a- compartiments et transport i 26dec
DESCRIPTION
ÂTRANSCRIPT


Compartiments et transport intracellulaires
A- Compartiments intracellulaires et tri des protéines
B-Transport vésiculaire dans les voies de sécrétion et d’endocytose





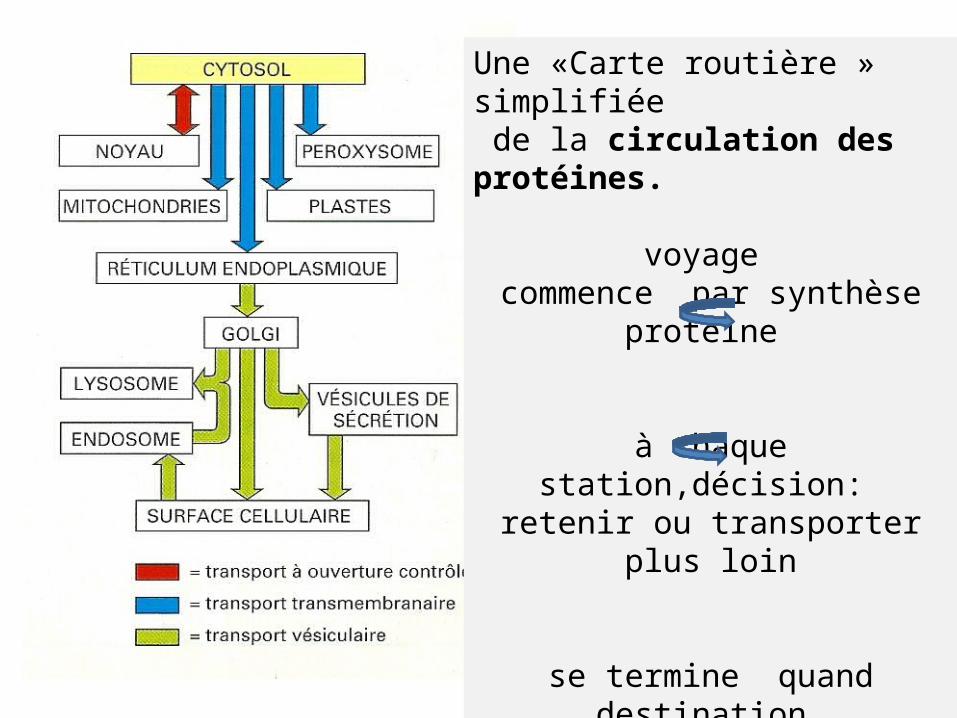
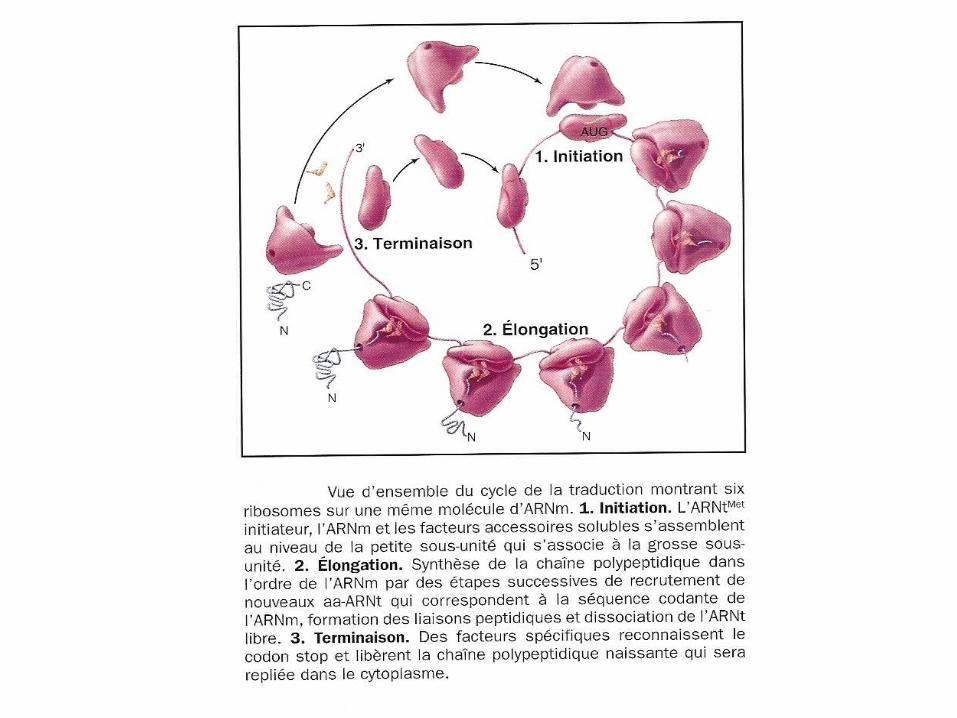
Une «Carte routière » simplifiée de la circulation des protéines.
voyage commence par synthèse
protéine
à chaque station,décision: retenir ou transporter plus loin
se termine quand destination finale atteinte
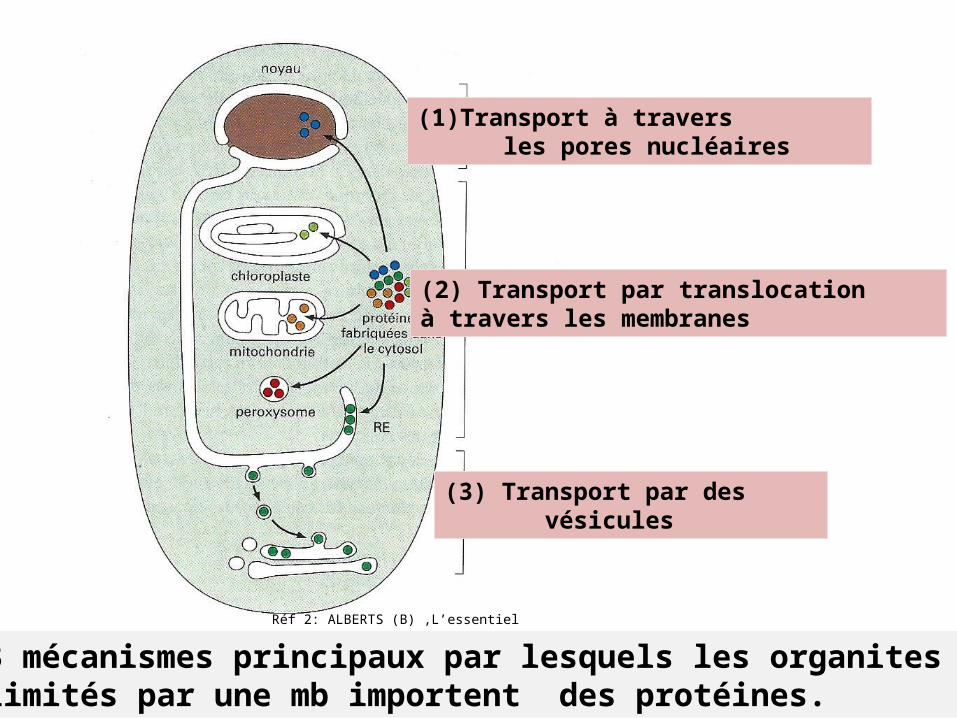
Réf 2: ALBERTS (B) ,L’essentiel
(1) Transport à travers les pores nucléaires
(2) Transport par translocationà travers les membranes
(3) Transport par des vésicules
3 mécanismes principaux par lesquels les organites limités par une mb importent des protéines.

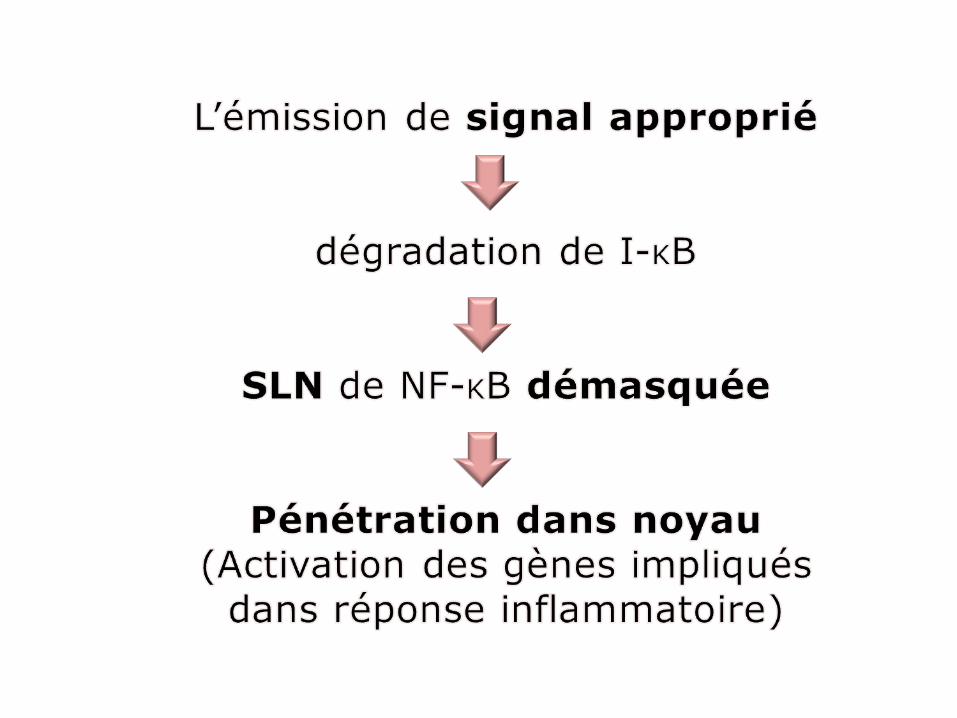
Interactions entre signaux et récepteursorientent les constituants cellulaires
vers leur destination correcte
des signaux de reconnaissance spécifique intégrés dans structure protéines et *AN, guident la migration vers le compartiment
auquel elles sont destinées
*AN : acides nucléiques

I . Compartiments membranaires


Bactéries
Compartiment intracell unique(complexes multi
- protéiques )
Eucaryotes
Compartiments limités par mb
fonctionnellement distincts
Cellule = compartiment qui isole un ensemble de macromolécules du reste de l’univers!
I-Avantages de la compartimentation

Le maintien de la compartimentation est la clé de la permanence de l’identité cellulaire
Or une cellule vit et interagit avec
un environnement
Il existe en plus des échanges avec l’extérieur dépendant de mb ,
un trafic mol entre les différents compartiments

Les diverses fonctions de biosynthèse, de dégradation et de production d’énergie
compartimentées dans cellules eucaryotes
Les ≠compartiments nombre , taille varient selon type cellu
orientés vers des fonctions distinctes et spécialisées . Un organite a souvent
le monopole d’une fonction donnée

Réf 2: ALBERTS (B) ,L’essentiel

Principales fonctions des compartiments d’une cellule eucaryote limités par une mb
Compartiment Principale fonction•Cytosol•Noyau•RE
•AG•Lysosomes•Endosomes •Mitochondrie•Chloroplastes
•Peroxysomes
• nombreuses voies métaboliques , synthèse protéique•contient génome , synthèse de ADN ,ARN• s’associe aux ribosomes: synthèse protéines sécrétées et protéines intégrées mb , synthèse lipides•modification , tri et empaquetage de protéines , lipides•dégradation intracellulaire (enzymes hydrolytiques)•Tri du matériel incorporé par endocytose •Synthèse de l’ATP par phosphorylation oxydative•Synthèse de l’ATP et fixation du carbone par photosynthèse•Oxydation des molécules toxiques ( contiennent la catalase et ≠ oxydases)

