a supramolecular endosomal escape approach for enhancing gene silencing of sirna using...
TRANSCRIPT

www.rsc.org/MaterialsBRegistered Charity Number 207890
Showcasing work from Professor Nobuhiko
Yui’s Laboratory, Institute of Biomaterials and
Bioengineering Tokyo Medical and Dental University,
Japan.
Title: A supramolecular endosomal escape approach for
enhancing gene silencing of siRNA using acid-degradable
cationic polyrotaxanes
This work demonstrates the acid-degradable cationic polyrotaxanes
bearing 3-sulfanylpropionyl ester linkages for the cytoplasmic
delivery of siRNA. In response to acidic pH in endosomes, the
ester linkages of the polyrotaxanes were hydrolyzed and released
threaded α-cyclodextrins to induce endosomal membrane
destabilization.
As featured in:
See A. Tamura et al.,
J. Mater. Chem. B, 2013, 1, 3535.

Journal ofMaterials Chemistry B
PAPER
Publ
ishe
d on
21
May
201
3. D
ownl
oade
d by
Uni
vers
ity o
f R
egin
a on
27/
10/2
014
01:2
4:19
.
View Article OnlineView Journal | View Issue
Department of Organic Biomaterials, Instit
Tokyo Medical and Dental University, 2-
101-0062, Japan. E-mail: [email protected]
5280-8020
† Electronic supplementary informationthe synthesis of acid-degradable polyrotaof acid-degradable PRXs at pH 7.4 andwith 21DMAE–COO–PRX/Alexa647–siRNA
Cite this: J. Mater. Chem. B, 2013, 1,3535
Received 10th April 2013Accepted 21st May 2013
DOI: 10.1039/c3tb20514b
www.rsc.org/MaterialsB
This journal is ª The Royal Society of
A supramolecular endosomal escape approach forenhancing gene silencing of siRNA using acid-degradable cationic polyrotaxanes†
Atsushi Tamura and Nobuhiko Yui*
Acid-degradable cationic polyrotaxanes (PRXs) composed of N,N-dimethylaminoethyl (DMAE) group-
modified a-cyclodextrins (CDs) that were threaded onto a poly(ethylene glycol) capped with a bulky
stopper via acid-cleavable 3-sulfanylpropionyl ester linkages (DMAE–COO–PRX) were developed for
improving the cytoplasmic transportation of PRX/siRNA polyplexes. In response to acidic pH in
endosomes, the ester linkages of the DMAE–COO–PRXs were hydrolyzed and an abundance of DMAE-
modified a-CDs was released from the PRXs. Wherein, the released DMAE-modified a-CDs showed
endosomal membrane destabilization through the removal of phospholipids from the membrane,
resulting in the endosomal escape of siRNA. The confocal fluorescence microscopic observation revealed
that the colocalization ratio of siRNAs to endosomes decreased for DMAE–COO–PRX in comparison with
disulfide-introduced PRX (DMAE–SS–PRX). As a result, the DMAE–COO–PRX/siRNA showed higher gene
silencing activity than DMAE–SS–PRX and linear poly(ethylenimine) (L-PEI) even at low siRNA
concentration (10 nM). Therefore, the DMAE–COO–PRXs are an effective candidate as a carrier for
cytoplasmic siRNA delivery.
