a novel human recombinant antibody fragment capable of neutralizing mexican scorpion toxins
TRANSCRIPT

ilable at ScienceDirect
Toxicon xxx (2013) 1–7
Contents lists ava
Toxicon
journal homepage: www.elsevier .com/locate/ toxicon
A novel human recombinant antibody fragment capable ofneutralizing Mexican scorpion toxins
Lidia Riaño-Umbarila, Timoteo Olamendi-Portugal, Citlalli Morelos-Juárez,Georgina B. Gurrola, Lourival D. Possani, Baltazar Becerril*
Departamento de Medicina Molecular y Bioprocesos, Instituto de Biotecnología, UNAM, Apartado Postal 510-3, Cuernavaca 62250,Morelos, Mexico
a r t i c l e i n f o
Article history:Received 18 March 2013Received in revised form 8 August 2013Accepted 12 September 2013Available online xxxx
Keywords:Human scFvIn vitro maturationScorpion toxin
Abbreviations: aa, amino acid(s); CDR, complemregion (s); C. limpidus, Centruroides limpidus limpidustoxins 1 and 2 from Centruroides limpidus limpiduCentruroides noxius Hoffman; Cn2 and Cn3, toxinstruroides noxius venom; C. suffusus, Centruroides sutoxin 2 of Centruroides suffusus suffusus; FW, framechain antibody fragment; VH, variable domain of hable domain of light chain.* Corresponding author. Present address: Av. U
Colonia Chamilpa, Cuernavaca 62210, Mexico. Tel.:E-mail address: [email protected] (B. Becerr
0041-0101/$ – see front matter � 2013 Elsevier Ltdhttp://dx.doi.org/10.1016/j.toxicon.2013.09.016
Please cite this article in press as: Riañoneutralizing Mexican scorpion toxins, Toxi
a b s t r a c t
Using phage display and directed evolution, our group has progressed in the constructionof a second family of human single chain variable fragments (scFv) which bind to scorpiontoxins dangerous to mammals. It was observed that scFv C1 only bound initially to toxinCn2, which constitutes 6.8% of whole venom from the scorpion Centruroides noxius Hoff-man. Only a few amino acid changes were necessary to extend its recognition to othersimilar toxins and without affecting the recognition for its primary antigen (Cn2 toxin).One variant of scFv C1 (scFv 202F) was selected after two cycles of directed evolutionagainst Cll1 toxin, the second major toxic component from the venom of the Mexicanscorpion Centruroides limpidus limpidus Karsh (0.5% of the whole venom). scFv 202F is alsocapable of recognizing Cn2 toxin. Despite not having the highest affinity for toxins Cll1(KD ¼ 25.1 � 10�9 M) or Cn2 (KD ¼ 8.1 � 10�9 M), this antibody fragment neutralized oneLD50 of each one of these toxins. Additionally, scFv 202F moderately recognized Cll2 toxinwhich constitutes 1.5% of the venom from C. limpidus. Based on our previous experience,we consider that these results are promising; consequently, we continue working ongenerating new optimized variants from scFv C1 that could be part of a recombinantscorpion anti-venom from human origin, that might reach the market in the near future.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
In order to improve the quality of anti-venoms used intherapy, several procedures have been developed to ensurethe effectiveness and safety of anti-venoms from non-
entarity determiningKarsh; Cll1 and Cll2,s venom; C. noxius,2 and 3 from Cen-
ffusus suffusus; Css2,work(s); scFv, singleeavy chain; VL, vari-
niversidad No. 2001,þ52 7773 291669.il).
. All rights reserved.
-Umbarila, L., et al., Acon (2013), http://dx.do
human origin. Some examples are: the optimization ofthe immune response of animals challenged with venoms;the purification of the horse immunoglobulins (IgGs) andtheir enzymatic digestion with pepsin to remove the crys-tallizable fragment (Fc) which is immunogenic for humans(reviewed in Espino-Solis et al. (2009)). Due to significantadvances in antibody engineering in recent decades, it hasbeen possible the generation of human antibody fragmentscapable of toxin neutralization, with the advantage of beingless immunogenic than whole immunoglobulins (Holligerand Hudson, 2005).
