a, patrick honga, sarah baika, oscar a. negrete , … · cell migration from the peripheral mucosa...
TRANSCRIPT

1
DC-SIGN binds to HIV-1 gp120 in a distinct but overlapping fashion compared to ICAM-2 and
ICAM-3.
Stephen V. Sua, Patrick Honga, Sarah Baika, Oscar A. Negretea, Kevin B. Gurneya, and *Benhur
Leea,b,c
aDept of Microbiology, Immunology and Molecular Genetics, bDept. of Pathology & Laboratory
Medicine, cUCLA AIDS Institute, David Geffen School of Medicine, UCLA, Los Angeles, CA,
USA, 90095
Running title: Differential binding of DC-SIGN to gp120, ICAM-2 and ICAM-3
*Corresponding Author:Benhur Lee, M.D.
Dept of Microbiology, Immunology and Molecular Genetics
3821 Molecular Sciences Building
609 Charles E. Young Drive East
Los Angeles, CA 90095
Office: 310-794-2132
Fax: 310-267-2580
Email: [email protected]
JBC Papers in Press. Published on February 16, 2004 as Manuscript M400184200
Copyright 2004 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
SUMMARY
DC-SIGN is a C-type lectin that binds to endogenous adhesion molecules ICAM-2 and
ICAM-3 as well as the viral envelope glycoprotein HIV-1 gp120. We wished to determine if DC-
SIGN binds differently to its endogenous ligands ICAM-2 and ICAM-3 versus HIV-1 gp120.
We found that recombinant soluble DC-SIGN bound to gp120-Fc more than 100- and 50-fold
better than ICAM-2-Fc and ICAM-3-Fc, respectively. This relative difference was maintained
using DC-SIGN expressed on 3 different CD4-negative cell lines. Although the cell surface
affinity for gp120 varied by up to 4-fold on the cell lines examined, the affinity for gp120 was
not a correlate of the cell line’s ability to transfer virus. Monosaccharides with equatorial 4-OH
groups competed as well as D-mannose for gp120 binding to DC-SIGN, regardless of how the
other hydroxyl groups were positioned. Disaccharide competitors and glycan chip analysis
showed that DC-SIGN has a preference for oligosaccharides linked in an alpha-anomeric
configuration. Alanine scanning mutagenesis of DC-SIGN revealed that highly conserved
residues which coordinate calcium (D366) and/or are involved in both calcium and specific
carbohydrate interactions (E347, N349, E354, D355) significantly compromised binding to all 3
ligands. Mutating non-conserved residues (N311, R345, V351, G352, E353, S360, G361, N362)
minimally affected binding except for the D367 mutant, which enhanced gp120 binding but
diminished ICAM-2 and ICAM-3 binding. Conversely, mutating the moderately conserved
residue (G346) abrogated gp120 binding but enhanced ICAM-2 and ICAM-3 binding. Thus,
DC-SIGN appears to bind in a distinct but overlapping manner to gp120 when compared to
ICAM-2 and ICAM-3.
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
INTRODUCTION
DC-SIGN is a mannose-specific calcium-dependent (C-type) lectin expressed on
dendritic cells (DCs) and certain subsets of macrophages (1-3). DC-SIGN binds with high
affinity to the envelope glycoprotein gp120 of HIV and has been shown in vitro to transfer HIV
from monocyte-derived dendritic cells (MDDCs) to permissive CD4+ T-cells (2,4-6). It is
believed that the high-affinity interaction of gp120 with DC-SIGN on DCs facilitates HIV
infection of CD4+ permissive T-cells in the lymph nodes during the natural course of dendritic
cell migration from the peripheral mucosa to the secondary lymphoid organs. DC-SIGN also
binds to the glycosylated envelopes of other viruses such as feline immunodeficient virus (FIV)
(Aymeric et. al., in press), simian immunodeficiency virus (SIV) (7-10), hepatitis C (11-13),
Ebola (14-18), cytomegalovirus (CMV) (19) and Dengue (20,21). Importantly, DC-SIGN has
been shown to be the receptor responsible for the productive infection of DCs by Dengue virus
(20). Recent studies also show that DC-SIGN binds to non-viral pathogens such as the
Leishmania pifanoi (22,23), Schistosoma mansoni (22,24), and Helicobacter pylori (22) and
Mycobacterium tuberculosis (22,25,26). The binding of M. tuberculosis to DC-SIGN on DCs is
believed to trigger IL-10 secretion and thus compromise the immunostimulatory function of the
targeted DCs (27).
In addition to ligands derived from pathogens, DC-SIGN has been shown to bind to two
endogenous human ligands, ΙCAM-2 and ΙCAM-3. ΙCAM-2 is highly expressed on endothelial
cells and is thought to bind to DC-SIGN on a subset of DC precursors and mediate DC
emigration from the blood (28). ΙCAM-3 is expressed on naive T-cells and its interaction with
DC-SIGN on DCs is thought to play a crucial role in DC-mediated T-cell activation (29,30).
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
Peptide sequence and structural analysis indicate that DC-SIGN contains the prototypical
110-140 residue carbohydrate recognition domain (CRD) (31-33) common to C-type lectins. All
known structures of C-type lectin CRDs show significant structural conservation characterized
by the presence of two pairs of highly conserved disulfide bonds (one of which joins the N- and
C-termini of the CRDs), a set of conserved amino acids that form the hydrophobic core of the
domain, the highly conserved Asp-Glu-Cys (EDC) amino acid motif involved in both calcium
coordination and sugar binding, and a highly conserved aspartic (E) residue that serves to
coordinate calcium and facilitate in the correct folding of the domain. The CRDs of different C-
type lectins can recognize various saccharide ligands. DC-SIGN contains the highly conserved
Glu-Pro-Asn (EPN) motif and thus belongs to the mannose-specific family of lectins (31).
Nonetheless, the CRDs of C-type lectins can recognize a remarkably broad range of ligands and
have been shown to specifically bind also to proteins (34), lipids (35) and inorganic molecules
such as calcium carbonate (36). Moreover, some CRDs are able to bind to both protein and
saccharide ligands (37,38).
Biochemical and structural studies using synthetic substrates demonstrate that DC-SIGN
preferentially binds to oligomannosides such as those found on N-linked high-mannose type
glycoproteins (30,39-41). Indeed, binding studies with envelope glycoproteins gp95 of FIV
(Aymeric, et al., in press) and gp120 of HIV-1 (2,40) along with ΙCAM-2 (28) and ΙCAM-3 (30)
confirms that DC-SIGN binds high mannose glycans on these glycoproteins. Additional
carbohydrate profiling has led to the identification of Lewis blood group Ags (Lex, Ley, Lea, Leb)
(22) as additional oligosaccharide structures that are specifically bound by DC-SIGN.
Furthermore, recent reports also identified the highly mannosylated surface lipoglycan
lipoarabinomannan of M. tuberculosis, as a ligand that specifically binds DC-SIGN (25,27).
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
Recently, a mucosally transmitted pathogenic SHIV variant (SHIV 162P) was shown to
bind to DC-SIGN 3-fold better than its parental derivative, which was non-pathogenic and poorly
transmissible. This gain in DC-SIGN binding function was mapped to an addition of a N-linked
glycosylation site in the V2 loop (42). In addition, highly purified DC-SIGN+ DCs from human
gut biopsies can bind and transfer HIV ten- to a hundred-fold more efficiently than DC-SIGN-
DCs from the same tissue (43). These data suggest that DC-SIGN may play a role in the
pathogenesis of mucosal HIV transmission and the DC-SIGN/gp120 interface may be a
legitimate target for antimicrobial therapeutics. However, since DC-SIGN also binds to
endogenous ΙCAM-2 and ΙCAM-3, we wished to determine if there were any diffferences that
distinguish DC-SIGN binding to gp120 from binding to the two endogenous ligands ΙCAM-2
and ΙCAM-3.
In this study, we showed that recombinant gp120 binds with much greater affinity to
DC-SIGN than ΙCAM-2 and ΙCAM-3 in in vitro and cell surface binding assays. The cell
surface affinity of DC-SIGN for gp120 varied by up to 4-fold among the cell lines examined, but
the cell-surface affinity for gp120 did not correlate with DC-SIGN’s ability to transfer virus
suggesting that cell-specific cofactors may be involved. Competition experiments using gp120,
ICAM-2 and ICAM-3 revealed a preference for saccharide ligands with an equatorial 4-hydroxyl
group. More extensive glycan array profiling also indicated that the presence of an alpha
anomeric glycosidic linkage contributed to the high affinity binding of cognate oligosaccharide
ligands. Lastly, targeted alanine-scanning mutagenesis of the CRD of DC-SIGN identified
critical residues that differentiate DC-SIGN binding to each of the three ligands.
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
EXPERIMENTAL PROCEDURES
Materials- DNA restriction enzymes were obtained from New England Biolabs (Beverly, MA).
High fidelity Pfu Turbo DNA polymerase was obtained from Invitrogen (Carlsbad, CA), and
oligonucleotides were supplied from MWG Biotech (High Point, NC). All chemicals and
monosaccharides were purchased from Sigma-Aldrich (St. Louis, MO). Disaccharides and high
binding certified 96-well plates were purchased from Fisher (Pittsburg, PA). Iso-propyl-β-D-
thiogalactoside (IPTG) was obtained from BioVectra (Oxford, CT). Anti-human Fc monoclonal
antibodies conjugated with horseradish-peroxidase (HRP) and the 1-step Ultra TMB substrate for
ELISA were purchased from Pierce Biotechnoogy (Rockford, IL) . Purified recombinant ΙCAM-
2-Fc and ΙCAM-3-Fc proteins were purchased from R&D Systems, Inc. (Minneapolis, MN).
Recombinant gp120-Fc was produced and purified as described (40).
Cloning and Purification of sDC-SIGN- The entire extracellular domain of DC-SIGN (sDC-
SIGN) beginning with Ala77 and ending with DC-SIGN’s natural stop codon was cloned into
pET15b (Novagen, Madison, WI) using flanking PCR primers with NdeI and BamHI sites. The
resulting in-frame fusion gene contains an N-terminal 6x-Histidine tag followed by the sDC-
SIGN gene.
Freshly transformed BL21/DE3 bacteria was inoculated and allowed to reach mid-log
phase at 30° C with shaking before induction with IPTG at a final concentration of 1mM.
Expression of sDC-SIGN was carried out in 1 L of Luria-broth supplemented with 100 µg/ml of
ampicillin. The culture was incubated further for at least 3 hrs. before being harvested by
centrifuging at 4000x g for 10 min at 4°C. The resulting pellet was resuspended in 10 ml of 100
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
mM NaH2P4,10 mM Tris-HCL and 6 M Guanidine HCL, pH 8, and lysed by repeated sonication
(5 bursts of 1 min. duration at 4°C). The lysate was then supplemented with 0.01% β-
mercaptoethanol and incubated at 4°C for 2 hrs. The lysate was then centrifuged at 150,000x g
for 30 min at 4°C in a Beckman 55.2Ti rotor, and the supernatant was incubated with 800 µl of
Ni-NTA agarose resin (Qiagen, Valencia, CA) (pre-equilibrated with denaturing buffer) at 4°C
overnight. The resin was loaded onto a 30 cm chromatography column and all subsequent
washes were done with a 30-fold resin-volume excess of wash buffer starting with 30 mM
TrisHCl, pH 8, 0.5 M NaCl, 1 mM CaCl2, 6 M urea and 10 mM imidazole. The column was
then washed again with same buffer except 15 mM imidazole was used. Successive washes of 30
mM TrisHCl, pH 8, 0.5 M NaCl in decreasing concentrations of urea starting with 5 M urea were
performed to renature the protein. The protein was eluted with 30 mM TrisHCl, pH 8, 0.5 M
NaCl, 1 mM CaCl2 and 1 M imidazole. The eluant was dialyzed twice with 500 ml of 30 mM
TrisHCl, pH 8, 1.0 M NaCl and 1 mM CaCl2 to remove excess imidazole.
Saccharide competition assays- 20 µl of a 200 nM sDC-SIGN concentrate were coated onto high
binding 96-well plates in the presence of 30 mM TrisHCl, 30 mM NaHCO3 and 1 mM CaCl2
overnight at 4°C (20 µl per well). The plates were then blocked in the same buffer as above but
supplemented with 5% BSA (50 µl per well) at 37 ºC for 3 hrs. The plates were washed 5 times
with wash buffer (TBS supplemented with 1 mM CaCl2 and 0.1% Tween). 2.4 nM, 300 nM and
150 nM of recombinant gp120-Fc, ΙCAM-2-Fc and ΙCAM-3-Fc, respectively, were incubated in
parallel with the indicated amounts of the various saccharides at room temperature for 2 hrs. The
plates were then washed 5 times with wash buffer and incubated with anti-human Fc monoclonal
antibody conjugated with HRP (Pierce Biotechnology, Rockford, IL) for 1 hr. at room
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
temperature. The plates were then washed 5 times and the amount of bound ligand assessed with
1-step Ultra TMB substrate (Pierce Biotechnology, Rockford, IL). The colorimetric reading was
performed on a spectrophotometer (Dynex Technologies, Chantilly, VA). For every saccharide,
each experiment was performed at least 3 times, each time in duplicate.
Direct Binding Assays- A 200 nM solution of sDC-SIGN concentrate was coated onto high
binding 96-well plates as described above. Increasing amounts of purified recombinant ligand
were added and incubated at room temperature for 1 hr. The plates are washed 5 times with
wash buffer and incubated with anti-human Fc monoclonal antibody conjugated with HRP as
above for 1 hr. at room temperature. The plates were washed 5 more times and the binding was
assessed by colorimetry as described above. For each ligand (gp120-Fc, ICAM-2-Fc or ICAM-
3-Fc), each experiment was performed at least 3 times in duplicates. Dissociation constants (KD)
were calculated using GraphPad™ Prism software (San Diego, CA). For the differential lectin
binding experiments, 0.2 µg/ml of the indicated lectins were coated onto streptavidin plates
(Pierce Biotechnology, Rockford, IL) for 2 hrs. Plates were washed 3 times with wash buffer
(PBS supplemented with 0.5% Tween and 2% BSA). Ligands were added and bound at room
temperature for 1 hr., followed by an additional 3 washes. HRP-conjugated goat anti-human Fc
antibodies were added for 30 min. to detect the amount of bound ligand. The activity of HRP
was quantified colorimetrically as described above.
Cell Surface Binding Assays- The B cell line HS Sultan and the monocytic cell line THP-1 stably
expressing DC-SIGN were derived from stable integration of a retroviral vector MIGR1/EGFP
vector containing DC-SIGN (6). 293T cells expressing DC-SIGN were produced by transient
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
CaPO4 transfections with pCDNA3-DCSIGN according to standard protocols (6). The cell-
surface binding reaction was done as described previously (40). The cell surface Kds were
calculated using the GraphPad™ Prism software and were obtained by titrating in the amount of
the indicated ligand and normalizing the highest Mean Fluorescent Intensity (MFI) value
obtained to 100%.
Saccharide Chip Array - 96-well format of mono and oligosaccharides was purchased from
Glycominds (Israel). Binding assays with sDC-SIGN were performed according to
manufacturer’s instructions. Briefly, the 96-well plate was washed 5 times with TBS/0.1%
Tween (wash buffer) and allowed to bind to 20 µl of a 200 nM solution of purified recombinant
His-tagged sDC-SIGN at room temperature for 1 hr. The plate was washed 5 times with wash
buffer and incubated with a mouse anti-DC-SIGN monoclonal antibody (DC028) for 1 hr. at
room temperature. DC028 recognizes the repeat region of DC-SIGN and does not interfere with
DC-SIGN’s ability to recognize glycans on gp120 (10). After 5 additional washes, the bound
DC028 was detected by a goat anti-mouse antibody conjugated with horseradish-peroxidase
(HRP). For increased sensitivity, HRP activity was detected using a chemiluminescence
substrate, SuperSignal ELISA Femto (Pierce Biotechnology, Rockford, IL). The relative light
units were measured using a luminometer (LJL Biosystems, Sunnyvale, CA).
Generation of DC-SIGN Mutants- Site-directed mutagenesis was carried out using the
QuickChange kit (Invitrogen, Carlsbad, CA) according to manufacturer’s instructions. Briefly,
for each mutant, a pair complimentary oligonucleotide primers containing the appropriate base-
pair change is used in a PCR reaction to generate a de novo vector containing the cognate DC-
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
SIGN mutant. Each mutant was verified by DNA sequencing. The expression level of each DC-
SIGN mutant was determined by FACS analysis using a conformationally independent (DC028)
and a conformationally dependent (#507) anti-DC-SIGN mouse monoclonal antibody (10).
RESULTS
Recombinant soluble DC-SIGN oligomerizes similarly to DC-SIGN on dendritic cells. To
delineate the differences in the binding affinities of gp120, ΙCAM-2 and ΙCAM-3 to DC-SIGN,
we first measured the dissociation constant (Kd) of the three ligands in an immobilized solid-
phase ELISA format. To this end, we expressed the extracellular domain of DC-SIGN (sDC-
SIGN) as an N-terminal 6X-Histidine-tagged fusion protein. To determine if sDC-SIGN
multimerizes into tetramers as has been previously demonstrated (39), we fractionated the
recombinant protein on a 5-20% sucrose gradient. As can seen in Figure 1, purified recombinant
sDC-SIGN readily formed trimers and tetramers in agreement with previous biochemical and
physical studies (39). Importantly, the higher ordered oligomers of purified sDC-SIGN
corresponded approximately to the oligomeric state of full-length DC-SIGN proteins isolated
from immature monocyte-derived dendritic cells (Fig. 1). To our knowledge, this is the first
demonstration that full-length DC-SIGN on primary dendritic cells can also exist in the
oligomeric state found for recombinant sDC-SIGN.
gp120 binds to DC-SIGN with greater affinity than ΙΙΙΙ CAM-2 and ΙΙΙΙCAM-3. In order to facilitate
our binding studies, HIV-1 gp120 was produced as an IgG1-Fc fusion protein (40). Using
purified sDC-SIGN, we found that gp120-Fc bound to sDC-SIGN with 100- and 50-fold higher
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

