a new sauropod dinosaur from the lower cretaceous cedar … · 2012-07-17 · cedar mountain...
TRANSCRIPT

A new sauropod dinosaur from the Lower CretaceousCedar Mountain Formation Utah USA
MICHAEL P TAYLOR MATHEW J WEDEL and RICHARD L CIFELLI
Taylor MP Wedel MJ and Cifelli RL 2011 A new sauropod dinosaur from the Lower Cretaceous Cedar MountainFormation Utah USA Acta Palaeontologica Polonica 56 (1) 75ndash98
Brontomerus mcintoshi is a new genus and species of sauropod dinosaur from the Hotel Mesa Quarry in Grand CountyUtah USA in the upper part of the Ruby Ranch Member (AptianndashAlbian) of the Lower Cretaceous Cedar Mountain Forminusmation It is known from at least two fragmentary specimens of different sizes The type specimen is OMNH 66430 theleft ilium of a juvenile individual tentatively referred specimens include a crushed presacral centrum a complete andwellminuspreserved midminustominusposterior caudal vertebra the partial centrum of a distal caudal vertebra a complete pneumatic anminusterior dorsal rib from the right side the nearly complete left scapula of a much larger presumably adult individual andtwo partial sternal plates Brontomerus is diagnosed by five autapomorphies of the type specimen preacetabular lobe55 of total ilium length longer than in any other sauropod preacetabular lobe directed anterolaterally at 30 to thesagittal but straight in dorsal view and vertically oriented postacetabular lobe reduced to near absence ischiadicpeduncle reduced to very low bulge ilium proportionally taller than in any other sauropod 52 as high as long In aphylogenetic analysis Brontomerus was recovered as a camarasauromorph in all most parsimonious trees but with unminuscertain position within that clade The large preacetabular lobe of the ilium anchored powerful protractor and abductormuscles but precise interpretation is impossible without functionally related elements such as femora and proximal cauminusdal vertebrae Brontomerus is the eighth sauropod genus named from the Early Cretaceous of North America and moreremain to be described North American sauropod diversity did not decline catastrophically at the end of the Jurassic asoften assumed The most striking differences between Late Jurassic and Early Cretaceous sauropod faunas in NorthAmerica is that the former are abundant and dominated by diplodocids whereas the latter are comparatively scarcemdashthough still diversemdashand dominated by macronarians
Key words Dinosauria Sauropoda Camarasauromorpha Brontomerus Brontomerus mcintoshi diversity Early Creminustaceous North America
Michael P Taylor [dinomiketaylororguk] Department of Earth Sciences University College London Gower StreetLondon WC1E 6BT United KingdomMathew J Wedel [mathewwedelgmailcom] College of Osteopathic Medicine of the Pacific and College of PodiatricMedicine Western University of Health Sciences 309 E Second Street Pomona California 91766minus1854 USARichard L Cifelli [rlcouedu] Sam Noble Oklahoma Museum of Natural History 2401 Chautauqua NormanOklahoma 73072 USA
Received 27 July 2010 accepted 23 December 2010 available online 23 February 2011
Introduction
The record of Early Cretaceous sauropod dinosaurs in NorthAmerica was for many years poorly represented with theonly generic names in use being Astrodon Leidy 1865 andPleurocoelus Marsh 1888 the former represented only byteeth and often considered synonymous with the latter (egHatcher 1903a Gilmore 1921 Carpenter and Tidwell 2005)In recent years this record has been greatly expanded andclarified by the discovery and description of SonorasaurusRatkevich 1998 Cedarosaurus Tidwell Carpenter andBrooks 1999 Sauroposeidon Wedel Cifelli and Sanders2000a Venenosaurus Tidwell Carpenter and Meyer 2001Paluxysaurus Rose 2007 and Abydosaurus Chure BrittWhitlock and Wilson 2010 Further material representing
yet more new sauropod taxa is known and awaits descripminustion for example two new taxa from the Dalton WellsQuarry a camarasaurid and a titanosaurian (Eberth et al2006 220) and a titanosaurian from the Yellow Cat Memberof the Cedar Mountain Formation represented by an articuminuslated sequence of five presacral vertebrae (Tidwell and Carminuspenter 2007)
Here we describe a new sauropod taxon which further exminustends the record of Early Cretaceous North American saurominuspods The ilium and scapula of this sauropod were figured butnot described by Kirkland et al (1997 93) who consideredthem as ldquocomparable to Pleurocoelusrdquo (at that time the onlyknown Early Cretaceous sauropod from North America)
Institutional abbreviationsmdashAMNH American Museum ofNatural History New York USA BMNH the Natural Hisminus
doi104202app20100073Acta Palaeontol Pol 56 (1) 75ndash98 2011
tory Museum London UK CCG Chengdu College of Geminusology Chengdu China CM Carnegie Museum of NaturalHistory Pittsburgh USA DMNH Denver Museum of Natuminusral History Denver USA DNM Dinosaur National Monuminusment Dinosaur USA FMNH Field Museum of NaturalHistory Chicago USA FWMSH Fort Worth Museum ofScience and History Fort Worth USA HMN HumboldtMuseum fuumlr Naturkunde Berlin Germany OMNH Oklaminushoma Museum of Natural History Norman USA ZDMZigong Dinosaur Museum Zigong China
Anatomical nomenclaturemdashWe follow Upchurch et al(2004a) in describing scapulae as though oriented horizonminustally the coracoid articular surface is designated anteriorand the end commonly called ldquodistalrdquo is designated posteminusrior See Taylor (2009 787) for a summary of other schemesthat have been used Names of clades are used as summaminusrized in Table 1
Geological and faunal contextThe holotype and referred specimens described herein werecollected at OMNH locality V857 known as the Hotel MesaQuarry (Kirkland et al 1997 96ndash97 fig 28) One of us(RLC) who had already been working in Lower Cretaceousrocks of the region was notified about the site through thecourtesy of James I Kirkland and was guided to it by BillHawes of Grand Junction Colorado in September 1994 Adminusditional collecting at the site for OMNH was conducted byRandall L Nydam and James I Kirkland in March 1995
OMNH V857 lies on the southwest flank of Hotel Mesain easternmost Grand County Utah (see Kirkland and Madminussen 2007 fig 2) about 13 km northminusnortheast of the juncminustion of the Colorado and Dolores rivers (sec 4 T23S R24E
Dewey 75rsquo quadrangle USGS provisional edition 1985precise locality data are on file at OMNH and are available toqualified investigators upon request) This site had previminusously been worked by private collectors the number idenminustity condition and disposition of the privately collected fosminussils cannot now be determined However given the densityof bone still present and exposed and the fact that the existminusing quarry was already some 5ndash6 m long and 3 m deep it apminuspears that a considerable number of elements were removedfrom the quarry and that the loss of valuable scientific informinusmation has unfortunately been considerable Bones left exminusposed by these previous collectors were in various states ofdisrepair some had been broken and their pieces used to holddown the remnants of a plastic tarpaulin The material reminusported herein was recovered in the course of the two briefOMNH surveysalvage visits to this site
Stratigraphically OMNH V857 lies in a sequence ofLower Cretaceous rocks interposed between the MorrisonFormation (Kimmeridgian) below and the Dakota Formation(Cenomanian) above Westward these rocks are recognizedas the Cedar Mountain Formation eastward the Burro Canminusyon Formation The arbitrary dividing line between these enminustities is generally placed at the Colorado River (Stokes 1952Tschudy et al 1984) which technically places OMNH V857within the Burro Canyon Formation However we will referto the locality as belonging to the more widely recognizedCedar Mountain Formation as it is in this formation thatcomparable specimens are known and the stratigraphy andsedimentology do not change across the arbitrary borderThe most extensive recent applicable stratigraphic work isthat of Kirkland et al (1997 1999) who recognized fivemembers within the Cedar Mountain Formation (in ascendminusing order) the Buckhorn Conglomerate Yellow Cat PoisonStrip Sandstone Ruby Ranch and Mussentuchit members
76 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 1 Clade names used in this study and the definitions used For simplicity specifiers are indicated by genus rather than species in each case thetype species of the genus is intended Nodeminusbased clades are indicated by ldquo+rdquo branchminusbased clades by ldquonotrdquo
Clade name As defined by DefinitionSauropoda Yates (2006 12) Saltasaurus not Melanorosaurus
Neosauropoda Wilson and Sereno (1998 46) Diplodocus + SaltasaurusDiplodocoidea Wilson and Sereno (1998 17) Diplodocus not Saltasaurus
Diplodocimorpha Calvo and Salgado (1995 14) Diplodocus + RebbachisaurusRebbachisauridae Salgado et al (2004 910) Rebbachisaurus not Diplodocus
Diplodocidae Sereno (1998 63) Diplodocus not DicraeosaurusDiplodocinae Taylor and Naish (2005 5) Diplodocus not ApatosaurusMacronaria Wilson and Sereno (1998 49) Saltasaurus not Diplodocus
Camarasauromorpha Upchurch et al (2004a306) Camarasaurus + SaltasaurusCamarasauridae Taylor and Naish (2007 1555) Camarasaurus not Saltasaurus
Titanosauriformes Wilson and Sereno (1998 51) Brachiosaurus + SaltasaurusBrachiosauridae Wilson and Sereno (1998 20ndash21) Brachiosaurus not SaltasaurusSomphospondyli Wilson and Sereno (1998 53) Saltasaurus not Brachiosaurus
Titanosauria Wilson and Upchurch (2003 156) Andesaurus + SaltasaurusLithostrotia Wilson and Upchurch (2003 156) Malawisaurus + Saltasaurus
Saltasauridae Sereno (1998 63) Saltasaurus + OpisthocoelicaudiaOpisthocoelicaudiinae Sereno (1998 63) Opisthocoelicaudia not Saltasaurus
(The Buckhorn Conglomerate Member has a nonminusoverlapminusping geographic distribution with respect to the Yellow Catand Poison Strip Sandstone members and may be a lateralequivalent of one or both see Kirkland et al 1999 figs 1 2)The existing paleontological record from these units ispatchy but there is evidence of three distinct dinosaur faunasfrom the Yellow Cat (Barremian or older) Poison StripSandstone plus Ruby Ranch (AptianndashAlbian) and Mussenminustuchit (AlbianndashCenomanian) members respectively (Kirkminusland et al 1997) While spanning a total of some 30 Ma thefaunal record from the Cedar Mountain Formation is punctuminusated by significant hiatuses (Sames et al 2010) and fossilhorizons that are stratigraphically close may therefore bewidely separated in time
Like most terrigenous Mesozoic units of the Western Inminusterior (see for example Ostrom 1970 14) the laterally equivminusalent Cedar Mountain and Burro Canyon formations areenormously variable At Hotel Mesa the lowest two (Buckminushorn Conglomerate Yellow Cat) and uppermost (Mussenminustuchit) members are absent and only two members of the Ceminusdar Mountain Formation may be recognized the Poison StripSandstone (below) and Ruby Ranch (above) membersStratigraphically OMNH locality V857 lies within the RubyRanch Member a few meters below the local contact withthe Dakota Formation (see Kirkland et al 1999 fig 8) As aworking hypothesis we regard the fossils from the HotelMesa site as being AptianndashAlbian and therefore approximinusmately equivalent in age to assemblages from the CloverlyFormation of Wyoming and Montana and the Trinity Groupof Texas and Oklahoma (eg Ostrom 1970 Winkler et al1990 Jacobs et al 1991 Brinkman et al 1998 Davis et al2008) As elsewhere the Ruby Ranch Member in this area iscomprised predominantly of drab pale green and mauvemudstones with abundant carbonate nodules The fossil horiminuszon which is mainly sandstone with some pebbles and interminusmittent irregular mudstone stringers is about 40ndash50 cmthick Locally these mudstones contain sparse microverteminusbrate fossils often rounded or broken About 100 kg of thismatrix was collected and transported back to the OMNH forfossil extraction using underwater screen washing and assominusciated concentration techniques (Cifelli et al 1996)
Although the microvertebrate assemblage from HotelMesa is neither very diverse nor particularly well repreminussented it does merit attention as being the only such site ofAptianndashAlbian age within the Colorado Plateau A faunal listis presented in Table 2 As is generally the case for such asminussemblages a number of aquatic taxa are represented thoughno remains of Testudines have been recovered yet Probablytwo chondrichthyans are present a hybodontid (possiblyHybodus which was listed in the Ruby Ranch fauna byKirkland et al 1999 table 2) and a polyacrodontid perhapsPolyacrodus The neoceratodontid form genus Ceratoduswidespread in AptianndashAlbian faunas of North America (seereview by Kirkland 1987) is represented by a poorly preminusserved tooth plate two actinopterygians are present one repminusresented by teeth and ganoid scales probably referable to
Lepisosteidae No mammalian or lissamphibian fossils havebeen recovered and Reptilia is represented solely byarchosaurians of which crocodyliforms (three or four taxaincluding a species of the widespread taxon Bernissartia andunidentified species of Goniopholididae and Atoposauridae)are most diverse Megafossils of the Hotel Mesa Quarrymainly represent Sauropoda (including many unidentifiedfragmentary specimens in addition to those describedherein) theropods are known by isolated teeth and rare inminuscomplete postcranial elements The fauna includes anornithopod represented by a single nonminusdiagnostic toothKirkland et al (1999) mentioned the presence in the RubyRanch fauna of Tenontosaurus otherwise known from theCloverly Formation of Wyoming and Montana (Ostrom1970) and the Trinity Group of Texas and Oklahoma (Langminusston 1974 Winkler et al 1997b Brinkman et al 1998)based on specimens from an unidentified site on the westernside of the San Rafael Swell Utah The ornithopod specimenfrom the Hotel Mesa Quarry OMNH 27824 probably does
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 77
Table 2 Vertebrate fauna of the Hotel Mesa Quarry (OMNH V857)Ruby Ranch Member Cedar Mountain Formation (AptianndashAlbian)Grand County Utah
ChondrichthyesCtenacanthiformes
HybodontidaeGen et sp indet
PolyacrodontidaePolyacrodus sp
OsteichthyesOrder indet
Family indetGen et sp indet
LepisosteiformesLepisosteidae
Gen et sp indetDipnoi
NeoceratodontidaeCeratodus sp
ReptiliaCrocodyliformes
Family indetGen et sp indet
BernissartiidaeBernissartia sp
GoniopholididaeGen et sp indet
AtoposauridaeGen et sp indet
TheropodaFamily indet
Gen et sp indetSauropoda
Camarasauromorpha family incertae sedisBrontomerus mcintoshi gen and sp nov
OrnithopodaFamily indet
Gen et sp indet
not represent Tenontosaurus but further identification canminusnot be firmly established at present
Although the quarry has not been worked in severalyears it is not exhausted and may contain additional relevantmaterial (James Kirkland personal communication October2007)
Systematic paleontology
Dinosauria Owen 1842Saurischia Seeley 1888Sauropoda Marsh 1878Neosauropoda Bonaparte 1986Camarasauromorpha Salgado Coria and Calvo 1997Genus Brontomerus novType species Brontomerus mcintoshi sp nov see below
Etymology From Greek bronto thunder Greek meroacutes thigh ldquothunminusderminusthighsrdquo in reference to the substantial femoral musculature impliedby the morphology of the ilium Intended English pronunciation BronminusminustoeminusMEERminusus
DiagnosismdashAs for type and only species (see below)
Brontomerus mcintoshi sp novFigs 1 2 5ndash8 10 12 Table 3
Etymology In honor of veteran sauropod worker John S McIntoshwhose seminal paleontological work done mostly unfunded and on hisown time has been an inspiration to all of us who follow
Holotype OMNH 66430 a left ilium
Tentatively referred material OMNH 66429 crushed presacralcentrum OMNH 61248 midminustominusposterior caudal vertebra OMNH27794 partial distal caudal centrum OMNH 27766 anterior right dorminussal rib OMNH 27761 nearly complete left scapula missing anteriorportion OMNH 66431 and 66432 two partial sternal plates other fragminusments as detailed in Table 3
Type locality Hotel Mesa Quarry (OMNH locality V857) GrandCounty eastern Utah
Type horizon Top of the Ruby Ranch Member of the Cedar MountainFormation (Lower Cretaceous AptianndashAlbian)
DiagnosismdashPreacetabular lobe 55 of total ilium lengthlonger than in any other sauropod preacetabular lobe directedanterolaterally at 30 relative to the sagittal plane but straightin dorsal view and vertically oriented postacetabular lobe reminusduced to near absence ischiadic peduncle reduced to very lowbulge ilium proportionally taller than in any other saurominuspodmdashheight is 52 of total length compared with a maximinusmum of 45 in other sauropods If the tentatively referred eleminus
78 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 3 Material referred to the camarasauromorph sauropod Brontomerus mcintoshi from the Lower Cretaceous Cedar Mountain Formation ofUtah
OMNH specimen number Description Length (cm) Width (cm) Thickness (cm)27761 partial scapula missing anterior part 98 55 527762 large flat rib shaft 101 75 227763 rib fragment 63 8 527764 rib fragment 60 25 227765 rib fragment 58 6 327766 complete pneumatic rib 76 12 427767 dorsal part of rib flattened 26 17 227768 rib fragment 31 6 1527769 fragment of ischium 10 5 2527770 rib fragment 14 35 127771 rib fragment 17 3 227772 rib fragment (in two parts) 13 7 227773 rib fragment 135 4 227773 flat scrap 7 5 0527773 flat scrap 115 9 1
27774ndash27783 fragments27784 collection of 21 fragments
27785ndash27793 fragments27794 partial distal caudal centrum
27795ndash27800 fragments61248 nearly complete midminuscaudal vertebra 11 6 5566429 crushed presacral centrum 14 14 366430 ilium 405 31 866430 dorsal fragment of ilium 14 3 166430 dorsal fragment of ilium 155 9 166431 incomplete sternal plate 15 75 1566432 incomplete sternal plate 115 7 1
ments do belong to the same species as the holotype then thefollowing additional characters also diagnose the new taxonpresacral vertebrae camellate midminustominusposterior caudal verteminusbrae with elongate preminus and postzygapophyseal rami havingthe postzygapophyseal facets hanging below the level of theramus first dorsal rib with expanded dorsally oriented articuminuslar facets laterally curving shaft and ventrally directed pneuminusmatic foramen in head acromion expansion of scapula prominusnounced and steep but not forming acromion fossa dorsal andventral margins of scapular blade ldquosteppedrdquo sternal platescrescentic and three times as long as broad
Unambiguous autapomorphies distinguishing Brontomeminusrus from the root of the polytomy in which it is recovered inthe strict consensus of most parsimonious trees in the phylominusgenetic analysis below character 184 ratio of centrumlengthheight in middle caudal vertebrae 20 185 sharpridge on lateral surface of middle caudal centra at archminusbodyjunction absent 212 posterior end of scapular body racminusquetminusshaped (dorsoventrally expanded) 261 in lateral viewthe most anteroventral point on the iliac preacetabular lobe isalso the most anterior point (preacetabular lobe is pointed)264 projected line connecting articular surfaces of ischiadicand pubic peduncles of ilium passes ventral to ventral marginof postacetabular lobe of ilium
Description
This taxon is based on a collection of elements all from thesame quarry all of them consistent with assignment to a sinminusgle taxon (Fig 1) However the elements were not found arminusticulated and their differing sizes do not permit interpretaminustion as belonging to a single individual For example the parminus
tial scapula is 98 cm long Reconstruction after the scapula ofGiraffatitan brancai (Janensch 1914) suggests that the comminusplete element was about 121 cm long In Rapetosaurus CurryRogers and Forster 2001 the scapula and ilium are about thesame length (Curry Rogers and Forster 2001 fig 3) but theilium of Brontomerus is only one third the reconstructedlength of the scapula The quarry therefore contains at leasttwo individuals of widely differing sizes The smaller prominusviding the holotype ilium and the referred presacral centrumand sternal plates is interpreted as a juvenile the remainingelements probably belonged to the larger mature individual
Among the sauropod elements recovered from the HotelMesa site at least three (the caudal vertebra scapula andilium) have characters not known in any other sauropodSince it is not possible to conclusively demonstrate that allthree elements belong to the same taxon (and size differminusences preclude their belonging to a single individual) it istherefore possible that the quarry contains as many as threenew taxa However since all the informative elements havecharacters that indicate a titanosauriform identity we conminusservatively consider it more likely that only a single newtaxon is present Groups of sauropods of mixed ages areknown for several taxa (eg Bird 1985 Coria 1994 Bader2003) so there is a precedent for finding adults and juveminusniles together Although we diagnose the new taxon solelyon the type ilium we also tentatively refer the other eleminusments to this taxon and note the additional characters thatpertain if this referral is correct we hope that subsequentwork in the Hotel Mesa Quarry or elsewhere in the CedarMountain Formation will yield specimens that allow us toconfirm or refute this referral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 79
Fig 1 Skeletal inventory of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah in left lateral view Preserved elements are white missing elements are reconstructed in gray After a Camarasaurus grandis reconstructionkindly provided by Scott Hartman
The assignment of specimen numbers to the material deminusscribed here is complex (Table 3) Specimen number OMNH27773 comprises three elements OMNH 27784 consists of 21small fragments of bone none of them informative all otherelements have their own specimen numbers in the range27761ndash27800 apart from the midminuscaudal vertebra OMNH61248 and the reassigned numbers 66429ndash66432 for eleminusments extracted from OMNH 27773
IliummdashThe most informative element is OMNH 66430 a leftilium (Fig 2) which has therefore been selected as the holominustype The ilium was preserved complete but lay hidden beminusneath the scapula and so was damaged in the field (JamesKirkland personal communication March 2008) The iliumis preserved in three parts one provides most of the boneincluding the well preserved preacetabular lobe pubic andischiadic peduncles and acetabular margin and the other two
provide most of the dorsal margin giving a good indication ofthe degree of curvature The relative positions and orientationof the two smaller fragments can not be definitely ascertainedbut they appear to be parts of a single large fragment broken atthe point where we have reconstructed them as touching andif this interpretation is correct then the curvature of the pair inminusdicates which side must be oriented laterally
The ilium is remarkable in that the preacetabular lobe isrelatively larger than in any other sauropod (Fig 3 Table 4measurement protocol is illustrated in Fig 4) the postaceminustabular lobe is reduced almost to the point of absence and theilium is proportionally taller than in any other sauropod Theischiadic peduncle is reduced to a very low ventral projectionfrom almost the most posterior point of the ilium The near abminussence of the ischiadic peduncle cannot be attributed to damageas the iliac articular surface is preserved Immediately posterominus
80 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
acetabulum
pubicpeduncle
10 cm
ischiadicpeduncle
pre-acetabularblade
Fig 2 Left ilium of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah type specimen OMNH 66430 in lateral view reconstructed from the three fragments (A) and ventral view (B)
dorsal to this surface is a subtle notch between the peduncleand the very reduced postacetabular lobe This notch and theareas either side of it are composed of finished bone demonminusstrating that the great reduction of the postacetabular lobe toois a genuine osteological feature and not due to damage In reminusgard to the proportionally large preacetabular lobe the iliumof Brontomerus resembles that of Rapetosaurus (Fig 3E Taminusble 4) However that taxon has a normal postacetabular lobeand is not proportionally tall In overall proportions the iliumof Brontomerus is more similar to the left ilium HMN J1 asminussigned to Giraffatitan brancai (Janensch 1961 pl E 2) whichalso has a reduced postacetabular lobe mdashsee also Fig 3DHowever the ilium of Brontomerus is proportionally tallerthan that of G brancai and its anterior margin comes to apoint rather than being smoothly rounded as in that taxon InBrontomerus the maximum height of the ilium above theacetabular margin when measured perpendicular to the lonminusgest axis is 52 in other sauropods examined this proportiondoes not exceed 45 and averages 40 (Table 4)
As with many sauropods the preacetabular lobe of theilium flares laterally However in most sauropods this flarminusing is progressive so that in dorsal or ventral view the mostposterior part of the preacetabular lobe is nearly parallel witha line drawn between the pubic and ischiadic peduncles andsmooth lateral curvature inclines the more anterior parts inminuscreasingly laterally so that the more anterior part is almost atright angles to this line and the ilium appears smoothly
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 81
Fig 3 Ilia of sauropod dinosaurs scaled to same total length A Mamenchisaurus hochuanensis Young and Zhao 1972 holotype CCG V 20401 right iliumreversed modified from Young and Zhao (1972 pl 6 1a) B Diplodocus carnegii Hatcher 1901 CM 94 right ilium reversed modified from Hatcher (1901pl 10 1) C Camarasaurus supremus Cope 1877 AMNH 5761 Il 1 left ilium modified from Osborn and Mook (1921 fig 87) D Giraffatitan brancai(Janensch 1914) HMN J1 left ilium modified from Janensch (1961 pl E 2) E Rapetosaurus krausi Curry Rogers and Forster 2001 holotype FMNH PR2209 left ilium modified from Curry Rogers (2009 fig 39B) F Brontomerus mcintoshi gen et sp nov holotype OMNH 66430 left ilium
Fig 4 Measurement protocol for sauropod ilia as illustrated in Fig 3 andshown in Table 4 Total length is measured along the longest axis of theilium lengths of preacetabular and postacetabular lobes are measured parminusallel to this axis and extend from the extremity of the lobe to the anteriormargin of the pubic peduncle and posterior margin of the ischiadic pedunclerespectively Supracetabular height is measured perpendicular to the lonminusgest axis and extends from the highest point of the acetabulum to the pointlevel with the highest part of the ilium
curved in dorsal or ventral viewmdashfor example ApatosaurusMarsh 1877 (Upchurch et al 2004b pl 4 D E) Haplominuscanthosaurus Hatcher 1903b (Hatcher 1903a pl 5 1) andSaltasaurus Bonaparte and Powell 1980 (Powell 1992 fig17) In Brontomerus by contrast the blade of the ilium apminuspears to be ldquohingedrdquomdashdeflected laterally at a point directlyanterior to the public pedunclemdashso that the preacetabularlobe is straight in dorsal or ventral view and directed anterominuslaterally by an angle of about 30 to the sagittal In this reminusspect it more closely resembles the ilium of Camarasaurus
Cope 1877 (Osborn and Mook 1921 fig 49) although it difminusfers in other respects
The Brontomerus ilium is laterally compressed and unlikemost sauropod ilia the dorsal margin is not deflected laterallyrelative to the more ventral part so that in ventral view it apminuspears very thin (Fig 2B) It is well established that the longbones of sauropods grow isometrically through ontogeny(Carpenter and McIntosh 1994 Wilhite 1999 2003 Ikejiri etal 2005 Tidwell and Wilhite 2005 Taylor 2009) while theirvertebrae undergo significant changes in proportions laminaminus
82 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 4 Relative measurements of ilia in sauropods Measurement protocol is illustrated in Fig 4
Taxon Specimen ReferenceTotallength(cm)
Length ofpreacetabular
lobe (cmproportion oftotal length)
Length ofpostacetabular
lobe (cmproportion oftotal length)
Supracetabularheight (cm
proportion oftotal length)
Mamenchisaurushochuanensis
CCG V 20401 Young and Zhao (1972 pl 6 1a) 102 39 38 18 18 41 40
Diplodocus carnegii CM 94 Hatcher (1901 pl 10 1) 1091 41 38 20 18 49 45Camarasaurus
supremusAMNH 5761 Il 1 Osborn and Mook (1921 fig 87) 115 36 31 19 17 48 42
Giraffatitan brancai2
HMN Aa 13Janensch (1961 pl E 1a) 119 47 39 21 18 46 39
Measured by SchwarzminusWings(personal communication 2009) 1143 535 47 299 26
HMN J1Janensch (1961 pl E 2) 1055 55 52 16 15 37 35
Measured by SchwarzminusWings(personal communication 2009) 1077 528 49 135 13
Rapetosaurus krausi FMNH PR 2209 Curry Rogers (2009 fig 39) 42 20 48 7 17 16 38Brontomerus mcintoshi OMNH 66430 (this study) 405 223 55 0 0 21 52
1 Hatcher (1901) did not state the length of the ilium of CM 94 and did not figure that of CM 84 We have assumed that the ilium of the figured specimen CM94 is the same size as that of CM 84 (Hatcher 1901 46) and calculated the proportions from the figured specimen
2 Janenschrsquos (1961 pl E) figures of the two ilia of Giraffatitan brancai are not executed from an orthogonal perspective the ilium of HMN Aa 13 is illusminustrated from a slightly anterolateral position foreshortening the preacetabular lobe and that of HMN J 1 from a slight posterolateral position foreshortenminusing the postacetabular lobe The true lengths of the two lobes are probably somewhere between the two percentages calculated from the figures about45 for the former and 16 for the latter
10
cm
Fig 5 Damaged presacral vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah OMNH 66429 in dorsal view as photograph (A) and interpretive drawing (B) Shading indicates air spaces
tion pneumatic excavations and neurocentral fusion (Wedel2003a 248 b 352ndash354) Ontogenetic changes in limbminusgirdleelements such as the ilium are less well understood due to apaucity of sufficiently well preserved specimens (Ray Wilhitepersonal communication October 2007) Therefore the lateralcompression of the Brontomerus ilium may be a juvenile charminusacter with the ilium thickening through ontogeny to supportthe growing weight of the animal or it may in fact be phylominusgenetically significant
Presacral centrummdashA single presacral centrum OMNH66429 was recovered (Fig 5) Unfortunately preservation isvery poor the neural arch and all processes have been lostand the centrum has been greatly crushed dorsoventrally sothat the remaining part is essentially flat the element is 14
cm in both anteroposterior length and transverse width butno more than 3 cm in dorsoventral depth The small size indiminuscates that this element belonged to a juvenile The internalstructure of the centrum is visible however and consists offine septa dividing a hollow internal space irregularly intomany small camellae This morphology is characteristic oftitanosauriforms (Wedel 2003b 354ndash355) Highly camellateinternal structure has not previously been observed in juveminusnile sauropod vertebrae but this may be due to samplingbias so far all juvenile sauropod vertebrae that have beenstudied for internal structure have been those of Camaraminussaurus and diplodocoids which follow an ontogenetic trajecminustory in which large shallow lateral fossae develop into cameminusrae from which smaller accessory camerae and camellae deminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 83
prezygapophysis
neural spine
postzygapophysealramus
postzygapophysealfacet
chevron facets
5 cm
Fig 6 Midminuscaudal vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 61248 in dorsal (A) anterior (B) left lateral (C) posterior (D) and ventral (E) views
velop (Wedel et al 2000a fig 11 Wedel 2003b 349) TheHotel Mesa presacral suggests that camellate vertebrae mayhave developed differently in titanosauriforms possibly byinminussitu formation of camellae during pneumatization as ocminuscurs in the camellate vertebrae of birds (personal observaminustion MJW)
Caudal vertebraemdashOMNH 61248 is a distinctive caudalvertebra with elongated preminus and postzygapophyseal rami(Fig 6) Apart from the tip of the right prezygapophysis theelement is complete and well preserved While the centrum isonly 11 cm in length the distance from the prezygapophysisto postzygapophysis is 145 cm The centrum is slightlybroader than tall (6 cm compared with 55 cm anteriorly 65cm compared with 5 cm posteriorly) and gently waisted Theneural arch is set forward on the centrum but does not reachthe anterior margin The neural spine is so reduced and sostrongly inclined posteriorly as to be all but indistinguishminusable and is apparent only as a very low eminence abovethe postzygapophyses The postzygapophyseal facets themminusselves are set on the posterolateral faces of a low process thathangs below the main postzygapophyseal ramus Chevronfacets are weakly present on the posterior margin of the venminustral surface of the centrum but not on the anterior marginThe elongation index of 22 indicates a midminustominusposterior pominussition in the caudal sequence for this element as similarcentrum proportions do not appear until about caudal 30 inGiraffatitan brancai (Janensch 1950 pl 3)
This vertebra most closely resembles the indeterminatesauropod vertebra BMNH 27500 from the Wessex Formationof the Isle of Wight figured by Naish and Martill (2001 pl33) The Barremian age of that specimen places it about 15 Ma
earlier than OMNH 61248 Its neural arch is less elevated thanthat of the Hotel Mesa specimen its postzygapophyses projectyet farther posteriorly and its prezygapophyses less far anteriminusorly and it is very mildly biconvex rather than procoelous butin other respects including absolute size it is a good match forthe Brontomerus caudal
Also included in the Hotel Mesa material is OMNH27794 a partial distal caudal centrum figured by Wedel(2005 fig 77) This centrum is approximately round incrossminussection about 4 cm in diameter and internally consistsof apneumatic cancellous bone
RibsmdashThe Hotel Mesa material contains several dorsal ribsin various states of preservation but no readily identifiablecervical ribs The dorsal rib elements include the shaft of alarge flat rib (OMNH 27762) portions of several smaller ribshafts (OMNH 27763ndash27765 27768 and others) a flattenedrib head (OMNH 27767) and most informatively a smallcomplete rib (OMNH 27766 Fig 7) Despite its excellentpreservation and apparent lack of distortion this element isdifficult to interpret Its shaft is straight for almost its wholelength and both articular facets are directed dorsally ratherthan being inclined medially The tuberculum is directly inline with the main part of the shaft of the rib and thecapitulum is at an angle of about 30 to it In these respectsthe rib resembles the most anterior dorsal rib of the Diplominusdocus carnegii holotype CM 84 (personal observation MPTthis element was not figured by Hatcher [1901]) We thereminusfore interpret this rib as having probably belonged to the rightside of the first dorsal vertebra
The rib is unusual in other respects however most notablythat the ventral part of the shaft curves laterally rather than meminus
84 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
10 cm
capitulum
pneumatic fossa
tuberculum
Fig 7 First right dorsal rib of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah OMNH 27766 in posterior view head of rib showing pneumatic invasion of shaft (A) and complete rib showing laterally directed curvature of shaft (B)
dially Careful inspection of the bone reveals no indication ofdistortion or of incorrect reconstruction It may be possiblethat in life the thorax was transversely compressed so that thedorsal part of the rib shaft was directed ventromedially and themore ventral part was vertical Both articular facets are subminuscircular in dorsal view and significantly expanded comparedwith the rami that bear them On the anterior face of the head alow ridge arises just below the capitulum and extends downthe medial edge of the rib for about 40 of its length
The head of the rib is also unusual in that a thin sheet ofbone connects the rami that support the articular facets andthis sheet extends much farther proximally than in mostsauropod ribs Its precise extent cannot be ascertained due tobreakage The sheet of bone is perforated close to the capituminuslar ramus and from this perforation a pneumatic cavity inminusvades the shaft of the rib extending ventrally from within ashallow fossa in the posterior face Pneumatization of thedorsal ribs is a synapomorphy of Titanosauriformes (Wilsonand Sereno 1998) although pneumatic dorsal ribs are also inminusfrequently present in diplodocids (Gilmore 1936 Lovelaceet al 2003 2008)
Pectoral girdlemdashOMNH 27761 is a partial scapula consistminusing of the blade and part of the anterior expansion but missminusing the glenoid region and the remainder of the anterior exminuspansion (Fig 8) As preserved the element is nearly flat butthis may be due to postminusmortem distortion and in any casethe most strongly curved part of most sauropod scapulae isthe anterior part that is missing from this specimen The genminustle curvature preserved in the posterior part of the blade indiminuscates that the element was from the left side The bone is surminusprisingly thin in all preserved parts never exceeding a few
cm in contrast to for example the scapula of Camarasaurussupremus which is thick even in midminusblade (Osborn andMook 1921 fig 74b) This suggests that the glenoid thickenminusing and the acromial ridge may have been located some disminustance anterior to the preserved portion and a reconstructionafter the proportions of Giraffatitan brancai (Fig 8) suggeststhat about 80 of the scapularsquos full length is preserved Theposterior part of the acromion expansion is preserved howminusever and is sufficient to show that this expansion was prominusnounced so that the maximum dorsoventral height of thescapula was more than two and half times its minimumheight at the midpoint of the blade The dorsal margin slopesup towards the anterior expansion rather than forming a posminusteriorly directed ldquohookrdquo or a distinct acromion fossa betweenthe blade and the acromion process
The posterior expansion of the scapula is distinctive Insome sauropods the posterior part of the scapular blade isexpanded not at all or only slightly for example in Omeiminussaurus Young 1939 (He et al 1988 fig 41) Apatosaurus(Upchurch et al 2004b fig 4) and Rapetosaurus (Fig 9E)In others the ventral margin of the scapular blade is straightor nearly so while the dorsal margin is deflected dorsally tocreate an asymmetric expansion for example in Camaraminussaurus (Fig 9C) and Giraffatitan brancai (Fig 9D) In a fewsauropods however the ventral margin of the blade is alsodeflected ventrally to form a ldquoracquetminusshapedrdquo posterior exminuspansion This is seen in rebbachisaurids and some titanominussaurians eg some specimens of Alamosaurus Gilmore1922 (Gilmore 1946 fig 6) In Brontomerus the posteriorpart of the scapular blade is expanded in a characteristic manminusner the ventral margin is straight except for a posteroventral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 85
50 cm
ventral ldquosteprdquo
acromion expansion
dorsal ldquostepsrdquo
Fig 8 Partial left scapula of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 27761 in lateral view tentatively reconstructed after Giraffatitan brancai HMN Sa 9 (Janensch 1961 pl 15 1)
excursion two thirds of the way along the preserved portionafter which the margin continues parallel to its original traminusjectory so that the excursion appears as a gentle ldquosteprdquo Thedorsal margin is also ldquosteppedrdquo in this manner though withtwo distinct steps rather than one of which the more anterioris most strongly pronounced The net result of these featuresis that the dorsal and ventral borders of the scapula are bothstraight near the posterior extremity and that they are subminusparallel diverging by only about five degrees in the regionjust anterior to the rounded end of the posterior expansionThe step in the ventral border is not known in any othersauropod however the scapula of Neuquensaurus Powell1992 has a stepped dorsal border similar to that of Brontominusmerus (Huene 1929 via Glut 1997 275)
These characters of the scapula must be treated with somecaution however since this bone appears subject to morevariation than any other in the sauropod skeleton see for exminusample the range of shapes in scapulae of Giraffatitan brancai(Janensch 1961 pl 15 1ndash3) and in Camarasaurus supremus(Osborn and Mook 1921 figs 74ndash80)
Two small flat elements OMNH 66431 and 66432 are inminusterpreted as partial sternal plates (Fig 10) The medial edge ofeach is identifiable due to its rugose texture which formed theattachment site for cartilage joining the plates to each otherand to the sternal ribs The sternals are anteroposteriorly elonminusgate and mediolaterally narrow when complete they wereprobably at least three times as long as broad as in ldquoSaltaminussaurusrdquo robustus Huene 1929 (Huene 1929 via McIntosh1990 fig 169L) and proportionally longer than in any othersauropod including Saltasaurus loricatus (McIntosh 1990fig 169 Powell 1992 fig 30) The sternals are crescentic in
shape the anterior and posterior extremities curving laterallyaway from the midline This state was considered a titanominussaurian synapomorphy by Wilson (2002 268) but its distribuminustion is more complex in the current analysis being synapominusmorphic for Neosauropoda with losses in Flagellicaudata andCamarasaurus
86 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
Fig 10 Partial paired sternal plates of the camarasauromorph sauropodBrontomerus mcintoshi gen et sp nov from the Lower Cretaceous CedarMountain Formation of Utah OMNH 66431 and 66432 in ventral view
Fig 9 Scapulocoracoids and scapulae of sauropod dinosaurs scaled to same length of scapular blade from posterior point of glenoid to posterior margin ofblade A Mamenchisaurus youngi Young and Zhao 1972 holotype ZDM0083 left scapulocoracoid modified from Ouyang and Ye (2002 fig 22)B Diplodocus longus Hatcher 1901 USNM 10865 right scapulocoracoid reversed photograph by MPT C Camarasaurus supremus Cope 1877 AMNH5761 Sc 1 left scapula and AMNH 5761 Cor 1 left coracoid probably associated modified from Osborn and Mook (1921 figs 75 81a) D Giraffatitanbrancai (Janensch 1914) HMN Sa 9 left scapula modified from Janensch (1961 pl 15 1) E Rapetosaurus krausi Curry Rogers and Forster 2001holotype FMNH PR 2209 right scapula reversed modified from Curry Rogers (2009 fig 32) F Brontomerus mcintoshi gen et sp nov OMNH 27761left scapula tentatively reconstructed after Giraffatitan brancai
Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

