a review of stock identification of haddock, · pdf filediv. 5z - usa), and 6) gulf of maine...
TRANSCRIPT

60(4), 1998 1
Introduction
Haddock, Melanogrammus aeglefinus(Fig. 1), is a commercially importantgroundfish species of the gadid familythat is distributed throughout the north-west Atlantic from Greenland to CapeHatteras (Bigelow and Schroeder,1953). This range is effectively dividedby the Fundian (“Northeast”) andLaurentian Channels, both in excess of180 m in depth (Fig. 2). Although had-
A Review of Stock Identification ofHaddock, Melanogrammus aeglefinus , in
the Northwest Atlantic Ocean
GAVIN A. BEGG
Gavin A. Begg is an NRC Research Associatewith the Northeast Fisheries Science Center,National Marine Fisheries Service, NOAA, 166Water Street, Woods Hole, MA 02543. Email:[email protected]
ABSTRACT—Haddock, Melanogrammusaeglefinus, is a principal commercial spe-cies distributed throughout the northwestAtlantic Ocean, with major aggregations oc-curring on Georges Bank and on the ScotianShelf. This review examines all availableinformation on stock structure of haddockto evaluate the suitability of current stockunits and to investigate areas that requirefurther research. Combined informationfrom tag-recapture, demographic, recruit-ment, meristic, parasitic, and genetic stud-ies provide evidence for the identificationof haddock stocks, with major populationdivisions occurring between New England,Nova Scotia, and Newfoundland waters.Within each of these major divisions a num-ber of discrete stocks appear to exist, al-though uncertainty remains in the amountof separation found within each region.Research utilizing more recent stock iden-tification techniques should refine and im-prove our understanding of haddock stockstructure in the northwest Atlantic.
dock are demersal, they are rarely foundin abundance below 180 m, resulting inthe channels acting as barriers to dis-persal (Needler, 1930). Haddock arefound in cool, temperate waters acrossthe continental shelf and over offshoresubmerged banks, with major commer-cial aggregations occurring on thesouthern Grand Bank, Scotian Shelf,and Georges Bank (Zwanenburg et al.,1992).
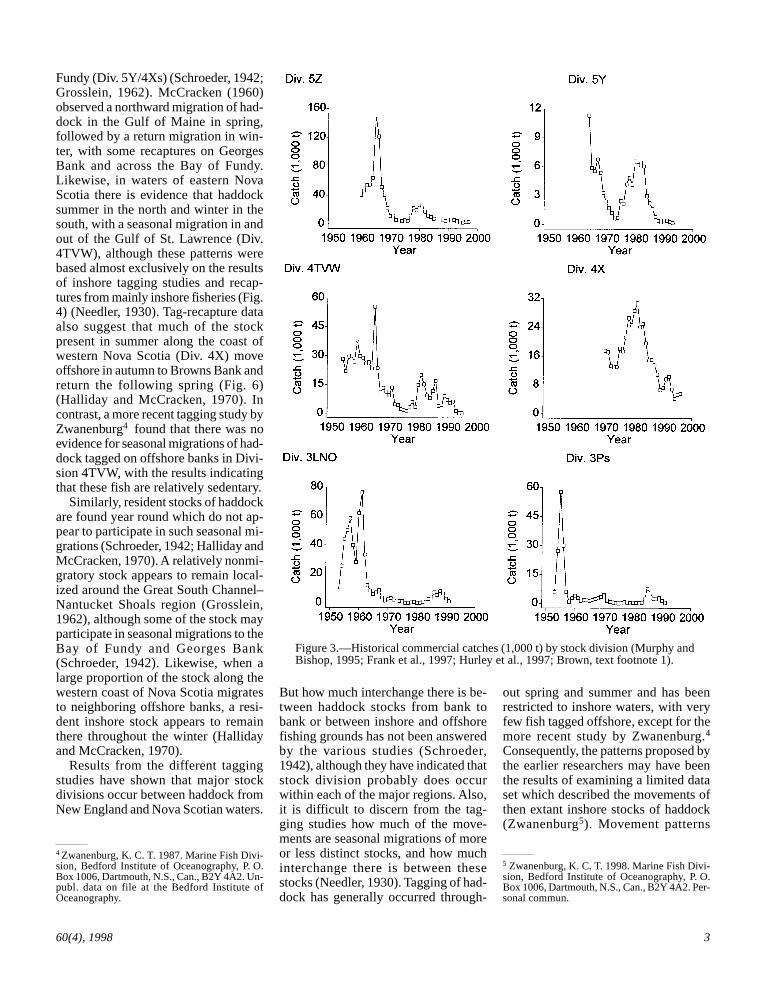
Historically, haddock has been a keyspecies in terms of abundance and con-tribution to the commercial fisheries inthe northwest Atlantic with peak land-ings of 249,000 metric tons (t) in 1965(ICNAF, 1967). In contrast, landingshave declined to 24,500 t in 1992 (NAFO,1995), with most haddock stocks acrossthe region in a depressed condition andthe focus of rebuilding plans (Fig. 3)(Murphy and Bishop, 1995; Hurley et
al., 1997; Frank et al., 1997; Gavarisand Van Eeckhaute, 1998; Brown1).
Beneficial to stock recovery and ef-fective fisheries management is the“stock concept,” as recruitment withineach stock must sustain each popu-lation’s catch (Kutkuhn, 1981). In fish-eries science, many claim that the mostuseful definition of a stock is one thathas a sound genetic basis because man-agement policies may not achieve long-term conservation goals without theknowledge of the number of noninter-breeding, self-recruiting populationscontained within an exploited speciesdistribution (Ovenden, 1990). Similarly,in accordance with the “biological stock
Figure 1.—Haddock, Melanogrammus aeglefinus.
1 Brown, R. W. 1998. U.S. assessment of theGeorges Bank (5Z) haddock stock, 1998. U.S. Dep.Commer, NOAA, Natl. Mar. Fish. Serv., NortheastFish. Sci. Cent., NEFSC Lab. Ref. Doc. 98.

2 Marine Fisheries Review
Figure 2.—ICNAF/NAFO scientific and statistical subareas, divisions, and subdi-visions of the northwest Atlantic.
concept,” fisheries management in thenorthwest Atlantic has been concernedwith the delineation of fishing areas thatcorrespond to geographic ranges of in-dependently reproducing populations(stock distribution areas) (Zwanenburget al., 1992). Interpretation of stockstructure in this review relates to theIhssen et al. (1981) definition of a stockas “a group of randomly mating, repro-ductively isolated individuals of a singlespecies with temporal or spatial integrity.”
The definition of management unitsin the northwest Atlantic upon whichhaddock stocks are assessed and regu-lated has been based not only on con-siderations of biological stock structure,but also on considerations of other spe-cies and fishery distributions, oceano-graphic features, submarine topography,political and administrative boundaries,homogeneity of international fisheriesparticipation, and practicalities of datacollection and fishery regulation (Halli-day and Pinhorn, 1990). In contrast tobiological stock units, fisheries manage-
ment units are geographic areas inwhich a suite of regulatory measurescan be applied to achieve specific man-agement objectives (Gavaris and VanEeckhaute, 1998). Currently, six stocksare recognized for haddock in the north-west Atlantic: 1) Grand Banks (Div.3LNO), 2) St. Pierre Bank (Div. 3Ps),3) Eastern Scotian Shelf and SouthernGulf of St. Lawrence (Div. 4TVW), 4)Western Scotian Shelf (Div. 4X), 5)Georges Bank (Div. 5Zjm - Canada;Div. 5Z - USA), and 6) Gulf of Maine(Div. 5Y) (Fig. 2).
Stock identification studies of had-dock in the northwest Atlantic havebeen conducted since the 1930’s, usinga variety of approaches including tag-recapture, demographic, recruitment,meristic, parasitic, and genetic tech-niques. Delineation of haddock stockshas been complicated by seasonal dif-ferences in the species’ spatial distribu-tions (Zwanenburg et al., 1992) and thelimitations associated with individualstock identification methods. This pa-
per reviews the literature on stock iden-tification of haddock in the northwestAtlantic in order to evaluate the suit-ability of the current management/stockunits by using all available biological in-formation, rather than that acquired froma single procedure in isolation. Areas forfuture research are also discussed.