11
affinity than ΙCAM-2-Fc and ΙCAM-3-Fc, respectively (Fig. 2A). We then asked if DC-SIGN’s
differential affinity for the three ligands was maintained when DC-SIGN was expressed on the
cell surface. We chose three cell lines (THP-1, HS-Sultan and 293T HEK cells) with no
appreciable binding to gp120, ICAM-2 and ICAM-3 in the absence of DC-SIGN (ref. 2 and data
not shown). THP-1 and HS-Sultan cells were retrovirally transduced and clones were isolated
that stably expressed DC-SIGN at similar levels. HS Sultan is a mature B-cell line and the
functionality of DC-SIGN on B cells has not been previously established. 293T HEK cells were
transiently transfected with DC-SIGN. In all three cell lines, much like the in vitro binding
studies, gp120-Fc bound to DC-SIGN with much greater affinity than ΙCAM-2-Fc and ΙCAM-3-
Fc (Fig. 2B). However, the relative differences in binding affinity between ΙCAM-2-Fc and
ΙCAM-3-Fc to DC-SIGN were cell-type dependent. For example, while ICAM-2 (Kd=6.66 +
0.88 nM) bound better than ICAM-3 (Kd=17.0 + 1.74 nM) on THP-1 DC-SIGN+ cells, it bound
less avidly than ICAM-3 on HS-Sultan DC-SIGN+ cells (Kd=57.9 + 4.09 nM vs 13.92 + 1.38
nM, respectively). While gp120 showed the highest affinity binding to all 3 cell lines, gp120
bound to HS Sultan DC-SIGN+ (Kd, 0.017 nM) cells 4-fold better than to 293T DC-SIGN+ (Kd,
0.078 nM) transfectants and the THP-1 DC-SIGN+ (Kd, 0.068 nM) cells (Fig. 2B). These data
suggest that the ligand binding behavior of DC-SIGN may be modulated in a cell-type specific
manner although we cannot formally exclude interactions of gp120, ΙCAM-2 and ΙCAM-3 with
other cell surface proteins.
Recent studies have noted that the efficiency of DC-SIGN’s ability to transfer virus to
permissive T-cells varies depending on the cell-type on which DC-SIGN is expressed (44).
Since HS Sultan DC-SIGN+ cells bound gp120 with greater affinity than THP-1 DC-SIGN+ cells
and HS Sultan cells also belong to an antigen-presenting cell lineage (as do dendritic cells and
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12
THP-1 cells), we tested whether HS-Sultan DC-SIGN+ cells would transfer HIV better than
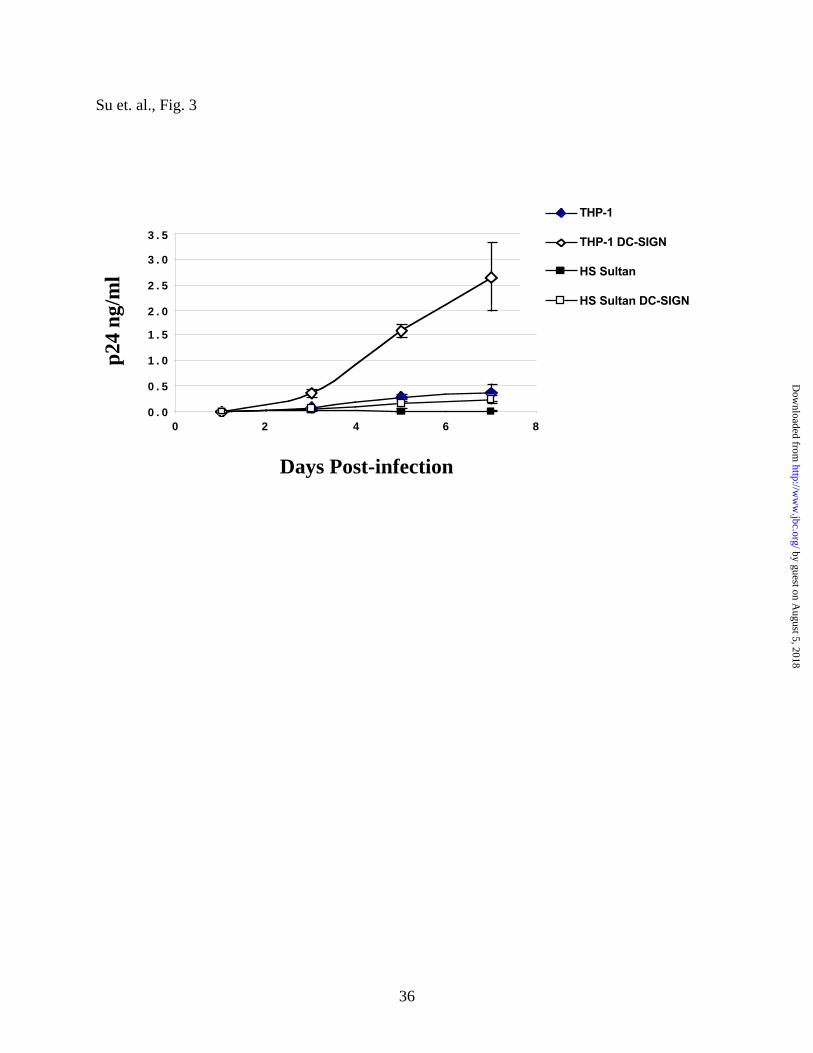
THP-1 DC-SIGN+ cells. As seen in Fig. 3, HS Sultan DC-SIGN+ cells were extremely
inefficient in transferring HIV-1 to permissive T-cells (compare with THP-1 DC-SIGN+ cells in
Fig. 3). This suggest that viral binding and transfer are dissociable functions of DC-SIGN, and
that cell-specific co-factors other than those that contribute to the cell surface affinity for gp120
are likely to contribute to DC-SIGN’s ability to facilitate infection in trans.
ΙΙΙΙCAM-2 and ΙΙΙΙCAM-3 contain less high-mannose oligosaccharides than gp120. Since DC-
SIGN is a mannose-specific lectin, one possible reason for the poor affinity of ΙCAM-2 and
ΙCAM-3 for DC-SIGN compared with gp120 is that ΙCAM-2 and ΙCAM-3 have fewer high
mannose oligosaccharides compared to gp120. Although gp120 has a larger number of potential
N-linked glycosylation sites than ICAM-2 and ICAM-3, the type of glycans (complex/hybrid
versus high mannose) on each site can only be determined empirically, Therefore, we used
different lectins of various specificities to determine the relative amounts of the major types of
glycans on these three ligands. In an equilibrium binding assay, we found that Galanthus nivalis
GNA bound to gp120 (Kd =0.15 mM) with about 150-fold higher affinity compared to ICAM-2
(Kd =22.7 mM) and ΙCAM-3 (Kd =20.4 mM), respectively (Fig. 4). Since GNA is a lectin that
reacts most strongly with multiple terminal α(1,3)mannose residues (45), the data showed that
gp120 contained more high-mannose type carbohydrates than ΙCAM-2 and ΙCAM-3. We also
found that Datura stramonium DSA bound to gp120 (Kd=0.74 mM) with a higher affinity than
either ΙCAM-2 (Kd=76.3 mM) or ΙCAM-3 (Kd=58.7 mM) (Fig. 4). On mammalian cells, DSA
binds to terminal N-acetyl-D-glucosamine (GlcNAc) moieties if they are not masked by
galactose or sialic acid. Thus, gp120 may contain more terminal GlcNAc residues compared to
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