A scFv is constituted by the variable domains of a lightchain and a heavy chain linked by a peptide linker, usu-ally 15 amino acids (aa) long (Bird et al., 1988; Hustonet al., 1991). This format has a similar functional activityas compared to that of a whole antibody. Due to its
novel human recombinant antibody fragment capable ofi.org/10.1016/j.toxicon.2013.09.016

L. Riaño-Umbarila et al. / Toxicon xxx (2013) 1–72
comparatively small size (30 kDa), a scFv would have agreater spreading in any organism, compared with an IgGor a F (ab0)2, which would be an important advantage inacute cases of poisoning by scorpions. It is worth notingthat the genetic information of scFvs is relatively easy tomanipulate. By means of phage display, directed evolu-tion and rational panning procedures, scFvs with opti-mized properties and level of expression can be isolatedagainst specific targets.
Scorpion venom toxicity for mammals is due to thepresence of toxins that affect the activity of the sodiumchannel (reviewed in Rodriguez de la Vega and Possani(2005)). Each venom contains one or several main toxiccomponents, in terms of its toxicity and abundance,meaning that neutralization of them is sufficient to achievethe effect of inhibiting whole venom. Three out of the 8most dangerous Mexican scorpions are: Centruroides suf-fusus suffususwhich is distributed in the Durango state andits major toxic component is Css2 toxin (2.8% from wholevenom); Centruroides noxius is distributed in Nayarit stateand its major toxic component is Cn2 (6.8% from de wholevenom); and Centruroides limpidus limpidus distributedmainly in the states of Morelos and Guerrero. This lastvenom contains two main components Cll1 and Cll2 withabundances of 0.5% and 1.5% respectively.
Total neutralization of the venom by means of specificantibodies depends on several factors such as complexity ofthe venom, cross reactivity and affinity. There are cases inwhich a single antibody is able of neutralizing up to twodifferent scorpion venoms (Riano-Umbarila et al., 2011). Inother reported cases two antibodies are required toneutralize one scorpion venom (Clot-Faybesse et al., 1999;Hmila et al., 2010). The goal of our group is to generate aset of human antibody fragments like those already devel-oped against the venoms of C. suffusus and C. noxius (Riano-Umbarila et al., 2011, 2005) against otherMexican scorpions.The whole set of scFvs should be equivalent to the com-mercial polyclonal and polyvalent anti-venom. The effec-tivity of each scFv is based on their cross reactivity level,which is favored by the high aa sequence identity of thesetoxins, and the conservation of the disulfide bridges (Fig. 1).
In this contribution, the parental scFv C1 (which rec-ognizes Cn2 toxin) was subjected to two cycles of directedevolution against Cll1 and Cll2 toxins. We obtained severalscFv C1 variants which showed better recognition to Cll1and Cll2. From this process, scFv 202F was isolated. Thisvariant was able of neutralizing toxins Cll1, and surpris-ingly, Cn2 toxin as well. The maturation process (throughmutagenic PCR and affinity selection) once again was suc-cessfully enough to increase the recognition capacity andthe improvement of the affinity in a second scFv family.
Fig. 1. Amino acid sequence alignment of homologous toxins. Cll1 and Cll2: toxinsvenom. Css2: toxin 2 from C. suffusus. Dots indicate identical residues. All sequenc
Please cite this article in press as: Riaño-Umbarila, L., et al., Aneutralizing Mexican scorpion toxins, Toxicon (2013), http://dx.do
2. Materials and methods
2.1. Toxins
Toxins were purified from scorpion venoms using pre-viously described methodologies: Cll1 and Cll2 were ob-tained from venom of C. limpidus (Dehesa-Davila et al.,1996; Ramirez et al., 1994); Cn2 and Cn3 toxins from C.noxius Hoffmann (Zamudio et al., 1992) and Css2 from C.suffusus (Hernandez-Salgado et al., 2009; Schiavon et al.,2006).
2.2. Directed evolution
2.2.1. Mutant libraries constructionEach maturation process included the construction of a
mutant library by random mutagenesis and three or fourrounds of bio-panning. Error-prone PCR reactions wereperformed under conditions that permit different mutationrates along the coding sequence of scFv C1 (Cadwell andJoyce, 1992; Leung et al., 1989). The primers used werethe following: Forward (50-ATACCTATTGCCTACGGC-30) andMyc (50-TCAGATCCTCTTCTGAGATG-30). The PCR productswere gel-purified, digested with restriction enzymes SfiIandNotI and ligated into pSyn2 phagemid predigestedwiththe same enzymes. The ligation product was inserted intoEscherichia coli strain TG1 electro-competent cells. The li-brary was characterized and evaluated by bio-panningagainst Cll1 and Cll2 toxins as described previously(Marks et al., 1991). The DNA segments of four clones iso-lated from the first cycle of directed evolutionwere used astemplates for a new step of random mutagenesis(Meyerhans et al., 1990). The construction of the secondlibrary was performed using the conditions alreadydescribed.