tory Museum London UK CCG Chengdu College of Geminusology Chengdu China CM Carnegie Museum of NaturalHistory Pittsburgh USA DMNH Denver Museum of Natuminusral History Denver USA DNM Dinosaur National Monuminusment Dinosaur USA FMNH Field Museum of NaturalHistory Chicago USA FWMSH Fort Worth Museum ofScience and History Fort Worth USA HMN HumboldtMuseum fuumlr Naturkunde Berlin Germany OMNH Oklaminushoma Museum of Natural History Norman USA ZDMZigong Dinosaur Museum Zigong China
Anatomical nomenclaturemdashWe follow Upchurch et al(2004a) in describing scapulae as though oriented horizonminustally the coracoid articular surface is designated anteriorand the end commonly called ldquodistalrdquo is designated posteminusrior See Taylor (2009 787) for a summary of other schemesthat have been used Names of clades are used as summaminusrized in Table 1
Geological and faunal contextThe holotype and referred specimens described herein werecollected at OMNH locality V857 known as the Hotel MesaQuarry (Kirkland et al 1997 96ndash97 fig 28) One of us(RLC) who had already been working in Lower Cretaceousrocks of the region was notified about the site through thecourtesy of James I Kirkland and was guided to it by BillHawes of Grand Junction Colorado in September 1994 Adminusditional collecting at the site for OMNH was conducted byRandall L Nydam and James I Kirkland in March 1995
OMNH V857 lies on the southwest flank of Hotel Mesain easternmost Grand County Utah (see Kirkland and Madminussen 2007 fig 2) about 13 km northminusnortheast of the juncminustion of the Colorado and Dolores rivers (sec 4 T23S R24E
Dewey 75rsquo quadrangle USGS provisional edition 1985precise locality data are on file at OMNH and are available toqualified investigators upon request) This site had previminusously been worked by private collectors the number idenminustity condition and disposition of the privately collected fosminussils cannot now be determined However given the densityof bone still present and exposed and the fact that the existminusing quarry was already some 5ndash6 m long and 3 m deep it apminuspears that a considerable number of elements were removedfrom the quarry and that the loss of valuable scientific informinusmation has unfortunately been considerable Bones left exminusposed by these previous collectors were in various states ofdisrepair some had been broken and their pieces used to holddown the remnants of a plastic tarpaulin The material reminusported herein was recovered in the course of the two briefOMNH surveysalvage visits to this site
Stratigraphically OMNH V857 lies in a sequence ofLower Cretaceous rocks interposed between the MorrisonFormation (Kimmeridgian) below and the Dakota Formation(Cenomanian) above Westward these rocks are recognizedas the Cedar Mountain Formation eastward the Burro Canminusyon Formation The arbitrary dividing line between these enminustities is generally placed at the Colorado River (Stokes 1952Tschudy et al 1984) which technically places OMNH V857within the Burro Canyon Formation However we will referto the locality as belonging to the more widely recognizedCedar Mountain Formation as it is in this formation thatcomparable specimens are known and the stratigraphy andsedimentology do not change across the arbitrary borderThe most extensive recent applicable stratigraphic work isthat of Kirkland et al (1997 1999) who recognized fivemembers within the Cedar Mountain Formation (in ascendminusing order) the Buckhorn Conglomerate Yellow Cat PoisonStrip Sandstone Ruby Ranch and Mussentuchit members
76 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 1 Clade names used in this study and the definitions used For simplicity specifiers are indicated by genus rather than species in each case thetype species of the genus is intended Nodeminusbased clades are indicated by ldquo+rdquo branchminusbased clades by ldquonotrdquo
Clade name As defined by DefinitionSauropoda Yates (2006 12) Saltasaurus not Melanorosaurus
Neosauropoda Wilson and Sereno (1998 46) Diplodocus + SaltasaurusDiplodocoidea Wilson and Sereno (1998 17) Diplodocus not Saltasaurus
Diplodocimorpha Calvo and Salgado (1995 14) Diplodocus + RebbachisaurusRebbachisauridae Salgado et al (2004 910) Rebbachisaurus not Diplodocus
Diplodocidae Sereno (1998 63) Diplodocus not DicraeosaurusDiplodocinae Taylor and Naish (2005 5) Diplodocus not ApatosaurusMacronaria Wilson and Sereno (1998 49) Saltasaurus not Diplodocus
Camarasauromorpha Upchurch et al (2004a306) Camarasaurus + SaltasaurusCamarasauridae Taylor and Naish (2007 1555) Camarasaurus not Saltasaurus
Titanosauriformes Wilson and Sereno (1998 51) Brachiosaurus + SaltasaurusBrachiosauridae Wilson and Sereno (1998 20ndash21) Brachiosaurus not SaltasaurusSomphospondyli Wilson and Sereno (1998 53) Saltasaurus not Brachiosaurus
Titanosauria Wilson and Upchurch (2003 156) Andesaurus + SaltasaurusLithostrotia Wilson and Upchurch (2003 156) Malawisaurus + Saltasaurus
Saltasauridae Sereno (1998 63) Saltasaurus + OpisthocoelicaudiaOpisthocoelicaudiinae Sereno (1998 63) Opisthocoelicaudia not Saltasaurus
(The Buckhorn Conglomerate Member has a nonminusoverlapminusping geographic distribution with respect to the Yellow Catand Poison Strip Sandstone members and may be a lateralequivalent of one or both see Kirkland et al 1999 figs 1 2)The existing paleontological record from these units ispatchy but there is evidence of three distinct dinosaur faunasfrom the Yellow Cat (Barremian or older) Poison StripSandstone plus Ruby Ranch (AptianndashAlbian) and Mussenminustuchit (AlbianndashCenomanian) members respectively (Kirkminusland et al 1997) While spanning a total of some 30 Ma thefaunal record from the Cedar Mountain Formation is punctuminusated by significant hiatuses (Sames et al 2010) and fossilhorizons that are stratigraphically close may therefore bewidely separated in time
Like most terrigenous Mesozoic units of the Western Inminusterior (see for example Ostrom 1970 14) the laterally equivminusalent Cedar Mountain and Burro Canyon formations areenormously variable At Hotel Mesa the lowest two (Buckminushorn Conglomerate Yellow Cat) and uppermost (Mussenminustuchit) members are absent and only two members of the Ceminusdar Mountain Formation may be recognized the Poison StripSandstone (below) and Ruby Ranch (above) membersStratigraphically OMNH locality V857 lies within the RubyRanch Member a few meters below the local contact withthe Dakota Formation (see Kirkland et al 1999 fig 8) As aworking hypothesis we regard the fossils from the HotelMesa site as being AptianndashAlbian and therefore approximinusmately equivalent in age to assemblages from the CloverlyFormation of Wyoming and Montana and the Trinity Groupof Texas and Oklahoma (eg Ostrom 1970 Winkler et al1990 Jacobs et al 1991 Brinkman et al 1998 Davis et al2008) As elsewhere the Ruby Ranch Member in this area iscomprised predominantly of drab pale green and mauvemudstones with abundant carbonate nodules The fossil horiminuszon which is mainly sandstone with some pebbles and interminusmittent irregular mudstone stringers is about 40ndash50 cmthick Locally these mudstones contain sparse microverteminusbrate fossils often rounded or broken About 100 kg of thismatrix was collected and transported back to the OMNH forfossil extraction using underwater screen washing and assominusciated concentration techniques (Cifelli et al 1996)
Although the microvertebrate assemblage from HotelMesa is neither very diverse nor particularly well repreminussented it does merit attention as being the only such site ofAptianndashAlbian age within the Colorado Plateau A faunal listis presented in Table 2 As is generally the case for such asminussemblages a number of aquatic taxa are represented thoughno remains of Testudines have been recovered yet Probablytwo chondrichthyans are present a hybodontid (possiblyHybodus which was listed in the Ruby Ranch fauna byKirkland et al 1999 table 2) and a polyacrodontid perhapsPolyacrodus The neoceratodontid form genus Ceratoduswidespread in AptianndashAlbian faunas of North America (seereview by Kirkland 1987) is represented by a poorly preminusserved tooth plate two actinopterygians are present one repminusresented by teeth and ganoid scales probably referable to
Lepisosteidae No mammalian or lissamphibian fossils havebeen recovered and Reptilia is represented solely byarchosaurians of which crocodyliforms (three or four taxaincluding a species of the widespread taxon Bernissartia andunidentified species of Goniopholididae and Atoposauridae)are most diverse Megafossils of the Hotel Mesa Quarrymainly represent Sauropoda (including many unidentifiedfragmentary specimens in addition to those describedherein) theropods are known by isolated teeth and rare inminuscomplete postcranial elements The fauna includes anornithopod represented by a single nonminusdiagnostic toothKirkland et al (1999) mentioned the presence in the RubyRanch fauna of Tenontosaurus otherwise known from theCloverly Formation of Wyoming and Montana (Ostrom1970) and the Trinity Group of Texas and Oklahoma (Langminusston 1974 Winkler et al 1997b Brinkman et al 1998)based on specimens from an unidentified site on the westernside of the San Rafael Swell Utah The ornithopod specimenfrom the Hotel Mesa Quarry OMNH 27824 probably does
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 77
Table 2 Vertebrate fauna of the Hotel Mesa Quarry (OMNH V857)Ruby Ranch Member Cedar Mountain Formation (AptianndashAlbian)Grand County Utah
ChondrichthyesCtenacanthiformes
HybodontidaeGen et sp indet
PolyacrodontidaePolyacrodus sp
OsteichthyesOrder indet
Family indetGen et sp indet
LepisosteiformesLepisosteidae
Gen et sp indetDipnoi
NeoceratodontidaeCeratodus sp
ReptiliaCrocodyliformes
Family indetGen et sp indet
BernissartiidaeBernissartia sp
GoniopholididaeGen et sp indet
AtoposauridaeGen et sp indet
TheropodaFamily indet
Gen et sp indetSauropoda
Camarasauromorpha family incertae sedisBrontomerus mcintoshi gen and sp nov
OrnithopodaFamily indet
Gen et sp indet
not represent Tenontosaurus but further identification canminusnot be firmly established at present
Although the quarry has not been worked in severalyears it is not exhausted and may contain additional relevantmaterial (James Kirkland personal communication October2007)
Systematic paleontology
Dinosauria Owen 1842Saurischia Seeley 1888Sauropoda Marsh 1878Neosauropoda Bonaparte 1986Camarasauromorpha Salgado Coria and Calvo 1997Genus Brontomerus novType species Brontomerus mcintoshi sp nov see below
Etymology From Greek bronto thunder Greek meroacutes thigh ldquothunminusderminusthighsrdquo in reference to the substantial femoral musculature impliedby the morphology of the ilium Intended English pronunciation BronminusminustoeminusMEERminusus
DiagnosismdashAs for type and only species (see below)
Brontomerus mcintoshi sp novFigs 1 2 5ndash8 10 12 Table 3
Etymology In honor of veteran sauropod worker John S McIntoshwhose seminal paleontological work done mostly unfunded and on hisown time has been an inspiration to all of us who follow
Holotype OMNH 66430 a left ilium
Tentatively referred material OMNH 66429 crushed presacralcentrum OMNH 61248 midminustominusposterior caudal vertebra OMNH27794 partial distal caudal centrum OMNH 27766 anterior right dorminussal rib OMNH 27761 nearly complete left scapula missing anteriorportion OMNH 66431 and 66432 two partial sternal plates other fragminusments as detailed in Table 3
Type locality Hotel Mesa Quarry (OMNH locality V857) GrandCounty eastern Utah
Type horizon Top of the Ruby Ranch Member of the Cedar MountainFormation (Lower Cretaceous AptianndashAlbian)
DiagnosismdashPreacetabular lobe 55 of total ilium lengthlonger than in any other sauropod preacetabular lobe directedanterolaterally at 30 relative to the sagittal plane but straightin dorsal view and vertically oriented postacetabular lobe reminusduced to near absence ischiadic peduncle reduced to very lowbulge ilium proportionally taller than in any other saurominuspodmdashheight is 52 of total length compared with a maximinusmum of 45 in other sauropods If the tentatively referred eleminus
78 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 3 Material referred to the camarasauromorph sauropod Brontomerus mcintoshi from the Lower Cretaceous Cedar Mountain Formation ofUtah
OMNH specimen number Description Length (cm) Width (cm) Thickness (cm)27761 partial scapula missing anterior part 98 55 527762 large flat rib shaft 101 75 227763 rib fragment 63 8 527764 rib fragment 60 25 227765 rib fragment 58 6 327766 complete pneumatic rib 76 12 427767 dorsal part of rib flattened 26 17 227768 rib fragment 31 6 1527769 fragment of ischium 10 5 2527770 rib fragment 14 35 127771 rib fragment 17 3 227772 rib fragment (in two parts) 13 7 227773 rib fragment 135 4 227773 flat scrap 7 5 0527773 flat scrap 115 9 1
27774ndash27783 fragments27784 collection of 21 fragments
27785ndash27793 fragments27794 partial distal caudal centrum
27795ndash27800 fragments61248 nearly complete midminuscaudal vertebra 11 6 5566429 crushed presacral centrum 14 14 366430 ilium 405 31 866430 dorsal fragment of ilium 14 3 166430 dorsal fragment of ilium 155 9 166431 incomplete sternal plate 15 75 1566432 incomplete sternal plate 115 7 1
ments do belong to the same species as the holotype then thefollowing additional characters also diagnose the new taxonpresacral vertebrae camellate midminustominusposterior caudal verteminusbrae with elongate preminus and postzygapophyseal rami havingthe postzygapophyseal facets hanging below the level of theramus first dorsal rib with expanded dorsally oriented articuminuslar facets laterally curving shaft and ventrally directed pneuminusmatic foramen in head acromion expansion of scapula prominusnounced and steep but not forming acromion fossa dorsal andventral margins of scapular blade ldquosteppedrdquo sternal platescrescentic and three times as long as broad
Unambiguous autapomorphies distinguishing Brontomeminusrus from the root of the polytomy in which it is recovered inthe strict consensus of most parsimonious trees in the phylominusgenetic analysis below character 184 ratio of centrumlengthheight in middle caudal vertebrae 20 185 sharpridge on lateral surface of middle caudal centra at archminusbodyjunction absent 212 posterior end of scapular body racminusquetminusshaped (dorsoventrally expanded) 261 in lateral viewthe most anteroventral point on the iliac preacetabular lobe isalso the most anterior point (preacetabular lobe is pointed)264 projected line connecting articular surfaces of ischiadicand pubic peduncles of ilium passes ventral to ventral marginof postacetabular lobe of ilium
Description
This taxon is based on a collection of elements all from thesame quarry all of them consistent with assignment to a sinminusgle taxon (Fig 1) However the elements were not found arminusticulated and their differing sizes do not permit interpretaminustion as belonging to a single individual For example the parminus
tial scapula is 98 cm long Reconstruction after the scapula ofGiraffatitan brancai (Janensch 1914) suggests that the comminusplete element was about 121 cm long In Rapetosaurus CurryRogers and Forster 2001 the scapula and ilium are about thesame length (Curry Rogers and Forster 2001 fig 3) but theilium of Brontomerus is only one third the reconstructedlength of the scapula The quarry therefore contains at leasttwo individuals of widely differing sizes The smaller prominusviding the holotype ilium and the referred presacral centrumand sternal plates is interpreted as a juvenile the remainingelements probably belonged to the larger mature individual
Among the sauropod elements recovered from the HotelMesa site at least three (the caudal vertebra scapula andilium) have characters not known in any other sauropodSince it is not possible to conclusively demonstrate that allthree elements belong to the same taxon (and size differminusences preclude their belonging to a single individual) it istherefore possible that the quarry contains as many as threenew taxa However since all the informative elements havecharacters that indicate a titanosauriform identity we conminusservatively consider it more likely that only a single newtaxon is present Groups of sauropods of mixed ages areknown for several taxa (eg Bird 1985 Coria 1994 Bader2003) so there is a precedent for finding adults and juveminusniles together Although we diagnose the new taxon solelyon the type ilium we also tentatively refer the other eleminusments to this taxon and note the additional characters thatpertain if this referral is correct we hope that subsequentwork in the Hotel Mesa Quarry or elsewhere in the CedarMountain Formation will yield specimens that allow us toconfirm or refute this referral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 79
Fig 1 Skeletal inventory of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah in left lateral view Preserved elements are white missing elements are reconstructed in gray After a Camarasaurus grandis reconstructionkindly provided by Scott Hartman
The assignment of specimen numbers to the material deminusscribed here is complex (Table 3) Specimen number OMNH27773 comprises three elements OMNH 27784 consists of 21small fragments of bone none of them informative all otherelements have their own specimen numbers in the range27761ndash27800 apart from the midminuscaudal vertebra OMNH61248 and the reassigned numbers 66429ndash66432 for eleminusments extracted from OMNH 27773
IliummdashThe most informative element is OMNH 66430 a leftilium (Fig 2) which has therefore been selected as the holominustype The ilium was preserved complete but lay hidden beminusneath the scapula and so was damaged in the field (JamesKirkland personal communication March 2008) The iliumis preserved in three parts one provides most of the boneincluding the well preserved preacetabular lobe pubic andischiadic peduncles and acetabular margin and the other two
provide most of the dorsal margin giving a good indication ofthe degree of curvature The relative positions and orientationof the two smaller fragments can not be definitely ascertainedbut they appear to be parts of a single large fragment broken atthe point where we have reconstructed them as touching andif this interpretation is correct then the curvature of the pair inminusdicates which side must be oriented laterally
The ilium is remarkable in that the preacetabular lobe isrelatively larger than in any other sauropod (Fig 3 Table 4measurement protocol is illustrated in Fig 4) the postaceminustabular lobe is reduced almost to the point of absence and theilium is proportionally taller than in any other sauropod Theischiadic peduncle is reduced to a very low ventral projectionfrom almost the most posterior point of the ilium The near abminussence of the ischiadic peduncle cannot be attributed to damageas the iliac articular surface is preserved Immediately posterominus
80 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
acetabulum
pubicpeduncle
10 cm
ischiadicpeduncle
pre-acetabularblade
Fig 2 Left ilium of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah type specimen OMNH 66430 in lateral view reconstructed from the three fragments (A) and ventral view (B)
dorsal to this surface is a subtle notch between the peduncleand the very reduced postacetabular lobe This notch and theareas either side of it are composed of finished bone demonminusstrating that the great reduction of the postacetabular lobe toois a genuine osteological feature and not due to damage In reminusgard to the proportionally large preacetabular lobe the iliumof Brontomerus resembles that of Rapetosaurus (Fig 3E Taminusble 4) However that taxon has a normal postacetabular lobeand is not proportionally tall In overall proportions the iliumof Brontomerus is more similar to the left ilium HMN J1 asminussigned to Giraffatitan brancai (Janensch 1961 pl E 2) whichalso has a reduced postacetabular lobe mdashsee also Fig 3DHowever the ilium of Brontomerus is proportionally tallerthan that of G brancai and its anterior margin comes to apoint rather than being smoothly rounded as in that taxon InBrontomerus the maximum height of the ilium above theacetabular margin when measured perpendicular to the lonminusgest axis is 52 in other sauropods examined this proportiondoes not exceed 45 and averages 40 (Table 4)
As with many sauropods the preacetabular lobe of theilium flares laterally However in most sauropods this flarminusing is progressive so that in dorsal or ventral view the mostposterior part of the preacetabular lobe is nearly parallel witha line drawn between the pubic and ischiadic peduncles andsmooth lateral curvature inclines the more anterior parts inminuscreasingly laterally so that the more anterior part is almost atright angles to this line and the ilium appears smoothly
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 81
Fig 3 Ilia of sauropod dinosaurs scaled to same total length A Mamenchisaurus hochuanensis Young and Zhao 1972 holotype CCG V 20401 right iliumreversed modified from Young and Zhao (1972 pl 6 1a) B Diplodocus carnegii Hatcher 1901 CM 94 right ilium reversed modified from Hatcher (1901pl 10 1) C Camarasaurus supremus Cope 1877 AMNH 5761 Il 1 left ilium modified from Osborn and Mook (1921 fig 87) D Giraffatitan brancai(Janensch 1914) HMN J1 left ilium modified from Janensch (1961 pl E 2) E Rapetosaurus krausi Curry Rogers and Forster 2001 holotype FMNH PR2209 left ilium modified from Curry Rogers (2009 fig 39B) F Brontomerus mcintoshi gen et sp nov holotype OMNH 66430 left ilium
Fig 4 Measurement protocol for sauropod ilia as illustrated in Fig 3 andshown in Table 4 Total length is measured along the longest axis of theilium lengths of preacetabular and postacetabular lobes are measured parminusallel to this axis and extend from the extremity of the lobe to the anteriormargin of the pubic peduncle and posterior margin of the ischiadic pedunclerespectively Supracetabular height is measured perpendicular to the lonminusgest axis and extends from the highest point of the acetabulum to the pointlevel with the highest part of the ilium
curved in dorsal or ventral viewmdashfor example ApatosaurusMarsh 1877 (Upchurch et al 2004b pl 4 D E) Haplominuscanthosaurus Hatcher 1903b (Hatcher 1903a pl 5 1) andSaltasaurus Bonaparte and Powell 1980 (Powell 1992 fig17) In Brontomerus by contrast the blade of the ilium apminuspears to be ldquohingedrdquomdashdeflected laterally at a point directlyanterior to the public pedunclemdashso that the preacetabularlobe is straight in dorsal or ventral view and directed anterominuslaterally by an angle of about 30 to the sagittal In this reminusspect it more closely resembles the ilium of Camarasaurus
Cope 1877 (Osborn and Mook 1921 fig 49) although it difminusfers in other respects
The Brontomerus ilium is laterally compressed and unlikemost sauropod ilia the dorsal margin is not deflected laterallyrelative to the more ventral part so that in ventral view it apminuspears very thin (Fig 2B) It is well established that the longbones of sauropods grow isometrically through ontogeny(Carpenter and McIntosh 1994 Wilhite 1999 2003 Ikejiri etal 2005 Tidwell and Wilhite 2005 Taylor 2009) while theirvertebrae undergo significant changes in proportions laminaminus
82 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 4 Relative measurements of ilia in sauropods Measurement protocol is illustrated in Fig 4
Taxon Specimen ReferenceTotallength(cm)
Length ofpreacetabular
lobe (cmproportion oftotal length)
Length ofpostacetabular
lobe (cmproportion oftotal length)
Supracetabularheight (cm
proportion oftotal length)
Mamenchisaurushochuanensis
CCG V 20401 Young and Zhao (1972 pl 6 1a) 102 39 38 18 18 41 40
Diplodocus carnegii CM 94 Hatcher (1901 pl 10 1) 1091 41 38 20 18 49 45Camarasaurus
supremusAMNH 5761 Il 1 Osborn and Mook (1921 fig 87) 115 36 31 19 17 48 42
Giraffatitan brancai2
HMN Aa 13Janensch (1961 pl E 1a) 119 47 39 21 18 46 39
Measured by SchwarzminusWings(personal communication 2009) 1143 535 47 299 26
HMN J1Janensch (1961 pl E 2) 1055 55 52 16 15 37 35
Measured by SchwarzminusWings(personal communication 2009) 1077 528 49 135 13
Rapetosaurus krausi FMNH PR 2209 Curry Rogers (2009 fig 39) 42 20 48 7 17 16 38Brontomerus mcintoshi OMNH 66430 (this study) 405 223 55 0 0 21 52
1 Hatcher (1901) did not state the length of the ilium of CM 94 and did not figure that of CM 84 We have assumed that the ilium of the figured specimen CM94 is the same size as that of CM 84 (Hatcher 1901 46) and calculated the proportions from the figured specimen
2 Janenschrsquos (1961 pl E) figures of the two ilia of Giraffatitan brancai are not executed from an orthogonal perspective the ilium of HMN Aa 13 is illusminustrated from a slightly anterolateral position foreshortening the preacetabular lobe and that of HMN J 1 from a slight posterolateral position foreshortenminusing the postacetabular lobe The true lengths of the two lobes are probably somewhere between the two percentages calculated from the figures about45 for the former and 16 for the latter
10
cm
Fig 5 Damaged presacral vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah OMNH 66429 in dorsal view as photograph (A) and interpretive drawing (B) Shading indicates air spaces
tion pneumatic excavations and neurocentral fusion (Wedel2003a 248 b 352ndash354) Ontogenetic changes in limbminusgirdleelements such as the ilium are less well understood due to apaucity of sufficiently well preserved specimens (Ray Wilhitepersonal communication October 2007) Therefore the lateralcompression of the Brontomerus ilium may be a juvenile charminusacter with the ilium thickening through ontogeny to supportthe growing weight of the animal or it may in fact be phylominusgenetically significant
Presacral centrummdashA single presacral centrum OMNH66429 was recovered (Fig 5) Unfortunately preservation isvery poor the neural arch and all processes have been lostand the centrum has been greatly crushed dorsoventrally sothat the remaining part is essentially flat the element is 14
cm in both anteroposterior length and transverse width butno more than 3 cm in dorsoventral depth The small size indiminuscates that this element belonged to a juvenile The internalstructure of the centrum is visible however and consists offine septa dividing a hollow internal space irregularly intomany small camellae This morphology is characteristic oftitanosauriforms (Wedel 2003b 354ndash355) Highly camellateinternal structure has not previously been observed in juveminusnile sauropod vertebrae but this may be due to samplingbias so far all juvenile sauropod vertebrae that have beenstudied for internal structure have been those of Camaraminussaurus and diplodocoids which follow an ontogenetic trajecminustory in which large shallow lateral fossae develop into cameminusrae from which smaller accessory camerae and camellae deminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 83
prezygapophysis
neural spine
postzygapophysealramus
postzygapophysealfacet
chevron facets
5 cm
Fig 6 Midminuscaudal vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 61248 in dorsal (A) anterior (B) left lateral (C) posterior (D) and ventral (E) views
velop (Wedel et al 2000a fig 11 Wedel 2003b 349) TheHotel Mesa presacral suggests that camellate vertebrae mayhave developed differently in titanosauriforms possibly byinminussitu formation of camellae during pneumatization as ocminuscurs in the camellate vertebrae of birds (personal observaminustion MJW)
Caudal vertebraemdashOMNH 61248 is a distinctive caudalvertebra with elongated preminus and postzygapophyseal rami(Fig 6) Apart from the tip of the right prezygapophysis theelement is complete and well preserved While the centrum isonly 11 cm in length the distance from the prezygapophysisto postzygapophysis is 145 cm The centrum is slightlybroader than tall (6 cm compared with 55 cm anteriorly 65cm compared with 5 cm posteriorly) and gently waisted Theneural arch is set forward on the centrum but does not reachthe anterior margin The neural spine is so reduced and sostrongly inclined posteriorly as to be all but indistinguishminusable and is apparent only as a very low eminence abovethe postzygapophyses The postzygapophyseal facets themminusselves are set on the posterolateral faces of a low process thathangs below the main postzygapophyseal ramus Chevronfacets are weakly present on the posterior margin of the venminustral surface of the centrum but not on the anterior marginThe elongation index of 22 indicates a midminustominusposterior pominussition in the caudal sequence for this element as similarcentrum proportions do not appear until about caudal 30 inGiraffatitan brancai (Janensch 1950 pl 3)
This vertebra most closely resembles the indeterminatesauropod vertebra BMNH 27500 from the Wessex Formationof the Isle of Wight figured by Naish and Martill (2001 pl33) The Barremian age of that specimen places it about 15 Ma
earlier than OMNH 61248 Its neural arch is less elevated thanthat of the Hotel Mesa specimen its postzygapophyses projectyet farther posteriorly and its prezygapophyses less far anteriminusorly and it is very mildly biconvex rather than procoelous butin other respects including absolute size it is a good match forthe Brontomerus caudal
Also included in the Hotel Mesa material is OMNH27794 a partial distal caudal centrum figured by Wedel(2005 fig 77) This centrum is approximately round incrossminussection about 4 cm in diameter and internally consistsof apneumatic cancellous bone
RibsmdashThe Hotel Mesa material contains several dorsal ribsin various states of preservation but no readily identifiablecervical ribs The dorsal rib elements include the shaft of alarge flat rib (OMNH 27762) portions of several smaller ribshafts (OMNH 27763ndash27765 27768 and others) a flattenedrib head (OMNH 27767) and most informatively a smallcomplete rib (OMNH 27766 Fig 7) Despite its excellentpreservation and apparent lack of distortion this element isdifficult to interpret Its shaft is straight for almost its wholelength and both articular facets are directed dorsally ratherthan being inclined medially The tuberculum is directly inline with the main part of the shaft of the rib and thecapitulum is at an angle of about 30 to it In these respectsthe rib resembles the most anterior dorsal rib of the Diplominusdocus carnegii holotype CM 84 (personal observation MPTthis element was not figured by Hatcher [1901]) We thereminusfore interpret this rib as having probably belonged to the rightside of the first dorsal vertebra
The rib is unusual in other respects however most notablythat the ventral part of the shaft curves laterally rather than meminus
84 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
10 cm
capitulum
pneumatic fossa
tuberculum
Fig 7 First right dorsal rib of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah OMNH 27766 in posterior view head of rib showing pneumatic invasion of shaft (A) and complete rib showing laterally directed curvature of shaft (B)
dially Careful inspection of the bone reveals no indication ofdistortion or of incorrect reconstruction It may be possiblethat in life the thorax was transversely compressed so that thedorsal part of the rib shaft was directed ventromedially and themore ventral part was vertical Both articular facets are subminuscircular in dorsal view and significantly expanded comparedwith the rami that bear them On the anterior face of the head alow ridge arises just below the capitulum and extends downthe medial edge of the rib for about 40 of its length
The head of the rib is also unusual in that a thin sheet ofbone connects the rami that support the articular facets andthis sheet extends much farther proximally than in mostsauropod ribs Its precise extent cannot be ascertained due tobreakage The sheet of bone is perforated close to the capituminuslar ramus and from this perforation a pneumatic cavity inminusvades the shaft of the rib extending ventrally from within ashallow fossa in the posterior face Pneumatization of thedorsal ribs is a synapomorphy of Titanosauriformes (Wilsonand Sereno 1998) although pneumatic dorsal ribs are also inminusfrequently present in diplodocids (Gilmore 1936 Lovelaceet al 2003 2008)
Pectoral girdlemdashOMNH 27761 is a partial scapula consistminusing of the blade and part of the anterior expansion but missminusing the glenoid region and the remainder of the anterior exminuspansion (Fig 8) As preserved the element is nearly flat butthis may be due to postminusmortem distortion and in any casethe most strongly curved part of most sauropod scapulae isthe anterior part that is missing from this specimen The genminustle curvature preserved in the posterior part of the blade indiminuscates that the element was from the left side The bone is surminusprisingly thin in all preserved parts never exceeding a few
cm in contrast to for example the scapula of Camarasaurussupremus which is thick even in midminusblade (Osborn andMook 1921 fig 74b) This suggests that the glenoid thickenminusing and the acromial ridge may have been located some disminustance anterior to the preserved portion and a reconstructionafter the proportions of Giraffatitan brancai (Fig 8) suggeststhat about 80 of the scapularsquos full length is preserved Theposterior part of the acromion expansion is preserved howminusever and is sufficient to show that this expansion was prominusnounced so that the maximum dorsoventral height of thescapula was more than two and half times its minimumheight at the midpoint of the blade The dorsal margin slopesup towards the anterior expansion rather than forming a posminusteriorly directed ldquohookrdquo or a distinct acromion fossa betweenthe blade and the acromion process
The posterior expansion of the scapula is distinctive Insome sauropods the posterior part of the scapular blade isexpanded not at all or only slightly for example in Omeiminussaurus Young 1939 (He et al 1988 fig 41) Apatosaurus(Upchurch et al 2004b fig 4) and Rapetosaurus (Fig 9E)In others the ventral margin of the scapular blade is straightor nearly so while the dorsal margin is deflected dorsally tocreate an asymmetric expansion for example in Camaraminussaurus (Fig 9C) and Giraffatitan brancai (Fig 9D) In a fewsauropods however the ventral margin of the blade is alsodeflected ventrally to form a ldquoracquetminusshapedrdquo posterior exminuspansion This is seen in rebbachisaurids and some titanominussaurians eg some specimens of Alamosaurus Gilmore1922 (Gilmore 1946 fig 6) In Brontomerus the posteriorpart of the scapular blade is expanded in a characteristic manminusner the ventral margin is straight except for a posteroventral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 85
50 cm
ventral ldquosteprdquo
acromion expansion
dorsal ldquostepsrdquo
Fig 8 Partial left scapula of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 27761 in lateral view tentatively reconstructed after Giraffatitan brancai HMN Sa 9 (Janensch 1961 pl 15 1)
excursion two thirds of the way along the preserved portionafter which the margin continues parallel to its original traminusjectory so that the excursion appears as a gentle ldquosteprdquo Thedorsal margin is also ldquosteppedrdquo in this manner though withtwo distinct steps rather than one of which the more anterioris most strongly pronounced The net result of these featuresis that the dorsal and ventral borders of the scapula are bothstraight near the posterior extremity and that they are subminusparallel diverging by only about five degrees in the regionjust anterior to the rounded end of the posterior expansionThe step in the ventral border is not known in any othersauropod however the scapula of Neuquensaurus Powell1992 has a stepped dorsal border similar to that of Brontominusmerus (Huene 1929 via Glut 1997 275)
These characters of the scapula must be treated with somecaution however since this bone appears subject to morevariation than any other in the sauropod skeleton see for exminusample the range of shapes in scapulae of Giraffatitan brancai(Janensch 1961 pl 15 1ndash3) and in Camarasaurus supremus(Osborn and Mook 1921 figs 74ndash80)
Two small flat elements OMNH 66431 and 66432 are inminusterpreted as partial sternal plates (Fig 10) The medial edge ofeach is identifiable due to its rugose texture which formed theattachment site for cartilage joining the plates to each otherand to the sternal ribs The sternals are anteroposteriorly elonminusgate and mediolaterally narrow when complete they wereprobably at least three times as long as broad as in ldquoSaltaminussaurusrdquo robustus Huene 1929 (Huene 1929 via McIntosh1990 fig 169L) and proportionally longer than in any othersauropod including Saltasaurus loricatus (McIntosh 1990fig 169 Powell 1992 fig 30) The sternals are crescentic in
shape the anterior and posterior extremities curving laterallyaway from the midline This state was considered a titanominussaurian synapomorphy by Wilson (2002 268) but its distribuminustion is more complex in the current analysis being synapominusmorphic for Neosauropoda with losses in Flagellicaudata andCamarasaurus
86 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
Fig 10 Partial paired sternal plates of the camarasauromorph sauropodBrontomerus mcintoshi gen et sp nov from the Lower Cretaceous CedarMountain Formation of Utah OMNH 66431 and 66432 in ventral view
Fig 9 Scapulocoracoids and scapulae of sauropod dinosaurs scaled to same length of scapular blade from posterior point of glenoid to posterior margin ofblade A Mamenchisaurus youngi Young and Zhao 1972 holotype ZDM0083 left scapulocoracoid modified from Ouyang and Ye (2002 fig 22)B Diplodocus longus Hatcher 1901 USNM 10865 right scapulocoracoid reversed photograph by MPT C Camarasaurus supremus Cope 1877 AMNH5761 Sc 1 left scapula and AMNH 5761 Cor 1 left coracoid probably associated modified from Osborn and Mook (1921 figs 75 81a) D Giraffatitanbrancai (Janensch 1914) HMN Sa 9 left scapula modified from Janensch (1961 pl 15 1) E Rapetosaurus krausi Curry Rogers and Forster 2001holotype FMNH PR 2209 right scapula reversed modified from Curry Rogers (2009 fig 32) F Brontomerus mcintoshi gen et sp nov OMNH 27761left scapula tentatively reconstructed after Giraffatitan brancai
Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