Les compartiments de la cellule:organites membranaires spécialisés
-Compartiments de traitement protéines(R E , A G, Vésicules de sécrétion )
-Compartiment des acides nucléiques(noyau , nucléole)
-Compartiments clos(mitochondries, chloroplastes , peroxysomes)
-Compartiment cytosolique
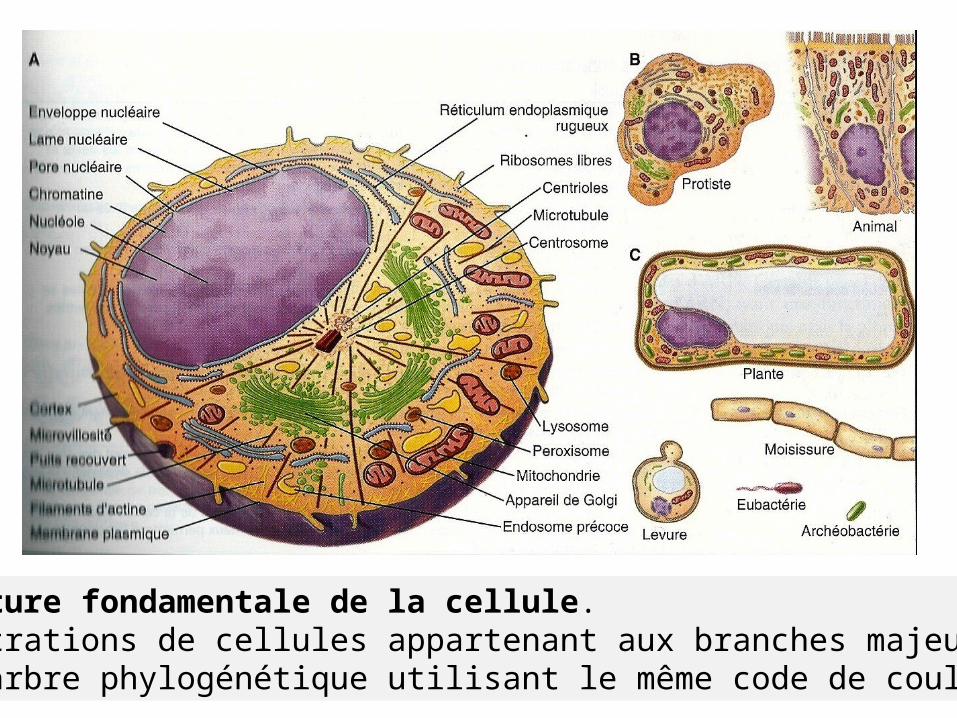
Structure fondamentale de la cellule.Illustrations de cellules appartenant aux branches majeures de l’arbre phylogénétique utilisant le même code de couleurs.
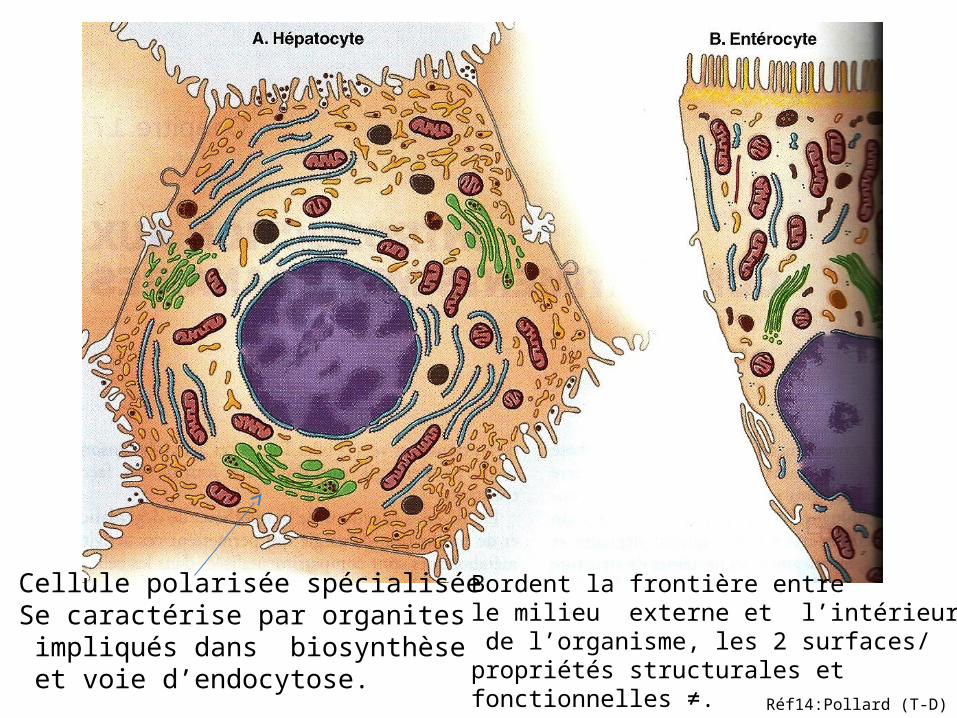
Cellule polarisée spécialisée Se caractérise par organites impliqués dans biosynthèse et voie d’endocytose.
Bordent la frontière entre le milieu externe et l’intérieur de l’organisme, les 2 surfaces/propriétés structurales et fonctionnelles ≠. Réf14:Pollard (T-D)
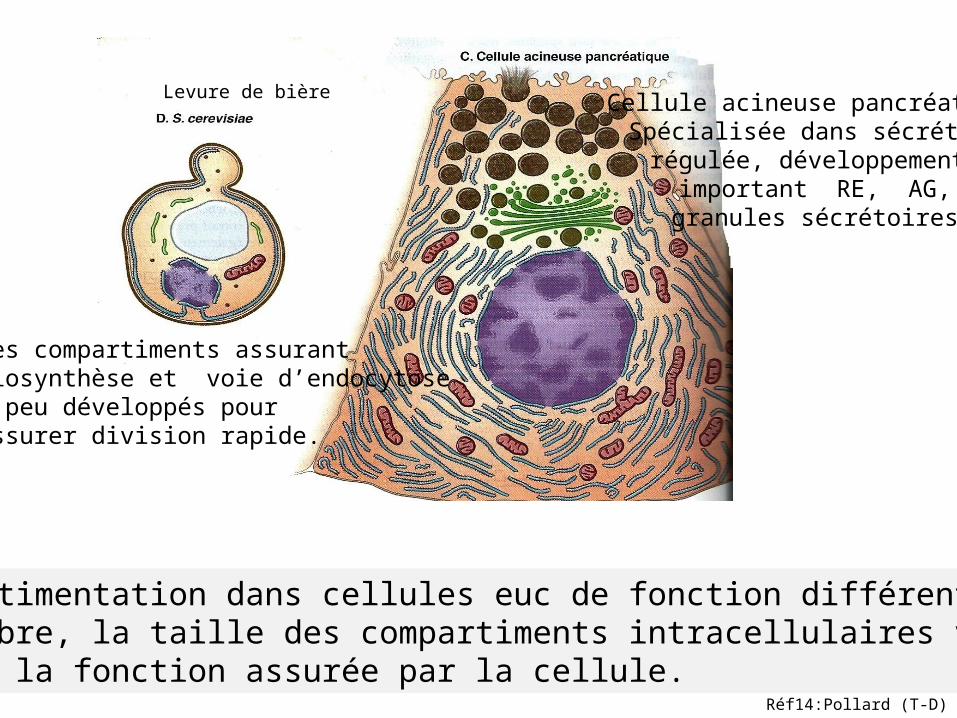
Compartimentation dans cellules euc de fonction différente.Le nombre, la taille des compartiments intracellulaires varient selon la fonction assurée par la cellule.
Cellule acineuse pancréatiqueSpécialisée dans sécrétionrégulée, développement
important RE, AG,granules sécrétoires
Levure de bière
Les compartiments assurantbiosynthèse et voie d’endocytose peu développés pour assurer division rapide.
Réf14:Pollard (T-D)

Les compartiments mb :-ne sont pas distribués au hasard- emplacements souvent caractéristiques
L’ensemble des compartiments (organites) occupe prés de ½ du volume cellulaire.
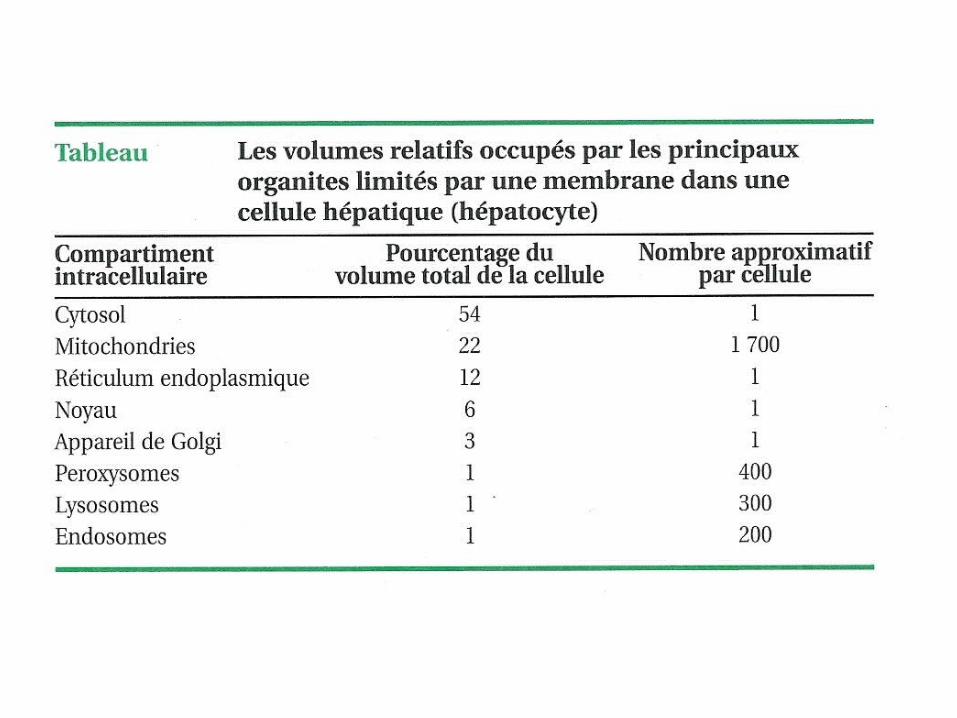

Les compartimentszones privilégiées dans volume restreint Microenvironnement
(enzymes, cofacteurs ,substrats concentrés)
Vitesse des interactions moléculaires

Environnement de part et d’autre de mb semi perméable modulée pour obtenir:
-milieu ionique adéquat ou -asymétrie ionique ( PH, gradient ionique et/ou électrique)
Mol hydrophobes à l’abri de l’eau
séquestration des activités à risque-enzymes dégradation des lysosomes -enzymes oxydatives ,peroxysomes

Comprendre la compartimentation au sein d’une cell euc !
-Savoir ce qui se passe dans ses compartiments?
-Comment les molécules circulent entre elles?
- Comment les compartiments sont créés et conservés?


[Origines des compartiments]
La compartimentation chez les cellules procaryotes a permis aux 1ers eucaryotes
- de taille- capter l’énergie plus efficacement-réguler expression gènes d’une manière plus complexe
Modèle hypothétique de l’évolution de la compartimentation intracellulaire.
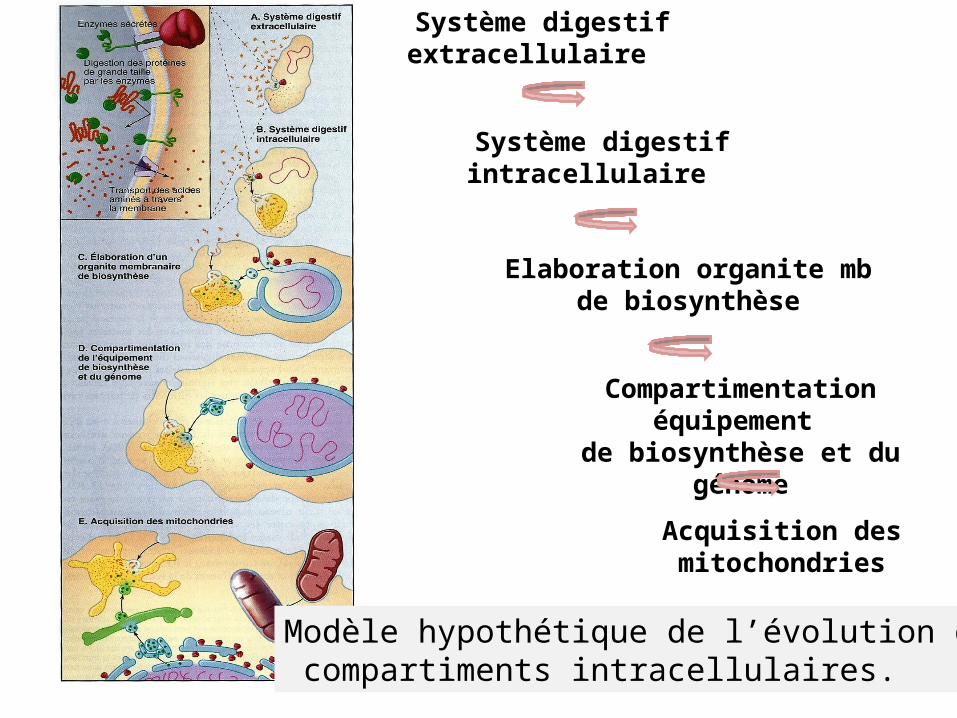
Modèle hypothétique de l’évolution des compartiments intracellulaires.
Système digestif extracellulaire
Système digestif intracellulaire
Elaboration organite mb de biosynthèse
Compartimentation équipement de biosynthèse et du génome
Acquisition des mitochondries

La description sommaire des principauxaspects de l’origine de la
compartimentation
Apprécier importance fonctionnelle des organites
et Identifier les corrélations
possibles

Procaryotes ancestraux compartimentés!
Biosynthèse à l’intérieurDigestion à l’extracellulaire
Exporter matériel de digestion(enzymes hydrolytiques / surface Ou produits de sécrétion libres)
et Capter produits de digestion (fig:A)

A .Procaryotes ancestraux, biosynthèse au sein de cellule digestion extracellulaire.B. Internalisation enzymes d’hydrolyse sécrétées avec nutriments provenant de l’environnement.
Matériel de digestion à la surface mb
Internalisation enzymes d’hydrolyse avec nutriments de l’environnement/1° étape vers la compartimentation, création d’un lysosome
Matériel génétique
Réf14:Pollard (T-D)
Canal
Transporteur

Adressage et transport protéines /mb
1° étape vers la compartimentation
Hypothèse
Invaginations des sous domaines mb impliqués dans synthèse des lipides mb et
translocation des protéines
RE :organite intracellulaire de biosynthèse

Internalisation
Enzymes d’hydrolyse sécrétées +
nutriments /environnement
Lysosome primitif (Fig:B)
l’efficacité de la digestion et l’absorption des nutriments
macromoléculaires

Présence de 2 organites intracellulaires
vésicules de transport dont éléments protéiques et lipidiques synthétisés
dans RE primitif!
Fonctionnent :-comme transporteurs (exportant vers
surface ou vacuoles et important matière 1°)-comme protecteurs (séparant enzymes
digestives du cytoplasme environnant.)

