a novel methodology for characterizing strand-break termini and damaged bases in plasmid dna exposed...
TRANSCRIPT

ANALYTICALBIOCHEMISTRY
Analytical Biochemistry 362 (2007) 229–235
www.elsevier.com/locate/yabio
A novel methodology for characterizing strand-break termini and damaged bases in plasmid DNA exposed to ionizing radiation
Ken Akamatsu ¤
Radiation EVect Analysis Group, Nuclear Science and Engineering Directorate, Japan Atomic Energy Agency, 2-4 Shirakatashirane, Tokai-mura, Naka-gun, Ibaraki 319-1195, Japan
Received 2 November 2006Available online 26 January 2007
Abstract
We have developed a de novo methodology to characterize radiation damage in DNA. An enzyme system consisting of the 3�!5� exo-nuclease snake venom phosphodiesterase (SVPD) and calf intestine alkaline phosphatase (CIAP) was used to examine the 3� termini ofstrand-break sites. In this study, we hypothesized that the strand-break termini can be divided into two categories: CIAP-independentSVPD sites and CIAP-dependent SVPD sites. The former consists of strand-break termini that can be recognized directly and digested bySVPD without CIAP pretreatment, whereas the latter includes the termini that cannot be digested by SVPD without CIAP pretreatment.In addition, the apparent radiation–chemical yield (G value) can be estimated using the level of intact 2�-deoxynucleotides produced dur-ing a 15-min incubation with SVPD. The G value for total strand breaks in fully dried DNA irradiated with 60Co �-rays was estimated tobe 0.1 �mol/J. Moreover, the G values of CIAP-dependent and CIAP-independent SVPD sites were estimated to be 0.078 and 0.024 �mol/J, respectively. These values suggest that 3�-phosphate termini are more likely to be produced than 3� termini without phosphate. Further-more, piperidine-treated irradiated plasmid DNA was also treated with the same enzyme system to examine the piperidine-labile sites. Asa result of the treatment, the G value of the CIAP-dependent SVPD sites increased to 0.16 �mol/J, whereas no signiWcant increase wasseen in the G value of the CIAP-independent SVPD sites. This observation implies that most piperidine-labile damaged bases can be elim-inated to form apurinic/apyrimidinic sites, which are completely removed by piperidine treatment to form 3� phosphate termini, and thatprompt CIAP-independent SVPD sites are piperidine resistant.© 2006 Elsevier Inc. All rights reserved.
Keywords: DNA damage; Strand-break terminus; Base damage; Ionizing radiation; Phosphodiesterase; Phosphatase; Enzyme kinetics
DNA damage is known to be a major cause of cell death,mutations, and carcinogenesis. In particular, if strandbreaks along the sugar–phosphate backbone are not com-pletely repaired, they can perturb cellular homeostasis. Pre-viously, the yield of single- or double-strand breaks inDNA induced by ionizing radiation has been calculatedusing agarose electrophoresis and is widely reported, e.g.,[1]. This method, however, does not produce any additionalinformation about the characteristics of the strand-breaktermini, such as the chemical structure or the size of theoverhangs at double-strand break sites. In fact, double-
* Fax: +81 29 282 6768.E-mail address: [email protected]
0003-2697/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.ab.2006.12.006
strand breaks produced by high linear energy transfer(LET)1 alpha particles are more diYcult to rejoin in livingcells than those produced by low LET 137Cs �-rays [2,3]. Inthe past few decades, some strand-break termini induced bychemicals or ionizing radiation and initiated by hydrogenabstraction from the sugar moiety have been characterizedusing a variety of techniques, including high-performanceliquid chromatography (HPLC), gas chromatography–mass spectrometry, and polyacrylamide electrophoresis [4].These studies, which were performed using DNA oligomers,
1 Abbreviations used: LET, linear energy transfer; pdN, 5�-mono-deoxynucleatide; SVPD, snake venom phosphodiesterase; AP, apurinic/apyramidinic; CIAP, calf intestine alkaline phosphatase.

