4.5 kb NcoI fragmentfrom cosmid 3D23 that hybridized with the probe was se-lected for further study and subcloned into pUC120 forDNA sequencing. This sequence analysis was made unex-pectedly challenging by the presence of 2 secondary struc-tures within the fragment that proved to be very refractory totypical cycle sequencing protocols. Sequence informationfrom the left-hand end of the fragment showed the presenceof the cas3 gene as well as genes designated pah3 and bls3,which are orthologues of the S. clavuligerus clavam biosyn-thetic genes pah1 and pah2, and bls1 and bls2 encoding pro-clavaminic acid amidinohydrolase and b-lactam synthetase,
respectively (Fig. 2). In contrast, sequence information fromthe right-hand end of the fragment showed the presence of agene, oxr, with no counterpart in the gene clusters associatedwith clavam biosynthesis in S. clavuligerus. Oxr resemblesmembers of a family of oxidoreductases that comprisesFMN- or FAD- and NAD(P)H-dependent enzymes includingnitroreductases and enoylreductases (Yanto et al. 2010). Ini-tially no sequence information could be obtained for the in-ternal region of the fragment, and it was only whensubfragments were amplified by PCR in reaction mixturescontaining increasing concentrations of 7-deaza-dGTP inplace of dGTP, that products were generated with secondarystructures sufficiently reduced in strength that they could besequenced by conventional cycle sequencing techniques.This region contained an oat1 and oat2 (Fig. 2) orthologue,oat3, flanked by 2 stem-loop structures. Analysis of the re-gion between oat3 and oxr indicated the presence of a secon-dary structure 76 nt in length with a free energy of –55 kJ(Fig. 4B), while the region between cas3 and oat3 indicated
Fig. 3. Amplification of a fragment of the clavaminate synthase-encoding gene from Streptomyces antibioticus by PCR. (A) Ethidium bro-mide stained agarose gel showing a 551 bp DNA fragment (arrow) of the cas3 gene amplified by PCR. (B) Alignment of the DNA sequenceof the amplified fragment of cas3 with the corresponding regions of cas1 and cas2.
Goomeshi Nobary and Jensen 417
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.
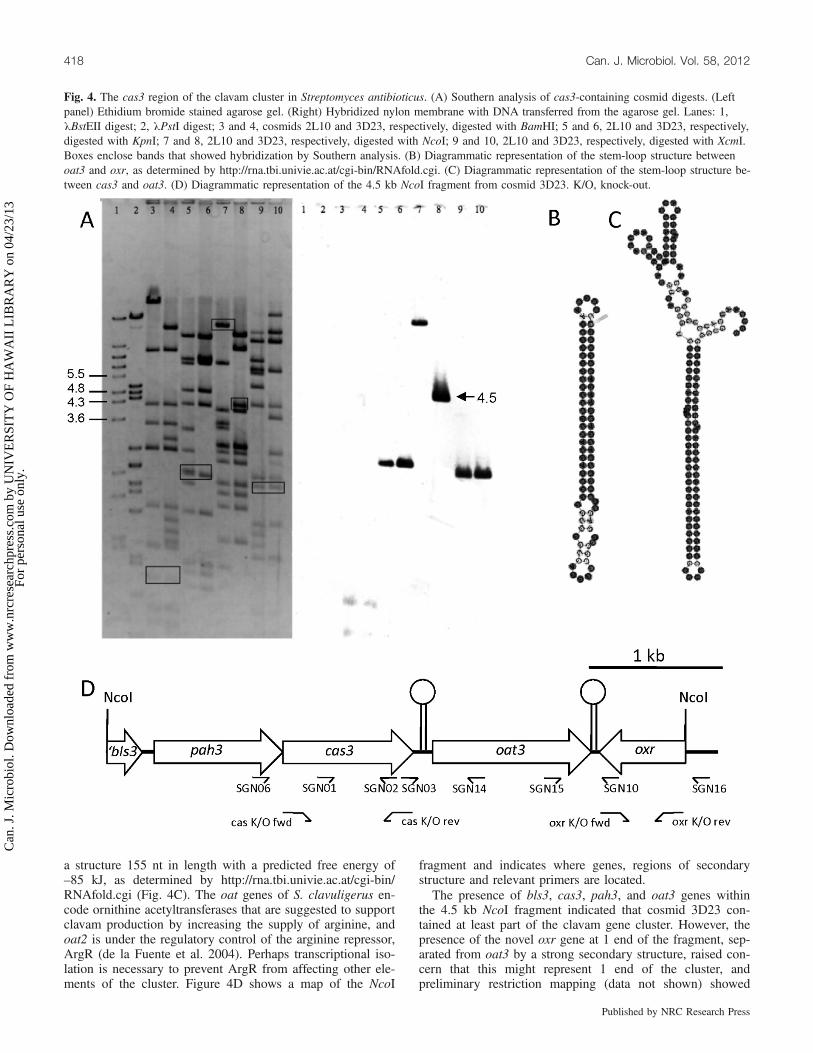
a structure 155 nt in length with a predicted free energy of–85 kJ, as determined by http://rna.tbi.univie.ac.at/cgi-bin/RNAfold.cgi (Fig. 4C). The oat genes of S. clavuligerus en-code ornithine acetyltransferases that are suggested to supportclavam production by increasing the supply of arginine, andoat2 is under the regulatory control of the arginine repressor,ArgR (de la Fuente et al. 2004). Perhaps transcriptional iso-lation is necessary to prevent ArgR from affecting other ele-ments of the cluster. Figure 4D shows a map of the NcoI
fragment and indicates where genes, regions of secondarystructure and relevant primers are located.The presence of bls3, cas3, pah3, and oat3 genes within
the 4.5 kb NcoI fragment indicated that cosmid 3D23 con-tained at least part of the clavam gene cluster. However, thepresence of the novel oxr gene at 1 end of the fragment, sep-arated from oat3 by a strong secondary structure, raised con-cern that this might represent 1 end of the cluster, andpreliminary restriction mapping (data not shown) showed
Fig. 4. The cas3 region of the clavam cluster in Streptomyces antibioticus. (A) Southern analysis of cas3-containing cosmid digests. (Leftpanel) Ethidium bromide stained agarose gel. (Right) Hybridized nylon membrane with DNA transferred from the agarose gel. Lanes: 1,lBstEII digest; 2, lPstI digest; 3 and 4, cosmids 2L10 and 3D23, respectively, digested with BamHI; 5 and 6, 2L10 and 3D23, respectively,digested with KpnI; 7 and 8, 2L10 and 3D23, respectively, digested with NcoI; 9 and 10, 2L10 and 3D23, respectively, digested with XcmI.Boxes enclose bands that showed hybridization by Southern analysis. (B) Diagrammatic representation of the stem-loop structure betweenoat3 and oxr, as determined by http://rna.tbi.univie.ac.at/cgi-bin/RNAfold.cgi. (C) Diagrammatic representation of the stem-loop structure be-tween cas3 and oat3. (D) Diagrammatic representation of the 4.5 kb NcoI fragment from cosmid 3D23. K/O, knock-out.