Stock Identification Techniques
Tag-Recapture
Stock identification of haddock in theGulf of Maine and Nova Scotian wa-ters was initially determined using tag-recapture methods (Needler, 1930;Schroeder, 1942; McCracken, 1960;Halliday and McCracken, 1970; Mc-Kenzie2; McCracken3). Needler (1930)hypothesized three main stocks of had-dock in the northwest Atlantic (NewEngland, Nova Scotia, and Newfound-land) based on divisions of shallow-water areas by the Fundian and Laur-entian Channels and movement patternsof the species. Tag-recapture data haveindicated that there is little interchangebetween haddock from New Englandand Nova Scotian waters (Fig. 4, 5)(Needler, 1930; Schroeder, 1942). How-ever, in the Bay of Fundy, where the tworegions are linked by shallow water,some mixing of haddock occurs (Fig.6) (Needler, 1930; McCracken, 1960;Halliday and McCracken, 1970).
Seasonal migrations have been ob-served in haddock from both New En-gland and Nova Scotian waters (Needler,1930; McCracken, 1960; Halliday andMcCracken, 1970). Typically, haddockmove to inshore shallow waters inspring and to deeper offshore watersthroughout winter and late summer(Fig. 6) (Needler, 1930). A large num-ber of local tag-returns a year or morelater show that most haddock return tothe same locality (Needler, 1930). Tag-recapture data have indicated that aseparate seasonally migrating stock ex-ists along the coast of the Gulf of Mainefrom Jeffreys Ledge to the Bay of
2 McKenzie, R. A. 1940. The spring haddock“run,” Jordan Harbour, N.S. Fish. Res. BoardCan., Atl. Prog. Rep. 28:9-13.3 McCracken, F. D. 1956. Cod and haddock tag-ging off Lockeport, N.S. Fish. Res. Board Can.,Atl. Prog. Rep. 64:10-15.
60(4), 1998 3
Figure 3.—Historical commercial catches (1,000 t) by stock division (Murphy andBishop, 1995; Frank et al., 1997; Hurley et al., 1997; Brown, text footnote 1).
Fundy (Div. 5Y/4Xs) (Schroeder, 1942;Grosslein, 1962). McCracken (1960)observed a northward migration of had-dock in the Gulf of Maine in spring,followed by a return migration in win-ter, with some recaptures on GeorgesBank and across the Bay of Fundy.Likewise, in waters of eastern NovaScotia there is evidence that haddocksummer in the north and winter in thesouth, with a seasonal migration in andout of the Gulf of St. Lawrence (Div.4TVW), although these patterns werebased almost exclusively on the resultsof inshore tagging studies and recap-tures from mainly inshore fisheries (Fig.4) (Needler, 1930). Tag-recapture dataalso suggest that much of the stockpresent in summer along the coast ofwestern Nova Scotia (Div. 4X) moveoffshore in autumn to Browns Bank andreturn the following spring (Fig. 6)(Halliday and McCracken, 1970). Incontrast, a more recent tagging study byZwanenburg4 found that there was noevidence for seasonal migrations of had-dock tagged on offshore banks in Divi-sion 4TVW, with the results indicatingthat these fish are relatively sedentary.
Similarly, resident stocks of haddockare found year round which do not ap-pear to participate in such seasonal mi-grations (Schroeder, 1942; Halliday andMcCracken, 1970). A relatively nonmi-gratory stock appears to remain local-ized around the Great South Channel–Nantucket Shoals region (Grosslein,1962), although some of the stock mayparticipate in seasonal migrations to theBay of Fundy and Georges Bank(Schroeder, 1942). Likewise, when alarge proportion of the stock along thewestern coast of Nova Scotia migratesto neighboring offshore banks, a resi-dent inshore stock appears to remainthere throughout the winter (Hallidayand McCracken, 1970).
Results from the different taggingstudies have shown that major stockdivisions occur between haddock fromNew England and Nova Scotian waters.
4 Zwanenburg, K. C. T. 1987. Marine Fish Divi-sion, Bedford Institute of Oceanography, P. O.Box 1006, Dartmouth, N.S., Can., B2Y 4A2. Un-publ. data on file at the Bedford Institute ofOceanography.
But how much interchange there is be-tween haddock stocks from bank tobank or between inshore and offshorefishing grounds has not been answeredby the various studies (Schroeder,1942), although they have indicated thatstock division probably does occurwithin each of the major regions. Also,it is difficult to discern from the tag-ging studies how much of the move-ments are seasonal migrations of moreor less distinct stocks, and how muchinterchange there is between thesestocks (Needler, 1930). Tagging of had-dock has generally occurred through-
out spring and summer and has beenrestricted to inshore waters, with veryfew fish tagged offshore, except for themore recent study by Zwanenburg.4
Consequently, the patterns proposed bythe earlier researchers may have beenthe results of examining a limited dataset which described the movements ofthen extant inshore stocks of haddock(Zwanenburg5). Movement patterns
5 Zwanenburg, K. C. T. 1998. Marine Fish Divi-sion, Bedford Institute of Oceanography, P. O.Box 1006, Dartmouth, N.S., Can., B2Y 4A2. Per-sonal commun.

4 Marine Fisheries Review
Figure 4.—Migration patterns of tagged haddock in New England and Nova Scotianwaters showing little exchange between tagged fish in the two regions (Needler, 1930).
presented by the recaptures, therefore,is incomplete and may be biased byvariable fishing effort. Also, the presentpatterns of distribution and movementmay be difficult to compare to previouswork in that the earlier population lev-els are so different from present verylow levels (Zwanenburg5).
Demographics
Demographic or biological popula-tion parameters have been used to dif-ferentiate a finer scale of haddock stockseparation in northwest Atlantic watersthan that achieved by tag-recapturepractices (Table 1). Growth differencesof haddock from New England andNova Scotian waters support the currentstock units throughout these regions(Fig. 7). Needler (1930) observed dif-ferences in growth rates of haddockfrom corresponding inshore and off-shore areas of New England and NovaScotia, providing further evidence to thetag-recapture data of stock division be-tween fish from these regions. InshoreGulf of Maine (Div. 5Y) and offshoreGeorges Bank (Div. 5Z) haddock grewfaster than those in coastal Nova Scotianwaters and Browns Bank, respectively(Div. 4X). Likewise, Schuck and Arnold(1951) found significant differences inmean length-at-age between haddockfrom Georges Bank and Browns Bank,suggesting that little mixing occurs be-tween fish from these regions.
As suggested by the tagging studies,stock structure was also apparent withineach of the major regions, with the pos-sibility of distinct inshore and offshorestocks. Haddock along the coast of theGulf of Maine appear to grow slowerthan those fish from Nantucket Shoalsand Georges Bank, indicating possiblestock division between fish from theseareas (Clark et al., 1982; Begg et al.6).Differential growth rates between had-dock from the eastern part of GeorgesBank and Nantucket Shoals suggestedthat fish from these areas may also bedistinct stocks (Begg et al.6). Similarly,faster growth rates of inshore haddockthan those from offshore waters of the
6 Begg, G. A., J. A. Hare, and D. D. Sheehan.The role of life history parameters as indicatorsof stock structure. Manuscr. in review.

60(4), 1998 5
Figure 5.—Migration patterns of tagged haddock in New England waters, showing little exchange with Nova Scotian stocks,except in the Bay of Fundy (Schroeder, 1942).
Scotian Shelf suggested stock divisionwithin this region (Needler, 1930).
Hennemuth et al. (1964) observed simi-lar growth rates and age composition of
haddock from the western (Div. 4X) andeastern (Div. 4VW) part of the Scotian

6 Marine Fisheries Review
Figure 6.—Seasonal migrations of haddock in New England and Nova Scotian waters,tagged along the eastern and western shores of the Bay of Fundy (McCracken, 1960).
Figure 7.—Growth differences inmean lengths at age among haddockstocks Div. 4TVW (Frank et al.,1997), 4X (Hurley et al., 1997), 5Yand 5Z (calculated from 1997 NMFSNortheast Fisheries Science Centerspring bottom-trawl survey data).