230 Novel method to characterize radiation damage in DNA / K. Akamatsu / Anal. Biochem. 362 (2007) 229–235
reveal that (i) hydrogen abstraction from the sugar moietyproduces a variety of alkaline-labile sites due to newly pro-duced carbonyl groups, (ii) a strand break that occurs via asugar radical is usually accompanied by the release of a freenucleobase, and (iii) a sugar residue related to a strandbreak is almost always located at the 3�-phosphate termi-nus (e.g., 3�-phosphoglycolate and 3�-phosphoglycalde-hyde). C-H hydrogen atoms in the sugar can be abstractednot only by hydroxyl radicals produced by the radiolysis ofwater molecules and Fenton-like reactions but also bysome enediyne antibiotics and anticancer agents, such asneocarzinostatin and bleomycin, respectively. Therefore,strand-break termini derived from the neutral sugar carbonradical have been extensively investigated. Considering theeVects of ionizing radiation on DNA, however, directenergy transfer from the photons and ion particles andreactions with hydroxyl radicals should be taken intoaccount because the probability of “direct-type eVects” isfairly high in a living cell [2,3]. The resulting breaks due todirect-type eVects, however, have not been suYciently eluci-dated, in part because almost all of the occupied electronsin DNA can be excited and ionized by these eVects. In otherwords, direct-type eVects cause ionizing- radiation-speciWcevents that are not induced by other agents. As mentionedabove, however, sugar neutral radicals can be produced bydirect-type eVects via C-H+ radical cations [5,6].
To obtain chemical information about strand-breaktermini caused by direct-type eVects, an artiWcial oligonu-cleotide a few tens of basepairs long is required forhigh-resolution analysis, though actual strand scission in acell occurs in high-molecular-weight DNA [7,8]. Moreover,32P-end-labeled polydeoxynucleotides are needed asirradiation samples [9]. In this report, we propose a newmethodology to investigate strand-break termini and base-damaged sites in plasmid DNA based on the rate at which5�-monodeoxynucleotides (pdNs) are produced from thetermini by the 3�!5� exonuclease SVPD. SVPD recognizesand removes damaged 5�-deoxynucleotide, dihydrothymi-dine-5�-phosphates at the same rate as that for diVerentintact 5�-deoxynucleotides, whereas thymidine glycol-5�-phosphate and some tandem lesions are refractory toSVPD [10,11]. Moreover, SVPD cannot digest the phos-phodiester linkage immediately 3� to the apurinic/apyrimi-dinic (AP) site [12] but instead will cleave the penultimatephosphodiester bond to produce a pdN with the AP site[13]. These characteristics of SVPD suggest that the three-dimensional structure of the nucleobase plays an importantrole in determining the cleavability of the 5�-mono-deoxynucleotide from the 3� terminus. Hence, under experi-mental conditions in which few tandem lesions areproduced, the rate of pdN production is related to the num-ber of 3� termini that lack a free phosphate. Additionally,the total number of 3� termini can be estimated using phos-phatase-pretreated damaged DNA. Thus, for this study, wehypothesized that the strand-break termini could bedivided into two categories: CIAP-independent SVPD sitesand CIAP-dependent SVPD sites. The former consists of
the strand-break termini that can be directly recognizedand digested by SVPD without CIAP pretreatment,whereas the latter includes the termini that cannot bedigested without CIAP pretreatment. Furthermore, to esti-mate the yield of base-damaged sites, the irradiated DNAwas incubated in an aqueous solution of piperidine at 90 °Cfor 30 min before the enzyme treatment to produce 3�-phos-phate termini at the damaged sites. Several damaged nucle-otides with oxidized bases except for at least 8-oxo-guanineand 8-oxo-adenine are known to be piperidine-labile sites(for a review see Burrows and Muller [14]). The proposedmethodology serves as a model system to examine the char-acteristics of strand-break termini and base-damaged sitesinduced in natural DNA. Strategic use of a variety of site-speciWc enzymes in this type of analysis should clarify thediVerences between the types of DNA damage induced byionizing radiation.
Materials and methods
Chemicals
The 2686-bp pUC19 DNA plasmid (1 mg/mL in 10 mMTE buVer at pH 8.0) was purchased from Bayou Biolabs(Haralan, LA). The 2�-deoxynucleosides (dNs), 2�-deoxy-cytidine (dC), 2�-deoxyguanosine (dG), thymidine (dT), and2�-deoxyadenosine (dA), and the nucleobases cytosine (C),guanine (G), thymine (T), and adenine (A) were purchasedfrom Sigma–Aldrich (Japan). All chemicals were used with-out further puriWcation.
Enzymes
Sma I (Serratia marcescens Sb, 12 units/�L), Sau3 A I(Staphylococcus aureus 3A, 10 units/�L), snake venomphosphodiesterase (Crotalus adamanteus, 39.0 units/mgsolid, 1.29 A280/mg/mL), and calf intestine alkaline phos-phatase were purchased from Amersham–Pharmacia Bio-tech (Piscataway, NJ). The unit deWnitions of the nucleasesare as follows: Sma I, 1 unit is deWned as the amount ofenzyme required to completely digest 1 �g of � DNA in 1 hat pH 7.5 and 30 °C in 50 �L of assay buVer; Sau3 A I,1 unit is deWned as the amount of enzyme required to com-pletely digest 1 �g of � DNA in 1 h at pH 7.5 and 30 °C in50 �L of assay buVer; SVPD, 1 unit is deWned as the amountof enzyme required to hydrolyze 1 �mol of p-nitrophenylthymidine 5�-phosphate per minute at pH 8.9 and 25 °C;CIAP, 1 unit is deWned as the amount of enzyme requiredto hydrolyze 1 �mol of p-nitrophenyl phosphate per minuteat pH 8.0 and 37 °C.