Apparition de plusieurs destinationspossibles dans cellule
Signaux d’adressagepour distinguer les ≠ circuits
les uns des autres
Stratégie "diviser et spécialiser" ulilisée grand nombre de fois
voies d’exportation et de digestion: machinerie de synthèse.(Fig:C)
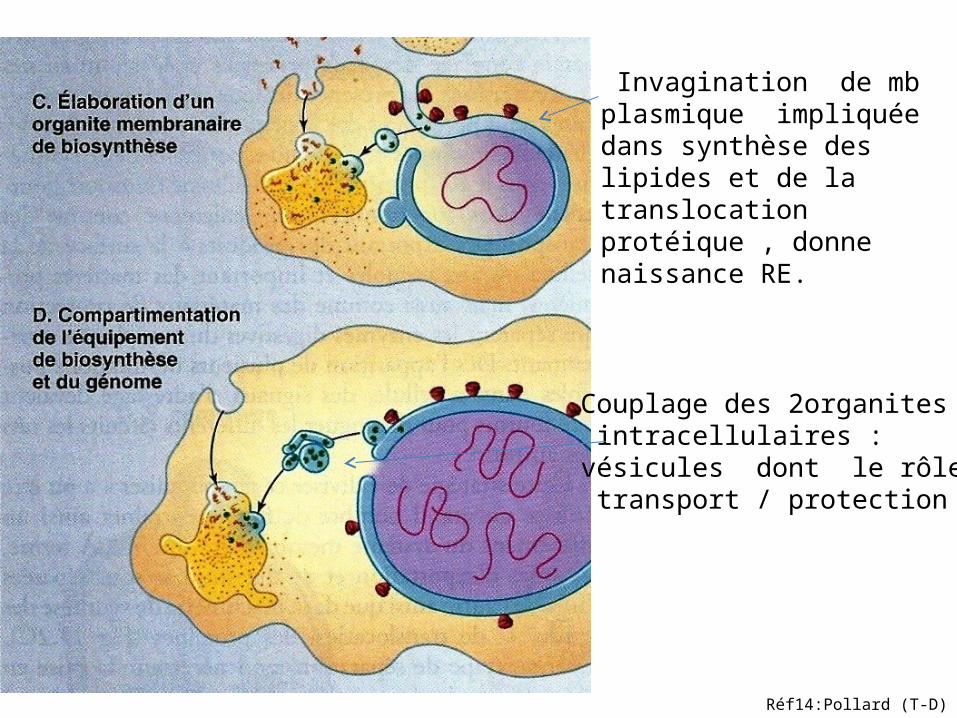
Invagination de mb plasmique impliquée dans synthèse des lipides et de la translocation protéique , donne naissance RE.
Couplage des 2organites intracellulaires :vésicules dont le rôle: transport / protection
Réf14:Pollard (T-D)
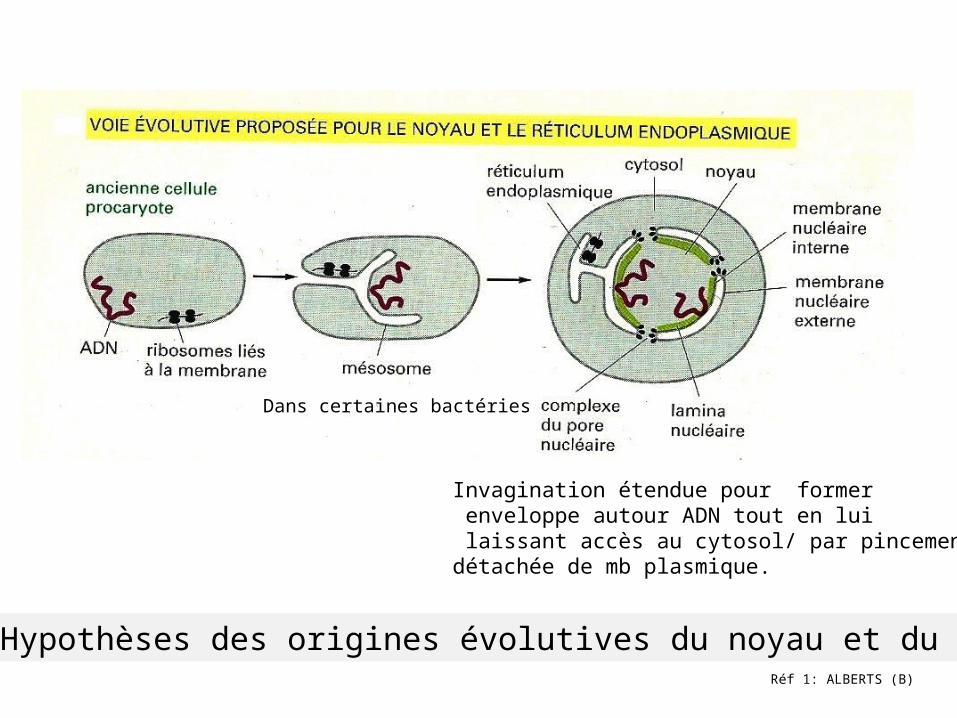
Réf 1: ALBERTS (B)
Hypothèses des origines évolutives du noyau et du RE
Dans certaines bactéries
Invagination étendue pour former enveloppe autour ADN tout en lui laissant accès au cytosol/ par pincementdétachée de mb plasmique.
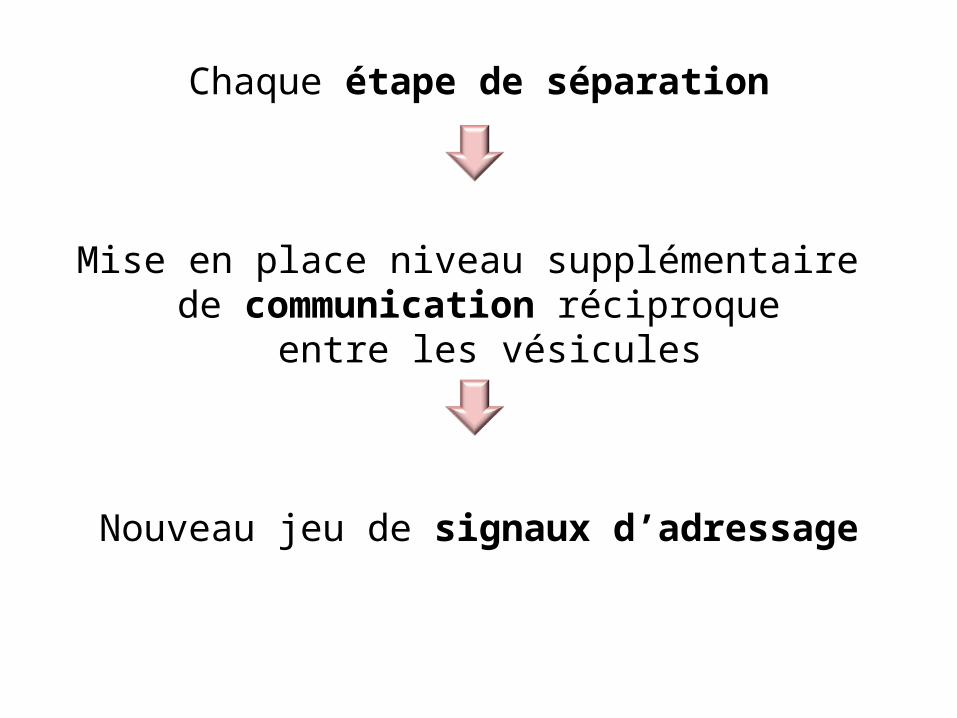
Chaque étape de séparation
Mise en place niveau supplémentaire de communication réciproque
entre les vésicules
Nouveau jeu de signaux d’adressage

Evolution/développement
système vacuolaire:RE (centre de la translocation
des protéines et synthèse des lipides), AG ,système endosome / lysosome
RE a donné naissance à enveloppe nucléaire
(génome plus complexe , séparation transcription/traduction) (Fig:D)
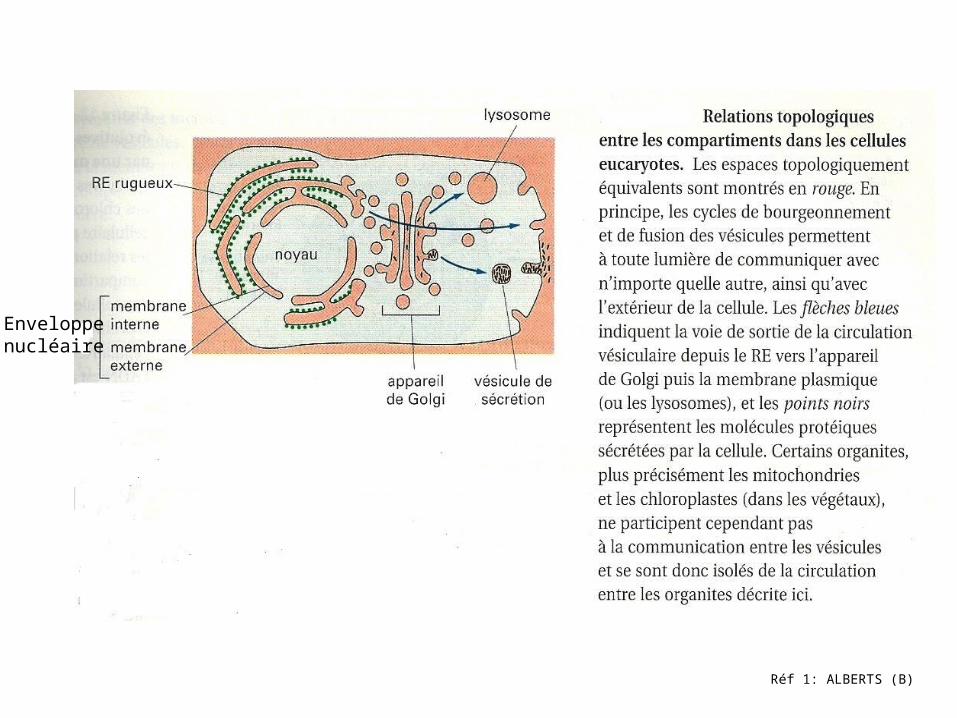
Réf 1: ALBERTS (B)
Enveloppenucléaire

Quand oxygène apparu dans atmosphère
pour exploiter ce puissant oxydant ,apparition peroxysomes (centre
dégradation oxydative)
Apparition mitochondriesSites d’utilisation d’oxygène ,
de l’extraction d’énergie libre des métabolites, et de sa conversion en ATP
(Fig:E)
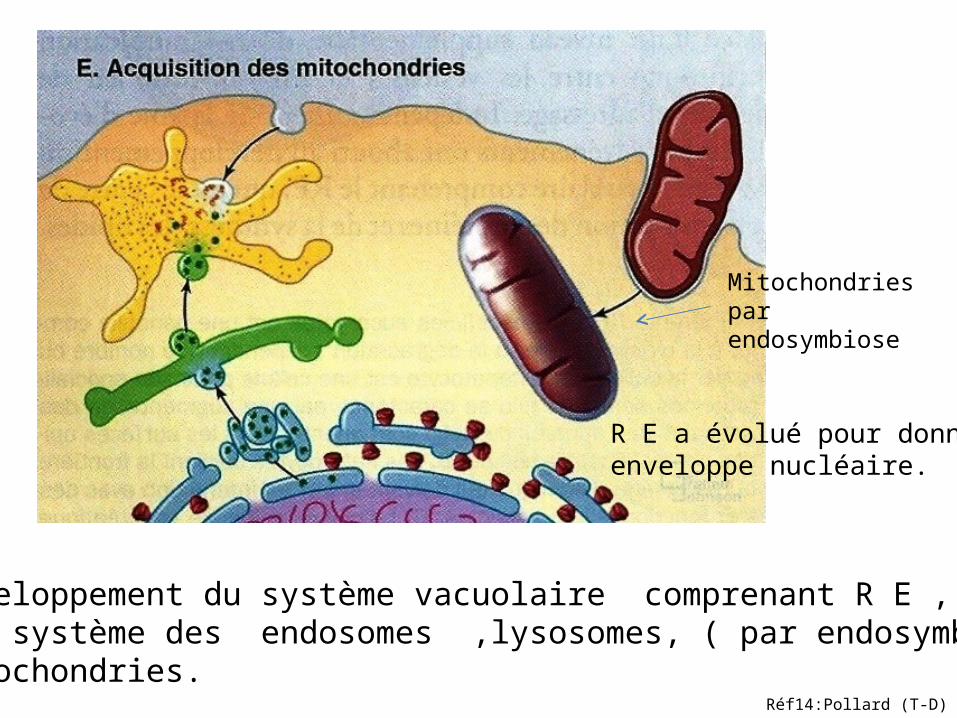
Développement du système vacuolaire comprenant R E , AG et système des endosomes ,lysosomes, ( par endosymbiose )mitochondries.
R E a évolué pour donner enveloppe nucléaire.
Mitochondriespar endosymbiose
Réf14:Pollard (T-D)
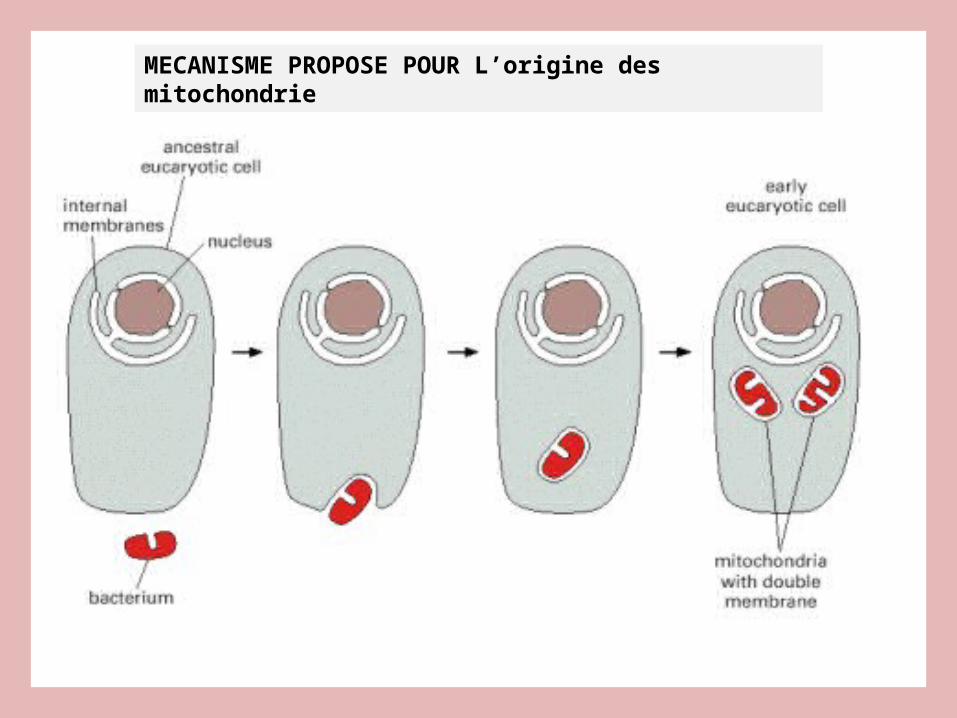
MECANISME PROPOSE POUR L’origine des mitochondrie
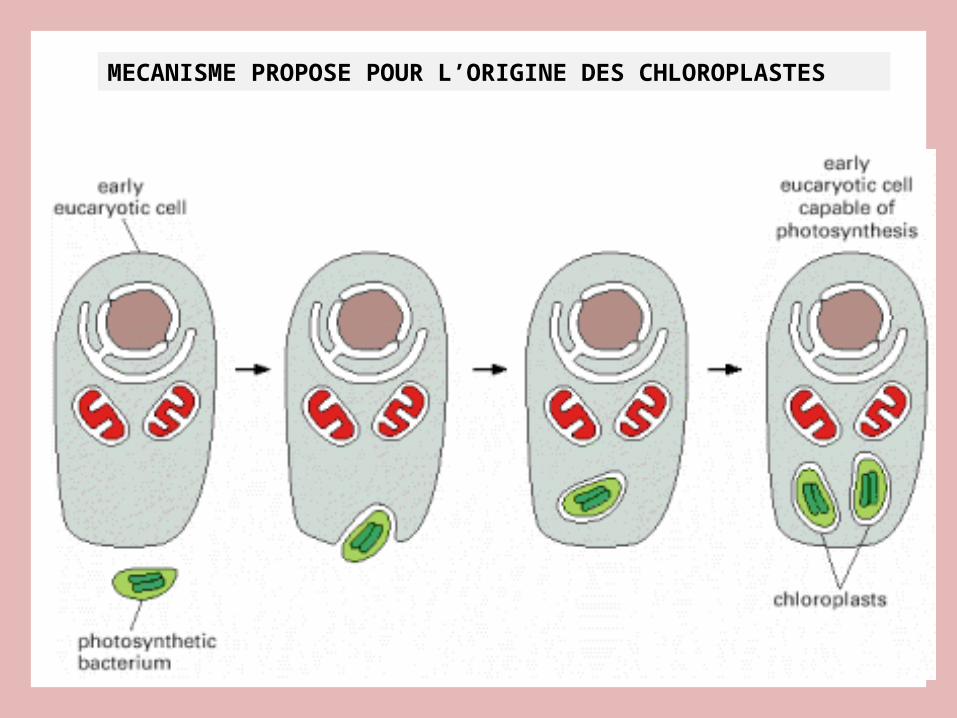
MECANISME PROPOSE POUR L’ORIGINE DES CHLOROPLASTES
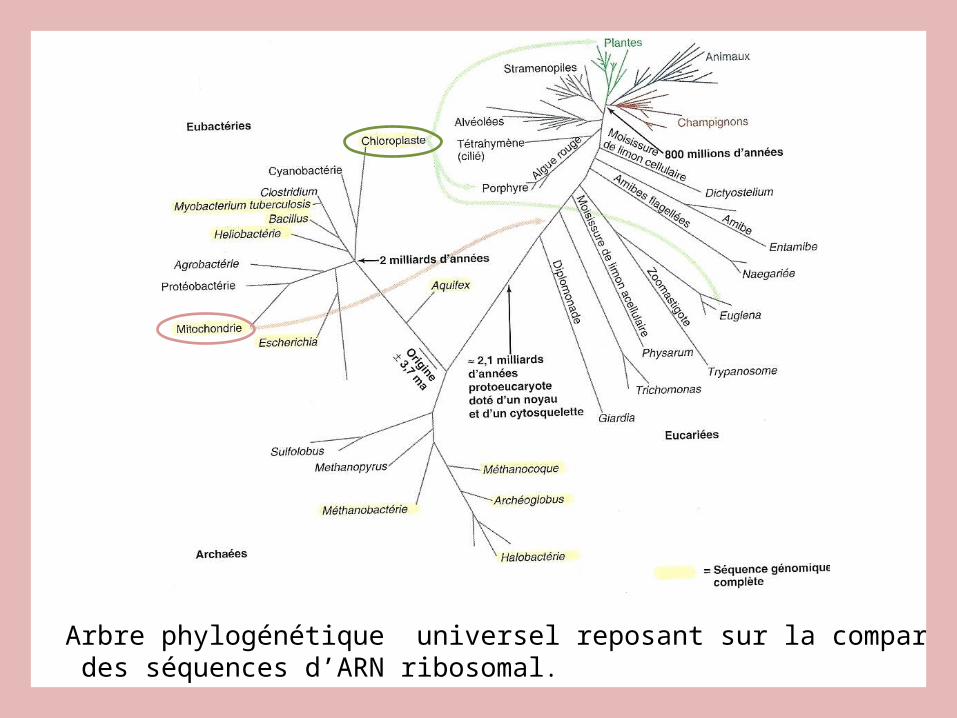
Arbre phylogénétique universel reposant sur la comparaison des séquences d’ARN ribosomal.