1 Introduction
RNA interference technology using synthetic small interferingRNA (siRNA) has received tremendous attention as an analyticaltool for genomic function or drug screening, and the centralpart of interest is a therapeutic use by the down-regulation ofpathogenic genes.1–3 Nowadays, several clinical trials areongoing in view of realizing the clinical application of siRNA.4
Since the successful gene silencing largely depends on thesiRNA delivery system, the establishment of effective siRNAcarriers is a challenging issue. To date, a wide variety of siRNAdelivery carriers based on a polyelectrolyte complex, a self-assembly of negatively charged siRNA and positively chargedpolymers, liposomes, or nanoparticles, have been developed.5–7
Previously, we developed cytocleavable cationic polyrotaxanes(PRXs) for improving the intracellular release efficiency ofplasmid DNA (pDNA) and siRNA.8–11 This PRX is composed ofN,N-dimethylaminoethyl (DMAE) group-modied a-cyclodex-trins (CDs) that are threaded onto a poly(ethylene glycol) (PEG)capped with a terminal bulky N-benzyloxycarbonyl-L-tyrosine
ute of Biomaterials and Bioengineering,
3-10 Kanda-Surugadai, Chiyoda, Tokyo
.jp; Fax: +81-3-5280-8027; Tel: +81-3-
(ESI) available: Detailed procedure forxanes, the time-course of SEC proles5.5, and CLSM image of cells treated. See DOI: 10.1039/c3tb20514b
Chemistry 2013
(Z-Tyr) through disulde linkages (DMAE–SS–PRX). In the caseof siRNA delivery, we have found that the stability of DMAE–SS–PRX/siRNA polyplexes against a polyanion exchange reaction,an intracellular uptake level of siRNA, and a gene silencingactivity can be signicantly improved by optimizing the numberof threading CDs on PRX. In particular, the DMAE–SS–PRX witha relatively high number of threading CDs (CD coverage of morethan ca. 40%) is found to form a stable polyplex with siRNA, andthe intracellular uptake level of siRNA with the aid of theDMAE–SS–PRX is approximately 27-fold higher than that withthe aid of linear poly(ethylenimine) (L-PEI) (i.e., commerciallyavailable transfection reagent, ExGen 500). Additionally, theintracellularly degradable character of the DMAE–SS–PRXfacilitates the gene silencing activity by the intracellular releaseof siRNA from the polyplex.11 However, despite the signicantdifference in intracellular uptake of siRNA, the DMAE–SS–PRXand L-PEI show comparable gene silencing activity, most likelydue to the lack of endosomal escape capability of the DMAE–SS–PRX. Since the disulde linkage is mainly cleaved by gluta-thione located in the cytoplasm, the release of siRNA frompolyplexes is limited in endosomes.11,12 Therefore, PRX/siRNApolyplexes are highly required to overcome the endosomalbarrier for improving the gene silencing efficiency.
To date, various endosomal escape methodologies based onpolymeric materials have been proposed. Inside the endosomesand lysosomes, the pH values are known to be lowered to 4.5 to6.5 by the function of vacuolar H+–ATPase.13–15 In this regard,polyelectrolytes with specic endosomal membrane disruptive
J. Mater. Chem. B, 2013, 1, 3535–3544 | 3535

Journal of Materials Chemistry B Paper
Publ
ishe
d on
21
May
201
3. D
ownl
oade
d by
Uni
vers
ity o
f R
egin
a on
27/
10/2
014
01:2
4:19
. View Article Online
activity in an acidic environment, such as polycations with lowpKa value to destabilize the membrane through the protonsponge effect16,17 and pH-responsive membrane disruptivepolyelectrolytes,18–20 are of interest for facilitating endosomalescape. Meanwhile, a- and b-CDs are also known to destabilizethe plasma membranes by the removal of phospholipids andcholesterols through the incorporation into their hydrophobiccavity.21–24 Arima and co-workers have reported that the conju-gation of a-CDs with poly(amidoamine) dendrimers signi-cantly enhanced the gene expression of pDNA and genesilencing of siRNA, presumably due to the promotion of endo-somal escape by the membrane destabilizing activity of conju-gated a-CDs.25,26We have also conrmed that gene expression ofpoly(3-lysine)/pDNA polyplexes was remarkably increased by theconjugation of b-CDs on the side chain of poly(3-lysine).27
Likewise, various CD-conjugated polycations are designed for invitro and in vivo delivery of pDNA and siRNA.28–30 Accordingly, itis a possible endosomal escape strategy in our system that thesite specic release of threaded a-CDs from PRXs in endosomesand lysosomes can induce the membrane destabilization toensure cytoplasmic siRNA transportation.