Preparation of linear pUC19 by Sma I digestion
pUC19 plasmid DNA (1 mg/mL in TE at pH 8.0) waspuriWed twice by ethanol precipitation. PuriWed EDTA-freepUC19 was incubated in buVer (10 mM Tris–HCl at pH7.5, 7 mM MgCl2, 20 mM KCl, and 7 mM 2-mercaptotha-

Novel method to characterize radiation damage in DNA / K. Akamatsu / Anal. Biochem. 362 (2007) 229–235 231
nol) with Sma I (84 units, 7 �L) at 30 °C for 7 h (»250�g ofDNA/200 �L of reaction buVer). The reaction mixture wasphenol extracted to remove the enzyme, and the DNA waspuriWed by ethanol precipitation. The concentration of theaqueous solution of Sma I-digested pUC19 (pUC19/Sma I)was determined by measuring the OD260. The purity of thesamples of pUC19/Sma I was conWrmed by calculating theOD260/OD280 ratio; samples of pUC19/Sma I with a ratiogreater than 1.8 were used in this study. Agarose electro-phoresis analysis showed that the degree of digestion wasmore than 95% (data not shown). The DNA solution wasused without further puriWcation.
Preparation of pUC19/Sau3A I
Fifteen double-stranded DNA fragments produced bythe treatment of pUC19 with Sau3 A I were needed toobtain a calibration curve that allowed the conversion ofthe concentrations of recovered dNs into those of the 3� ter-mini. These fragments with a total of 30 recessed 3�-OH ter-mini were expected to function as the model molecules ofdouble-stranded DNA.
The preparation process was similar to that of pUC19/Sma I. In brief, puriWed EDTA-free pUC19 (»175�g) wasincubated in 200 �L of reaction buVer (10 mM Tris–HCl atpH 7.5, 10 mM MgCl2, 50 mM NaCl, and 1 mM dithiothrei-tol) with Sau3 A I (50 units, 5 �L) at 37 °C for 2 h. An addi-tional 50 units of the enzyme was added to the reactionmixture and the sample was incubated at 37 °C for 4 h tocomplete the digestion reaction. The DNA solution wasused without further puriWcation.
Preparation of Xuorinated polymer-coated silicon plates
A silicon plate (15£ 6.5 mm) was washed thoroughlywith ethanol using ultrasonication. A piece of mending tape(12£ 3 mm) was placed onto the plate. The partially sealedplate was dipped in a solution of Xuorinated resin andpetroleum ether. After drying in a draft chamber at roomtemperature, the tape was removed. Twenty microliters of1 N sodium hydroxide was placed on the uncoated area for1 min to increase the hydrophilicity of the surface. The platewas then washed thoroughly with water to remove the alka-line solution.
Preparation of pUC19/Sma I for gamma irradiation
Twenty-four microliters of pUC19/Sma I aqueous solu-tion (2 �g/�L) was mounted on the uncoated area of a plateand the plate was partially dried in a draft chamber. Theplates carrying pUC19/Sma I were thoroughly dried undervacuum over P2O5 for a few days. Each sample on the platewas put into a glass tube (external diameter 10 mm; internaldiameter 6.8 mm), which was sealed under a vacuum(»0.1 Pa). The glass tube with a plate was placed into alarger glass tube (total glass thickness 4.0 mm) to achievesecondary electron equilibrium.
Gamma irradiation of pUC19/Sma I
Gamma irradiation (60Co, 1.17 and 1.33 MeV) of thepUC19/Sma I samples was performed at the TakasakiAdvanced Radiation Research Institute (Japan AtomicEnergy Agency). The level of radioactivity was 10,800 TBqat the beginning of the experiment. Each sample was placedat positions 277, 158, 56, 23, and 3 cm away from the 60Cosource (exposure rates 9.9, 24.8, 99.1, 222.9, and 445.8 C/kg/h, respectively) and irradiated for 120 h at room tempera-ture (doses 40, 100, 400, 900, and 1800 kGy, respectively).The dosimetry was also checked using an alanine–polysty-rene dosimeter [15] (data not shown). It should be notedthat approximately 90% of the 2�-deoxynucleotide unitsthat constitute a pUC19/Sma I plasmid remain intact evenafter irradiation at a dose of 1800 kGy (data not shown).A control sample was prepared with the same method exceptthat it remained outside of the irradiation room. Each samplewas kept at 0–4 °C until it was dissolved in water.