Shelf and the southern Gulf of St.Lawrence (Div. 4T). In contrast, had-dock in the Bay of Fundy (Div. 4Xs)had a faster growth rate and younger agecomposition than those from waters offwestern Nova Scotia (Hennemuth et al.,1964), although these differences mayhave been influenced by gear selectiv-ity problems and mixing of haddockfrom New England and Nova Scotian
stocks within the bay. Beacham (1983)and Trippel et al. (1997) found differ-ences in median lengths at sexual ma-turity among haddock from eastern andwestern Scotian Shelf waters, althoughthese results were not statistically com-pared (Table 1). Haddock from the west-ern Scotian Shelf (Div. 4X) tend to belarger and older at maturation than thosefrom Georges Bank (Div. 5Z) (Over-
holtz, 1987), but smaller than thosefrom St. Pierre Bank (Div. 3Ps) (Table1). Stock separation of haddock withinNewfoundland waters has been basedmainly on differences in growth rates(Templeman et al., 1978a). Haddock onSt. Pierre Bank typically grow faster andare of a greater mean length-at-age andlength at first-maturity than those on theGrand Bank (Div. 3LNO) (Table 1)(Templeman et al., 1978a,b; Temple-man and Bishop, 1979a,b).
Stock division of haddock inferredfrom several demographic characteris-tics has tended to agree with patternsdeveloped from the tag-recapture stud-ies. However, for the most part, indi-vidual regional growth curves have notbeen compared in a statistically rigor-ous fashion to determine the signifi-cance, if any, of the apparent growthdifferences used to separate the stocks(Schuck and Arnold, 1951). Needler(1930) only presented average length-at-age data, and samples were taken bycommercial fishing gear which ex-cluded younger haddock, while prob-ably selecting for larger sizes of spe-cific aged fish. Consequently, discrep-ancies in growth rates estimated byNeedler (1930) and Hennemuth et al.(1964) may have been related to differ-ences in sampling and analytical meth-odologies. Differences in samplingtimes and gear types between the vari-ous studies has resulted in a lack of ho-mogeneity in the origin of the samplesused to provide demographic charactersto differentiate haddock stocks. Also,

60(4), 1998 7
Table 1.—Summary of biological parameters (length/age at 50% maturity: L 50, A50; and von Bertalanffy growthcoefficients: L∞, K, t0) derived for each haddock stock Div. 3LNO, 3Ps, 4TVW, 4X, 5Y, and 5Z.
Stock division
Parameter 3LNO 3Ps 4TVW 4X 5Y 5Z
L50 (cm) females 511 503 304 384 397 397
L50 (cm) males 441 403 294 334 277 317
A50 (years) females 5.11 4.33 3.44 3.64 2.87 2.47
A50 (years) males 4.01 3.33 2.94 3.34 1.97 1.87
L∞ 74.82 84.62 51.35 76.66 72.98 73.88
K 0.102 0.242 0.265 1.126 0.358 0.388
t0 –3.62 0.472 –0.975 0.186 0.308 0.178
1 Templeman et al. (1978b).2 Templeman and Bishop (1979a).3 Templeman and Bishop (1979b).4 Trippel et al. (1997).5 Calculated from mean lengths-at-age reported in Frank et al. (1997).6 O’Boyle et al. (1988).7 Begg et al., text footnote 6.8 O’Brien et al. (1993).
Figure 8.—Principal spawning locations of haddock in the northwest Atlantic (Pageand Frank, 1989).
while the magnitude of the differencesin growth rates that have been detectedamong haddock from some regions maybe sufficiently large to indicate real dif-ferences in the stocks (Hennemuth etal., 1964), these differences may be theresult of demographic plasticity withinstocks as a response to changing ambi-ent abiotic and biotic conditions.
Spawning and Recruitment Patterns
Spawning Times and Locations
Further evidence for stock discrimi-nation of haddock within New Englandand Nova Scotian waters has been pro-vided by spawning and recruitment pat-terns of the species in relation to thedistinct oceanographic conditions ofeach region. Peak spawning of haddockoccurs in late March and early Aprilwithin New England (Marak and Living-stone, 1970; Grosslein and Hennemuth,1973; Lough and Bolz, 1989), from lateApril to June in Nova Scotia (Hurley andCampana, 1989; Page and Frank, 1989;Waiwood and Buzeta, 1989), andthroughout June and July in Newfound-land waters (Templeman et al., 1978b;Templeman and Bishop, 1979b). Spawn-ing periodicity of haddock is highly vari-able and appears to be correlated withwater temperature, resulting in delayedspawning during colder years and ad-vanced spawning during warmer years(Page and Frank, 1989).
Throughout the northwest Atlanticthe main spawning grounds of haddockoccur over Georges Bank (Div. 5Z), thewestern part of the Scotian Shelf (Div.4X), Gulf of Maine (Div. 5Y), Nan-tucket Shoals (Div. 5Z), Emerald-West-ern Banks (Div. 4VW), Grand Bank(Div. 3LNO), and St. Pierre Bank (Div.3Ps) (Fig. 8) (Colton and Temple, 1961;Smith and Morse, 1985; Page andFrank, 1989). Along the Scotian Shelfthe primary spawning grounds occur onBrowns Bank (Div. 4X) and Emerald-Western Banks (Div. 4VW), with lowerlevels on adjacent banks and in inshorecoastal areas (Hurley and Campana,1989; Campana et al., 1989). Buoyancycharacteristics of haddock eggs and lo-cal physical and oceanographic condi-tions may result in hatching failurethroughout these inshore waters (Frank
8 Marine Fisheries Review
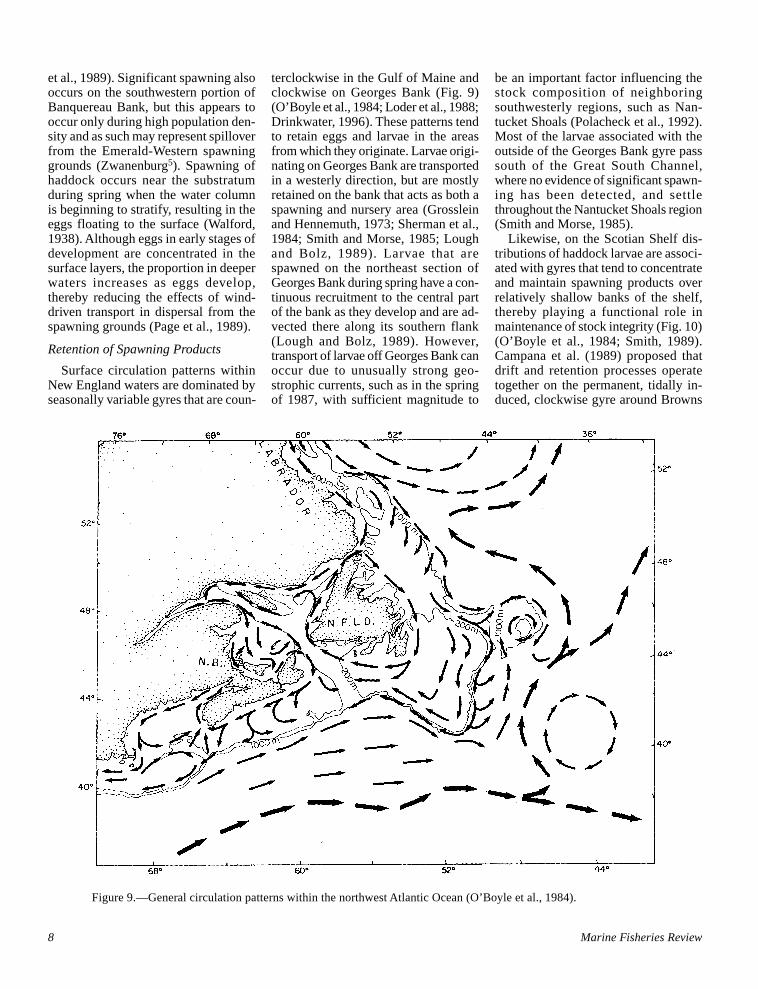
Figure 9.—General circulation patterns within the northwest Atlantic Ocean (O’Boyle et al., 1984).
et al., 1989). Significant spawning alsooccurs on the southwestern portion ofBanquereau Bank, but this appears tooccur only during high population den-sity and as such may represent spilloverfrom the Emerald-Western spawninggrounds (Zwanenburg5). Spawning ofhaddock occurs near the substratumduring spring when the water columnis beginning to stratify, resulting in theeggs floating to the surface (Walford,1938). Although eggs in early stages ofdevelopment are concentrated in thesurface layers, the proportion in deeperwaters increases as eggs develop,thereby reducing the effects of wind-driven transport in dispersal from thespawning grounds (Page et al., 1989).