(The Buckhorn Conglomerate Member has a nonminusoverlapminusping geographic distribution with respect to the Yellow Catand Poison Strip Sandstone members and may be a lateralequivalent of one or both see Kirkland et al 1999 figs 1 2)The existing paleontological record from these units ispatchy but there is evidence of three distinct dinosaur faunasfrom the Yellow Cat (Barremian or older) Poison StripSandstone plus Ruby Ranch (AptianndashAlbian) and Mussenminustuchit (AlbianndashCenomanian) members respectively (Kirkminusland et al 1997) While spanning a total of some 30 Ma thefaunal record from the Cedar Mountain Formation is punctuminusated by significant hiatuses (Sames et al 2010) and fossilhorizons that are stratigraphically close may therefore bewidely separated in time
Like most terrigenous Mesozoic units of the Western Inminusterior (see for example Ostrom 1970 14) the laterally equivminusalent Cedar Mountain and Burro Canyon formations areenormously variable At Hotel Mesa the lowest two (Buckminushorn Conglomerate Yellow Cat) and uppermost (Mussenminustuchit) members are absent and only two members of the Ceminusdar Mountain Formation may be recognized the Poison StripSandstone (below) and Ruby Ranch (above) membersStratigraphically OMNH locality V857 lies within the RubyRanch Member a few meters below the local contact withthe Dakota Formation (see Kirkland et al 1999 fig 8) As aworking hypothesis we regard the fossils from the HotelMesa site as being AptianndashAlbian and therefore approximinusmately equivalent in age to assemblages from the CloverlyFormation of Wyoming and Montana and the Trinity Groupof Texas and Oklahoma (eg Ostrom 1970 Winkler et al1990 Jacobs et al 1991 Brinkman et al 1998 Davis et al2008) As elsewhere the Ruby Ranch Member in this area iscomprised predominantly of drab pale green and mauvemudstones with abundant carbonate nodules The fossil horiminuszon which is mainly sandstone with some pebbles and interminusmittent irregular mudstone stringers is about 40ndash50 cmthick Locally these mudstones contain sparse microverteminusbrate fossils often rounded or broken About 100 kg of thismatrix was collected and transported back to the OMNH forfossil extraction using underwater screen washing and assominusciated concentration techniques (Cifelli et al 1996)
Although the microvertebrate assemblage from HotelMesa is neither very diverse nor particularly well repreminussented it does merit attention as being the only such site ofAptianndashAlbian age within the Colorado Plateau A faunal listis presented in Table 2 As is generally the case for such asminussemblages a number of aquatic taxa are represented thoughno remains of Testudines have been recovered yet Probablytwo chondrichthyans are present a hybodontid (possiblyHybodus which was listed in the Ruby Ranch fauna byKirkland et al 1999 table 2) and a polyacrodontid perhapsPolyacrodus The neoceratodontid form genus Ceratoduswidespread in AptianndashAlbian faunas of North America (seereview by Kirkland 1987) is represented by a poorly preminusserved tooth plate two actinopterygians are present one repminusresented by teeth and ganoid scales probably referable to
Lepisosteidae No mammalian or lissamphibian fossils havebeen recovered and Reptilia is represented solely byarchosaurians of which crocodyliforms (three or four taxaincluding a species of the widespread taxon Bernissartia andunidentified species of Goniopholididae and Atoposauridae)are most diverse Megafossils of the Hotel Mesa Quarrymainly represent Sauropoda (including many unidentifiedfragmentary specimens in addition to those describedherein) theropods are known by isolated teeth and rare inminuscomplete postcranial elements The fauna includes anornithopod represented by a single nonminusdiagnostic toothKirkland et al (1999) mentioned the presence in the RubyRanch fauna of Tenontosaurus otherwise known from theCloverly Formation of Wyoming and Montana (Ostrom1970) and the Trinity Group of Texas and Oklahoma (Langminusston 1974 Winkler et al 1997b Brinkman et al 1998)based on specimens from an unidentified site on the westernside of the San Rafael Swell Utah The ornithopod specimenfrom the Hotel Mesa Quarry OMNH 27824 probably does
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 77
Table 2 Vertebrate fauna of the Hotel Mesa Quarry (OMNH V857)Ruby Ranch Member Cedar Mountain Formation (AptianndashAlbian)Grand County Utah
ChondrichthyesCtenacanthiformes
HybodontidaeGen et sp indet
PolyacrodontidaePolyacrodus sp
OsteichthyesOrder indet
Family indetGen et sp indet
LepisosteiformesLepisosteidae
Gen et sp indetDipnoi
NeoceratodontidaeCeratodus sp
ReptiliaCrocodyliformes
Family indetGen et sp indet
BernissartiidaeBernissartia sp
GoniopholididaeGen et sp indet
AtoposauridaeGen et sp indet
TheropodaFamily indet
Gen et sp indetSauropoda
Camarasauromorpha family incertae sedisBrontomerus mcintoshi gen and sp nov
OrnithopodaFamily indet
Gen et sp indet
not represent Tenontosaurus but further identification canminusnot be firmly established at present
Although the quarry has not been worked in severalyears it is not exhausted and may contain additional relevantmaterial (James Kirkland personal communication October2007)
Systematic paleontology
Dinosauria Owen 1842Saurischia Seeley 1888Sauropoda Marsh 1878Neosauropoda Bonaparte 1986Camarasauromorpha Salgado Coria and Calvo 1997Genus Brontomerus novType species Brontomerus mcintoshi sp nov see below
Etymology From Greek bronto thunder Greek meroacutes thigh ldquothunminusderminusthighsrdquo in reference to the substantial femoral musculature impliedby the morphology of the ilium Intended English pronunciation BronminusminustoeminusMEERminusus
DiagnosismdashAs for type and only species (see below)
Brontomerus mcintoshi sp novFigs 1 2 5ndash8 10 12 Table 3
Etymology In honor of veteran sauropod worker John S McIntoshwhose seminal paleontological work done mostly unfunded and on hisown time has been an inspiration to all of us who follow
Holotype OMNH 66430 a left ilium
Tentatively referred material OMNH 66429 crushed presacralcentrum OMNH 61248 midminustominusposterior caudal vertebra OMNH27794 partial distal caudal centrum OMNH 27766 anterior right dorminussal rib OMNH 27761 nearly complete left scapula missing anteriorportion OMNH 66431 and 66432 two partial sternal plates other fragminusments as detailed in Table 3
Type locality Hotel Mesa Quarry (OMNH locality V857) GrandCounty eastern Utah
Type horizon Top of the Ruby Ranch Member of the Cedar MountainFormation (Lower Cretaceous AptianndashAlbian)
DiagnosismdashPreacetabular lobe 55 of total ilium lengthlonger than in any other sauropod preacetabular lobe directedanterolaterally at 30 relative to the sagittal plane but straightin dorsal view and vertically oriented postacetabular lobe reminusduced to near absence ischiadic peduncle reduced to very lowbulge ilium proportionally taller than in any other saurominuspodmdashheight is 52 of total length compared with a maximinusmum of 45 in other sauropods If the tentatively referred eleminus
78 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 3 Material referred to the camarasauromorph sauropod Brontomerus mcintoshi from the Lower Cretaceous Cedar Mountain Formation ofUtah
OMNH specimen number Description Length (cm) Width (cm) Thickness (cm)27761 partial scapula missing anterior part 98 55 527762 large flat rib shaft 101 75 227763 rib fragment 63 8 527764 rib fragment 60 25 227765 rib fragment 58 6 327766 complete pneumatic rib 76 12 427767 dorsal part of rib flattened 26 17 227768 rib fragment 31 6 1527769 fragment of ischium 10 5 2527770 rib fragment 14 35 127771 rib fragment 17 3 227772 rib fragment (in two parts) 13 7 227773 rib fragment 135 4 227773 flat scrap 7 5 0527773 flat scrap 115 9 1
27774ndash27783 fragments27784 collection of 21 fragments
27785ndash27793 fragments27794 partial distal caudal centrum
27795ndash27800 fragments61248 nearly complete midminuscaudal vertebra 11 6 5566429 crushed presacral centrum 14 14 366430 ilium 405 31 866430 dorsal fragment of ilium 14 3 166430 dorsal fragment of ilium 155 9 166431 incomplete sternal plate 15 75 1566432 incomplete sternal plate 115 7 1
ments do belong to the same species as the holotype then thefollowing additional characters also diagnose the new taxonpresacral vertebrae camellate midminustominusposterior caudal verteminusbrae with elongate preminus and postzygapophyseal rami havingthe postzygapophyseal facets hanging below the level of theramus first dorsal rib with expanded dorsally oriented articuminuslar facets laterally curving shaft and ventrally directed pneuminusmatic foramen in head acromion expansion of scapula prominusnounced and steep but not forming acromion fossa dorsal andventral margins of scapular blade ldquosteppedrdquo sternal platescrescentic and three times as long as broad
Unambiguous autapomorphies distinguishing Brontomeminusrus from the root of the polytomy in which it is recovered inthe strict consensus of most parsimonious trees in the phylominusgenetic analysis below character 184 ratio of centrumlengthheight in middle caudal vertebrae 20 185 sharpridge on lateral surface of middle caudal centra at archminusbodyjunction absent 212 posterior end of scapular body racminusquetminusshaped (dorsoventrally expanded) 261 in lateral viewthe most anteroventral point on the iliac preacetabular lobe isalso the most anterior point (preacetabular lobe is pointed)264 projected line connecting articular surfaces of ischiadicand pubic peduncles of ilium passes ventral to ventral marginof postacetabular lobe of ilium
Description
This taxon is based on a collection of elements all from thesame quarry all of them consistent with assignment to a sinminusgle taxon (Fig 1) However the elements were not found arminusticulated and their differing sizes do not permit interpretaminustion as belonging to a single individual For example the parminus
tial scapula is 98 cm long Reconstruction after the scapula ofGiraffatitan brancai (Janensch 1914) suggests that the comminusplete element was about 121 cm long In Rapetosaurus CurryRogers and Forster 2001 the scapula and ilium are about thesame length (Curry Rogers and Forster 2001 fig 3) but theilium of Brontomerus is only one third the reconstructedlength of the scapula The quarry therefore contains at leasttwo individuals of widely differing sizes The smaller prominusviding the holotype ilium and the referred presacral centrumand sternal plates is interpreted as a juvenile the remainingelements probably belonged to the larger mature individual
Among the sauropod elements recovered from the HotelMesa site at least three (the caudal vertebra scapula andilium) have characters not known in any other sauropodSince it is not possible to conclusively demonstrate that allthree elements belong to the same taxon (and size differminusences preclude their belonging to a single individual) it istherefore possible that the quarry contains as many as threenew taxa However since all the informative elements havecharacters that indicate a titanosauriform identity we conminusservatively consider it more likely that only a single newtaxon is present Groups of sauropods of mixed ages areknown for several taxa (eg Bird 1985 Coria 1994 Bader2003) so there is a precedent for finding adults and juveminusniles together Although we diagnose the new taxon solelyon the type ilium we also tentatively refer the other eleminusments to this taxon and note the additional characters thatpertain if this referral is correct we hope that subsequentwork in the Hotel Mesa Quarry or elsewhere in the CedarMountain Formation will yield specimens that allow us toconfirm or refute this referral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 79
Fig 1 Skeletal inventory of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah in left lateral view Preserved elements are white missing elements are reconstructed in gray After a Camarasaurus grandis reconstructionkindly provided by Scott Hartman
The assignment of specimen numbers to the material deminusscribed here is complex (Table 3) Specimen number OMNH27773 comprises three elements OMNH 27784 consists of 21small fragments of bone none of them informative all otherelements have their own specimen numbers in the range27761ndash27800 apart from the midminuscaudal vertebra OMNH61248 and the reassigned numbers 66429ndash66432 for eleminusments extracted from OMNH 27773
IliummdashThe most informative element is OMNH 66430 a leftilium (Fig 2) which has therefore been selected as the holominustype The ilium was preserved complete but lay hidden beminusneath the scapula and so was damaged in the field (JamesKirkland personal communication March 2008) The iliumis preserved in three parts one provides most of the boneincluding the well preserved preacetabular lobe pubic andischiadic peduncles and acetabular margin and the other two
provide most of the dorsal margin giving a good indication ofthe degree of curvature The relative positions and orientationof the two smaller fragments can not be definitely ascertainedbut they appear to be parts of a single large fragment broken atthe point where we have reconstructed them as touching andif this interpretation is correct then the curvature of the pair inminusdicates which side must be oriented laterally
The ilium is remarkable in that the preacetabular lobe isrelatively larger than in any other sauropod (Fig 3 Table 4measurement protocol is illustrated in Fig 4) the postaceminustabular lobe is reduced almost to the point of absence and theilium is proportionally taller than in any other sauropod Theischiadic peduncle is reduced to a very low ventral projectionfrom almost the most posterior point of the ilium The near abminussence of the ischiadic peduncle cannot be attributed to damageas the iliac articular surface is preserved Immediately posterominus
80 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
acetabulum
pubicpeduncle
10 cm
ischiadicpeduncle
pre-acetabularblade
Fig 2 Left ilium of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah type specimen OMNH 66430 in lateral view reconstructed from the three fragments (A) and ventral view (B)
dorsal to this surface is a subtle notch between the peduncleand the very reduced postacetabular lobe This notch and theareas either side of it are composed of finished bone demonminusstrating that the great reduction of the postacetabular lobe toois a genuine osteological feature and not due to damage In reminusgard to the proportionally large preacetabular lobe the iliumof Brontomerus resembles that of Rapetosaurus (Fig 3E Taminusble 4) However that taxon has a normal postacetabular lobeand is not proportionally tall In overall proportions the iliumof Brontomerus is more similar to the left ilium HMN J1 asminussigned to Giraffatitan brancai (Janensch 1961 pl E 2) whichalso has a reduced postacetabular lobe mdashsee also Fig 3DHowever the ilium of Brontomerus is proportionally tallerthan that of G brancai and its anterior margin comes to apoint rather than being smoothly rounded as in that taxon InBrontomerus the maximum height of the ilium above theacetabular margin when measured perpendicular to the lonminusgest axis is 52 in other sauropods examined this proportiondoes not exceed 45 and averages 40 (Table 4)
As with many sauropods the preacetabular lobe of theilium flares laterally However in most sauropods this flarminusing is progressive so that in dorsal or ventral view the mostposterior part of the preacetabular lobe is nearly parallel witha line drawn between the pubic and ischiadic peduncles andsmooth lateral curvature inclines the more anterior parts inminuscreasingly laterally so that the more anterior part is almost atright angles to this line and the ilium appears smoothly
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 81
Fig 3 Ilia of sauropod dinosaurs scaled to same total length A Mamenchisaurus hochuanensis Young and Zhao 1972 holotype CCG V 20401 right iliumreversed modified from Young and Zhao (1972 pl 6 1a) B Diplodocus carnegii Hatcher 1901 CM 94 right ilium reversed modified from Hatcher (1901pl 10 1) C Camarasaurus supremus Cope 1877 AMNH 5761 Il 1 left ilium modified from Osborn and Mook (1921 fig 87) D Giraffatitan brancai(Janensch 1914) HMN J1 left ilium modified from Janensch (1961 pl E 2) E Rapetosaurus krausi Curry Rogers and Forster 2001 holotype FMNH PR2209 left ilium modified from Curry Rogers (2009 fig 39B) F Brontomerus mcintoshi gen et sp nov holotype OMNH 66430 left ilium
Fig 4 Measurement protocol for sauropod ilia as illustrated in Fig 3 andshown in Table 4 Total length is measured along the longest axis of theilium lengths of preacetabular and postacetabular lobes are measured parminusallel to this axis and extend from the extremity of the lobe to the anteriormargin of the pubic peduncle and posterior margin of the ischiadic pedunclerespectively Supracetabular height is measured perpendicular to the lonminusgest axis and extends from the highest point of the acetabulum to the pointlevel with the highest part of the ilium
curved in dorsal or ventral viewmdashfor example ApatosaurusMarsh 1877 (Upchurch et al 2004b pl 4 D E) Haplominuscanthosaurus Hatcher 1903b (Hatcher 1903a pl 5 1) andSaltasaurus Bonaparte and Powell 1980 (Powell 1992 fig17) In Brontomerus by contrast the blade of the ilium apminuspears to be ldquohingedrdquomdashdeflected laterally at a point directlyanterior to the public pedunclemdashso that the preacetabularlobe is straight in dorsal or ventral view and directed anterominuslaterally by an angle of about 30 to the sagittal In this reminusspect it more closely resembles the ilium of Camarasaurus
Cope 1877 (Osborn and Mook 1921 fig 49) although it difminusfers in other respects
The Brontomerus ilium is laterally compressed and unlikemost sauropod ilia the dorsal margin is not deflected laterallyrelative to the more ventral part so that in ventral view it apminuspears very thin (Fig 2B) It is well established that the longbones of sauropods grow isometrically through ontogeny(Carpenter and McIntosh 1994 Wilhite 1999 2003 Ikejiri etal 2005 Tidwell and Wilhite 2005 Taylor 2009) while theirvertebrae undergo significant changes in proportions laminaminus
82 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 4 Relative measurements of ilia in sauropods Measurement protocol is illustrated in Fig 4
Taxon Specimen ReferenceTotallength(cm)
Length ofpreacetabular
lobe (cmproportion oftotal length)
Length ofpostacetabular
lobe (cmproportion oftotal length)
Supracetabularheight (cm
proportion oftotal length)
Mamenchisaurushochuanensis
CCG V 20401 Young and Zhao (1972 pl 6 1a) 102 39 38 18 18 41 40
Diplodocus carnegii CM 94 Hatcher (1901 pl 10 1) 1091 41 38 20 18 49 45Camarasaurus
supremusAMNH 5761 Il 1 Osborn and Mook (1921 fig 87) 115 36 31 19 17 48 42
Giraffatitan brancai2
HMN Aa 13Janensch (1961 pl E 1a) 119 47 39 21 18 46 39
Measured by SchwarzminusWings(personal communication 2009) 1143 535 47 299 26
HMN J1Janensch (1961 pl E 2) 1055 55 52 16 15 37 35
Measured by SchwarzminusWings(personal communication 2009) 1077 528 49 135 13
Rapetosaurus krausi FMNH PR 2209 Curry Rogers (2009 fig 39) 42 20 48 7 17 16 38Brontomerus mcintoshi OMNH 66430 (this study) 405 223 55 0 0 21 52
1 Hatcher (1901) did not state the length of the ilium of CM 94 and did not figure that of CM 84 We have assumed that the ilium of the figured specimen CM94 is the same size as that of CM 84 (Hatcher 1901 46) and calculated the proportions from the figured specimen
2 Janenschrsquos (1961 pl E) figures of the two ilia of Giraffatitan brancai are not executed from an orthogonal perspective the ilium of HMN Aa 13 is illusminustrated from a slightly anterolateral position foreshortening the preacetabular lobe and that of HMN J 1 from a slight posterolateral position foreshortenminusing the postacetabular lobe The true lengths of the two lobes are probably somewhere between the two percentages calculated from the figures about45 for the former and 16 for the latter
10
cm
Fig 5 Damaged presacral vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah OMNH 66429 in dorsal view as photograph (A) and interpretive drawing (B) Shading indicates air spaces
tion pneumatic excavations and neurocentral fusion (Wedel2003a 248 b 352ndash354) Ontogenetic changes in limbminusgirdleelements such as the ilium are less well understood due to apaucity of sufficiently well preserved specimens (Ray Wilhitepersonal communication October 2007) Therefore the lateralcompression of the Brontomerus ilium may be a juvenile charminusacter with the ilium thickening through ontogeny to supportthe growing weight of the animal or it may in fact be phylominusgenetically significant
Presacral centrummdashA single presacral centrum OMNH66429 was recovered (Fig 5) Unfortunately preservation isvery poor the neural arch and all processes have been lostand the centrum has been greatly crushed dorsoventrally sothat the remaining part is essentially flat the element is 14
cm in both anteroposterior length and transverse width butno more than 3 cm in dorsoventral depth The small size indiminuscates that this element belonged to a juvenile The internalstructure of the centrum is visible however and consists offine septa dividing a hollow internal space irregularly intomany small camellae This morphology is characteristic oftitanosauriforms (Wedel 2003b 354ndash355) Highly camellateinternal structure has not previously been observed in juveminusnile sauropod vertebrae but this may be due to samplingbias so far all juvenile sauropod vertebrae that have beenstudied for internal structure have been those of Camaraminussaurus and diplodocoids which follow an ontogenetic trajecminustory in which large shallow lateral fossae develop into cameminusrae from which smaller accessory camerae and camellae deminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 83
prezygapophysis
neural spine
postzygapophysealramus
postzygapophysealfacet
chevron facets
5 cm
Fig 6 Midminuscaudal vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 61248 in dorsal (A) anterior (B) left lateral (C) posterior (D) and ventral (E) views
velop (Wedel et al 2000a fig 11 Wedel 2003b 349) TheHotel Mesa presacral suggests that camellate vertebrae mayhave developed differently in titanosauriforms possibly byinminussitu formation of camellae during pneumatization as ocminuscurs in the camellate vertebrae of birds (personal observaminustion MJW)
Caudal vertebraemdashOMNH 61248 is a distinctive caudalvertebra with elongated preminus and postzygapophyseal rami(Fig 6) Apart from the tip of the right prezygapophysis theelement is complete and well preserved While the centrum isonly 11 cm in length the distance from the prezygapophysisto postzygapophysis is 145 cm The centrum is slightlybroader than tall (6 cm compared with 55 cm anteriorly 65cm compared with 5 cm posteriorly) and gently waisted Theneural arch is set forward on the centrum but does not reachthe anterior margin The neural spine is so reduced and sostrongly inclined posteriorly as to be all but indistinguishminusable and is apparent only as a very low eminence abovethe postzygapophyses The postzygapophyseal facets themminusselves are set on the posterolateral faces of a low process thathangs below the main postzygapophyseal ramus Chevronfacets are weakly present on the posterior margin of the venminustral surface of the centrum but not on the anterior marginThe elongation index of 22 indicates a midminustominusposterior pominussition in the caudal sequence for this element as similarcentrum proportions do not appear until about caudal 30 inGiraffatitan brancai (Janensch 1950 pl 3)
This vertebra most closely resembles the indeterminatesauropod vertebra BMNH 27500 from the Wessex Formationof the Isle of Wight figured by Naish and Martill (2001 pl33) The Barremian age of that specimen places it about 15 Ma
earlier than OMNH 61248 Its neural arch is less elevated thanthat of the Hotel Mesa specimen its postzygapophyses projectyet farther posteriorly and its prezygapophyses less far anteriminusorly and it is very mildly biconvex rather than procoelous butin other respects including absolute size it is a good match forthe Brontomerus caudal
Also included in the Hotel Mesa material is OMNH27794 a partial distal caudal centrum figured by Wedel(2005 fig 77) This centrum is approximately round incrossminussection about 4 cm in diameter and internally consistsof apneumatic cancellous bone
RibsmdashThe Hotel Mesa material contains several dorsal ribsin various states of preservation but no readily identifiablecervical ribs The dorsal rib elements include the shaft of alarge flat rib (OMNH 27762) portions of several smaller ribshafts (OMNH 27763ndash27765 27768 and others) a flattenedrib head (OMNH 27767) and most informatively a smallcomplete rib (OMNH 27766 Fig 7) Despite its excellentpreservation and apparent lack of distortion this element isdifficult to interpret Its shaft is straight for almost its wholelength and both articular facets are directed dorsally ratherthan being inclined medially The tuberculum is directly inline with the main part of the shaft of the rib and thecapitulum is at an angle of about 30 to it In these respectsthe rib resembles the most anterior dorsal rib of the Diplominusdocus carnegii holotype CM 84 (personal observation MPTthis element was not figured by Hatcher [1901]) We thereminusfore interpret this rib as having probably belonged to the rightside of the first dorsal vertebra
The rib is unusual in other respects however most notablythat the ventral part of the shaft curves laterally rather than meminus
84 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
10 cm
capitulum
pneumatic fossa
tuberculum
Fig 7 First right dorsal rib of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah OMNH 27766 in posterior view head of rib showing pneumatic invasion of shaft (A) and complete rib showing laterally directed curvature of shaft (B)
dially Careful inspection of the bone reveals no indication ofdistortion or of incorrect reconstruction It may be possiblethat in life the thorax was transversely compressed so that thedorsal part of the rib shaft was directed ventromedially and themore ventral part was vertical Both articular facets are subminuscircular in dorsal view and significantly expanded comparedwith the rami that bear them On the anterior face of the head alow ridge arises just below the capitulum and extends downthe medial edge of the rib for about 40 of its length
The head of the rib is also unusual in that a thin sheet ofbone connects the rami that support the articular facets andthis sheet extends much farther proximally than in mostsauropod ribs Its precise extent cannot be ascertained due tobreakage The sheet of bone is perforated close to the capituminuslar ramus and from this perforation a pneumatic cavity inminusvades the shaft of the rib extending ventrally from within ashallow fossa in the posterior face Pneumatization of thedorsal ribs is a synapomorphy of Titanosauriformes (Wilsonand Sereno 1998) although pneumatic dorsal ribs are also inminusfrequently present in diplodocids (Gilmore 1936 Lovelaceet al 2003 2008)
Pectoral girdlemdashOMNH 27761 is a partial scapula consistminusing of the blade and part of the anterior expansion but missminusing the glenoid region and the remainder of the anterior exminuspansion (Fig 8) As preserved the element is nearly flat butthis may be due to postminusmortem distortion and in any casethe most strongly curved part of most sauropod scapulae isthe anterior part that is missing from this specimen The genminustle curvature preserved in the posterior part of the blade indiminuscates that the element was from the left side The bone is surminusprisingly thin in all preserved parts never exceeding a few
cm in contrast to for example the scapula of Camarasaurussupremus which is thick even in midminusblade (Osborn andMook 1921 fig 74b) This suggests that the glenoid thickenminusing and the acromial ridge may have been located some disminustance anterior to the preserved portion and a reconstructionafter the proportions of Giraffatitan brancai (Fig 8) suggeststhat about 80 of the scapularsquos full length is preserved Theposterior part of the acromion expansion is preserved howminusever and is sufficient to show that this expansion was prominusnounced so that the maximum dorsoventral height of thescapula was more than two and half times its minimumheight at the midpoint of the blade The dorsal margin slopesup towards the anterior expansion rather than forming a posminusteriorly directed ldquohookrdquo or a distinct acromion fossa betweenthe blade and the acromion process
The posterior expansion of the scapula is distinctive Insome sauropods the posterior part of the scapular blade isexpanded not at all or only slightly for example in Omeiminussaurus Young 1939 (He et al 1988 fig 41) Apatosaurus(Upchurch et al 2004b fig 4) and Rapetosaurus (Fig 9E)In others the ventral margin of the scapular blade is straightor nearly so while the dorsal margin is deflected dorsally tocreate an asymmetric expansion for example in Camaraminussaurus (Fig 9C) and Giraffatitan brancai (Fig 9D) In a fewsauropods however the ventral margin of the blade is alsodeflected ventrally to form a ldquoracquetminusshapedrdquo posterior exminuspansion This is seen in rebbachisaurids and some titanominussaurians eg some specimens of Alamosaurus Gilmore1922 (Gilmore 1946 fig 6) In Brontomerus the posteriorpart of the scapular blade is expanded in a characteristic manminusner the ventral margin is straight except for a posteroventral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 85
50 cm
ventral ldquosteprdquo
acromion expansion
dorsal ldquostepsrdquo
Fig 8 Partial left scapula of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 27761 in lateral view tentatively reconstructed after Giraffatitan brancai HMN Sa 9 (Janensch 1961 pl 15 1)
excursion two thirds of the way along the preserved portionafter which the margin continues parallel to its original traminusjectory so that the excursion appears as a gentle ldquosteprdquo Thedorsal margin is also ldquosteppedrdquo in this manner though withtwo distinct steps rather than one of which the more anterioris most strongly pronounced The net result of these featuresis that the dorsal and ventral borders of the scapula are bothstraight near the posterior extremity and that they are subminusparallel diverging by only about five degrees in the regionjust anterior to the rounded end of the posterior expansionThe step in the ventral border is not known in any othersauropod however the scapula of Neuquensaurus Powell1992 has a stepped dorsal border similar to that of Brontominusmerus (Huene 1929 via Glut 1997 275)
These characters of the scapula must be treated with somecaution however since this bone appears subject to morevariation than any other in the sauropod skeleton see for exminusample the range of shapes in scapulae of Giraffatitan brancai(Janensch 1961 pl 15 1ndash3) and in Camarasaurus supremus(Osborn and Mook 1921 figs 74ndash80)
Two small flat elements OMNH 66431 and 66432 are inminusterpreted as partial sternal plates (Fig 10) The medial edge ofeach is identifiable due to its rugose texture which formed theattachment site for cartilage joining the plates to each otherand to the sternal ribs The sternals are anteroposteriorly elonminusgate and mediolaterally narrow when complete they wereprobably at least three times as long as broad as in ldquoSaltaminussaurusrdquo robustus Huene 1929 (Huene 1929 via McIntosh1990 fig 169L) and proportionally longer than in any othersauropod including Saltasaurus loricatus (McIntosh 1990fig 169 Powell 1992 fig 30) The sternals are crescentic in
shape the anterior and posterior extremities curving laterallyaway from the midline This state was considered a titanominussaurian synapomorphy by Wilson (2002 268) but its distribuminustion is more complex in the current analysis being synapominusmorphic for Neosauropoda with losses in Flagellicaudata andCamarasaurus
86 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
Fig 10 Partial paired sternal plates of the camarasauromorph sauropodBrontomerus mcintoshi gen et sp nov from the Lower Cretaceous CedarMountain Formation of Utah OMNH 66431 and 66432 in ventral view
Fig 9 Scapulocoracoids and scapulae of sauropod dinosaurs scaled to same length of scapular blade from posterior point of glenoid to posterior margin ofblade A Mamenchisaurus youngi Young and Zhao 1972 holotype ZDM0083 left scapulocoracoid modified from Ouyang and Ye (2002 fig 22)B Diplodocus longus Hatcher 1901 USNM 10865 right scapulocoracoid reversed photograph by MPT C Camarasaurus supremus Cope 1877 AMNH5761 Sc 1 left scapula and AMNH 5761 Cor 1 left coracoid probably associated modified from Osborn and Mook (1921 figs 75 81a) D Giraffatitanbrancai (Janensch 1914) HMN Sa 9 left scapula modified from Janensch (1961 pl 15 1) E Rapetosaurus krausi Curry Rogers and Forster 2001holotype FMNH PR 2209 right scapula reversed modified from Curry Rogers (2009 fig 32) F Brontomerus mcintoshi gen et sp nov OMNH 27761left scapula tentatively reconstructed after Giraffatitan brancai
Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