Treatment of the irradiated pUC19/Sma I samples with CIAP and SVPD
An irradiated pUC19/Sma I sample was transferred to amicrotube and lyophilized. The residue was resuspendedwith 150 �L of cold water (stock solution, 0.32 �g/�L). Tenmicroliters of the irradiated DNA stock solution was addedinto 50-mL reaction assembled on ice with the Wnal reac-tion conditions of 1£SVPD buVer (0.1 M Tris–HCl, 0.1 MNaCl, and 14 mM MgCl2 at pH 8.0), 0.0001 unit/�L SVPD,and 0.08 unit/�L CIAP. The microtube containing the reac-tion mixture was transferred to a water bath, incubated at31.4 °C for 15 min, and terminated by cooling the reactionto 0 °C and adding 5 �L of 0.5 M EDTA (pH 8.0). The reac-tion mixture containing the dNs was analyzed by HPLC asdescribed below. The brief protocol is shown in Fig. 1.
Treatment of the irradiated pUC19/Sma I samples with SVPD followed by CIAP treatment
An irradiated pUC19/Sma I sample was transferred to amicrotube and lyophilized. The residue was resuspendedwith 150 �L of cold water (stock solution, 0.32 �g/�L). Tenmicroliters of the irradiated DNA stock solution was addedinto 50-�L reaction assembled on ice with the Wnal reactionconditions of 1£SVPD buVer and 0.0001 unit/�L SVPD.The microtube containing the reaction mixture was trans-ferred to a water bath, incubated at 31.4 °C for a givenperiod of time, and terminated by transferring the tube intoliquid nitrogen (77 K) for 10 s. The solution at »0 °C wastransferred to a 5-kDa molecular-weight cutoV Wlter (Milli-pore) and centrifuged at 6000g at 0 °C to remove the SVPD.The Wlter was washed with 100 �L of cold water three timesto recover the digested 2�-deoxynucleotide-5�-phosphate.The Wltered solution (»350�L) was lyophilized. Theobtained residue was resuspended with 50�L of water,mixed with 2�L of CIAP, and incubated at 37 °C for

232 Novel method to characterize radiation damage in DNA / K. Akamatsu / Anal. Biochem. 362 (2007) 229–235
30 min. The reaction mixture was transferred to a 10-kDamolecular-weight cutoV Wlter (Millipore) and centrifuged at6000g at 0 °C to remove the CIAP. The Wlter was washedwith 100 �L of cold water three times to recover dNs. TheWltered solution containing the dNs was analyzed by HPLCas described below. The brief protocol is shown in Fig. 1.
Quantitative analysis of the 2�-deoxynucleosides using HPLC
The enzymes in the reaction mixture were removed usinga 5-kDa molecular-weight cutoV Wlter and centrifugation at0 °C and 6000g. The tube and the Wltration membrane werewashed thoroughly with cold water (>18M�cm) at 0°C(total volume 300�L). The Wltered solution (total volume»350�L) was lyophilized. The residue was dissolved in 26�Lof cold water at 0°C. Twenty microliters of the sample wasanalyzed by HPLC. The HPLC system (CLASS-VP, Shi-madzu) was equipped with a C-30 reverse-phase column(Develosil RPAQUEOUS; 4.6£150 mm; Nomura Chemi-cal), an autosampler (SIL-10ADvp, Shimadzu), and a photo-diode alley UV–VIS detector (SPD-M10Avp, Shimadzu)that monitored at 210 and 260nm. Samples were eluted with