Retention of Spawning Products
Surface circulation patterns withinNew England waters are dominated byseasonally variable gyres that are coun-
terclockwise in the Gulf of Maine andclockwise on Georges Bank (Fig. 9)(O’Boyle et al., 1984; Loder et al., 1988;Drinkwater, 1996). These patterns tendto retain eggs and larvae in the areasfrom which they originate. Larvae origi-nating on Georges Bank are transportedin a westerly direction, but are mostlyretained on the bank that acts as both aspawning and nursery area (Grossleinand Hennemuth, 1973; Sherman et al.,1984; Smith and Morse, 1985; Loughand Bolz, 1989). Larvae that arespawned on the northeast section ofGeorges Bank during spring have a con-tinuous recruitment to the central partof the bank as they develop and are ad-vected there along its southern flank(Lough and Bolz, 1989). However,transport of larvae off Georges Bank canoccur due to unusually strong geo-strophic currents, such as in the springof 1987, with sufficient magnitude to
be an important factor influencing thestock composition of neighboringsouthwesterly regions, such as Nan-tucket Shoals (Polacheck et al., 1992).Most of the larvae associated with theoutside of the Georges Bank gyre passsouth of the Great South Channel,where no evidence of significant spawn-ing has been detected, and settlethroughout the Nantucket Shoals region(Smith and Morse, 1985).
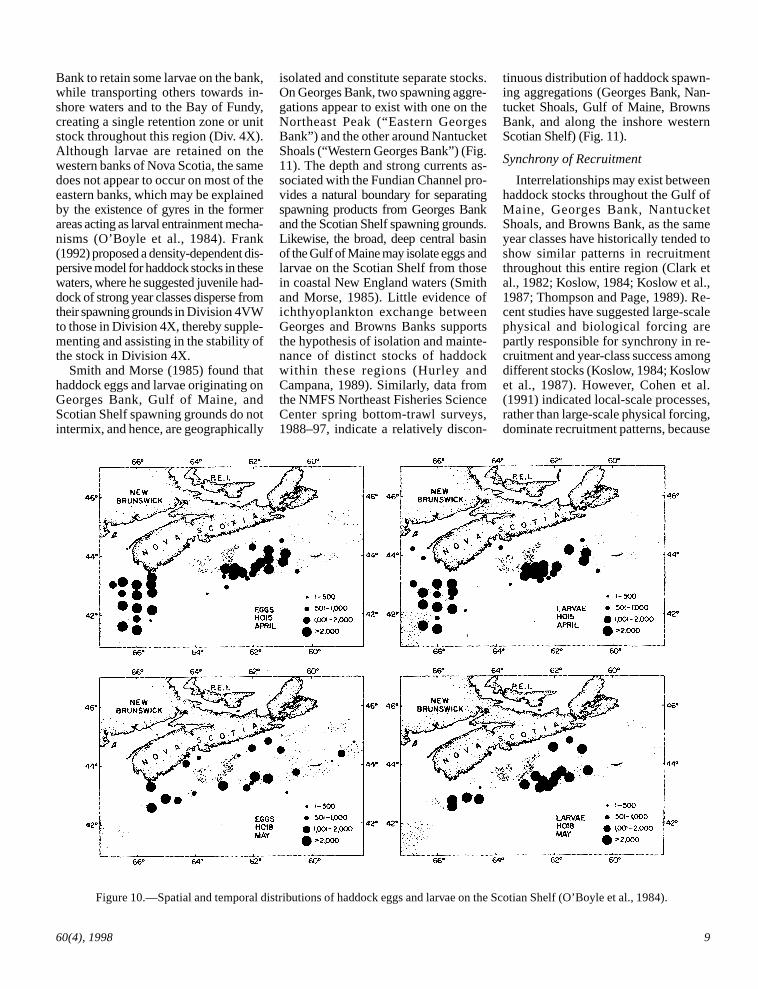
Likewise, on the Scotian Shelf dis-tributions of haddock larvae are associ-ated with gyres that tend to concentrateand maintain spawning products overrelatively shallow banks of the shelf,thereby playing a functional role inmaintenance of stock integrity (Fig. 10)(O’Boyle et al., 1984; Smith, 1989).Campana et al. (1989) proposed thatdrift and retention processes operatetogether on the permanent, tidally in-duced, clockwise gyre around Browns
60(4), 1998 9
Figure 10.—Spatial and temporal distributions of haddock eggs and larvae on the Scotian Shelf (O’Boyle et al., 1984).
Bank to retain some larvae on the bank,while transporting others towards in-shore waters and to the Bay of Fundy,creating a single retention zone or unitstock throughout this region (Div. 4X).Although larvae are retained on thewestern banks of Nova Scotia, the samedoes not appear to occur on most of theeastern banks, which may be explainedby the existence of gyres in the formerareas acting as larval entrainment mecha-nisms (O’Boyle et al., 1984). Frank(1992) proposed a density-dependent dis-persive model for haddock stocks in thesewaters, where he suggested juvenile had-dock of strong year classes disperse fromtheir spawning grounds in Division 4VWto those in Division 4X, thereby supple-menting and assisting in the stability ofthe stock in Division 4X.
Smith and Morse (1985) found thathaddock eggs and larvae originating onGeorges Bank, Gulf of Maine, andScotian Shelf spawning grounds do notintermix, and hence, are geographically
isolated and constitute separate stocks.On Georges Bank, two spawning aggre-gations appear to exist with one on theNortheast Peak (“Eastern GeorgesBank”) and the other around NantucketShoals (“Western Georges Bank”) (Fig.11). The depth and strong currents as-sociated with the Fundian Channel pro-vides a natural boundary for separatingspawning products from Georges Bankand the Scotian Shelf spawning grounds.Likewise, the broad, deep central basinof the Gulf of Maine may isolate eggs andlarvae on the Scotian Shelf from thosein coastal New England waters (Smithand Morse, 1985). Little evidence ofichthyoplankton exchange betweenGeorges and Browns Banks supportsthe hypothesis of isolation and mainte-nance of distinct stocks of haddockwithin these regions (Hurley andCampana, 1989). Similarly, data fromthe NMFS Northeast Fisheries ScienceCenter spring bottom-trawl surveys,1988–97, indicate a relatively discon-
tinuous distribution of haddock spawn-ing aggregations (Georges Bank, Nan-tucket Shoals, Gulf of Maine, BrownsBank, and along the inshore westernScotian Shelf) (Fig. 11).
Synchrony of Recruitment
Interrelationships may exist betweenhaddock stocks throughout the Gulf ofMaine, Georges Bank, NantucketShoals, and Browns Bank, as the sameyear classes have historically tended toshow similar patterns in recruitmentthroughout this entire region (Clark etal., 1982; Koslow, 1984; Koslow et al.,1987; Thompson and Page, 1989). Re-cent studies have suggested large-scalephysical and biological forcing arepartly responsible for synchrony in re-cruitment and year-class success amongdifferent stocks (Koslow, 1984; Koslowet al., 1987). However, Cohen et al.(1991) indicated local-scale processes,rather than large-scale physical forcing,dominate recruitment patterns, because

10 Marine Fisheries Review
Figure 11.—NMFS Northeast Fisheries Science Center spring bottom-trawl surveycatch (number) per tow of sexually mature haddock, 1988–97.
most significant correlations were foundbetween neighboring stocks.
Meristics and Morphometrics
Meristic characters, in the form ofvertebral number, have been used toprovide insights into stock structure ofhaddock (Clark and Vladykov, 1960;Tremblay et al., 1984; Vladykov7).Vladykov7 used vertebral counts ofadult haddock to confirm the threepopulation groups suggested by Needler(1930): New England, Nova Scotia, andNewfoundland. After updating this mer-istic analysis, Clark and Vladykov(1960) proposed that the Nova Scotiastock be divided into three stocks: east-ern, central, and western Nova Scotia.Mean number of vertebrae differed sig-nificantly between the five stocks andincreased with latitude from New En-gland to the eastern Scotian Shelf, sug-gesting an inverse relationship withwater temperature (Fig. 12). In contrast,Tremblay et al. (1984) found lower ver-tebral numbers for juvenile haddockfrom the northeastern part of the ScotianShelf (Div. 4V) than those from central(Div. 4W) and western Nova Scotianwaters (Div 4X). Based on these differ-ences, they proposed that the haddockstock occupying the northeastern andcentral area (Div. 4VW) of the ScotianShelf be divided into two stock compo-nents: eastern Scotian Shelf (Div. 4V)and central Scotian Shelf (Div. 4W).