418 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.

that the insert in cosmid 3D23 did not extend far beyondbls3. Therefore an overlapping cosmid, 6J11, was sent forcomplete DNA sequence analysis (SeqWright, Houston,Texas). The sequence of the cosmid insert (36 847 bp) hasbeen deposited in GenBank (accession No. JN705801), andthe DNA sequence of the pOJ436 vector component (8598bp) has also been deposited (accession No. JN705802). Alist of the ORFs for the complete clavam cluster of S. antibio-ticus is shown (Table 1), and a list of all of the ORFs in thecosmid 6J11 insert is given in Table S21.
The clavam gene cluster of S. antibioticusGenes encoding all of the early enzymes of clavam biosyn-
thesis, ceaS3, bls3, pah3, cas3, and oat3, were located in theclavam gene cluster of S. antibioticus, and they are orientedand arranged in the same order as the corresponding genesfrom the clavulanic acid cluster of S. clavuligerus (Fig. 5).They show high sequence identity to their correspondinggenes from S. clavuligerus (74%–88% at the amino acidlevel), and in several cases even higher identity to genesfrom Streptomyces flavogriseus, a species not previouslyknown but now presumed to produce clavam metabolites.Flanking this region are several other genes also recognizableas clavam biosynthetic genes based on their similarity togenes from S. clavuligerus that have been demonstrated bymutational analysis to be essential for 5S clavam biosynthesisbut not for clavulanic acid biosynthesis (Fig. 5). One suchgene, cvm1, encodes a putative aldo–keto reductase, and acounterpart designated akr is located in the flank upstreamof ceaS3 in the S. antibioticus clavam cluster. Aldo–keto re-ductase shows 73% similarity to Cvm1 at the amino acidlevel. This same flank of the cluster contains the gene hmt,encoding a hydroxymethyltransferase that shows 78% similar-ity to OrfA, the hydroxymethyltransferase essential for ala-nylclavam biosynthesis in S. clavuligerus. A potentialregulatory gene, psr, showing similarity to the pathway-specific transcriptional regulator encoded by the cvm7p gene
of S. clavuligerus is located at the extreme end of the cluster,and just adjacent to it, atr encodes a putative aminotransfer-ase similar to the aminotransferase encoded by the cvm6pgene of S. clavuligerus. Both cvm6p and cvm7p are essentialfor 5S clavam biosynthesis in S. clavuligerus.Surprisingly, the other flank of the clavam gene cluster of
S. antibioticus contains a second copy of a pah gene, desig-nated pah4, located 6.7 kb downstream and oriented in theopposite direction from the pah3 gene. Pah4 shows 62%identity with and 75% similarity to Pah3 by BLAST analysis,and so we presume that it is a second proclavaminic acidamidinohydrolase, although it also shows lower levels of sim-ilarity to a wide range of proteins annotated as amidino-hydrolases and arginases. Assuming that pah4 is part of theclavam cluster, this flank therefore also includes the oxrgene and another unusual gene, lig. Lig belongs to the ATP-grasp fold superfamily, a group that includes D-alanine-D-alanine ligase, carbamoyl phosphate synthetase, and enzymesof secondary metabolism (Galperin and Koonin 1997).Orf17, encoded by orf17 in the clavulanic acid gene cluster,is also a member of the ATP-grasp fold superfamily, andthere is 42% overall similarity between Lig and Orf17 at theamino acid level but no obvious similarity at the nucleotidelevel. Orf17 is a N-glycyl-clavaminic acid synthetase that cat-alyzes the conversion of clavaminic acid to N-glycyl-clavaminic acid, and it is specifically required for the latestages of clavulanic acid but not 5S clavam biosynthesis inS. clavuligerus (Arulanantham et al. 2006). Therefore, despitesome similarity to orf17, lig is a gene like oxr, which has nocounterpart in 5S clavam biosynthesis in S. clavuligerus.Conversely, no orthologue of cvm5, the flavin-dependentoxygenase-encoding gene that is essential for production ofall 5S clavams in S. clavuligerus, was found in the S. antibio-ticus clavam cluster.In addition to genes encoding putative pathway enzymes,
this flank also includes 2 ancillary genes that may play roles.Immediately adjacent to oxr lies trn, encoding an apparent
Table 1. Proteins encoded by the clavam gene cluster of Streptomyces antibioticus.