Mitochondries apparues par endosymbiose!
On ne sait pas si peroxysomes
ou
apparus de même manière que mitochondrie
spécialisation complémentairedu compartiment vacuolaire
(Aucune trace de procaryote dans peroxysomes.)

L’apparition de l’O2 dans l’environnement
a permis la synthèse du cholestérol et son intégration mb
Épaissit sans modifier fluidité mb ,en absence de paroi cellulaire,
solide mais flexible
Facteur important pourvolume cellu au début de l’évolution.

La compartimentation et division du travail qui l’accompagne :
Bénéfices mais aussi Problèmes
Compartiments non autonomes,leurs activités doivent être intégrées
pour bénéficier à l’ensembles de la cell,
nécessite mécanismes de transport-entre compartiments -à travers mb qui les délimitent

Réalisation d’échange entre systèmes mb
Quelles sont les molécules concernées,les compartiments et les mécanismes
impliqués?
(L’essentiel du trafic cellulaire semble basé sur les protéines)


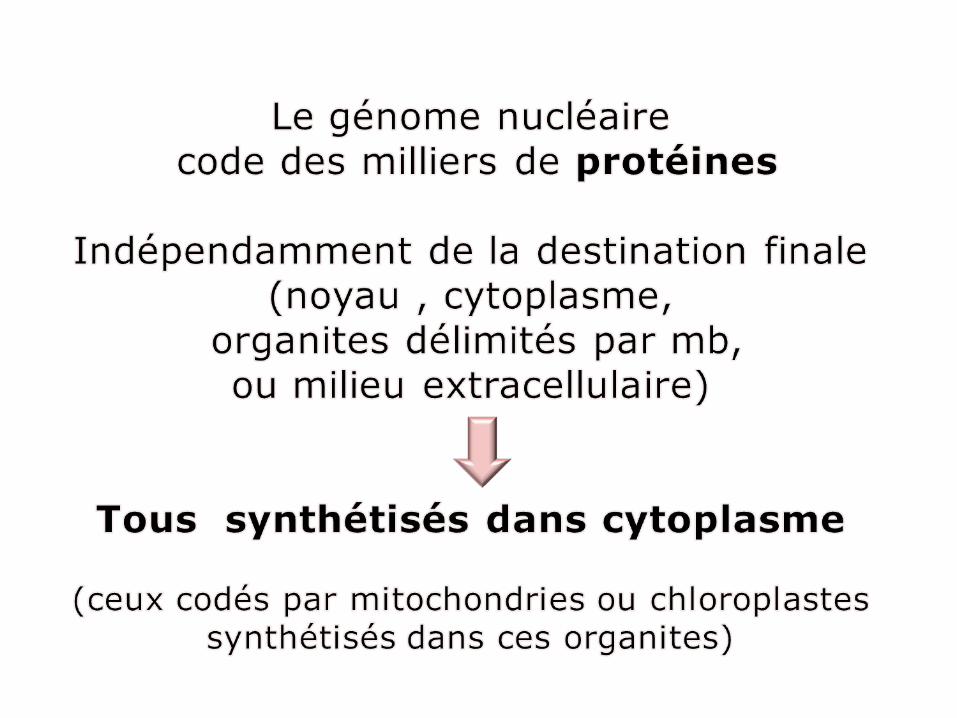
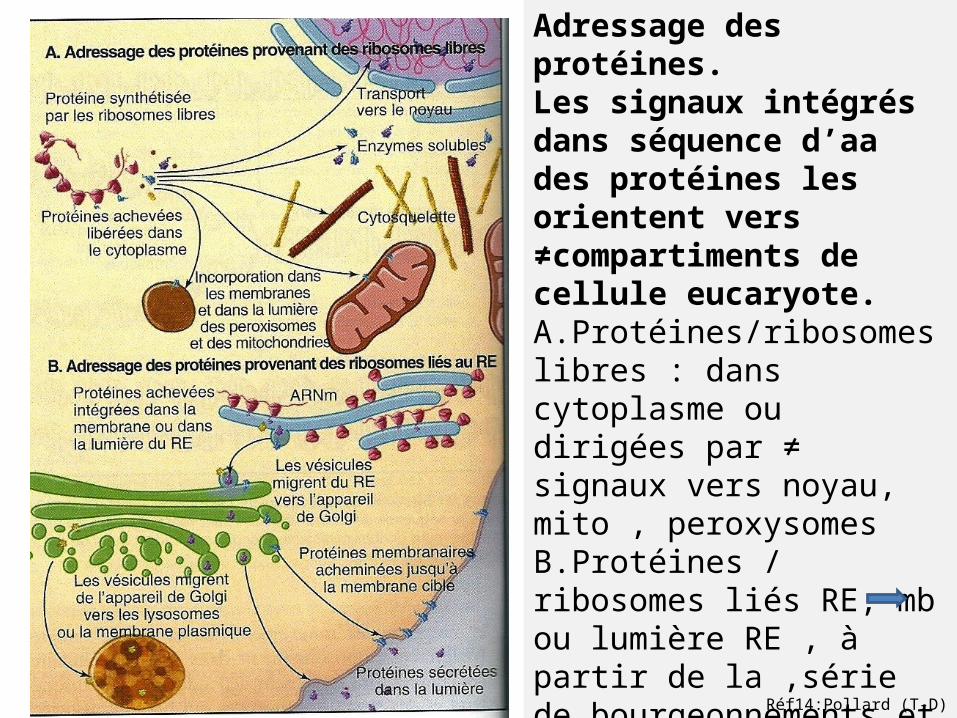
Adressage des protéines.Les signaux intégrés dans séquence d’aa des protéines les orientent vers ≠compartiments de cellule eucaryote.A.Protéines/ribosomes libres : dans cytoplasme ou dirigées par ≠ signaux vers noyau, mito , peroxysomesB.Protéines / ribosomes liés RE; mb ou lumière RE , à partir de la ,série de bourgeonnements et de fusions vésiculaires AG ,lysosomes ou mb plasmique.
Réf14:Pollard (T-D)

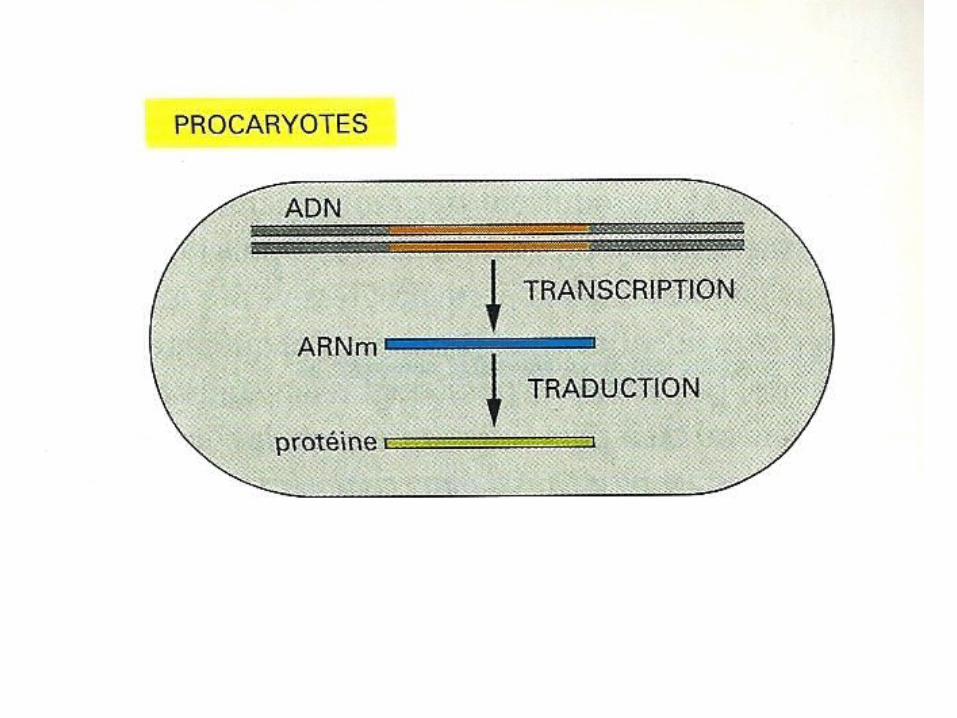
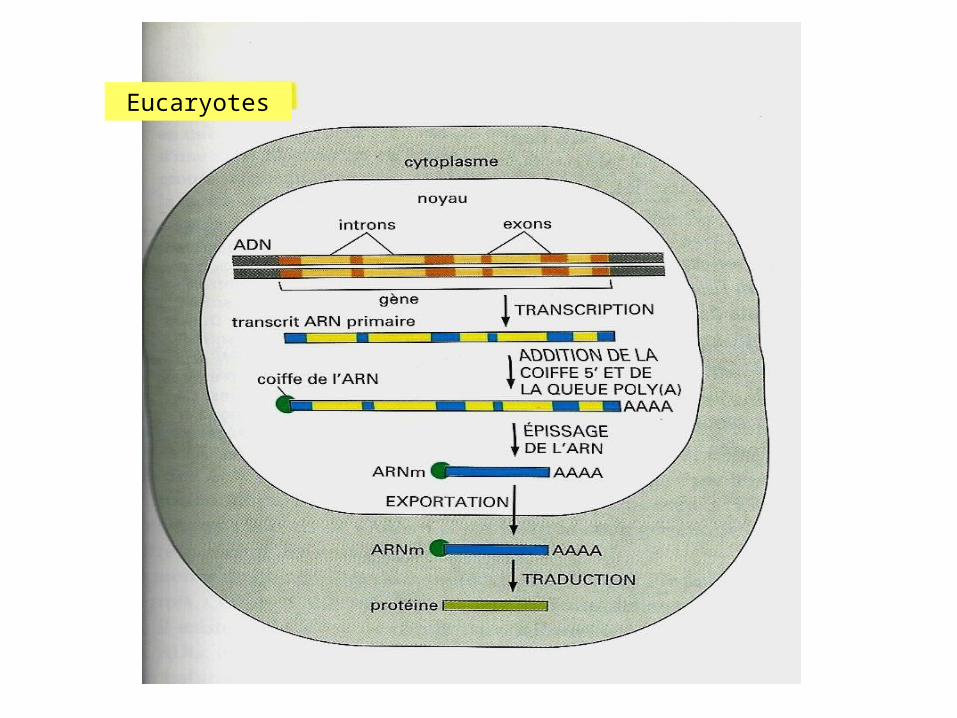
Eucaryotes