13
ICAM-2 or ICAM-3. In toto, the lectin binding profiles suggest that gp120 is glycosylated
differently from ICAM-2 and ICAM-3 but that the similar Kds of ICAM-2 and ICAM-3 for GNA
and DSA, respectively, indicate that ICAM-2 and ICAM-3 are glycosylated similarly.
Glycan specificities of DC-SIGN-mediated binding. To further assess whether there are any
differences in the type of glycans on gp120, ΙCAM-2 and ΙCAM-3 that mediate DC-SIGN
binding, we used various monosaccharides and disaccharides to compete for ligand binding to
immobilized sDC-SIGN. Monosaccharides with equatorial 4-hydroxyl groups (e.g. mannose,
mannose derivatives, glucose and fucose) preferentially competed for gp120 binding to DC-
SIGN over galactose and galactosides, which have axial 4-hydroxyl groups (Table 1; Fig 5).
gp120 and ΙCAM-3 binding to DC-SIGN were effectively competed by both D-mannose (Ki,
5.6 mM and 1.6 mM, respectively) and D-glucose (Ki, 8.08 mM and 2.35 mM, respectively) but
not D-galactose (Ki, 63.56 mM and 68.83 mM, respectively), confirming DC-SIGN’s relative
preference for binding to equatorial 4-OH groups (Table I). As a negative control, L-mannose,
the stereoisomer of D-mannose, failed to be an effective competitor for gp120 and ICAM-3
binding (see Table I and Fig. 5A). This preference for a free equatorial 4-hydroxyl group closely
mimics another mannose specific C-type lectin, macrophage mannose receptor (46).
For gp120, DC-SIGN binding was also preferentially competed by oligosaccharides
since the disaccharide maltose Gluc(α1,4)Gluc (Ki, 0.27 mM),was a much better competitor than
the monosaccharide D-glucose (Table 1; Fig. 5B). Finally, using disaccharide competitors that
differ only in the alpha- or beta- positioning of their glycosidic linkage, such as maltose
(Gluc(α1,4)Gluc) and its anomer cellobiose (Gluc(β1,4)Gluc) (Ki, 1.43 mM), we found that DC-
SIGN may prefer to bind oligosaccharides linked in an alpha-anomeric configuration (Table I,
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14
Maltose Ki=0.2 mM; Cellobiose Ki=1.4 mM). This preference was later confirmed by saccharide
chip analysis (see below and Table II). We understand that dissacharides of glucose are not
found in mammalian systems, and therefore may not represent a physiological ligand for DC-
SIGN. However, since our data suggest that there is no significant difference between D-
mannose, L-fucose and D-glucose binding to DC-SIGN (Table I), and that both competition and
direct binding assays suggest a preference for an alpha-anomeric configuration in glucose based
disaccharides, we believe that this preference will hold in more detailed future studies using
oligomers of mannose.
Like gp120, D-mannose best competed for ΙCAM-2 and ΙCAM-3 binding to sDC-SIGN
(Table I). Since it took less D-mannose sugars to compete for the binding of the two endogenous
ligands than for gp120, the data suggests that there are less D-mannose sugars on ΙCAM-2 and
ΙCAM-3 than gp120, consistent with the differential lectin binding data shown in Figure 4. The
monosaccharide competition data of ΙCAM-3 closely followed the trend of gp120, suggesting
that DC-SIGN binding to ΙCAM-3 was also dependent on the equatorial 4-OH group. However,
unlike gp120, maltose (Ki, 13.0 mM) competed approximately 8-fold and 6-fold less well than
D-mannose (Ki, 1.6 mM) and D-glucose (Ki, 2.3 mM), respectively (Table I). It is plausible that
the binding of sDC-SIGN on the sparsely mannosylated ΙCAM-3 is unsaturated and more
flexible via the widely spaced terminal mannose moieties. Consequently, more maltose is
needed to saturate all the binding sites in the DC-SIGN oligomer before it is able to fully
compete for the sites bound by ΙCAM-3. Similarly, the disaccharide maltose also did not
compete more effectively than the cognate monosaccharides for DC-SIGN binding to ΙCAM-2.
However, it is peculiar that D-glucose is such a poor competitor of ΙCAM-2 binding since both
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