ORF (aa) Most similar protein (BLAST analysis) Similarity (%) Predicted functionStreptomyces clavu-ligerus counterpart
Psr (843) SanG from Streptomyces ansochromogenes 44 Pathway specific transcriptionalregulator
Cvm7p
Atr (467) 4-Aminobutyrate aminotransferase fromSegniliparus rotundus
78 Aminotransferase Cvm6p
Akr (340) Cvm1 from S. clavuligerus 73 Aldo–keto reductase Cvm1Hmt (416) OrfA from S. clavuligerus 78 Hydroxymethyltransferase OrfACeaS3 (571) CeaS-like protein from Streptomyces
flavogriseus92 Carboxyethylarginine synthase CeaS1, CeaS2
Bls3 (507) Bls2 from S. clavuligerus 74 b-Lactam synthetase Bls1, Bls2Pah3 (320) Pah-like protein from S. flavogriseus 90 Proclavaminate amidinohydrolase Pah1, Pah2Cas3 (324) Cas1 from S. clavuligerus 88 Clavaminate synthase Cas1, Cas2Oat3 (394) Oat2 from S. clavuligerus 87 Ornithine acetyltransferase Oat1, Oat2Oxr (214) Oxidoreductase from Rhodococcus erythropolis 80 Enoylreductase NoneTrn (405) Major facilitator superfamily MFS_1
Streptomyces violaceusniger58 Clavam transporter None
Lig (478) LigA protein from Streptomyces hygroscopicus 62 Ligase NonePah4 (309) Pah-like protein from S. flavogriseus 72 Proclavaminate amidinohydrolase Pah1, Pah2Ctr (456) Hypothetical protein from Streptomyces sp. 69 Transcriptional regulator None
Goomeshi Nobary and Jensen 419
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.

transport-related protein of the major facilitator superfamilyMFS_1 (Pao et al. 1998), which may provide transport or re-sistance functions related to clavam production. The genecluster is flanked on its downstream edge by ctr, encodingan apparent regulatory protein from the Xre family of tran-scriptional regulators. Genes lying outside of the 2 transcrip-tional regulators, psr and ctr, show no obvious relationship toclavam biosynthesis (Table S21).
Mutation of cas3To establish unequivocally that this region of the S. anti-
bioticus chromosome represents the clavam gene cluster, thecas3 gene was subjected to PCR-targeted mutagenesis. The4.5 kb NcoI fragment cloned in pUC120 was used as theplatform for targeted mutagenesis and was introduced intoE. coli BW25113, a host strain carrying the lRED recombi-nation functions. Subsequent introduction by electroporationof a PCR-amplified apramycin resistance (Aprr) cassette spe-cifically designed to target cas3, led to replacement of thecas3 gene by the Aprr cassette. This mutant version of theplasmid was then transferred into S. antibioticus by inter-generic conjugation where it replaced the resident cas3 geneby homologous recombination. Four putative cas3 mutants(retained Aprr in the absence of selection) were recovered,and the authenticity of the mutations was confirmed by PCR(Fig. S2A1). All 4 were true double crossover mutants, andso a representative was selected for further analysis.
Mutation of oxrThe involvement of oxr in clavam production in S. antibioti-
cus was also investigated by mutagenesis, since it is centrallylocated in the cluster but has no counterpart in S. clavuligerus.A 4.2 kb NotI–EcoRI fragment encompassing the oxr genewas subcloned into plasmid pUC119, and oxr was replacedby an Aprr cassette by PCR-targeted mutagenesis. The muta-tion was then transferred into the chromosome of S. antibioti-cus as described above for cas3, and putative mutants werevalidated by PCR (Fig. S2B1). Of 5 potential mutants tested,2 were true double crossover mutants (lanes 5 and 7), while 3
were single crossover mutants. A representative oxr doublecrossover mutant was selected for further analysis.
Metabolite production by wild-type and mutant strains ofS. antibioticusWild-type and mutant strains of S. antibioticus were culti-
vated in a mannitol–soy based fermentation medium for up to72 h, and 25 µL samples of culture supernatants were takenat intervals and analyzed by agar disk diffusion bioassay formethionine antimetabolite activity. Bioassays showed that thewild-type culture supernatants contained substantial levels ofmethionine antimetabolite activity. Activity peaked at 48 h inthe wild-type culture supernatant (zone of inhibition =35 mm in the absence of methionine, no zone in the presenceof methionine), consistent with the presence of valclavamand (or) 2-hydroxyethylclavam (Zelyas et al. 2008).No methionine antimetabolite bioactivity was seen in culture
supernatants of cas3 mutants, confirming the predicted func-tion of cas3 as an early gene essential for clavam production.Surprisingly, however, methionine antimetabolite bioactivitywas also absent from culture supernatants of oxr mutants.Therefore, despite its lack of a counterpart in S. clavuligerus,oxr is essential for clavam production in S. antibioticus.Culture supernatants were also treated with imidazole to
derivatize clavam metabolites specifically and were then sub-jected to analysis by HPLC. Chromatographic profiles forwild-type culture supernatants showed a very prominentpeak at retention time 5.0 min, with an absorption maximumat 311 nm, typical of imidazole-derivatized clavam com-pounds (Fig. 6A). When this derivatized supernatant was an-alyzed by HPLC with on-line MS detection, the prominentpeak gave a total mass ion of 398, fragmented mass ion of330, consistent with the presence of valclavam (Fig. 6D). Asmaller A311 absorbing peak seen in the HPLC profile at re-tention time 3.5 min did not show a mass corresponding to2-hydroxyethylclavam, and so its identity is unknown, andno peak corresponding to 2-hydroxyethylclavam was detectedin culture supernatants under the growth conditions used.When cas3 mutant strains were cultivated under identical
conditions and subjected to HPLC analysis, no evidence of
Fig. 5. Gene clusters for clavam biosynthesis in Streptomyces antibioticus and Streptomyces clavuligerus. Genes encoding early steps of cla-vam biosynthesis are shaded black. Genes essential for synthesis of all 5S clavams are shaded dark grey. Genes essential for alanylclavambiosynthesis only are shaded light gray.