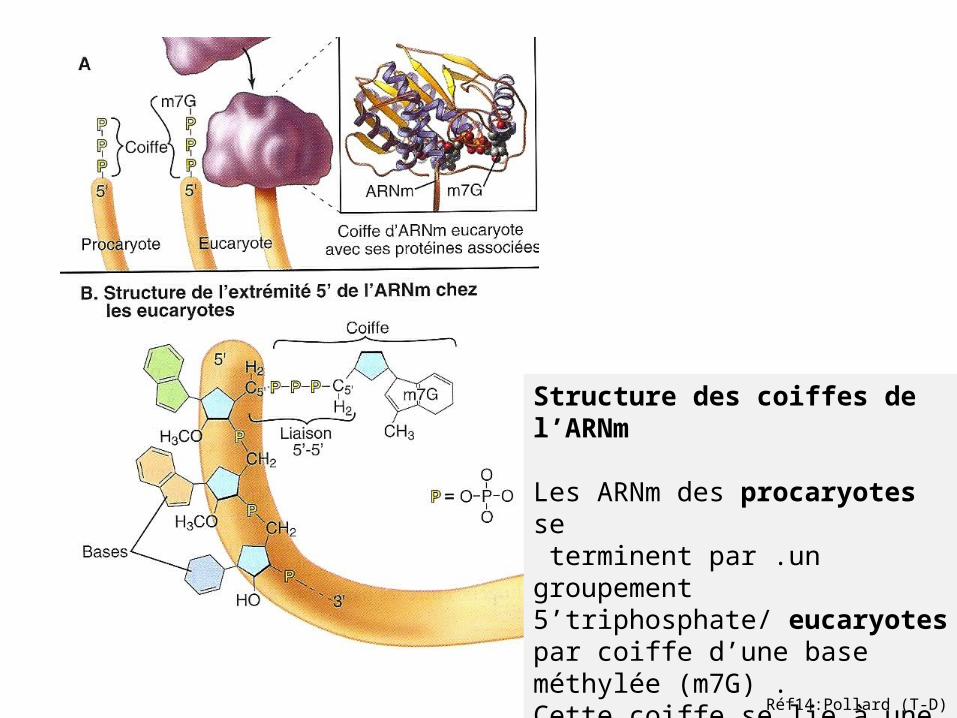
Structure des coiffes de l’ARNm Les ARNm des procaryotes se terminent par .un groupement 5’triphosphate/ eucaryotes par coiffe d’une base méthylée (m7G) .Cette coiffe se lie à une protéinequi protège la dégradation par les nucléases.
Réf14:Pollard (T-D)

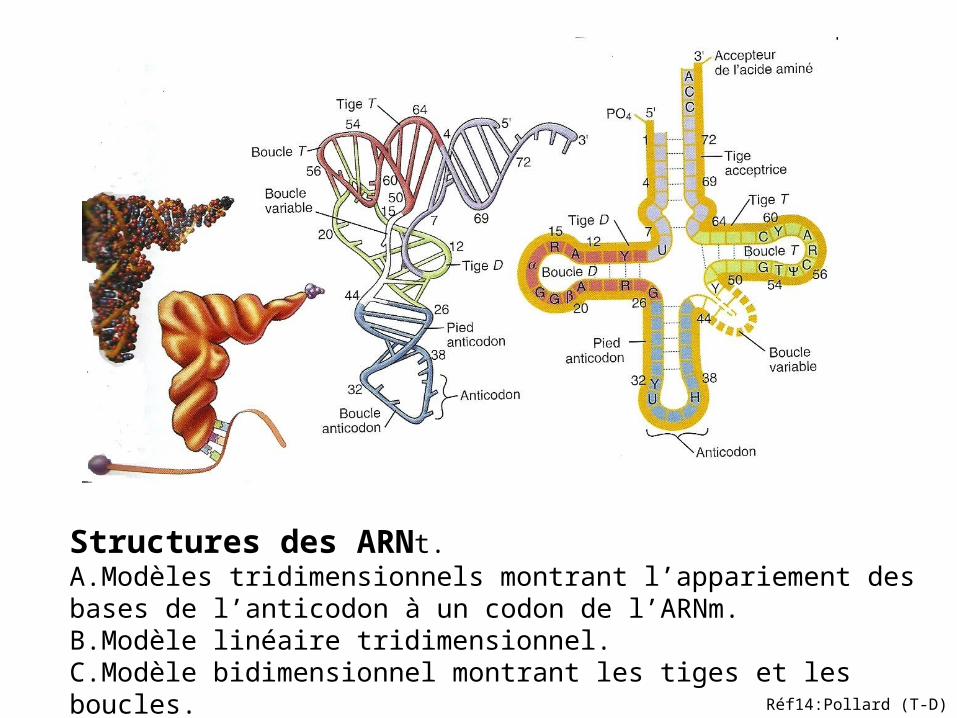
Structures des ARNt. A.Modèles tridimensionnels montrant l’appariement des bases de l’anticodon à un codon de l’ARNm.B.Modèle linéaire tridimensionnel.C.Modèle bidimensionnel montrant les tiges et les boucles.
Réf14:Pollard (T-D)
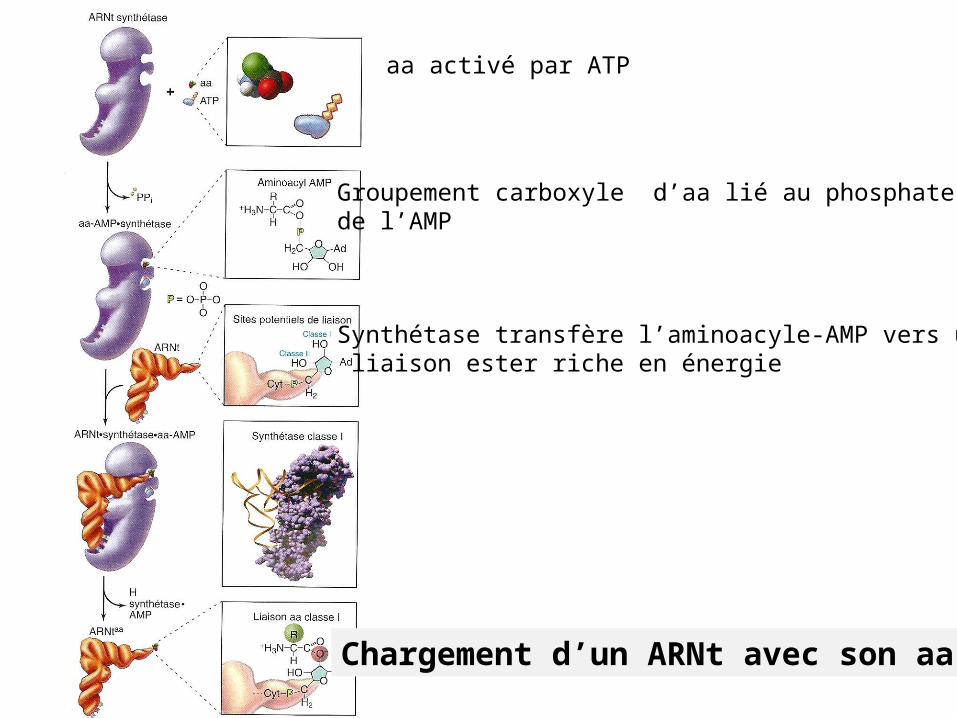
Chargement d’un ARNt avec son aa .
aa activé par ATP
Groupement carboxyle d’aa lié au phosphate αde l’AMP
Synthétase transfère l’aminoacyle-AMP vers une liaison ester riche en énergie

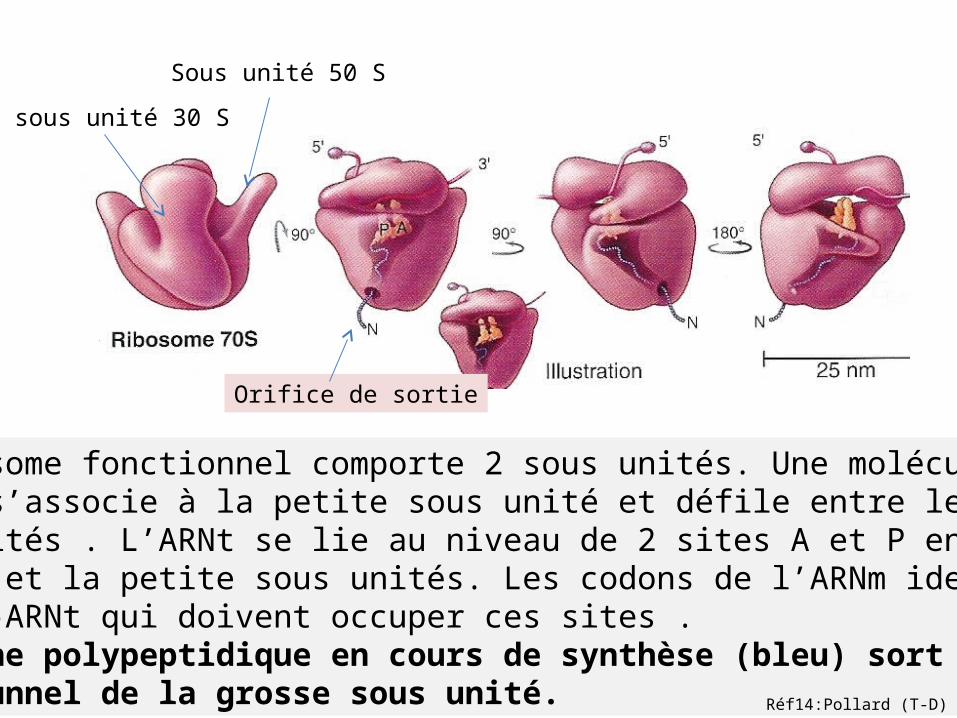
sous unité 30 S
Sous unité 50 S
Le ribosome fonctionnel comporte 2 sous unités. Une molécule d’ARNm s’associe à la petite sous unité et défile entre les 2 sous unités . L’ARNt se lie au niveau de 2 sites A et P entre la grosse et la petite sous unités. Les codons de l’ARNm identifient les aa-ARNt qui doivent occuper ces sites .La chaine polypeptidique en cours de synthèse (bleu) sort d’un tunnel de la grosse sous unité.
Orifice de sortie
Réf14:Pollard (T-D)

S: coefficient de sédimentation par ultracentrifugation. Réf14:Pollard (T-D)




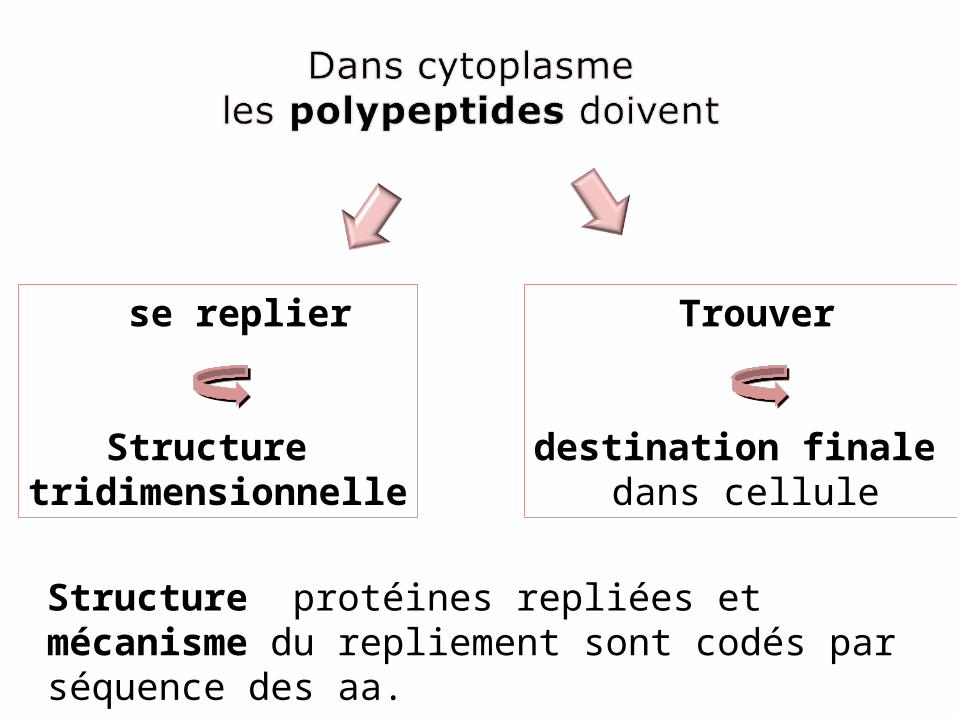
se replier
Structure tridimensionnelle
Trouver
destination finale dans cellule
Structure protéines repliées et mécanisme du repliement sont codés par séquence des aa.

In vitro, nombreuses petites protéines se replient rapidement.
Mais beaucoup de protéines nouvellement
synthétisés ne peuvent se replier spontanément
ont besoin d’aide pour surmonter: variations intermédiaires de repliement,
éviter leur dénaturation , leur agrégation ou la protéolyse.

