a mouse dna repair enzyme (apex nuclease) having exonuclease and apurinic/apyrimidinic endonuclease...
TRANSCRIPT

Biochit, lica et Biophysica Acta, 1079 (19ql) 57-64 © 1991 Elsevier Science Publishers B.V. 0167-4838/ql/$03.50 ADONIS 0167483891002536
BBAPRO 33980
A mouse DNA repair enzyme (APEX nuclease) having exonuclease and apurinic/apyrimidinic endonuclease activities:
purification and characterization
Shuji Seki, Shogo Ikeda *, Sekiko Watanabe, Masao Hatsushika, Ken Tsutsui, Kosuke Akiyama and Bo Zhang
Department of Molecular Biology. Institute o[ Cellular and Molecular Biolo~', Okayama Uniuersiry Medical School. Okayama ¢Japan)
(Received 18 March 1991)
57
Key words: DNA repair enzyme; Exonuclease: Apurinic/apyrimidinic endoauclease; DNA phosphatase; Amino acid sequc0ce; Priming enzyme
A mouse repair enzyme having priming activity on bleomycin-damaged DNA for DNA polymerase was purified to apparent homogeneity and characterized. The enzyme extracted from permeabilized mouse ascites sarcoma (SR-C3H/He) cells with 0.2 M potassium phosphate buffer (pH 7.5) was purified by successive chramatograpbies on phosphocellulose, DEAE-cellulose, phosphocellulose (a second time), ~phadex G.100, single.stranded DNA cellulose and hydroxyapatite. The purified enzyme has an M, of 34000 as determined by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Enzymatical studies indicated that it is a multifunctional enzyme having exonuclease, apurinic/apyrimidinic endonuclease and phosphatase activities, similar to Escher/ch/a co/i exonucle- ase !I!. This enzyme is tentatively designated as APEX nuclease for apurinic/apyrimidinic endonuclease and exonuclease activities. The amino acid composition, amino-terminal amino acid sequence and an internal amino acid sequence of APEX nuclease are determined.
Introduction
The DNA damage that occurs daily as a conse- quence of spontaneous hydrolysis under physiological conditions or by chemical, physical or biological stresses is substantial and of various types [I]. Among the types of damage, apurinic/apyrimidinic (AP} sites are gener- ated at several thousand residues per genome per day in a mammalian cell [2]. Single-strand DNA breaks with 3' termini blocked I%' nucleotide fragments are also known to be frequently induced by free radical pathways initiated by ionizing radiation, bleomycin and other sources of oxygen radicals [1,3-6]. In our studies on DNA repair synthesis after bleomycin-induced dam-
* Present address: Department of Bu~logical Chcmi,try, Facully ¢~[ Science. Okayama Univer~a~ of Science. Okayama 7(10. ~'apan.
Correspondence: S. Seki. Department of Molecular BioloK,. Insti- tute of Cellular and Molecular Biolog'y, Oka~ama Universily Medical School. 2-5-1. Shikata-cho. Okayama 711~). Japan
age in mammalian cells, we found and partially puri- fied a priming enzyme (an exonuclease) that can initi- ate repair of bleomycin-induced and X-ray-induced single-strand breaks [7-11]. This enzyme also showed priming activiD' for DNA polymerase on acid-de- purinated DNA and micrococcal nuclease-treated DNA, .suggesting that it is a multifunctional enD'rae having exonuclease. AP endonuclease and phosphatase act vities [9.12]. Thus this enzsme exhibits properties sir dlar to those of E. coli ,:xonuclea~ !I1 and yeast DNA 3'-repair diesterase [IA3,1a]. Mammalian AP endonucleases possibly involved in the repair of AP sites have been purified from various species including mice [15-171. rats [181, cattle [t9] and humans [20-221. As far as we know, however, none of the previously reported mammalian AP endonucleases possessed ex- onuclease actMty.
In the present paper we describe the purification, characterization and partial amino acid sequences of the mouse repair enzyme (designated as APEX nuele- ase) having exonuclease and AP endonuclease activi- ties.

58
Materials and Methods
Materials The reagents used in these experiments were ob-
tained from the following sources: [~H]dTTP from Amersham Japan, Tokyo, Japan: [a~'P]phos~hate and [a-3-'P]dCTP from ICN Biochemicals, CA, U.S.A.; ribonucleotides {NTPs) and deoxyribonuclcotides (dNTPs) from Seikagakv. Kngyou, Tokyo, Jap,m: E. colt exonuclease Ill, DNA polymerase I and the large fragment (Klenow polymcrase) of DNA polymerase I from Takara Shuzo. Kyoto, Japan; calf thymus DNA and DNA alternate copolymer poly(dA-dT)-poly(dA- dT) from Pharmacia, Uppsala, Sweden. The other reagents used were obtained from the previously de- scribed sources [Q]. Mouse ascites sarcoma [SR- C3H/He) cells were obtained and maintained as de- scribed previously [23]. pUC18 DNA was prepared as described previously [2,1]. Bleomycin-Fc(ll)-trcated ¢calf thymus and pUCI8) DNAs and X-irradiated DNA were preparcd as described previously [9-11].