15
competition experiments with gp120 and ΙCAM-3 show that D-glucose is as good a competitor
as D-mannose (Table I). This suggests that DC-SIGN binding to ICAM-2 involves unique
elements, which may include protein-protein in addition to protein-carbohydrate interactions.
Saccharide chip analysis. To further gain insights into DC-SIGN's glycan binding specificities,
we employed a saccharide micro-array analysis of mono and oligosaccharides displayed in a 96-
well ELISA plate format. The carbohydrates were covalently coupled via a flexible linker to the
surface, and this technology has been used to profile the saccharide binding specificities of
carbohydrate binding proteins such as lectins and antibodies (47-49). Here, we used sDC-SIGN
to bind to the immobilized saccharides in an attempt to characterize DC-SIGN’s glycan binding
specificities. These direct binding studies are consistent with the competition data presented in
Fig. 5 and Table I. Namely, sDC-SIGN does not bind to any galactose, galactosides or any
various galactose linked to another sugar (Table II and data not shown). Rather, sDC-SIGN
exhibited the greatest binding to glucose and mannose moieties, showing a strict specificity for
the 4 -OH group in an equatorial position (data not shown and ref (39)). In this assay, sDC-SIGN
also tends to prefer glycosidic bonds linked in the alpha anomeric configuration (Table II),
consistent with the competition data shown in Table I.
Mutational analysis of DC-SIGN. The competition and binding data suggest that DC-SIGN may
bind differentially to gp120, ICAM-2 and ICAM-3. Therefore, we next sought to determine if
specific residues on DC-SIGN diferentially contribute to gp120, ΙCAM-2 and ΙCAM-3 binding.
To this end, we generated alanine scan mutants that correspond to most of the solvent exposed
amino acids implicated in the maintenance of calcium co-ordination or carbohydrate contacts in
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

16
the crystal structure of DC-SIGN’s CRD (Fig. 6A). The sixteen different mutations created
represent various degrees of conservation amongst C-type mannose specific lectins. All alleles
were expressed in the HEK 293T cell line and ligand binding was done similarly to that noted in
Figure 2B. A panel of conformation-independent and conformation-dependent anti-DC-SIGN
monoclonal antibodies was used to assess the cell surface expression level and gross structural
integrity of each DC-SIGN mutant. mAB 507 is a conformation-dependent antibody directed
against the CRD and has the ability to block gp120 binding to DC-SIGN (50) although its exact
epitope is not known. mAB DC028 (10) is a conformation-independent antibody directed
against the repeat domain of DC-SIGN, and should not be directly affected by mutations in the
CRD. As seen in Figure 6B, the expression of wild-type DC-SIGN and its alleles was compared
by normalizing the mean channel fluorescence obtained for each allele (and for each antibody) to
the wild-type expression, which was set at 100%. mAb DC028 will necessarily measure the
absolute amounts of cell surface DC-SIGN whereas mAb 507, whose epitope is unknown could
potentially be affected by a particular mutation. Therefore, the gp120, ICAM-2 and ICAM-3
binding to DC-SIGN and its alleles were normalized to the expression level as determined by
mAb DC028 staining (Fig. 6C). In addition, since all the alleles except for P348A were
recognized by mAB 507 at 50% or more of wild-type level, we believe there none of these
mutants exhibit gross perturbation of DC-SIGN’s stucture. We believe the decreased expression
of P348A is due to the absolute decrease in cell surface DC-SIGN as both DC028 and 507
staining revealed the same percent decrease in expression (Fig. 6B).
The observed phenotypes of the 16 mutants fall into several classes. Firstly, as expected,
mutations in residues highly conserved in all C-type lectins, E354, D355 and D366, severely
compromised binding to all three ligands. E354 coordinates with calcium and interacts with 4-
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

17
OH of mannose in the cocrystal structures of an oligosaccharide with both DC-SIGN and DC-
SIGNR(41). The amino acid D366 of DC-SIGN coordinates extensively with the calcium ion
only. Residues E347, P348 and N349 represent the tripeptide sequence that is a highly conserved
signature motif for mannose-specific lectins. Both E347 and N349 residues make extensive
contacts with the calcium ion and 3-OH of mannose as observed in the DC-SIGN
CRD/oligosaccharide cocrystal structure. Interestingly, mutating E347, P348 and N349 to
alanine reduced gp120 binding much more substantially than ICAM-2 and ICAM-3 binding.
Notably, the P348A mutant appeared to exhibit enhanced binding to the two adhesion molecules
(Fig. 6C) although this could be an artificial effect of normalization as P348A was the only
mutant that was expressed at significantly lower levels than all the other alleles (Fig. 6B). In
sum, mutagenesis of these conserved residues suggests that the primary binding determinants of
all three ligands to DC-SIGN was calcium and mannose-dependent to a certain degree but also
that DC-SIGN binds to gp120 in a similar but distinct fashion from ICAM-2 and ICAM-3.
Mutations of amino acids less conserved in C-type mannose lectins show a wide range of
informative phenotypes. Mutating residues N311, R345, V351, S360, G361, and N362 to
alanines showed only a minimal effect on binding to all three ligands (Fig. 6C). Some of these
results were somewhat surprising. For example, N311 was postulated in the cocrystal structure
to, alongside F313, form a fitting groove for high mannose recognition that specifically
discriminates binding against the inner branch point mannose (41) (Fig. 7D), and S360
maintains extensive contacts with Man3 and Man4 in the pentasaccharide co-crystalized with
DC-SIGN’s CRD. On the other hand, the mutants G346A and E353A diminished binding to
gp120 by at least 50%, while not having a negative effect on ICAM-2 and ICAM-3 binding.
Indeed, the G346A mutation appeared to enhance ICAM-2 and ICAM-3 binding by up to 2-fold.
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

18
Surprisingly, G346 and E353 did not have direct interactions with the pentasaccharide moiety in
the co-crystal structure with the CRD of DC-SIGN. Once again, these data suggest that DC-
SIGN binding to the two endogenous ligands (ICAM-2 and ICAM-3) may involve unique
elements from that required for gp120 binding, and that our mutagenic analysis using the
“natural”ligands of DC-SIGN can provide information about the binding behavior of DC-SIGN
that is not immediately obvious even from structural data.Finally, the mutant D367A diminished
ΙCAM-2 and ΙCAM-3 binding by about 50% while enhancing gp120 binding by up to 2-fold,
suggesting yet again that DC-SIGN binding to gp120 is qualitatively different from its binding to
ICAM-2 and ICAM-3.
DISCUSSION
The present study provides evidence for the molecular determinants of DC-SIGN binding
to a viral ligand, gp120, and its two endogenous ligands, ΙCAM-2 and ΙCAM-3. Firstly, using
purified components in an in vitro equilibrium binding assay, we showed that gp120 binds to
sDC-SIGN with nanomolar affinity, 100- and 50-fold better than ΙCAM-2 and ΙCAM-3,
respectively. When DC-SIGN is expressed on the cell surface, gp120 still bound with relatively
much greater affinity than ΙCAM-2 and ΙCAM-3 on all three cell types examined. It is
interesting to note that the affinity is much higher when binding is done on cell surface expressed
DC-SIGN. This difference in affinity may be due to cellular factor(s) affecting the binding
behavior of DC-SIGN and/or the presence of other cognate binding partners that contribute to the
total cell surface affinity of examined ligands. We note, however, that all three cell lines were
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

19
CD4-negative and lack appreciable gp120, ICAM-2 and ICAM-3 binding activity in the absence
of DC-SIGN expression.
We also note that the cell surface affinity of DC-SIGN for gp120 does not correlate with
its ability to transfer HIV-1 to permissive T cells. It has been reported that the capacity of DC-
SIGN to transmit HIV to permissive T-cells is greater when it is expressed in THP-1 versus 293
cells (44), suggesting the presence of contributory cellular factors to DC-SIGN’s ability to
facilitate infection in trans. Here, we show that the difference in transmission between these cell
lines was not due to the cell surface affinity for gp120 since the measured Kd for the two cell
types were similar. Intriguingly, we also show that even on an antigen presenting cell line (HS-
Sultan) with a 4-fold higher affinity for gp120, DC-SIGN was still not able to facilitate virus
infection in trans. For the moment, it appears that DC-SIGN’s ability to transfer virus is
restricted to a cellular factor(s) common to dendritic cells and THP-1 cells.
Our competition data provide further insights into the molecular specificities of DC-
SIGN glycan binding. Unlike the rat mannose binding protein (MBP) (51,52) and the
macrophage mannose receptor (MR) (46), structural and functional data all support the notion
that DC-SIGN prefers to bind mannose or glucose-derived oligosaccharides linked in an alpha
anomeric configuration. Since our data indicate that gp120 contains more high-mannose type
sugars than ICAM-2 and ICAM-3, and it has been previously shown that most of the high-
mannose moieties in gp120 are linked in the alpha anomeric configuration (53-55), it is therefore
not surprising that DC-SIGN binds with so much higher affinity to gp120 than to ΙCAM-2 or
ΙCAM-3 (Fig. 2). Our competition data also indicates that DC-SIGN has a relatively high
affinity for L-fucose. This is consistent with published reports of DC-SIGN’s ability to bind to
Lewis blood group antigens (22), which are composed of glycosphingolipids, largely defined by
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