not represent Tenontosaurus but further identification canminusnot be firmly established at present
Although the quarry has not been worked in severalyears it is not exhausted and may contain additional relevantmaterial (James Kirkland personal communication October2007)
Systematic paleontology
Dinosauria Owen 1842Saurischia Seeley 1888Sauropoda Marsh 1878Neosauropoda Bonaparte 1986Camarasauromorpha Salgado Coria and Calvo 1997Genus Brontomerus novType species Brontomerus mcintoshi sp nov see below
Etymology From Greek bronto thunder Greek meroacutes thigh ldquothunminusderminusthighsrdquo in reference to the substantial femoral musculature impliedby the morphology of the ilium Intended English pronunciation BronminusminustoeminusMEERminusus
DiagnosismdashAs for type and only species (see below)
Brontomerus mcintoshi sp novFigs 1 2 5ndash8 10 12 Table 3
Etymology In honor of veteran sauropod worker John S McIntoshwhose seminal paleontological work done mostly unfunded and on hisown time has been an inspiration to all of us who follow
Holotype OMNH 66430 a left ilium
Tentatively referred material OMNH 66429 crushed presacralcentrum OMNH 61248 midminustominusposterior caudal vertebra OMNH27794 partial distal caudal centrum OMNH 27766 anterior right dorminussal rib OMNH 27761 nearly complete left scapula missing anteriorportion OMNH 66431 and 66432 two partial sternal plates other fragminusments as detailed in Table 3
Type locality Hotel Mesa Quarry (OMNH locality V857) GrandCounty eastern Utah
Type horizon Top of the Ruby Ranch Member of the Cedar MountainFormation (Lower Cretaceous AptianndashAlbian)
DiagnosismdashPreacetabular lobe 55 of total ilium lengthlonger than in any other sauropod preacetabular lobe directedanterolaterally at 30 relative to the sagittal plane but straightin dorsal view and vertically oriented postacetabular lobe reminusduced to near absence ischiadic peduncle reduced to very lowbulge ilium proportionally taller than in any other saurominuspodmdashheight is 52 of total length compared with a maximinusmum of 45 in other sauropods If the tentatively referred eleminus
78 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 3 Material referred to the camarasauromorph sauropod Brontomerus mcintoshi from the Lower Cretaceous Cedar Mountain Formation ofUtah
OMNH specimen number Description Length (cm) Width (cm) Thickness (cm)27761 partial scapula missing anterior part 98 55 527762 large flat rib shaft 101 75 227763 rib fragment 63 8 527764 rib fragment 60 25 227765 rib fragment 58 6 327766 complete pneumatic rib 76 12 427767 dorsal part of rib flattened 26 17 227768 rib fragment 31 6 1527769 fragment of ischium 10 5 2527770 rib fragment 14 35 127771 rib fragment 17 3 227772 rib fragment (in two parts) 13 7 227773 rib fragment 135 4 227773 flat scrap 7 5 0527773 flat scrap 115 9 1
27774ndash27783 fragments27784 collection of 21 fragments
27785ndash27793 fragments27794 partial distal caudal centrum
27795ndash27800 fragments61248 nearly complete midminuscaudal vertebra 11 6 5566429 crushed presacral centrum 14 14 366430 ilium 405 31 866430 dorsal fragment of ilium 14 3 166430 dorsal fragment of ilium 155 9 166431 incomplete sternal plate 15 75 1566432 incomplete sternal plate 115 7 1
ments do belong to the same species as the holotype then thefollowing additional characters also diagnose the new taxonpresacral vertebrae camellate midminustominusposterior caudal verteminusbrae with elongate preminus and postzygapophyseal rami havingthe postzygapophyseal facets hanging below the level of theramus first dorsal rib with expanded dorsally oriented articuminuslar facets laterally curving shaft and ventrally directed pneuminusmatic foramen in head acromion expansion of scapula prominusnounced and steep but not forming acromion fossa dorsal andventral margins of scapular blade ldquosteppedrdquo sternal platescrescentic and three times as long as broad
Unambiguous autapomorphies distinguishing Brontomeminusrus from the root of the polytomy in which it is recovered inthe strict consensus of most parsimonious trees in the phylominusgenetic analysis below character 184 ratio of centrumlengthheight in middle caudal vertebrae 20 185 sharpridge on lateral surface of middle caudal centra at archminusbodyjunction absent 212 posterior end of scapular body racminusquetminusshaped (dorsoventrally expanded) 261 in lateral viewthe most anteroventral point on the iliac preacetabular lobe isalso the most anterior point (preacetabular lobe is pointed)264 projected line connecting articular surfaces of ischiadicand pubic peduncles of ilium passes ventral to ventral marginof postacetabular lobe of ilium
Description
This taxon is based on a collection of elements all from thesame quarry all of them consistent with assignment to a sinminusgle taxon (Fig 1) However the elements were not found arminusticulated and their differing sizes do not permit interpretaminustion as belonging to a single individual For example the parminus
tial scapula is 98 cm long Reconstruction after the scapula ofGiraffatitan brancai (Janensch 1914) suggests that the comminusplete element was about 121 cm long In Rapetosaurus CurryRogers and Forster 2001 the scapula and ilium are about thesame length (Curry Rogers and Forster 2001 fig 3) but theilium of Brontomerus is only one third the reconstructedlength of the scapula The quarry therefore contains at leasttwo individuals of widely differing sizes The smaller prominusviding the holotype ilium and the referred presacral centrumand sternal plates is interpreted as a juvenile the remainingelements probably belonged to the larger mature individual
Among the sauropod elements recovered from the HotelMesa site at least three (the caudal vertebra scapula andilium) have characters not known in any other sauropodSince it is not possible to conclusively demonstrate that allthree elements belong to the same taxon (and size differminusences preclude their belonging to a single individual) it istherefore possible that the quarry contains as many as threenew taxa However since all the informative elements havecharacters that indicate a titanosauriform identity we conminusservatively consider it more likely that only a single newtaxon is present Groups of sauropods of mixed ages areknown for several taxa (eg Bird 1985 Coria 1994 Bader2003) so there is a precedent for finding adults and juveminusniles together Although we diagnose the new taxon solelyon the type ilium we also tentatively refer the other eleminusments to this taxon and note the additional characters thatpertain if this referral is correct we hope that subsequentwork in the Hotel Mesa Quarry or elsewhere in the CedarMountain Formation will yield specimens that allow us toconfirm or refute this referral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 79
Fig 1 Skeletal inventory of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah in left lateral view Preserved elements are white missing elements are reconstructed in gray After a Camarasaurus grandis reconstructionkindly provided by Scott Hartman
The assignment of specimen numbers to the material deminusscribed here is complex (Table 3) Specimen number OMNH27773 comprises three elements OMNH 27784 consists of 21small fragments of bone none of them informative all otherelements have their own specimen numbers in the range27761ndash27800 apart from the midminuscaudal vertebra OMNH61248 and the reassigned numbers 66429ndash66432 for eleminusments extracted from OMNH 27773
IliummdashThe most informative element is OMNH 66430 a leftilium (Fig 2) which has therefore been selected as the holominustype The ilium was preserved complete but lay hidden beminusneath the scapula and so was damaged in the field (JamesKirkland personal communication March 2008) The iliumis preserved in three parts one provides most of the boneincluding the well preserved preacetabular lobe pubic andischiadic peduncles and acetabular margin and the other two
provide most of the dorsal margin giving a good indication ofthe degree of curvature The relative positions and orientationof the two smaller fragments can not be definitely ascertainedbut they appear to be parts of a single large fragment broken atthe point where we have reconstructed them as touching andif this interpretation is correct then the curvature of the pair inminusdicates which side must be oriented laterally
The ilium is remarkable in that the preacetabular lobe isrelatively larger than in any other sauropod (Fig 3 Table 4measurement protocol is illustrated in Fig 4) the postaceminustabular lobe is reduced almost to the point of absence and theilium is proportionally taller than in any other sauropod Theischiadic peduncle is reduced to a very low ventral projectionfrom almost the most posterior point of the ilium The near abminussence of the ischiadic peduncle cannot be attributed to damageas the iliac articular surface is preserved Immediately posterominus
80 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
acetabulum
pubicpeduncle
10 cm
ischiadicpeduncle
pre-acetabularblade
Fig 2 Left ilium of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah type specimen OMNH 66430 in lateral view reconstructed from the three fragments (A) and ventral view (B)
dorsal to this surface is a subtle notch between the peduncleand the very reduced postacetabular lobe This notch and theareas either side of it are composed of finished bone demonminusstrating that the great reduction of the postacetabular lobe toois a genuine osteological feature and not due to damage In reminusgard to the proportionally large preacetabular lobe the iliumof Brontomerus resembles that of Rapetosaurus (Fig 3E Taminusble 4) However that taxon has a normal postacetabular lobeand is not proportionally tall In overall proportions the iliumof Brontomerus is more similar to the left ilium HMN J1 asminussigned to Giraffatitan brancai (Janensch 1961 pl E 2) whichalso has a reduced postacetabular lobe mdashsee also Fig 3DHowever the ilium of Brontomerus is proportionally tallerthan that of G brancai and its anterior margin comes to apoint rather than being smoothly rounded as in that taxon InBrontomerus the maximum height of the ilium above theacetabular margin when measured perpendicular to the lonminusgest axis is 52 in other sauropods examined this proportiondoes not exceed 45 and averages 40 (Table 4)
As with many sauropods the preacetabular lobe of theilium flares laterally However in most sauropods this flarminusing is progressive so that in dorsal or ventral view the mostposterior part of the preacetabular lobe is nearly parallel witha line drawn between the pubic and ischiadic peduncles andsmooth lateral curvature inclines the more anterior parts inminuscreasingly laterally so that the more anterior part is almost atright angles to this line and the ilium appears smoothly
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 81
Fig 3 Ilia of sauropod dinosaurs scaled to same total length A Mamenchisaurus hochuanensis Young and Zhao 1972 holotype CCG V 20401 right iliumreversed modified from Young and Zhao (1972 pl 6 1a) B Diplodocus carnegii Hatcher 1901 CM 94 right ilium reversed modified from Hatcher (1901pl 10 1) C Camarasaurus supremus Cope 1877 AMNH 5761 Il 1 left ilium modified from Osborn and Mook (1921 fig 87) D Giraffatitan brancai(Janensch 1914) HMN J1 left ilium modified from Janensch (1961 pl E 2) E Rapetosaurus krausi Curry Rogers and Forster 2001 holotype FMNH PR2209 left ilium modified from Curry Rogers (2009 fig 39B) F Brontomerus mcintoshi gen et sp nov holotype OMNH 66430 left ilium
Fig 4 Measurement protocol for sauropod ilia as illustrated in Fig 3 andshown in Table 4 Total length is measured along the longest axis of theilium lengths of preacetabular and postacetabular lobes are measured parminusallel to this axis and extend from the extremity of the lobe to the anteriormargin of the pubic peduncle and posterior margin of the ischiadic pedunclerespectively Supracetabular height is measured perpendicular to the lonminusgest axis and extends from the highest point of the acetabulum to the pointlevel with the highest part of the ilium
curved in dorsal or ventral viewmdashfor example ApatosaurusMarsh 1877 (Upchurch et al 2004b pl 4 D E) Haplominuscanthosaurus Hatcher 1903b (Hatcher 1903a pl 5 1) andSaltasaurus Bonaparte and Powell 1980 (Powell 1992 fig17) In Brontomerus by contrast the blade of the ilium apminuspears to be ldquohingedrdquomdashdeflected laterally at a point directlyanterior to the public pedunclemdashso that the preacetabularlobe is straight in dorsal or ventral view and directed anterominuslaterally by an angle of about 30 to the sagittal In this reminusspect it more closely resembles the ilium of Camarasaurus
Cope 1877 (Osborn and Mook 1921 fig 49) although it difminusfers in other respects
The Brontomerus ilium is laterally compressed and unlikemost sauropod ilia the dorsal margin is not deflected laterallyrelative to the more ventral part so that in ventral view it apminuspears very thin (Fig 2B) It is well established that the longbones of sauropods grow isometrically through ontogeny(Carpenter and McIntosh 1994 Wilhite 1999 2003 Ikejiri etal 2005 Tidwell and Wilhite 2005 Taylor 2009) while theirvertebrae undergo significant changes in proportions laminaminus
82 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 4 Relative measurements of ilia in sauropods Measurement protocol is illustrated in Fig 4
Taxon Specimen ReferenceTotallength(cm)
Length ofpreacetabular
lobe (cmproportion oftotal length)
Length ofpostacetabular
lobe (cmproportion oftotal length)
Supracetabularheight (cm
proportion oftotal length)
Mamenchisaurushochuanensis
CCG V 20401 Young and Zhao (1972 pl 6 1a) 102 39 38 18 18 41 40
Diplodocus carnegii CM 94 Hatcher (1901 pl 10 1) 1091 41 38 20 18 49 45Camarasaurus
supremusAMNH 5761 Il 1 Osborn and Mook (1921 fig 87) 115 36 31 19 17 48 42
Giraffatitan brancai2
HMN Aa 13Janensch (1961 pl E 1a) 119 47 39 21 18 46 39
Measured by SchwarzminusWings(personal communication 2009) 1143 535 47 299 26
HMN J1Janensch (1961 pl E 2) 1055 55 52 16 15 37 35
Measured by SchwarzminusWings(personal communication 2009) 1077 528 49 135 13
Rapetosaurus krausi FMNH PR 2209 Curry Rogers (2009 fig 39) 42 20 48 7 17 16 38Brontomerus mcintoshi OMNH 66430 (this study) 405 223 55 0 0 21 52
1 Hatcher (1901) did not state the length of the ilium of CM 94 and did not figure that of CM 84 We have assumed that the ilium of the figured specimen CM94 is the same size as that of CM 84 (Hatcher 1901 46) and calculated the proportions from the figured specimen
2 Janenschrsquos (1961 pl E) figures of the two ilia of Giraffatitan brancai are not executed from an orthogonal perspective the ilium of HMN Aa 13 is illusminustrated from a slightly anterolateral position foreshortening the preacetabular lobe and that of HMN J 1 from a slight posterolateral position foreshortenminusing the postacetabular lobe The true lengths of the two lobes are probably somewhere between the two percentages calculated from the figures about45 for the former and 16 for the latter
10
cm
Fig 5 Damaged presacral vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah OMNH 66429 in dorsal view as photograph (A) and interpretive drawing (B) Shading indicates air spaces
tion pneumatic excavations and neurocentral fusion (Wedel2003a 248 b 352ndash354) Ontogenetic changes in limbminusgirdleelements such as the ilium are less well understood due to apaucity of sufficiently well preserved specimens (Ray Wilhitepersonal communication October 2007) Therefore the lateralcompression of the Brontomerus ilium may be a juvenile charminusacter with the ilium thickening through ontogeny to supportthe growing weight of the animal or it may in fact be phylominusgenetically significant
Presacral centrummdashA single presacral centrum OMNH66429 was recovered (Fig 5) Unfortunately preservation isvery poor the neural arch and all processes have been lostand the centrum has been greatly crushed dorsoventrally sothat the remaining part is essentially flat the element is 14
cm in both anteroposterior length and transverse width butno more than 3 cm in dorsoventral depth The small size indiminuscates that this element belonged to a juvenile The internalstructure of the centrum is visible however and consists offine septa dividing a hollow internal space irregularly intomany small camellae This morphology is characteristic oftitanosauriforms (Wedel 2003b 354ndash355) Highly camellateinternal structure has not previously been observed in juveminusnile sauropod vertebrae but this may be due to samplingbias so far all juvenile sauropod vertebrae that have beenstudied for internal structure have been those of Camaraminussaurus and diplodocoids which follow an ontogenetic trajecminustory in which large shallow lateral fossae develop into cameminusrae from which smaller accessory camerae and camellae deminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 83
prezygapophysis
neural spine
postzygapophysealramus
postzygapophysealfacet
chevron facets
5 cm
Fig 6 Midminuscaudal vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 61248 in dorsal (A) anterior (B) left lateral (C) posterior (D) and ventral (E) views
velop (Wedel et al 2000a fig 11 Wedel 2003b 349) TheHotel Mesa presacral suggests that camellate vertebrae mayhave developed differently in titanosauriforms possibly byinminussitu formation of camellae during pneumatization as ocminuscurs in the camellate vertebrae of birds (personal observaminustion MJW)
Caudal vertebraemdashOMNH 61248 is a distinctive caudalvertebra with elongated preminus and postzygapophyseal rami(Fig 6) Apart from the tip of the right prezygapophysis theelement is complete and well preserved While the centrum isonly 11 cm in length the distance from the prezygapophysisto postzygapophysis is 145 cm The centrum is slightlybroader than tall (6 cm compared with 55 cm anteriorly 65cm compared with 5 cm posteriorly) and gently waisted Theneural arch is set forward on the centrum but does not reachthe anterior margin The neural spine is so reduced and sostrongly inclined posteriorly as to be all but indistinguishminusable and is apparent only as a very low eminence abovethe postzygapophyses The postzygapophyseal facets themminusselves are set on the posterolateral faces of a low process thathangs below the main postzygapophyseal ramus Chevronfacets are weakly present on the posterior margin of the venminustral surface of the centrum but not on the anterior marginThe elongation index of 22 indicates a midminustominusposterior pominussition in the caudal sequence for this element as similarcentrum proportions do not appear until about caudal 30 inGiraffatitan brancai (Janensch 1950 pl 3)
This vertebra most closely resembles the indeterminatesauropod vertebra BMNH 27500 from the Wessex Formationof the Isle of Wight figured by Naish and Martill (2001 pl33) The Barremian age of that specimen places it about 15 Ma
earlier than OMNH 61248 Its neural arch is less elevated thanthat of the Hotel Mesa specimen its postzygapophyses projectyet farther posteriorly and its prezygapophyses less far anteriminusorly and it is very mildly biconvex rather than procoelous butin other respects including absolute size it is a good match forthe Brontomerus caudal
Also included in the Hotel Mesa material is OMNH27794 a partial distal caudal centrum figured by Wedel(2005 fig 77) This centrum is approximately round incrossminussection about 4 cm in diameter and internally consistsof apneumatic cancellous bone
RibsmdashThe Hotel Mesa material contains several dorsal ribsin various states of preservation but no readily identifiablecervical ribs The dorsal rib elements include the shaft of alarge flat rib (OMNH 27762) portions of several smaller ribshafts (OMNH 27763ndash27765 27768 and others) a flattenedrib head (OMNH 27767) and most informatively a smallcomplete rib (OMNH 27766 Fig 7) Despite its excellentpreservation and apparent lack of distortion this element isdifficult to interpret Its shaft is straight for almost its wholelength and both articular facets are directed dorsally ratherthan being inclined medially The tuberculum is directly inline with the main part of the shaft of the rib and thecapitulum is at an angle of about 30 to it In these respectsthe rib resembles the most anterior dorsal rib of the Diplominusdocus carnegii holotype CM 84 (personal observation MPTthis element was not figured by Hatcher [1901]) We thereminusfore interpret this rib as having probably belonged to the rightside of the first dorsal vertebra
The rib is unusual in other respects however most notablythat the ventral part of the shaft curves laterally rather than meminus
84 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
10 cm
capitulum
pneumatic fossa
tuberculum
Fig 7 First right dorsal rib of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah OMNH 27766 in posterior view head of rib showing pneumatic invasion of shaft (A) and complete rib showing laterally directed curvature of shaft (B)
dially Careful inspection of the bone reveals no indication ofdistortion or of incorrect reconstruction It may be possiblethat in life the thorax was transversely compressed so that thedorsal part of the rib shaft was directed ventromedially and themore ventral part was vertical Both articular facets are subminuscircular in dorsal view and significantly expanded comparedwith the rami that bear them On the anterior face of the head alow ridge arises just below the capitulum and extends downthe medial edge of the rib for about 40 of its length
The head of the rib is also unusual in that a thin sheet ofbone connects the rami that support the articular facets andthis sheet extends much farther proximally than in mostsauropod ribs Its precise extent cannot be ascertained due tobreakage The sheet of bone is perforated close to the capituminuslar ramus and from this perforation a pneumatic cavity inminusvades the shaft of the rib extending ventrally from within ashallow fossa in the posterior face Pneumatization of thedorsal ribs is a synapomorphy of Titanosauriformes (Wilsonand Sereno 1998) although pneumatic dorsal ribs are also inminusfrequently present in diplodocids (Gilmore 1936 Lovelaceet al 2003 2008)
Pectoral girdlemdashOMNH 27761 is a partial scapula consistminusing of the blade and part of the anterior expansion but missminusing the glenoid region and the remainder of the anterior exminuspansion (Fig 8) As preserved the element is nearly flat butthis may be due to postminusmortem distortion and in any casethe most strongly curved part of most sauropod scapulae isthe anterior part that is missing from this specimen The genminustle curvature preserved in the posterior part of the blade indiminuscates that the element was from the left side The bone is surminusprisingly thin in all preserved parts never exceeding a few
cm in contrast to for example the scapula of Camarasaurussupremus which is thick even in midminusblade (Osborn andMook 1921 fig 74b) This suggests that the glenoid thickenminusing and the acromial ridge may have been located some disminustance anterior to the preserved portion and a reconstructionafter the proportions of Giraffatitan brancai (Fig 8) suggeststhat about 80 of the scapularsquos full length is preserved Theposterior part of the acromion expansion is preserved howminusever and is sufficient to show that this expansion was prominusnounced so that the maximum dorsoventral height of thescapula was more than two and half times its minimumheight at the midpoint of the blade The dorsal margin slopesup towards the anterior expansion rather than forming a posminusteriorly directed ldquohookrdquo or a distinct acromion fossa betweenthe blade and the acromion process
The posterior expansion of the scapula is distinctive Insome sauropods the posterior part of the scapular blade isexpanded not at all or only slightly for example in Omeiminussaurus Young 1939 (He et al 1988 fig 41) Apatosaurus(Upchurch et al 2004b fig 4) and Rapetosaurus (Fig 9E)In others the ventral margin of the scapular blade is straightor nearly so while the dorsal margin is deflected dorsally tocreate an asymmetric expansion for example in Camaraminussaurus (Fig 9C) and Giraffatitan brancai (Fig 9D) In a fewsauropods however the ventral margin of the blade is alsodeflected ventrally to form a ldquoracquetminusshapedrdquo posterior exminuspansion This is seen in rebbachisaurids and some titanominussaurians eg some specimens of Alamosaurus Gilmore1922 (Gilmore 1946 fig 6) In Brontomerus the posteriorpart of the scapular blade is expanded in a characteristic manminusner the ventral margin is straight except for a posteroventral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 85
50 cm
ventral ldquosteprdquo
acromion expansion
dorsal ldquostepsrdquo
Fig 8 Partial left scapula of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 27761 in lateral view tentatively reconstructed after Giraffatitan brancai HMN Sa 9 (Janensch 1961 pl 15 1)
excursion two thirds of the way along the preserved portionafter which the margin continues parallel to its original traminusjectory so that the excursion appears as a gentle ldquosteprdquo Thedorsal margin is also ldquosteppedrdquo in this manner though withtwo distinct steps rather than one of which the more anterioris most strongly pronounced The net result of these featuresis that the dorsal and ventral borders of the scapula are bothstraight near the posterior extremity and that they are subminusparallel diverging by only about five degrees in the regionjust anterior to the rounded end of the posterior expansionThe step in the ventral border is not known in any othersauropod however the scapula of Neuquensaurus Powell1992 has a stepped dorsal border similar to that of Brontominusmerus (Huene 1929 via Glut 1997 275)
These characters of the scapula must be treated with somecaution however since this bone appears subject to morevariation than any other in the sauropod skeleton see for exminusample the range of shapes in scapulae of Giraffatitan brancai(Janensch 1961 pl 15 1ndash3) and in Camarasaurus supremus(Osborn and Mook 1921 figs 74ndash80)
Two small flat elements OMNH 66431 and 66432 are inminusterpreted as partial sternal plates (Fig 10) The medial edge ofeach is identifiable due to its rugose texture which formed theattachment site for cartilage joining the plates to each otherand to the sternal ribs The sternals are anteroposteriorly elonminusgate and mediolaterally narrow when complete they wereprobably at least three times as long as broad as in ldquoSaltaminussaurusrdquo robustus Huene 1929 (Huene 1929 via McIntosh1990 fig 169L) and proportionally longer than in any othersauropod including Saltasaurus loricatus (McIntosh 1990fig 169 Powell 1992 fig 30) The sternals are crescentic in
shape the anterior and posterior extremities curving laterallyaway from the midline This state was considered a titanominussaurian synapomorphy by Wilson (2002 268) but its distribuminustion is more complex in the current analysis being synapominusmorphic for Neosauropoda with losses in Flagellicaudata andCamarasaurus
86 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
Fig 10 Partial paired sternal plates of the camarasauromorph sauropodBrontomerus mcintoshi gen et sp nov from the Lower Cretaceous CedarMountain Formation of Utah OMNH 66431 and 66432 in ventral view
Fig 9 Scapulocoracoids and scapulae of sauropod dinosaurs scaled to same length of scapular blade from posterior point of glenoid to posterior margin ofblade A Mamenchisaurus youngi Young and Zhao 1972 holotype ZDM0083 left scapulocoracoid modified from Ouyang and Ye (2002 fig 22)B Diplodocus longus Hatcher 1901 USNM 10865 right scapulocoracoid reversed photograph by MPT C Camarasaurus supremus Cope 1877 AMNH5761 Sc 1 left scapula and AMNH 5761 Cor 1 left coracoid probably associated modified from Osborn and Mook (1921 figs 75 81a) D Giraffatitanbrancai (Janensch 1914) HMN Sa 9 left scapula modified from Janensch (1961 pl 15 1) E Rapetosaurus krausi Curry Rogers and Forster 2001holotype FMNH PR 2209 right scapula reversed modified from Curry Rogers (2009 fig 32) F Brontomerus mcintoshi gen et sp nov OMNH 27761left scapula tentatively reconstructed after Giraffatitan brancai
Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

ments do belong to the same species as the holotype then thefollowing additional characters also diagnose the new taxonpresacral vertebrae camellate midminustominusposterior caudal verteminusbrae with elongate preminus and postzygapophyseal rami havingthe postzygapophyseal facets hanging below the level of theramus first dorsal rib with expanded dorsally oriented articuminuslar facets laterally curving shaft and ventrally directed pneuminusmatic foramen in head acromion expansion of scapula prominusnounced and steep but not forming acromion fossa dorsal andventral margins of scapular blade ldquosteppedrdquo sternal platescrescentic and three times as long as broad
Unambiguous autapomorphies distinguishing Brontomeminusrus from the root of the polytomy in which it is recovered inthe strict consensus of most parsimonious trees in the phylominusgenetic analysis below character 184 ratio of centrumlengthheight in middle caudal vertebrae 20 185 sharpridge on lateral surface of middle caudal centra at archminusbodyjunction absent 212 posterior end of scapular body racminusquetminusshaped (dorsoventrally expanded) 261 in lateral viewthe most anteroventral point on the iliac preacetabular lobe isalso the most anterior point (preacetabular lobe is pointed)264 projected line connecting articular surfaces of ischiadicand pubic peduncles of ilium passes ventral to ventral marginof postacetabular lobe of ilium
Description
This taxon is based on a collection of elements all from thesame quarry all of them consistent with assignment to a sinminusgle taxon (Fig 1) However the elements were not found arminusticulated and their differing sizes do not permit interpretaminustion as belonging to a single individual For example the parminus
tial scapula is 98 cm long Reconstruction after the scapula ofGiraffatitan brancai (Janensch 1914) suggests that the comminusplete element was about 121 cm long In Rapetosaurus CurryRogers and Forster 2001 the scapula and ilium are about thesame length (Curry Rogers and Forster 2001 fig 3) but theilium of Brontomerus is only one third the reconstructedlength of the scapula The quarry therefore contains at leasttwo individuals of widely differing sizes The smaller prominusviding the holotype ilium and the referred presacral centrumand sternal plates is interpreted as a juvenile the remainingelements probably belonged to the larger mature individual
Among the sauropod elements recovered from the HotelMesa site at least three (the caudal vertebra scapula andilium) have characters not known in any other sauropodSince it is not possible to conclusively demonstrate that allthree elements belong to the same taxon (and size differminusences preclude their belonging to a single individual) it istherefore possible that the quarry contains as many as threenew taxa However since all the informative elements havecharacters that indicate a titanosauriform identity we conminusservatively consider it more likely that only a single newtaxon is present Groups of sauropods of mixed ages areknown for several taxa (eg Bird 1985 Coria 1994 Bader2003) so there is a precedent for finding adults and juveminusniles together Although we diagnose the new taxon solelyon the type ilium we also tentatively refer the other eleminusments to this taxon and note the additional characters thatpertain if this referral is correct we hope that subsequentwork in the Hotel Mesa Quarry or elsewhere in the CedarMountain Formation will yield specimens that allow us toconfirm or refute this referral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 79
Fig 1 Skeletal inventory of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah in left lateral view Preserved elements are white missing elements are reconstructed in gray After a Camarasaurus grandis reconstructionkindly provided by Scott Hartman
The assignment of specimen numbers to the material deminusscribed here is complex (Table 3) Specimen number OMNH27773 comprises three elements OMNH 27784 consists of 21small fragments of bone none of them informative all otherelements have their own specimen numbers in the range27761ndash27800 apart from the midminuscaudal vertebra OMNH61248 and the reassigned numbers 66429ndash66432 for eleminusments extracted from OMNH 27773
IliummdashThe most informative element is OMNH 66430 a leftilium (Fig 2) which has therefore been selected as the holominustype The ilium was preserved complete but lay hidden beminusneath the scapula and so was damaged in the field (JamesKirkland personal communication March 2008) The iliumis preserved in three parts one provides most of the boneincluding the well preserved preacetabular lobe pubic andischiadic peduncles and acetabular margin and the other two
provide most of the dorsal margin giving a good indication ofthe degree of curvature The relative positions and orientationof the two smaller fragments can not be definitely ascertainedbut they appear to be parts of a single large fragment broken atthe point where we have reconstructed them as touching andif this interpretation is correct then the curvature of the pair inminusdicates which side must be oriented laterally
The ilium is remarkable in that the preacetabular lobe isrelatively larger than in any other sauropod (Fig 3 Table 4measurement protocol is illustrated in Fig 4) the postaceminustabular lobe is reduced almost to the point of absence and theilium is proportionally taller than in any other sauropod Theischiadic peduncle is reduced to a very low ventral projectionfrom almost the most posterior point of the ilium The near abminussence of the ischiadic peduncle cannot be attributed to damageas the iliac articular surface is preserved Immediately posterominus
80 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
acetabulum
pubicpeduncle
10 cm
ischiadicpeduncle
pre-acetabularblade
Fig 2 Left ilium of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah type specimen OMNH 66430 in lateral view reconstructed from the three fragments (A) and ventral view (B)
dorsal to this surface is a subtle notch between the peduncleand the very reduced postacetabular lobe This notch and theareas either side of it are composed of finished bone demonminusstrating that the great reduction of the postacetabular lobe toois a genuine osteological feature and not due to damage In reminusgard to the proportionally large preacetabular lobe the iliumof Brontomerus resembles that of Rapetosaurus (Fig 3E Taminusble 4) However that taxon has a normal postacetabular lobeand is not proportionally tall In overall proportions the iliumof Brontomerus is more similar to the left ilium HMN J1 asminussigned to Giraffatitan brancai (Janensch 1961 pl E 2) whichalso has a reduced postacetabular lobe mdashsee also Fig 3DHowever the ilium of Brontomerus is proportionally tallerthan that of G brancai and its anterior margin comes to apoint rather than being smoothly rounded as in that taxon InBrontomerus the maximum height of the ilium above theacetabular margin when measured perpendicular to the lonminusgest axis is 52 in other sauropods examined this proportiondoes not exceed 45 and averages 40 (Table 4)
As with many sauropods the preacetabular lobe of theilium flares laterally However in most sauropods this flarminusing is progressive so that in dorsal or ventral view the mostposterior part of the preacetabular lobe is nearly parallel witha line drawn between the pubic and ischiadic peduncles andsmooth lateral curvature inclines the more anterior parts inminuscreasingly laterally so that the more anterior part is almost atright angles to this line and the ilium appears smoothly
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 81
Fig 3 Ilia of sauropod dinosaurs scaled to same total length A Mamenchisaurus hochuanensis Young and Zhao 1972 holotype CCG V 20401 right iliumreversed modified from Young and Zhao (1972 pl 6 1a) B Diplodocus carnegii Hatcher 1901 CM 94 right ilium reversed modified from Hatcher (1901pl 10 1) C Camarasaurus supremus Cope 1877 AMNH 5761 Il 1 left ilium modified from Osborn and Mook (1921 fig 87) D Giraffatitan brancai(Janensch 1914) HMN J1 left ilium modified from Janensch (1961 pl E 2) E Rapetosaurus krausi Curry Rogers and Forster 2001 holotype FMNH PR2209 left ilium modified from Curry Rogers (2009 fig 39B) F Brontomerus mcintoshi gen et sp nov holotype OMNH 66430 left ilium
Fig 4 Measurement protocol for sauropod ilia as illustrated in Fig 3 andshown in Table 4 Total length is measured along the longest axis of theilium lengths of preacetabular and postacetabular lobes are measured parminusallel to this axis and extend from the extremity of the lobe to the anteriormargin of the pubic peduncle and posterior margin of the ischiadic pedunclerespectively Supracetabular height is measured perpendicular to the lonminusgest axis and extends from the highest point of the acetabulum to the pointlevel with the highest part of the ilium
curved in dorsal or ventral viewmdashfor example ApatosaurusMarsh 1877 (Upchurch et al 2004b pl 4 D E) Haplominuscanthosaurus Hatcher 1903b (Hatcher 1903a pl 5 1) andSaltasaurus Bonaparte and Powell 1980 (Powell 1992 fig17) In Brontomerus by contrast the blade of the ilium apminuspears to be ldquohingedrdquomdashdeflected laterally at a point directlyanterior to the public pedunclemdashso that the preacetabularlobe is straight in dorsal or ventral view and directed anterominuslaterally by an angle of about 30 to the sagittal In this reminusspect it more closely resembles the ilium of Camarasaurus
Cope 1877 (Osborn and Mook 1921 fig 49) although it difminusfers in other respects
The Brontomerus ilium is laterally compressed and unlikemost sauropod ilia the dorsal margin is not deflected laterallyrelative to the more ventral part so that in ventral view it apminuspears very thin (Fig 2B) It is well established that the longbones of sauropods grow isometrically through ontogeny(Carpenter and McIntosh 1994 Wilhite 1999 2003 Ikejiri etal 2005 Tidwell and Wilhite 2005 Taylor 2009) while theirvertebrae undergo significant changes in proportions laminaminus
82 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 4 Relative measurements of ilia in sauropods Measurement protocol is illustrated in Fig 4
Taxon Specimen ReferenceTotallength(cm)
Length ofpreacetabular
lobe (cmproportion oftotal length)
Length ofpostacetabular
lobe (cmproportion oftotal length)
Supracetabularheight (cm
proportion oftotal length)
Mamenchisaurushochuanensis
CCG V 20401 Young and Zhao (1972 pl 6 1a) 102 39 38 18 18 41 40
Diplodocus carnegii CM 94 Hatcher (1901 pl 10 1) 1091 41 38 20 18 49 45Camarasaurus
supremusAMNH 5761 Il 1 Osborn and Mook (1921 fig 87) 115 36 31 19 17 48 42
Giraffatitan brancai2
HMN Aa 13Janensch (1961 pl E 1a) 119 47 39 21 18 46 39
Measured by SchwarzminusWings(personal communication 2009) 1143 535 47 299 26
HMN J1Janensch (1961 pl E 2) 1055 55 52 16 15 37 35
Measured by SchwarzminusWings(personal communication 2009) 1077 528 49 135 13
Rapetosaurus krausi FMNH PR 2209 Curry Rogers (2009 fig 39) 42 20 48 7 17 16 38Brontomerus mcintoshi OMNH 66430 (this study) 405 223 55 0 0 21 52
1 Hatcher (1901) did not state the length of the ilium of CM 94 and did not figure that of CM 84 We have assumed that the ilium of the figured specimen CM94 is the same size as that of CM 84 (Hatcher 1901 46) and calculated the proportions from the figured specimen
2 Janenschrsquos (1961 pl E) figures of the two ilia of Giraffatitan brancai are not executed from an orthogonal perspective the ilium of HMN Aa 13 is illusminustrated from a slightly anterolateral position foreshortening the preacetabular lobe and that of HMN J 1 from a slight posterolateral position foreshortenminusing the postacetabular lobe The true lengths of the two lobes are probably somewhere between the two percentages calculated from the figures about45 for the former and 16 for the latter
10
cm
Fig 5 Damaged presacral vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah OMNH 66429 in dorsal view as photograph (A) and interpretive drawing (B) Shading indicates air spaces
tion pneumatic excavations and neurocentral fusion (Wedel2003a 248 b 352ndash354) Ontogenetic changes in limbminusgirdleelements such as the ilium are less well understood due to apaucity of sufficiently well preserved specimens (Ray Wilhitepersonal communication October 2007) Therefore the lateralcompression of the Brontomerus ilium may be a juvenile charminusacter with the ilium thickening through ontogeny to supportthe growing weight of the animal or it may in fact be phylominusgenetically significant
Presacral centrummdashA single presacral centrum OMNH66429 was recovered (Fig 5) Unfortunately preservation isvery poor the neural arch and all processes have been lostand the centrum has been greatly crushed dorsoventrally sothat the remaining part is essentially flat the element is 14
cm in both anteroposterior length and transverse width butno more than 3 cm in dorsoventral depth The small size indiminuscates that this element belonged to a juvenile The internalstructure of the centrum is visible however and consists offine septa dividing a hollow internal space irregularly intomany small camellae This morphology is characteristic oftitanosauriforms (Wedel 2003b 354ndash355) Highly camellateinternal structure has not previously been observed in juveminusnile sauropod vertebrae but this may be due to samplingbias so far all juvenile sauropod vertebrae that have beenstudied for internal structure have been those of Camaraminussaurus and diplodocoids which follow an ontogenetic trajecminustory in which large shallow lateral fossae develop into cameminusrae from which smaller accessory camerae and camellae deminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 83
prezygapophysis
neural spine
postzygapophysealramus
postzygapophysealfacet
chevron facets
5 cm
Fig 6 Midminuscaudal vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 61248 in dorsal (A) anterior (B) left lateral (C) posterior (D) and ventral (E) views
velop (Wedel et al 2000a fig 11 Wedel 2003b 349) TheHotel Mesa presacral suggests that camellate vertebrae mayhave developed differently in titanosauriforms possibly byinminussitu formation of camellae during pneumatization as ocminuscurs in the camellate vertebrae of birds (personal observaminustion MJW)
Caudal vertebraemdashOMNH 61248 is a distinctive caudalvertebra with elongated preminus and postzygapophyseal rami(Fig 6) Apart from the tip of the right prezygapophysis theelement is complete and well preserved While the centrum isonly 11 cm in length the distance from the prezygapophysisto postzygapophysis is 145 cm The centrum is slightlybroader than tall (6 cm compared with 55 cm anteriorly 65cm compared with 5 cm posteriorly) and gently waisted Theneural arch is set forward on the centrum but does not reachthe anterior margin The neural spine is so reduced and sostrongly inclined posteriorly as to be all but indistinguishminusable and is apparent only as a very low eminence abovethe postzygapophyses The postzygapophyseal facets themminusselves are set on the posterolateral faces of a low process thathangs below the main postzygapophyseal ramus Chevronfacets are weakly present on the posterior margin of the venminustral surface of the centrum but not on the anterior marginThe elongation index of 22 indicates a midminustominusposterior pominussition in the caudal sequence for this element as similarcentrum proportions do not appear until about caudal 30 inGiraffatitan brancai (Janensch 1950 pl 3)
This vertebra most closely resembles the indeterminatesauropod vertebra BMNH 27500 from the Wessex Formationof the Isle of Wight figured by Naish and Martill (2001 pl33) The Barremian age of that specimen places it about 15 Ma
earlier than OMNH 61248 Its neural arch is less elevated thanthat of the Hotel Mesa specimen its postzygapophyses projectyet farther posteriorly and its prezygapophyses less far anteriminusorly and it is very mildly biconvex rather than procoelous butin other respects including absolute size it is a good match forthe Brontomerus caudal
Also included in the Hotel Mesa material is OMNH27794 a partial distal caudal centrum figured by Wedel(2005 fig 77) This centrum is approximately round incrossminussection about 4 cm in diameter and internally consistsof apneumatic cancellous bone
RibsmdashThe Hotel Mesa material contains several dorsal ribsin various states of preservation but no readily identifiablecervical ribs The dorsal rib elements include the shaft of alarge flat rib (OMNH 27762) portions of several smaller ribshafts (OMNH 27763ndash27765 27768 and others) a flattenedrib head (OMNH 27767) and most informatively a smallcomplete rib (OMNH 27766 Fig 7) Despite its excellentpreservation and apparent lack of distortion this element isdifficult to interpret Its shaft is straight for almost its wholelength and both articular facets are directed dorsally ratherthan being inclined medially The tuberculum is directly inline with the main part of the shaft of the rib and thecapitulum is at an angle of about 30 to it In these respectsthe rib resembles the most anterior dorsal rib of the Diplominusdocus carnegii holotype CM 84 (personal observation MPTthis element was not figured by Hatcher [1901]) We thereminusfore interpret this rib as having probably belonged to the rightside of the first dorsal vertebra
The rib is unusual in other respects however most notablythat the ventral part of the shaft curves laterally rather than meminus
84 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
10 cm
capitulum
pneumatic fossa
tuberculum
Fig 7 First right dorsal rib of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah OMNH 27766 in posterior view head of rib showing pneumatic invasion of shaft (A) and complete rib showing laterally directed curvature of shaft (B)
dially Careful inspection of the bone reveals no indication ofdistortion or of incorrect reconstruction It may be possiblethat in life the thorax was transversely compressed so that thedorsal part of the rib shaft was directed ventromedially and themore ventral part was vertical Both articular facets are subminuscircular in dorsal view and significantly expanded comparedwith the rami that bear them On the anterior face of the head alow ridge arises just below the capitulum and extends downthe medial edge of the rib for about 40 of its length
The head of the rib is also unusual in that a thin sheet ofbone connects the rami that support the articular facets andthis sheet extends much farther proximally than in mostsauropod ribs Its precise extent cannot be ascertained due tobreakage The sheet of bone is perforated close to the capituminuslar ramus and from this perforation a pneumatic cavity inminusvades the shaft of the rib extending ventrally from within ashallow fossa in the posterior face Pneumatization of thedorsal ribs is a synapomorphy of Titanosauriformes (Wilsonand Sereno 1998) although pneumatic dorsal ribs are also inminusfrequently present in diplodocids (Gilmore 1936 Lovelaceet al 2003 2008)
Pectoral girdlemdashOMNH 27761 is a partial scapula consistminusing of the blade and part of the anterior expansion but missminusing the glenoid region and the remainder of the anterior exminuspansion (Fig 8) As preserved the element is nearly flat butthis may be due to postminusmortem distortion and in any casethe most strongly curved part of most sauropod scapulae isthe anterior part that is missing from this specimen The genminustle curvature preserved in the posterior part of the blade indiminuscates that the element was from the left side The bone is surminusprisingly thin in all preserved parts never exceeding a few
cm in contrast to for example the scapula of Camarasaurussupremus which is thick even in midminusblade (Osborn andMook 1921 fig 74b) This suggests that the glenoid thickenminusing and the acromial ridge may have been located some disminustance anterior to the preserved portion and a reconstructionafter the proportions of Giraffatitan brancai (Fig 8) suggeststhat about 80 of the scapularsquos full length is preserved Theposterior part of the acromion expansion is preserved howminusever and is sufficient to show that this expansion was prominusnounced so that the maximum dorsoventral height of thescapula was more than two and half times its minimumheight at the midpoint of the blade The dorsal margin slopesup towards the anterior expansion rather than forming a posminusteriorly directed ldquohookrdquo or a distinct acromion fossa betweenthe blade and the acromion process
The posterior expansion of the scapula is distinctive Insome sauropods the posterior part of the scapular blade isexpanded not at all or only slightly for example in Omeiminussaurus Young 1939 (He et al 1988 fig 41) Apatosaurus(Upchurch et al 2004b fig 4) and Rapetosaurus (Fig 9E)In others the ventral margin of the scapular blade is straightor nearly so while the dorsal margin is deflected dorsally tocreate an asymmetric expansion for example in Camaraminussaurus (Fig 9C) and Giraffatitan brancai (Fig 9D) In a fewsauropods however the ventral margin of the blade is alsodeflected ventrally to form a ldquoracquetminusshapedrdquo posterior exminuspansion This is seen in rebbachisaurids and some titanominussaurians eg some specimens of Alamosaurus Gilmore1922 (Gilmore 1946 fig 6) In Brontomerus the posteriorpart of the scapular blade is expanded in a characteristic manminusner the ventral margin is straight except for a posteroventral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 85
50 cm
ventral ldquosteprdquo
acromion expansion
dorsal ldquostepsrdquo
Fig 8 Partial left scapula of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 27761 in lateral view tentatively reconstructed after Giraffatitan brancai HMN Sa 9 (Janensch 1961 pl 15 1)
excursion two thirds of the way along the preserved portionafter which the margin continues parallel to its original traminusjectory so that the excursion appears as a gentle ldquosteprdquo Thedorsal margin is also ldquosteppedrdquo in this manner though withtwo distinct steps rather than one of which the more anterioris most strongly pronounced The net result of these featuresis that the dorsal and ventral borders of the scapula are bothstraight near the posterior extremity and that they are subminusparallel diverging by only about five degrees in the regionjust anterior to the rounded end of the posterior expansionThe step in the ventral border is not known in any othersauropod however the scapula of Neuquensaurus Powell1992 has a stepped dorsal border similar to that of Brontominusmerus (Huene 1929 via Glut 1997 275)
These characters of the scapula must be treated with somecaution however since this bone appears subject to morevariation than any other in the sauropod skeleton see for exminusample the range of shapes in scapulae of Giraffatitan brancai(Janensch 1961 pl 15 1ndash3) and in Camarasaurus supremus(Osborn and Mook 1921 figs 74ndash80)
Two small flat elements OMNH 66431 and 66432 are inminusterpreted as partial sternal plates (Fig 10) The medial edge ofeach is identifiable due to its rugose texture which formed theattachment site for cartilage joining the plates to each otherand to the sternal ribs The sternals are anteroposteriorly elonminusgate and mediolaterally narrow when complete they wereprobably at least three times as long as broad as in ldquoSaltaminussaurusrdquo robustus Huene 1929 (Huene 1929 via McIntosh1990 fig 169L) and proportionally longer than in any othersauropod including Saltasaurus loricatus (McIntosh 1990fig 169 Powell 1992 fig 30) The sternals are crescentic in
shape the anterior and posterior extremities curving laterallyaway from the midline This state was considered a titanominussaurian synapomorphy by Wilson (2002 268) but its distribuminustion is more complex in the current analysis being synapominusmorphic for Neosauropoda with losses in Flagellicaudata andCamarasaurus
86 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
Fig 10 Partial paired sternal plates of the camarasauromorph sauropodBrontomerus mcintoshi gen et sp nov from the Lower Cretaceous CedarMountain Formation of Utah OMNH 66431 and 66432 in ventral view
Fig 9 Scapulocoracoids and scapulae of sauropod dinosaurs scaled to same length of scapular blade from posterior point of glenoid to posterior margin ofblade A Mamenchisaurus youngi Young and Zhao 1972 holotype ZDM0083 left scapulocoracoid modified from Ouyang and Ye (2002 fig 22)B Diplodocus longus Hatcher 1901 USNM 10865 right scapulocoracoid reversed photograph by MPT C Camarasaurus supremus Cope 1877 AMNH5761 Sc 1 left scapula and AMNH 5761 Cor 1 left coracoid probably associated modified from Osborn and Mook (1921 figs 75 81a) D Giraffatitanbrancai (Janensch 1914) HMN Sa 9 left scapula modified from Janensch (1961 pl 15 1) E Rapetosaurus krausi Curry Rogers and Forster 2001holotype FMNH PR 2209 right scapula reversed modified from Curry Rogers (2009 fig 32) F Brontomerus mcintoshi gen et sp nov OMNH 27761left scapula tentatively reconstructed after Giraffatitan brancai
Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