Preparation of acid-depurinated DNA and assay for AP endonuclease actie i.,y
The superhelical (form 1) pUCIS DNA and calf thymus DNA were depurinated by i~lcubating them in 3 vol. of 50 mM sodium citrate (pH 3.5) at 60°C for 15 rain essentially as described by Niwa and Moses [9.25]. After the incubation, the mixture was chilled to 0°C and dialysed against 50 mM Tris-HCI (pH 7.5) for 3 h and t,en against distilled water overnight. The dialysed DNA solution was used to measure AP endonucleasc activity. The acid treatment produced approximately six alkali-sensitive sites per pUCI8 DNA molecule, as determined by alkaline gel eleetrophoresis [26]. The assay mixture (15 tal final vol.) co:retained 0.25 ~,g (0.14 pmol) acid-depurinated pUCI8 DNA and 0-7.5 ~tl of an appropriate dilution of the enzyme in Triton-buffer B (0.0175% Triton X-ID0. 0.25 M sucrose, 10 mM Tris-HCl, 4 mM MgCI:, [ mM EDTA and 6 mM 2-mereaptoethano[, pH 8JI adjusted at 25°C1. After incubating the assay raixture at 37°C for 20 rain, the reaction was stopped by chilling it to 0°C and then adding 3 tal of 6-fold-concentrated gel loading buffer (0.25% bromphenol blue, 0.25% xylenc cyanol and 30% glycerol in H20). The mixture was loaded into a slot of a submerged 0.8% agarose gel. Electrophoretic analyses of conformation of pUC18 DNA were con- ducted as described previously [10,11,24].
Assay of the enzyme that enhances the template-primer actit'i~" of bleomycin-damaged, X-irradiated or acid.de- purinated DNA for DNA polymemse 13 or Klenow poly- merase
In the present paper the term "template-primer ac- tivity' of DNA is used to indicate the ability to ser~c as
a template-primer for DNA polymerase /~ or Klenow polymerase. The term 'priming activity (or enzyme)" on bteomycin-damaged, X-irradiated or acid-depurinated DNA is used to denote the activity (or enzyme) that enhances the template-primer activity of bleomycin- damaged, X-irradiated or acid-dcpurinated DNA by removing the 3' tags or by introducing single-strand breaks. The priming activity was measured by the two- step- {priming and repair DNA synthesis step-} method [U, ll]. For measuring the priming activity on bleomycin-damaged DNA, for example, 6 tag calf thy- mus DNA was incubated at 37°C for 30 min with 20 #M Fe(ll), 0.2 ~.g bleomycin and an appropriate an~ount of a priming enzyme fraction in a 39-/~I reac- tion mixture made up with Tritor,-buffer B. After the incubation, the reaction mixture was incubated at 60°C fo~ I0 min to inactivate enzymes in the priming enzyme fraction and then chilled to 0°C. A 21-~1 aliquot of the substrate mixture (labeled substrate: [~H]dTTP) for DNA synthesis supplemented with 0.04 unit of Klenow polymerase or DNA polymerase /3 was added to the reaction mixture, DNA synthesis carried out at 37°C for 30 rain. The ractioactivity incorporated into acid-in- soluble materials was mcasured by a disc method [91. One unit of pri,ning enzyme in the present assay is defined as the amount of the enzyme causing 1 nmol of [3H]dTMP incorporation per 30 rain into bleomyein- damaged DNA under the standard assay conditions.
Assay of DNA double-stranded exonuclease and phos- phatase acticitws
Radioactive DNA for exonuelease assays was pre- pared by growing Escherichia colt HBI01 in Luria- Bertani medium (125 ml) supplemented with [3H] thvmidine (i m C i / l ~ ml, 50 Ci/mmol). When the cclls reached the late log phase, they were harvested and their DNA was purified by standard phenol extrac- tion techniques. The radioactivit~ of the labeled DNA was 744 cpm/nmol of nucleotide. Bleomycin-, micro- coccal nuclcase- or DNase l-treatment of the labeled DNA was conducted as described previously [9,27]. Exonuclease activity of the priming enzyme was mea- sured using the bleomycin-, micrococcal nuclease- or DNasc l-treated, ~H-labeled DNA essentially as de- scribed [9]. To demonstrate 3'-5' exonuclease activity, the alternate copolymer poly(dA-dT) - poly(dA-dT) was 3'-end-labeled by incubating it with [3H]d'i~P and Klenow polymerase. The purified, 3'-end-labeled copolymers were incubated at 37°C for varying times with the priming enzyme in Triton-buffer B. Nu- cleotides released by exonucleolytic digestion were an- alyzed by polyethylene imine-cclluiose thin layer chro- matography [28]. To measure DNA 3' phosphatase activity, 3'[3ZP]phosphoryl-terminated DNA (specific activity: 8.9.104 cpm/nmol of phosphate) was pre- pared by partially digesting ~:P-labeled E. colt DNA
5O
with micrococcal nucleasc [29,30]. The priming enzyme was incubated at 37°C with the 3'-[~:P]phosphor3l - terminated DNA and released [':P]inorganic phos- phate (acid-soluble. Norit-non-adsorbable ":P) ~as measured as dcscribcd pre~4ously [31)].
Detennhmtion of partial amim~ acid sequences o[ the enzyme
When the purified priming enzyme 1,APEX nucle- ase) was treated at 37°C for 12.5 h with 70(~ formic acid. which hydrolyzes peptidc bonds between aspar- tate and proline, the enzyme was cleaved into two fragments with molecular masses of 30 and 4 kDa. ]'he amino-terminal amino acid sequence of the en~'me and that of the large formic acid fragment purified by SDS-polyaerylamide gel elcctrophoresis (SDS-PAGE) were determined by an Applied Biosystems Model 477A automated protein sequencer.
Other methods DNA polymerase/3 was purified [9,31] and assayed
[9] as described previously. Agmose gel electrophoresis and SDS-PAGE were performed as described previ- ously [10-12,32]. Repair of pUCI8 DN,'~ with bleo- mycin-, peplomycin- or X-ray-induced single-strand breaks was measured by monitoring conformational changes of the plasmid DNA [10,11,26]. Protein con- centrations were determined by the BCA Protein As- say (Pierce, IIfinois. U.S.A), using bovine serum albu- min as the standard. The amino acid composition of the enzyme was determined using an Applied Biosys- terns Derivatizer. Model 420A.