A major limitation of these meristicstudies has been the lack of temporalcomparisons to examine the consistencyof stock structure patterns over time.Spatial comparisons have also been re-stricted, with few inshore samples hav-ing been collected. Results presented byTremblay et al. (1984) were mean val-ues and were not accompanied by datashowing the amount of variability bothwithin and between samples. Conse-quently, it is difficult to comment on thevalidity of the differences in their re-sults compared to those of Clark andVladykov (1960). The actual degree ofmixing between stocks cannot be esti-mated from any of the meristic studies,and the minor differences used to sepa-
Figure 12.—Mean vertebrae number of haddock from fishing grounds of the north-west Atlantic Ocean (Clark and Vladykov, 1960).
rate stocks should be viewed with cau-tion, particularly given the high degreeof environmentally influenced plastic-ity that can exist in meristic characters.
More recently, otolith shape analy-sis was investigated to determine its util-ity as a tool for stock discrimination of
haddock stocks in the northwest Atlan-tic (Begg8). Several shape variables
7 Vladykov, V. D. 1935. Haddock races along theNorth American coast. Biol. Board Can., Atl.Prog. Rep. 14:3-7.
8 Begg, G. A. 1998. Northeast Fisheries ScienceCenter, National Marine Fisheries Service,NOAA, 166 Water Street, Woods Hole, MA02543. Unpublished data on file at the NortheastFisheries Science Center.

60(4), 1998 11
Figure 13.—Relationship betweengenetic difference (Nei’s D) and geo-graphic distance separating haddockon northwest Atlantic banks (Zwan-enburg et al., 1992).
Table 2.—Results of stock identification studies to differentiate haddock stocks in the Northwest Atlantic. 1
Stock structure (no. of stocks identified within)
Stock I.D. method New England Nova Scotia Newfoundland
Tag-recapture 1-2 2(5Y; 5Z / N.Sh.?) (4X; 4TVW) N/A
Growth rates 2-3 2 2(5Y; 5Z; N.Sh.?) (4X; 4TVW) (3Ps; 3LNO)
Spawning patterns 2-3 2 2(5Y; 5Z; N.Sh.?) (4X; 4TVW) (3Ps; 3LNO)
Meristics N/A 3-4 N/A(4X; 4V; 4W; 4T?)
Parasites N/A 3 N/A(4Xs; 4X; 4W)
Genetics ? ? N/ACurrent management units 1(Canada) / 2 (USA) 2 2
(5Zjm - Canada; 5Z, 5Y-USA) (4X; 4TVW) (3Ps; 3LNO)
1 Key: Div. 5Zjm - Georges Bank, Canada; Div. 5Z - Georges Bank, USA; Div. 5Y - Gulf of Maine; N.Sh. - NantucketShoals; Div. 4X - Western Nova Scotia; Div. 4Xs - Bay of Fundy; Div. 4W - Central Nova Scotia; Div. 4V - Eastern NovaScotia; Div. 4T - Gulf of St. Lawrence; Div. 3Ps - St. Pierre Bank; Div. 3LNO - Grand Bank. N/A = No data available. ? =Results uncertain.
were statistically significant betweenhaddock samples from Georges Bank,the Gulf of Maine, and the ScotianShelf, providing a phenotypic measureof stock separation.
Parasites
The use of parasites for stock identi-fication of haddock in the northwestAtlantic has been limited to a prelimi-nary study by Scott (1981). A total of19 species of alimentary-tract parasitesof haddock from the Scotian Shelf wereidentified, although most of them wereubiquitous, occurring in a variety ofhosts over wide geographical areas. Onlytwo species (Digenea: Lepidapedonrachion and Myxosporida: Myxidiumbergense) showed the degree of hostspecificity, abundance, and correlationwith known haddock stock delineationneeded to be considered prospects forbiological tags. Infestation of L. rachionin haddock from Browns Bank (Div.4X) was higher than those from the Bayof Fundy (Div. 4Xs) and lower thanthose from the Emerald-Banquereauarea (Div. 4W), providing indirect evi-dence for separate stocks in these areas(Scott, 1981). Of interest is that para-sites have been used successfully to dis-tinguish haddock stocks in the north-east Atlantic. Lubieniecki (1977) ana-lyzed the incidence and intensity of in-festation with Grillotia erinaceusplerocerci to indicate a number of sepa-rate stocks of haddock within this re-gion. In contrast, Scott (1981) foundthat G. erinaceus was not a major para-site in haddock from the northwest At-lantic, thereby reducing its utility as abiological tag in these waters.
Genetics
Genetic information obtained frommitochondrial DNA procedures has notbeen considered in construction of thepresent management units of haddockstock structure in the northwest Atlan-tic (Zwanenburg et al., 1992), althoughgenetically discrete stocks have beenidentified using electrophoretic tech-niques in northeastern Atlantic waters(Jamieson and Birley, 1988). Impor-tantly, the stock structure of haddockidentified by the genetic results ofJamieson and Birley (1988) was con-
sistent with the parasitic study ofLubieniecki (1977).
Examination of mitochondrial DNAin haddock from the northwest Atlantichas provided conflicting information onthe stock structure of the species in thisregion. Zwanenburg et al. (1992) deter-mined that haddock sampled from off-shore banks of New England (Georges,Div. 5Z), Nova Scotia (Browns, Div.4X; Western, Div. 4W; Banquereau,Div. 4V), and Newfoundland (St. Pierre,Div. 3Ps) were comprised of a mixtureof divergent genotypes that may havearisen in past populations that weremore isolated than those at present. Al-though, no statistically significant dif-ferences in pair-wise comparisons ofgenotype frequencies among haddockfrom any of the banks were detected,gene flow among the population wasconsidered to be restricted. A geo-graphic cline in genotype frequency,increasing genetic differences with geo-graphic distance (Fig. 13), and the deepocean channels acting as barriers togene flow formed the basis for their rea-soning of stock separation. In contrast,Purcell et al. (1996) suggested that had-dock on Georges Bank may not com-prise a genetically discrete stock. Theyhypothesized that significant heteroge-neity in haplotype frequencies observedin haddock samples from the 1975 and1985 cohorts from Georges Bank wascaused by episodic intrusions of ScotianShelf surface water onto the bank re-sulting in larvae from different regionscontributing to the gene pool of haddockin this area. However, Purcell et al.
(1996) also recognized that heteroge-neity in mtDNA markers may have beendue to mixing of the spawning aggre-gations on the Northeast Peak and Nan-tucket Shoals.
Discussion
Current management units for had-dock in the northwest Atlantic tend toencompass discrete populations identi-fied on a biological stock basis, al-though further investigation into thestock structure of haddock in New En-gland and Nova Scotian waters is re-quired (Table 2). Correspondence of thespecies’ biological stock structure withthat of its current management units waslargely the result of the overriding im-portance of haddock and Atlantic cod,Gadus morhua, to the fishery when thestock boundaries in the northwest At-

12 Marine Fisheries Review
lantic were originally identified in 1932by the North American Council on Fish-ery Investigations, and in 1951 by theInternational Commission for theNorthwest Atlantic Fisheries, as infor-mation on stock separation was onlyavailable for these two species at thosetimes (Halliday and Pinhorn, 1990).
Stock Structure
Throughout the northwest Atlantic,haddock stock structure is divided intothree components (New England, NovaScotia, Newfoundland) by the Fundianand Laurentian Channels that act asbarriers to dispersal (Needler, 1930).Within each of these major populationdivisions, a number of separate haddockstocks exist.
Tag-recapture data, growth rate infor-mation, and spawning and circulationpatterns within New England watersindicate that there is a resident stock ofhaddock on Georges Bank (Div. 5Z) anda separate seasonally migrating stockalong the coast of the Gulf of Maine(Div. 5Y) (Needler, 1930; Schroeder,1942; Grosslein, 1962; Smith andMorse, 1985). Growth and spawningdata suggest that a discrete stock mayalso be present in the Nantucket Shoalsregion (Needler, 1930; Smith andMorse, 1985), although haddock fromthis area may be a mixture of fish fromthe Gulf of Maine and Georges Bank.Separation of these stocks may be en-hanced by the Great South Channel, al-though it probably does not have thesame effect on dispersal as the othermuch deeper channels in the northwestAtlantic. A considerable degree of un-certainty still remains in the currentviews of haddock stock structure withinNew England waters, particularly overthe discreteness of the Nantucket Shoalspopulation and, to a lesser extent, thatin the Gulf of Maine.