The assignment of specimen numbers to the material deminusscribed here is complex (Table 3) Specimen number OMNH27773 comprises three elements OMNH 27784 consists of 21small fragments of bone none of them informative all otherelements have their own specimen numbers in the range27761ndash27800 apart from the midminuscaudal vertebra OMNH61248 and the reassigned numbers 66429ndash66432 for eleminusments extracted from OMNH 27773
IliummdashThe most informative element is OMNH 66430 a leftilium (Fig 2) which has therefore been selected as the holominustype The ilium was preserved complete but lay hidden beminusneath the scapula and so was damaged in the field (JamesKirkland personal communication March 2008) The iliumis preserved in three parts one provides most of the boneincluding the well preserved preacetabular lobe pubic andischiadic peduncles and acetabular margin and the other two
provide most of the dorsal margin giving a good indication ofthe degree of curvature The relative positions and orientationof the two smaller fragments can not be definitely ascertainedbut they appear to be parts of a single large fragment broken atthe point where we have reconstructed them as touching andif this interpretation is correct then the curvature of the pair inminusdicates which side must be oriented laterally
The ilium is remarkable in that the preacetabular lobe isrelatively larger than in any other sauropod (Fig 3 Table 4measurement protocol is illustrated in Fig 4) the postaceminustabular lobe is reduced almost to the point of absence and theilium is proportionally taller than in any other sauropod Theischiadic peduncle is reduced to a very low ventral projectionfrom almost the most posterior point of the ilium The near abminussence of the ischiadic peduncle cannot be attributed to damageas the iliac articular surface is preserved Immediately posterominus
80 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
acetabulum
pubicpeduncle
10 cm
ischiadicpeduncle
pre-acetabularblade
Fig 2 Left ilium of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah type specimen OMNH 66430 in lateral view reconstructed from the three fragments (A) and ventral view (B)
dorsal to this surface is a subtle notch between the peduncleand the very reduced postacetabular lobe This notch and theareas either side of it are composed of finished bone demonminusstrating that the great reduction of the postacetabular lobe toois a genuine osteological feature and not due to damage In reminusgard to the proportionally large preacetabular lobe the iliumof Brontomerus resembles that of Rapetosaurus (Fig 3E Taminusble 4) However that taxon has a normal postacetabular lobeand is not proportionally tall In overall proportions the iliumof Brontomerus is more similar to the left ilium HMN J1 asminussigned to Giraffatitan brancai (Janensch 1961 pl E 2) whichalso has a reduced postacetabular lobe mdashsee also Fig 3DHowever the ilium of Brontomerus is proportionally tallerthan that of G brancai and its anterior margin comes to apoint rather than being smoothly rounded as in that taxon InBrontomerus the maximum height of the ilium above theacetabular margin when measured perpendicular to the lonminusgest axis is 52 in other sauropods examined this proportiondoes not exceed 45 and averages 40 (Table 4)
As with many sauropods the preacetabular lobe of theilium flares laterally However in most sauropods this flarminusing is progressive so that in dorsal or ventral view the mostposterior part of the preacetabular lobe is nearly parallel witha line drawn between the pubic and ischiadic peduncles andsmooth lateral curvature inclines the more anterior parts inminuscreasingly laterally so that the more anterior part is almost atright angles to this line and the ilium appears smoothly
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 81
Fig 3 Ilia of sauropod dinosaurs scaled to same total length A Mamenchisaurus hochuanensis Young and Zhao 1972 holotype CCG V 20401 right iliumreversed modified from Young and Zhao (1972 pl 6 1a) B Diplodocus carnegii Hatcher 1901 CM 94 right ilium reversed modified from Hatcher (1901pl 10 1) C Camarasaurus supremus Cope 1877 AMNH 5761 Il 1 left ilium modified from Osborn and Mook (1921 fig 87) D Giraffatitan brancai(Janensch 1914) HMN J1 left ilium modified from Janensch (1961 pl E 2) E Rapetosaurus krausi Curry Rogers and Forster 2001 holotype FMNH PR2209 left ilium modified from Curry Rogers (2009 fig 39B) F Brontomerus mcintoshi gen et sp nov holotype OMNH 66430 left ilium
Fig 4 Measurement protocol for sauropod ilia as illustrated in Fig 3 andshown in Table 4 Total length is measured along the longest axis of theilium lengths of preacetabular and postacetabular lobes are measured parminusallel to this axis and extend from the extremity of the lobe to the anteriormargin of the pubic peduncle and posterior margin of the ischiadic pedunclerespectively Supracetabular height is measured perpendicular to the lonminusgest axis and extends from the highest point of the acetabulum to the pointlevel with the highest part of the ilium
curved in dorsal or ventral viewmdashfor example ApatosaurusMarsh 1877 (Upchurch et al 2004b pl 4 D E) Haplominuscanthosaurus Hatcher 1903b (Hatcher 1903a pl 5 1) andSaltasaurus Bonaparte and Powell 1980 (Powell 1992 fig17) In Brontomerus by contrast the blade of the ilium apminuspears to be ldquohingedrdquomdashdeflected laterally at a point directlyanterior to the public pedunclemdashso that the preacetabularlobe is straight in dorsal or ventral view and directed anterominuslaterally by an angle of about 30 to the sagittal In this reminusspect it more closely resembles the ilium of Camarasaurus
Cope 1877 (Osborn and Mook 1921 fig 49) although it difminusfers in other respects
The Brontomerus ilium is laterally compressed and unlikemost sauropod ilia the dorsal margin is not deflected laterallyrelative to the more ventral part so that in ventral view it apminuspears very thin (Fig 2B) It is well established that the longbones of sauropods grow isometrically through ontogeny(Carpenter and McIntosh 1994 Wilhite 1999 2003 Ikejiri etal 2005 Tidwell and Wilhite 2005 Taylor 2009) while theirvertebrae undergo significant changes in proportions laminaminus
82 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 4 Relative measurements of ilia in sauropods Measurement protocol is illustrated in Fig 4
Taxon Specimen ReferenceTotallength(cm)
Length ofpreacetabular
lobe (cmproportion oftotal length)
Length ofpostacetabular
lobe (cmproportion oftotal length)
Supracetabularheight (cm
proportion oftotal length)
Mamenchisaurushochuanensis
CCG V 20401 Young and Zhao (1972 pl 6 1a) 102 39 38 18 18 41 40
Diplodocus carnegii CM 94 Hatcher (1901 pl 10 1) 1091 41 38 20 18 49 45Camarasaurus
supremusAMNH 5761 Il 1 Osborn and Mook (1921 fig 87) 115 36 31 19 17 48 42
Giraffatitan brancai2
HMN Aa 13Janensch (1961 pl E 1a) 119 47 39 21 18 46 39
Measured by SchwarzminusWings(personal communication 2009) 1143 535 47 299 26
HMN J1Janensch (1961 pl E 2) 1055 55 52 16 15 37 35
Measured by SchwarzminusWings(personal communication 2009) 1077 528 49 135 13
Rapetosaurus krausi FMNH PR 2209 Curry Rogers (2009 fig 39) 42 20 48 7 17 16 38Brontomerus mcintoshi OMNH 66430 (this study) 405 223 55 0 0 21 52
1 Hatcher (1901) did not state the length of the ilium of CM 94 and did not figure that of CM 84 We have assumed that the ilium of the figured specimen CM94 is the same size as that of CM 84 (Hatcher 1901 46) and calculated the proportions from the figured specimen
2 Janenschrsquos (1961 pl E) figures of the two ilia of Giraffatitan brancai are not executed from an orthogonal perspective the ilium of HMN Aa 13 is illusminustrated from a slightly anterolateral position foreshortening the preacetabular lobe and that of HMN J 1 from a slight posterolateral position foreshortenminusing the postacetabular lobe The true lengths of the two lobes are probably somewhere between the two percentages calculated from the figures about45 for the former and 16 for the latter
10
cm
Fig 5 Damaged presacral vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah OMNH 66429 in dorsal view as photograph (A) and interpretive drawing (B) Shading indicates air spaces
tion pneumatic excavations and neurocentral fusion (Wedel2003a 248 b 352ndash354) Ontogenetic changes in limbminusgirdleelements such as the ilium are less well understood due to apaucity of sufficiently well preserved specimens (Ray Wilhitepersonal communication October 2007) Therefore the lateralcompression of the Brontomerus ilium may be a juvenile charminusacter with the ilium thickening through ontogeny to supportthe growing weight of the animal or it may in fact be phylominusgenetically significant
Presacral centrummdashA single presacral centrum OMNH66429 was recovered (Fig 5) Unfortunately preservation isvery poor the neural arch and all processes have been lostand the centrum has been greatly crushed dorsoventrally sothat the remaining part is essentially flat the element is 14
cm in both anteroposterior length and transverse width butno more than 3 cm in dorsoventral depth The small size indiminuscates that this element belonged to a juvenile The internalstructure of the centrum is visible however and consists offine septa dividing a hollow internal space irregularly intomany small camellae This morphology is characteristic oftitanosauriforms (Wedel 2003b 354ndash355) Highly camellateinternal structure has not previously been observed in juveminusnile sauropod vertebrae but this may be due to samplingbias so far all juvenile sauropod vertebrae that have beenstudied for internal structure have been those of Camaraminussaurus and diplodocoids which follow an ontogenetic trajecminustory in which large shallow lateral fossae develop into cameminusrae from which smaller accessory camerae and camellae deminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 83
prezygapophysis
neural spine
postzygapophysealramus
postzygapophysealfacet
chevron facets
5 cm
Fig 6 Midminuscaudal vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 61248 in dorsal (A) anterior (B) left lateral (C) posterior (D) and ventral (E) views
velop (Wedel et al 2000a fig 11 Wedel 2003b 349) TheHotel Mesa presacral suggests that camellate vertebrae mayhave developed differently in titanosauriforms possibly byinminussitu formation of camellae during pneumatization as ocminuscurs in the camellate vertebrae of birds (personal observaminustion MJW)
Caudal vertebraemdashOMNH 61248 is a distinctive caudalvertebra with elongated preminus and postzygapophyseal rami(Fig 6) Apart from the tip of the right prezygapophysis theelement is complete and well preserved While the centrum isonly 11 cm in length the distance from the prezygapophysisto postzygapophysis is 145 cm The centrum is slightlybroader than tall (6 cm compared with 55 cm anteriorly 65cm compared with 5 cm posteriorly) and gently waisted Theneural arch is set forward on the centrum but does not reachthe anterior margin The neural spine is so reduced and sostrongly inclined posteriorly as to be all but indistinguishminusable and is apparent only as a very low eminence abovethe postzygapophyses The postzygapophyseal facets themminusselves are set on the posterolateral faces of a low process thathangs below the main postzygapophyseal ramus Chevronfacets are weakly present on the posterior margin of the venminustral surface of the centrum but not on the anterior marginThe elongation index of 22 indicates a midminustominusposterior pominussition in the caudal sequence for this element as similarcentrum proportions do not appear until about caudal 30 inGiraffatitan brancai (Janensch 1950 pl 3)
This vertebra most closely resembles the indeterminatesauropod vertebra BMNH 27500 from the Wessex Formationof the Isle of Wight figured by Naish and Martill (2001 pl33) The Barremian age of that specimen places it about 15 Ma
earlier than OMNH 61248 Its neural arch is less elevated thanthat of the Hotel Mesa specimen its postzygapophyses projectyet farther posteriorly and its prezygapophyses less far anteriminusorly and it is very mildly biconvex rather than procoelous butin other respects including absolute size it is a good match forthe Brontomerus caudal
Also included in the Hotel Mesa material is OMNH27794 a partial distal caudal centrum figured by Wedel(2005 fig 77) This centrum is approximately round incrossminussection about 4 cm in diameter and internally consistsof apneumatic cancellous bone
RibsmdashThe Hotel Mesa material contains several dorsal ribsin various states of preservation but no readily identifiablecervical ribs The dorsal rib elements include the shaft of alarge flat rib (OMNH 27762) portions of several smaller ribshafts (OMNH 27763ndash27765 27768 and others) a flattenedrib head (OMNH 27767) and most informatively a smallcomplete rib (OMNH 27766 Fig 7) Despite its excellentpreservation and apparent lack of distortion this element isdifficult to interpret Its shaft is straight for almost its wholelength and both articular facets are directed dorsally ratherthan being inclined medially The tuberculum is directly inline with the main part of the shaft of the rib and thecapitulum is at an angle of about 30 to it In these respectsthe rib resembles the most anterior dorsal rib of the Diplominusdocus carnegii holotype CM 84 (personal observation MPTthis element was not figured by Hatcher [1901]) We thereminusfore interpret this rib as having probably belonged to the rightside of the first dorsal vertebra
The rib is unusual in other respects however most notablythat the ventral part of the shaft curves laterally rather than meminus
84 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
10 cm
capitulum
pneumatic fossa
tuberculum
Fig 7 First right dorsal rib of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah OMNH 27766 in posterior view head of rib showing pneumatic invasion of shaft (A) and complete rib showing laterally directed curvature of shaft (B)
dially Careful inspection of the bone reveals no indication ofdistortion or of incorrect reconstruction It may be possiblethat in life the thorax was transversely compressed so that thedorsal part of the rib shaft was directed ventromedially and themore ventral part was vertical Both articular facets are subminuscircular in dorsal view and significantly expanded comparedwith the rami that bear them On the anterior face of the head alow ridge arises just below the capitulum and extends downthe medial edge of the rib for about 40 of its length
The head of the rib is also unusual in that a thin sheet ofbone connects the rami that support the articular facets andthis sheet extends much farther proximally than in mostsauropod ribs Its precise extent cannot be ascertained due tobreakage The sheet of bone is perforated close to the capituminuslar ramus and from this perforation a pneumatic cavity inminusvades the shaft of the rib extending ventrally from within ashallow fossa in the posterior face Pneumatization of thedorsal ribs is a synapomorphy of Titanosauriformes (Wilsonand Sereno 1998) although pneumatic dorsal ribs are also inminusfrequently present in diplodocids (Gilmore 1936 Lovelaceet al 2003 2008)
Pectoral girdlemdashOMNH 27761 is a partial scapula consistminusing of the blade and part of the anterior expansion but missminusing the glenoid region and the remainder of the anterior exminuspansion (Fig 8) As preserved the element is nearly flat butthis may be due to postminusmortem distortion and in any casethe most strongly curved part of most sauropod scapulae isthe anterior part that is missing from this specimen The genminustle curvature preserved in the posterior part of the blade indiminuscates that the element was from the left side The bone is surminusprisingly thin in all preserved parts never exceeding a few
cm in contrast to for example the scapula of Camarasaurussupremus which is thick even in midminusblade (Osborn andMook 1921 fig 74b) This suggests that the glenoid thickenminusing and the acromial ridge may have been located some disminustance anterior to the preserved portion and a reconstructionafter the proportions of Giraffatitan brancai (Fig 8) suggeststhat about 80 of the scapularsquos full length is preserved Theposterior part of the acromion expansion is preserved howminusever and is sufficient to show that this expansion was prominusnounced so that the maximum dorsoventral height of thescapula was more than two and half times its minimumheight at the midpoint of the blade The dorsal margin slopesup towards the anterior expansion rather than forming a posminusteriorly directed ldquohookrdquo or a distinct acromion fossa betweenthe blade and the acromion process
The posterior expansion of the scapula is distinctive Insome sauropods the posterior part of the scapular blade isexpanded not at all or only slightly for example in Omeiminussaurus Young 1939 (He et al 1988 fig 41) Apatosaurus(Upchurch et al 2004b fig 4) and Rapetosaurus (Fig 9E)In others the ventral margin of the scapular blade is straightor nearly so while the dorsal margin is deflected dorsally tocreate an asymmetric expansion for example in Camaraminussaurus (Fig 9C) and Giraffatitan brancai (Fig 9D) In a fewsauropods however the ventral margin of the blade is alsodeflected ventrally to form a ldquoracquetminusshapedrdquo posterior exminuspansion This is seen in rebbachisaurids and some titanominussaurians eg some specimens of Alamosaurus Gilmore1922 (Gilmore 1946 fig 6) In Brontomerus the posteriorpart of the scapular blade is expanded in a characteristic manminusner the ventral margin is straight except for a posteroventral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 85
50 cm
ventral ldquosteprdquo
acromion expansion
dorsal ldquostepsrdquo
Fig 8 Partial left scapula of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 27761 in lateral view tentatively reconstructed after Giraffatitan brancai HMN Sa 9 (Janensch 1961 pl 15 1)
excursion two thirds of the way along the preserved portionafter which the margin continues parallel to its original traminusjectory so that the excursion appears as a gentle ldquosteprdquo Thedorsal margin is also ldquosteppedrdquo in this manner though withtwo distinct steps rather than one of which the more anterioris most strongly pronounced The net result of these featuresis that the dorsal and ventral borders of the scapula are bothstraight near the posterior extremity and that they are subminusparallel diverging by only about five degrees in the regionjust anterior to the rounded end of the posterior expansionThe step in the ventral border is not known in any othersauropod however the scapula of Neuquensaurus Powell1992 has a stepped dorsal border similar to that of Brontominusmerus (Huene 1929 via Glut 1997 275)
These characters of the scapula must be treated with somecaution however since this bone appears subject to morevariation than any other in the sauropod skeleton see for exminusample the range of shapes in scapulae of Giraffatitan brancai(Janensch 1961 pl 15 1ndash3) and in Camarasaurus supremus(Osborn and Mook 1921 figs 74ndash80)
Two small flat elements OMNH 66431 and 66432 are inminusterpreted as partial sternal plates (Fig 10) The medial edge ofeach is identifiable due to its rugose texture which formed theattachment site for cartilage joining the plates to each otherand to the sternal ribs The sternals are anteroposteriorly elonminusgate and mediolaterally narrow when complete they wereprobably at least three times as long as broad as in ldquoSaltaminussaurusrdquo robustus Huene 1929 (Huene 1929 via McIntosh1990 fig 169L) and proportionally longer than in any othersauropod including Saltasaurus loricatus (McIntosh 1990fig 169 Powell 1992 fig 30) The sternals are crescentic in
shape the anterior and posterior extremities curving laterallyaway from the midline This state was considered a titanominussaurian synapomorphy by Wilson (2002 268) but its distribuminustion is more complex in the current analysis being synapominusmorphic for Neosauropoda with losses in Flagellicaudata andCamarasaurus
86 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
Fig 10 Partial paired sternal plates of the camarasauromorph sauropodBrontomerus mcintoshi gen et sp nov from the Lower Cretaceous CedarMountain Formation of Utah OMNH 66431 and 66432 in ventral view
Fig 9 Scapulocoracoids and scapulae of sauropod dinosaurs scaled to same length of scapular blade from posterior point of glenoid to posterior margin ofblade A Mamenchisaurus youngi Young and Zhao 1972 holotype ZDM0083 left scapulocoracoid modified from Ouyang and Ye (2002 fig 22)B Diplodocus longus Hatcher 1901 USNM 10865 right scapulocoracoid reversed photograph by MPT C Camarasaurus supremus Cope 1877 AMNH5761 Sc 1 left scapula and AMNH 5761 Cor 1 left coracoid probably associated modified from Osborn and Mook (1921 figs 75 81a) D Giraffatitanbrancai (Janensch 1914) HMN Sa 9 left scapula modified from Janensch (1961 pl 15 1) E Rapetosaurus krausi Curry Rogers and Forster 2001holotype FMNH PR 2209 right scapula reversed modified from Curry Rogers (2009 fig 32) F Brontomerus mcintoshi gen et sp nov OMNH 27761left scapula tentatively reconstructed after Giraffatitan brancai
Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

dorsal to this surface is a subtle notch between the peduncleand the very reduced postacetabular lobe This notch and theareas either side of it are composed of finished bone demonminusstrating that the great reduction of the postacetabular lobe toois a genuine osteological feature and not due to damage In reminusgard to the proportionally large preacetabular lobe the iliumof Brontomerus resembles that of Rapetosaurus (Fig 3E Taminusble 4) However that taxon has a normal postacetabular lobeand is not proportionally tall In overall proportions the iliumof Brontomerus is more similar to the left ilium HMN J1 asminussigned to Giraffatitan brancai (Janensch 1961 pl E 2) whichalso has a reduced postacetabular lobe mdashsee also Fig 3DHowever the ilium of Brontomerus is proportionally tallerthan that of G brancai and its anterior margin comes to apoint rather than being smoothly rounded as in that taxon InBrontomerus the maximum height of the ilium above theacetabular margin when measured perpendicular to the lonminusgest axis is 52 in other sauropods examined this proportiondoes not exceed 45 and averages 40 (Table 4)
As with many sauropods the preacetabular lobe of theilium flares laterally However in most sauropods this flarminusing is progressive so that in dorsal or ventral view the mostposterior part of the preacetabular lobe is nearly parallel witha line drawn between the pubic and ischiadic peduncles andsmooth lateral curvature inclines the more anterior parts inminuscreasingly laterally so that the more anterior part is almost atright angles to this line and the ilium appears smoothly
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 81
Fig 3 Ilia of sauropod dinosaurs scaled to same total length A Mamenchisaurus hochuanensis Young and Zhao 1972 holotype CCG V 20401 right iliumreversed modified from Young and Zhao (1972 pl 6 1a) B Diplodocus carnegii Hatcher 1901 CM 94 right ilium reversed modified from Hatcher (1901pl 10 1) C Camarasaurus supremus Cope 1877 AMNH 5761 Il 1 left ilium modified from Osborn and Mook (1921 fig 87) D Giraffatitan brancai(Janensch 1914) HMN J1 left ilium modified from Janensch (1961 pl E 2) E Rapetosaurus krausi Curry Rogers and Forster 2001 holotype FMNH PR2209 left ilium modified from Curry Rogers (2009 fig 39B) F Brontomerus mcintoshi gen et sp nov holotype OMNH 66430 left ilium
Fig 4 Measurement protocol for sauropod ilia as illustrated in Fig 3 andshown in Table 4 Total length is measured along the longest axis of theilium lengths of preacetabular and postacetabular lobes are measured parminusallel to this axis and extend from the extremity of the lobe to the anteriormargin of the pubic peduncle and posterior margin of the ischiadic pedunclerespectively Supracetabular height is measured perpendicular to the lonminusgest axis and extends from the highest point of the acetabulum to the pointlevel with the highest part of the ilium
curved in dorsal or ventral viewmdashfor example ApatosaurusMarsh 1877 (Upchurch et al 2004b pl 4 D E) Haplominuscanthosaurus Hatcher 1903b (Hatcher 1903a pl 5 1) andSaltasaurus Bonaparte and Powell 1980 (Powell 1992 fig17) In Brontomerus by contrast the blade of the ilium apminuspears to be ldquohingedrdquomdashdeflected laterally at a point directlyanterior to the public pedunclemdashso that the preacetabularlobe is straight in dorsal or ventral view and directed anterominuslaterally by an angle of about 30 to the sagittal In this reminusspect it more closely resembles the ilium of Camarasaurus
Cope 1877 (Osborn and Mook 1921 fig 49) although it difminusfers in other respects
The Brontomerus ilium is laterally compressed and unlikemost sauropod ilia the dorsal margin is not deflected laterallyrelative to the more ventral part so that in ventral view it apminuspears very thin (Fig 2B) It is well established that the longbones of sauropods grow isometrically through ontogeny(Carpenter and McIntosh 1994 Wilhite 1999 2003 Ikejiri etal 2005 Tidwell and Wilhite 2005 Taylor 2009) while theirvertebrae undergo significant changes in proportions laminaminus
82 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 4 Relative measurements of ilia in sauropods Measurement protocol is illustrated in Fig 4
Taxon Specimen ReferenceTotallength(cm)
Length ofpreacetabular
lobe (cmproportion oftotal length)
Length ofpostacetabular
lobe (cmproportion oftotal length)
Supracetabularheight (cm
proportion oftotal length)
Mamenchisaurushochuanensis
CCG V 20401 Young and Zhao (1972 pl 6 1a) 102 39 38 18 18 41 40
Diplodocus carnegii CM 94 Hatcher (1901 pl 10 1) 1091 41 38 20 18 49 45Camarasaurus
supremusAMNH 5761 Il 1 Osborn and Mook (1921 fig 87) 115 36 31 19 17 48 42
Giraffatitan brancai2
HMN Aa 13Janensch (1961 pl E 1a) 119 47 39 21 18 46 39
Measured by SchwarzminusWings(personal communication 2009) 1143 535 47 299 26
HMN J1Janensch (1961 pl E 2) 1055 55 52 16 15 37 35
Measured by SchwarzminusWings(personal communication 2009) 1077 528 49 135 13
Rapetosaurus krausi FMNH PR 2209 Curry Rogers (2009 fig 39) 42 20 48 7 17 16 38Brontomerus mcintoshi OMNH 66430 (this study) 405 223 55 0 0 21 52
1 Hatcher (1901) did not state the length of the ilium of CM 94 and did not figure that of CM 84 We have assumed that the ilium of the figured specimen CM94 is the same size as that of CM 84 (Hatcher 1901 46) and calculated the proportions from the figured specimen
2 Janenschrsquos (1961 pl E) figures of the two ilia of Giraffatitan brancai are not executed from an orthogonal perspective the ilium of HMN Aa 13 is illusminustrated from a slightly anterolateral position foreshortening the preacetabular lobe and that of HMN J 1 from a slight posterolateral position foreshortenminusing the postacetabular lobe The true lengths of the two lobes are probably somewhere between the two percentages calculated from the figures about45 for the former and 16 for the latter
10
cm
Fig 5 Damaged presacral vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah OMNH 66429 in dorsal view as photograph (A) and interpretive drawing (B) Shading indicates air spaces
tion pneumatic excavations and neurocentral fusion (Wedel2003a 248 b 352ndash354) Ontogenetic changes in limbminusgirdleelements such as the ilium are less well understood due to apaucity of sufficiently well preserved specimens (Ray Wilhitepersonal communication October 2007) Therefore the lateralcompression of the Brontomerus ilium may be a juvenile charminusacter with the ilium thickening through ontogeny to supportthe growing weight of the animal or it may in fact be phylominusgenetically significant
Presacral centrummdashA single presacral centrum OMNH66429 was recovered (Fig 5) Unfortunately preservation isvery poor the neural arch and all processes have been lostand the centrum has been greatly crushed dorsoventrally sothat the remaining part is essentially flat the element is 14
cm in both anteroposterior length and transverse width butno more than 3 cm in dorsoventral depth The small size indiminuscates that this element belonged to a juvenile The internalstructure of the centrum is visible however and consists offine septa dividing a hollow internal space irregularly intomany small camellae This morphology is characteristic oftitanosauriforms (Wedel 2003b 354ndash355) Highly camellateinternal structure has not previously been observed in juveminusnile sauropod vertebrae but this may be due to samplingbias so far all juvenile sauropod vertebrae that have beenstudied for internal structure have been those of Camaraminussaurus and diplodocoids which follow an ontogenetic trajecminustory in which large shallow lateral fossae develop into cameminusrae from which smaller accessory camerae and camellae deminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 83
prezygapophysis
neural spine
postzygapophysealramus
postzygapophysealfacet
chevron facets
5 cm
Fig 6 Midminuscaudal vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 61248 in dorsal (A) anterior (B) left lateral (C) posterior (D) and ventral (E) views
velop (Wedel et al 2000a fig 11 Wedel 2003b 349) TheHotel Mesa presacral suggests that camellate vertebrae mayhave developed differently in titanosauriforms possibly byinminussitu formation of camellae during pneumatization as ocminuscurs in the camellate vertebrae of birds (personal observaminustion MJW)
Caudal vertebraemdashOMNH 61248 is a distinctive caudalvertebra with elongated preminus and postzygapophyseal rami(Fig 6) Apart from the tip of the right prezygapophysis theelement is complete and well preserved While the centrum isonly 11 cm in length the distance from the prezygapophysisto postzygapophysis is 145 cm The centrum is slightlybroader than tall (6 cm compared with 55 cm anteriorly 65cm compared with 5 cm posteriorly) and gently waisted Theneural arch is set forward on the centrum but does not reachthe anterior margin The neural spine is so reduced and sostrongly inclined posteriorly as to be all but indistinguishminusable and is apparent only as a very low eminence abovethe postzygapophyses The postzygapophyseal facets themminusselves are set on the posterolateral faces of a low process thathangs below the main postzygapophyseal ramus Chevronfacets are weakly present on the posterior margin of the venminustral surface of the centrum but not on the anterior marginThe elongation index of 22 indicates a midminustominusposterior pominussition in the caudal sequence for this element as similarcentrum proportions do not appear until about caudal 30 inGiraffatitan brancai (Janensch 1950 pl 3)
This vertebra most closely resembles the indeterminatesauropod vertebra BMNH 27500 from the Wessex Formationof the Isle of Wight figured by Naish and Martill (2001 pl33) The Barremian age of that specimen places it about 15 Ma
earlier than OMNH 61248 Its neural arch is less elevated thanthat of the Hotel Mesa specimen its postzygapophyses projectyet farther posteriorly and its prezygapophyses less far anteriminusorly and it is very mildly biconvex rather than procoelous butin other respects including absolute size it is a good match forthe Brontomerus caudal
Also included in the Hotel Mesa material is OMNH27794 a partial distal caudal centrum figured by Wedel(2005 fig 77) This centrum is approximately round incrossminussection about 4 cm in diameter and internally consistsof apneumatic cancellous bone
RibsmdashThe Hotel Mesa material contains several dorsal ribsin various states of preservation but no readily identifiablecervical ribs The dorsal rib elements include the shaft of alarge flat rib (OMNH 27762) portions of several smaller ribshafts (OMNH 27763ndash27765 27768 and others) a flattenedrib head (OMNH 27767) and most informatively a smallcomplete rib (OMNH 27766 Fig 7) Despite its excellentpreservation and apparent lack of distortion this element isdifficult to interpret Its shaft is straight for almost its wholelength and both articular facets are directed dorsally ratherthan being inclined medially The tuberculum is directly inline with the main part of the shaft of the rib and thecapitulum is at an angle of about 30 to it In these respectsthe rib resembles the most anterior dorsal rib of the Diplominusdocus carnegii holotype CM 84 (personal observation MPTthis element was not figured by Hatcher [1901]) We thereminusfore interpret this rib as having probably belonged to the rightside of the first dorsal vertebra
The rib is unusual in other respects however most notablythat the ventral part of the shaft curves laterally rather than meminus
84 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
10 cm
capitulum
pneumatic fossa
tuberculum
Fig 7 First right dorsal rib of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah OMNH 27766 in posterior view head of rib showing pneumatic invasion of shaft (A) and complete rib showing laterally directed curvature of shaft (B)
dially Careful inspection of the bone reveals no indication ofdistortion or of incorrect reconstruction It may be possiblethat in life the thorax was transversely compressed so that thedorsal part of the rib shaft was directed ventromedially and themore ventral part was vertical Both articular facets are subminuscircular in dorsal view and significantly expanded comparedwith the rami that bear them On the anterior face of the head alow ridge arises just below the capitulum and extends downthe medial edge of the rib for about 40 of its length
The head of the rib is also unusual in that a thin sheet ofbone connects the rami that support the articular facets andthis sheet extends much farther proximally than in mostsauropod ribs Its precise extent cannot be ascertained due tobreakage The sheet of bone is perforated close to the capituminuslar ramus and from this perforation a pneumatic cavity inminusvades the shaft of the rib extending ventrally from within ashallow fossa in the posterior face Pneumatization of thedorsal ribs is a synapomorphy of Titanosauriformes (Wilsonand Sereno 1998) although pneumatic dorsal ribs are also inminusfrequently present in diplodocids (Gilmore 1936 Lovelaceet al 2003 2008)
Pectoral girdlemdashOMNH 27761 is a partial scapula consistminusing of the blade and part of the anterior expansion but missminusing the glenoid region and the remainder of the anterior exminuspansion (Fig 8) As preserved the element is nearly flat butthis may be due to postminusmortem distortion and in any casethe most strongly curved part of most sauropod scapulae isthe anterior part that is missing from this specimen The genminustle curvature preserved in the posterior part of the blade indiminuscates that the element was from the left side The bone is surminusprisingly thin in all preserved parts never exceeding a few
cm in contrast to for example the scapula of Camarasaurussupremus which is thick even in midminusblade (Osborn andMook 1921 fig 74b) This suggests that the glenoid thickenminusing and the acromial ridge may have been located some disminustance anterior to the preserved portion and a reconstructionafter the proportions of Giraffatitan brancai (Fig 8) suggeststhat about 80 of the scapularsquos full length is preserved Theposterior part of the acromion expansion is preserved howminusever and is sufficient to show that this expansion was prominusnounced so that the maximum dorsoventral height of thescapula was more than two and half times its minimumheight at the midpoint of the blade The dorsal margin slopesup towards the anterior expansion rather than forming a posminusteriorly directed ldquohookrdquo or a distinct acromion fossa betweenthe blade and the acromion process
The posterior expansion of the scapula is distinctive Insome sauropods the posterior part of the scapular blade isexpanded not at all or only slightly for example in Omeiminussaurus Young 1939 (He et al 1988 fig 41) Apatosaurus(Upchurch et al 2004b fig 4) and Rapetosaurus (Fig 9E)In others the ventral margin of the scapular blade is straightor nearly so while the dorsal margin is deflected dorsally tocreate an asymmetric expansion for example in Camaraminussaurus (Fig 9C) and Giraffatitan brancai (Fig 9D) In a fewsauropods however the ventral margin of the blade is alsodeflected ventrally to form a ldquoracquetminusshapedrdquo posterior exminuspansion This is seen in rebbachisaurids and some titanominussaurians eg some specimens of Alamosaurus Gilmore1922 (Gilmore 1946 fig 6) In Brontomerus the posteriorpart of the scapular blade is expanded in a characteristic manminusner the ventral margin is straight except for a posteroventral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 85
50 cm
ventral ldquosteprdquo
acromion expansion
dorsal ldquostepsrdquo
Fig 8 Partial left scapula of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 27761 in lateral view tentatively reconstructed after Giraffatitan brancai HMN Sa 9 (Janensch 1961 pl 15 1)
excursion two thirds of the way along the preserved portionafter which the margin continues parallel to its original traminusjectory so that the excursion appears as a gentle ldquosteprdquo Thedorsal margin is also ldquosteppedrdquo in this manner though withtwo distinct steps rather than one of which the more anterioris most strongly pronounced The net result of these featuresis that the dorsal and ventral borders of the scapula are bothstraight near the posterior extremity and that they are subminusparallel diverging by only about five degrees in the regionjust anterior to the rounded end of the posterior expansionThe step in the ventral border is not known in any othersauropod however the scapula of Neuquensaurus Powell1992 has a stepped dorsal border similar to that of Brontominusmerus (Huene 1929 via Glut 1997 275)
These characters of the scapula must be treated with somecaution however since this bone appears subject to morevariation than any other in the sauropod skeleton see for exminusample the range of shapes in scapulae of Giraffatitan brancai(Janensch 1961 pl 15 1ndash3) and in Camarasaurus supremus(Osborn and Mook 1921 figs 74ndash80)
Two small flat elements OMNH 66431 and 66432 are inminusterpreted as partial sternal plates (Fig 10) The medial edge ofeach is identifiable due to its rugose texture which formed theattachment site for cartilage joining the plates to each otherand to the sternal ribs The sternals are anteroposteriorly elonminusgate and mediolaterally narrow when complete they wereprobably at least three times as long as broad as in ldquoSaltaminussaurusrdquo robustus Huene 1929 (Huene 1929 via McIntosh1990 fig 169L) and proportionally longer than in any othersauropod including Saltasaurus loricatus (McIntosh 1990fig 169 Powell 1992 fig 30) The sternals are crescentic in
shape the anterior and posterior extremities curving laterallyaway from the midline This state was considered a titanominussaurian synapomorphy by Wilson (2002 268) but its distribuminustion is more complex in the current analysis being synapominusmorphic for Neosauropoda with losses in Flagellicaudata andCamarasaurus
86 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
Fig 10 Partial paired sternal plates of the camarasauromorph sauropodBrontomerus mcintoshi gen et sp nov from the Lower Cretaceous CedarMountain Formation of Utah OMNH 66431 and 66432 in ventral view
Fig 9 Scapulocoracoids and scapulae of sauropod dinosaurs scaled to same length of scapular blade from posterior point of glenoid to posterior margin ofblade A Mamenchisaurus youngi Young and Zhao 1972 holotype ZDM0083 left scapulocoracoid modified from Ouyang and Ye (2002 fig 22)B Diplodocus longus Hatcher 1901 USNM 10865 right scapulocoracoid reversed photograph by MPT C Camarasaurus supremus Cope 1877 AMNH5761 Sc 1 left scapula and AMNH 5761 Cor 1 left coracoid probably associated modified from Osborn and Mook (1921 figs 75 81a) D Giraffatitanbrancai (Janensch 1914) HMN Sa 9 left scapula modified from Janensch (1961 pl 15 1) E Rapetosaurus krausi Curry Rogers and Forster 2001holotype FMNH PR 2209 right scapula reversed modified from Curry Rogers (2009 fig 32) F Brontomerus mcintoshi gen et sp nov OMNH 27761left scapula tentatively reconstructed after Giraffatitan brancai
Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
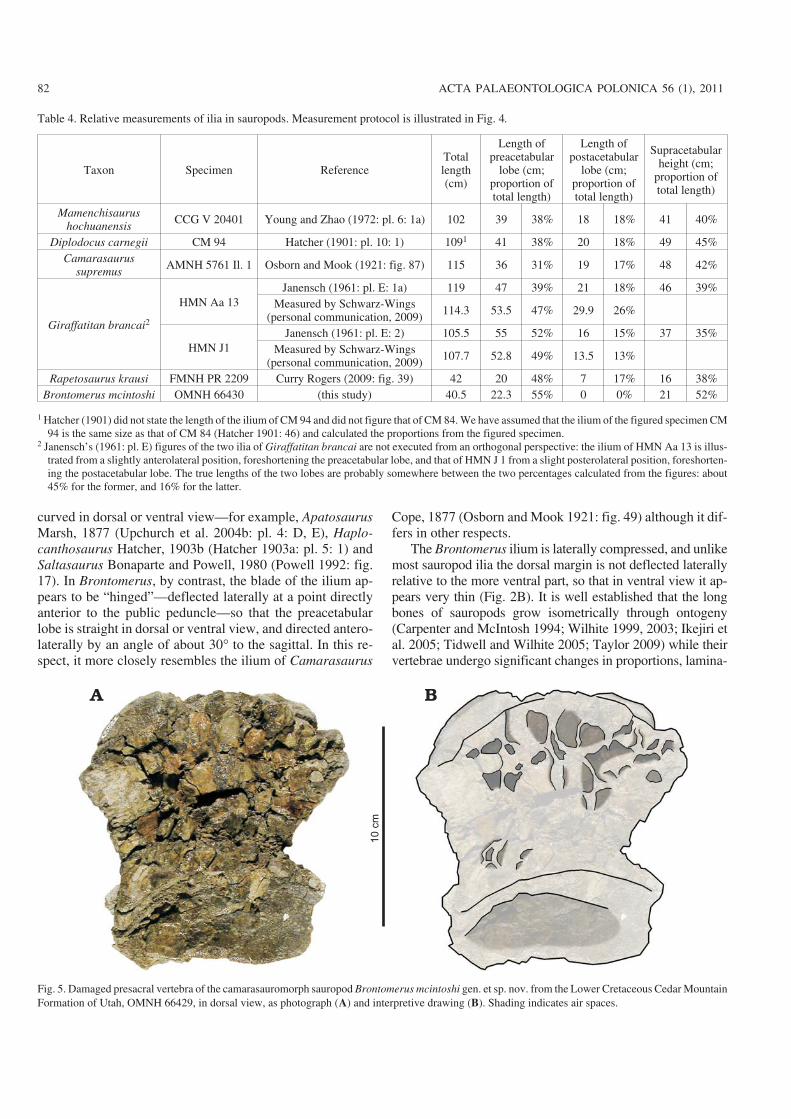
curved in dorsal or ventral viewmdashfor example ApatosaurusMarsh 1877 (Upchurch et al 2004b pl 4 D E) Haplominuscanthosaurus Hatcher 1903b (Hatcher 1903a pl 5 1) andSaltasaurus Bonaparte and Powell 1980 (Powell 1992 fig17) In Brontomerus by contrast the blade of the ilium apminuspears to be ldquohingedrdquomdashdeflected laterally at a point directlyanterior to the public pedunclemdashso that the preacetabularlobe is straight in dorsal or ventral view and directed anterominuslaterally by an angle of about 30 to the sagittal In this reminusspect it more closely resembles the ilium of Camarasaurus
Cope 1877 (Osborn and Mook 1921 fig 49) although it difminusfers in other respects
The Brontomerus ilium is laterally compressed and unlikemost sauropod ilia the dorsal margin is not deflected laterallyrelative to the more ventral part so that in ventral view it apminuspears very thin (Fig 2B) It is well established that the longbones of sauropods grow isometrically through ontogeny(Carpenter and McIntosh 1994 Wilhite 1999 2003 Ikejiri etal 2005 Tidwell and Wilhite 2005 Taylor 2009) while theirvertebrae undergo significant changes in proportions laminaminus
82 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 4 Relative measurements of ilia in sauropods Measurement protocol is illustrated in Fig 4
Taxon Specimen ReferenceTotallength(cm)
Length ofpreacetabular
lobe (cmproportion oftotal length)
Length ofpostacetabular
lobe (cmproportion oftotal length)
Supracetabularheight (cm
proportion oftotal length)
Mamenchisaurushochuanensis
CCG V 20401 Young and Zhao (1972 pl 6 1a) 102 39 38 18 18 41 40
Diplodocus carnegii CM 94 Hatcher (1901 pl 10 1) 1091 41 38 20 18 49 45Camarasaurus
supremusAMNH 5761 Il 1 Osborn and Mook (1921 fig 87) 115 36 31 19 17 48 42
Giraffatitan brancai2
HMN Aa 13Janensch (1961 pl E 1a) 119 47 39 21 18 46 39
Measured by SchwarzminusWings(personal communication 2009) 1143 535 47 299 26
HMN J1Janensch (1961 pl E 2) 1055 55 52 16 15 37 35
Measured by SchwarzminusWings(personal communication 2009) 1077 528 49 135 13
Rapetosaurus krausi FMNH PR 2209 Curry Rogers (2009 fig 39) 42 20 48 7 17 16 38Brontomerus mcintoshi OMNH 66430 (this study) 405 223 55 0 0 21 52
1 Hatcher (1901) did not state the length of the ilium of CM 94 and did not figure that of CM 84 We have assumed that the ilium of the figured specimen CM94 is the same size as that of CM 84 (Hatcher 1901 46) and calculated the proportions from the figured specimen
2 Janenschrsquos (1961 pl E) figures of the two ilia of Giraffatitan brancai are not executed from an orthogonal perspective the ilium of HMN Aa 13 is illusminustrated from a slightly anterolateral position foreshortening the preacetabular lobe and that of HMN J 1 from a slight posterolateral position foreshortenminusing the postacetabular lobe The true lengths of the two lobes are probably somewhere between the two percentages calculated from the figures about45 for the former and 16 for the latter
10
cm
Fig 5 Damaged presacral vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah OMNH 66429 in dorsal view as photograph (A) and interpretive drawing (B) Shading indicates air spaces
tion pneumatic excavations and neurocentral fusion (Wedel2003a 248 b 352ndash354) Ontogenetic changes in limbminusgirdleelements such as the ilium are less well understood due to apaucity of sufficiently well preserved specimens (Ray Wilhitepersonal communication October 2007) Therefore the lateralcompression of the Brontomerus ilium may be a juvenile charminusacter with the ilium thickening through ontogeny to supportthe growing weight of the animal or it may in fact be phylominusgenetically significant
Presacral centrummdashA single presacral centrum OMNH66429 was recovered (Fig 5) Unfortunately preservation isvery poor the neural arch and all processes have been lostand the centrum has been greatly crushed dorsoventrally sothat the remaining part is essentially flat the element is 14
cm in both anteroposterior length and transverse width butno more than 3 cm in dorsoventral depth The small size indiminuscates that this element belonged to a juvenile The internalstructure of the centrum is visible however and consists offine septa dividing a hollow internal space irregularly intomany small camellae This morphology is characteristic oftitanosauriforms (Wedel 2003b 354ndash355) Highly camellateinternal structure has not previously been observed in juveminusnile sauropod vertebrae but this may be due to samplingbias so far all juvenile sauropod vertebrae that have beenstudied for internal structure have been those of Camaraminussaurus and diplodocoids which follow an ontogenetic trajecminustory in which large shallow lateral fossae develop into cameminusrae from which smaller accessory camerae and camellae deminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 83
prezygapophysis
neural spine
postzygapophysealramus
postzygapophysealfacet
chevron facets
5 cm
Fig 6 Midminuscaudal vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 61248 in dorsal (A) anterior (B) left lateral (C) posterior (D) and ventral (E) views
velop (Wedel et al 2000a fig 11 Wedel 2003b 349) TheHotel Mesa presacral suggests that camellate vertebrae mayhave developed differently in titanosauriforms possibly byinminussitu formation of camellae during pneumatization as ocminuscurs in the camellate vertebrae of birds (personal observaminustion MJW)
Caudal vertebraemdashOMNH 61248 is a distinctive caudalvertebra with elongated preminus and postzygapophyseal rami(Fig 6) Apart from the tip of the right prezygapophysis theelement is complete and well preserved While the centrum isonly 11 cm in length the distance from the prezygapophysisto postzygapophysis is 145 cm The centrum is slightlybroader than tall (6 cm compared with 55 cm anteriorly 65cm compared with 5 cm posteriorly) and gently waisted Theneural arch is set forward on the centrum but does not reachthe anterior margin The neural spine is so reduced and sostrongly inclined posteriorly as to be all but indistinguishminusable and is apparent only as a very low eminence abovethe postzygapophyses The postzygapophyseal facets themminusselves are set on the posterolateral faces of a low process thathangs below the main postzygapophyseal ramus Chevronfacets are weakly present on the posterior margin of the venminustral surface of the centrum but not on the anterior marginThe elongation index of 22 indicates a midminustominusposterior pominussition in the caudal sequence for this element as similarcentrum proportions do not appear until about caudal 30 inGiraffatitan brancai (Janensch 1950 pl 3)
This vertebra most closely resembles the indeterminatesauropod vertebra BMNH 27500 from the Wessex Formationof the Isle of Wight figured by Naish and Martill (2001 pl33) The Barremian age of that specimen places it about 15 Ma
earlier than OMNH 61248 Its neural arch is less elevated thanthat of the Hotel Mesa specimen its postzygapophyses projectyet farther posteriorly and its prezygapophyses less far anteriminusorly and it is very mildly biconvex rather than procoelous butin other respects including absolute size it is a good match forthe Brontomerus caudal
Also included in the Hotel Mesa material is OMNH27794 a partial distal caudal centrum figured by Wedel(2005 fig 77) This centrum is approximately round incrossminussection about 4 cm in diameter and internally consistsof apneumatic cancellous bone
RibsmdashThe Hotel Mesa material contains several dorsal ribsin various states of preservation but no readily identifiablecervical ribs The dorsal rib elements include the shaft of alarge flat rib (OMNH 27762) portions of several smaller ribshafts (OMNH 27763ndash27765 27768 and others) a flattenedrib head (OMNH 27767) and most informatively a smallcomplete rib (OMNH 27766 Fig 7) Despite its excellentpreservation and apparent lack of distortion this element isdifficult to interpret Its shaft is straight for almost its wholelength and both articular facets are directed dorsally ratherthan being inclined medially The tuberculum is directly inline with the main part of the shaft of the rib and thecapitulum is at an angle of about 30 to it In these respectsthe rib resembles the most anterior dorsal rib of the Diplominusdocus carnegii holotype CM 84 (personal observation MPTthis element was not figured by Hatcher [1901]) We thereminusfore interpret this rib as having probably belonged to the rightside of the first dorsal vertebra
The rib is unusual in other respects however most notablythat the ventral part of the shaft curves laterally rather than meminus
84 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
10 cm
capitulum
pneumatic fossa
tuberculum
Fig 7 First right dorsal rib of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah OMNH 27766 in posterior view head of rib showing pneumatic invasion of shaft (A) and complete rib showing laterally directed curvature of shaft (B)
dially Careful inspection of the bone reveals no indication ofdistortion or of incorrect reconstruction It may be possiblethat in life the thorax was transversely compressed so that thedorsal part of the rib shaft was directed ventromedially and themore ventral part was vertical Both articular facets are subminuscircular in dorsal view and significantly expanded comparedwith the rami that bear them On the anterior face of the head alow ridge arises just below the capitulum and extends downthe medial edge of the rib for about 40 of its length
The head of the rib is also unusual in that a thin sheet ofbone connects the rami that support the articular facets andthis sheet extends much farther proximally than in mostsauropod ribs Its precise extent cannot be ascertained due tobreakage The sheet of bone is perforated close to the capituminuslar ramus and from this perforation a pneumatic cavity inminusvades the shaft of the rib extending ventrally from within ashallow fossa in the posterior face Pneumatization of thedorsal ribs is a synapomorphy of Titanosauriformes (Wilsonand Sereno 1998) although pneumatic dorsal ribs are also inminusfrequently present in diplodocids (Gilmore 1936 Lovelaceet al 2003 2008)
Pectoral girdlemdashOMNH 27761 is a partial scapula consistminusing of the blade and part of the anterior expansion but missminusing the glenoid region and the remainder of the anterior exminuspansion (Fig 8) As preserved the element is nearly flat butthis may be due to postminusmortem distortion and in any casethe most strongly curved part of most sauropod scapulae isthe anterior part that is missing from this specimen The genminustle curvature preserved in the posterior part of the blade indiminuscates that the element was from the left side The bone is surminusprisingly thin in all preserved parts never exceeding a few
cm in contrast to for example the scapula of Camarasaurussupremus which is thick even in midminusblade (Osborn andMook 1921 fig 74b) This suggests that the glenoid thickenminusing and the acromial ridge may have been located some disminustance anterior to the preserved portion and a reconstructionafter the proportions of Giraffatitan brancai (Fig 8) suggeststhat about 80 of the scapularsquos full length is preserved Theposterior part of the acromion expansion is preserved howminusever and is sufficient to show that this expansion was prominusnounced so that the maximum dorsoventral height of thescapula was more than two and half times its minimumheight at the midpoint of the blade The dorsal margin slopesup towards the anterior expansion rather than forming a posminusteriorly directed ldquohookrdquo or a distinct acromion fossa betweenthe blade and the acromion process
The posterior expansion of the scapula is distinctive Insome sauropods the posterior part of the scapular blade isexpanded not at all or only slightly for example in Omeiminussaurus Young 1939 (He et al 1988 fig 41) Apatosaurus(Upchurch et al 2004b fig 4) and Rapetosaurus (Fig 9E)In others the ventral margin of the scapular blade is straightor nearly so while the dorsal margin is deflected dorsally tocreate an asymmetric expansion for example in Camaraminussaurus (Fig 9C) and Giraffatitan brancai (Fig 9D) In a fewsauropods however the ventral margin of the blade is alsodeflected ventrally to form a ldquoracquetminusshapedrdquo posterior exminuspansion This is seen in rebbachisaurids and some titanominussaurians eg some specimens of Alamosaurus Gilmore1922 (Gilmore 1946 fig 6) In Brontomerus the posteriorpart of the scapular blade is expanded in a characteristic manminusner the ventral margin is straight except for a posteroventral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 85
50 cm
ventral ldquosteprdquo
acromion expansion
dorsal ldquostepsrdquo
Fig 8 Partial left scapula of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 27761 in lateral view tentatively reconstructed after Giraffatitan brancai HMN Sa 9 (Janensch 1961 pl 15 1)
excursion two thirds of the way along the preserved portionafter which the margin continues parallel to its original traminusjectory so that the excursion appears as a gentle ldquosteprdquo Thedorsal margin is also ldquosteppedrdquo in this manner though withtwo distinct steps rather than one of which the more anterioris most strongly pronounced The net result of these featuresis that the dorsal and ventral borders of the scapula are bothstraight near the posterior extremity and that they are subminusparallel diverging by only about five degrees in the regionjust anterior to the rounded end of the posterior expansionThe step in the ventral border is not known in any othersauropod however the scapula of Neuquensaurus Powell1992 has a stepped dorsal border similar to that of Brontominusmerus (Huene 1929 via Glut 1997 275)
These characters of the scapula must be treated with somecaution however since this bone appears subject to morevariation than any other in the sauropod skeleton see for exminusample the range of shapes in scapulae of Giraffatitan brancai(Janensch 1961 pl 15 1ndash3) and in Camarasaurus supremus(Osborn and Mook 1921 figs 74ndash80)
Two small flat elements OMNH 66431 and 66432 are inminusterpreted as partial sternal plates (Fig 10) The medial edge ofeach is identifiable due to its rugose texture which formed theattachment site for cartilage joining the plates to each otherand to the sternal ribs The sternals are anteroposteriorly elonminusgate and mediolaterally narrow when complete they wereprobably at least three times as long as broad as in ldquoSaltaminussaurusrdquo robustus Huene 1929 (Huene 1929 via McIntosh1990 fig 169L) and proportionally longer than in any othersauropod including Saltasaurus loricatus (McIntosh 1990fig 169 Powell 1992 fig 30) The sternals are crescentic in
shape the anterior and posterior extremities curving laterallyaway from the midline This state was considered a titanominussaurian synapomorphy by Wilson (2002 268) but its distribuminustion is more complex in the current analysis being synapominusmorphic for Neosauropoda with losses in Flagellicaudata andCamarasaurus
86 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
Fig 10 Partial paired sternal plates of the camarasauromorph sauropodBrontomerus mcintoshi gen et sp nov from the Lower Cretaceous CedarMountain Formation of Utah OMNH 66431 and 66432 in ventral view
Fig 9 Scapulocoracoids and scapulae of sauropod dinosaurs scaled to same length of scapular blade from posterior point of glenoid to posterior margin ofblade A Mamenchisaurus youngi Young and Zhao 1972 holotype ZDM0083 left scapulocoracoid modified from Ouyang and Ye (2002 fig 22)B Diplodocus longus Hatcher 1901 USNM 10865 right scapulocoracoid reversed photograph by MPT C Camarasaurus supremus Cope 1877 AMNH5761 Sc 1 left scapula and AMNH 5761 Cor 1 left coracoid probably associated modified from Osborn and Mook (1921 figs 75 81a) D Giraffatitanbrancai (Janensch 1914) HMN Sa 9 left scapula modified from Janensch (1961 pl 15 1) E Rapetosaurus krausi Curry Rogers and Forster 2001holotype FMNH PR 2209 right scapula reversed modified from Curry Rogers (2009 fig 32) F Brontomerus mcintoshi gen et sp nov OMNH 27761left scapula tentatively reconstructed after Giraffatitan brancai
Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