Results
Purification of a mouse repair enzj'me for bleomvcin- damaged DNA
Partial purification of a repair cnz,yme having ex- onucleolytic activity on bleomycin-damaged DNA and thereby providing priming sites for DNA polymerase was reported previously [9]. In the present stud}', the enzyme was further purified to apparent homogeneity. The purification was followed by measuring its DNA polymerase /3 prim!rig activity on bleomycin-damaged DNA. The enzyme was extracted from permeable mouse ascites sarcoma cells with 0.2 M potassium phosphate (KPi) buffer (pH 7.5) [9]. After adjustment of the KPi concentration to 0.1 M, the extract (fraction Nil was mixed with packed phosphtx;ellulose equili- brated with 0.1 M KPi. Enzymes bound to phospho- cellulose were eluted with (;.3 M KPi. The eluant (fraction N 2) was passed through DEAE-celluk)se mainly to remove nuelcic acid as described previously 19,31]. Material (fraction N3) not retained by lhe DEAE-cellulose column was loaded onto tne ~cond phosphocellulose column equilibrated with 0.15 M KPi
J ~
, 3d.
10 [- J
0 13 30 40 50 60 70 FIq'ACT ION
8 MW 44 46 48 50 52 54 56 58 60
k D Q
2 9 ~ ~ ~ ~ - - ~ ~ -
2/.. ~. ~.- ~: . . . .
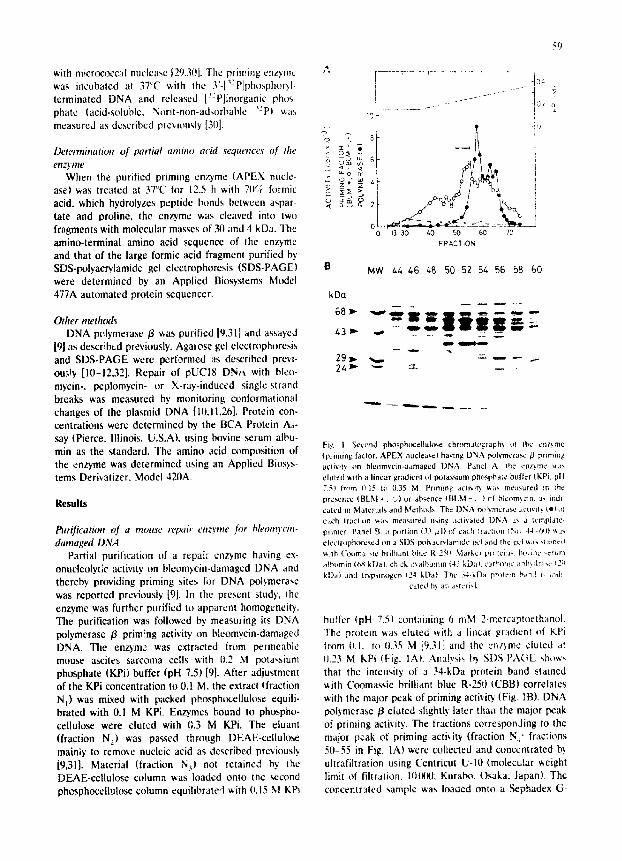
Fig I Second phosphocellukvse chromatogtzphs ot {he enzymc (priming IacIoL APEX n u c l e a r ) having DNA polvmerasc/~ primin~ actKily on bleomycin-dama~ed DNA. Panel A: the enzyme ,aas eluted ~tlh a linear gri~ienl eft potassium phosphate buffer (KPi. pit 75~ fmra IF 15 to 035 M. Priming acti',.t b v,a,, measured m the presence ( B t . M + . ~1 or absence (BLM - . .) cl bteom,F:m. :J- indl cared in Malerials and Methods The DNA ~x~L'~n3crase ~cttvtb ~1+ ot t:ach fraclion ",,,,as measured n,,ing aCli','~tcd DNA as a t,~mplale primer. Panel B: a pq~rti~m (33 ,all of eac,~ Waction (";(~ 44 6qi '.',a'., ei¢c~[,,lphorescd on a SI)S-Ix~t~a~.ryt-~mide gel and d~e gt:l ~as .,laJncd
~ l l h ( ' t~ma ' ~e brilhant blue R-25it M,Jtkcl pr<~tcin, b ~ i n c ~erur:, albumin (6S k D O chick ,~'.alhumm (43 kDa). t i the, nit ar~h,dra~c ~2 ~
kt)a) .and t~psmtrgen t24 kD:.O The 34-kDa prolcm b,md i, iadi cated b'. an az, lcri ,k
buffer (pH 7.5) containing 6 mM 2-mercaptoethanol. The protein was eluted with a linear gradient of KPi from 0.1. to I).35 M [9,31] and the enl}me cluted at (I.23 M kPi (Fig. lA). Analysis b} SDS-FA(;L show~ that the intensity of a 34-kDa protein band stained with Coomassie brilliant blue R-250 (CBB) correlates with the major peak of priming aetivi~ (Fig. IB). DNA polymerase/3 eluted slightly later than the major peak of priming activity. The fractions corresponJing to the major peak of priming activity (fraction N~' fractions 50-55 in Fig. lA) were collected and concentrated b.~ uhrafiltration using Centricut U-10 (molecular weight limit of filtration. 100011; Kurabo, Osaka. Japan). The concentrated sample was loaded onto a Sephadex G-
60
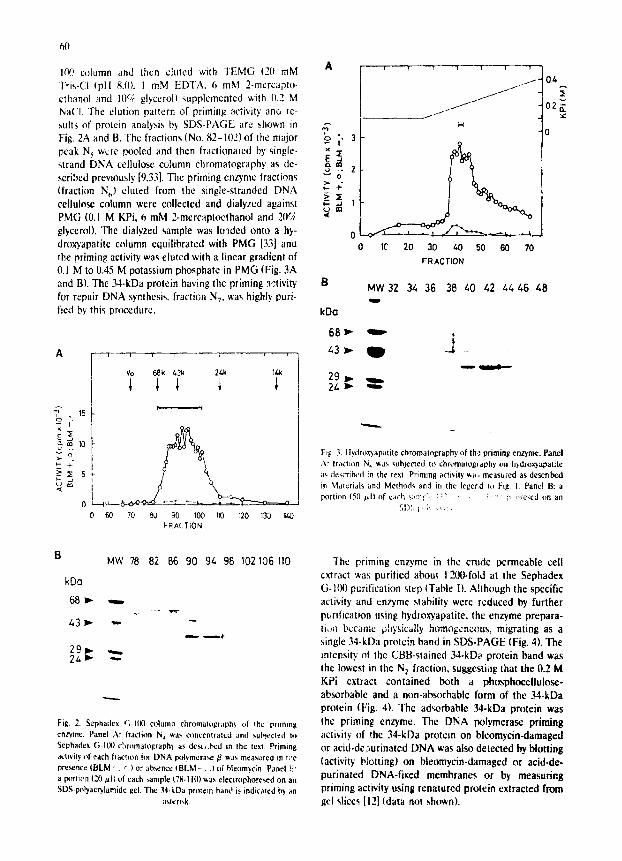