A range of stock identification tech-niques suggests a complex stock struc-ture is evident for haddock along theScotian Shelf, although there is dis-agreement over the amount of separa-tion (Table 2). At least two major stockdivisions occur in Nova Scotian waters,comprising haddock from an eastern(Div. 4TVW) and western Scotian Shelfstock (Div. 4X) (Martin, 1953; Grosslein,
1962; Bowen, 1987). These stocks ap-pear to be relatively distinct with lim-ited mixing between them except alongthe coast during summer when haddockfrom inshore waters of the western stockmove eastward, and in winter whenthose from the eastern stock movesouthwest (McCracken3). However,Frank (1992) found that the spatial dy-namics of haddock year classes on theScotian Shelf are consistent with a pat-tern of unidirectional mixing of stocksduring the pelagic juvenile stage, thena mixed-stock composition up to the ageof maturation, followed by a return mi-gration of adult fish to their natal sites.Consequently, Frank (1992) suggestedthat the management unit on the ScotianShelf should include both Division 4Xand Division 4VW stocks until the po-tential flux across these stock boundariesis resolved.
Haddock from the eastern ScotianShelf stock appear to be closely relatedto those in the southern Gulf of St.Lawrence, and probably belong to thesame stock (Halliday, 1971), althoughcomparative data for these regions arelimited. Lack of biological data forstock discrimination from the easternScotian Shelf and the southern Gulf ofSt. Lawrence precludes confirmation ofthese purported stock units. Few had-dock have been tagged in these watersto determine movement patterns, andsamples required for demographic andspawning patterns have been limited.Uncertainty exists over the stock statusof inshore haddock from the southernScotian Shelf and Bay of Fundy stock(Div. 4X); however, haddock in the lat-ter area may be a mixture from the Gulfof Maine and western Scotian Shelfstocks.
Although little tagging data exist forhaddock in Newfoundland waters, east-ern Scotian Shelf waters, and the south-ern Gulf of St. Lawrence, it is reason-able to assume that there is little inter-change between haddock from theseareas and those to the southwest, as theLaurentian Channel is probably an evenmore effective barrier than the FundianChannel since it is considerably widerand deeper (Needler, 1930). WithinNewfoundland waters, two main had-dock stocks exist, one on Grand Bank
(Div. 3LNO) and the other on St. PierreBank (Div. 3Ps) (Table 2). Haddockfrom these two areas are thought to notmix extensively and are considered tobe separate stocks based on persistentdifferences in growth rates and yearclass compositions (Templeman, 1953;Grosslein, 1962; Templeman andBishop, 1979a; Halliday and Pinhorn,1990).
Stock Identification Techniques
Combined information from tag-re-capture, demographic, spawning, re-cruitment, meristic, parasitic, and ge-netic studies have provided evidence forthe identification of haddock stocksthroughout the northwest Atlantic(Table 2). Tag-recapture data identifieda broad-scale pattern of haddock stockstructure in the northwest Atlantic, butprovided no conclusive information onstock structure within each of the ma-jor regions (Needler, 1930; Schroeder,1942; McCracken, 1960; Halliday andMcCracken, 1970). Recapture data,however, first indicated that New En-gland and Nova Scotian stocks may notbe homogeneous units, but instead maybe comprised of a number of separatestocks (Needler, 1930; Halliday andMcCracken, 1970).
Growth differences between haddockfrom regions within the northwest At-lantic provided evidence in support oftagging results, which indicated thathaddock may return to the same local-ity to spawn (Needler, 1930; Schuck andArnold, 1951), resulting in stocks be-ing reproductively isolated and demo-graphic differences subsequently main-tained. Zwanenburg et al. (1992) alsosuggested that haddock home with highfidelity to their natal banks to spawn,thereby maintaining stock separation.Consequently, the amount of mixingbetween different parts of the popula-tion of the different regions may not besufficient to mask local stock differ-ences (Needler, 1930).
Meristic studies agreed with the gen-eral stock structure of haddock pro-posed by Needler (1930), although theyindicated that there may be three to fourseparate stocks within Nova Scotianwaters (Clark and Vladykov, 1960;Tremblay et al., 1984; Vladykov7). In

60(4), 1998 13
9 Begg, G. A., and J. R. Waldman. An holisticapproach to fish stock identification. Manuscr.in review.
contrast, conflicting genetic results havenot improved our understanding of had-dock stock structure in the northwestAtlantic, because mtDNA proceduresmay not be sensitive enough to identifyfine-scale population structure (Purcellet al., 1996). The incongruity betweenthe studies of Zwanenburg et al. (1992)and Purcell et al. (1996) is probablypartly responsible for the lack of con-sideration of genetics in the delineationand management of the species stocks.The utility of genetic-based methods inproviding a clearer resolution of had-dock stock structure will require con-siderably more samples (Purcell et al.,1996), both of a temporal and spatialcomponent. In addition, a detailed un-derstanding of the extent of both cur-rent and historical divergence of had-dock stocks will enable assessment ofhistorical changes in stock structure(Purcell et al., 1996).
The various techniques used to iden-tify stock structure of haddock in thenorthwest Atlantic have tended to agreeon the major stock divisions betweenNew England, Nova Scotia, and New-foundland waters, but they have differedpartly in the degree of separation foundwithin each of these regions (Table 2).These differences are probably relatedto the sensitivity of each method in de-tecting stock separation and the limita-tions associated with each technique.Tagging studies of haddock have gen-erally been restricted to inshore watersin spring and summer, with few fishhaving been tagged on important off-shore banks where major spawning ag-gregations occur (Needler, 1930;Schroeder, 1942; McCracken, 1960;Halliday and McCracken, 1970). Con-sequently, the degree of interchangebetween stocks from inshore and off-shore fishing grounds and their relativediscreteness cannot be determined fromthese studies. Typically, the demo-graphic studies have only presented av-erage length-at-age data, thereby pre-cluding examination of the amount ofvariability in the data and the level ofsignificance used to determine differ-ences in stock dynamics (Needler,1930). Gear selectivity problems, dif-ferences in sampling times, and the gen-eral lack of homogeneity in samples
have also tended to confound the resultsof these studies (Schuck and Arnold,1951; Hennemuth et al., 1964).
Spawning (Grosslein and Henne-muth, 1973; Sherman et al., 1984; Smithand Morse, 1985; Lough and Bolz,1989), recruitment (Clark et al., 1982;Koslow, 1984; Koslow et al., 1987; Th-ompson and Page, 1989), meristic(Clark and Vladykov, 1960; Tremblayet al., 1984; Vladykov7), parasitic(Scott, 1981), and genetic (Zwanenburget al., 1992; Purcell et al., 1996) stockidentification studies of haddock haveusually been restricted in their tempo-ral and spatial comparisons, effectivelypreventing examination of the tempo-ral persistency in stock structure pat-terns. Inadequate spatial samples, par-ticularly from inshore waters, havemade it difficult to determine the con-nectivity and relative separation ofstocks found on inshore and offshorefishing grounds, a topic that requiresconsiderably more investigation if afiner resolution of haddock stock struc-ture in the northwest Atlantic is to beachieved.
Future Research
Stock identification is a necessaryprecursor for effective fisheries man-agement (Kutkuhn, 1981). Although anumber of haddock stocks in the north-west Atlantic have been identified us-ing a combination of traditional tech-niques, uncertainty remains in the dis-creteness of stocks in New England andCanadian waters. This lack of under-standing of haddock stock structure canlimit the ability to develop and imple-ment effective stock rebuilding pro-grams throughout the region. Suchknowledge is useful for setting manage-ment restrictions in fisheries which con-tain several stocks with different levelsof exploitation, as less productive stocksmay be seriously depleted or eliminatedif exploited with fishing rates that ad-equately exploit more productive stocksin a mixed fishery (Ricker, 1958).
Future research examining the stockstructure of haddock in these regionsshould advance existing studies by uti-lizing more recent, innovative stockidentification techniques such as chemi-cal analysis of calcified structures that
have proved useful for differentiatingamong Atlantic cod stocks (Campanaet al., 1994). Contemporary stock iden-tification tools such as otolith markingand image analysis procedures shouldalso be investigated to determine theirutility for discrimination of haddockstocks in the northwest Atlantic. A com-bination of techniques should be usedin unison to strengthen and confirm anysuggested stock structure provided bya single procedure in isolation owing tothe inadequacies associated with anyparticular method (i.e. an integratedholistic approach to stock identification)(Begg and Waldman9).