420 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.
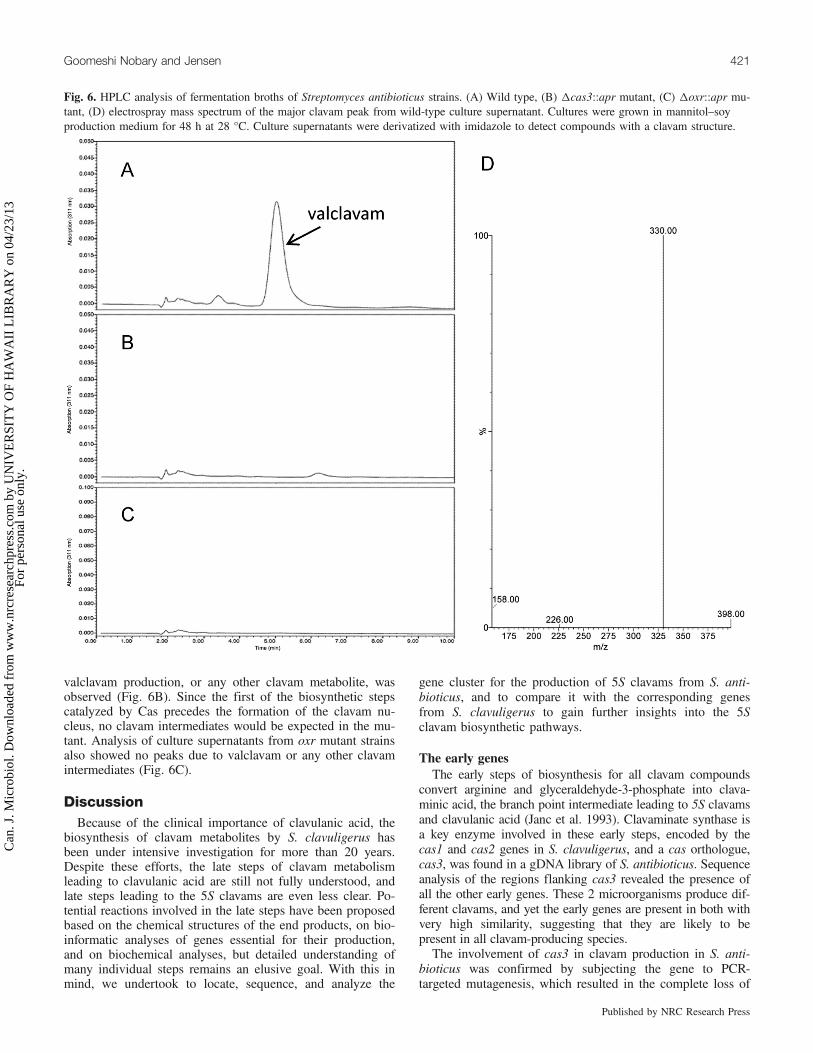
valclavam production, or any other clavam metabolite, wasobserved (Fig. 6B). Since the first of the biosynthetic stepscatalyzed by Cas precedes the formation of the clavam nu-cleus, no clavam intermediates would be expected in the mu-tant. Analysis of culture supernatants from oxr mutant strainsalso showed no peaks due to valclavam or any other clavamintermediates (Fig. 6C).
DiscussionBecause of the clinical importance of clavulanic acid, the
biosynthesis of clavam metabolites by S. clavuligerus hasbeen under intensive investigation for more than 20 years.Despite these efforts, the late steps of clavam metabolismleading to clavulanic acid are still not fully understood, andlate steps leading to the 5S clavams are even less clear. Po-tential reactions involved in the late steps have been proposedbased on the chemical structures of the end products, on bio-informatic analyses of genes essential for their production,and on biochemical analyses, but detailed understanding ofmany individual steps remains an elusive goal. With this inmind, we undertook to locate, sequence, and analyze the
gene cluster for the production of 5S clavams from S. anti-bioticus, and to compare it with the corresponding genesfrom S. clavuligerus to gain further insights into the 5Sclavam biosynthetic pathways.
The early genesThe early steps of biosynthesis for all clavam compounds
convert arginine and glyceraldehyde-3-phosphate into clava-minic acid, the branch point intermediate leading to 5S clavamsand clavulanic acid (Janc et al. 1993). Clavaminate synthase isa key enzyme involved in these early steps, encoded by thecas1 and cas2 genes in S. clavuligerus, and a cas orthologue,cas3, was found in a gDNA library of S. antibioticus. Sequenceanalysis of the regions flanking cas3 revealed the presence ofall the other early genes. These 2 microorganisms produce dif-ferent clavams, and yet the early genes are present in both withvery high similarity, suggesting that they are likely to bepresent in all clavam-producing species.The involvement of cas3 in clavam production in S. anti-
bioticus was confirmed by subjecting the gene to PCR-targeted mutagenesis, which resulted in the complete loss of
Fig. 6. HPLC analysis of fermentation broths of Streptomyces antibioticus strains. (A) Wild type, (B) Dcas3::apr mutant, (C) Doxr::apr mu-tant, (D) electrospray mass spectrum of the major clavam peak from wild-type culture supernatant. Cultures were grown in mannitol–soyproduction medium for 48 h at 28 °C. Culture supernatants were derivatized with imidazole to detect compounds with a clavam structure.