2.2.2. Bio-panning against toxinsFor the first cycle of directed evolution three bio-
panning rounds were performed. For the first bio-panningstep, 2 mg mL�1 of Cll1 or Cll2 toxins were used to coat eachimmuno-tube. BSA (1%) was used as blocking agent and1�1013 mL�1 phage-antibodies were poured into immune-tubes. The unspecific or weak binding phages were elimi-nated by several washes with phosphate-buffered saline(PBS)-Tween 20 (Marks et al., 1991). Recovering of specificphages was made by adding TG1 cells to the washedimmuno-tubes (Riano-Umbarila et al., 2005). Each phagesample was subjected to independent rounds of bio-panning. Toxin concentration was kept in the followingrounds, changing the blocking agents which were gelatin(second round) and milk (third round). Several clones from
1 and 2 from C. limpidus venom; Cn2 and Cn3: toxins 2 and 3 from C. noxiuses contain 66 aa residues and the same array of cysteine residues.
novel human recombinant antibody fragment capable ofi.org/10.1016/j.toxicon.2013.09.016

Fig. 2. Maturation process of scFv C1 against toxins Cll1 (in black) and Cll2(in gray). Antibody variants isolated from two cycles of directed evolution.Final amino acid changes in each variant are shown indicating their struc-tural location.
L. Riaño-Umbarila et al. / Toxicon xxx (2013) 1–7 3
the last round were evaluated for recognition to Cll1 or Cll2by phage-ELISA. The best clones were evaluated andsequenced.
A second cycle of directed evolution and affinity-selection was carried out under similar conditions. Thesecond library was subjected to four bio-panning roundsagainst Cll1 and Cll2 toxins. Toxin concentrations were1 mg mL�1 for the first and second rounds, 0.5 mg mL�1 and0.25 mg mL�1 for the third and fourth rounds respectively.Selected clones from the last round were evaluated assoluble protein in ELISA. Antibodies that showed thehighest signals were characterized.
2.3. ELISAS
Phage-Antibody and soluble protein ELISA assays wereperformed as described (Riano-Umbarila et al., 2005).
2.4. Antibody expression and purification
scFv 202F-encoding DNA was cloned into pSyn1 vectorfor protein expression in E. coli strain TG1 cells. Proteinexpression and purification were carried out as describedpreviously (Riano-Umbarila et al., 2005). The final purifi-cation procedure was performed by gel-filtration chroma-tography on a Superdex� 75 column (Pharmacia BiotechAB, Uppsala, Sweden). The buffer used was 1� PBS and theflow rate was 0.6 mL min�1. The protein concentration wasdetermined spectrophotometrically at l ¼ 280 nm.
2.5. Surface plasmon resonance measurements
The kinetic constants of the scFv 202F binding toimmobilized toxins Cll1 or Cll2 were determined using aBiacore biosensor system (Biacore X, Uppsala, Sweden).One hundred and seventy ng of Cll1 or Cn2 were dissolvedin 100 mL of 10 mM 2-(N-morpholino) ethanesulfonic acid(pH6). Twenty microliters of toxin solution was bound tocell 2 of a CM5 sensor chip using the Amine Coupling Kit(Biacore) at a flow rate of 5 mL min�1. Approximately 200resonance units (RU) were coupled. Cell 1 (nothing bound)was used as a control. A protein solution of scFv 202F wasserially diluted in HBS-EP buffer (Biacore); samples of100 mL were injected over each chip (Cll1 or Cn2 coupled) ata flow rate 50 mL min�1. Biosensor measurements wereperformed at 25 �C. scFv 202F protein concentrationsranged from 1 nM to 250 nM for both assays. The delayphase lasted 800 s. The chip surface was regenerated with10 mM HCl. The kinetic constants were determined usingthe corresponding sensorgrams, which were corrected bysubtracting the values from both the reference flow cell andthe blank buffer injection. The Langmuir (1:1) model fromBIA-evaluation software version 3.1 was used for kineticconstants determination.
2.6. Neutralization tests
In order to evaluate the neutralization ability of the scFv202F, one LD50 of the toxin Cll1 (1.7 mg of Cll1 toxin per 20 gof mouse) or one LD50 of the toxin Cn2 (0.25 mg of Cn2 toxinper 20 g of mouse) were used. Groups of 6 or10 female CD1
Please cite this article in press as: Riaño-Umbarila, L., et al., Aneutralizing Mexican scorpion toxins, Toxicon (2013), http://dx.do
mice weighting w20 g were injected by intra-peritonealvia, with 1 LD50 of Cll1 or 2 LD50 of Cn2 (control groups).In the experimental groups, the neutralization activity ofscFv 202F was tested by the administration of a mix of 202Fand each toxin at a molar ratio of 1:10 of toxin:antibody.The mixtures were pre-incubated for 30 min at roomtemperature (w25 �C) prior to injection. Intoxicationsymptoms were observed over a period of 24 h.