Les chaperons
Facilitent le repliement Inhibent l’agrégation des protéines
ne libèrent polypeptides que si état de repliement favorable
2 classes de molécules :- HSP70 et leurs régulateurs
- Chaperonines (cylindriques)

Protéines nouvellement synthétisés
85°/°
se replient spontanément
ou avec assistance
des HSP70
15°/°
nécessitent un isolement au seindes chaperonines
pour se replier

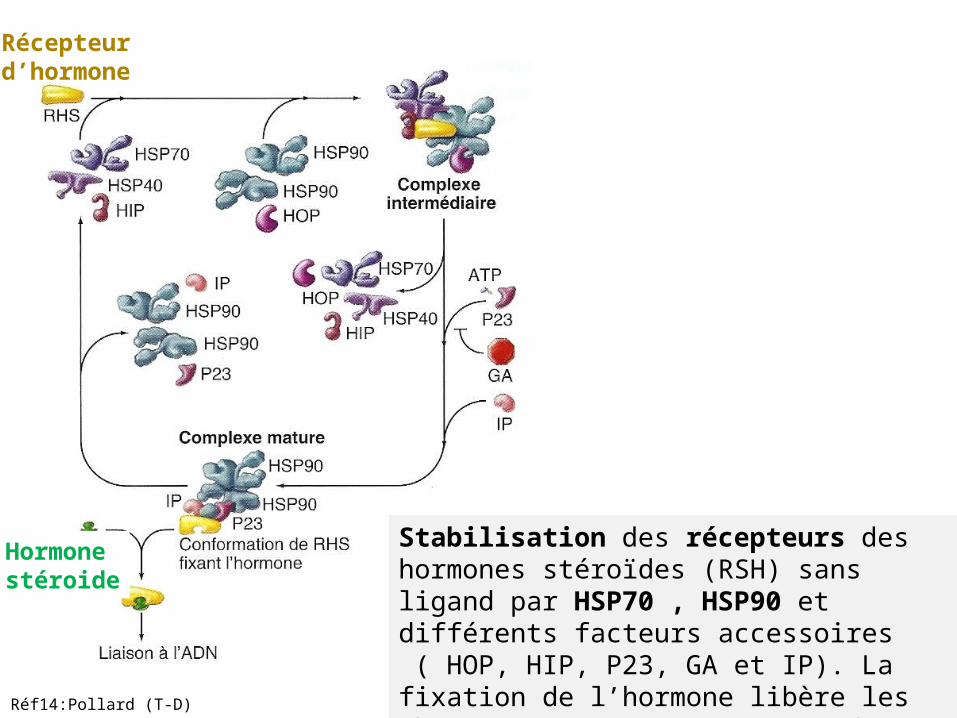
Stabilisation des récepteurs des hormones stéroïdes (RSH) sans ligand par HSP70 , HSP90 et différents facteurs accessoires ( HOP, HIP, P23, GA et IP). La fixation de l’hormone libère les chaperons et permet au RSH de migrer vers le noyau.
Récepteur d’hormone
Hormonestéroide
Réf14:Pollard (T-D)

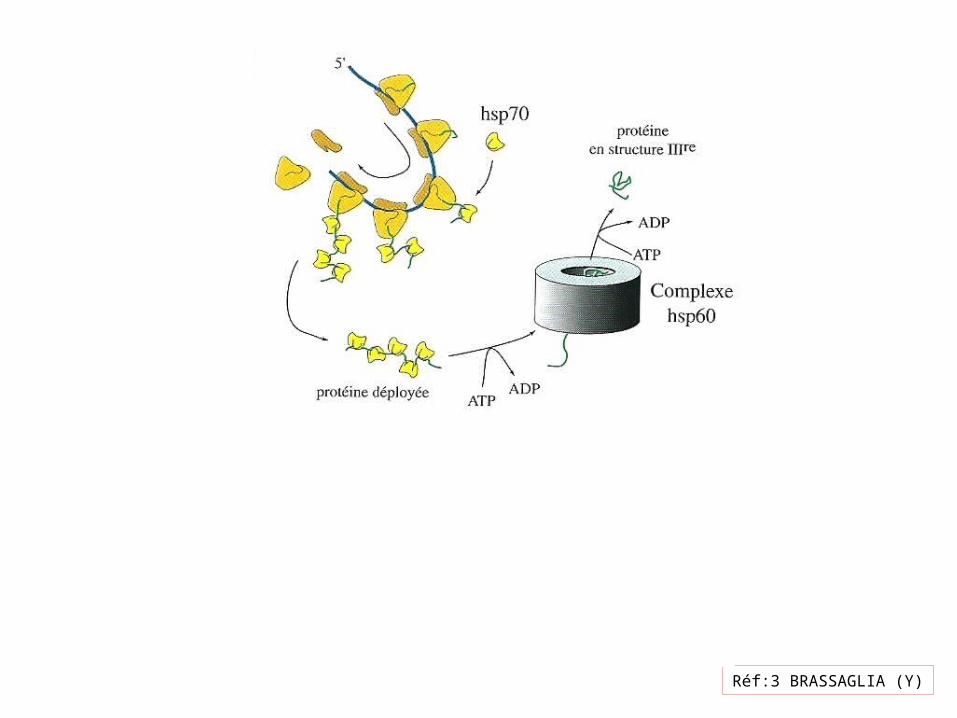
Réf:3 BRASSAGLIA (Y)
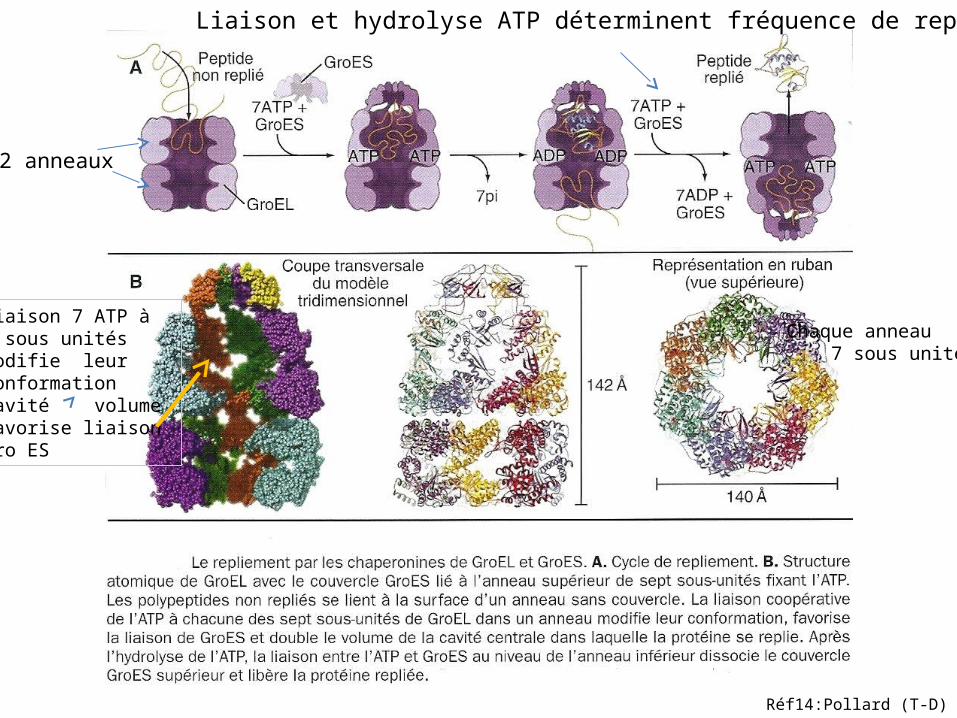
2 anneaux
Chaque anneau 7 sous unités
Liaison 7 ATP à 7 sous unités modifie leur ConformationCavité volumeFavorise liaison Gro ES
Liaison et hydrolyse ATP déterminent fréquence de repliement
Réf14:Pollard (T-D)

III- Echanges entre noyau / cytoplasme

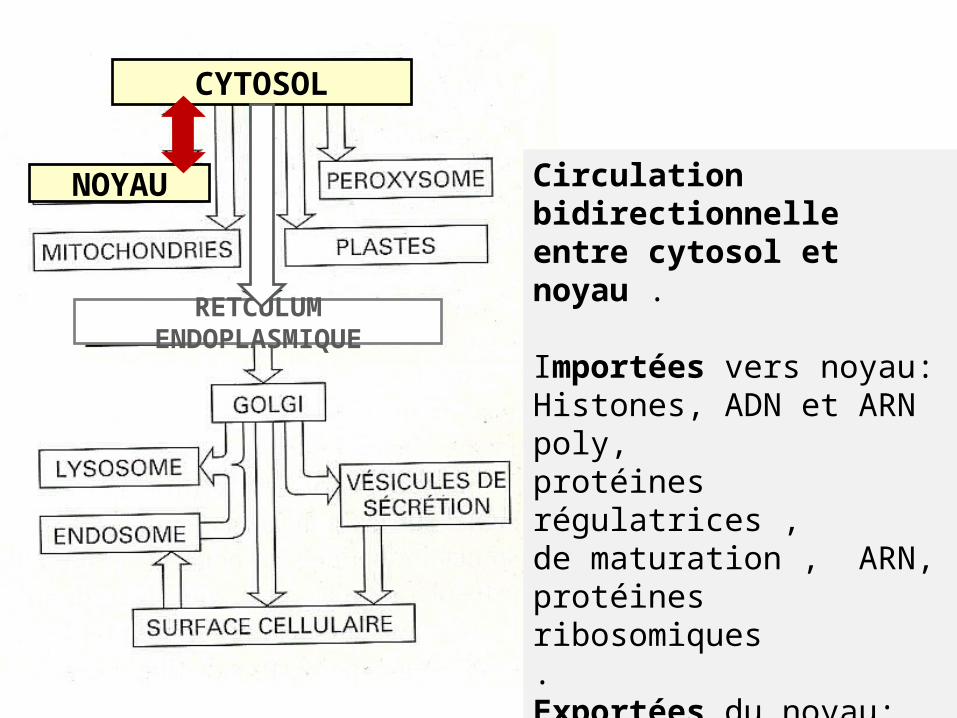
RETCULUM ENDOPLASMIQUE
CYTOSOL
NOYAU Circulation bidirectionnelleentre cytosol et noyau .
Importées vers noyau:Histones, ADN et ARN poly,protéines régulatrices ,de maturation , ARN, protéines ribosomiques.Exportées du noyau:ARNm , ARNt ,sous unités ribosomiques.

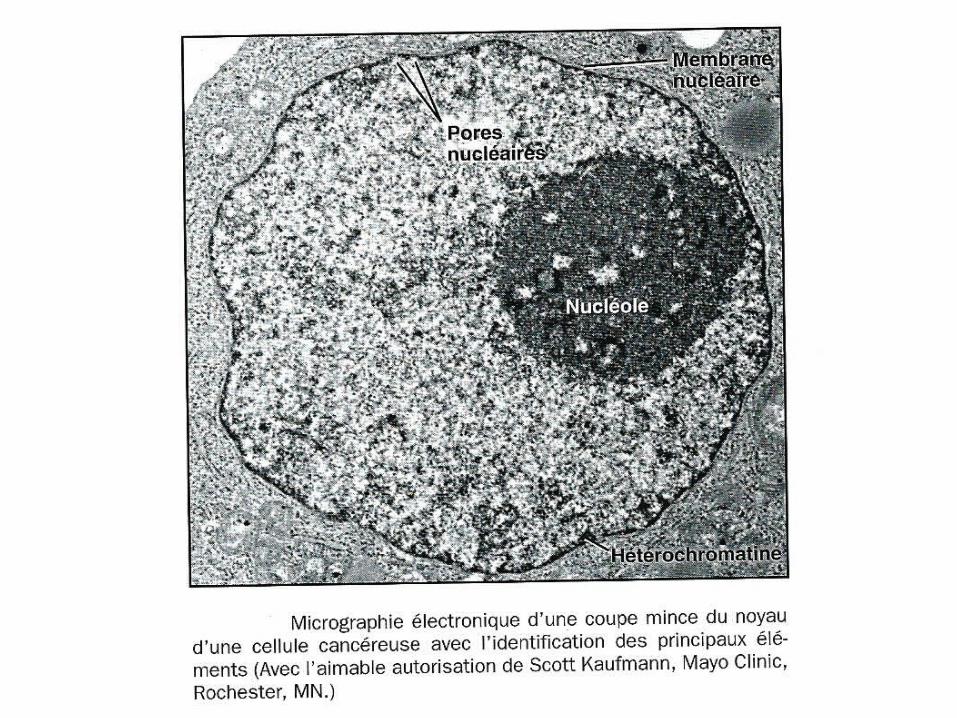
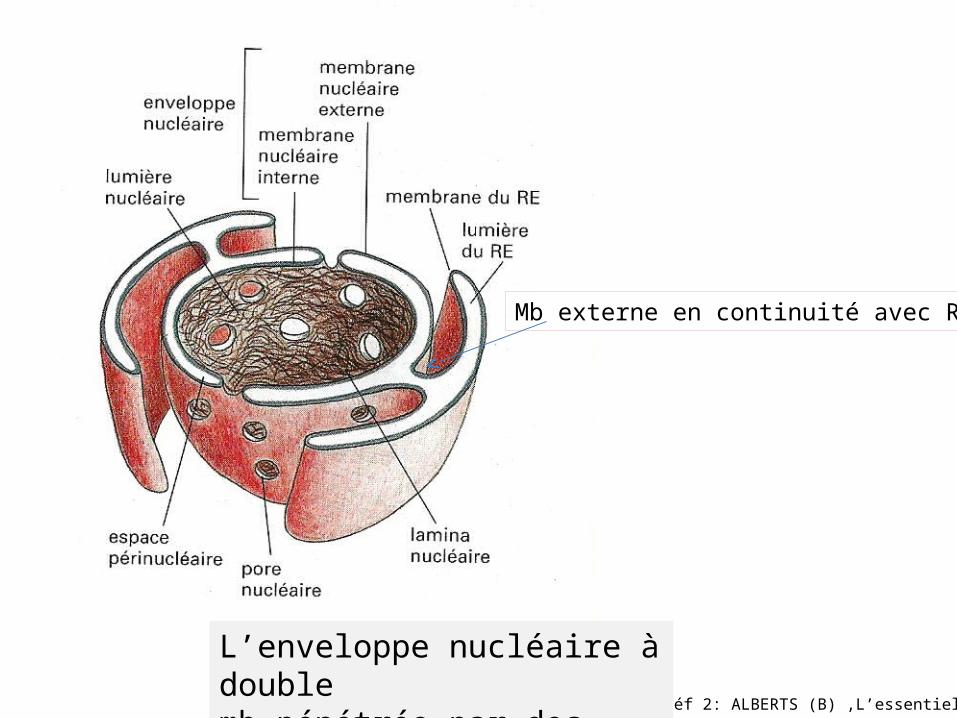
Réf 2: ALBERTS (B) ,L’essentiel
L’enveloppe nucléaire à doublemb pénétrée par des pores.
Mb externe en continuité avec RE

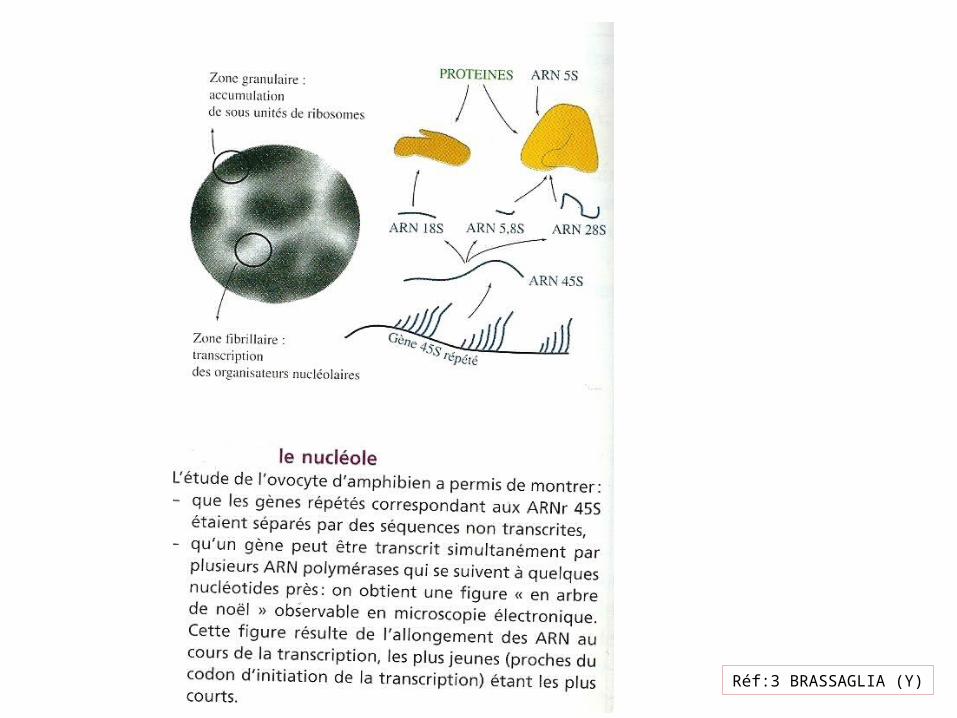
Réf:3 BRASSAGLIA (Y)