I(~) column and thcn cluted with TEMG (20 mM Tris-Cl (pH 8.01. I mM EDTA, 6 mM 2-mcrcapto- ethanol and 10% glycerol) supplemented with 11.2 M NaCI. The elution pattern of priming activity ano re- sults of protcin analysis by SDS-PAGE are shown in Fig. 2A and B. The fractions (No. 82-1021 of the major peak N 5 w,:re pooled and then fractionatcd by single- strand DNA cellulose column chromatography as de- scribed previously [9.33]. The priming enzyme fractions (fraction N~,) eluted from the single-stranded DNA cellulose column were collected and dialyzed against PMG (0.1 M KPi, 6 mM 2-mercaptocthanol and 2115; glycerol). The dialyzed sample was Io'~ded onto a hy- dmxyapatite column equilibrated with PMG [33] ann the priming activity was eluted with a linear gradient of 0.1 M to 0.45 M potassium phosphate in PMG (Fig. 3A and B). The 34-kDa protein having the priming activity for repair DNA synthesis, fraction NT, was highly puri- fied by this procedure.
';';" 15
~ 0
o
i i [ - ~ 1 t i
vo 6Ek 43k 24k 14k
60 70 80 90 100 ~tO ~20 133 140 FRACTION
MW 78
kDa
68 ~
43 ~
29~. .,..,. 2/.1~ -,,.,
02 86 90 9/. 98 102 106 110
Fig. 2. Sephadex G I l l 0 ct)luma chromatog!;!ph~ of the prtmmg
en~me. Panel A: fraction N 4 wa- c~mcentratcd and "~uhtecled to
Sephadcx G-1U0 c~aromatoglaph~ as d¢,,t,.bcd in the text. Priming
aclwity at each Iraction ~t)r DNA Ix~tymera~/3 was measured in t:~e
pr¢~nce (BLM -' . c;) or absence tBLM - . .1 of bleornycin. Panel V.:
a portion (20/~1~ of each m m # e (78.1101 ',,,as electrophore,,ed on an SDS-p~lyac~'lamide gel. The 34-kDa protem band i:, indicaled by an
asterisk,
'~'3! O I"
~,.m u .- 2
O
U m <[
B
kDa
68 ~"
1,3
29 2z;
, ! t 1' I I' "
M
IC 20 30 40 50 60 70
FRACTION
MW32 34 36 38 40 42 L./.,46 /.8
l ip 4,
0 4
0,2 K.
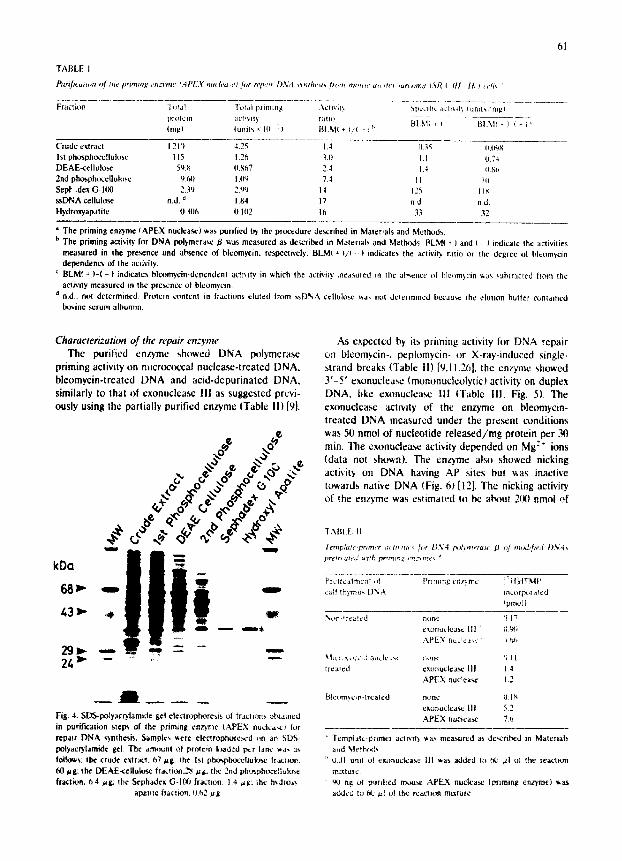
Fig. 3 tlydrox2,apatite chromatography ~f th,." priming enzyme. Panel A: Iraclton Nf, v, as snblected to chromalogJaphy t,n hydro~'apalJte as de,,cribcd in the text. Priming activity ',',a, measured as described
in Mdtcdals and Method,~ and in the legerd Io Fig. 1. Panel B: a portiun {5{} ,~11 of each ,,~'r:~-i~ !:~ ' ! ' ,~!:¢~re,.ed on an
The priming enzyme in the crude permeable cell extract was purified about 1 200-fold at the Sephadex G-Ill0 purification step {Table I). Although the specific activity and enzyme stability were reduced by further pt~rification using hydroxyapatite, the enzyme prepara- ti.~ became physically homogeneous, migrating as a single 34-kDa protein band in SDS-PAGE (Fig. 4). The intensity of the CBB-stained 34-kDa protein band was the lowest in the N: fraction, suggesting that the 0.2 M KPi extract contained both a phosphocellulose- absorbable and a non-absorbable form of the 34-kDa protein (Fig. 41, The adsorbable 34-kDa protein was the priming enzyme, The DNA pol vmerase priming activity of the 34-kDa protein on bleomycin-damaged or acid-de,-~urinated DNA was also detected by blotting (activity blotting) on bleomyein-damaged or acid-de- purinated DNA-fixed membranes or by measuring priming activity using renatured protein extracted from gel slices [12] (data not shown).