20
a difference in the position of the linkage following a difference in the number of fucose sugars
present (56). Indeed, DC-SIGN specifically binds the bacterium H. pylori (22) and the parasitic
worm S. mansoni (24) both of which express the Lewis x antigen.
However, it has become increasingly evident that discerning DC-SIGN’s glycan binding
specificities is a bit more complex with respect to what glycans it is predicted to bind versus the
actual glycan moieties it does bind on its “natural” ligands. Namely, although DC-SIGN seems
to bind to the vector-borne parasite Leishmania pifanoi, the interaction is not inhibitable by
mannan (23), which to date has inhibited DC-SIGN binding to all known ligands. It is possible
that DC-SIGN binds via another high affinity glycan found on L. pifanoi that is yet to be
characterized, or that protein-protein interactions are actually involved in binding. Also, our data
showed that maltose is a highly effective inhibitor of gp120 binding to DC-SIGN suggesting that
dimannosides would be as good or better ligands for DC-SIGN. Yet, synthetic glycoclusters
harboring dimannoside substrates failed to bind to cells expressing DC-SIGN (57). Nonetheless,
convincing evidence shows that the binding of DC-SIGN to M. tuberculosis is due to the
dimannoside, Man(α1,2)Man, cap of the lipoglycan lipoarabinomannan (ManLam) (25,27). The
binding of this native substrate ManLam is specific to the dimannoside cap since AraLam
(ManLam devoid of the mannose cap) fails to bind to DC-SIGN, and enzymatic cleavage of the
dimannosides from ManLam abrogated its ability to compete for DC-SIGN binding to M.
tuberculosis. Thus, it prudent to bear in mind the difference between what ligands DC-SIGN can
or cannot bind when presented as neo-glycoconjugates versus what ligands it does bind when
presented on a biologically relevant molecule.
Our alanine-scanning mutagenesis of most of the solvent exposed amino acids of DC-
SIGN CRD reveals several insights into DC-SIGN function. Using the schematic presented in
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

21
Feinberg et al. (41), (Fig. 7A) we have mutations at the highly conserved end of the
“oligosaccharide binding valley” as exemplified by residues E347, N349 E354, E355 and D366
that significantly diminished binding to all three ligands presumably by destroying calcium
coordination and hydrogen bonding to the 4-OH of Man2 (Fig. 7B). It is most likely that this
one end of the valley contributes the greatest amount of binding energy to the DC-SIGN-ligand
interaction since the consensus motif for all mannose-specific and C-type lectins resides here
(Fig. 6A and Fig. 7B). The binding phenotype of P348A mutant is interesting since the
homologous residue in rat serum mannose-binding protein (MBP) has been shown to undergo
cis-trans isomerization in the presence and absence of calcium, respectively (58). In rat serum
MBP, the peptide bond of the highly conserved glutamic acid preceding proline adopts a cis
conformation in the presence of calcium; and this is absolutely required in positioning critical
amino acids, such as the highly conserved residue E354 in loop 4, for ligation of calcium ions
and subsequent substrate binding. In the absence of calcium, the peptide bond adopts a trans
conformation, and causes dramatic conformational changes in loops 3 and 4 that consequently
move side chain atoms by as much as 12 Å(58). Since the P348A mutant affected gp120 binding
to DC-SIGN much more significantly than ICAM-2 or ICAM-3 binding, we speculate that
gp120's interactions with DC-SIGN occur in a more calcium and mannose-dependent fashion.
The same mutant appears to enhance ICAM-2 and ICAM-3 binding when corrected for
expression levels, suggesting that DC-SIGN’s interaction with these adhesion molecules may
involve unique elements in addition to protein-carbohydrate contacts.
Moving down towards the other end (flanked by residues N311 and F313, see Fig. 7D) in
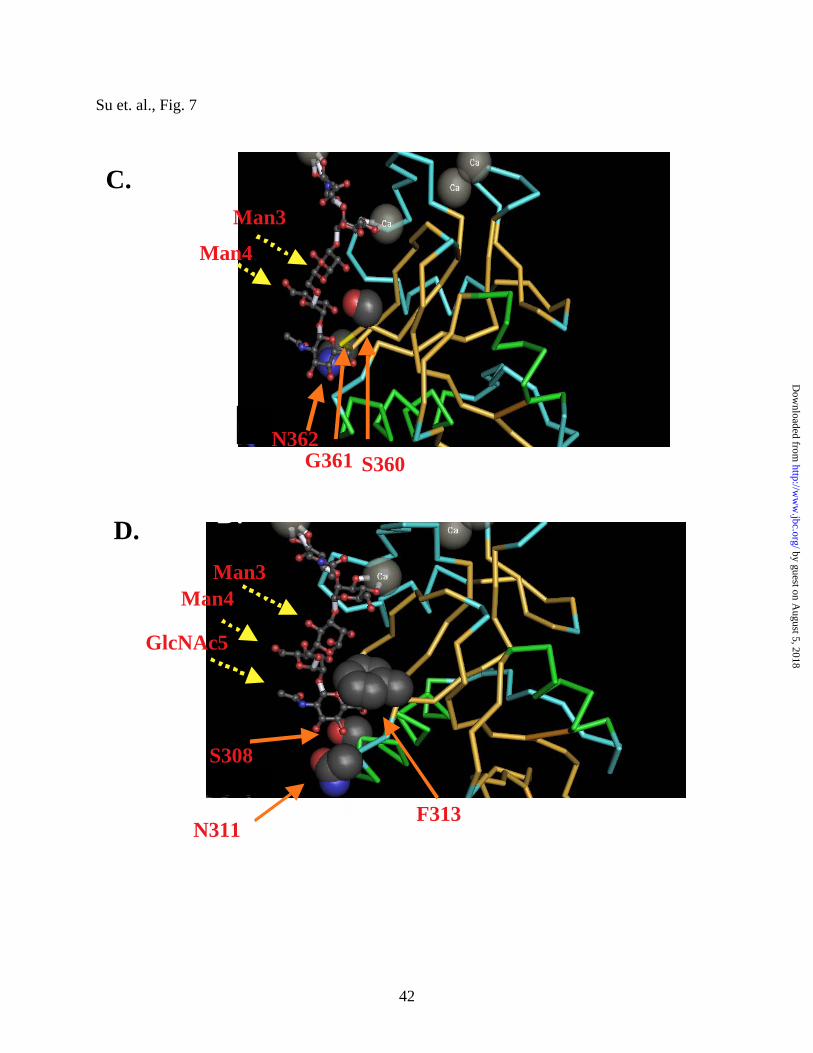
the valley of the binding groove, we found three alanine mutants S360A, G361A and N362A that
have minimal effect on DC-SIGN’s binding to all three ligands (Fig. 6C). This was surprising
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

22
since in the co-crystal structure, these residues exhibit numerous 3-OH and water-hydrogen
bonding with the pentasaccharide (Fig. 7C) (41). At best, ICAM 2 binding to the S360A mutant
was decreased by about 30%. Amino acid S360, which packs closely against F313, is interesting
since its interactions with two sugar molecules seem to allow for the proposed discriminatory
role of F313. It is possible that other interactions mediated by the dense clusters of cognate
glycans in the highly mannosylated gp120 can compensate for the slight loss of binding
contributed by an “accessory” residue such as S360. We speculate that the sparsely
mannosylated ΙCAM-3 and ΙCAM-2 may be more sensitive to these mutations and have less
“carbohydrate reserve” to compensate for the loss of such binding energy (especially with the
S360A mutant). Lastly, the crystal structure of DC-SIGN predicts that N311 and F313 play
important roles in forming the other end of the binding pocket and contribute to the ligand
specificity of DC-SIGN by favoring the binding to the outer branch mannose as a result of the
steric clash impose by F311 on the beta anomeric linkage of the inner trimannose branch point
(41). Our data indicate that the N311A mutant still binds to all three ligands, suggesting that
F313 may play the dominant critical role in high mannose binding of this end of the pocket.
Further mutational analysis at this end of the oligomannose binding valley will determine if this
hypothesis is true for which specific glycoprotein ligand.
It is interesting to note that D367A mutant abolishes binding to ΙCAM-2 and ΙCAM-3 but
enhances gp120 binding. In the crystal structure, D367 resides in the highly conserved end of
the pocket and makes extensive water-mediated bonding with the 6th carbon hydroxyl group of
Man2. The mutant suggests that D367 is important for ΙCAM-2 and ΙCAM-3 since substituting
alanine at this position reduced binding by more than 50% and also suggests that D367 may
partially obscure more important interactions with gp120 (as changing this residue to alanine
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