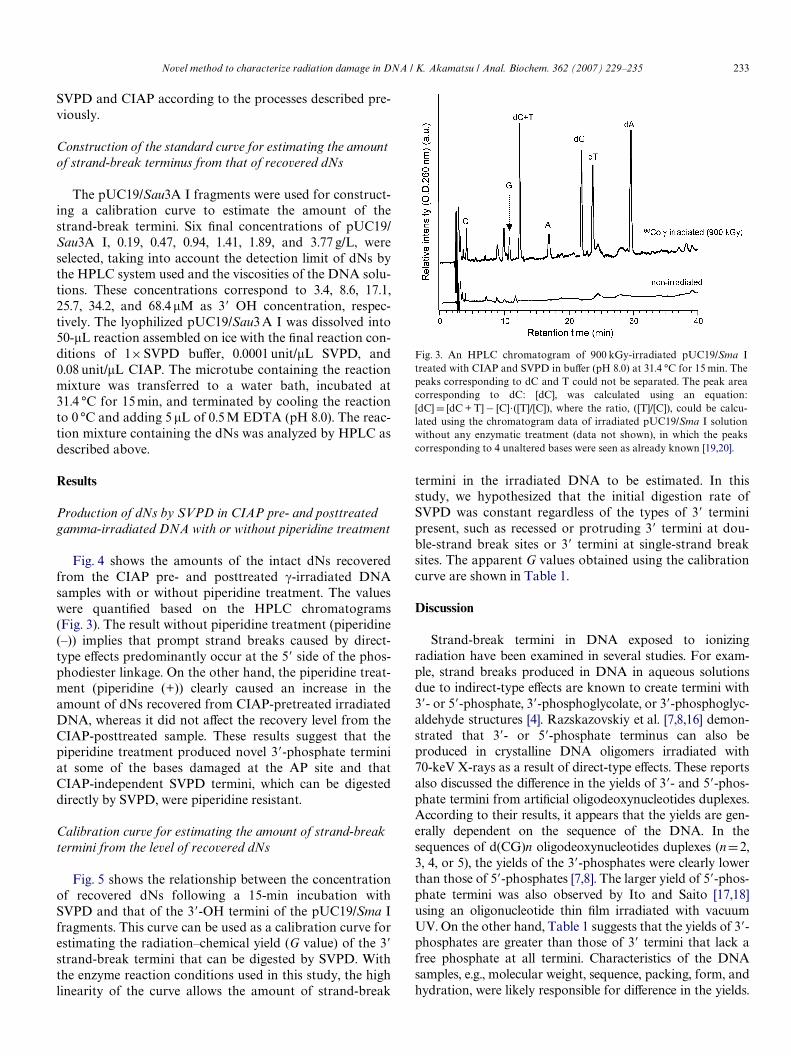
a linear gradient of acetonitrile (0–20% over 40 min) in50 mM ammonium acetate at 40 °C with a Xow rate of0.8 mL/min. The typical chromatogram is shown in Fig. 3.Authentic dNs solutions were prepared for quantitative anal-ysis as follows: dA monohydrate (FW 251.24, 25.6 mg), dCmonohydrate (FW 227.22, 22.6 mg), dG containing 6% water(FW 251.24, 25.6 mg), and anhydrous dT (FW 242.23,24.2 mg) were dissolved together in 1000 mL of 50 mMammonium formate (pH 6.86). The Wnal concentration ofeach dN in the standard solution was 0.1019, 0.0995, 0.0958,and 0.0999 mM, respectively. No undigested pdN wasdetected following treatment with an excess amount ofCIAP.
Treatment of the irradiated pUC19/Sma I sample with piperidine
An irradiated pUC19/Sma I sample was dissolved into1 M piperidine. The solution was incubated for 30 min at90 °C, which removed most of the piperidine-labile dam-aged nucleosides to produce 3�-phosphate termini via APsite formation (Fig. 2). The piperidine-treated samples werecooled with liquid nitrogen, lyophilized, and treated with
Fig. 1. The analytical scheme to estimate the yields of CIAP-independent and CIAP-dependent SVPD sites.
Fig. 2. Strand-break formation at piperidine-labile damaged bases as a result of piperidine treatment. It is known that SchiV’s base is reversibly formedbetween C1� and the amine, which increase the basicity of H2� [14]. The mechanism of the �- and �-elimination reactions, that also occur by, e.g., formam-idopyrimidine N-glycosylase (Fpg), was investigated in detail by Tchou [24] and Bhagwat [25].
Novel method to characterize radiation damage in DNA / K. Akamatsu / Anal. Biochem. 362 (2007) 229–235 233
SVPD and CIAP according to the processes described pre-viously.
Construction of the standard curve for estimating the amount of strand-break terminus from that of recovered dNs
The pUC19/Sau3A I fragments were used for construct-ing a calibration curve to estimate the amount of thestrand-break termini. Six Wnal concentrations of pUC19/Sau3A I, 0.19, 0.47, 0.94, 1.41, 1.89, and 3.77 g/L, wereselected, taking into account the detection limit of dNs bythe HPLC system used and the viscosities of the DNA solu-tions. These concentrations correspond to 3.4, 8.6, 17.1,25.7, 34.2, and 68.4 �M as 3� OH concentration, respec-tively. The lyophilized pUC19/Sau3 A I was dissolved into50-�L reaction assembled on ice with the Wnal reaction con-ditions of 1£SVPD buVer, 0.0001 unit/�L SVPD, and0.08 unit/�L CIAP. The microtube containing the reactionmixture was transferred to a water bath, incubated at31.4 °C for 15 min, and terminated by cooling the reactionto 0 °C and adding 5 �L of 0.5 M EDTA (pH 8.0). The reac-tion mixture containing the dNs was analyzed by HPLC asdescribed above.