3. Results
scFv C1 parental antibody fragment, was previouslyisolated by means of phage display from a non-immunelibrary of antibody fragments using Cn2 toxin from thevenom of C. noxius as target (Riano-Umbarila et al., 2005).The initial affinity of this antibody fragment for toxin Cn2was low and consequently unable of neutralizing its toxiceffect. Evaluations of cross-reactivity of scFv C1 by phageand soluble protein-ELISA showed that C1 did not recog-nize Cll1, Cll2 toxins from C. limpidus scorpion venom(Riano-Umbarila et al., 2005) or blocking agents (BSA,gelatin or milk). Previously to this work we performed adirected evolution cycle against Cn2 toxin, although somescFv C1 variants increased the recognition to toxin Cn2(evaluated by phage-ELISA); the most important observa-tion was that these scFvs also recognized Cll1 and Cll2toxins. This cross-reactivity indicates the plasticity of theparental scFv C1 for recognition of toxins with similarsequence. Based on this result, we decided to direct therecognition of scFv C1 towards toxins Cll1 and Cll2.
3.1. scFv C1 directed evolution
For the first cycle, a mutagenic library of 1�107 variantsfrom parental scFv C1 was constructed with a mutagenicrate of 0.36%. This library was displayed on phages andscreened against toxins Cll1 and Cll2. Four scFvs were iso-lated (2 against Cll1 and 2 against Cll2) (Fig. 2) which
novel human recombinant antibody fragment capable ofi.org/10.1016/j.toxicon.2013.09.016

L. Riaño-Umbarila et al. / Toxicon xxx (2013) 1–74
recognized toxin Cn2, as well as toxins Cll1 and Cll2. DNAsequencing allowed to determine that the selected variantsagainst toxin Cll1, have in common the change D54G, whiletwo variants selected against Cll2 toxin, have the changeM105L. Since these changes extended scFvs recognition foradditional toxins, we decided to perform a second round ofdirected evolution. The four selected variants were used astemplates in mutagenic PCR reactions, in order to allow therecombination among the selected changes. The size of thelibrary was 3 � 107 variants (mutagenic rate of 0.4%) andscreened against toxins Cll1 and Cll2 (four rounds). Fromthe last round of bio-panning, two variants against Cll1 andone against Cll2 were selected (Fig. 2). These scFvs showedan important increase of recognition for the antigens.
3.2. Characterization of isolated scFvs
3.2.1. Cross-reactivity assayThe selected variants from the first cycle (Fig. 2) by
ELISA of phage-antibody, showed the first traces of recog-nition against Cll1 and Cll2 toxins (parental scFv scFv do notrecognize them). Additionally, all clones increased theirrecognition to Cn2 compared to the parental scFv C1. Thevariants from the second cycle were evaluated in an ELISAtest using soluble proteins. The clone scFv 305E generatedagainst Cll2 showed no significant improvements in itsrecognition to Cll2 toxin, while scFv 202F selected againstCll1 toxin showed a higher level of recognition than scFv206B (also selected against Cll1 toxin). A culture of E. coliTG1containing the scFv 202F-pSyn2was induced overnightand the supernatant was used to test cross-reactivity withthe toxins Cn2, Cn3, Cll1, Cll2 and Css2 (Fig. 3). The resultsrevealed that this variant has a high recognition to Cn2 andCll1, and cross-react with other similar toxins.
3.2.2. Sequences analysesWith the purpose of knowing which mutations were
responsible of this behavior, the selected variants weresequenced. The mutations D54G and M105L (located atCDRs 2 and 3 of the heavy chain (VH)) were present in allvariants, which suggests a recombination of the two orig-inally predominant changes (Fig. 2) (Meyerhans et al.,1990). The best variant, scFv 202F showed an additionalchange (S56G CDR2 VH) (Fig. 4), that is presumably
Fig. 3. Cross-reactivity of scFv 202F against closely related Mexican scorpiontoxins. Toxins were used at a concentration of 3 mg mL�1.
Please cite this article in press as: Riaño-Umbarila, L., et al., Aneutralizing Mexican scorpion toxins, Toxicon (2013), http://dx.do
responsible of the higher recognition for Cll1 and Cn2toxins (Fig. 3).