tion pneumatic excavations and neurocentral fusion (Wedel2003a 248 b 352ndash354) Ontogenetic changes in limbminusgirdleelements such as the ilium are less well understood due to apaucity of sufficiently well preserved specimens (Ray Wilhitepersonal communication October 2007) Therefore the lateralcompression of the Brontomerus ilium may be a juvenile charminusacter with the ilium thickening through ontogeny to supportthe growing weight of the animal or it may in fact be phylominusgenetically significant
Presacral centrummdashA single presacral centrum OMNH66429 was recovered (Fig 5) Unfortunately preservation isvery poor the neural arch and all processes have been lostand the centrum has been greatly crushed dorsoventrally sothat the remaining part is essentially flat the element is 14
cm in both anteroposterior length and transverse width butno more than 3 cm in dorsoventral depth The small size indiminuscates that this element belonged to a juvenile The internalstructure of the centrum is visible however and consists offine septa dividing a hollow internal space irregularly intomany small camellae This morphology is characteristic oftitanosauriforms (Wedel 2003b 354ndash355) Highly camellateinternal structure has not previously been observed in juveminusnile sauropod vertebrae but this may be due to samplingbias so far all juvenile sauropod vertebrae that have beenstudied for internal structure have been those of Camaraminussaurus and diplodocoids which follow an ontogenetic trajecminustory in which large shallow lateral fossae develop into cameminusrae from which smaller accessory camerae and camellae deminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 83
prezygapophysis
neural spine
postzygapophysealramus
postzygapophysealfacet
chevron facets
5 cm
Fig 6 Midminuscaudal vertebra of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 61248 in dorsal (A) anterior (B) left lateral (C) posterior (D) and ventral (E) views
velop (Wedel et al 2000a fig 11 Wedel 2003b 349) TheHotel Mesa presacral suggests that camellate vertebrae mayhave developed differently in titanosauriforms possibly byinminussitu formation of camellae during pneumatization as ocminuscurs in the camellate vertebrae of birds (personal observaminustion MJW)
Caudal vertebraemdashOMNH 61248 is a distinctive caudalvertebra with elongated preminus and postzygapophyseal rami(Fig 6) Apart from the tip of the right prezygapophysis theelement is complete and well preserved While the centrum isonly 11 cm in length the distance from the prezygapophysisto postzygapophysis is 145 cm The centrum is slightlybroader than tall (6 cm compared with 55 cm anteriorly 65cm compared with 5 cm posteriorly) and gently waisted Theneural arch is set forward on the centrum but does not reachthe anterior margin The neural spine is so reduced and sostrongly inclined posteriorly as to be all but indistinguishminusable and is apparent only as a very low eminence abovethe postzygapophyses The postzygapophyseal facets themminusselves are set on the posterolateral faces of a low process thathangs below the main postzygapophyseal ramus Chevronfacets are weakly present on the posterior margin of the venminustral surface of the centrum but not on the anterior marginThe elongation index of 22 indicates a midminustominusposterior pominussition in the caudal sequence for this element as similarcentrum proportions do not appear until about caudal 30 inGiraffatitan brancai (Janensch 1950 pl 3)
This vertebra most closely resembles the indeterminatesauropod vertebra BMNH 27500 from the Wessex Formationof the Isle of Wight figured by Naish and Martill (2001 pl33) The Barremian age of that specimen places it about 15 Ma
earlier than OMNH 61248 Its neural arch is less elevated thanthat of the Hotel Mesa specimen its postzygapophyses projectyet farther posteriorly and its prezygapophyses less far anteriminusorly and it is very mildly biconvex rather than procoelous butin other respects including absolute size it is a good match forthe Brontomerus caudal
Also included in the Hotel Mesa material is OMNH27794 a partial distal caudal centrum figured by Wedel(2005 fig 77) This centrum is approximately round incrossminussection about 4 cm in diameter and internally consistsof apneumatic cancellous bone
RibsmdashThe Hotel Mesa material contains several dorsal ribsin various states of preservation but no readily identifiablecervical ribs The dorsal rib elements include the shaft of alarge flat rib (OMNH 27762) portions of several smaller ribshafts (OMNH 27763ndash27765 27768 and others) a flattenedrib head (OMNH 27767) and most informatively a smallcomplete rib (OMNH 27766 Fig 7) Despite its excellentpreservation and apparent lack of distortion this element isdifficult to interpret Its shaft is straight for almost its wholelength and both articular facets are directed dorsally ratherthan being inclined medially The tuberculum is directly inline with the main part of the shaft of the rib and thecapitulum is at an angle of about 30 to it In these respectsthe rib resembles the most anterior dorsal rib of the Diplominusdocus carnegii holotype CM 84 (personal observation MPTthis element was not figured by Hatcher [1901]) We thereminusfore interpret this rib as having probably belonged to the rightside of the first dorsal vertebra
The rib is unusual in other respects however most notablythat the ventral part of the shaft curves laterally rather than meminus
84 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
10 cm
capitulum
pneumatic fossa
tuberculum
Fig 7 First right dorsal rib of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah OMNH 27766 in posterior view head of rib showing pneumatic invasion of shaft (A) and complete rib showing laterally directed curvature of shaft (B)
dially Careful inspection of the bone reveals no indication ofdistortion or of incorrect reconstruction It may be possiblethat in life the thorax was transversely compressed so that thedorsal part of the rib shaft was directed ventromedially and themore ventral part was vertical Both articular facets are subminuscircular in dorsal view and significantly expanded comparedwith the rami that bear them On the anterior face of the head alow ridge arises just below the capitulum and extends downthe medial edge of the rib for about 40 of its length
The head of the rib is also unusual in that a thin sheet ofbone connects the rami that support the articular facets andthis sheet extends much farther proximally than in mostsauropod ribs Its precise extent cannot be ascertained due tobreakage The sheet of bone is perforated close to the capituminuslar ramus and from this perforation a pneumatic cavity inminusvades the shaft of the rib extending ventrally from within ashallow fossa in the posterior face Pneumatization of thedorsal ribs is a synapomorphy of Titanosauriformes (Wilsonand Sereno 1998) although pneumatic dorsal ribs are also inminusfrequently present in diplodocids (Gilmore 1936 Lovelaceet al 2003 2008)
Pectoral girdlemdashOMNH 27761 is a partial scapula consistminusing of the blade and part of the anterior expansion but missminusing the glenoid region and the remainder of the anterior exminuspansion (Fig 8) As preserved the element is nearly flat butthis may be due to postminusmortem distortion and in any casethe most strongly curved part of most sauropod scapulae isthe anterior part that is missing from this specimen The genminustle curvature preserved in the posterior part of the blade indiminuscates that the element was from the left side The bone is surminusprisingly thin in all preserved parts never exceeding a few
cm in contrast to for example the scapula of Camarasaurussupremus which is thick even in midminusblade (Osborn andMook 1921 fig 74b) This suggests that the glenoid thickenminusing and the acromial ridge may have been located some disminustance anterior to the preserved portion and a reconstructionafter the proportions of Giraffatitan brancai (Fig 8) suggeststhat about 80 of the scapularsquos full length is preserved Theposterior part of the acromion expansion is preserved howminusever and is sufficient to show that this expansion was prominusnounced so that the maximum dorsoventral height of thescapula was more than two and half times its minimumheight at the midpoint of the blade The dorsal margin slopesup towards the anterior expansion rather than forming a posminusteriorly directed ldquohookrdquo or a distinct acromion fossa betweenthe blade and the acromion process
The posterior expansion of the scapula is distinctive Insome sauropods the posterior part of the scapular blade isexpanded not at all or only slightly for example in Omeiminussaurus Young 1939 (He et al 1988 fig 41) Apatosaurus(Upchurch et al 2004b fig 4) and Rapetosaurus (Fig 9E)In others the ventral margin of the scapular blade is straightor nearly so while the dorsal margin is deflected dorsally tocreate an asymmetric expansion for example in Camaraminussaurus (Fig 9C) and Giraffatitan brancai (Fig 9D) In a fewsauropods however the ventral margin of the blade is alsodeflected ventrally to form a ldquoracquetminusshapedrdquo posterior exminuspansion This is seen in rebbachisaurids and some titanominussaurians eg some specimens of Alamosaurus Gilmore1922 (Gilmore 1946 fig 6) In Brontomerus the posteriorpart of the scapular blade is expanded in a characteristic manminusner the ventral margin is straight except for a posteroventral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 85
50 cm
ventral ldquosteprdquo
acromion expansion
dorsal ldquostepsrdquo
Fig 8 Partial left scapula of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 27761 in lateral view tentatively reconstructed after Giraffatitan brancai HMN Sa 9 (Janensch 1961 pl 15 1)
excursion two thirds of the way along the preserved portionafter which the margin continues parallel to its original traminusjectory so that the excursion appears as a gentle ldquosteprdquo Thedorsal margin is also ldquosteppedrdquo in this manner though withtwo distinct steps rather than one of which the more anterioris most strongly pronounced The net result of these featuresis that the dorsal and ventral borders of the scapula are bothstraight near the posterior extremity and that they are subminusparallel diverging by only about five degrees in the regionjust anterior to the rounded end of the posterior expansionThe step in the ventral border is not known in any othersauropod however the scapula of Neuquensaurus Powell1992 has a stepped dorsal border similar to that of Brontominusmerus (Huene 1929 via Glut 1997 275)
These characters of the scapula must be treated with somecaution however since this bone appears subject to morevariation than any other in the sauropod skeleton see for exminusample the range of shapes in scapulae of Giraffatitan brancai(Janensch 1961 pl 15 1ndash3) and in Camarasaurus supremus(Osborn and Mook 1921 figs 74ndash80)
Two small flat elements OMNH 66431 and 66432 are inminusterpreted as partial sternal plates (Fig 10) The medial edge ofeach is identifiable due to its rugose texture which formed theattachment site for cartilage joining the plates to each otherand to the sternal ribs The sternals are anteroposteriorly elonminusgate and mediolaterally narrow when complete they wereprobably at least three times as long as broad as in ldquoSaltaminussaurusrdquo robustus Huene 1929 (Huene 1929 via McIntosh1990 fig 169L) and proportionally longer than in any othersauropod including Saltasaurus loricatus (McIntosh 1990fig 169 Powell 1992 fig 30) The sternals are crescentic in
shape the anterior and posterior extremities curving laterallyaway from the midline This state was considered a titanominussaurian synapomorphy by Wilson (2002 268) but its distribuminustion is more complex in the current analysis being synapominusmorphic for Neosauropoda with losses in Flagellicaudata andCamarasaurus
86 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
Fig 10 Partial paired sternal plates of the camarasauromorph sauropodBrontomerus mcintoshi gen et sp nov from the Lower Cretaceous CedarMountain Formation of Utah OMNH 66431 and 66432 in ventral view
Fig 9 Scapulocoracoids and scapulae of sauropod dinosaurs scaled to same length of scapular blade from posterior point of glenoid to posterior margin ofblade A Mamenchisaurus youngi Young and Zhao 1972 holotype ZDM0083 left scapulocoracoid modified from Ouyang and Ye (2002 fig 22)B Diplodocus longus Hatcher 1901 USNM 10865 right scapulocoracoid reversed photograph by MPT C Camarasaurus supremus Cope 1877 AMNH5761 Sc 1 left scapula and AMNH 5761 Cor 1 left coracoid probably associated modified from Osborn and Mook (1921 figs 75 81a) D Giraffatitanbrancai (Janensch 1914) HMN Sa 9 left scapula modified from Janensch (1961 pl 15 1) E Rapetosaurus krausi Curry Rogers and Forster 2001holotype FMNH PR 2209 right scapula reversed modified from Curry Rogers (2009 fig 32) F Brontomerus mcintoshi gen et sp nov OMNH 27761left scapula tentatively reconstructed after Giraffatitan brancai
Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

velop (Wedel et al 2000a fig 11 Wedel 2003b 349) TheHotel Mesa presacral suggests that camellate vertebrae mayhave developed differently in titanosauriforms possibly byinminussitu formation of camellae during pneumatization as ocminuscurs in the camellate vertebrae of birds (personal observaminustion MJW)
Caudal vertebraemdashOMNH 61248 is a distinctive caudalvertebra with elongated preminus and postzygapophyseal rami(Fig 6) Apart from the tip of the right prezygapophysis theelement is complete and well preserved While the centrum isonly 11 cm in length the distance from the prezygapophysisto postzygapophysis is 145 cm The centrum is slightlybroader than tall (6 cm compared with 55 cm anteriorly 65cm compared with 5 cm posteriorly) and gently waisted Theneural arch is set forward on the centrum but does not reachthe anterior margin The neural spine is so reduced and sostrongly inclined posteriorly as to be all but indistinguishminusable and is apparent only as a very low eminence abovethe postzygapophyses The postzygapophyseal facets themminusselves are set on the posterolateral faces of a low process thathangs below the main postzygapophyseal ramus Chevronfacets are weakly present on the posterior margin of the venminustral surface of the centrum but not on the anterior marginThe elongation index of 22 indicates a midminustominusposterior pominussition in the caudal sequence for this element as similarcentrum proportions do not appear until about caudal 30 inGiraffatitan brancai (Janensch 1950 pl 3)
This vertebra most closely resembles the indeterminatesauropod vertebra BMNH 27500 from the Wessex Formationof the Isle of Wight figured by Naish and Martill (2001 pl33) The Barremian age of that specimen places it about 15 Ma
earlier than OMNH 61248 Its neural arch is less elevated thanthat of the Hotel Mesa specimen its postzygapophyses projectyet farther posteriorly and its prezygapophyses less far anteriminusorly and it is very mildly biconvex rather than procoelous butin other respects including absolute size it is a good match forthe Brontomerus caudal
Also included in the Hotel Mesa material is OMNH27794 a partial distal caudal centrum figured by Wedel(2005 fig 77) This centrum is approximately round incrossminussection about 4 cm in diameter and internally consistsof apneumatic cancellous bone
RibsmdashThe Hotel Mesa material contains several dorsal ribsin various states of preservation but no readily identifiablecervical ribs The dorsal rib elements include the shaft of alarge flat rib (OMNH 27762) portions of several smaller ribshafts (OMNH 27763ndash27765 27768 and others) a flattenedrib head (OMNH 27767) and most informatively a smallcomplete rib (OMNH 27766 Fig 7) Despite its excellentpreservation and apparent lack of distortion this element isdifficult to interpret Its shaft is straight for almost its wholelength and both articular facets are directed dorsally ratherthan being inclined medially The tuberculum is directly inline with the main part of the shaft of the rib and thecapitulum is at an angle of about 30 to it In these respectsthe rib resembles the most anterior dorsal rib of the Diplominusdocus carnegii holotype CM 84 (personal observation MPTthis element was not figured by Hatcher [1901]) We thereminusfore interpret this rib as having probably belonged to the rightside of the first dorsal vertebra
The rib is unusual in other respects however most notablythat the ventral part of the shaft curves laterally rather than meminus
84 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
10 cm
capitulum
pneumatic fossa
tuberculum
Fig 7 First right dorsal rib of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formation ofUtah OMNH 27766 in posterior view head of rib showing pneumatic invasion of shaft (A) and complete rib showing laterally directed curvature of shaft (B)
dially Careful inspection of the bone reveals no indication ofdistortion or of incorrect reconstruction It may be possiblethat in life the thorax was transversely compressed so that thedorsal part of the rib shaft was directed ventromedially and themore ventral part was vertical Both articular facets are subminuscircular in dorsal view and significantly expanded comparedwith the rami that bear them On the anterior face of the head alow ridge arises just below the capitulum and extends downthe medial edge of the rib for about 40 of its length
The head of the rib is also unusual in that a thin sheet ofbone connects the rami that support the articular facets andthis sheet extends much farther proximally than in mostsauropod ribs Its precise extent cannot be ascertained due tobreakage The sheet of bone is perforated close to the capituminuslar ramus and from this perforation a pneumatic cavity inminusvades the shaft of the rib extending ventrally from within ashallow fossa in the posterior face Pneumatization of thedorsal ribs is a synapomorphy of Titanosauriformes (Wilsonand Sereno 1998) although pneumatic dorsal ribs are also inminusfrequently present in diplodocids (Gilmore 1936 Lovelaceet al 2003 2008)
Pectoral girdlemdashOMNH 27761 is a partial scapula consistminusing of the blade and part of the anterior expansion but missminusing the glenoid region and the remainder of the anterior exminuspansion (Fig 8) As preserved the element is nearly flat butthis may be due to postminusmortem distortion and in any casethe most strongly curved part of most sauropod scapulae isthe anterior part that is missing from this specimen The genminustle curvature preserved in the posterior part of the blade indiminuscates that the element was from the left side The bone is surminusprisingly thin in all preserved parts never exceeding a few
cm in contrast to for example the scapula of Camarasaurussupremus which is thick even in midminusblade (Osborn andMook 1921 fig 74b) This suggests that the glenoid thickenminusing and the acromial ridge may have been located some disminustance anterior to the preserved portion and a reconstructionafter the proportions of Giraffatitan brancai (Fig 8) suggeststhat about 80 of the scapularsquos full length is preserved Theposterior part of the acromion expansion is preserved howminusever and is sufficient to show that this expansion was prominusnounced so that the maximum dorsoventral height of thescapula was more than two and half times its minimumheight at the midpoint of the blade The dorsal margin slopesup towards the anterior expansion rather than forming a posminusteriorly directed ldquohookrdquo or a distinct acromion fossa betweenthe blade and the acromion process
The posterior expansion of the scapula is distinctive Insome sauropods the posterior part of the scapular blade isexpanded not at all or only slightly for example in Omeiminussaurus Young 1939 (He et al 1988 fig 41) Apatosaurus(Upchurch et al 2004b fig 4) and Rapetosaurus (Fig 9E)In others the ventral margin of the scapular blade is straightor nearly so while the dorsal margin is deflected dorsally tocreate an asymmetric expansion for example in Camaraminussaurus (Fig 9C) and Giraffatitan brancai (Fig 9D) In a fewsauropods however the ventral margin of the blade is alsodeflected ventrally to form a ldquoracquetminusshapedrdquo posterior exminuspansion This is seen in rebbachisaurids and some titanominussaurians eg some specimens of Alamosaurus Gilmore1922 (Gilmore 1946 fig 6) In Brontomerus the posteriorpart of the scapular blade is expanded in a characteristic manminusner the ventral margin is straight except for a posteroventral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 85
50 cm
ventral ldquosteprdquo
acromion expansion
dorsal ldquostepsrdquo
Fig 8 Partial left scapula of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 27761 in lateral view tentatively reconstructed after Giraffatitan brancai HMN Sa 9 (Janensch 1961 pl 15 1)
excursion two thirds of the way along the preserved portionafter which the margin continues parallel to its original traminusjectory so that the excursion appears as a gentle ldquosteprdquo Thedorsal margin is also ldquosteppedrdquo in this manner though withtwo distinct steps rather than one of which the more anterioris most strongly pronounced The net result of these featuresis that the dorsal and ventral borders of the scapula are bothstraight near the posterior extremity and that they are subminusparallel diverging by only about five degrees in the regionjust anterior to the rounded end of the posterior expansionThe step in the ventral border is not known in any othersauropod however the scapula of Neuquensaurus Powell1992 has a stepped dorsal border similar to that of Brontominusmerus (Huene 1929 via Glut 1997 275)
These characters of the scapula must be treated with somecaution however since this bone appears subject to morevariation than any other in the sauropod skeleton see for exminusample the range of shapes in scapulae of Giraffatitan brancai(Janensch 1961 pl 15 1ndash3) and in Camarasaurus supremus(Osborn and Mook 1921 figs 74ndash80)
Two small flat elements OMNH 66431 and 66432 are inminusterpreted as partial sternal plates (Fig 10) The medial edge ofeach is identifiable due to its rugose texture which formed theattachment site for cartilage joining the plates to each otherand to the sternal ribs The sternals are anteroposteriorly elonminusgate and mediolaterally narrow when complete they wereprobably at least three times as long as broad as in ldquoSaltaminussaurusrdquo robustus Huene 1929 (Huene 1929 via McIntosh1990 fig 169L) and proportionally longer than in any othersauropod including Saltasaurus loricatus (McIntosh 1990fig 169 Powell 1992 fig 30) The sternals are crescentic in
shape the anterior and posterior extremities curving laterallyaway from the midline This state was considered a titanominussaurian synapomorphy by Wilson (2002 268) but its distribuminustion is more complex in the current analysis being synapominusmorphic for Neosauropoda with losses in Flagellicaudata andCamarasaurus
86 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
Fig 10 Partial paired sternal plates of the camarasauromorph sauropodBrontomerus mcintoshi gen et sp nov from the Lower Cretaceous CedarMountain Formation of Utah OMNH 66431 and 66432 in ventral view
Fig 9 Scapulocoracoids and scapulae of sauropod dinosaurs scaled to same length of scapular blade from posterior point of glenoid to posterior margin ofblade A Mamenchisaurus youngi Young and Zhao 1972 holotype ZDM0083 left scapulocoracoid modified from Ouyang and Ye (2002 fig 22)B Diplodocus longus Hatcher 1901 USNM 10865 right scapulocoracoid reversed photograph by MPT C Camarasaurus supremus Cope 1877 AMNH5761 Sc 1 left scapula and AMNH 5761 Cor 1 left coracoid probably associated modified from Osborn and Mook (1921 figs 75 81a) D Giraffatitanbrancai (Janensch 1914) HMN Sa 9 left scapula modified from Janensch (1961 pl 15 1) E Rapetosaurus krausi Curry Rogers and Forster 2001holotype FMNH PR 2209 right scapula reversed modified from Curry Rogers (2009 fig 32) F Brontomerus mcintoshi gen et sp nov OMNH 27761left scapula tentatively reconstructed after Giraffatitan brancai
Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

dially Careful inspection of the bone reveals no indication ofdistortion or of incorrect reconstruction It may be possiblethat in life the thorax was transversely compressed so that thedorsal part of the rib shaft was directed ventromedially and themore ventral part was vertical Both articular facets are subminuscircular in dorsal view and significantly expanded comparedwith the rami that bear them On the anterior face of the head alow ridge arises just below the capitulum and extends downthe medial edge of the rib for about 40 of its length
The head of the rib is also unusual in that a thin sheet ofbone connects the rami that support the articular facets andthis sheet extends much farther proximally than in mostsauropod ribs Its precise extent cannot be ascertained due tobreakage The sheet of bone is perforated close to the capituminuslar ramus and from this perforation a pneumatic cavity inminusvades the shaft of the rib extending ventrally from within ashallow fossa in the posterior face Pneumatization of thedorsal ribs is a synapomorphy of Titanosauriformes (Wilsonand Sereno 1998) although pneumatic dorsal ribs are also inminusfrequently present in diplodocids (Gilmore 1936 Lovelaceet al 2003 2008)
Pectoral girdlemdashOMNH 27761 is a partial scapula consistminusing of the blade and part of the anterior expansion but missminusing the glenoid region and the remainder of the anterior exminuspansion (Fig 8) As preserved the element is nearly flat butthis may be due to postminusmortem distortion and in any casethe most strongly curved part of most sauropod scapulae isthe anterior part that is missing from this specimen The genminustle curvature preserved in the posterior part of the blade indiminuscates that the element was from the left side The bone is surminusprisingly thin in all preserved parts never exceeding a few
cm in contrast to for example the scapula of Camarasaurussupremus which is thick even in midminusblade (Osborn andMook 1921 fig 74b) This suggests that the glenoid thickenminusing and the acromial ridge may have been located some disminustance anterior to the preserved portion and a reconstructionafter the proportions of Giraffatitan brancai (Fig 8) suggeststhat about 80 of the scapularsquos full length is preserved Theposterior part of the acromion expansion is preserved howminusever and is sufficient to show that this expansion was prominusnounced so that the maximum dorsoventral height of thescapula was more than two and half times its minimumheight at the midpoint of the blade The dorsal margin slopesup towards the anterior expansion rather than forming a posminusteriorly directed ldquohookrdquo or a distinct acromion fossa betweenthe blade and the acromion process
The posterior expansion of the scapula is distinctive Insome sauropods the posterior part of the scapular blade isexpanded not at all or only slightly for example in Omeiminussaurus Young 1939 (He et al 1988 fig 41) Apatosaurus(Upchurch et al 2004b fig 4) and Rapetosaurus (Fig 9E)In others the ventral margin of the scapular blade is straightor nearly so while the dorsal margin is deflected dorsally tocreate an asymmetric expansion for example in Camaraminussaurus (Fig 9C) and Giraffatitan brancai (Fig 9D) In a fewsauropods however the ventral margin of the blade is alsodeflected ventrally to form a ldquoracquetminusshapedrdquo posterior exminuspansion This is seen in rebbachisaurids and some titanominussaurians eg some specimens of Alamosaurus Gilmore1922 (Gilmore 1946 fig 6) In Brontomerus the posteriorpart of the scapular blade is expanded in a characteristic manminusner the ventral margin is straight except for a posteroventral
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 85
50 cm
ventral ldquosteprdquo
acromion expansion
dorsal ldquostepsrdquo
Fig 8 Partial left scapula of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar Mountain Formaminustion of Utah OMNH 27761 in lateral view tentatively reconstructed after Giraffatitan brancai HMN Sa 9 (Janensch 1961 pl 15 1)
excursion two thirds of the way along the preserved portionafter which the margin continues parallel to its original traminusjectory so that the excursion appears as a gentle ldquosteprdquo Thedorsal margin is also ldquosteppedrdquo in this manner though withtwo distinct steps rather than one of which the more anterioris most strongly pronounced The net result of these featuresis that the dorsal and ventral borders of the scapula are bothstraight near the posterior extremity and that they are subminusparallel diverging by only about five degrees in the regionjust anterior to the rounded end of the posterior expansionThe step in the ventral border is not known in any othersauropod however the scapula of Neuquensaurus Powell1992 has a stepped dorsal border similar to that of Brontominusmerus (Huene 1929 via Glut 1997 275)
These characters of the scapula must be treated with somecaution however since this bone appears subject to morevariation than any other in the sauropod skeleton see for exminusample the range of shapes in scapulae of Giraffatitan brancai(Janensch 1961 pl 15 1ndash3) and in Camarasaurus supremus(Osborn and Mook 1921 figs 74ndash80)
Two small flat elements OMNH 66431 and 66432 are inminusterpreted as partial sternal plates (Fig 10) The medial edge ofeach is identifiable due to its rugose texture which formed theattachment site for cartilage joining the plates to each otherand to the sternal ribs The sternals are anteroposteriorly elonminusgate and mediolaterally narrow when complete they wereprobably at least three times as long as broad as in ldquoSaltaminussaurusrdquo robustus Huene 1929 (Huene 1929 via McIntosh1990 fig 169L) and proportionally longer than in any othersauropod including Saltasaurus loricatus (McIntosh 1990fig 169 Powell 1992 fig 30) The sternals are crescentic in
shape the anterior and posterior extremities curving laterallyaway from the midline This state was considered a titanominussaurian synapomorphy by Wilson (2002 268) but its distribuminustion is more complex in the current analysis being synapominusmorphic for Neosauropoda with losses in Flagellicaudata andCamarasaurus
86 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
Fig 10 Partial paired sternal plates of the camarasauromorph sauropodBrontomerus mcintoshi gen et sp nov from the Lower Cretaceous CedarMountain Formation of Utah OMNH 66431 and 66432 in ventral view
Fig 9 Scapulocoracoids and scapulae of sauropod dinosaurs scaled to same length of scapular blade from posterior point of glenoid to posterior margin ofblade A Mamenchisaurus youngi Young and Zhao 1972 holotype ZDM0083 left scapulocoracoid modified from Ouyang and Ye (2002 fig 22)B Diplodocus longus Hatcher 1901 USNM 10865 right scapulocoracoid reversed photograph by MPT C Camarasaurus supremus Cope 1877 AMNH5761 Sc 1 left scapula and AMNH 5761 Cor 1 left coracoid probably associated modified from Osborn and Mook (1921 figs 75 81a) D Giraffatitanbrancai (Janensch 1914) HMN Sa 9 left scapula modified from Janensch (1961 pl 15 1) E Rapetosaurus krausi Curry Rogers and Forster 2001holotype FMNH PR 2209 right scapula reversed modified from Curry Rogers (2009 fig 32) F Brontomerus mcintoshi gen et sp nov OMNH 27761left scapula tentatively reconstructed after Giraffatitan brancai
Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