ARNr 16 S ARNr 18 S
Réf14:Pollard (T-D)
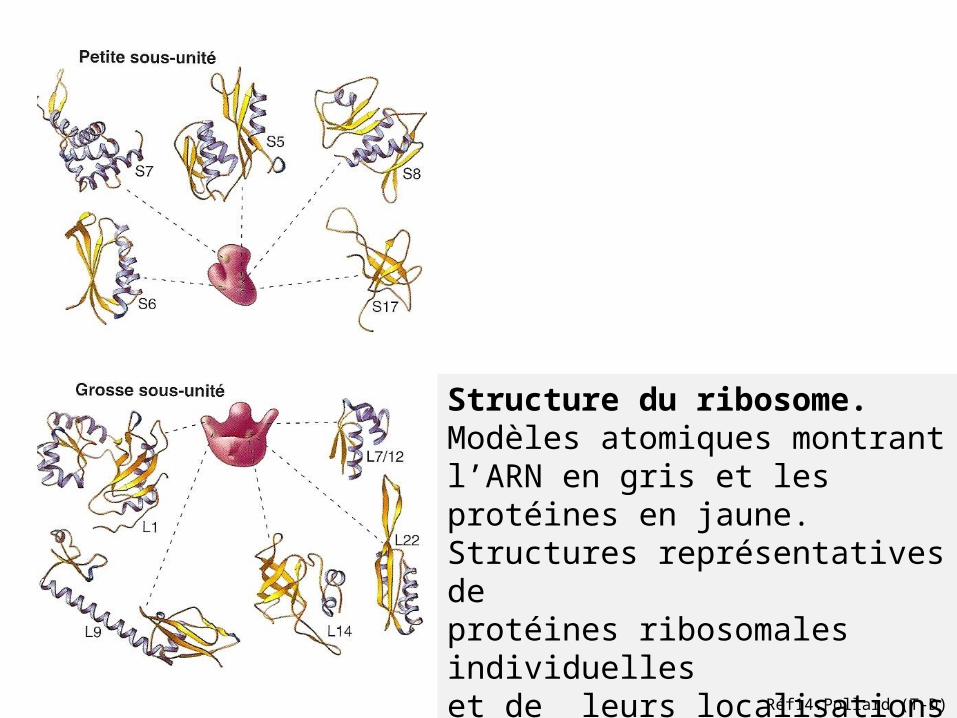
Structure du ribosome.Modèles atomiques montrant l’ARN en gris et les protéines en jaune.Structures représentatives de protéines ribosomales individuelleset de leurs localisations sur la petite et la grosse sous unité.
Réf14:Pollard (T-D)


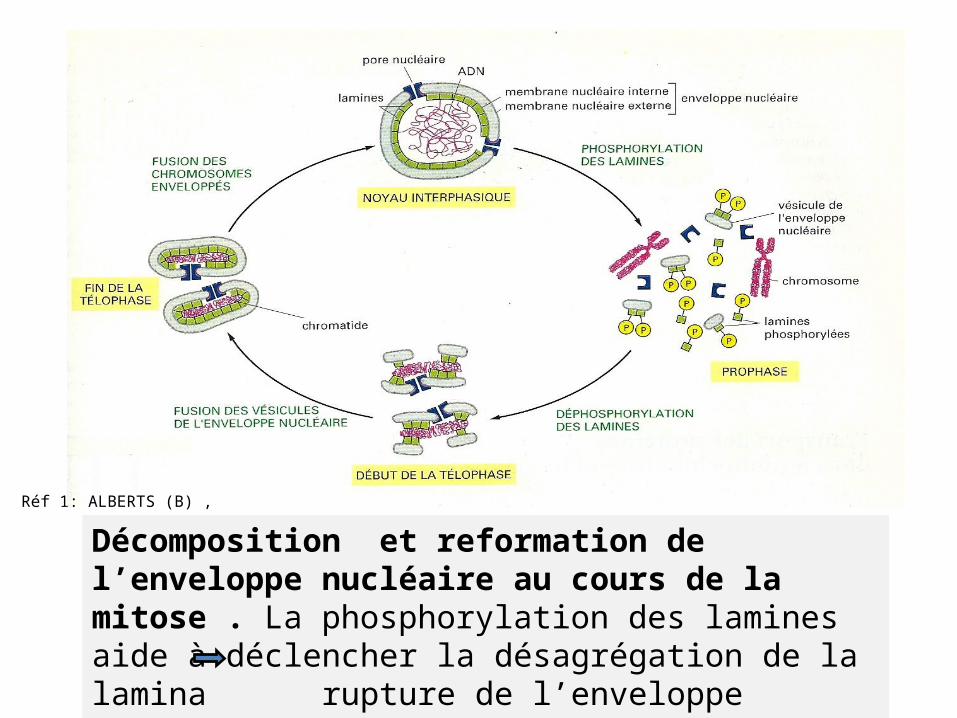
Réf 1: ALBERTS (B) ,
Décomposition et reformation de l’enveloppe nucléaire au cours de la mitose . La phosphorylation des lamines aide à déclencher la désagrégation de la lamina rupture de l’enveloppe nucléaire en vésicules La déphosphorylation : processus inverse.

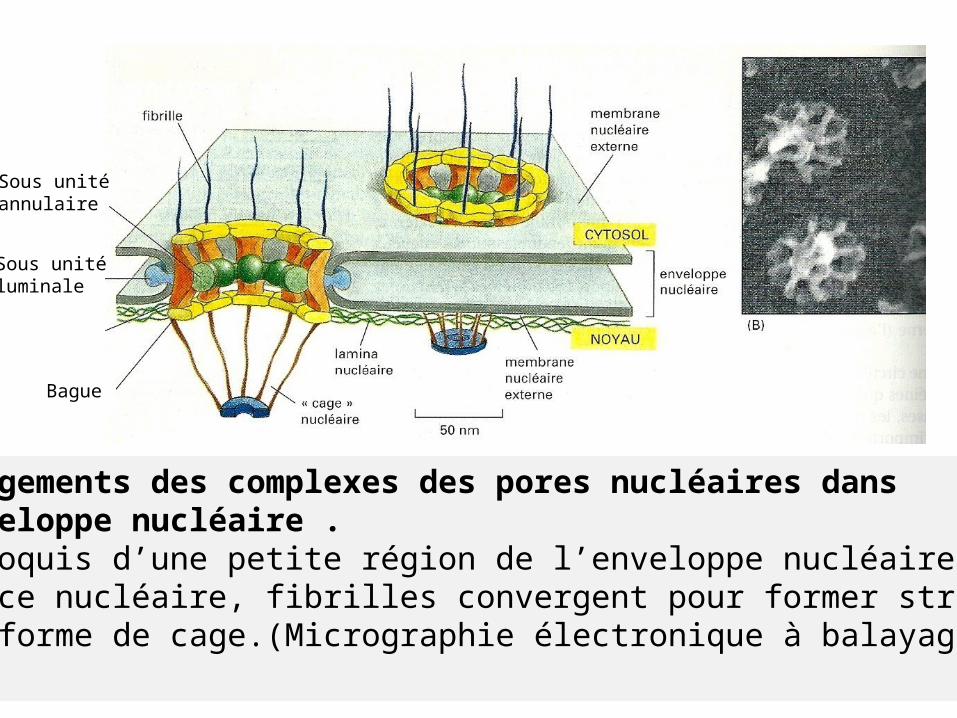
Arrangements des complexes des pores nucléaires dans l’enveloppe nucléaire .(A)Croquis d’une petite région de l’enveloppe nucléaire.(B)Face nucléaire, fibrilles convergent pour former structure en forme de cage.(Micrographie électronique à balayage.)
Sous unitéannulaire
Sous unité luminale
Bague
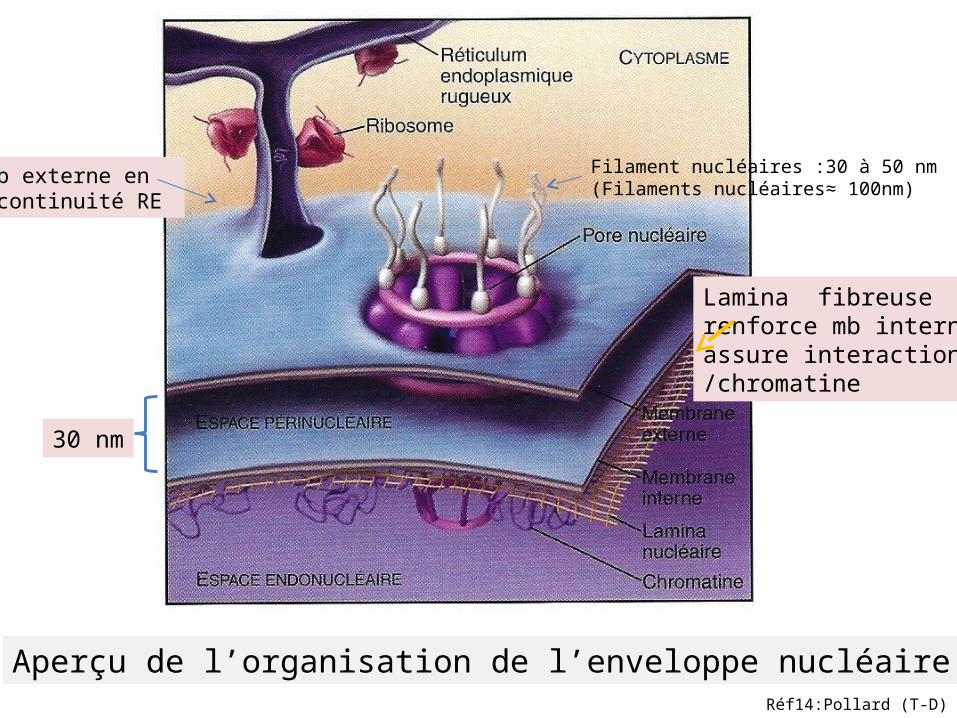
Aperçu de l’organisation de l’enveloppe nucléaire
Mb externe en continuité RE
30 nm
Lamina fibreuserenforce mb interne,assure interaction /chromatine
Filament nucléaires :30 à 50 nm(Filaments nucléaires≈ 100nm)
Réf14:Pollard (T-D)

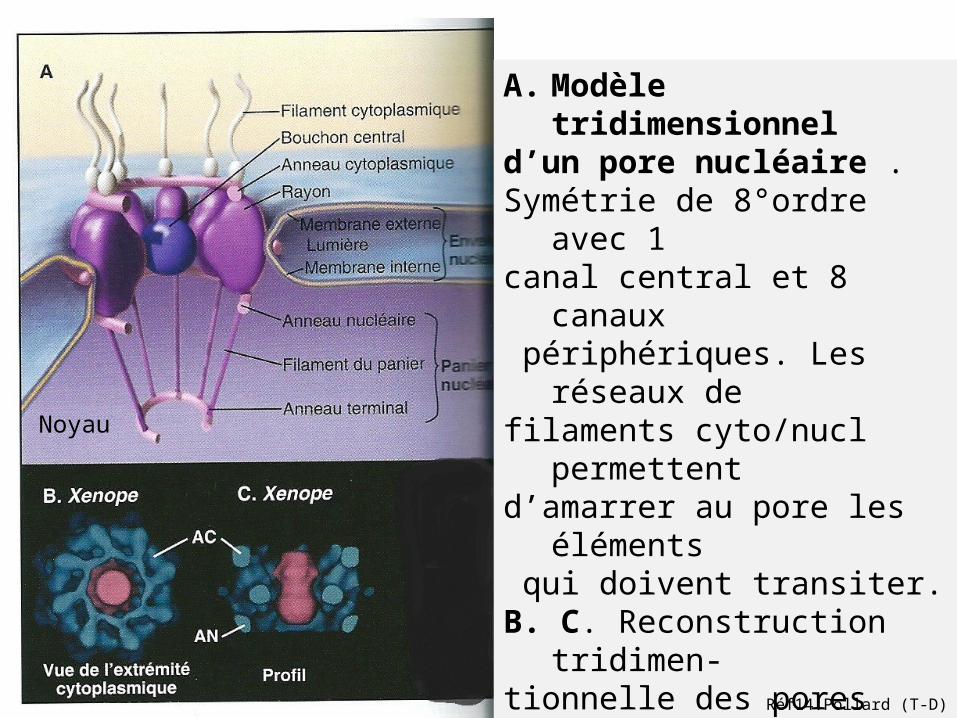
A. Modèle tridimensionnel d’un pore nucléaire .Symétrie de 8°ordre avec 1 canal central et 8 canaux périphériques. Les réseaux de filaments cyto/nucl permettent d’amarrer au pore les éléments qui doivent transiter.B. C. Reconstruction tridimen-tionnelle des pores nucléaires de grenouille Xenopus laevis.
AC: anneau cytoplasmiqueAN: anneau nucléaire
Noyau
Réf14:Pollard (T-D)



A. Micrographie électronique .Coupe mince d’ enveloppe nucléaire avec lamina et pores nucléaires.
B. Micrographie électronique à balayage de la surface interne de l’enveloppe nucléaire.
Panier nucléaire
Réf14:Pollard (T-D)