23
enhances gp120 binding). Conversely, the G346A mutant almost abrogated gp120 binding while
enhancing ICAM-2 and ΙCAM-3 binding, suggesting that ΙCAM-2 and ICAM-3 binding to DC-
SIGN may be qualitatively different from gp120. This same residue in DC-SIGNR, a
homologue of DC-SIGN, makes water-mediated contacts with the 6th carbon hydroxyl group and
may be important for saccharide binding. This observation, coupled with the fact that D-glucose
competes more than 100-fold less efficiently than D-mannose for ΙCAM-2-Fc binding to DC-
SIGN, argues once again that ΙCAM-2 and ICAM-3 binding to DC-SIGN may involve
protein/protein in addition to protein-carbohydrate interactions. It is not uncommon that lectins
bind to both saccharide and protein determinants concurrently (37,38). Further experimentation
is warranted to determine the validity of this hypothesis.
In this study, we show that DC-SIGN binds to HIV-1 gp120, ICAM-2 and ICAM-3 in a
distinct but overlapping fashion. We also provide biochemical and genetic evidence that DC-
SIGN binding to ICAM-2 is qualitatively different from DC-SIGN binding to gp120 and ΙCAM-
3. Importantly, we have identified residues that differentially contribute to binding all three
ligands. Delineation of such differences may be useful for future therapeutic intervention
targeting the gp120-DC-SIGN interface.
Acknowledgements
We would like to thank Linda Baum for her critical review of this manuscript. We would
like to thank Kurt Drickamer for his initial gift of recombinant soluble DC-SIGN and Sophia
Young for her technical assistance in making recombinant soluble DC-SIGN in the Lee Lab.
B.L. is a Charles E. Culpepper Medical Scholar supported by the Rockefeller Brothers Fund and
a recipient of the Burroughs Wellcome Fund Career Development Award, and is supported by
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

24
NIH grants RO1-AI52021 and R21-AI055305. We also acknowledge support of the UCLA
AIDS Institute and the flow cytometry core (UCLA CFAR grant, NIH AI-28697), and the James
B. Pendleton Charitable Trust.
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

25
REFERENCES
1. Soilleux, E. J., Morris, L. S., Leslie, G., Chehimi, J., Luo, Q., Levroney, E., Trowsdale,J., Montaner, L. J., Doms, R. W., Weissman, D., Coleman, N., and Lee, B. (2002) JLeukoc Biol 71, 445-457
2. Geijtenbeek, T. B., Kwon, D. S., Torensma, R., van Vliet, S. J., van Duijnhoven, G. C.,Middel, J., Cornelissen, I. L., Nottet, H. S., KewalRamani, V. N., Littman, D. R., Figdor,C. G., and van Kooyk, Y. (2000) Cell 100, 587-597
3. Jameson, B., Baribaud, F., Pohlmann, S., Ghavimi, D., Mortari, F., Doms, R. W., andIwasaki, A. (2002) J Virol 76, 1866-1875
4. Curtis, B. M., Scharnowske, S., and Watson, A. J. (1992) Proc Natl Acad Sci U S A 89,8356-8360
5. Geijtenbeek, T. B., and van Kooyk, Y. (2003) Curr Top Microbiol Immunol 276, 31-546. Lee, B., Leslie, G., Soilleux, E., O'Doherty, U., Baik, S., Levroney, E., Flummerfelt, K.,
Swiggard, W., Coleman, N., Malim, M., and Doms, R. W. (2001) J Virol 75, 12028-12038
7. Clapham, P. R., and McKnight, A. (2001) Br Med Bull 58, 43-598. Baribaud, F., Pohlmann, S., and Doms, R. W. (2001) Virology 286, 1-69. Schwartz, A. J., Alvarez, X., and Lackner, A. A. (2002) AIDS Res Hum Retroviruses 18,
1021-102910. Baribaud, F., Pohlmann, S., Sparwasser, T., Kimata, M. T., Choi, Y. K., Haggarty, B. S.,
Ahmad, N., Macfarlan, T., Edwards, T. G., Leslie, G. J., Arnason, J., Reinhart, T. A.,Kimata, J. T., Littman, D. R., Hoxie, J. A., and Doms, R. W. (2001) J Virol 75, 10281-10289
11. Gardner, J. P., Durso, R. J., Arrigale, R. R., Donovan, G. P., Maddon, P. J., Dragic, T.,and Olson, W. C. (2003) Proc Natl Acad Sci U S A 100, 4498-4503
12. Lozach, P. Y., Lortat-Jacob, H., de Lacroix de Lavalette, A., Staropoli, I., Foung, S.,Amara, A., Houles, C., Fieschi, F., Schwartz, O., Virelizier, J. L., Arenzana-Seisdedos,F., and Altmeyer, R. (2003) J Biol Chem 278, 20358-20366
13. Pohlmann, S., Zhang, J., Baribaud, F., Chen, Z., Leslie, G. J., Lin, G., Granelli-Piperno,A., Doms, R. W., Rice, C. M., and McKeating, J. A. (2003) J Virol 77, 4070-4080
14. Baribaud, F., Pohlmann, S., Leslie, G., Mortari, F., and Doms, R. W. (2002) J Virol 76,9135-9142
15. Alvarez, C. P., Lasala, F., Carrillo, J., Muniz, O., Corbi, A. L., and Delgado, R. (2002) JVirol 76, 6841-6844
16. Lasala, F., Arce, E., Otero, J. R., Rojo, J., and Delgado, R. (2003) Antimicrob AgentsChemother 47, 3970-3972
17. Lin, G., Simmons, G., Pohlmann, S., Baribaud, F., Ni, H., Leslie, G. J., Haggarty, B. S.,Bates, P., Weissman, D., Hoxie, J. A., and Doms, R. W. (2003) J Virol 77, 1337-1346
18. Simmons, G., Reeves, J. D., Grogan, C. C., Vandenberghe, L. H., Baribaud, F.,Whitbeck, J. C., Burke, E., Buchmeier, M. J., Soilleux, E. J., Riley, J. L., Doms, R. W.,Bates, P., and Pohlmann, S. (2003) Virology 305, 115-123
19. Halary, F., Amara, A., Lortat-Jacob, H., Messerle, M., Delaunay, T., Houles, C., Fieschi,F., Arenzana-Seisdedos, F., Moreau, J. F., and Dechanet-Merville, J. (2002) Immunity 17,653-664
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

26
20. Navarro-Sanchez, E., Altmeyer, R., Amara, A., Schwartz, O., Fieschi, F., Virelizier, J. L.,Arenzana-Seisdedos, F., and Despres, P. (2003) EMBO Rep 4, 723-728
21. Tassaneetrithep, B., Burgess, T. H., Granelli-Piperno, A., Trumpfheller, C., Finke, J.,Sun, W., Eller, M. A., Pattanapanyasat, K., Sarasombath, S., Birx, D. L., Steinman, R.M., Schlesinger, S., and Marovich, M. A. (2003) J Exp Med 197, 823-829
22. Appelmelk, B. J., van Die, I., van Vliet, S. J., Vandenbroucke-Grauls, C. M.,Geijtenbeek, T. B., and van Kooyk, Y. (2003) J Immunol 170, 1635-1639
23. Colmenares, M., Puig-Kroger, A., Pello, O. M., Corbi, A. L., and Rivas, L. (2002) J BiolChem 277, 36766-36769
24. van Die, I., van Vliet, S. J., Nyame, A. K., Cummings, R. D., Bank, C. M., Appelmelk,B., Geijtenbeek, T. B., and van Kooyk, Y. (2003) Glycobiology 13, 471-478
25. Maeda, N., Nigou, J., Herrmann, J. L., Jackson, M., Amara, A., Lagrange, P. H., Puzo,G., Gicquel, B., and Neyrolles, O. (2003) J Biol Chem 278, 5513-5516
26. Tailleux, L., Schwartz, O., Herrmann, J. L., Pivert, E., Jackson, M., Amara, A., Legres,L., Dreher, D., Nicod, L. P., Gluckman, J. C., Lagrange, P. H., Gicquel, B., andNeyrolles, O. (2003) J Exp Med 197, 121-127
27. Geijtenbeek, T. B., Van Vliet, S. J., Koppel, E. A., Sanchez-Hernandez, M.,Vandenbroucke-Grauls, C. M., Appelmelk, B., and Van Kooyk, Y. (2003) J Exp Med197, 7-17
28. Geijtenbeek, T. B., Krooshoop, D. J., Bleijs, D. A., van Vliet, S. J., van Duijnhoven, G.C., Grabovsky, V., Alon, R., Figdor, C. G., and van Kooyk, Y. (2000) Nat Immunol 1,353-357
29. Geijtenbeek, T. B., van Duijnhoven, G. C., van Vliet, S. J., Krieger, E., Vriend, G.,Figdor, C. G., and van Kooyk, Y. (2002) J Biol Chem 277, 11314-11320
30. Geijtenbeek, T. B., Torensma, R., van Vliet, S. J., van Duijnhoven, G. C., Adema, G. J.,van Kooyk, Y., and Figdor, C. G. (2000) Cell 100, 575-585
31. Weis, W. I., Taylor, M. E., and Drickamer, K. (1998) Immunol Rev 163, 19-3432. Zelensky, A. N., and Gready, J. E. (2003) Proteins 52, 466-47733. Drickamer, K. (1999) Curr Opin Struct Biol 9, 585-59034. Natarajan, K., Dimasi, N., Wang, J., Mariuzza, R. A., and Margulies, D. H. (2002) Annu
Rev Immunol 20, 853-88535. Sano, H., Kuroki, Y., Honma, T., Ogasawara, Y., Sohma, H., Voelker, D. R., and Akino,
T. (1998) J Biol Chem 273, 4783-478936. Geider, S., Baronnet, A., Cerini, C., Nitsche, S., Astier, J. P., Michel, R., Boistelle, R.,
Berland, Y., Dagorn, J. C., and Verdier, J. M. (1996) J Biol Chem 271, 26302-2630637. Kijimoto-Ochiai, S. (2002) Cell Mol Life Sci 59, 648-66438. Matsumoto, N., Ribaudo, R. K., Abastado, J. P., Margulies, D. H., and Yokoyama, W. M.
(1998) Immunity 8, 245-25439. Mitchell, D. A., Fadden, A. J., and Drickamer, K. (2001) J Biol Chem 276, 28939-2894540. Hong, P. W., Flummerfelt, K. B., de Parseval, A., Gurney, K., Elder, J. H., and Lee, B.
(2002) J Virol 76, 12855-1286541. Feinberg, H., Mitchell, D. A., Drickamer, K., and Weis, W. I. (2001) Science 294, 2163-
216642. Lue, J., Hsu, M., Yang, D., Marx, P., Chen, Z., and Cheng-Mayer, C. (2002) J Virol 76,
10299-10306
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