Herein, we described a unique supramolecular approachtoward endosomal escape of PRX/siRNA polyplexes using the acid-degradable cationic PRX bearing terminal 3-sulfanylpropionylesters (DMAE–COO–PRX) (Fig. 1A). The 3-sulfanylpropionyl
Fig. 1 (A) Chemical structure of acid-degradable DMAE–COO–PRX. (B) SchematicsiRNA, the dissociation of the DMAE–COO–PRX/siRNA complex by the hydrolysisdestabilization by released a-CDs.
3536 | J. Mater. Chem. B, 2013, 1, 3535–3544
(or b-thiopropionate) linkage is sufficiently stable at physiologicalpH, whereas it is hydrolyzed at acidic pH.31–33 Since the PRXstructure hinders the exposure of the hydrophobic cavity of a-CDsthrough the threading of a PEG chain, it is expected to mask themembrane destabilizing activity of a-CDs under extracellularconditions. In contrast, aer cellular internalization of theDMAE–COO–PRXs through endocytosis, the DMAE–COO–PRXsare thought to be degraded into constituent molecules (i.e., PEGand a-CD) by hydrolysis and release an abundance of threadeda-CDs in acidic endosomes and lysosomes. Therefore, selectiveendosomal membrane destabilization of a-CDs may be achievedby increasing the local concentration of a-CD in endosomes. Inthis study, the pH-dependent membrane destabilization and theendosomal escape capabilities of acid-degradable DMAE–COO–PRXs were investigated, and the gene silencing activity of DMAE–COO–PRX/siRNA polyplexes was discussed in comparison withour previous studies on reduction-degradable DMAE–SS–PRXs.
2 Materials and methods2.1 Reagents
The siRNA duplexes against rey luciferase (sense: 50-CUUACG CUG AGU ACU UCG AdTdT-30, antisense: 50-UCG AAG UACUCA GCG UAA GdTdT-30) and the siRNA duplexes withscrambled sequence (sense: 50-GGC UAC GUC CAG GAG CGC
illustration of polyelectrolyte complex formation between DMAE–COO–PRX andof DMAE–COO–PRX in acidic endosomes/lysosomes, and endosomal membrane
This journal is ª The Royal Society of Chemistry 2013

Paper Journal of Materials Chemistry B
Publ
ishe
d on
21
May
201
3. D
ownl
oade
d by
Uni
vers
ity o
f R
egin
a on
27/
10/2
014
01:2
4:19
. View Article Online
AdTdT-30, antisense: 50-UGC GCU CCU GGA CGU AGC CdTdT-30) were obtained from Thermo Fisher Scientic (Lafayette, CO,USA). The sense 50-Alexa Fluor 647-labeled siRNA duplex againstrey luciferase (Alexa647–siRNA) (sense: 50-Alexa647-CUU ACGCUG AGU ACU UCG AdTdT-30, antisense: 50-UCG AAG UAC UCAGCG UAA GdTdT-30) was obtained from Japan Bio Services(Saitama, Japan). The plasmid DNA (pDNA) encoding reyluciferase (pGL3-control; 5269 bp) and renilla luciferase (pRL-CMV; 4079 bp) were obtained from Promega (Madison, WI,USA). The pDNAs were amplied in competent DH5a Escher-ichia coli (Takara Bio, Shiga, Japan) and then puried with aQIAlter Plasmid Giga Kit (Qiagen, Hilden, Germany). ExGen500 reagent (22 kDa linear poly(ethylenimine) (L-PEI))was obtained from Fermentes (Ontario, Canada). Ethidiumbromide (EtBr) solution was obtained from Nippon Gene(Tokyo, Japan). 2-[4-(2-Hydroxyethyl)-1-piperazinyl]ethane-sulfonic acid (HEPES) and 2-morpholinoethanesulfonic acidmonohydrate (MES) were obtained from Dojindo Laboratories(Kumamoto, Japan). The Milli-Q water used in this studywas prepared using an Academic A10 ultrapure water system(Millipore, Billerica, MA, USA).