3.3. Functional characterization of scFv 202F
3.3.1. ExpressionGiven the recognition ability of scFv variant 202F, it was
decided to subclone the DNA sequence that encodes thatantibody into the expression vector pSyn1. The proteinexpressed in E. coli strain TG1 cells was purified asdescribed in materials and methods. The final purificationof the protein by molecular exclusion chromatographyshowed a single peak, recovered after 21 min with abehavior similar to previously reported single chain anti-bodies (Riano-Umbarila et al., 2005; Rodriguez-Rodriguezet al., 2012). The yield was approximately 1.5 mg L�1 cul-ture, similar to previous results of our group (Riano-Umbarila et al., 2011).
3.3.2. Determinations of affinity by BiacoreThe level of recognition by scFv 202F of toxins Cn2 and
Cll1 was evaluated in a sensor of molecular interactions inreal time (Biacore), where the kinetic constants weredetermined (Table 1). Each toxin was coupled to a separatechip, using the same protocol in order to obtain a similarlevel of binding to the chip (w200 RU). This approachhelped to compare the sensorgrams (Fig. 5), for evaluatingthe level of recognition by scFv 202F to the two toxins (Cll1and Cn2). The level of recognition for each toxin by scFv202F was different. For example, at a concentration of250 nM the maximum response for Cll1 was 200 RU, whilefor Cn2, it was 370 RU. In the dissociation phase, the signaldecay response was greater for Cll1 toxin, while for Cn2, itwas lower at all tested concentrations of scFv (Fig. 5). Thisbehavior is reflected in the values of the kinetic constantsKon, Koff and KD (Table 1). The affinity of scFv 202F is higherfor toxin Cn2 (KD ¼ 8.1 � 10�9 M) as compared to the onefor Cll1 (KD ¼ 2.5 � 10�8 M). scFv 202F affinity values aresignificantly better compared to parental scFv C1, which didnot recognize Cll1 toxin and its affinity (KD) for Cn2 toxinwas only 5.4 � 10�7 M (Table 1). Considering the affinityincrement of scFv 202F to those toxins, we performed someneutralization assays in mice.
3.3.3. Neutralization assaysThe neutralizing capacity of scFv 202F was evaluated
with toxins Cll1 and Cn2 (Table 2). The results were unex-pected since the antibody protectedmice challengedwith 1LD50 of toxins Cll1 and Cn2 at a molar ratio 1:10 (tox-in:antibody). The neutralization level was surprising,because the affinity values were lower as compared toother scFvs which neutralize Cn2 toxin (Juarez-Gonzalezet al., 2005; Riano-Umbarila et al., 2005).
4. Discussion
The scFv C1 is composed by a VH domain from VH3
family, and a VL domain corresponding to the family Vl1(Riano-Umbarila et al., 2005). Due to the thermodynamiccharacteristics of these two variable domain families, theyare classified among the most stable domains (Ewert et al.,
novel human recombinant antibody fragment capable ofi.org/10.1016/j.toxicon.2013.09.016

Fig. 4. Amino acid sequences of scFvs C1 and 202F. scFv C1 is the parental antibody fragment of this family. The linker between the VH and VL domains isindicated in italic letters. The mutations of scFv 202F are indicated in boldface in CDRs 2 and 3 of VH domain. Dots in scFv 202F mean conserved residues ascompare to scFv C1.
L. Riaño-Umbarila et al. / Toxicon xxx (2013) 1–7 5
2003). In particular the combination of domains VH3-Vl1appears to be very robust (expression and stability) (Ewertet al., 2003), therefore the use of this scFv scaffold astemplate for directed evolution, could guarantee the se-lection of optimized toxin binders. The strategy of randommutagenesis for affinity maturation has been the mostsuccessful with respect to other maturation processespreviously tested in our group.
Two cycles of directed evolution and phage displayallowed the selection of a series of changes in scFv C1variants which improved the recognition toward Cll1 and
Table 1Affinity Constants. Biosensor assays were performed at 25 �C with a flow rate of
scFv Toxin Kon (M�1 s�1) � 104 SE Kon
C1 Cll1 NDCn2 2.0 230.0
202F Cll1 7.6 7.3Cn2 11.3 477.0
Please cite this article in press as: Riaño-Umbarila, L., et al., Aneutralizing Mexican scorpion toxins, Toxicon (2013), http://dx.do
Cll2 toxins. These changes are located at the VH domain, inCDR2 and CDR3, which allowed their binding to severalscorpion toxins. They also maintained the recognition totoxin Cn2, with a remarkable affinity improvement whencompared to toxin Cll1. This type of results is the secondone reported by our group, which we consider very strik-ing, since we could have expected that during an in vitromaturation process, the resulting variants from consecutivedirected evolution cycles, might have diminished or lost therecognition to their primary target (Dubreuil et al., 2005;Miyazaki et al., 1999). We consider that it was not the
50 mL min�1. SE ¼ standard deviation. ND ¼ not determined.