excursion two thirds of the way along the preserved portionafter which the margin continues parallel to its original traminusjectory so that the excursion appears as a gentle ldquosteprdquo Thedorsal margin is also ldquosteppedrdquo in this manner though withtwo distinct steps rather than one of which the more anterioris most strongly pronounced The net result of these featuresis that the dorsal and ventral borders of the scapula are bothstraight near the posterior extremity and that they are subminusparallel diverging by only about five degrees in the regionjust anterior to the rounded end of the posterior expansionThe step in the ventral border is not known in any othersauropod however the scapula of Neuquensaurus Powell1992 has a stepped dorsal border similar to that of Brontominusmerus (Huene 1929 via Glut 1997 275)
These characters of the scapula must be treated with somecaution however since this bone appears subject to morevariation than any other in the sauropod skeleton see for exminusample the range of shapes in scapulae of Giraffatitan brancai(Janensch 1961 pl 15 1ndash3) and in Camarasaurus supremus(Osborn and Mook 1921 figs 74ndash80)
Two small flat elements OMNH 66431 and 66432 are inminusterpreted as partial sternal plates (Fig 10) The medial edge ofeach is identifiable due to its rugose texture which formed theattachment site for cartilage joining the plates to each otherand to the sternal ribs The sternals are anteroposteriorly elonminusgate and mediolaterally narrow when complete they wereprobably at least three times as long as broad as in ldquoSaltaminussaurusrdquo robustus Huene 1929 (Huene 1929 via McIntosh1990 fig 169L) and proportionally longer than in any othersauropod including Saltasaurus loricatus (McIntosh 1990fig 169 Powell 1992 fig 30) The sternals are crescentic in
shape the anterior and posterior extremities curving laterallyaway from the midline This state was considered a titanominussaurian synapomorphy by Wilson (2002 268) but its distribuminustion is more complex in the current analysis being synapominusmorphic for Neosauropoda with losses in Flagellicaudata andCamarasaurus
86 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
10 cm
Fig 10 Partial paired sternal plates of the camarasauromorph sauropodBrontomerus mcintoshi gen et sp nov from the Lower Cretaceous CedarMountain Formation of Utah OMNH 66431 and 66432 in ventral view
Fig 9 Scapulocoracoids and scapulae of sauropod dinosaurs scaled to same length of scapular blade from posterior point of glenoid to posterior margin ofblade A Mamenchisaurus youngi Young and Zhao 1972 holotype ZDM0083 left scapulocoracoid modified from Ouyang and Ye (2002 fig 22)B Diplodocus longus Hatcher 1901 USNM 10865 right scapulocoracoid reversed photograph by MPT C Camarasaurus supremus Cope 1877 AMNH5761 Sc 1 left scapula and AMNH 5761 Cor 1 left coracoid probably associated modified from Osborn and Mook (1921 figs 75 81a) D Giraffatitanbrancai (Janensch 1914) HMN Sa 9 left scapula modified from Janensch (1961 pl 15 1) E Rapetosaurus krausi Curry Rogers and Forster 2001holotype FMNH PR 2209 right scapula reversed modified from Curry Rogers (2009 fig 32) F Brontomerus mcintoshi gen et sp nov OMNH 27761left scapula tentatively reconstructed after Giraffatitan brancai
Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

Comparison with other EarlyCretaceous North Americansauropods
Brontomerus cannot be directly compared with AstrodonPleurocoelus Sonorasaurus or Sauroposeidon due to theabsence of overlapping material between these genera norwith Abydosaurus as the overlapping elements have not yetbeen adequately described (Carpenter and Tidwell 2005Ratkevich 1998 Wedel et al 2000b Chure et al 2010) Twoother sauropod dinosaurs are already known from the CedarMountain Formation Cedarosaurus from the Yellow CatMember and Venenosaurus from the Poison Strip MemberBrontomerus is from the stratigraphically higher RubyRanch Member and can be distinguished from both of thesetaxa as discussed below Since the Yellow Cat Member isBarremian in age the Poison Strip Member is Aptian and theupper part of the Ruby Ranch Member (where the HotelMesa Quarry is located) is AptianndashAlbian these three saurominuspods together probably span the last three ages of the EarlyCretaceous Brontomerus is also distinct from Paluxysaurusas shown below
CedarosaurusmdashCedarosaurus is known from a single parminustial semiminusdisarticulated skeleton DMNH 39045 described byTidwell et al (1999) Although much of the skeleton is preminusserved relatively few elements overlap with the material ofBrontomerus described above only dorsal vertebrae midminustominusposterior caudal vertebrae dorsal ribs partial scapulae andsternal plates are in common The single crushed presacralcentrum of Brontomerus cannot be usefully compared withthe dorsal vertebrae of Cedarosaurus beyond the observationthat the presacral bone texture described by Tidwell et al(1999 23) as ldquonumerous matrix filled chambers which areseparated by thin walls of bonerdquo is a good match Tidwell et al(1999) did not figure either the ribs or sternal plates of Cedarominussaurus but photographs supplied by Virginia Tidwell showthat its sternals are generally similar in shape to those ofBrontomerus though much larger and somewhat less elonminusgate Tidwell et al (1999 25) noted the absence of pneumaticforamina in the two preserved rib heads while recognizing thepossibility that anterior ribs might be pneumatic while posteminusrior ribs of the same individual lack this feature Photographsof a rib head were supplied by Virginia Tidwell and show littleresemblance to that of Brontomerus but damage to both articminusular facets hinders comparison The preserved portions ofCedarosaurus scapulae are from the anterior end and thereminusfore do not greatly overlap with the more posterior preservedportion of the Brontomerus scapula however the mid scapuminuslar region of Cedarosaurus differs in the possession of a morepronounced acromion process less straight ventral border andrelatively narrower scapular shaft Finally the midminustominusposteminusrior caudal vertebra of Brontomerus lacks the distinctive sharpridge extending along the edge of the neural arch described by
Tidwell et al (1999 25 fig 5) but other differences such asits greater elongation and greatly reduced neural spine are notinconsistent with the caudals of Cedarosaurus taking into acminuscount that the Brontomerus caudal is from a more distal posiminustion in the caudal sequence than any of those figured byTidwell et al (1999) In conclusion the preponderance of thescant morphological evidence supports the generic separationof Brontomerus from Cedarosaurus Furthermore the YellowCat Member is no younger than Barremian in age and may beBerriasianndashValanginian (Sames and Madsen 2007) givingCedarosaurus a minimum age of 121 Mya and a maximumage of ~140 Mya while the AptianndashAlbian position of the Hominustel Mesa Quarry at the top of the Ruby Ranch Member sugminusgests a much younger age While this gap does not in itselfprove generic separation it corroborates this conclusion
VenenosaurusmdashVenenosaurus was originally describedfrom a single small adult specimen DMNH 40932 althoughelements from one or more juveniles were also present in thequarry (Tidwell et al 2001 140) Some of the juvenile mateminusrial was subsequently described by Tidwell and Wilhite(2005) but none of this material overlaps with that of Brontominusmerus so comparisons with Venenosaurus must be made onthe basis of the type specimen alone The overlapping materialconsists of caudal vertebrae dorsal ribs and a left scapulaThe ldquodistalrdquo caudal of Venenosaurus figured by Tidwell et al(2001 fig 114C) is similar to the midminustominusposterior caudal ofBrontomerus in the proportions of the centrum and in the elonminusgation of the posteriorly directed postzygapophyseal ramusHowever the Venenosaurus distal caudal has very muchshorter prezygapophyses and a much less tall neural archwhich is set forward almost to the margin of the centrum ratherthan set back 10 of the centrumrsquos length It also lacks thecharacteristic ventral process that hangs from the postzygapominusphyseal ramus in Brontomerus and bears the postzygapominusphyseal facets The scapula of Venenosaurus figured byTidwell et al (2001 fig 115A) does not resemble that ofBrontomerus having a more curved ventral border a muchless steep ascent of the dorsal border towards the anterior exminuspansion a less expanded posterior blade and no sign of theldquostepsrdquo apparent on both borders of the blade of BrontomerusThe illustrated dorsal rib head of Venenosaurus (Tidwell et al2001 fig 119) differs from that of Brontomerus having avery short tuberculum a capitulum no broader than the ramusthat supports it and a very different pneumatic excavationwhich invades the bone in a dorsal direction penetrating thecapitulum rather than ventrally penetrating the shaft Thesedifferences of the ribs however may be less significant thanthey appear the tuberculum of the Venenosaurus rib is ldquosomeminuswhat erodedrdquo (Tidwell et al 2001 153) which may explain itsshortness the degree of expansion of the capitular head mayvary serially with the Venenosaurus rib being from a moreposterior position than the Brontomerus rib and pneumaticfeatures tend to vary both serially and between individualsand even on occasion between the two sides of a single eleminusment (eg in Xenoposeidon see Taylor and Naish 2007
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 87
1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

1552ndash1553) Nevertheless the balance of evidence stronglyindicates that Brontomerus is distinct from Venenosaurus
PaluxysaurusmdashThe internal structure of the presacral verteminusbrae of Paluxysaurus seems to be camellate like that ofBrontomerus based on the referred isolated dorsal centrumFWMSH 93Bminus10minus48 (Rose 2007 17 44ndash45) Some of theldquodistalrdquo caudal vertebrae of Paluxysaurus (Rose 2007 fig 17)somewhat resemble the Brontomerus midminustominusposterior caudalbut none has the very elongate prezygapophyses or ventralprocess of the postzygapophyseal ramus that characterize thelatter and the Paluxysaurus caudals have distinct dorsallyprojecting neural spines The figured ribs of Paluxysaurus(Rose 2007 fig 15) differ from that of Brontomerus in everyrespect save the pneumatic invasion of the ribminushead in the diminusrection of the shaft but these ribs are too badly damaged foruseful comparison and in any case are probably from a moreposterior position The prepared scapulae of Paluxysaurus(Rose 2007 fig 20) differ from that of Brontomerus in theirmore concave ventral border narrower blade less expandedposterior extremity and lack of ldquostepsrdquo on the anterior andposterior borders The sternal plates of Paluxysaurus are muchless proportionally elongate than those of Brontomerus Theilium of Paluxysaurus is not figured but according to the deminus
scription it differs in almost every respect from that of Brontominusmerus In conclusion significant differences in the midminuscaudalvertebrae scapulae and sternal plates and probably in the ribsdemonstrate that these two genera are separate
Phylogenetic analysisOn the assumption that the referred material belongs toBrontomerus we attempted to establish the phylogenetic pominussition of the new taxon by means of a phylogenetic analysisWe used the matrix of Harris (2006) as a basis adding thesingle new taxon to yield a matrix of 31 taxa (29 ingroupsand two outgroups) and 331 characters The only otherchange made was the rescoring of character 261 for Rapetominussaurus (ldquoin lateral view the anteroventralmost point on theiliac preacetabular processrdquo) changing it from state 1 (ldquoisposterior to the anteriormost part of the process (process issemicircular with posteroventral excursion of cartilage caprdquo)to state 0 (ldquois also the anteriormost point (preacetabular prominuscess is pointedrdquo) Brontomerus could be scored for 20 of the331 characters 6 of the total (Table 5)
Following Harris (2006) PAUP 40b10 (Swofford 2002)was used to perform a heuristic search using random stepminus
88 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Table 5 Character scores for the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah in the matrix used for the phylogenetic analysis of this paper Apart from the addition of Brontomerus and the rescoring of characminuster 261 for Rapetosaurus the matrix is identical to that of Harris (2006) Brontomerus is scored only for the listed characters Conventional anatomiminuscal nomenclature is here used in place of the avian nomenclature of Harris (2006)
Character Score184 Ratio of centrum lengthheight in middle caudal vertebrae 1 20185 Sharp ridge on lateral surface of middle caudal centra at archminusbody junction 0 absent186 Morphology of articular surfaces in middle caudal centra 0 subcircular187 Ventral longitudinal excavation on anterior and middle caudal centra 0 absent188 Morphology of anterior articular face of middle and posterior caudal centra 0 amphicoelousamphiplatyan
189 Position of neural arches over centra on middle caudal vertebrae 1 located mostly or entirely over anterior halfof centrum
191 Morphology of posterior caudal centra 0 cylindrical197 Proximal pneumatic foramina on dorsal ribs 1 present
198 Morphology of proximal ends of anterior dorsal ribs 1 strongly convex anteriorly and deeply concaveposteriorly
199 Crossminussectional shape of anterior dorsal ribs 0 subcircular
208 Size of scapular acromion 1 broad (dorsoventral width more than 150minimum width of scapular body)
210 Morphology of portion of acromion posterior to deltoid crest 0 flat or convex and decreases in mediolateralthickness toward posterior margin
212 Morphology of scapular body 2 posterior end racquetminusshaped (dorsoventrallyexpanded)
221 Morphology of sternal plate 2 elliptical with concave lateral margin259 Morphology of dorsal margin of ilium body (in lateral view) 1 semicircular (markedly convex)260 Position of dorsalmost point on ilium 1 anterior to base of pubic process
261 In lateral view the most anteroventral point on the iliac preacetabularprocess 0 is also the most anterior point
(preacetabular lobe is pointed)262 Orientation of preacetabular lobe of ilium with respect to axis of body 1 anterolateral in vertical plane
263 Size of ischiadic peduncle of ilium 1 low and rounded (long axis of ilium orientedanterodorsallyminusposteroventrally)
264 Projected line connecting articular surfaces of ischiadic and pubicpeduncles of ilium 0 passes ventral to ventral margin of postacetabular
lobe of ilium
wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

wise addition with 50 replicates and with maximum trees =500000 The analysis yielded 180 equally parsimonioustrees with length = 788 consistency index (CI) = 05228 reminustention index (RI) = 06848 and rescaled consistency index(RC) = 03581
The strict consensus tree (Fig 11A) is poorly resolvedwith Titanosauriformes collapsing into a broad polytomywithin which only Saltasauridae is differentiated This repreminussents a dramatic loss of resolution compared to the resultswithout Brontomerus (Harris 2006 fig 5A) A posteriori deleminustion of Brontomerus however yields a well resolved Macrominusnaria similar to that of Harrisrsquos (2006) analysis with Camaraminussaurus Brachiosaurus Riggs 1903 Euhelopus Romer 1956and Malawisaurus Jacobs Winkler Downs and Gomani1993 as successive singleton outgroups to a group of more deminusrived titanosaurians This demonstrates that the addition ofBrontomerus to the matrix does not cause instability in the reminuslationships between these more fully represented taxa and thatit is only the position of Brontomerus itself that is unstableAmong the equally most parsimonious positions of Brontominusmerus are as a nonminustitanosauriform camarasauromorph abasal titanosauriform a basal somphospondyl the sister taxonto Euhelopus a basal titanosaurian a basal lithostrotian and aderived nonminussaltasaurid lithostrotian One further step is suffiminuscient to place Brontomerus as a brachiosaurid a basal
(nonminuscamarasauromorph) macronarian a basal (nonminusdiplodominuscid) diplodocoid or even a nonminusneosauropod Three furthersteps are required for Brontomerus to be recovered as a saltaminussaurid specifically an opisthocoelicaudiine
In the 50 majority rule tree (Fig 11B) all the standardsauropod clades are recovered This tree shows the mostlikely position of Brontomerus as a basal somphospondyl ina trichotomy with Euhelopus and Titanosauria
In order to investigate a possible source of instability wealso reminusran the analysis with the Rapetosaurus ilium characminuster restored to the state given by Harris (2006) and found thatthe value of this character made no difference to the resultsall trees are one step longer with the new value but the topolminusogy of all consensus trees (strict semistrict and majorityrule) is unaffected by the changed scoring
When Brontomerus is scored for the holotype ilium onlyand the referred elements are ignored the strict consensusconstrains Brontomerus only to be a eusauropod more derivedthan Shunosaurus Dong Zhou and Zhang 1983
DiscussionFunctional anatomymdashThe functional significance of theunusual ilium of Brontomerus is difficult to interpret due to
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 89
Fig 11 Phylogenetic relationships of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah produced using PAUP 40b10 on the matrix of Harris (2006) augmented by Brontomerus having 31 taxa and 331 characters Left sidestrict consensus of 180 most parsimonious trees (length = 788 CI = 05228 RI = 06848 RC = 03581) right side 50 majority rule consensus
the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

the absence of functionally related skeletal elements such asthe pubis and ischium posterior dorsal and anterior caudalvertebrae and femur In life these elements work together asa functional complex each affecting the function of the othminusers Some speculation is warranted however
The large preacetabular blade of the ilium provides an anminuschor for large protraction muscles which would have alminuslowed the leg to be moved forwards powerfully this is surminusprising as femoral retraction is required for forward locomominustion requiring large muscles to pull the femur backwardsand the ilium of Brontomerus offers almost no attachmentarea for posteriorly located muscles The caudofemoralismuscle is the main power generator in locomotion for extantreptiles and osteological correlates such as the fourth trominuschanter of the femur indicate that this was also true ofnonminusavian dinosaurs This muscle connects the femur to thetail so in the absence of proximal caudal vertebrae and chevminusrons of Brontomerus it is not possible to determine whetherthe femoral retractors were enlarged in proportion with theprotractors If so then this increase in musculature would inminusdicate that Brontomerus may have been unusually athleticfor a sauropod Another possibility is that powerful femoralprotraction was useful for delivering a strong kick (Fig 12)
As well as protractors the preacetabular blade of the
ilium also anchors abductors (ie muscles which draw theleg laterally away from the median plane) These muscles areimportant for creating abduction torque when standing andmay have facilitated occasional bipedal stance or even limminusited bipedal walking
A third possibility is that the proportionally large legmuscles were required to drive unusually long legs Thelarge anterior expansion of the scapula provides weak addiminustional support for this hypothesis If this interpretation werecorrect Brontomerus might have resembled a giraffe in grossmorphology
Did sauropods really decline in the Early Cretaceous ofNorth AmericamdashApart from Cetiosaurus oxoniensis Philminuslips 1871 the first reasonably complete remains of sauropodswere discovered in the Morrison Formation of the AmericanWest in the 1870s and 1880s and these remains provided thefirst accurate understanding of the size and bauplan of theclade (Cope 1877 1878 Marsh 1877) Sauropod remains hadpreviously been assumed to represent gigantic crocodiles(Owen 1859) or pterosaurminusbird intermediates (Seeley 1870)Nineteenth century workers also quickly recognized thatsauropods were common in the Morrison Formation and esminussentially absent in the Early Cretaceous ldquoDakotardquo Formations
90 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Fig 12 Speculative life restoration of the camarasauromorph sauropod Brontomerus mcintoshi gen et sp nov from the Lower Cretaceous Cedar MountainFormation of Utah Adult individual (sized according to the referred scapula) protects juvenile (sized according to the holotype ilium) from a Utahraptorthe enlarged femoral protractors may have enabled a powerful kick Executed by Francisco Gascoacute under direction from MPT and MJW reproduced withpermission
which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

which at the time included the Cloverly and Cedar MountainFormations in addition to the Dakota Formation itself (Witzkeand Ludvigson 1994) In fact the absence of sauropods fromEarly Cretaceous rocks was so wellminusestablished that Marsh(1888) erroneously identified the age of the Potomac Formaminustion (now the Potomac Group) as Late Jurassic based on thepresence of the sauropod Pleurocoelus (now synonymizedwith Astrodon Carpenter and Tidwell 2005)
In the following century the infrequent recovery of saurominuspod material from Early Cretaceous deposits across NorthAmerica (eg Larkin 1910 Ostrom 1970 Langston 1974) reminusinforced the perception that the Late Jurassic was the apexof sauropod success and the Early Cretaceous its nadir Thisapparent dichotomy became entrenched in both scientific(Bakker 1978 Behrensmeyer et al 1992) and popular (Bakker1986) accounts of the evolutionary history and paleoecologyof dinosaurs including previous publications by the junior auminusthors (Wedel et al 2000a b Wedel and Cifelli 2005) Howminusever a wave of discovery of new material of Early Cretaceoussauropods (Britt and Stadtman 1996 1997 Britt et al 19971998 2004 Winkler et al 1997a Gomani et al 1999 Burgeet al 2000 Maxwell and Cifelli 2000 Burge and Bird 2001Coulson et al 2004 Bird 2005 Tidwell and Carpenter 2007)and the description of many new taxa in the past decade (reminusviewed below and in Table 6) prompts us to reconsider the
question of whether sauropods really did decline in the EarlyCretaceous of North America
The diversity of Early Cretaceous North American saurominuspodsmdashTo date eight sauropod genera have been describedfrom the Early Cretaceous of North America Astrodon inminuscluding Pleurocoelus (Leidy 1865 Marsh 1888) Sonoraminussaurus (Ratkevich 1998) Cedarosaurus (Tidwell et al 1999)Sauroposeidon (Wedel et al 2000a) Venenosaurus (Tidwellet al 2001) Paluxysaurus (Rose 2007) Abydosaurus (Chureet al 2010) and now Brontomerus (this paper)
The pool of named taxa will always be only a subset ofthe taxa that have been or are being studied and the taxa unminusder study are in turn only a subset of the pool of potentiallynew taxa that have been discovered in the field excavatedand prepared Almost all of the Early Cretaceous NorthAmerican sauropods that have been recently described or arecurrently under study were discovered and excavated in the1990s or 2000s Sauropod fossils are often logistically chalminuslenging to excavate prepare manipulate photograph andeven measure simply because they are so large and heavy Inour experience the process of preparation and description ofa new sauropod can be very protracted compared to the prepminusaration and description of other smaller vertebrates The reminuscently named taxa represent the first ldquograduatesrdquo of the arduminus
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 91
Table 6 Early Cretaceous North American sauropod diversity MNI = Minimum Number of Individuals
Unit Stage Taxa MNI References
Arundel Formation AptianndashAlbian Astrodon (= Pleurocoelus) 2+ Johnston (1859) Leidy (1865) Marsh (1888) Lull(1911) Kingham (1962) Tidwell and Carpenter (2005)
Antlers Formation AptianndashAlbianSauroposeidon proteles 1 Wedel et al (2000a b)
isolated coracoid 1 Larkin (1910)teeth Cifelli et al (1997a)
Trinity Group AptianndashAlbian ldquoPleurocoelusrdquo 2+ Langston (1974) Gallup (1989)
Twin MountainsFormation Aptian
Paluxysaurus 4 Winkler et al (1997a) Gomani et al (1999)Rose (2007)
unnamed braincase 1 Carpenter and Tidwell (2003)Turney Ranch Formation AlbianndashCenomanian Sonorasaurus thompsoni 1 Ratkevich (1998) Curtice (2000)
CloverlyFormation
Unit V
AptianndashAlbian
cf Brachiosaurus 3 Wedel (2000)Unit VI titanosaurian humerus 1 Ostrom (1970)
Unit VIIunnamed basaltitanosauriform 1 Ostrom (1970) DrsquoEmic and Britt (2008)
cf Sauroposeidon 1 Ostrom (1970) Wedel et al (2000a b)
CedarMountainFormation
Yellow Cat Barremian
Dalton Wells titanosaurian 14 Britt and Stadtman (1996 1997)Britt et al (1997 1998 2004)
Dalton Wells camarasaur 2 Britt et al (1997 2004)Dalton Wells brachiosaur 3 Britt et al (2004)Cedarosaurus weiskopfae 1 Tidwell et al (1999)
titanosaurian cervicals 1 Tidwell and Carpenter (2007)Poison Strip Aptian Venenosaurus dicrocei 2 Tidwell et al (2001) Tidwell and Wilhite (2005)
Ruby Ranch AptianndashAlbian
Long Walk brachiosaur 2 DeCourten (1991)
CEU brachiosaur 6 Burge et al (2000) Burge and Bird (2001)Coulson et al (2004) Bird (2005)
Brontomerus mcintoshi 2 this study
Mussentuchit AlbianndashCenomanianAbydosaurus mcintoshi 4 Chure et al (2010)
dwarf teeth Maxwell and Cifelli (2000)
ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

ous descriptive process and many more new taxa will probaminusbly be described in the coming decade These includendash From the Yellow Cat Member of the Cedar Mountain Forminus
mation a camarasaurid a brachiosaurid and a titanominussaurian from the Dalton Wells Quarry all represented bymultiple individuals (Britt and Stadtman 1996 1997 Brittet al 1997 1998 2004)
ndash From the Ruby Ranch Member of the Cedar Mountain Forminusmation the Long Walk Quarry brachiosaurid (DeCourten1991) which is currently under study by Virginia Tidwelland Kenneth Carpenter and may represent a new taxon andthe CEU brachiosaurid which is represented by remains ofat least 6 individuals including a cervical vertebra 128 cmlong and which is apparently similar to or perhaps evensynonymous with Sauroposeidon (Burge et al 2000 Burgeand Bird 2001 Coulson et al 2004 Bird 2005)
ndash From Unit VII of the Cloverly Formation numerous verteminusbrae and appendicular elements of a possibly new basaltitanosauriform (interpreted as a titanosaurian by Ostrom1970) which is currently under study by Michael DrsquoEmicat the University of Michigan and possibly a second asminusyetunidentified sauropod taxon (DrsquoEmic and Britt 2008)
Additional sauropod remains that are extremely incomminusplete or not diagnostic and therefore unfit for naming areknown from several formations Some of these may eventuminusally be referred to existing or forthcoming named taxa ormight attain holotype status following the discovery of morecomplete material or additional study This material includesndash From the Antlers Formation the isolated coracoid descriminus
bed by Larkin (1910) and isolated teeth (Cifelli et al1997a) Available evidence neither supports nor contradictsthe possibility that this material pertains to Sauroposeidon(for which the coracoid is unknown) although the coracoiddoes not resemble those of other brachiosaurids includingBrachiosaurus altithorax (Riggs 1904) and Giraffatitanbrancai (Janensch 1950)
ndash From the Yellow Cat Member of the Cedar Mountain Forminusmation a series of partial cervical vertebrae of a titanominussaurian (Tidwell and Carpenter 2007)
ndash From the Mussentuchit Member of the Cedar MountainFormation numerous teeth similar to those of Astrodonthat may represent a dwarf titanosauriform (Maxwell andCifelli 2000)
ndash From Unit V of the Cloverly Formation a brachiosauridvery similar to Brachiosaurus (Wedel 2000)
ndash From Unit VI of the Cloverly Formation a humerus simiminuslar to those of titanosaurians (Ostrom 1970)
ndash From Unit VII of the Cloverly Formation a very elongatecervical vertebra of a juvenile sauropod that shares severalapomorphies with Sauroposeidon (Ostrom 1970 Wedel etal 2000a b Wedel and Cifelli 2005)
ndash From the Trinity Group the titanosauriform braincase deminusscribed by Tidwell and Carpenter (2003) and postcranialmaterial from multiple localities that cannot be referred toPaluxysaurus with certainty (Langston 1974 Gallup 1989Rose 2007)
If all of the apparently diagnostic sets of material (firstlist above) were named as new genera the generic diversityof Early Cretaceous North American sauropods could risefrom the current total of 8 to as many as 15 On the otherhand some of this undescribed material may eventually bereferred to existing genera and even some of the eight existminusing genera might eventually be synonymized For examplethe Sauroposeidon type material might represent the neck ofCedarosaurus or of Venenosaurus in the former case Saurominusposeidon would become a junior synonym of Cedarosaurusand in the latter Venenosaurus would become a junior synminusonym of Sauroposeidon Note however that no existing eviminusdence supports these hypothetical synonymizations and theyseem unlikely given the temporal and geographic distanceseparating the taxa concerned
The statistical analysis of Mannion et al (2010 17ndash18)reported that when apparent diversity is corrected for availminusability of relevantlyminusaged rocks the Early Cretaceous repreminussents a time of low diversity for sauropods However theiranalysis was based only on validly named genera and asshown here there is evidence for many additional genera thatare yet to be formally recognized
The diversity of Early Cretaceous North American saurominuspods has traditionally been contrasted with that of the sauropodtaxa of the Late Jurassic Morrison Formation which is reminusviewed below
The diversity of Morrison sauropodsmdashSix wellminusknowngenera have been recognized from the Morrison Formation forover a century Apatosaurus Barosaurus Brachiosaurus Caminusmarasaurus Diplodocus and Haplocanthosaurus Severalother taxa that have long been recognized are poorly repreminussented but nevertheless probably valid Dystrophaeus Amphiminuscoelias and ldquoApatosaurusrdquo minimus which is not referable toApatosaurus and is probably not even a diplodocoid(McIntosh 1990 398 Upchurch et al 2004a 298) In addiminustion several new taxa have been described in recent decadesor are currently under study Supersaurus EobrontosaurusSuuwassea and an apparently new diplodocoid announced byVietti and Hartman (2004) Upchurch et al (2004b 307) sugminusgested that Eobrontosaurus is referrable to Camarasaurus butthis assertion was not based on the identification of any sharedapomorphies The holotype is distinct from both Camaraminussaurus and Apatosaurus and probably represents a valid taxon(Ray Wilhite personal communication May 2008) Finally atleast five Morrison Formation genera have been synonymizedin recent years Cathetosaurus is now classified as a distinctspecies of Camarasaurus (C lewisi McIntosh et al 1996)Ultrasauros and Dystylosaurus are both junior synonyms ofSupersaurus (Curtice et al 1996 Curtice and Stadtman 2001)Elosaurus is a juvenile Apatosaurus (Upchurch et al 2004b)and Seismosaurus has been synonymized with Diplodocus asa distinct species D hallorum by Lucas et al (2006) and asthe existing species D longus by Lovelace et al (2008) Thisleaves a current total of 12 named genera plus one potentiallynew genus under study
92 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

Early Cretaceous and Morrison sauropods comparedmdashIf we simply count existing genera then Early CretaceousNorth American sauropods are only two thirds as diverse astheir Morrison forebears (8 genera compared to 12) If we inminusclude all of the potentially new taxa that are currently understudy the diversity of sauropods in the Early Cretaceous ofNorth America may eventually exceed that of the Morrison(16 genera to 13) However so simple a comparison of thesauropod faunas of the North American Late Jurassic andEarly Cretaceous is misleading for the following reasons
First the North American Early Cretaceous rock unitsseparately or together are simply dwarfed by the MorrisonFormation Foster (2003) listed more than 270 vertebrate lominuscalities in the Morrison Formation in Arizona ColoradoIdaho Kansas Montana New Mexico Oklahoma SouthDakota Utah and Wyoming Based on the sheer volume andoutcrop area of the Morrison Formation we might expect itsvertebrate fauna to appear more diverse than those of theEarly Cretaceous units
Second the Morrison Formation is exposed along theRocky Mountains and in part of the Great Basin whereas theNorth American Lower Cretaceous units are scattered acrossthe continent from the Arundel Formation on the eastern seaminusboard to the Turney Ranch Formation in the desert southwestand from the Cloverly Formation in southern Montana to theTrinity Group in central Texas Although there is some eviminusdence for biogeographic differentiation within the MorrisonFormation (Harris and Dodson 2004) the Lower Cretaceousunits span much more of the continent and much more timeTemporal differences among the Lower Cretaceous dinosaurfaunas are well established (Kirkland et al 1997) and theremay have been biogeographic differentiation as well (Jacobsand Winkler 1998 Nydam and Cifelli 2002) Most relevant inthis context are the AptianndashAlbian assemblages of the TrinityGroup Texas (Winkler et al 1990) and Oklahoma (Brinkmanet al 1998) and the Arundel Clay facies of the PotomacGroup Maryland (Kranz 1998) On this basis we might thereminusfore expect the combined fauna of the Lower Cretaceous unitsto be more diverse than that of the Morrison Formation
Third the Morrison Formation and the various Lower Creminustaceous terrestrial units do not represent equivalent amounts oftime The Morrison Formation represents about seven millionyears of deposition from 155ndash148 Mya (Kowallis et al 1998)The most temporally extensive North American Lower Cretaminusceous unit is the Cedar Mountain Formation the dinosaurminusbearing parts of which span at least 28 million years from theBarremian (126 plusmn 25 Mya Kirkland and Madsen 2007) to theearliest Cenomanian (9837 plusmn 007 Mya Cifelli et al 1997b)If the Yellow Cat Member of the Cedar Mountain Formationis indeed BerriasianndashValanginian (Sames and Madsen 2007)the Cedar Mountain Formation may span 40 million years ormore (albeit with large depositional hiatuses) All of the otherLower Cretaceous units fall into the span of time representedby the Cedar Mountain Formation which is four to six timesas long as that represented by the Morrison Formation On thisbasis we might also expect the faunas of the Lower Cretaminus
ceous units or even that of the Cedar Mountain Formation byitself to be more diverse than that of the Morrison Formation
Fourth the Morrison Formation has been intensively samminuspled for more than 130 years and numerous expeditions havegone to the Morrison Formation in the express hope of findingcomplete skeletons of sauropods The less spectacular mateminusrial of the Lower Cretaceous units has drawn much less attenminustion from both paleontologists and the public The CloverlyFormation has only been intensively sampled since the 1960sand the Antlers and Cedar Mountain Formations since the1990s (although earlier collections had been made from all ofthese formations) The explosive growth of our knowledge ofEarly Cretaceous sauropods in North America is a direct resultof more intensive sampling of all of the North AmericanLower Cretaceous units in the past two to three decades Thisnew pulse of collecting is mitigating the collector bias that faminusvored the Morrison Formation but a serious imbalance stillpersists and may never be completely alleviated This is partlybecause fieldwork in the Morrison Formation has not stoppedand probably will not in the foreseeable future and new saurominuspods continue to be discovered (eg Vietti and Hartman2004) and described (eg Suuwassea Harris and Dodson2004) from the Morrison Formation
In summary four factors bias our perception of sauropoddiversity in the Late Jurassic and Early Cretaceous of NorthAmerica the Morrison Formation is larger than all of theLower Cretaceous units combined and has been more intenminussively sampled and for much longer but the Lower Cretaminusceous units span more space and time It is not clear howthese various biases should be weighted Nevertheless threemajor conclusions can be drawn
First the North American sauropod fauna shifted from beminusing diplodocoidminusdominated during the Late Jurassic with alow diversity of macronarians to being exclusively composedof macronarians during the Early Cretaceous In particular theMorrison Formation is dominated by diplodocids which seemto have hit their global peak of diversity during the Kimmeminusridgian and Tithonian (including Tornieria and Australodocusin Africa Remes 2006 2007) and then swiftly disappearedNo definitive Cretaceous diplodocid is known from anywherein the world although other diplodocoid clades (Rebbachiminussauridae and Dicraeosauridae) persisted on southern contiminusnents (Salgado and Bonaparte 1991 Sereno et al 1999 2007Rauhut et al 2005) Upchurch and Mannion (2009) referredan isolated partial anterior caudal vertebra from the Early Creminustaceous of China to Diplodocidae but this referral has beenquestioned (Whitlock et al 2011) Thus the clade Diplominusdocidae generally considered successful seems to have beenlimited in both time and space (Taylor 2006)
Second per fauna and per quarry diversity of sauropods ismuch lower in the Lower Cretaceous than in the Morrison Forminusmation The Cedar Mountain Formation may eventually haveas many as 10 named genera of sauropods but these are spreadacross at least three members each with their own distinctdinosaurian faunas and no single fauna is yet known to conminustain more than three sauropod genera Large quarries in the
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 93
Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