Goomeshi Nobary and Jensen 421
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.
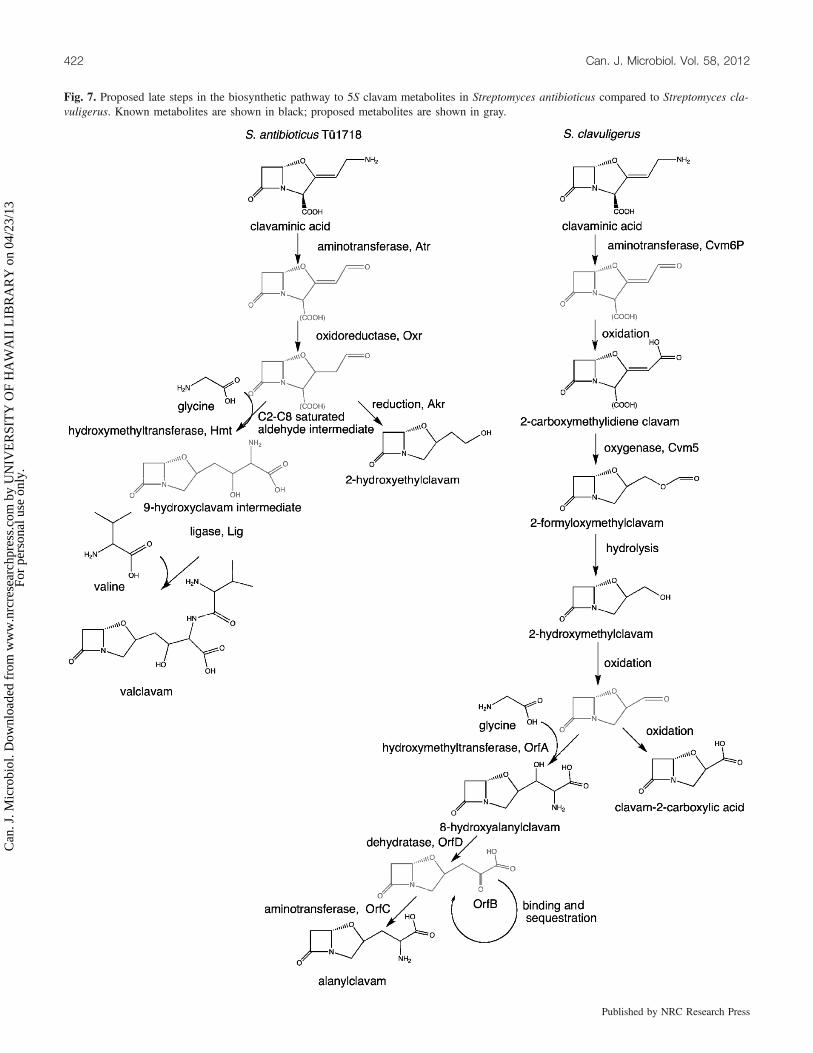
Fig. 7. Proposed late steps in the biosynthetic pathway to 5S clavam metabolites in Streptomyces antibioticus compared to Streptomyces cla-vuligerus. Known metabolites are shown in black; proposed metabolites are shown in gray.
422 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.

valclavam production in the mutant. The complete loss ofclavam production in this cas3 mutant strongly suggests thatthere is only 1 cas gene in the genome of S. antibioticus.One of the reasons offered for the presence of each of theearly genes at 2 copies in S. clavuligerus is the participationof these enzymes in 2 pathways, leading to clavulanic acidand to the 5S clavams (Marsh et al. 1992). Since S. antibioti-cus produces only 5S clavams, single copies of the earlyclavam biosynthetic genes were predicted. This expectationwas largely borne out, although a second copy of pah ispresent in the cluster. Pah removes the guanidino group fromguanidinoproclavaminate (Wu et al. 1995) and is an essentialenzyme in the pathway, but all other essential genes arepresent as single copies only, and so there is no clear explan-ation for the 2 copies of pah.