Koff (s�1) � 10�4 SE Koff � 10�6 KD (M) � 10�9
ND ND140.0 0.69 540.019.0 0.18 25.19.2 3.24 8.1
novel human recombinant antibody fragment capable ofi.org/10.1016/j.toxicon.2013.09.016
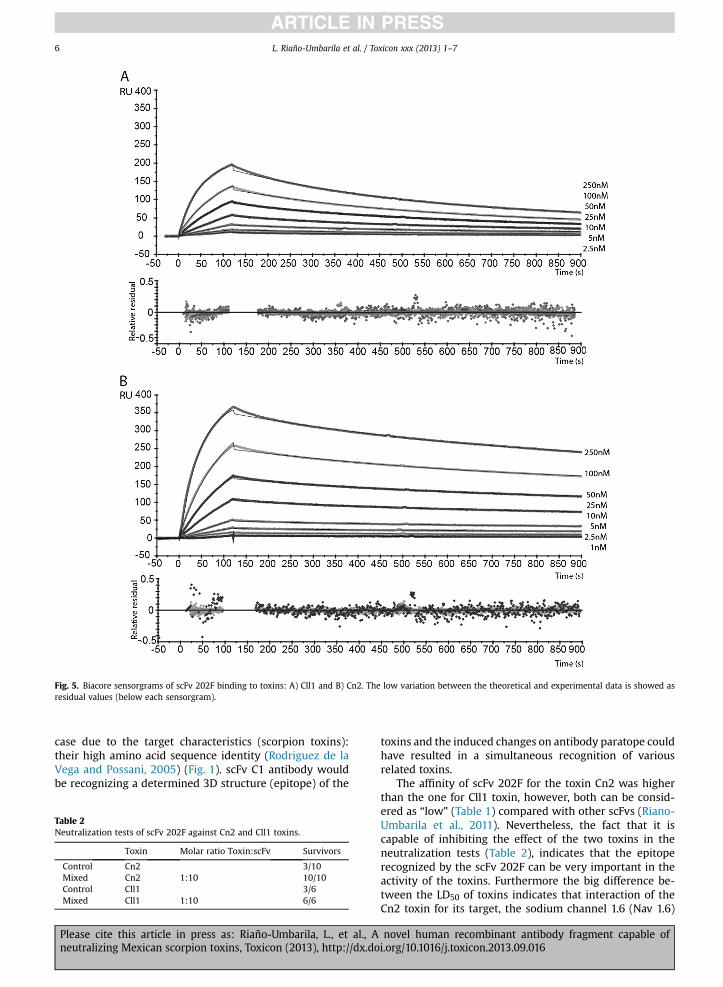
Fig. 5. Biacore sensorgrams of scFv 202F binding to toxins: A) Cll1 and B) Cn2. The low variation between the theoretical and experimental data is showed asresidual values (below each sensorgram).
L. Riaño-Umbarila et al. / Toxicon xxx (2013) 1–76
case due to the target characteristics (scorpion toxins):their high amino acid sequence identity (Rodriguez de laVega and Possani, 2005) (Fig. 1). scFv C1 antibody wouldbe recognizing a determined 3D structure (epitope) of the
Table 2Neutralization tests of scFv 202F against Cn2 and Cll1 toxins.
Toxin Molar ratio Toxin:scFv Survivors
Control Cn2 3/10Mixed Cn2 1:10 10/10Control Cll1 3/6Mixed Cll1 1:10 6/6
Please cite this article in press as: Riaño-Umbarila, L., et al., Aneutralizing Mexican scorpion toxins, Toxicon (2013), http://dx.do
toxins and the induced changes on antibody paratope couldhave resulted in a simultaneous recognition of variousrelated toxins.