Morrison Formation often have more than one sauropod genuspresent and the MarshminusFelch and Dry Mesa quarries eachcontain five sauropod generamdashuntil recently more than wereknown from the entire Lower Cretaceous of North America
Third whereas sauropods are the most abundant large verminustebrates in the Morrison Formation they are among the rarestelements in their respective faunas in the North AmericanLower Cretaceous The ldquobig sixrdquo Morrison Formation genminuseramdashApatosaurus Barosaurus Brachiosaurus Camarasauminusrus Diplodocus and Haplocanthosaurusmdashare all representedby more than 10 individuals and Camarasaurus Diplodocusand Apatosaurus are represented by close to 100 or more (Fosminuster 2003 table 5) In contrast the Early Cretaceous sauropodsknown from the most specimens are the Dalton Wells titanominussaurian (14) the CEU brachiosaur (6) Paluxysaurus (4) andAbydosaurus (4) of which the first two are currently undesminuscribed Most of the named taxa are known from only one ortwo individuals
SummarymdashSauropods are both diverse and abundant in theMorrison Formation multiple sauropod genera are oftenfound together in a quarry and these sauropods are mostlydiplodocids and Camarasaurus Representatives of othersauropod clades are sparse In particular macronarians arenot diverse although Camarasaurus is abundant and titanominussauriforms are represented only by Brachiosaurus (Haplominuscanthosaurus may also be a macronarian having been recovminusered within this clade by the phylogenetic analyses of Wilsonand Sereno [1998] and Upchurch et al [2004a] but as anonminusdiplodocimorph diplodocoid by Wilson [2002] Howminusever it is among the rarest of Morrison Formation saurominuspods)
In contrast during the Early Cretaceous of North Amerminusica sauropods were relatively rare (at least compared to theirMorrison Formation abundance) Although the total diverminussity of sauropods across all North American Early Cretaminusceous formations is high the diversity within each fauna islow and cominusoccurrence of multiple taxa at a single locality isthe exception rather than the rule All known Early Cretaminusceous North American sauropods are macronarians andmost are brachiosaurids or other basal titanosauriforms alminusthough a few titanosaurians and one probable camarasauridare also present The implications of these differences forMesozoic paleoecology especially energy flow and nutrientcycling in terrestrial ecosystems are only beginning to be exminusplored (eg Farlow et al 2010)
Earliest Cretaceous sauropods and the fate of DiplodominuscidaemdashUnderstanding of the history and evolution of saurominuspods in the midminusMesozoic has been impaired by the unavailminusability of rocks from the earliest Cretaceous in many parts ofthe world Possibly for this reason the fossil record of diplominusdocids is extremely limited with all known definitive diplominusdocid genera worldwide having arisen in the KimmeriminusdgianndashTithonian (Taylor 2006 137)mdashalthough nonminusdiplodominuscid diplodocoids notably rebbachisaurids are known to havepersisted well into the Cretaceous However it is also possible
that rather than becoming extinct at the end of the Jurassicdiplodocids persisted into the earliest Cretaceous and wereonly gradually replaced by the macronarian sauropod faunathat characterizes the Cedar Mountain Formation and otherNorth American Lower Cretaceous units
This idea can be investigated by searching for lateminussurvivminusing diplodocids in the lowest parts of the Cedar MountainFormation and in earliest Cretaceous strata outside NorthAmerica Until the recognition of the possible diplodocidDinheirosaurus Bonaparte and Mateus 1999 from the LateJurassic of Portugal no diplodocid genus had been namedfrom outside North America although the type species ofLourinhasaurus Dantas Sanz Silva Ortega Santos andCachatildeo 1998 ldquoApatosaurusrdquo alenquerensis Lapparent andZbyszewski 1957 was considered by its describers to repreminussent a diplodocid and the referred species ldquoBarosaurusrdquoafricanus Fraas 1908 was known from Tendaguru in Tanzaminusnia The African ldquoBarosaurusrdquo material is now recognised ascomprising two distinct new diplodocid genera TornieriaSternfeld 1911 (Remes 2006) and Australodocus Remes2007 both in fact belonging to Diplodocinae so the exisminustence of Late Jurassic diplodocids is now well establishedoutside North America with representatives in both Europeand Africa Both the Portuguese Lourinhă Formation and theAfrican Tendaguru Formation end at the JurassicCretaceousboundary but other latestminusJurassic formations in Portugal areconformably overlain by Lower Cretaceous strata correlativewith the Wealden Supergroup of England It is in these stratathat Early Cretaceous diplodocids may most usefully besought and there are signs that diplodocids may indeed havebeen present in the Wealden Taylor and Naish (2007 1560)reported the presence of a large sauropod metacarpal fromthe Hastings Group of the Wealden which has been identifiedas diplodocid and Naish and Martill (2001 232ndash234) disminuscussed other putative though not definitive Wealden diplominusdocid material Thus it seems likely that diplodocids did inminusdeed survive into the Cretaceous at least in Europe and posminussibly also in North America and that their apparent endminusJuminusrassic extinction is actually an artifact produced by the lackof representative strata from the earliest Cretaceous
ConclusionsBrontomerus mcintoshi is a new genus and species of macrominusnarian sauropod from the Ruby Ranch Member of the CedarMountain Formation of Utah The new taxon is represented byat least two individuals of different sizes probably a juvenileand an adult It is clearly separate from all previously knownCedar Mountain Formation sauropods and is distinguishedfrom all other sauropods by several unique characters of theilium and the scapula The new taxon is probably a fairly basalcamarasauromorph although resolution is poor due to the inminuscompleteness of the material The distinctive characters of theilium (eg huge preacetabular blade no postacetabular bladevery tall overall transversely thin) probably have functional
94 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

significance but this is difficult to assess in the absence ofother pelvic elements femora and proximal caudals
The improving record of Early Cretaceous sauropods inNorth America is extended by the new genus so that geminusnericminuslevel diversity of sauropods in this epoch now apminusproaches that of the Late Jurassic The most striking differminusences between Late Jurassic and Early Cretaceous sauropodsin North America is that the former are abundant and domiminusnated by diplodocids whereas the latter are comparativelyscarce and dominated by macronarians It is currently imposminussible to determine whether this shift happened suddenly orgradually over many millions of years in the earliest Cretaminusceous It is natural to assume that if the shift was sudden ithappened at the end of the Jurassic but that is not necessarilythe case The timing and tempo of this faunal shift remain unminuscertain future inferences will have to be based on improvedunderstanding of global changes in conditions in the earliestCretaceous and careful analysis of faunal changes on neighminusbouring continents especially Europe
AcknowledgementsWe thank Nicholas Czaplewski (OMNH Norman USA) and Jeff Person(OMNH) for access to the Brontomerus mcintoshi type material andKyle Davies (OMNH) for assistance with logistical matters Randall BIrmis (Utah Museum of Natural History Salt Lake City USA) suppliedthe photographs of the Brontomerus caudal vertebra Virginia Tidwell(Denver Museum of Natural History Denver USA) kindly provided unminuspublished photographs of Cedarosaurus and Venenosaurus material Wethank James I Kirkland (Utah Geological Survey Salt Lake City USA)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and Ray Wilhite (Louisiana State University Baton Rouge USA) forpermission to cite personal communications and John R Hutchinson(Royal Veterinary College London UK) for discussions on the possiblefunctional significance of the new taxonrsquos anatomy Nick Pharris (Uniminusversity of Michigan) helped us to arrive at the name Brontomerus afterwe had proposed an inferior version with the same meaning Ben Creislerand T Michael Keesey (both independent Seattle and Glendale USArespectively) provided assistance with the etymology and pronunciationScott Hartman (independent Madison USA) allowed us to use hisCamarasaurus grandis reconstruction as the basis for the skeletal invenminustory (Fig 1) and Francisco Gascoacute (Fundacion Dinopolis Teruel Spain)executed the life restoration (Fig 12) Mike DrsquoEmic (University ofMichigan Ann Arbor USA) and an anonymous reviewer commented onan earlier version of this work the current version was ably and helpfullyreviewed by Andreas Christian (University of Flensburg Germany)Daniela SchwarzminusWings (Museum fuumlr Naturkunde Berlin Germany)and another anonymous reviewer We also thank Brian Davis (Univerminussity of Oklahoma Norman USA) for his timely editorial handling of themanuscript Most of all we thank John S McIntosh (Wesleyan Univerminussity Middletown USA) in whose honor the new species is named forhis many decades of dedicated and enthusiastic work on sauropods andfor inspiring all of us who follow in his footsteps
ReferencesBader KS 2003 Insect Trace Fossils on Dinosaur Bones from the Upper
Jurassic Morrison Formation Northeastern Wyoming and their Use in
Vertebrate Taphonomy 119 pp + x Unpublished MSc thesis Departminusment of Geology University of Kansas Lawrence
Bakker RT 1978 Dinosaur feeding behavior and the origin of floweringplants Nature 274 661ndash663 [CrossRef]
Bakker RT 1986 The Dinosaur Heresies New Theories Unlocking theMystery of the Dinosaurs and their Extinction 481 pp William Morrowamp Company New York
Behrensmeyer AK Damuth JD DiMichele WA Potts R SuesHminusD and Wing SL 1992 Terrestrial Ecosystems Through TimeEvolutionary Paleoecology of Terrestrial Plants and Animals 568 ppUniversity of Chicago Press Chicago
Bird J 2005 New finds at the Price River II site Cedar Mountain Formaminustion in eastern Utah Journal of Vertebrate Paleontology 25 (Suppleminusment to No 3) 37A
Bird RT 1985 Bones for Barnum Brown Adventures of a DinosaurHunter 225 pp Texas Christian University Press Fort Worth
Bonaparte JF 1986 Les dinosaures (Carnosaures Allosaurideacutes Saurominuspodes Ceacutetiosaurideacutes) du Jurassique moyen de Cerro Coacutendor (ChubutArgentina) Annales de Paleacuteontologie 72 325ndash386
Bonaparte JF and Mateus O 1999 A new diplodocid Dinheirosauruslourinhanensis gen et sp nov from the Late Jurassic beds of PortugalRevista del Museo Argentino de Ciencias Naturales 5 13ndash29
Bonaparte JF and Powell JE 1980 A continental assemblage of tetraminuspods from the Upper Cretaceous beds of El Brete northwestern Argenminustina (SauropodandashCoelurosauriandashCarnosauriandashAves) Meacutemoires de laSocieacuteteacute Geacuteologique de France Nouvelle Seacuterie 139 19ndash28
Brinkman DL Cifelli RL and Czaplewski NJ 1998 First occurrenceof Deinonychus antirrhopus (Dinosauria Theropoda) from the AntlersFormation (Lower Cretaceous AptianndashAlbian) of Oklahoma BulletinOklahoma Geological Survey 146 1ndash26
Britt BB and Stadtman KL 1996 The Early Cretaceous Dalton Wells diminusnosaur fauna and the earliest North American titanosaurid sauropodJournal of Vertebrate Paleontology 16 (Supplement to No 3) 24A
Britt BB and Stadtman KL 1997 Dalton Wells Quarry In PJ Currieand K Padian (eds) The Encyclopedia of Dinosaurs 165ndash166 Acaminusdemic Press San Diego
Britt B Eberth D Scheetz R and Greenhalgh B 2004 Taphonomy ofthe Dalton Wells dinosaur quarry (Cedar Mountain Formation LowerCretaceous Utah) Journal of Vertebrate Paleontology 24 (Supplementto No 3) 41A
Britt BB Scheetz RD McIntosh JS and Stadtman KL 1998 Osteominuslogical characters of an Early Cretaceous titanosaurid sauropod dinominussaur from the Cedar Mountain Formation of Utah Journal of VertebratePaleontology 18 (Supplement to No 3) 29A
Britt BB Stadtman KL Scheetz RD and McIntosh JS 1997 Camaraminussaurid and titanosaurid sauropods from the Early Cretaceous Dalton WellsQuarry (Cedar Mountain Formation) Utah Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 34A
Burge DL and Bird JH 2001 Fauna of the Price River II Quarry of eastminusern Utah Journal of Vertebrate Paleontology 21 (Supplement to No 3)37A
Burge DL Bird JH Britt BB Chure DJ and Scheetz RL 2000 Abrachiosaurid from the Ruby Ranch Mbr (Cedar Mountain Fm) nearPrice Utah and sauropod faunal change across the JurassicndashCretaceousboundary of North America Journal of Vertebrate Paleontology 20(Supplement to No 3) 32A
Calvo JO and Salgado L 1995 Rebbachisaurus tessonei sp nov a newSauropoda from the AlbianndashCenomanian of Argentina new evidenceon the origin of the Diplodocidae Gaia 11 13ndash33
Carpenter K and McIntosh JS 1994 Upper Jurassic sauropod babiesfrom the Morrison Formation In K Carpenter KF Hirsch and JRHorner (eds) Dinosaur Eggs and Babies 265ndash278 Cambridge Uniminusversity Press Cambridge
Carpenter K and Tidwell V 2005 Reassessment of the Early CretaceousSauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes) In VTidwell and K Carpenter (eds) Thunder Lizards the SauropodomorphDinosaurs 78ndash114 Indiana University Press Bloomington Indiana
Chure D Britt BB Whitlock JA and Wilson JA 2010 First complete
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 95
sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

sauropod dinosaur skull from the Cretaceous of the Americas and theevolution of sauropod dentition Naturwissenschaften 97 379ndash391[CrossRef]
Cifelli RL Madsen SK and Larson EM 1996 Screenwashing and asminussociated techniques for the recovery of microvertebrate fossils In RLCifelli (ed) Techniques for Recovery and Preparation of Microminusvertebrate Fossils 1ndash24 Oklahoma Geological Survey Special Publiminuscation 96minus4
Cifelli RL Gardner JD Nydam RL and Brinkman DL 1997a Addiminustions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous)southeastern Oklahoma Oklahoma Geology Notes 57 124ndash131
Cifelli RL Kirkland JI Weil A Deino AL and Kowallis BJ 1997bHighminusprecision 40Ar39Ar geochronology and the advent of North Amerminusicarsquos Late Cretaceous terrestrial fauna Proceedings of the National Acadminusemy of Sciences USA 94 11163ndash11167 [CrossRef]
Cope ED 1877 On a gigantic saurian from the Dakota epoch of ColoradoPaleontology Bulletin 25 5ndash10
Cope ED 1878 On the saurians recently discovered in the Dakota beds ofColorado American Naturalist 12 71ndash85 [CrossRef]
Coria RA 1994 On a monospecific assemblage of sauropod dinosaurs fromPatagonia implications for gregarious behaviour Gaia 10 209ndash213
Coulson A Barrick R Straight W Decherd S and Bird J 2004 Descripminustion of the new brachiosaurid (Dinosauria Sauropoda) from the RubyRanch Member (Cretaceous Albian) of the Cedar Mountain FormationUtah Journal of Vertebrate Paleontology 24 (Supplement to No 3) 48A
Curry Rogers K 2009 The postcranial osteology of Rapetosaurus krausei(Sauropoda Titanosauria) from the Late Cretaceous of MadagascarJournal of Vertebrate Paleontology 29 1046ndash1086 [CrossRef]
Curry Rogers K and Forster CA 2001 Last of the dinosaur titans a newsauropod from Madagascar Nature 412 530ndash534 [CrossRef]
Curtice BD 2000 The axial skeleton of Sonorasaurus thompsoni Ratkevich1998 Mesa Southwest Museum Bulletin 7 83ndash87
Curtice BD and Stadtman KL 2001 The demise of Dystylosaurus edwiniand a revision of Supersaurus vivianae In RD McCord and D Boaz(eds) Western Association of Vertebrate Paleontologists and SouthwestPaleontological Symposium Proceedings 2001 Mesa Southwest MuseumBulletin 8 33ndash40
Curtice BD Stadtman KL and Curtice L 1996 A reassessment ofUltrasauros macintoshi (Jensen 1985) Museum of Northern ArizonaBulletin 60 87ndash95
Dantas P Sanz JL Silva CM Ortega F Santos VF and Cachăo M1998 Lourinhasaurus n gen novo dinossaacuteurio sauroacutepode do Juraacutessicosuperior (Kimmeridgiano superiorTitoniano inferior) de Portugal Actasdo V Congresso de Geologia 84 91ndash94
Davis BM Cifelli RL and KielanminusJaworowska Z 2008 Earliest evidenceof Deltatheroida (Mammalia) from the Early Cretaceous of North Amerminusica In EJ Sargis and M Dagosto (eds) Mammalian Evolutionary Morminusphology A Tribute to Frederick S Szalay 3ndash24 Springer Dordrecht
DeCourten FL 1991 New data on Early Cretaceous dinosaurs from the LongWalk Quarry and tracksite Emery County Utah In TC Chidsey Jr(ed) Geology of EastminusCentral Utah Utah Geological Association Publiminuscation 19 311ndash325
DrsquoEmic M and Britt B 2008 Reevaluation of the phylogenetic affinitiesand age of a basal titanosauriform (Sauropoda Dinosauria) from theEarly Cretaceous Cloverly Formation of North America Journal ofVertebrate Paleontology 28 (Supplement to No 3) 68A
Dong Z Zhou S and Zhang Y 1983 Dinosaurs from the Jurassic ofSichuan [in Chinese] Palaeontologica Sinica New Series C 162 1ndash136
Eberth DA Britt BB Scheetz R Stadtman KL and Brinkman DB2006 Dalton Wellsmdashgeology and significance of debrisminusflowminushosteddinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaminusceous) of eastern Utah USA Paleogeography PaleoclimatologyPaleoecology 236 217ndash245 [CrossRef]
Farlow JO Coroian ID and Foster JR 2010 Giants on the landscapemodelling the abundance of megaherbivorous dinosaurs of the Morriminusson Formation (Late Jurassic western USA) Historical Biology 22403ndash429 [CrossRef]
Foster JR 2003 Paleoecological analysis of the vertebrate fauna of the
Morrison Formation (Upper Jurassic) Rocky Mountain region USANew Mexico Museum of Natural History and Science Bulletin 23 1ndash95
Fraas E 1908 Ostafrikanische Dinosaurier Palaeontographica 55 105ndash144Gallup MR 1989 Functional morphology of the hindfoot of the Texas
sauropod Pleurocoelus sp indet In JO Farlow (ed) Paleobiology of theDinosaurs Geological Society of America Special Paper 238 71ndash74
Gilmore CW 1921 The fauna of the Arundel Formation of Maryland USNational Museum Proceedings 59 581ndash594
Gilmore CW 1922 Discovery of a sauropod dinosaur from the Ojo Alminusamo Formation of New Mexico Smithsonian Miscellaneous Collecminustions 81 1ndash9
Gilmore CW 1936 Osteology of Apatosaurus with special reference tospecimens in the Carnegie Museum Memoirs of the Carnegie Museum11 175ndash300
Gilmore CW 1946 Reptilian fauna from the North Horn Formation of cenminustral Utah US Geological Survey Professional Paper 210minusC 29ndash52
Glut DF 1997 Dinosaurs the Encyclopedia 1076 pp McFarland amp Comminuspany Inc Jefferson North Carolina
Gomani EM Jacobs LL and Winkler DA 1999 Comparison of theAfrican titanosaurian Malawisaurus with a North America Early Creminustaceous sauropod In Y Tomida TH Rich and P VickersminusRich (eds)Proceedings of the Second Gondwanan Dinosaur Symposium TokyoNational Science Museum Monograph 15 223ndash233
Harris JD 2006 The significance of Suuwassea emiliae (DinosauriaSauropoda) for flagellicaudatan intrarelationships and evolution Jourminusnal of Systematic Palaeontology 4 185ndash198 [CrossRef]
Harris JD and Dodson P 2004 A new diplodocoid sauropod (Dinosauria)from the Morrison Formation (Upper Jurassic) of Montana USA ActaPalaeontologica Polonica 49 197ndash210
Hatcher JB 1901 Diplodocus (Marsh) its osteology taxonomy and probminusable habits with a restoration of the skeleton Memoirs of the CarnegieMuseum 1 1ndash63
Hatcher JB 1903a Osteology of Haplocanthosaurus with description of anew species and remarks on the probable habits of the Sauropoda andthe age and origin of the Atlantosaurus beds Memoirs of the CarnegieMuseum 2 1ndash72
Hatcher JB 1903b A new name for the dinosaur Haplocanthus HatcherProceedings of the Biological Society of Washington 16 100
He X Li K and Cai K 1988 The Middle Jurassic Dinosaur Fauna fromDashanpu Zigong Sichuan Vol IV Sauropod Dinosaurs (2) Omeiminussaurus tianfuensis 143 pp Sichuan Publishing House of Science andTechnology Chengdu
Huene F von 1929 Los Saurisquios y Ornitisquios del Cretaceo ArgentinaAnnales Museo de La Plata 3 (Serie 2a) 1ndash196
Ikejiri T Tidwell V and Trexler DL 2005 New adult specimens ofCamarasaurus lentus highlight ontogenetic variation within the speminuscies In V Tidwell and K Carpenter (eds) Thunder Lizards theSauropodomorph Dinosaurs 154ndash179 Indiana University Press Bloominusmington Indiana
Jacobs LL and Winkler DA 1998 Mammals archosaurs and the Earlyto Late Cretaceous transition in northminuscentral Texas In Y Tomida LJFlynn and LL Jacobs (eds) Advances in Vertebrate Paleontology andGeochronology 253ndash280 National Science Museum Tokyo
Jacobs LL Winkler DA and Murry PA 1991 On the age and correlaminustion of the Trinity mammals Early Cretaceous of Texas USA Newsletminusters on Stratigraphy 24 35ndash43
Jacobs LL Winkler DA Downs WR and Gomani EM 1993 Newmaterial of an Early Cretaceous titanosaurid sauropod dinosaur fromMalawi Palaeontology 36 523ndash534
Janensch W 1914 Uumlbersicht uumlber der Wirbeltierfauna der TendaguruminusSchichten nebst einer kurzen Charakterisierung der neu aufgefuhrtenArten von Sauropoden Archiv fur Biontologie 3 81ndash110
Janensch W 1950 Die Wirbelsaumlule von Brachiosaurus brancai Palaeontominusgraphica Supplement 7 (3) 27ndash93
Janensch W 1961 Die Gliedmaszen und Gliedmaszenguumlrtel der Saurominuspoden der TendaguruminusSchichten Palaeontographica Supplement 7(3) 177ndash235
96 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011
Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

Johnston C 1859 Note on odontography American Journal of Dental Sciminusence 9 337ndash343
Kingham RF 1962 Studies of the sauropod dinosaur Astrodon Leidy Prominusceedings of the Washington Junior Academy of Science 1 38ndash44
Kirkland JI 1987 Upper Jurassic and Cretaceous lungfish tooth platesfrom the Western Interior the last dipnoan faunas of North AmericaHunteria 2 1ndash16
Kirkland JI and Madsen SK 2007 The Lower Cretaceous Cedar Mounminustain Formation Eastern Utah the View up an Always InterestingLearning Curve Fieldtrip Guidebook 108 pp Geological Society ofAmerica Rocky Mountain Section Boulder Colorado
Kirkland JI Britt B Burge D Carpenter K Cifelli R DeCourten FEaton J Hasiotis S and Lawton T 1997 Lower to middle Cretaminusceous dinosaur faunas of the Central Colorado Plateau a key to underminusstanding 35 million years of tectonics sedimentology evolution andbiogeography Brigham Young University Geology Studies 42 69ndash103
Kirkland JI Cifelli RL Britt BB Burge DL DeCourten FLEaton JG and Parrish JM 1999 Distribution of vertebrate faunas inthe Cedar Mountain Formation eastminuscentral Utah In DD Gillette(ed) Vertebrate Paleontology in Utah Utah Geological Survey Misminuscellaneous Publication 99minus1 201ndash218
Kowallis BJ Christiansen EH Deino AL Peterson F Turner CEKunk MJ and Obradovich JD 1998 The age of the Morrison Forminusmation Modern Geology 22 235ndash260
Kranz PM 1998 Mostly dinosaurs a review of the vertebrates of the Potominusmac Group (Aptian Arundel Formation) USA In SG Lucas JIKirkland and JW Estep (eds) Lower and Middle Cretaceous Terresminustrial Ecosystems 235ndash238 New Mexico Museum of Natural Historyand Science Albuquerque
Langston W Jr 1974 Nonminusmammalian Commanchean tetrapods Geominusscience and Man 8 77ndash102
Lapparent AF and Zbyszewski G 1957 Les Dinosauriens du PortugalServices Geacuteologiques du Portugal Meacutemoire 2 (nouvelle seacuterie) 1ndash63
Larkin P 1910 The occurrence of a sauropod dinosaur in the Trinity Cretaminusceous of Oklahoma Journal of Geology 18 93ndash98 [CrossRef]
Leidy J 1865 Cretaceous reptiles of the United States Smithsonian Contriminusbution to Knowledge 192 1ndash135
Lovelace D Wahl WR and Hartman SA 2003 Evidence for costalpneumaticity in a diplodocid dinosaur (Supersaurus vivianae) Journalof Vertebrate Paleontology 23 (Supplement to No 3) 73A
Lovelace DM Hartman SA and Wahl WR 2008 Morphology of aspecimen of Supersaurus (Dinosauria Sauropoda) from the MorrisonFormation of Wyoming and a reminusevaluation of diplodocid phylogenyArquivos do Museu Nacional Rio de Janeiro 65 (4) 527ndash544
Lucas SG Spielmann JA Rinehart LF Heckert AB Herne MCHunt AP Foster JR and Sullivan RM 2006 Taxonomic status ofSeismosaurus hallorum a Late Jurassic sauropod dinosaur from NewMexico New Mexico Museum of Natural History and Science Bulletin36 149ndash162
Lull RS 1911 Systematic Paleontology Lower Cretaceous Vertebrata183ndash210 and pls 11ndash20 Maryland Geological Survey Baltimore
Mannion PD Upchurch P Carrano MT and Barrett PM 2010 Testingthe effect of the rock record on diversity a multidisciplinary approach toelucidating the generic richness of sauropodomorph dinosaurs throughtime Biological Reviews online Early View 27 pp [CrossRef]
Marsh OC 1877 Notice of new dinosaurian reptiles from the Jurassic Forminusmation American Journal of Science Series 3 14 514ndash516
Marsh OC 1878 Principal characters of American Jurassic dinosaurs Part IAmerican Journal of Science Series 3 16 411ndash416
Marsh OC 1888 Notice of a new genus of Sauropoda and other new dinominussaurs from the Potomac Formation American Journal of Science Series 335 89ndash94
Maxwell WD and Cifelli RL 2000 Last evidence of sauropod dinosaurs(Saurischia Sauropodomorpha) in the North American midminusCretaminusceous Brigham Young University Geology Studies 45 19ndash24
McIntosh JS 1990 Sauropoda In DB Weishampel P Dodson and HOsmoacutelska (eds) The Dinosauria 345ndash401 University of CaliforniaPress Berkeley
McIntosh JS Miller WE Stadtman KL and Gillette DD 1996 Theosteology of Camarasaurus lewisi (Jensen 1988) Brigham Young Uniminusversity Geology Studies 41 73ndash115
Naish D and Martill DM 2001 Saurischian dinosaurs 1 Sauropods InDM Martill and D Naish (eds) Dinosaurs of the Isle of Wight185ndash241 The Palaeontological Association London
Nydam RL and Cifelli RL 2002 Lizards from the Lower Cretaceous(AptianndashAlbian) Antlers and Cloverly Formations Journal of Verteminusbrate Paleontology 22 286ndash298 [CrossRef]
Osborn HF and Mook CC 1921 Camarasaurus Amphicoelias and othersauropods of Cope Memoirs of the American Museum of Natural Hisminustory new series 3 247ndash387
Ostrom JH 1970 Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area Wyoming and MontanaBulletin of the Peabody Museum of Natural History 35 1ndash234
Ouyang H and Ye Y 2002 The First Mamenchisaurian Skeleton withComplete Skull Mamenchisaurus youngi 111 pp Sichuan Science andTechnology Press Chengdu China
Owen R 1842 Report on British fossil reptiles Part II Reports of the BritishAssociation for the Advancement of Sciences 11 60ndash204
Owen R 1859 Monograph on the fossil Reptilia of the Wealden andPurbeck Formations Supplement No 2 Crocodilia (Streptospondylusampc) Palaeontographical Society Monograph 11 20ndash44
Phillips J 1871 Geology of Oxford and the Valley of the Thames 529 ppClarendon Press Oxford
Powell JE 1992 Osteologiacutea de Saltasaurus loricatus (SauropodandashTitanominussauridae) del Cretaacutecico Superior del Noroeste Argentino In JL Sanz andAD Buscalioni (eds) Los Dinosaurios y su Entorno Biotico 165ndash230Actas del Segundo Curso de Paleontologia en Cuenca Instituto Juan deValdeacutes Ayuntamiento de Cuenca
Ratkevich R 1998 The Cretaceous brachiosaurid dinosaur Sonorasaurusthompsoni gen et sp nov from Arizona Journal of the ArizonaminusNeminusvada Academy of Science 31 71ndash82
Rauhut OWM Remes K Fechner R Gladera G and Puerta P 2005Discovery of a shortminusnecked sauropod dinosaur from the Late Jurassicperiod of Patagonia Nature 435 670ndash672 [CrossRef]
Remes K 2006 Revision of the Tendaguru sauropod dinosaur Tornieriaafricana (Fraas) and its relevance for sauropod paleobiogeographyJournal of Vertebrate Paleontology 26 651ndash669 [CrossRef]
Remes K 2007 A second Gondwanan diplodocid dinosaur from the UpperJurassic Tendaguru beds of Tanzania East Africa Palaeontology 50653ndash667 [CrossRef]
Riggs ES 1903 Brachiosaurus altithorax the largest known dinosaurAmerican Journal of Science 15 299ndash306 [CrossRef]
Riggs ES 1904 Structure and relationships of opisthocoelian dinosaursPart II the Brachiosauridae Field Columbian Museum Geological Seminusries 2 229ndash247
Romer AS 1956 Osteology of the Reptiles 772 pp University of ChicagoPress Chicago
Rose PJ 2007 A new titanosauriform sauropod (Dinosauria Saurischia)from the Early Cretaceous of Central Texas and its phylogenetic relaminustionships Palaeontologia Electronica 10 (8A) 1ndash65
Salgado L and Bonaparte JF 1991 Un nuevo sauroacutepodo DicraeosauridaeAmargasaurus cazaui gen et sp nov de la Formacioacuten La AmargaNeocomiano de la Provincia del Neuqueacuten Argentina Ameghiniana 28333ndash346
Salgado L Coria RA and Calvo JO 1997 Evolution of titanosauridsauropods I Phylogenetic analysis based on the postcranial evidenceAmeghiniana 34 3ndash32
Salgado L Garrido A Cocca SE and Cocca JR 2004 Lower Cretaminusceous rebbachisaurid sauropods from Cerro Aguada Del Leon (LohanCura Formation) Neuquen Province Northwestern Patagonia Argenminustina Journal of Vertebrate Paleontology 24 903ndash912 [CrossRef]
Sames B and Madsen SK 2007 Ostracod evidence of a Berriasian Agefor the lower Yellow Cat Member of the Cedar Mountain FormationUtah Fiftyminusninth Annual Meeting of the Rocky Mountain Section of theGeological Society of America 7ndash9 May 2007 Abstracts with Prominusgrams 39 (5) 10
doi104202app20100073
TAYLOR ET ALmdashNEW SAUROPOD FROM THE EARLY CRETACEOUS OF UTAH 97
Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011

Sames B Cifelli RL and Schudack M 2010 The nonmarine LowerCretaceous of the North American Western Interior foreland basin newbiostratigraphic results from ostracod correlations and early mammalsand their implications for paleontology and geology of the basinndashanoverview Earth Science Reviews 101 207ndash224 [CrossRef]
Seeley HG 1870 On Ornithopsis a gigantic animal of the pterodactylekind from the Wealden Annals and Magazine of Natural History Seminusries 4 5 279ndash283
Seeley HG 1888 On the classification of the fossil animals commonlynamed Dinosauria Proceedings of the Royal Society of London 43165ndash171 [CrossRef]
Sereno PC 1998 A rationale for phylogenetic definitions with applicationto the higherminuslevel taxonomy of Dinosauria Neues Jahrbuch fuumlr Geominuslogie und Palaumlontologie Abhandlungen 210 41ndash83
Sereno PC Beck AL Dutheil DB Larsson HCE Lyon GHMoussa B Sadleir RW Sidor CA Varricchio DJ Wilson GPand Wilson JA 1999 Cretaceous sauropods and the uneven rate of skeleminustal evolution among dinosaurs Science 286 1342ndash1347 [CrossRef]
Sereno PC Wilson JA Witmer LM Whitlock JA Maga A Ide Oand Rowe TA 2007 Structural extremes in a Cretaceous dinosaurPLoS ONE 2 (11) e1230 [CrossRef]
Sternfeld R 1911 Zur Nomenklatur der Gattung Gigantosaurus Fraas Sitminuszungesberichte der Gesellschaft Naturforschender Freunde zu Berlin1911 398ndash398
Stokes WL 1952 Lower Cretaceous in Colorado Plateau Bulletin of theAmerican Association of Petroleum Geologists 36 1766ndash1776
Swofford DL 2002 PAUP Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer Associates Sunderland Mass
Taylor MP 2006 Dinosaur diversity analysed by clade age place andyear of description In PM Barrett (ed) Ninth International Sympominussium on Mesozoic Terrestrial Ecosystems and Biota Manchester UK134ndash138 Cambridge Publications Cambridge
Taylor MP 2009 A reminusevaluation of Brachiosaurus altithorax Riggs1903 (Dinosauria Sauropoda) and its generic separation from Girafminusfatitan brancai (Janensch 1914) Journal of Vertebrae Palaeontology29 (3) 787ndash806 [CrossRef]
Taylor MP and Naish D 2005 The phylogenetic taxonomy of Diplominusdocoidea (Dinosauria Sauropoda) PaleoBios 25 1ndash7
Taylor MP and Naish D 2007 An unusual new neosauropod dinosaurfrom the Lower Cretaceous Hastings Beds Group of East Sussex Engminusland Palaeontology 50 1547ndash1564 [CrossRef]
Tidwell V and Carpenter K 2003 Braincase of an Early Cretaceoustitanosauriform sauropod from Texas Journal of Vertebrate Paleontolminusogy 23 176ndash180 [CrossRef]
Tidwell V and Carpenter K 2007 First description of cervical vertebraefor an Early Cretaceous titanosaur from North America Journal of Verminustebrate Paleontology 27 (Supplement to No 3) 158A
Tidwell V and Wilhite DR 2005 Ontogenetic variation and isometricgrowth in the forelimb of the Early Cretaceous sauropod VenenosaurusIn V Tidwell and K Carpenter (eds) Thunder Lizards the Saurominuspodomorph Dinosaurs 187ndash198 Indiana University Press Bloomingminuston Indiana
Tidwell V Carpenter K and Brooks W 1999 New sauropod from theLower Cretaceous of Utah USA Oryctos 2 21ndash37
Tidwell V Carpenter K and Meyer S 2001 New Titanosauriform (Saurominuspoda) from the Poison Strip Member of the Cedar Mountain Formation(Lower Cretaceous) Utah In DH Tanke and K Carpenter (eds) Mesominuszoic Vertebrate Life New Research Inspired by the Paleontology of PhilipJ Currie 139ndash165 Indiana University Press Bloomington
Tschudy RH Tschudy BD and Craig LC 1984 Palynological evaluaminustion of Cedar Mountain and Burro Canyon formations Colorado Plaminusteau United States Geological Survey Professional Paper 1281 1ndash21
Upchurch P and Mannion PD 2009 The first diplodocid from Asia andits implications for the evolutionary history of sauropod dinosaursPalaeontology 52 1195ndash1207 [CrossRef]
Upchurch P Barrett PM and Dodson P 2004a Sauropoda In DBWeishampel P Dodson and H Osmoacutelska (eds) The Dinosauria 2ndedition 259ndash322 University of California Press Berkeley
Upchurch P Tomida Y and Barrett PM 2004b A new specimen ofApatosaurus ajax (Sauropoda Diplodocidae) from the Morrison Forminusmation (Upper Jurassic) of Wyoming USA National Science Muminusseum Monographs 26 1ndash110
Vietti L and Hartman S 2004 A new diplodocid braincase (DinosauriaSauropoda) from the Morrison Formation of northminuswestern WyomingJournal of Vertebrate Paleontology 24 (Supplement to No 3) 68A
Wedel MJ 2000 New material of sauropod dinosaurs from the CloverlyFormation Journal of Vertebrate Paleontology 20 (Supplement to No3) 77A
Wedel MJ 2003a Vertebral pneumaticity air sacs and the physiology ofsauropod dinosaurs Paleobiology 29 243ndash255 [CrossRef]
Wedel MJ 2003b The evolution of vertebral pneumaticity in sauropod diminusnosaurs Journal of Vertebrate Paleontology 23 344ndash357 [CrossRef]
Wedel MJ 2005 Postcranial skeletal pneumaticity in sauropods and its imminusplications for mass estimates In JA Wilson and K Curry Rogers(eds) The Sauropods Evolution and Paleobiology 201ndash228 Univerminussity of California Press Berkeley
Wedel MJ and Cifelli RL 2005 Sauroposeidon Oklahomarsquos native giminusant Oklahoma Geology Notes 65 40ndash57
Wedel MJ Cifelli RL and Sanders RK 2000a Osteology paleominusbiology and relationships of the sauropod dinosaur SauroposeidonActa Palaeontologica Polonica 45 343ndash388
Wedel MJ Cifelli RL and Sanders RK 2000b Sauroposeidon proteles anew sauropod from the Early Cretaceous of Oklahoma Journal of Verteminusbrate Paleontology 20 109ndash114 [CrossRef]
Whitlock JA DrsquoEmic MD and Wilson JA 2011 Cretaceous diplominusdocoids in Asia Reminusevaluating the phylogenetic affinities of a fragminusmentary specimen Palaeontology online Early View [CrossRef]
Wilhite R 1999 Ontogenetic Variation in the Appendicular Skeleton of theGenus Camarasaurus 28 pp Unpublished MSc thesis Brigham YoungUniversity Utah
Wilhite R 2003 Biomechanical Reconstruction of the Appendicular Skeletonin Three North American Jurassic Sauropods 225 pp + vi UnpublishedPhD thesis Department of Geology and Geophysics Louisiana StateUniversity httpetdlsuedudocsavailableetdminus0408103minus003549
Wilson JA 2002 Sauropod dinosaur phylogeny critique and cladisticanalysis Zoological Journal of the Linnean Society 136 217ndash276[CrossRef]
Wilson JA and Sereno PC 1998 Early evolution and higherminuslevel phyminuslogeny of sauropod dinosaurs Society of Vertebrate PaleontologyMemoir 5 1ndash68 [CrossRef]
Wilson JA and Upchurch P 2003 A revision of Titanosaurus Lydekker(Dinosauria ndash Sauropoda) the first dinosaur genus with a lsquoGondwananrsquodistribution Journal of Systematic Palaeontology 1 125ndash160 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1990 Early Cretaceous(Comanchean) vertebrates of central Texas Journal of Vertebrate Paleminusontology 10 95ndash116 [CrossRef]
Winkler DA Murry PA and Jacobs LL 1997b A new species ofTenontosaurus (Dinosauria Ornithopoda) from the Early Cretaceous ofTexas Journal of Vertebrate Paleontology 17 330ndash348 [CrossRef]
Winkler DA Jacobs LL and Murry PA 1997a Jones Ranch an EarlyCretaceous sauropod boneminusbed in Texas Journal of Vertebrate Paleonminustology 17 (Supplement to No 3) 85A
Witzke BJ and Ludvigson GA 1994 The Dakota Formation in Iowa andits type area In GW Shurr GA Ludvigsonand and RH Hammond(eds) Perspectives on the eastern margin of the Cretaceous Western Inminusterior Basin Geological Society of America Special Paper 287 43ndash78
Yates AM 2006 Solving a dinosaurian puzzle the identity of Aliwalia rexGalton Historical Biology 19 93ndash123 [CrossRef]
Young CminusC 1939 On the new Sauropoda with notes on other fragmentaryreptiles from Szechuan Bulletin of the Geological Society of China 19279ndash315 [CrossRef]
Young CminusC and Zhao X 1972 Description of the type material ofMamenchisaurus hochuanensis [in Chinese] Institute of VertebratePaleontology and Paleoanthropology Monograph Series I 8 1ndash30
98 ACTA PALAEONTOLOGICA POLONICA 56 (1) 2011