The late genesGenes essential for the late steps of 5S clavam production
are distributed across the paralogue and clavam gene clustersin S. clavuligerus (Fig. 5; Tahlan et al. 2007). The paraloguegene cluster is so named because it includes second copies ofall of the early genes except for cas2 (Tahlan et al. 2004a) aswell as second copies of cvm6 and cvm7, 2 late genes fromthe clavam cluster. These latter 2 genes, cvm6p and cvm7p,are both essential for production of all 5S clavams, and theparalogue cluster also includes orfA, orfB, orfC, and orfD,encoding proteins resembling enzymes of amino acid biosyn-thesis, which are essential for alanylclavam production only(Zelyas et al. 2008). The second copy of cas2 (cas1) is lo-cated in the clavam cluster flanked by additional genes im-portant or essential for the late steps of 5S clavamproduction (Marsh et al. 1992; Mosher et al. 1999).Organization of the clavam gene cluster is much more
straightforward in S. antibioticus. Southern analysis of ourcosmid library yielded a single group of clones with overlap-ping inserts, indicating only 1 cluster. Several of the ORFssurrounding the early genes in the clavam gene cluster inS. antibioticus appear to encode enzymes for the late stepsof 5S clavam production. By comparing these ORFs withtheir corresponding genes from S. clavuligerus and takinginto account the differences in structure of the metabolitesproduced by each species, we have proposed a biosyntheticpathway to 8-hydroxyethylclavam and valclavam that is con-sistent with these data (Fig. 7). Although speculative at thisstage, the pathway is meant to serve as a framework to pointout the differences between the 2 species.The proposed pathway leading to valclavam in S. antibioti-
cus is much simpler than that to alanylclavam in S. clavulige-rus. It begins with an aminotransferase reaction common toboth pathways, but then the unsaturated aldehyde productcould serve directly as substrate for the enoylreductase-typeactivity of Oxr in S. antibioticus. In contrast, since S. clavuli-gerus apparently employs the Cvm5 oxygenase in place ofOxr, a more circuitous route is followed giving rise to a dif-ferent suite of clavam products. The proposed central role forOxr in 5S clavam biosynthesis in S. antibioticus is consistentwith our finding that mutation of oxr resulted in completeloss of clavam production. Bicyclic intermediates of clavambiosynthesis are typically unstable, and aldehyde intermedi-ates have especially short half-lives (MacKenzie et al. 2007),
which may explain why accumulation of clavam intermedi-ates was not observed in the oxr mutants.The C2–C8 saturated aldehyde product of Oxr could then
serve as substrate for a hydroxymethyltransferase reactioncatalyzed by Htr, entirely analogous to the hydroxymethyl-transferase reaction catalyzed by OrfA in alanylclavam bio-synthesis, except that Htr yields a 9-hydroxyclavamintermediate, whereas OrfA yields an 8-hydroxyalanylclavamintermediate (Zelyas et al. 2008). Since there is no evidencefor additional amino acid biosynthetic-type genes in theS. antibioticus cluster, we propose that the final step in val-clavam biosynthesis may involve addition of preformed va-line to the 9-hydroxyclavam intermediate, perhaps catalyzedby the ligase encoded by the novel lig gene. In contrast, thealanyl moiety of alanylclavam is apparently assembled on the8-hydroxyalanylclavam intermediate by de novo synthesis,consistent with the presence of the orfB, orfC, and orfDgenes in S. clavuligerus, which are essential for alanylclavamproduction (Zelyas et al. 2008).Clavamycin-type 5S clavam metabolites produced by other
species reported in the literature (King et al. 1986; Naegeli etal. 1986), have substituent groups at C-2 apparently derivedfrom 8-hydroxyalanylclavam, suggesting that the “alanyl-clavam” version of the 5S clavam pathway is involved intheir synthesis and thus may be more common in naturethan the “valclavam” pathway of S. antibioticus. Further ge-netic analyses of a broader group of clavam-producing spe-cies should resolve this question. In that regard,bioinformatic examination of the ORFs surrounding the earlyclavam biosynthetic genes in the genome of S. flavogriseusdid not provide any clarification because only clavulanicacid-type genes rather than 5S clavam-type genes were evi-dent, suggesting that this species produces clavulanic acidbut not 5S clavams.Validation of the proposed pathway to 5S clavams in
S. antibioticus will require the generation of additional mu-tants and in vitro biochemical analyses of the proposed indi-vidual steps. The inherent instability and lack of commercialavailability of putative biosynthetic intermediates makes suchstudies very challenging, but they should ultimately give afull understanding of the pathways for this important class ofmetabolites.
AcknowledgementsThis study was funded by Discovery grant 2824-06 from
the Natural Sciences and Engineering Research Council ofCanada.
ReferencesArulanantham, H., Kershaw, N.J., Hewitson, K.S., Hughes, C.E.,
Thirkettle, J.E., and Schofield, C.J. 2006. ORF17 from theclavulanic acid biosynthesis gene cluster catalyzes the ATP-dependent formation of N-glycyl-clavaminic acid. J. Biol. Chem.281(1): 279–287. doi:10.1074/jbc.M507711200. PMID:16251194.
Baggaley, K.H., Brown, A.G., and Schofield, C.J. 1997. Chemistry andbiosynthesis of clavulanic acid and other clavams. Nat. Prod. Rep.14(4): 309–333. doi:10.1039/np9971400309. PMID:9281835.
Baldwin, J.E., Fujishima, Y., Goh, K.-C., and Schofield, C.J. 1994.Enzymes of valclavam biosynthesis. Tetrahedron Lett. 359(17):2783–2786. doi:10.1016/S0040-4039(00)77032-8.
Goomeshi Nobary and Jensen 423
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.

Bierman, M., Logan, R., O’Brien, K., Seno, E.T., Rao, R.N., andSchoner, B.E. 1992. Plasmid cloning vectors for the conjugaltransfer of DNA from Escherichia coli to Streptomyces spp. Gene,116(1): 43–49. doi:10.1016/0378-1119(92)90627-2. PMID:1628843.
Birnboim, H.C., and Doly, J. 1979. A rapid alkaline extractionprocedure for screening recombinant plasmid DNA. Nucleic AcidsRes. 7(6): 1513–1523. doi:10.1093/nar/7.6.1513. PMID:388356.
Caines, M.E.C., Elkins, J.M., Hewitson, K.S., and Schofield, C.J.2004. Crystal structure and mechanistic implications of N2-(2-carboxyethyl)arginine synthase, the first enzyme in the clavulanicacid biosynthesis pathway. J. Biol. Chem. 279(7): 5685–5692.doi:10.1074/jbc.M310803200. PMID:14623876.
de la Fuente, A., Martin, J.F., Rodriguez-Garcia, A., and Liras, P.2004. Two proteins with ornithine acetyltransferase activity showdifferent functions in Streptomyces clavuligerus: Oat2 modulatesclavulanic acid biosynthesis in response to arginine. J. Bacteriol.186(19): 6501–6507. doi:10.1128/JB.186.19.6501-6507.2004.PMID:15375131.