61
TABLE 1
~ r i f i ( a l , m o f tile priming enzyme t.4P/:,Y , le . 'h , . ,e l for lgp!l, tr DP,24 ~ mhe~t~ ~r .m mtar, c a~ ~n'~ ~ur¢ . m a I{~R ( l i t " I t ~ c/t, "
Fraction 1 .t~! Total p, liming ,~cll~'it} ~DC~.IIIC aCti~ It'. bruit,, ' rngl
(me} (units~ 10 2} BI.M{ + ) / ( - 1 ~"
Crude extract 1210 4.25 1.4 0.3g 0.1~,~ l st phosphocellulose l 15 1.2~ 3 .[} 1. t 0.74 DEAE-cellnlose 59.~ 0,~67 24 1.4 IJ~h 2nd phosph{~ellufilse 9.60 I .l~ 7.4 ] 1 Ill Sept ,dex G-100 2.3 {} 2.0 0 14 12'~ I I s ssDNA cellulose n.d. d I .M I 7 n.d n d. Hydrox'/apatite 11.3116 0.102 Ib 33 32
a The priming enzsme ~APEX nuclea.~) was purified ~ the procedure de~ribed in Materials and Metba}ds. t, The priming activily for DNA polymera.,.e # was measured as described in Materials and Methods, I~LMI + ) and t - P indicate the activities
measured in the presence and absence of bl¢omycin, respectively. BLMI + )/~ -- I indicates the aetwit!, ratio or the degree of Neomycin dependency of the ao iv ib .
r BLM~ + ) - ( - ) indicates bleomycin-der~ndcn! OCtB Ity in which the actr,'ily ,neasured in the ab,.ence of Neomycin ~as ,~abtracted kom the activity measured in the presence of bleom)cin
a rid.. not determined Protein content in fractions eluted from ~,DNA edlnlo,.e ~,as nol d¢lermmed becau~ the elation baiter contained ~}vine serum albumin.
Characterization of the repair enzyme The purified enzyme showed DNA polymerase
priming activity on mierococcal nuclease-treated DNA. Neomycin-treated DNA and acid-depurinated DNA, similarly to that of cxonuclease 111 as suggested previ- ously using the partially purified enzyme (Table I1} [9].
, .o
oO on o.O .o oo o*.e
e
88}" ~ ,
43 ~- ~
291~ -=~ ~ ----" -- 1--
24 )" - - . . . . '
., l Fig. 4. SDS-pol~.acqtamide gel electrophorests ol fractions c.b~amed in purification steps of the priming ea t 'me /.APEX nuclea~¢~ for repa~r DNA s)mhesis. Samples gere electrophote,,ed ~rn an SDS- pobacr~lamide gel. The arr~tunt of protein hmded per lane ~a- as folk~ws: the crude extract, 87 tag. the 1st pht~p-bocdluk~e fr~c~cm. 60 ~g: the DEAE~ellulose fraction28 ,ag~ the 2rid phos~:,celtuk.~*: fraction, 6a #g; the Scphadex G-100 fr~l ton. 14 p,g: the hydmr<~
apatlte fTactlon. 062 tag
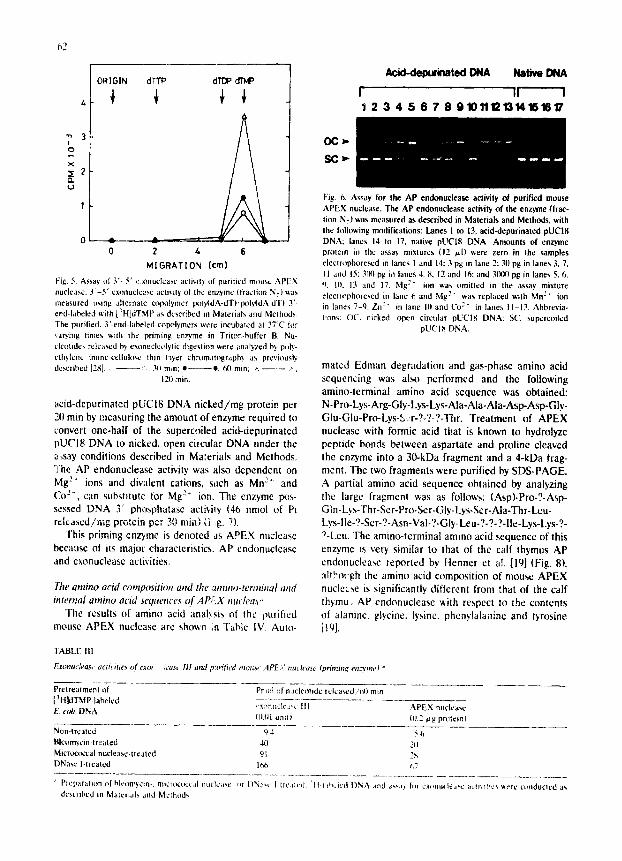
As expected by its priming activity for DNA repair on Neomycin-, peplomycin- or X-ray-induced single- strand breaks (]'able il)[9,11,26], the enzyme showed 3'-5' exonuclea~ (mononucleolytic) activity on duplex DNA, like exonuclcase II1 {'Fable III. Fig. 5). The exonuclease activity of the enzyme on bleomycin- treated DNA measured under the present conditions was 50 nmol of nucleotide released/rag protein per min. The exonuclea~ activity depended on Me:" ions (data not sho~n). The enzyme also showed nicking activity on DNA having AP sites but gas inactive towards native DNA (Fig. 6} [12]. The nicking activity of the enzyme was estimated to be about 20(} ~mol of
1ABLL It
Tcmplaf{..pr~mer ,~c Itr mr~ f . r 11~,2,1 pcd),rterasc fl
prelrt atrff axth pr* tn~ , l,t~2~mc$ *
oj nuMif ied 0 % ,i
Pruli'catmt-nt ~1~ Pr*ming cn/'~mc !~HldTMP c;~!f t~)-mu~ DNA incorf'n!t ated
! pmol
Non-*reat*d ~me [I ! ? c~onude a~,c Ill ~' t}~i
{~ea!ed e~.onudea~ Ill 1.4
APEX nuc!ea~¢ 1.2
Bleom~cmqrea|ed none {}18 exonudeast l i t 5.2 APEX n u d e a ~ ?.b
]empl~t~,pnmcl acH',iI) '~.as measured as de~Tribcd in Materia|~
LJ dl urn| of ¢~om~ckray._- Ill was added v_, 60 ,al of '~he rea,cti(m mL~tur¢
• ',~J ng of p~r~f~ed rr~ms{ APEX nudease lemming enzyme} ~a$ added to 6{J .=l o{ the reactron mixture
62
ORIGIN dTTP dTDP dTMP ¢ ¢
I O
~2
0 2 /,
MIGRATION (cm)