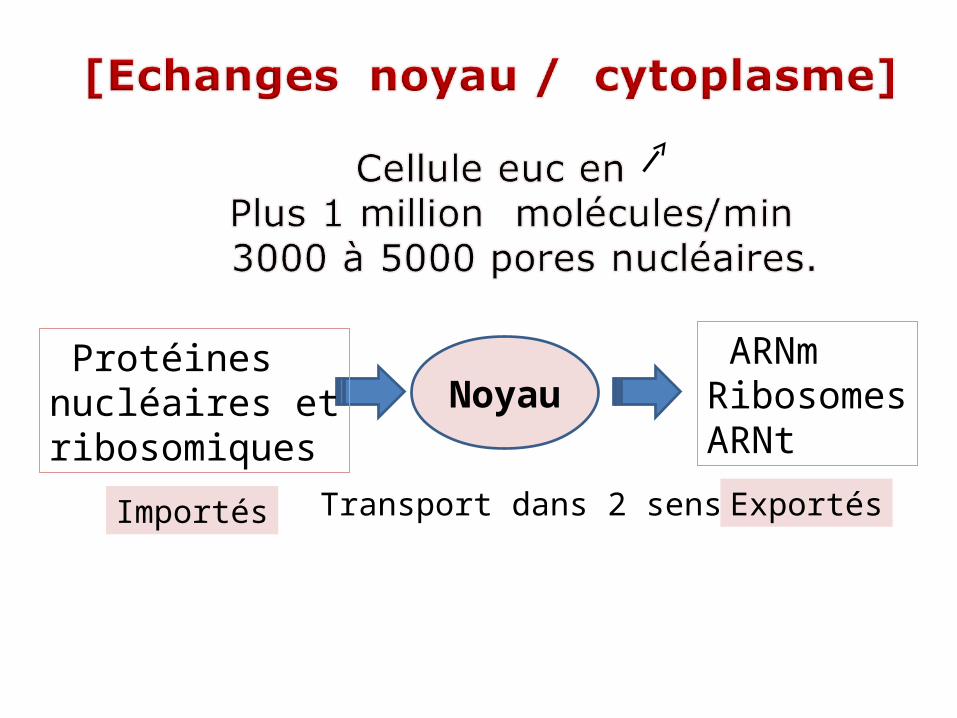
Noyau ARNmRibosomesARNt
Protéinesnucléaires etribosomiques
Transport dans 2 sensImportés Exportés

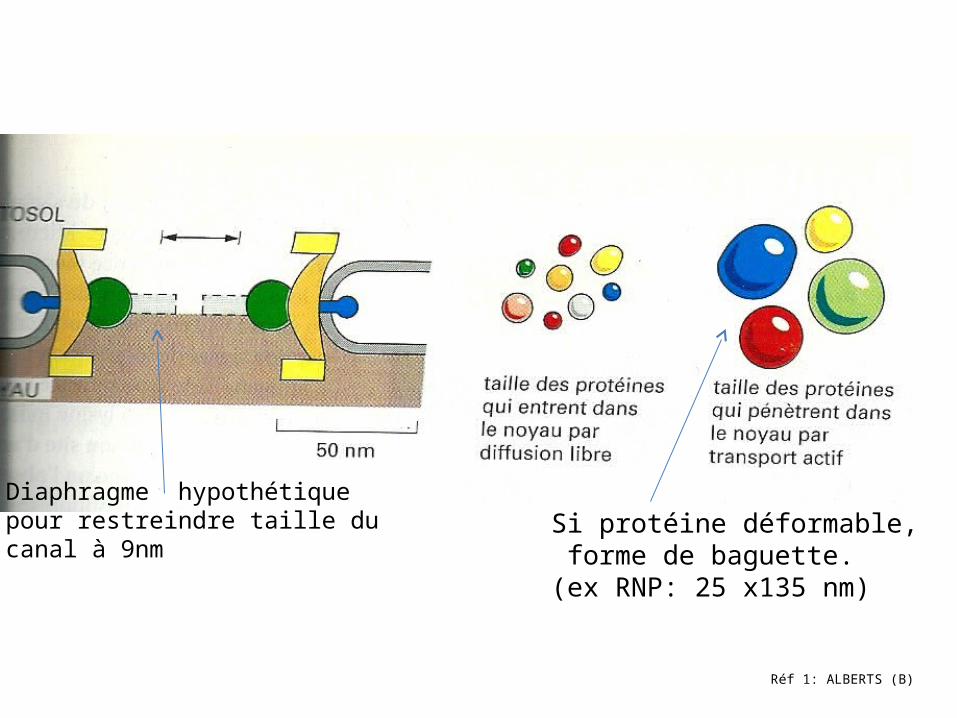
Réf 1: ALBERTS (B)
Diaphragme hypothétique pour restreindre taille du canal à 9nm
Si protéine déformable, forme de baguette. (ex RNP: 25 x135 nm)
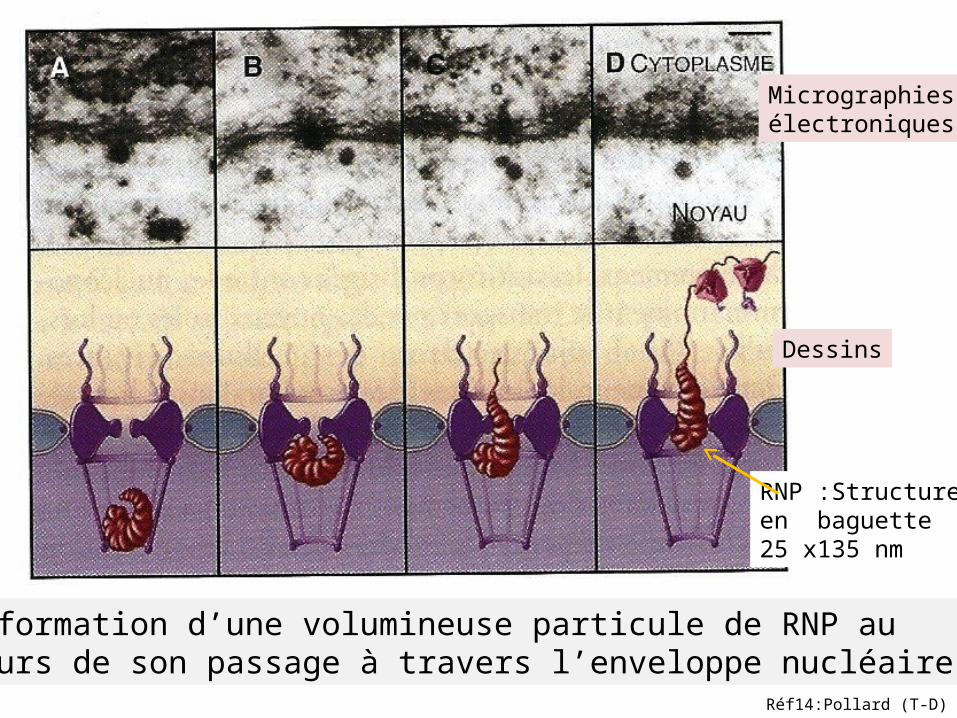
Micrographies électroniques
Dessins
Déformation d’une volumineuse particule de RNP au cours de son passage à travers l’enveloppe nucléaire.
RNP :Structureen baguette25 x135 nm
Réf14:Pollard (T-D)


0,1 µm
Particules d’or Petites maisséquestrées







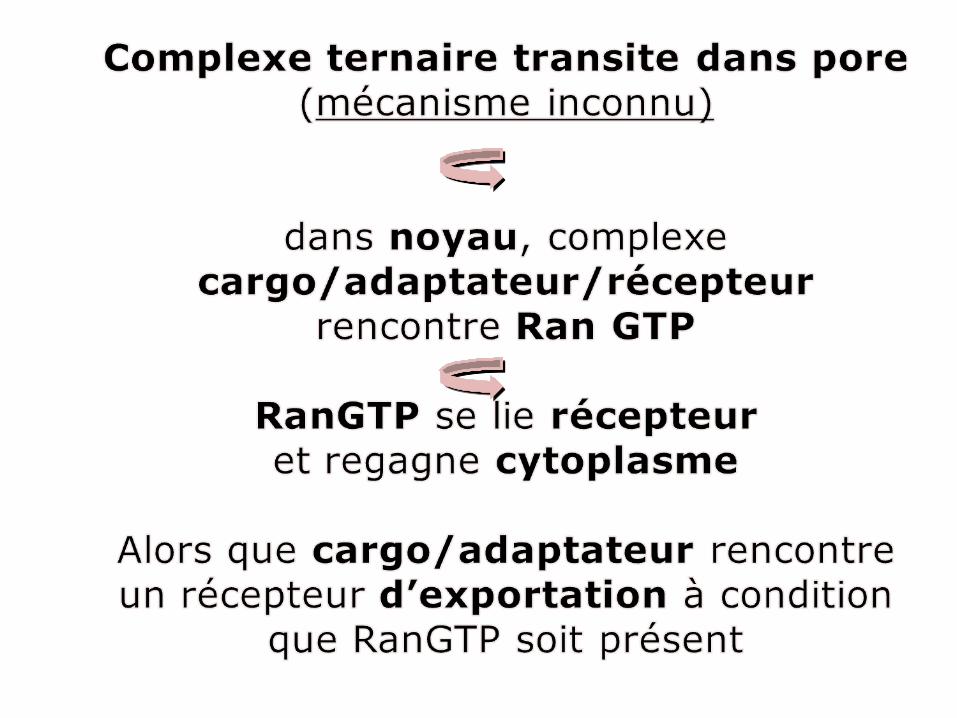
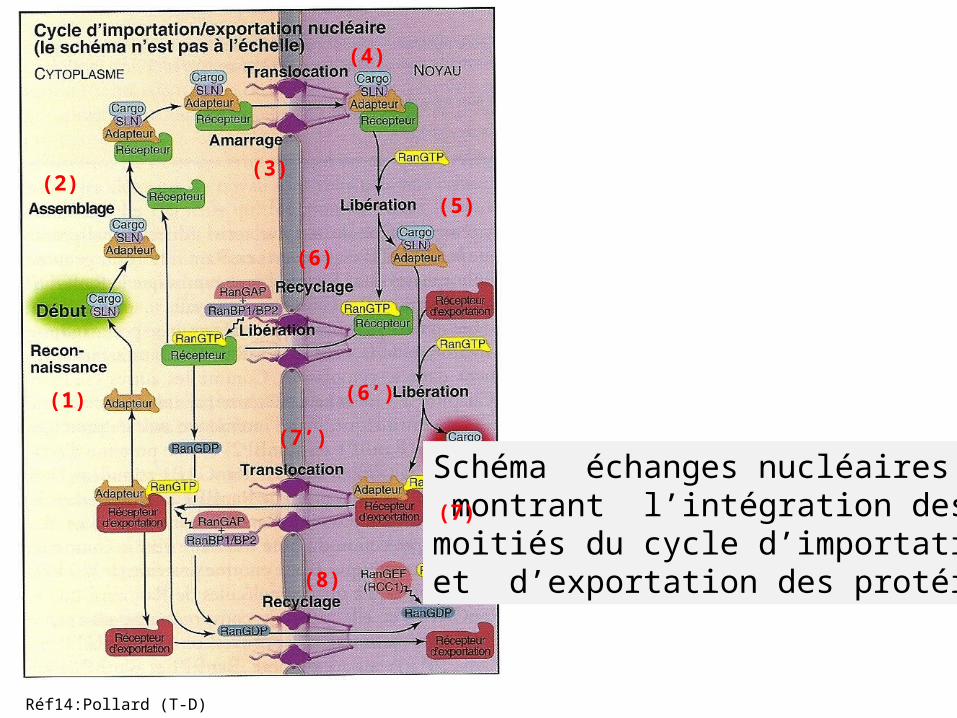
Schéma échanges nucléaires montrant l’intégration des 2 moitiés du cycle d’importation et d’exportation des protéines.
(1)
(5)(3)(2)
(4)
(6’)
(6)
(7)
(7’)
(8)
Réf14:Pollard (T-D)


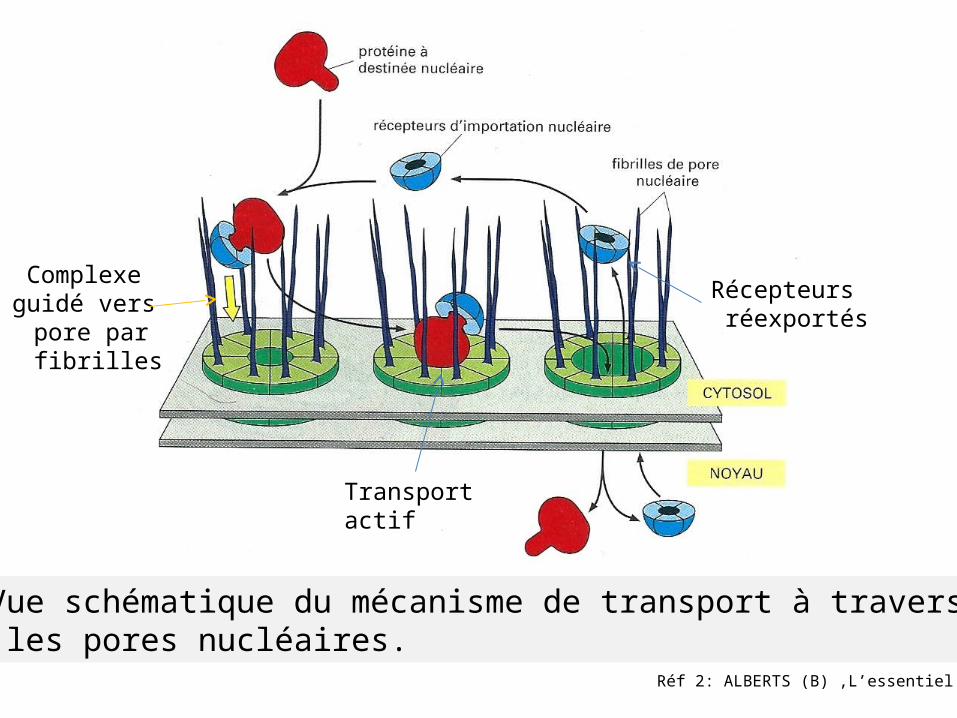
Réf 2: ALBERTS (B) ,L’essentiel
Vue schématique du mécanisme de transport à travers les pores nucléaires.
Complexe guidé vers
pore par fibrilles
Transport actif
Récepteurs réexportés

Réf 1: ALBERTS (B) ,
Représentation très schématique du mécanisme de transport actif à travers les pores nucléaires.
Fibrilles ne sont pas montrées