Population dynamic models used infisheries management need to be devel-oped that incorporate multistock com-plexes and demographic consequencesof dispersal, such as the more recentmetapopulation models (Frank, 1992).Such models may be particularly rel-evant for haddock stocks along theScotian Shelf and in other regions of thenorthwest Atlantic that appear to mi-grate and mix with other stocks at cer-tain times of the year. Although mostfisheries are managed on a single-spe-cies basis, there is a general recogni-tion that multispecies interactions arean important component of marine eco-system dynamics which should be con-sidered in contemporary managementplans (Mahon and Smith, 1989). Like-wise, multistock models, as is evidentfor haddock in the northwest Atlantic,should be developed and applied in asimilar context to multispecies models.Conceptually, species and stocks can beviewed as the same unit or level of hi-erarchy in these models, although bydefinition of a species, more gene flowwould be expected between stocks ofthe same species than between species,but treatment of the two units could bethe same.
Stock structure information providesa basis for understanding the dynamicsof fish populations that assists scientistsand managers in predicting how a stockmay respond to different managementstrategies. Investigation into the tempo-

14 Marine Fisheries Review
ral stability and historical shifts in stockstructure of haddock throughout thenorthwest Atlantic via archived biologi-cal samples and data sets in relation toenvironmental fluctuations and effectsof the fisheries, will enable an histori-cal perspective of stock structure to bedeveloped. This would assist in under-standing the collapse of haddock stocksthroughout their distribution and theimplications of these effects to currentstock assessment, rebuilding, and man-agement plans. Consequently, identifi-cation of haddock stocks and their ratesof mixing should continue to undergoinvestigation in order to refine our un-derstanding of haddock stock structurein the northwest Atlantic.
Acknowledgments
Thanks to Fred Serchuk, StephenClark, Ralph Halliday, Ken Frank, KeesZwanenburg, Russell Brown, WilliamOverholtz, and John Boreman for criti-cal comments, suggestions, and reviewsof this manuscript; to Peter Hurley forDivision 4X parameters; and to BrendaFiguerido for assistance with the fig-ures. This work was performed whilethe author held a National ResearchCouncil (NMFS, Northeast Fisheries Sci-ence Center) Research Associateship.
Literature CitedAnonymous. 1995. Status of the fishery resources
off the northeastern United States for 1994.U.S. Dep. Commer., NOAA Tech. Memo.NMFS-NE-108, 140 p.
Beacham, T. D. 1983. Variability in size and ageat sexual maturity of haddock (Melano-grammus aeglefinus) on the Scotian Shelf inthe Northwest Atlantic. Can. Tech. Rep. Fish.Aquat. Sci. 1168, 33 p.
Bigelow, H. B., and W. C. Schroeder. 1953. Fishesof the Gulf of Maine. Fish. Bull. 53, 533 p.
Bowen, W. D. 1987. A review of stock structurein the Gulf of Maine area: a workshop report.Can. Atl. Fish. Sci. Advis. Comm. Res. Doc.87/21, 51 p.
Campana, S. E., A. J. Fowler, and C. M. Jones.1994. Otolith elemental fingerprinting forstock identification of Atlantic cod (Gadusmorhua) using laser ablation ICPMS. Can. J.Fish. Aquat. Sci. 51:1942–1950.
________ , S. J. Smith, and P. C. F. Hurley. 1989.A drift-retention dichotomy for larval haddock(Melanogrammus aeglefinus) spawned onBrowns Bank. Can. J. Fish. Aquat. Sci. 46:93–102.
Clark, J. R., and V. D. Vladykov. 1960. Defini-tion of haddock stocks of the northwesternAtlantic. Fish. Bull. 60:283–296.
Clark, S. H., W. J. Overholtz, and R. C.Hennemuth. 1982. Review and assessment ofthe Georges Bank and Gulf of Maine haddock
fishery. J. Northwest Atl. Fish. Sci. 3:1–27.Cohen, E. B., D. G. Mountain, and R. O’Boyle.
1991. Local-scale versus large-scale factorsaffecting recruitment. Can. J. Fish. Aquat. Sci.48:1003–1006.
Colton, J. B., Jr., and R. F. Temple. 1961. Theenigma of Georges Bank spawning. Limnol.Oceanogr. 6:280–291.
Drinkwater, K. F. 1996. Atmospheric and oce-anic variability in the Northwest Atlantic dur-ing the 1980s and early 1990s. J. NorthwestAtl. Fish. Sci. 18:77–97.
Frank, K. T. 1992. Demographic consequencesof age-specific dispersal in marine fish popu-lations. Can. J. Fish. Aquat. Sci. 49:2222–2231.
________ , R. K. Mohn, and J. E. Simon. 1997.Assessment of 4TVW haddock in 1996. Can.Dep. Fish. Oceans, Atl. Fish. Res. Doc. 97/107, 90 p.
________ , F. H. Page, and J. K. McRuer. 1989.Hydrographic effects on the vertical distribu-tion of haddock (Melanogrammus aeglefinus)eggs and larvae on the southwestern ScotianShelf. Can. J. Fish. Aquat. Sci. 46 (Suppl.1):82–92.
Gavaris, S., and L. Van Eeckhaute. 1998. Assess-ment of haddock on eastern Georges Bank.Can. Dep. Fish. Oceans, DFO Atl. Fish. Res.Doc. 98/66, 75 p.
Grosslein, M. D. 1962. Haddock stocks in theICNAF convention area. ICNAF RedbookIII:124–131.
________ and R. C. Hennemuth. 1973. Spawn-ing stock and other factors related to recruit-ment of haddock on Georges Bank. Rapp. P.-v. Reun. Cons. Int. Explor. Mer. 164:77–88.
Halliday, R. G. 1971. Recent events in the had-dock fishery of the eastern Scotian Shelf. Int.Comm. Northwest Atl. Fish. Res. Bull. 8:49–58.
________ and F. D. McCracken. 1970. Move-ments of haddock tagged off Digby, NovaScotia. Int. Comm. Northwest Atl. Fish. Res.Bull. 7:8–14.
________ and A. T. Pinhorn. 1990. The delimi-tation of fishing areas in the northwest Atlan-tic. J. Northwest Atl. Fish. Sci. 10:1–51.
Hennemuth, R. C., M. D. Grosslein, and F. D.McCracken. 1964. Abundance, age composi-tion of landings, and total mortality of had-dock caught off southern Nova Scotia, 1956–1961. Int. Comm. Northwest Atl. Fish. Res.Bull. 1:43–73.
Hurley, P. C. F., G. A. P. Black, R. K. Mohn, andP. Comeau. 1997. Assessment of 4X haddockin 1996 and the first half of 1997. Can. Dep.Fish. Oceans, Atl. Fish. Res. Doc. 97/108,101 p.
________ and S. E. Campana. 1989. Distribu-tion and abundance of haddock (Melano-grammus aeglefinus) and Atlantic cod (Ga-dus morhua) eggs and larvae in the waters offsouthwest Nova Scotia. Can. J. Fish. Aquat.Sci. 46 (Suppl. 1):103–112.
ICNAF. 1967. Statistical bulletin. Int. Comm.Northwest Atl. Fish. Stat. Bull. 15, 95 p.
Ihssen, P. E., H. E. Booke, J. M. Casselman, J.M. McGlade, N. R. Payne, and F. M. Utter.1981. Stock identification: materials andmethods. Can. J. Fish. Aquat. Sci. 38:1838–1855.
Jamieson, A., and A. J. Birley. 1989. The distri-bution of transferrin alleles in haddock stocks.J. Cons. Cons. Int. Explor. Mer 45:248–262.
Koslow, J. A. 1984. Recruitment patterns innorthwest Atlantic fish stocks. Can. J. Fish.Aquat. Sci. 41:1722–1729.
________ , K. R. Thompson, and W. Silvert.1987. Recruitment to northwest Atlantic cod(Gadus morhua) and haddock (Melano-grammus aeglefinus) stocks: influence ofstock size and climate. Can. J. Fish. Aquat.Sci. 44:26–39.
Kutkuhn, J. H. 1981. Stock definition as a nec-essary basis for cooperative management ofGreat Lakes fish resources. Can. J. Fish.Aquat. Sci. 38:1476–1478.