iv Fig. 5. Assay of 3 ' -5 ' e.:onuclease activity of purified mouse A P E X nuclease. 3 ' -5 ' exonucte~rse acti,,ilY of the enzyme (fraction N 7) v.as measured using alternate ¢opolymer l~ly(dA-dT)'lX~b4dA-dTl 3'- end-labeled ~ith [3H]dTMP as described in Materials and Methods The purified. 3'-end-labeled copolymers were incubated at 37°C for ~a~'ing limes v.ith the priming enzyme in Triton-buffer B. Nu- cleotides released h) exonuclct,lylic digestion ~ere analyzed by poly- ethylene /mine-cellulosc thin l*lyer chromatography as previously described [2S] :. 311 min: • e, 60 rain: ~ ~.
12tl rain.
a c i d - d e p u r i n a t e d p U C I 8 D N A n i c k e d / m g p ro t e in p e r
21t rain by m e a s u r i n g t he a m o u n t o f e n z y m e r e q u i r e d to
conve r t o n e - h a l f o f the supe rco i l ed a c i d - d e p u r i n a t e d
p U C I 8 D N A to n icked , o p e n c i rcu la r D N A u n d e r t he
~,say condi t ions de sc r i bed in Ma :e r i a l s a n d M e t h o d s .
The A P e n d o n u c l e a s c act ivi ty was a lso d e p e n d e n t on
Mg 2. ions and d iva len t cat ions , sach as M n 2" and
Lo- , can substitute for Mg 2~ ion. The enzyme l~s- sessed DNA 3' phosphatase activity (46 nmol of Pi relcased/~;ig protein pcr 3',) min) (i:g. 71.
"rhis priming enzyme is denoted as APEX nuclease becau~ of its major characteristics. AP cndonucleasc and exonuclease activities.
The amino acid composttion and the atnino-terminat and internal amino acid seqttence~ of AP;~X nmh'as,'
The results of amino acid anal~si~ of the purified mouse APEX nueleasc arc shown in Table IV. Auto-
FABLE Ill
Exonuch,as~' aeli~itie~ o f t:to~ :cast III and p:v,fied memse APE.,," lltl¢ h'asC
Acid-dep~inated ONA Native DNA
f II I I 2 3 4 5 6 7 8 910111213141511617
OCl=.
SC
Fig~ ft. Assay for the AP endonuclease activity of purified mouse A P E X nuclease. The AP endonuclease activity of the enzyme (frac- tion N 71 was measured as described in Materials and Methods, with the following modifications: Lanes 1 to 13, a¢id-depurinaled pUCI8 DNA: lanes 14 to 17. native pUCI8 DNA. Amounts of enzyme protein in the assa~, mixtures (12 V.I) were zero in the samples electrophore~d in lanes I and 14:3 !0g in lane 2:30 pg in lanes 3, 7. 11 and 15:3t10 pg in lanes 4. 8, 12 and 16: and 3(~10 pg in lanes 5. 6. 9. l(t. 13 and 17. Mg 2" ion was omitted in the assay mixture electr*~phoresed in lane 6 and Mg 2" was replaced with Mn2" ion in lanes 7-q. Zn 2" in lane 10 and Co:" in lanes 11-13. Abbrevia- tions: PC. nicked, open circular pUCI8 DNA: St', supctcoiled
pU('18 DNA.
matcd Edman degradation and gas-phase amino acid sequencing was a l ~ performed and the following amino-terminal amino acid sequence was obtained: N-Pro-Lys-Arg-Gly-Lys-Lys-Ala-Ala-Ala-Asp-Asp-Gly- Glu-Glu-Pro-Lys-S=r-?-?-?-Thr. Treatment of APEX nuclease with formic acid that is known to hydrolyze peptide bonds between aspartate and proline cleaved the enzyme into a 30-kDa fragment and a 4-kDa frag- ment. The two fragments were purified by SDS-PAGE. A partial amino acid sequence obtained by analyzing the largc fragment was as follows: (Asp)-Pro-?-Asp- GIn-Lys-Th r-Se r- Pro-Ser-Gly-Lys-Sc r-Ala-Thr-Leu- Lys-lle-?-Ser-?-Asn-Val-?-Gly-Leu-?-?-?-lle-Lys-Lys-?- ?-Lcu. The amino-terminal amino acid sequence of this enzyme is very similar to that of the calf thymus AP endonueleasc reported by Henner et al. [19] (Fig. 8), altbot.gh the amino acid composition of mouse APEX nuele~se is significantly different from that of the calf thymu, AP endonuclease "~,ith respect to the contents of alaninc, glycine, lysinc, phcnylalanine and tyr(~ine [191.