Egan, L.A., Busby, R.W., Iwata-Reuyl, D., and Townsend, C.A.1997. Probable role of clavaminic acid as the terminal intermediatein the common pathway to clavulanic acid and the antipodalclavam metabolites. J. Am. Chem. Soc. 119(10): 2348–2355.doi:10.1021/ja963107o.
Elkins, J.M., Clifton, I.J., Hernández, H., Doan, L.X., Robinson, C.V.,Schofield, C.J., and Hewitson, K.S. 2002. Oligomeric structureof proclavaminic acid amidino hydrolase: evolution of ahydrolytic enzyme in clavulanic acid biosynthesis. Biochem. J.366(2): 423–434. doi:10.1042/BJ20020125. PMID:12020346.
Foulstone, M., and Reading, C. 1982. Assay of amoxicillin andclavulanic acid, the components of augmentin, in biological fluidswith high-performance liquid chromatography. Antimicrob.Agents Chemother. 22(5): 753–762. PMID:7181486.
Galperin, M.Y., and Koonin, E.V. 1997. A diverse superfamily ofenzymes with ATP-dependent carboxylate-amine/thiol ligaseactivity. Protein Sci. 6(12): 2639–2643. doi:10.1002/pro.5560061218. PMID:9416615.
Gust, B., Challis, G.L., Fowler, K., Kieser, T., and Chater, K.F. 2003.PCR-targeted Streptomyces gene replacement identifies a proteindomain needed for biosynthesis of the sesquiterpene soil odorgeosmin. Proc. Natl. Acad. Sci. U.S.A. 100(4): 1541–1546.doi:10.1073/pnas.0337542100. PMID:12563033.
Janc, J.W., Egan, L.A., and Townsend, C.A. 1993. Emergingevidence for a shared biosynthetic pathway among clavulanicacid and the structurally diverse clavam metabolites. Bioorg. Med.Chem. Lett. 3(11): 2313–2316. doi:10.1016/S0960-894X(01)80946-X.
Janc, J.W., Egan, L.A., and Townsend, C.A. 1995. Purification andcharacterization of clavaminate synthase from Streptomycesantibioticus. A multifunctional enzyme of clavam biosynthesis.J. Biol. Chem. 270(10): 5399–5404. doi:10.1074/jbc.270.10.5399.PMID:7890654.
Jensen, S.E., and Paradkar, A.S. 1999. Biosynthesis and moleculargenetics of clavulanic acid. Antonie van Leeuwenhoek, 75(1–2):125–133. PMID:10422585. doi:10.1023/A:1001755724055.
Jensen, S.E., Elder, K.J., Aidoo, K.A., and Paradkar, A.S. 2000.Enzymes catalyzing the early steps of clavulanic acid biosynthesisare encoded by two sets of paralogous genes in Streptomycesclavuligerus. Antimicrob. Agents Chemother. 44(3): 720–726.doi:10.1128/AAC.44.3.720-726.2000. PMID:10681345.
Jensen, S.E., Paradkar, A.S., Mosher, R.H., Anders, C., Beatty, P.H.,Brumlik, M.J., et al. 2004a. Five additional genes are involved inclavulanic acid biosynthesis in Streptomyces clavuligerus. Anti-
microb. Agents Chemother. 48(1): 192–202. doi:10.1128/AAC.48.1.192-202.2004. PMID:14693539.
Jensen, S.E., Wong, A., Griffin, A., and Barton, B. 2004b.Streptomyces clavuligerus has a second copy of the proclavami-nate amidinohydrolase gene. Antimicrob. Agents Chemother.48(2): 514–520. doi:10.1128/AAC.48.2.514-520.2004. PMID:14742203.
Kieser, T., Bibb, M.J., Buttner, M.J., Chater, K.F., and Hopwood, D.A.2000. Practical Streptomyces genetics. The John Innes Founda-tion, Norwich, England.
King, H.D., Langharig, J., and Sanglier, J.J. 1986. Clavamycins, newclavam antibiotics from two variants of Streptomyces hygroscopi-cus. I. Taxonomy of the producing organisms, fermentation, andbiological activities. J. Antibiot. (Tokyo), 39(4): 510–515. PMID:3710912.
Li, R., Khaleeli, N., and Townsend, C.A. 2000. Expansion of theclavulanic acid gene cluster: identification and in vivo functionalanalysis of three new genes required for biosynthesis of clavulanicacid by Streptomyces clavuligerus. J. Bacteriol. 182(14): 4087–4095. doi:10.1128/JB.182.14.4087-4095.2000. PMID:10869089.
MacKenzie, A.K., Kershaw, N.J., Hernandez, H., Robinson, C.V.,Schofield, C.J., and Andersson, I. 2007. Clavulanic aciddehydrogenase: structural and biochemical analysis of the finalstep in the biosynthesis of the b-lactamase inhibitor clavulanicacid. Biochemistry, 46(6): 1523–1533. doi:10.1021/bi061978x.PMID:17279617.
Marsh, E.N., Chang, M.D., and Townsend, C.A. 1992. Two isozymesof clavaminate synthase central to clavulanic acid formation:cloning and sequencing of both genes from Streptomycesclavuligerus. Biochemistry, 31(50): 12 648 – 12 657. doi:10.1021/bi00165a015. PMID:1472501.
Medema, M.H., Trefzer, A., Kovalchuk, A., van den Berg, M.,Müller, U., Heijne, W., et al. 2010. The sequence of a 1.8-Mbbacterial linear plasmid reveals a rich evolutionary reservoir ofsecondary metabolic pathways. Genome Biol. Evol. 2(0): 212–224. doi:10.1093/gbe/evq013. PMID:20624727.