2.2 Synthesis of acid-degradable cationic polyrotaxanes
The homobifunctional PEG–COO–NH2 was synthesized fromPEG–OH (Mn ¼ 4550) by terminal acryloylation and the subse-quent Michael addition with 2-aminoethanethiol hydrochlo-ride. The PEG–COO–NH2 was used for the synthesis of acid-degradable PRXs.8–11 The DMAE–COO–PRXs with variousnumbers of modied DMAE groups were synthesized by varyingthe feed amount of N,N0-carbonyldiimidazole, and subsequentnucleophilic reaction with N,N-dimethylaminoethyl amine aspreviously reported.8–11 The detailed procedure and character-ization were described in the ESI.† The reduction-degradableDMAE–SS–PRX was synthesized as previously described.11
2.3 Hydrolysis kinetics of DMAE–COO–PRX
The 72DMAE–COO–PRX was dissolved in 50 mM HEPES buffersolution at pH 7.4 or 50 mM MES buffer solution at pH 5.5 at aconcentration of 5 mg mL�1 and incubated at 37 �C. Aerincubation for a predetermined time period, aqueous phaseSEC was carried out to determine the amount of remainingPRXs. SEC measurements were carried out on a Gulliver system(Jasco, Tokyo, Japan) equipped with an internal refractive indexdetector (RI-2031 plus, Jasco) and a combination of TSKgela-4000 and a-2500 columns (Tosoh, Tokyo, Japan), eluted with10 mM phosphate buffer containing 700 mM NaCl (pH 7.4) at aow rate of 0.7 mL min�1 at 35 �C. The number-averagedmolecular weight (Mn,SEC) and polydispersity index (Mw/Mn)were calculated based on the standard PEG (Agilent Technolo-gies, Wilmington, DE, USA).
2.4 Preparation and characterization of DMAE–COO–PRX/siRNA polyplexes
The siRNA (siLuc) and the DMAE–COO–PRXs were separatelydissolved in 10 mM HEPES buffer solution at pH 7.4 or 10 mMMES buffer solution at pH 5.5 (siRNA concentration: 10 mM).
This journal is ª The Royal Society of Chemistry 2013
These solutions were mixed at various N/P ratios (molar ratio ofthe amino groups of the DMAE–COO–PRX to the phosphates ofsiRNA) to prepare the polyplexes. The formation of the polyplexwas conrmed by agarose gel electrophoresis aer incubationfor 6 h at 37 �C. The polyplex solutions (10 mL) prepared atvarious N/P ratios were added to 2% agarose gel containing0.2 mg mL�1 of EtBr, and electrophoresis was performed in TAE(40 mM Tris, 20 mM acetic acid, and 1 mM EDTA) buffer for 20min at 100 V. The gel was visualized under a UV trans-illuminator (UVP, Upland, CA, USA) at the excitation wavelengthof 302 nm. The uorescence intensity of the bands was analyzedusing ImageJ soware ver. 1.45 s (National Institutes of Health,Bethesda, MD).
The diameter and zeta potential of the DMAE–COO–PRX/siRNA polyplexes prepared at various N/P ratios were determinedon a Zetasizer Nano ZS (Malvern Instruments, Malvern, UK)equipped with a 4 mWHe–Ne laser (633 nm). The concentrationof siRNA in DMAE–COO–PRX/siRNA polyplex solutions (10 mMHEPES buffer, pH 7.4) was adjusted to 4 mM. The dynamic lightscattering measurements of the DMAE–COO–PRX/siRNA poly-plexes were carried out at 25 �C at a detection angle of 173�. Theobtained autocorrelation functions were analyzed by the cumu-lant method to determine the hydrodynamic diameters. The zetapotential measurements of the DMAE–COO–PRX/siRNA poly-plexes were performed at 25 �C.