(pritr2ll?g e/lztprlt,~ a
Pretreatment i~f Pr ~ol tfl n.=cleotidc relca,,,:d/t-~t rain [ ~ HklTMP-labeled /:' ('~dt DNA ,,x~t~d¢ ,,c 11l APEX nuclease
t().{) | Unll I (0.2 U g prl~tein)
Non-tr¢~led 94 56 Bleumycin-treated 40 2It Micr~x:t~cal nucleasc-trcated 91 2S DNasc I-lleatcd 1¢~6 ~,7
" Preparation of hlcc~m~,cin-, micr~.occ,iI nl=cle~,~c or I)N;~c I Ir~';tlcd : l t~ d',,,:k'd [)NA ,~ntl a~,,,;~, to~ ¢x~mudeJ,e ~ = ~ . ~ l l ~ . ' . , v, erc corldnoud as described m M~le=hds and Mcfl~ods
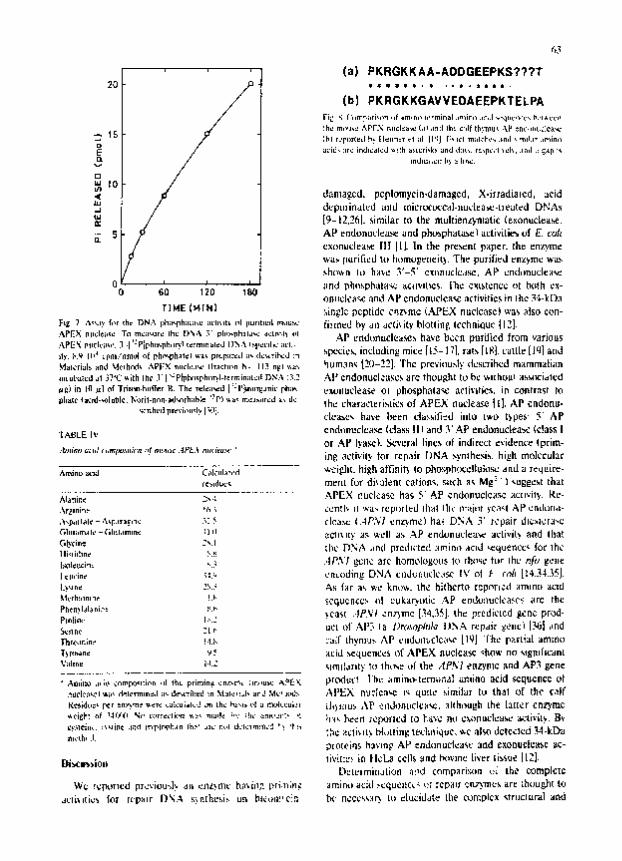
(,3
]c~ ~ .*.~.,? f,,r Ih r DN . "~, I:~;e-ph;mL;m,u ;,L'lmlmkk ,~1 ImurmbmL r~.mu~" 6 i~m~d ~ a l l a¢(i~i(~' hlotl i~l~'. I ¢ ¢ h r l ~ u ¢ ~1Z].
~uleriul~ and ~Jclhrd~ ,~,PF)C ~.c l ,u~. IIT~cht~n h- 113 n[Zl ' ~ klUnl3r~4 l~l---~-~]. ' r h : prcVi<lu~l~" dlJXL~'ih~d mxmmzEi~ lT,
~)in I~l~zl of) ' r iu~n.h.~er l~ .The Tele; l~L]J: : l '~m"~a~iL 'F~;~ UX4JIILICld~.'I~ ¢1 I J l l c ~ p h a h l ~ ~ ,c l i¢ i r~ . , ir~ con t ras t Do
"L'~BLE I~' cn r61 iTuc lca~ (~-~a~ I l l a n d 3 ' . A P ~ n d L ~ L L C ~ . ~ (c!a~,~ I
. ~ , i i I ~ I ¢ -.~.~F.Ir*~'n~ .%: ~ clca,.= ( .4P. 'v ' I c n z ) m ¢ ) ha~ D)%~. 3 ' ~cpaJr dl¢*]¢r;4~c
G b T i ~ S~.l ch~" ]3?~t ~, . rod pscdlc'tc£L : ~m im+ acid ~qUeT~L~¢ I'O[' lh¢ 1 Ik l i lbn¢ ~'.~
I¢l l l ' ine ~:.:, , . 'RLodin~ D N , ~, Cl )J~r ;u~lL 'd~" I V ¢11 ~ rr~'l [!¢.~-~...~5J. I Iy~ne 2~..; ~,)l fHr a~ ~'L" ]~nL:'~I t he hi(DJ~r~o C':prYFI¢~t Hmmn[~ ~LC3d )'l~'rh!Ilnlle ! 'b ~cc~ucn¢¢~ 411 L'Uk~LryLktic ..s.p CDc~Jl)tlclcasC'~ ~ r c r) l¢ Pl~li¢." I . , . : ~.CSS[ ..)~.%:d . :n .~Tnc [.'~4,3~,J. t h= ? red i c [ cd , ~ n ¢ prc~] I
")l,dr61r.;n~ )IRI ~ : l )~r Lhym:J~ i 'ke ¢lmduml)clcak~" Ll~)J r h ~ , pPIrLib~L a~;~l¢) T)r<hane ' ~ E i~L ~ . U ¢ ~ , : S Or" A P E X . ~ucmc~lc q~, n o %I~T[I[B--JITIL
~!'.~1~'~11~. :~JiP¢ .l~d I~l~lr~h~n I~:" Jl~" f:uI L]'.'IClm!II='~ ~ ! '~h
L~,=Lummi[mJtio¢l ;i,. ld comDar isc)r l ~ [ t l~e oori lpmcz¢

64
functional relationships between the various species of mammalian repair enzymes including AP endonucle- asc. We are now attempting eDNA cloning for APEX nuclease using probes of oligonucleotides deduced from the determined partial amino acid sequences. Such an approach is also important for studying repair deft- cient-inherited diseases.