The affinity of scFv 202F for the toxin Cn2 was higherthan the one for Cll1 toxin, however, both can be consid-ered as “low” (Table 1) compared with other scFvs (Riano-Umbarila et al., 2011). Nevertheless, the fact that it iscapable of inhibiting the effect of the two toxins in theneutralization tests (Table 2), indicates that the epitoperecognized by the scFv 202F can be very important in theactivity of the toxins. Furthermore the big difference be-tween the LD50 of toxins indicates that interaction of theCn2 toxin for its target, the sodium channel 1.6 (Nav 1.6)
novel human recombinant antibody fragment capable ofi.org/10.1016/j.toxicon.2013.09.016

L. Riaño-Umbarila et al. / Toxicon xxx (2013) 1–7 7
(Schiavon et al., 2006) is stronger than the one of toxin Cll1on all different target channels (Schiavon et al., 2012).Given the recognition plasticity of scFv C1 for its targettoxins, only two cycles of directed evolution, allowed theisolation of a neutralizing antibody. We are intending toimprove the affinity properties and expression levels ofscFv 202F by means of new cycles of directed evolution.This process will allow to identify key aa residues impor-tant for increasing affinity and stability. These residues canbe segregated and/or combined by site-directed mutagen-esis. We also expect to obtain entirely neutralizing variantsagainst Cll2 toxin, as well as to other related scorpiontoxins.
Ethical statement
The authors of this manuscript declare that they all havefollowed the ethical requirements for this communication.The experiments in which animals alive were used andreported in this manuscript were approved by EthicalCommittee of our Institute of Biotechnology-UNAM.
Acknowledgments
This work was supported by grants P-254 from theInstituto Bioclon S.A. de C.V. (Mexico) and CB 2010-155099-Q from CONACyTand DGAPAIN-201913 to B.B. We thank Dr.Humberto Flores for critical reading of the manuscript.Sergio Gonzalez Trujillo, Marcela Ramírez Yarza y MVElizabeth Mata is recognized for assistance with the animalprovision and care. Finally, we thank the unit of oligonu-cleotide synthesis from the Instituto de Biotecnología,UNAM (M.Sc. Santiago Becerra, M.Sc. Eugenio Bustos andDr. Paúl Gaytan) and the following colleagues: Mario Trejo,Martín Patiño and Cipriano Balderas for technical assis-tance in various aspects of this work.
Conflict of interest
The authors of this manuscript declare that there is noconflict of interest.
References
Bird, R.E., Hardman, K.D., Jacobson, J.W., Johnson, S., Kaufman, B.M.,Lee, S.M., Lee, T., Pope, S.H., Riordan, G.S., Whitlow, M., 1988. Single-chain antigen-binding proteins. Science 242, 423–426.
Cadwell, R.C., Joyce, G.F., 1992. Randomization of genes by PCR muta-genesis. PCR Methods Appl. 2, 28–33.
Clot-Faybesse, O., Juin, M., Rochat, H., Devaux, C., 1999. Monoclonal an-tibodies against the Androctonus australis hector scorpion neurotoxinI: characterisation and use for venom neutralisation. FEBS Lett. 458,313–318.
Dehesa-Davila, M., Ramirez, A.N., Zamudio, F.Z., Gurrola-Briones, G.,Lievano, A., Darszon, A., Possani, L.D., 1996. Structural and functionalcomparison of toxins from the venom of the scorpions Centruroidesinfamatus infamatus, Centruroides limpidus limpidus and Centruroidesnoxius. Comp. Biochem. Physiol. Biochem. Mol. Biol. 113, 331–339.
Dubreuil, O., Bossus, M., Graille, M., Bilous, M., Savatier, A., Jolivet, M.,Menez, A., Stura, E., Ducancel, F., 2005. Fine tuning of the specificity of
Please cite this article in press as: Riaño-Umbarila, L., et al., Aneutralizing Mexican scorpion toxins, Toxicon (2013), http://dx.do
an anti-progesterone antibody by first and second sphere residueengineering. J. Biol. Chem. 280, 24880–24887.
Espino-Solis, G.P., Riano-Umbarila, L., Becerril, B., Possani, L.D., 2009.Antidotes against venomous animals: state of the art and pro-spectives. J. Proteomics 72, 183–199.
Ewert, S., Huber, T., Honegger, A., Pluckthun, A., 2003. Biophysical prop-erties of human antibody variable domains. J. Mol. Biol. 325, 531–553.
Hernandez-Salgado, K., Estrada, G., Olvera, A., Coronas, F.I., Possani, L.D.,Corzo, G., 2009. Heterologous expressed toxic and non-toxic peptidevariants of toxin CssII are capable to produce neutralizing antibodiesagainst the venom of the scorpion Centruroides suffusus suffusus.Immunol. Lett. 125, 93–99.
Hmila, I., Saerens, D., Ben Abderrazek, R., Vincke, C., Abidi, N., Benlasfar, Z.,Govaert, J., El Ayeb, M., Bouhaouala-Zahar, B., Muyldermans, S., 2010.A bispecific nanobody to provide full protection against lethal scor-pion envenoming. FASEB J. 24, 3479–3489.