Loder, J. W., C. K. Ross, and P. C. Smith. 1988.A space- and time-scale characterization ofcirculation and mixing over submarine banks,with application to the Northwestern AtlanticContinental Shelf. Can. J. Fish. Aquat. Sci.45:1860–1885.
Lough, R. G., and G. R. Bolz. 1989. The move-ment of cod and haddock larvae onto theshoals of Georges Bank. J. Fish Biol. 35(Suppl. A):71–79.
Lubieniecki, B. 1977. The plerocercus ofGrillotia erinaceus as a biological tag for had-dock Melanogrammus aeglefinus in the NorthSea and north-east Atlantic. J. Fish Biol.11:555–565.
Mahon, R., and R. W. Smith. 1989. Demersal fishassemblages on the Scotian Shelf, northwestAtlantic: spatial distribution and persistence.Can. J. Fish. Aquat. Sci. 46 (Suppl. 1):134–152.
Marak, R. R., and R. Livingstone, Jr. 1970.Spawning dates of Georges Bank haddock.Int. Comm. Northwest Atl. Fish. Res. Bull.7:56–58.
Martin, W. R. 1953. Identification of majorgroundfish stocks in subarea 4 of the North-west Atlantic convention area. Int. Comm.Northwest Atl. Fish. Annu. Proc. 3:57–61.
McCracken, F. D. 1960. Studies of haddock inthe Passamaquoddy Bay region. J. Fish. Res.Board Can. 17:175–180.
Murphy, E. F., and C. A. Bishop. 1995. The sta-tus of 2GH cod, 3LNO haddock and 3Ps pol-lock. Can. Dep. Fish. Oceans, Atl. Fish. Res.Doc. 95/33, 30 p.
NAFO. 1995. Statistical bulletin. Northwest Atl.Fish. Organ. Stat. Bull. 42, 310 p.
Needler, A. W. H. 1930. The migrations of had-dock and the interrelationships of haddockpopulations in North American waters.Contrib. Can. Biol. 6:241–313.
O’Boyle, R. N., J. Simon, and K. T. Frank. 1988.An evaluation of the population dynamics of4X haddock during 1962–88 with yield pro-jected to 1989. Can. Atl. Fish. Sci. Advis.Comm. Res. Doc. 88/72, 59 p.
________ , M. Sinclair, R. J. Conover, K. H.Mann, and A. C. Kohler. 1984. Temporal andspatial distribution of ichthyoplankton com-munities of the Scotian Shelf in relation tobiological, hydrological, and physiographicfeatures. Rapp. P.-v. Reun. Cons. Int. Explor.Mer 183:27–40.
O’Brien, L., J. Burnett, and R. K. Mayo. 1993.Maturation of nineteen species of finfish offthe northeast coast of the United States, 1985–1990. U.S. Dep. Commer., NOAA Tech. Rep.NMFS 113, 66 p.
Ovenden, J. R. 1990. Mitochondrial DNA andmarine stock assessment: a review. Aust. J.Mar. Freshwater Res. 41:835–853.
Overholtz, W. J. 1987. Factors relating to the re-productive biology of Georges Bank haddock(Melanogrammus aeglefinus) in 1977–83. J.Northwest Atl. Fish. Sci. 7:145–154.
Page, F. H., and K. T. Frank. 1989. Spawningtime and egg stage duration in Northwest At-lantic haddock (Melanogrammus aeglefinus)

60(4), 1998 15
stocks with emphasis on Georges and BrownsBank. Can. J. Fish. Aquat. Sci. 46 (Suppl. 1):68–81.
________ , ________ , and K. R. Thompson.1989. Stage dependent vertical distribution ofhaddock (Melanogrammus aeglefinus) eggs ina stratified water column: observations andmodel. Can. J. Fish. Aquat. Sci. 46 (Suppl.1):55–67.
Polacheck, T., D. Mountain, D. McMillan, W.Smith, and P. Berrien. 1992. Recruitment ofthe 1987 year class of Georges Bank haddock(Melanogrammus aeglefinus): the influence ofunusual larval transport. Can. J. Fish. Aquat.Sci. 49:484–496.
Purcell, M. K., I. Kornfield, M. Fogarty, and A.Parker. 1996. Interdecadal heterogeneity in mi-tochondrial DNA of Atlantic haddock (Melano-grammus aeglefinus) from Georges Bank. Mo-lecular Mar. Biol. Biotech. 5:185–192.
Ricker, W. E. 1958. Maximum sustained yieldfrom fluctuating environments and mixedstocks. J. Fish. Res. Board Can. 15:991–1006.
Schroeder, W. C. 1942. Results of haddock tag-ging in the Gulf of Maine from 1923 to 1932.J. Mar. Res. 5:1–19.
Schuck, H. A., and E. L. Arnold, Jr. 1951. Com-parison of haddock from Georges and Brownsbanks. Fish. Bull. 52:177–185.
Scott, J. S. 1981. Alimentary tract parasites ofhaddock (Melanogrammus aeglefinus L.) onthe Scotian Shelf. Can. J. Zool. 59:2244–2252.
Sherman, K., W. Smith, W. Morse, M. Berman,J. Green, and L. Ejsymont. 1984. Spawning
strategies of fishes in relation to circulation,phytoplankton production, and pulses in zoop-lankton off the northeastern United States.Mar. Ecol. Prog. Ser. 18:1–19.
Smith, P. C. 1989. Circulation and dispersion onBrowns Bank. Can. J. Fish. Aquat. Sci.46:539–559.
Smith, W. G., and W. W. Morse. 1985. Retentionof larval haddock Melanogrammus aeglefinusin the Georges Bank region, a gyre-influencedspawning area. Mar. Ecol. Prog. Ser. 24:1–13.
Templeman, W. 1953. II. Knowledge of divisionsof stocks of cod, haddock, redfish and Ameri-can plaice in Subareas 3 and 2 of the North-west Atlantic convention area. Int. Comm.Northwest Atl. Fish. Annu. Proc. 3:62–66.
________ and C. A. Bishop. 1979a. Age, growth,year-class strength, and mortality of haddock,Melanogrammus aeglefinus, on St. PierreBank in 1948–75 and their relation to the had-dock fishery of this area. Int. Comm. North-west Atl. Fish. Res. Bull. 14:85–99.
________ and ________ . 1979b. Sexual maturityand spawning in haddock, Melanogrammusaeglefinus, of St. Pierre Bank. Int. Comm. North-west Atl. Fish. Res. Bull. 14:77–84.
________ , V. M. Hodder, and R. Wells. 1978a.Age, growth, year-class strength, and mortal-ity of the haddock, Melanogrammus aegle-finus, on the southern Grand Bank and theirrelation to the haddock fishery of this area.Int. Comm. Northwest Atl. Fish. Res. Bull.13:31–52.
________ , ________ , and ________ . 1978b.Sexual maturity and spawning in haddock,Melanogrammus aeglefinus, of the southernGrand Bank. Int. Comm. Northwest Atl. Fish.Res. Bull. 13:53–65.
Thompson, K. R., and F. H. Page. 1989. Detect-ing synchrony of recruitment using short,autocorrelated time series. Can. J. Fish. Aquat.Sci. 46:1831–1838.
Tremblay, M. J., R. Mahon, and R. O’Boyle.1984. Vertebral numbers in larval and juve-nile haddock of the Scotian Shelf and impli-cations for stock discrimination. J. NorthwestAtl. Fish. Sci. 5:213–217.
Trippel, E. A., M. J. Morgan, A. Fréchet, C.Rollet, A. Sinclair, C. Annand, D. Beanlands,and L. Brown. 1997. Changes in age andlength at sexual maturity of northwest Atlan-tic cod, haddock and pollock stocks, 1972–1995. Can. Tech. Rep. Fish. Aquat. Sci. 2157,120 p.
Waiwood, K. G., and M.-I. Buzeta. 1989. Re-productive biology of southwest Scotian Shelfhaddock (Melanogrammus aeglefinus). Can.J. Fish. Aquat. Sci. 46 (Suppl. 1):153–170.
Walford, L. A. 1938. Effects of currents on dis-tribution and survival of the eggs and larvaeof the haddock (Melanogrammus aeglefinus)on Georges Bank. Fish. Bull. 49:1–73.
Zwanenburg, K. C. T., P. Bentzen, and J. M.Wright. 1992. Mitochondrial DNA differen-tiation in western north Atlantic populationsof haddock (Melanogrammus aeglefinus).Can. J. Fish. Aquat. Sci. 49:2527–2537.