Acknowledgements
We thank Mr. T. Nakamura and Ms. T. Yasul lor their technical assistance and thank Nippon Kayaku for providing copper-free bleomycin A,. This work was supported in part by a Grant-in-Aid for Scientific Re- search from the Ministry' of Education, Science and Cultured of Japan.
References
I Friedberg, E.C. (1985) DNA repair. W H Freeman and Com- pany, New York.
2 Lindahl, T. (1982) Annu. Roy. Biochcm. 51 61-87. 3 Yeoule, R. (1987) Int. J. Radlat. Biol. 51,573-589 4 Giloni, L., Takeshita, M., Johnson, F., Iden. C. and Grollman,
A.P. (1981} J. BioL Chem. 256. 8608-8~15. 5 Murugesan, N., Xu. C., Ehrentcld, GM.. Sugiyama. II.. Kilkuskic,
R.E., Rottriguez, L.O., Chang, L,-II. and Hecht, SM. (1985) Biochemistry 24, 5735-5744.
6 Burger, R.M,, Prolan, S.J., Ho~itz. S.B. and Peisach. J. (1988)J. Biol. Chem. 26t, 15955-15959.
7 Seki, S. and Oda, T (1986)Carcinogenesis 7. 77-82. 8 Seki, S,, Mori, S., Nakashima, A. and Oda, T. (1987) Carcino.
genesis 8, 1391-1394. 9 Scki, S, and Oda, T. (1~8) Carcinogenesis 9, 2239-2244
10 Seki, S., Arakaki, Y. and Od~t. T (1989) Acla Med, Okayaroa 43, 73-80.
11 Seki, S, Ikeda. S., Tsulsui, K. and Tcraoka. H (1990) Carcino- genesis II , 1213-12t6.
12 Ikeda. S., Seki, S.. Watana~re, S.+ Hatsushika, M and Tsutsui, K. (19911 Anal. Biochem. 192, 96-103.
13 ~'eiss, B. and Grossman, L. (19871AdvJ EnzymoL 6(I, 1-34. 14 John,)n, A.W. and Demple. B. (19881 J, Biol. Chem. 263+ 18017-
18022. 15 Ludwig. G. and Thidmann+ H.W. (19791 Nucleic Acids Res, 6.
2tK) l - 2917. 16 Nes, I.F. i1980) Nucleic Acids Rcs. 8, 1575-1589. 17 Toml, inson, AE.. Bonk, R.T. and Linn. S. (1988) J. Biol. Chem.
2¢)3, 12532-12537. 18 (?esar, R. and Verly, W.G. (19831 Eur. J. Biochem. 129, 50~-517. 19 Ilenncr, W.D., Kiker, N.P.. Jorgensen, T.J. and Munck. J.-N.
(1987) Nucleic Acids Res. 15. 5529-5544. 20 Kane. C.M and Linn. S. (1981} J. Biol, Chem. 256, 3405-3414. 21 Shaper, N,L., Grafstrom, R,H. and Grossman, L. (1982) J. Biol.
Chem. 257. 13455-13458. 22 Brent, T.P (1983} Biochemistry 22, 4507o4512. 23 Seki, S. and Oda, T. (19771 Cancer Res. 37, 137-144. 24 Sambrook, J., Fritsch, EF. and Maniatis, T. (1989) Molecular
Clonig, A Laboratory Manual, 2nd Edn. Cold Spring Harbor Laboralory Press, Cold Spring Harbor.
25 Niw~. O. and Moses. R.E. (1981) Biochemistry 20, 238-244. 26 Zhang 13o, Seki, S. and Ikeda, S. 11991) Int. J, Biochem. 23,
703-711 27 Schlabach, A.. FridlendeL B.. Bolden. A. and Weissbach, A.
(1971) Biochem. Biophys. Res. Commun. 44, 879-885. 2X Seki. S.. Mori, S. and Oda, T. (1989) Bioehem. Int. 18, 235-242. 29 Hagen, U F.W. (1981) in DNA Repair (Friedbcrg, E,C. and
Hanawalt. P.C., eds.) Vol. I, pp. 431-445, Marcel Dekker. New York
,~) Richardram, C.C. and Kornberg, A. (19641 J. Biol. Chem. 239. 242-~0.
31 Chang, L.MS. (1973) J. Biol. Chem. 248, 3789-3795. 32 Lacmmli. U,K. (1970} Nalurc 227, 680-885. 33 Mosbaugh, D.W. and Meyer, R.R. {1980) J. Biol. Chem. 255,
111239-11}247. 34 Johnson, A.W. and Dempie, B. (1988)J. Biol. Chem. 25.3, 18009-
18016. 35 Popoff, S.C.. Spira, A.I., Johnson, A.W. and Demple, B. (1990}
Prc,¢. Natl. Acad. Sci. USA 87, 4193-4197 3b Kelley, M R , Venugopal. S.. Harless, J. and Deutsch, W.A.
(1989) Mol Cell. Blol 9. 965-973.