Holliger, P., Hudson, P.J., 2005. Engineered antibody fragments and therise of single domains. Nat. Biotechnol. 23, 1126–1136.
Huston, J.S., Mudgett-Hunter, M., Tai, M.S., McCartney, J., Warren, F.,Haber, E., Oppermann, H., 1991. Protein engineering of single-chain Fvanalogs and fusion proteins. Methods Enzymol. 203, 46–88.
Juarez-Gonzalez, V.R., Riano-Umbarila, L., Quintero-Hernandez, V., Ola-mendi-Portugal, T., Ortiz- Leon, M., Ortiz, E., Possani, L.D., Becerril, B.,2005. Directed evolution, phage display and combination of evolvedmutants: a strategy to recover the neutralization properties of thescFv version of BCF2 a neutralizing monoclonal antibody specific toscorpion toxin Cn2. J. Mol. Biol. 346, 1287–1297.
Leung, D.W., Chen, E., Goeddel, D.V., 1989. A method for random muta-genesis of a defined DNA segment using a modified polymerase chainreaction. Technique 1, 11–15.
Marks, J.D., Hoogenboom, H.R., Bonnert, T.P., McCafferty, J., Griffiths, A.D.,Winter, G., 1991. By-passing immunization. Human antibodies fromV-gene libraries displayed on phage. J. Mol. Biol. 222, 581–597.
Meyerhans, A., Vartanian, J.P., Wain-Hobson, S., 1990. DNA recombinationduring PCR. Nucleic Acids Res. 18, 1687–1691.
Miyazaki, C., Iba, Y., Yamada, Y., Takahashi, H., Sawada, J., Kurosawa, Y.,1999. Changes in the specificity of antibodies by site-specific muta-genesis followed by random mutagenesis. Protein Eng. 12, 407–415.
Ramirez, A.N., Martin, B.M., Gurrola, G.B., Possani, L.D., 1994. Isolation andcharacterization of a novel toxin from the venom of the scorpionCentruroides limpidus limpidus Karsch. Toxicon 32, 479–490.
Riano-Umbarila, L., Contreras-Ferrat, G., Olamendi-Portugal, T., Morelos-Juarez, C., Corzo, G., Possani, L.D., Becerril, B., 2011. Exploiting cross-reactivity to neutralize two different scorpion venoms with one sin-gle chain antibody fragment. J. Biol. Chem. 286, 6143–6151.
Riano-Umbarila, L., Juarez-Gonzalez, V.R., Olamendi-Portugal, T., Ortiz-Leon, M., Possani, L.D., Becerril, B., 2005. A strategy for the generationof specific human antibodies by directed evolution and phage display.An example of a single-chain antibody fragment that neutralizes amajor component of scorpion venom. FEBS J. 272, 2591–2601.
Rodriguez-Rodriguez, E.R., Ledezma-Candanoza, L.M., Contreras-Ferrat, L.G., Olamendi- Portugal, T., Possani, L.D., Becerril, B., Riano-Umbarila, L., 2012. A single mutation in framework 2 of the heavyvariable domain improves the properties of a diabody and a relatedsingle-chain antibody. J. Mol. Biol. 423, 337–350.
Rodriguez de la Vega, R.C., Possani, L.D., 2005. Overview of scorpiontoxins specific for Naþ channels and related peptides: biodiversity,structure-function relationships and evolution. Toxicon 46, 831–844.
Schiavon, E., Pedraza-Escalona, M., Gurrola, G.B., Olamendi-Portugal, T.,Corzo, G., Wanke, E., Possani, L.D., 2012. Negative-shift activation,current reduction and resurgent currents induced by beta-toxinsfrom Centruroides scorpions in sodium channels. Toxicon 59, 283–293.
Schiavon, E., Sacco, T., Cassulini, R.R., Gurrola, G., Tempia, F., Possani, L.D.,Wanke, E., 2006. Resurgent current and voltage sensor trappingenhanced activation by a beta-scorpion toxin solely in Nav1.6 chan-nel. Significance in mice Purkinje neurons. J. Biol. Chem. 281, 20326–20337.
Zamudio, F., Saavedra, R., Martin, B.M., Gurrola-Briones, G., Herion, P.,Possani, L.D., 1992. Amino acid sequence and immunological char-acterization with monoclonal antibodies of two toxins from thevenom of the scorpion Centruroides noxius Hoffmann. Eur. J. Biochem.204, 281–292.
novel human recombinant antibody fragment capable ofi.org/10.1016/j.toxicon.2013.09.016