Mellado, E., Lorenzana, L.M., Rodríguez-Sáiz, M., Díez, B., Liras,P., and Barredo, J.L. 2002. The clavulanic acid biosynthetic clusterof Streptomyces clavuligerus: genetic organization of the regionupstream of the car gene. Microbiology, 148(5): 1427–1438.PMID:11988517.
Miller, M.T., Bachmann, B.O., Townsend, C.A., and Rosenzweig, A.C.2001. Structure of b-lactam synthetase reveals how to synthesizeantibiotics instead of asparagine. Nat. Struct. Biol. 8(8): 684–689. doi:10.1038/90394. PMID:11473258.
Mosher, R.H., Paradkar, A.S., Anders, C., Barton, B., and Jensen, S.E.1999. Genes specific for the biosynthesis of clavam metabolitesantipodal to clavulanic acid are clustered with the gene forclavaminate synthase 1 in Streptomyces clavuligerus. Antimi-crob. Agents Chemother. 43(5): 1215–1224. PMID:10223939.
Naegeli, H.U., Loosli, H.R., and Nussbaumer, A. 1986. Clavamycins,new clavam antibiotics from two variants of Streptomyceshygroscopicus. II. Isolation and structures of clavamycins A, Band C from Streptomyces hygroscopicus NRRL 15846, and ofclavamycins D, E and F from Streptomyces hygroscopicus NRRL15879. J. Antibiot. (Tokyo), 39(4): 516–524. PMID:3710913.
Pao, S.S., Paulsen, I.T., and Saier, M.H., Jr. 1998. Major facilitatorsuperfamily. Microbiol. Mol. Biol. Rev. 62(1): 1–34. PMID:9529885.
Paradkar, A.S., and Jensen, S.E. 1995. Functional analysis of the geneencoding the clavaminate synthase 2 isoenzyme involved inclavulanic acid biosynthesis in Streptomyces clavuligerus. J.Bacteriol. 177(5): 1307–1314. PMID:7868606.
Paradkar, A.S., Mosher, R.H., Anders, C., Griffin, A., Griffin, J.,
424 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.

Hughes, C., et al. 2001. Applications of gene replacementtechnology to Streptomyces clavuligerus strain development forclavulanic acid production. Appl. Environ. Microbiol. 67(5):2292–2297. doi:10.1128/AEM.67.5.2292-2297.2001. PMID:11319114.
Pruess, D.L., and Kellett, M. 1983. Ro 22–5417, a new clavamantibiotic from Streptomyces clavuligerus. I. Discovery andbiological activity. J. Antibiot. (Tokyo), 36(3): 208–212. PMID:6833140.
Röhl, F., Rabenhorst, J., and Zähner, H. 1987. Biological propertiesand mode of action of clavams. Arch. Microbiol. 147(4): 315–320.doi:10.1007/BF00406126. PMID:3304182.
Sambrook, J., Fritsch, E.F., and Maniatas, T. 1989. Molecularcloning: a laboratory manual. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, N.Y., USA.
Tahlan, K., Anders, C., and Jensen, S.E. 2004a. The paralogous pairsof genes involved in clavulanic acid and clavam metabolitebiosynthesis are differently regulated in Streptomyces clavuligerus.J. Bacteriol. 186(18): 6286–6297. doi:10.1128/JB.186.18.6286-6297.2004. PMID:15342599.
Tahlan, K., Park, H.U., and Jensen, S.E. 2004b. Three unlinked geneclusters are involved in clavam metabolite biosynthesis inStreptomyces clavuligerus. Can. J. Microbiol. 50(10): 803–810.doi:10.1139/w04-070. PMID:15644894.
Tahlan, K., Anders, C., Wong, A., Mosher, R.H., Beatty, P.H.,Brumlik, M.J., et al. 2007. 5S Clavam biosynthetic genes arelocated in both the clavam and paralog gene clusters inStreptomyces clavuligerus. Chem. Biol. 14(2): 131–142. doi:10.1016/j.chembiol.2006.11.012. PMID:17317567.
Wu, T.K., Busby, R.W., Houston, T.A., McIlwaine, D.B., Egan, L.A.,and Townsend, C.A. 1995. Identification, cloning, sequencing,and overexpression of the gene encoding proclavaminate amidinohydrolase and characterization of protein function in clavulanicacid biosynthesis. J. Bacteriol. 177(13): 3714–3720. PMID:7601835.
Yanto, Y., Hall, M., and Bommarius, A.S. 2010. Nitroreductase fromSalmonella typhimurium: characterization and catalytic activity.Org. Biomol. Chem. 8(8): 1826–1832. doi:10.1039/b926274a.PMID:20449486.
Zelyas, N.J., Cai, H., Kwong, T., and Jensen, S.E. 2008.Alanylclavam biosynthetic genes are clustered together with onegroup of clavulanic acid biosynthetic genes in Streptomycesclavuligerus. J. Bacteriol. 190(24): 7957–7965. doi:10.1128/JB.00698-08. PMID:18931110.
Zhang, Z., Ren, J., Stammers, D.K., Baldwin, J.E., Harlos, K., andSchofield, C.J. 2000. Structural origins of the selectivity of thetrifunctional oxygenase clavaminic acid synthase. Nat. Struct.Biol. 7(2): 127–133. doi:10.1038/72398. PMID:10655615.
Goomeshi Nobary and Jensen 425
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
HA
WA
II L
IBR
AR
Y o
n 04
/23/
13Fo
r pe
rson
al u
se o
nly.