Results
Production of dNs by SVPD in CIAP pre- and posttreated gamma-irradiated DNA with or without piperidine treatment
Fig. 4 shows the amounts of the intact dNs recoveredfrom the CIAP pre- and posttreated �-irradiated DNAsamples with or without piperidine treatment. The valueswere quantiWed based on the HPLC chromatograms(Fig. 3). The result without piperidine treatment (piperidine(–)) implies that prompt strand breaks caused by direct-type eVects predominantly occur at the 5� side of the phos-phodiester linkage. On the other hand, the piperidine treat-ment (piperidine (+)) clearly caused an increase in theamount of dNs recovered from CIAP-pretreated irradiatedDNA, whereas it did not aVect the recovery level from theCIAP-posttreated sample. These results suggest that thepiperidine treatment produced novel 3�-phosphate terminiat some of the bases damaged at the AP site and thatCIAP-independent SVPD termini, which can be digesteddirectly by SVPD, were piperidine resistant.
Calibration curve for estimating the amount of strand-break termini from the level of recovered dNs
Fig. 5 shows the relationship between the concentrationof recovered dNs following a 15-min incubation withSVPD and that of the 3�-OH termini of the pUC19/Sma Ifragments. This curve can be used as a calibration curve forestimating the radiation–chemical yield (G value) of the 3�strand-break termini that can be digested by SVPD. Withthe enzyme reaction conditions used in this study, the highlinearity of the curve allows the amount of strand-break
termini in the irradiated DNA to be estimated. In thisstudy, we hypothesized that the initial digestion rate ofSVPD was constant regardless of the types of 3� terminipresent, such as recessed or protruding 3� termini at dou-ble-strand break sites or 3� termini at single-strand breaksites. The apparent G values obtained using the calibrationcurve are shown in Table 1.
Discussion
Strand-break termini in DNA exposed to ionizingradiation have been examined in several studies. For exam-ple, strand breaks produced in DNA in aqueous solutionsdue to indirect-type eVects are known to create termini with3�- or 5�-phosphate, 3�-phosphoglycolate, or 3�-phosphoglyc-aldehyde structures [4]. Razskazovskiy et al. [7,8,16] demon-strated that 3�- or 5�-phosphate terminus can also beproduced in crystalline DNA oligomers irradiated with70-keV X-rays as a result of direct-type eVects. These reportsalso discussed the diVerence in the yields of 3�- and 5�-phos-phate termini from artiWcial oligodeoxynucleotides duplexes.According to their results, it appears that the yields are gen-erally dependent on the sequence of the DNA. In thesequences of d(CG)n oligodeoxynucleotides duplexes (nD2,3, 4, or 5), the yields of the 3�-phosphates were clearly lowerthan those of 5�-phosphates [7,8]. The larger yield of 5�-phos-phate termini was also observed by Ito and Saito [17,18]using an oligonucleotide thin Wlm irradiated with vacuumUV. On the other hand, Table 1 suggests that the yields of 3�-phosphates are greater than those of 3� termini that lack afree phosphate at all termini. Characteristics of the DNAsamples, e.g., molecular weight, sequence, packing, form, andhydration, were likely responsible for diVerence in the yields.
Fig. 3. An HPLC chromatogram of 900 kGy-irradiated pUC19/Sma Itreated with CIAP and SVPD in buVer (pH 8.0) at 31.4 °C for 15 min. Thepeaks corresponding to dC and T could not be separated. The peak areacorresponding to dC: [dC], was calculated using an equation:[dC] D [dC + T]¡ [C].([T]/[C]), where the ratio, ([T]/[C]), could be calcu-lated using the chromatogram data of irradiated pUC19/Sma I solutionwithout any enzymatic treatment (data not shown), in which the peakscorresponding to 4 unaltered bases were seen as already known [19,20].

234 Novel method to characterize radiation damage in DNA / K. Akamatsu / Anal. Biochem. 362 (2007) 229–235
Another possible reason for the diVerences is that the rate atwhich SVPD digests a 3� terminus bearing a sugar fragmentwithout a phosphate (CIAP-independent SVPD site) may belower than the rate at which the enzyme digests intact 3�-OHtermini. Stuart and Chambers [13] indicated that SVPD doesnot hydrolyze the Wrst phosphodiester bond of a 3� terminusproduced by the �-elimination of an AP site well but insteadcleaves the penultimate phosphodiester bond as an endonu-clease. The SVPD digestion rate for a 3� terminus with asugar residue, however, could be lower than that for a nor-mal 3�-OH terminus.