27
43. Gurney, K., Elloitt, J., Nassanian, H., Song, S., MacGowan, I., Anton, P., and Lee, B.(2003) Keystone Symposia, abstract no. 221
44. Trumpfheller, C., Park, C. G., Finke, J., Steinman, R. M., and Granelli-Piperno, A.(2003) Int Immunol 15, 289-298
45. Hester, G., Kaku, H., Goldstein, I. J., and Wright, C. S. (1995) Nat Struct Biol 2, 472-47946. Zamze, S., Martinez-Pomares, L., Jones, H., Taylor, P. R., Stillion, R. J., Gordon, S., and
Wong, S. Y. (2002) J Biol Chem 277, 41613-4162347. Nimrichter, L., Gargir, A., Gortler, M., Altstock, R. T., Shtevi, A., Weisshaus, O., Fire,
E., Dotan, N., and Schnaar, R. L. (2003) Glycobiology48. Schwarz, M., Spector, L., Gargir, A., Shtevi, A., Gortler, M., Altstock, R. T., Dukler, A.
A., and Dotan, N. (2003) Glycobiology 13, 749-75449. Dove, A. (2001) Nat Biotechnol 19, 913-91750. Wu, L., Martin, T. D., Vazeux, R., Unutmaz, D., and KewalRamani, V. N. (2002) J Virol
76, 5905-591451. Weis, W. I., Drickamer, K., and Hendrickson, W. A. (1992) Nature 360, 127-13452. Ng, K. K., Drickamer, K., and Weis, W. I. (1996) J Biol Chem 271, 663-67453. Sanders, R. W., Venturi, M., Schiffner, L., Kalyanaraman, R., Katinger, H., Lloyd, K. O.,
Kwong, P. D., and Moore, J. P. (2002) J Virol 76, 7293-730554. Astoul, C. H., Peumans, W. J., Van Damme, E. J., and Roug. (2000) Biochem Biophys
Res Commun 274, 455-46055. Mizuochi, T., Matthews, T. J., Kato, M., Hamako, J., Titani, K., Solomon, J., and Feizi,
T. (1990) J Biol Chem 265, 8519-852456. Clausen, H., Levery, S. B., McKibbin, J. M., and Hakomori, S. (1985) Biochemistry 24,
3578-358657. Frison, N., Taylor, M. E., Soilleux, E., Bousser, M. T., Mayer, R., Monsigny, M.,
Drickamer, K., and Roche, A. C. (2003) J Biol Chem 278, 23922-2392958. Ng, K. K., Park-Snyder, S., and Weis, W. I. (1998) Biochemistry 37, 17965-17976
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

28
FIGURE LEGENDS
Figure 1. Recombinant soluble DC-SIGN oligmerizes similarly to DC-SIGN on monocyte-
derived dendritic cells. Detergent solubilized lysates from monocyte derived dendritic cells (top
panel) and the purified recombinant 6X-histidine tagged extracellular domain of DC-SIGN
(sDC-SIGN) were resolved on a 5-20% sucrose density gradient. Gradient fractions were
immunoprecipitated with rabbit polyclonal antibodies made against DC-SIGN peptides from the
carboxy-terminus and repeat domain of DC-SIGN and ran on a non-denaturing gel. Native
molecular weight markers (MW) are indicated on the left-hand side of the top panel.
Immunoprecipitated DC-SIGN was then detected in a western blot with a mouse monoclonal
antibody against the repeat domain of DC-SIGN (DC028). Approximate positions of DC-SIGN
monomers and the higher ordered oligomers are indicated by the shaded bars above the top
panel.
Figure 2. gp120 binds with much higher affinity to DC-SIGN than ICAM2 and ICAM3.
(A) Binding affinity of gp120-Fc, ICAM2-Fc and ICAM3-Fc to recombinant sDC-SIGN. 200
nM of sDC-SIGN was coated on each well and increasing concentration of the indicated ligand
was added. The amount of ligand bound was detected colormetrically using a human Fc-specific
antibody conjugated to HRP. (B). Binding of gp120, ICAM-2 and ICAM-3 to cell surface DC-
SIGN. Binding of the three ligands to cell surface DC-SIGN expressed on THP-1, HS-Sultan,
and 293T cells was assessed by flow cytometry using PE-conjugated anti-Fc antibodies (see
methods). Kds were obtained by titrating in the amount of ligand added and normalizing the
maximal binding seen (in geometric mean fluorescence intensities) to 100%. Binding curves and
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

29
Kd values were generated via Graphpad Prism™.
Figure 3. HS Sultan DC-SIGN+ cells do not facilitate virus infection in trans despite having
a higher affinity for HIV-1 gp120. 250 pg p24 equivalents of JR-CSF (an R5 HIV isolate)
were added to 2.5 x104 of the indicated cells for 2 hours at 37 ºC. Excess virus was washed away
(4X) by media, and 2.5 x104 T-cell blasts were subsequently added to each well with the
respective cells. Supernatants were half-exchanged with fresh media on day 0, 3, 5, and 7, and
p24 levels in the supernatant were determined by a commercial p24 ELISA kit. Increased p24
levels over the course of 7 days is indicative of virus transfer and replication in the T-cell blasts
as the parental HS Sultan and THP-1 cells are not permissive for viral replication.
Figure 4. gp120 contains more mannose sugars than ICAM2 and ICAM3. 0.2 µg/ml of the
indicated biotinylated lectins were coated onto streptavidin plates and increasing amounts of
gp120-Fc, ICAM-2-Fc or ICAM-3-Fc were added. The amount of ligand bound was detected by
adding HRP-conjugated anti-human Fc antibodies. The highest O.D. value obtained was
normalized to 100% and binding curves and Kd values were generated via Graphpad Prism™.
GNA, Galanthus nivalis; DSA, Datura stramonium.
Figure 5. Inhibition curves for gp120-Fc binding to sDC-SIGN in the presence of the
various monosaccharides and disaccharides. (A) A 200 nM solution of sDC-SIGN was first
coated onto ELISA plates and 5 nM of gp120 (JR-CSF) was added in the absence or presence of
increasing amounts of the indicated monosaccharides. Competition curves and Ki values were
generated by Graphpad Prism™. Each data point was done in duplicate and each experiment for
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

30
every saccharide was repeated at least 3 times. (B) A schematic summary of DC-SIGN’s
saccharide binding preferences as elucidated in this work and others. Note the position of the
free 4-OH of each saccharide and the anomeric linkages of each disaccharide. Similar
competition curves were performed for both ICAM-2 and ICAM-3. See Table I for complete list
of Ki values.
Figure. 6. Alanine scanning mutagenesis reveals differential DC-SIGN binding
determinants to gp120, ICAM-2 and ICAM-3. In A, the alignment and the amino acid number
shown is that of human DC-SIGN. The sixteen amino acids that we have mutated to alanines are
indicated by stars. Each of the mutated residues are underscored differentially to highlight the
degree of sequence conservations amongst C-type lectins. The degree of conservation is based
on sequence alignments of 10 different C-type mannose-specific lectins: human CD23, DC-
SIGN, DC-SIGNR, DCIR, endo180, Langerin, mannose-binding lectin, mincle, macrophage
mannose receptor and rat serum mannose binding protein. In B, cell surface expression of
seventeen DC-SIGN alleles on 293T was assessed by binding with a DC-SIGN-specific
conformation-independent mAb DC028 that recognizes the repeat domain of DC-SIGN. The
conformation-dependent mAb 507 that recognizes the CRD of DC-SIGN was also used to
monitor whether there was any gross perturbation of DC-SIGN’s conformation. The mean
channel fluorescence of each DC-SIGN allele for each antibody (mAb DC028 and mAb 507)
was obtained and normalized to that observed for the wild-type allele, which was set at 100%.
The data are shown as the normalized mean +/- S.E.M. of 4 independent experiments. In C,
wildtype and the sixteen different DC-SIGN mutants were expressed on 293T and allowed to
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