Direct-type radiation eVects also cause certain types ofdamaged bases. In fact, unaltered base release occurs as aresult of these eVects [19,20]. In this study, we hypothesized
Fig. 5. The relationship between the 3�-OH terminus concentration in thepUC19/Sau3A I fragments and total dNs recovered following SVPD andCIAP treatment in buVer (pH 8.0) for 15 min at 31.4 °C. The slope of theregression line, which was obtained by a least square Wt without the datapoint at the 3�-OH terminus concentration of 68.4 �M, is 0.723 (correla-tion coeYcient: 0.999).
that most nucleotides with damaged bases and nucleotideswithout bases due to unaltered base release events should bealkaline labile, resulting in strand breaks from, e.g., �-elimina-tion around the carbonyl moiety bearing in the sugar residue.We have previously identiWed the carbonyl groups in X-irra-diated 2-deoxyribose Wlm by means of X-ray absorption near-edge structure spectroscopy with oxygen K-edge [21,22] andinfrared spectroscopy [23]. In the case of an AP site, the Wnalstructure at a 3� terminus created by alkaline treatment is a3�-phosphate, which can be a CIAP-dependent SVPD site.Actually, the data shown in Fig. 4 and Table 1 indicate that3�-phosphate termini are produced by piperidine treatment,suggesting that the sugar residues that remained after theremoval of the damaged bases were eliminated. Unalteredbase released from irradiated dried DNA can easily be investi-gated using an HPLC system [19,20]. As shown in Fig. 3, threepeaks corresponding to C, G, and A nucleotides were clearlyvisible. There is some controversy about the mechanism bywhich this type of base release occurs. One hypothesis is thatthe base release results from sugar radicals [20]. According tothis hypothesis, chemical rearrangements after the productionof neutral sugar radicals are expected to be similar to those
Table 1Apparent strand break yields (G values, �mol/J) in 60Co �-irradiatedpUC19/Sma I based on SVPD digestion kinetics to 3�OH termini inpUC19/Sau3A I fragments
a The yield was obtained by HPLC analysis of low-molecular-weightfraction of the irradiated DNA without any enzymatic treatments.
Category of 3� terminus G value (�mol/J)
piperidine (¡) piperidine (+)
CIAP-independent SVPD site 0.024 0.026CIAP-dependent SVPD site 0.078 0.160Total 0.102 0.186Unaltered base released 0.018a —
Fig. 4. dNs recovered following SVPD treatment (15 min at 31.4 °C and pH 8.0) from pUC19/Sma I irradiated with 900 and 1800 kGy of 60Co �-rays withor without CIAP pretreatment. EVect of piperidine pretreatment was also investigated at 900 kGy. The bars and error bars represent the mean § 1 SDfrom two experiments.

Novel method to characterize radiation damage in DNA / K. Akamatsu / Anal. Biochem. 362 (2007) 229–235 235
produced by hydrogen abstraction from the sugar by ahydroxyl radical [4]. The neutral sugar radicals result in therelease of unaltered bases and the production of sugar rem-nants without phosphates, which can be digested at the penul-timate phosphodiester linkage [13]. Indeed, the G value ofunaltered base released is similar to the apparent G value ofCIAP-independent SVPD sites (Table 1). The sugar rem-nants, however, are not entirely removed because the G valueof the CIAP-independent SVPD site was not aVected by thepiperidine treatment (Table 1). If the sugar remnants wereentirely eliminated by this treatment, the termini shouldchange to 3�-phosphate CIAP-dependent SVPD sites (Fig. 2).
Damaged bases produced by direct-type eVects were iden-tiWed by Swarts et al. [6] using a 137Cs irradiator. They foundthat mainly 8-oxo-guanine, 8-OH-adenine, and 5,6-dihydro-thymine were produced in fully dried DNA (2.5 water mole-cules per nucleotide). The G values of base damage and baserelease from their data were 0.156 and 0.073�mol/J, respec-tively, whereas those from the data estimated from the valuesobtained using the 60Co irradiator (Table 1) were 0.082 and0.018�mol/J. The diVerence in these values may be correlatedwith the diVerence in the strand-break yields observed withthe diVerent light sources [1] (Fig. 5).
In conclusion, the proposed analytical method usingSVPD and CIAP to characterize radiation-damaged DNAwill allow the analysis of the diVerences in the eVects of avariety of ionizing radiations on the damage spectra. If neces-sary, the digested damaged 2�-deoxynucleosides and thedamaged bases removed by piperidine treatment can be iden-tiWed using mass spectrometry. Furthermore, this methodol-ogy can potentially characterize other types of DNA damageusing a variety of functional enzymes. The use of 5�!3�exonucleases, such as calf spleen phosphodiesterase, to pro-duce 2�-deoxynucleoside-3�-phosphate would enable quanti-Wcation of 5�-termini without free phosphates. Pretreatmentof damaged DNA with a ligase would repair simple single-strand breaks that have a structure that is the same as thestructure produced by hydrolysis of the phosphodiesterbond. The elucidation of the direct radiation eVects on DNAwill be enhanced with this analytical system.
Acknowledgments
This research was supported by a Grant-in-Aid for Sci-ence Research for Young Scientists (No. 15710048) from
the Japan Society for the Promotion of Science. I thank Dr.Yasuhiko Kobayashi (JAEA) for help with the 60Co �-irra-diation. I also thank Drs. Kentaro Fujii, Akinari Yokoya,and Komei Saito (JAEA) for discussing our results andtheir meaningful insights.
References
[1] C. Leloup, G. Garty, G. Assaf, A. Cristovao, A. Breskin, R. Chec-hik, S. Shchemelinin, T. Paz-Elizur, Z. Livneh, R.W. Schulte, V.Bashkirov, J.R. Milligan, B. Grosswendt, Int. J. Radiat. Biol. 81(2005) 41–54.
[2] T.J. Jenner, C.M. deLara, P. O’Neill, D.L. Stevens, Int. J. Radiat. Biol.64 (1993) 265–273.
[3] C.M. deLala, T.J. Jenner, K.M. Stewart Townsent, S.J. Marsden, P.O’Neill, Radiat. Res. 144 (1995) 43–49.
[4] W.K. Pogozelski, T.D. Tullius, Chem. Rev. 98 (1998) 1089–1107.[5] K. Fujii, K. Akamatsu, A. Yokoya, Radiat. Res. 161 (2004) 435–441.[6] S.G. Swarts, D. Becker, M. Sevilla, K.T. Wheeler, Radiat. Res. 145
(1996) 304–314.[7] M.G. Debije, Y. Razskazovskiy, W.A. Bernhard, J. Am. Chem. Soc.
123 (2001) 2917–2918.[8] Y. Razskazovskiy, M.G. Debije, S.B. Howerton, L.D. Williams, W.A.
Bernhard, Radiat. Res. 160 (2003) 334–339.[9] W.D. Henner, S.M. Grunberg, W.A. Haseltine, J. Biol. Chem. 257
(1982) 11750–11754.[10] M. Weinfeld, K.M. Soderlind, G.W. Buchko, Nucleic Acids Res. 21
(1993) 621–626.[11] K.J. Bowman, R. LePla, Y. Guichard, P.B. Farmer, G.D.D. Jones,
Nucleic Acids Res. 29 (2001) e101.[12] M. Weinfeld, M. Liuzzi, M.C. Paterson, Nucleic Acids Res. 17 (1989)
3735–3745.[13] G.R. Stuart, R.W. Chambers, Nucleic. Acids Res. 18 (1987) 7451–
7462.[14] C.J. Burrows, J.G. Muller, Chem. Rev. 98 (1998) 1109–1151.[15] T. Kojima, Y. Haruyama, H. Tachibana, R. Tanaka, J. Okamoto, K.
Yagi, N. Tamura, H. Hara, S. Kashiwazaki, Radiat. Phys. Chem. 42(1993) 757–760.
[16] Y. Razskazovskiy, M.G. Debije, W.A. Bernhard, Radiat. Res. 159(2003) 663–669.
[17] T. Ito, M. Saito, Photochem. Photobiol. 48 (1998) 567–572.[18] T. Ito, M. Saito, Radiat. Phys. Chem. 37 (1991) 681–690.[19] S.G. Swarts, M. Sevilla, D. Becker, C.J. Tokar, K.T. Wheeler, Radiat.
Res. 129 (1992) 333–344.[20] Y. Razskazovskiy, M.G. Debije, W.A. Bernhard, Radiat. Res. 155
(2000) 436–441.[21] K. Akamatsu, A. Yokoya, Radiat. Res. 155 (2001) 449–452.[22] K. Akamatsu, A. Yokoya, J. Synchrotron Rad. 8 (2001) 1001–1002.[23] K. Akamatsu, K. Fujii, A. Yokoya, Nucl. Instr. Met. Phys. Res. B 199
(2003) 328–331.[24] J. Tchou, A.P. Grollman, J. Biol. Chem. 270 (1995) 11671–11677.[25] M. Bhagwat, J.A. Gerlt, Biochemistry 35 (1996) 659–665.