31
bind to gp120-Fc, ICAM-2-Fc or ICAM-3-Fc. PE-conjugated anti-human Fc secondary
antibodies were used to detect the amount of bound ligand. The Mean Fluorescent Intensity of
each ligand bound to wild-type DC-SIGN was normalized to 100%. This normalized ligand
binding data was in turned normalized to the expression level of each DC-SIGN mutant as
determined by DC028 staining in B. Thus, if a ligand bound to a particular mutant at 50% of
wild-type levels but the expression of the mutant was only 50% of wild-type levels, the
expression-normalized binding of the ligand would be 100% of wild-type levels. The data is
plotted as the expression-normalized percentage bound of gp120, ICAM-2 and ICAM-3 for each
mutant allele compared to wildtype DC-SIGN.
Figure 7. The oligosaccharide binding groove of DC-SIGN. In A, the co-crystal of the CRD
of DC-SIGN complexed with the pentasaccharide is shown (adapted from ref. (41) ). The arrows
point to the three mannose residues and N-acetylgucosamine (GlcNAc5) that line the binding
groove of DC-SIGN. The extensive oligosaccharide binding valley is accented in white. In B,
the highly conserved binding pocket and the amino acid residues that have most drastic effects in
binding to all three ligands (i.e., E347 and N349) are shown. Note the extensive sugar and
calcium coordination contacts in this binding pocket. In C, the residues in our mutagenesis
studies that span the middle of the binding groove are shown. “Accessory” residues such as
S360, shown to have contacts with a mannose moiety in the crystal structure, appears to be
dispensable for gp120 binding but not ICAM-2 or ICAM-3 binding. In D, the other end of the
“oligomannose binding valley” enclosed by three amino acids S308, F313 and N311 is shown.
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

32
Table I. Saccharide competition data for DC-SIGN binding to gp120, ICAM-2 and
ICAM-3 . Inhibition constants (Ki) for each monosaccharide and disaccharide were determined
by a solid-phase competition assay. At least three independent experiments were done for each
condition stated and each experiment was done in duplicates. 95% confidence intervals (95%
C.I.) are indicated in parentheses. n.d., not done. The Ki of each saccharide tested relative to the
Ki of D-mannose is shown by the ratio of Ki saccharide/Ki mannose.
Ki (95% C.I.) Ki Saccharide/ Ki (95% C.I.) Ki Saccharide/ Ki (95% C.I.) Ki SaccharKi Mannose Ki Mannose Ki Mannose
MonosaccharidesmM mM mM
D-mannose 5.6 (3.7-8.7) 1.0 3.5 (2.5-5.5) 1.0 1.6 (0.6-3.8) 1.0L-mannose 350.0 (105.3-1164) 61.6 n.d. 110.9 (57.2-214.9) 68.8L-fucose 3.3 (2.0-5.4) 0.5 n.d. n.d.CH3-mannoside 3.0 (1.0-8.9) 1.5 19.3 (8.1-46.5) 5.5 0.9 (0.5-1.9) 0.6D-galactose 63.5 (32.3-124.8) 11.1 16.8 (4.5-63.3) 4.7 68.8 (22.3-211.6) 42.7CH3-galactose 163.0 (78.3-339.0) 28.7 196.3 (84.3-457.1) 55.7 117.9 (56.5-246.0) 73.2D-glucose 8.0 (4.5-14.4) 1.4 423.3 (149.5-1198) 120.2 2.3 (0.6-8.3) 1.4
DisaccharidesmM mM mM
Maltose 0.2 (<0.1-0.7) 0.1 5.5 (3.2-9.4) 1.5 13.0 (4.2-40.1) 8.1Cellobiose 1.4 (0.6-3.0) 0.2 n.d. n.d.Lactose 22.5 (11.5-43.9) 3.9 21.3 (10.0-45.3) 6.0 39.0 (8.5-178.7) 19.8Sucrose 11.2 (5.6-22.6) 1.9 76.8 (33.7-175) 21.8 6.8 (3.1-15.2) 4.2
gp120-Fc ICAM-2-Fc ICAM-3-Fc
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

33
Table II. Oligosaccharide array analysis of DC-SIGN binding specificity. Various mono-
and oligosaccharides were covalently linked on a 96-well ELISA plate as described in Materials
and Methods. The saccharide array was incubated with 200 nM of sDC-SIGN and the amount
of bound sDC-SIGN was detected with a mouse monoclonal antibody (DC028) against the
repeat region of DC-SIGN. HRP-conjugated anti-mouse antibodies were used as secondary
detection agents along with a chemiluminescent substrate. Data is shown in terms of relative
light units (RLUs) and the luminometer used exhibits a 5-log linearity from 102 –107 RLUs. The
saccharide binding data is consistent with the competition data presented in Table I. A subset of
the binding data indicating a preference of DC-SIGN for an alpha-anomeric glycosidic linkage is
shown. Binding data to galactose-based saccharides are shown as negative controls.
Saccharide Relative Light Units
Gal(ββββ1-3) GalNAc(ββββ) <100 Gal(ββββ1-4) Glc(ββββ) <100Gal(ββββ1-4) GlcNAc(ββββ) <100Gal(ββββ1-6) Gal(ββββ) <100
Glc(αααα1-4) Glc(αααα ) 2.87 X 106 Glc(αααα1-4) Glc(ββββ) 2.51 X 106
Glc(ββββ) 1.40 X 106 Glc(ββββ1-3) Glc(ββββ) 5.29 X 106 Glc(ββββ1-4) Glc(ββββ) 5.51 X 106
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

34
Su et. al. , Fig. 1
monomersdimers
trimerstetramers
MW
18412185
6858
40
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

35
Su et. al. Fig. 2
A
Kd (nM)
gp120-Fc 0.068 ± 0.006ICAM2-Fc 6.66 ± 0.88ICAM3-Fc 17.0 ± 1.74
gp120-Fc 0.017 ± 0.001ICAM2-Fc 24.99 ± 2.34ICAM3-Fc 26.35 ± 1.74
gp120-Fc 0.078 ± 0.006ICAM2-Fc 57.90 ± 4.09ICAM3-Fc 13.92 ± 1.38
-12 -11 -10 -9 -8 -7 -6 -5-50
0
50
100
150gp120-Fc
ICAM2-Fc
ICAM3-Fc
[Ligand] Log10 (M)
-12 -11 -10 -9 -8 -7 -6-50
0
50
100
150gp120-Fc
ICAM2-Fc
ICAM3-Fc
[Ligand] Log10 M
Cell Line Expressing DC-SIGN
-12 -11 -10 -9 -8 -7 -6-50
0
50
100
150
gp120-Fc
ICAM2-Fc
ICAM3-Fc
[Ligand] Log10 (M)
THP-1
HS Sultan
293T
B.
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from
36
Su et. al., Fig. 3
0 . 0
0 . 5
1 . 0
1 . 5
2 . 0
2 . 5
3 . 0
3 . 5
0 2 4 6 8
THP-1
THP-1 DC-SIGN
HS Sultan
HS Sultan DC-SIGN
p24
ng/m
l
Days Post-infection
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

37
Su et. al., Fig. 4
gp120ICAM2ICAM3
58.7
Kd mM
0.1522.720.4
Lectin
GNA
DSA
-7 -6 -5 -4 -3 -2 -1 00
50
100
150gp120ICAM-2ICAM-3
[Ligand] log10 M
gp120ICAM2ICAM3
76.30.74
-7 -6 -5 -4 -3 -2 -1 00
50
100
150gp120ICAM-2ICAM-3
[Ligand] log10 M
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

38
Su et. al., Fig. 5
B.
Maltose
OH
CH2OH CH2OH
αααα -anomericLinkage
CH2OH CH2OH
ββββ-anomericLinkage
OH
CH2OH
OH
CH2OH
OH
CH2OH CH2OH
ββββ-anomericLinkage
OHCH2OH
OH
Cellobiose D-Mannose D-Glucose Lactose D-Galactose
> > > >>≥≥≥≥
Saccharides with free equatorial 4’-OH groups Axial 4’-OH groups
A.%
of M
axim
al B
indi
ng
-4 -3 -2 -1 0-50
0
50
100
150
D-mannose
CH3-mannose
L-fucose
D-galactoseCH3-galactoseL-mannose
Log10 M[Saccharide]
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

39
Su et. al., Fig. 6A, B
SSRSNRFT // WNRGEPNNVGEEDCAEFSGNGWNDDKCNLAK �
307 314 343 345 350 355 360 365 370A.
Sequence Conservation AmongstC-type lectins
Low (< 40%)
Medium (40-80%)
High (80-100%)
Low (< 40%)
Medium (40-80%)
High (80-100%)
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
w t N 3 1 1 R 3 4 5 G 3 4 6 E 3 4 7 P 3 4 8 N 3 4 9 V 3 5 1 G 3 5 2 E 3 5 3 E 3 5 4 D 3 5 5 S 3 6 0 G 3 6 1 N 3 6 2 D 3 6 6 D 3 6 7
DC028 mAB Conformation-Independent507 mAB Conformation-Dependent
% o
f w
ildtp
ye b
indi
ng
B.
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

40
Su et. al., Fig. 6C
C.
Low (< 40%)
Medium (40-80%)
High (80-100%)
% o
f wild
-typ
e B
indi
ng
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
w t N311 R345 G346 E347 P348 N349 V351 G352 E353 E354 D355 S360 G361 N362 D366 D367
gp120FcICAM2FcICAM3Fc
DC-SIGN Mutant Binding to gp120, ICAM-2 and ICAM-3
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

41
Su et. al., Fig. 7
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from
42
Su et. al., Fig. 7
S360G361N362
Man3
Man4
F313
S308
Man3Man4
GlcNAc5
D.
F313
C.
D.
N311
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

LeeStephen V. Su, Patrick Hong, Sarah Baik, Oscar A. Negrete, Kevin B. Gurney and Benhur
ICAM-2 and ICAM-3DC-SIGN binds to HIV-1 gp120 in a distinct but overlapping fashion compared to
published online February 16, 2004J. Biol. Chem.
10.1074/jbc.M400184200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on August 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from