2.5 Hemolysis assay
Rat erythrocytes were obtained from Kohjin Bio (Saitama,Japan) and were washed three times with 10 mM HEPES buffercontaining 150 mM NaCl (pH 7.4) or 10 mM MES buffer con-taining 150 mM NaCl (pH 5.5) for adjusting the pH. Theerythrocyte suspensions (2 � 108 cells per mL, 100 mL) wereincubated with PRX solution (100 mL) for 6 h at 37 �C. Aer theincubation, the erythrocytes were separated by centrifugation(2000 rpm for 5 min) and the supernatant (100 mL) wascollected. The amount of released hemoglobin was determinedby reading the absorbance at 544 nm on an ARVOMXmultilabelcounter (Perkin Elmer, Wellesley, MA, USA) using an appro-priate lter set. To determine the absorbance of 100% hemo-lysis, erythrocytes were lysed with 1% Triton-X (Sigma-Aldrich,Milwaukee, WI, USA). The hemolytic activities of DMAE–COO–PRX at pH 7.4 and 5.5 were calculated relative to the 1% Triton-X-treated samples.
2.6 Intracellular uptake analysis by ow cytometry
HeLa cells derived from human cervical carcinoma were grownin minimum essential medium (MEM) (Gibco BRL, GrandIsland, NY, USA) containing 10% fetal bovine serum (FBS)(Gibco), 100 units per mL of penicillin, and 100 mg mL�1 strep-tomycin (Gibco) in a humidied 5% CO2 atmosphere at 37 �C.HeLa cells were seeded on a 24-well plate (BD Falcon, FranklinLakes, NJ, USA) at a density of 2.5 � 104 cells per cm2 andincubated at 37 �C for 24 h in MEM containing 10% FBS. Aerthe medium was exchanged with fresh medium (225 mL), theDMAE–COO–PRX/Alexa647–siRNA polyplex solutions (25 mL)were added to each well (concentration of Alexa647–siRNA:
J. Mater. Chem. B, 2013, 1, 3535–3544 | 3537

Journal of Materials Chemistry B Paper
Publ
ishe
d on
21
May
201
3. D
ownl
oade
d by
Uni
vers
ity o
f R
egin
a on
27/
10/2
014
01:2
4:19
. View Article Online
100 nM). Aer incubation for 48 h, the cells were washed threetimes with phosphate buffered saline (PBS) (Gibco). The cellswere harvested by the treatment with 0.25% trypsin containing0.26 mM EDTA (Gibco), collected by centrifugation (1000 rpm, 4�C, 5min), and suspended in PBS containing 0.1% bovine serumalbumin (BSA) (Sigma-Aldrich) (400 mL). Aer the cells werepassed through a 35 mm cell strainer (BD Falcon), ow cytometrywas performed on a FACSCanto II (BD Biosciences). TheAlexa647–siRNA was excited using a 17 mW He–Ne laser(633 nm) and detected with a 650–670 nm bandpass lter. Intotal 10 000 cells were acquired for each sample and the meanuorescence intensity of cell population was analyzed by DIVAsoware (BD Biosciences).
2.7 In vitro gene silencing by dual luciferase reporter assay
HeLa cells were seeded on a 24-well plate at a density of 2.5 �104 cells per cm2 and incubated overnight in MEM containing10% FBS. Aer the medium was exchanged with Opti-MEM I(Invitrogen, San Diego, CA, USA) (200 mL), pDNA solution (50mL) containing pGL3-control (360 ng), pRL-CMV (40 ng), andLipofectamine 2000 (Invitrogen) (1 mL) were added to each welland the cells were incubated at 37 �C for 4 h. Aer the mediumwas exchanged with fresh MEM containing 10% FBS (225 mL),the DMAE–COO–PRX/siRNA polyplex solutions (25 mL) wereadded to each well (concentration of siRNA: 100 nM). Aerfurther incubation for 48 h, the cells were washed twice withPBS and solubilized with passive lysis buffer (Promega)(100 mL). The relative light unit (RLU) of both the rey and therenilla luciferase were evaluated using a Dual LuciferaseReporter System (Promega) on an ARVOMXmultilabel counter(Perkin Elmer). The relative gene silencing effect of rey lucif-erase was calculated as follows: relative rey luciferase expres-sion (%) ¼ ([RLU of rey luc.]sample/[RLU of renilla luc.]sample)/([RLU of rey luc.]control/[RLU of renilla luc.]control) � 100.The subscripts “sample” and “control” indicate the RLU ofsiRNA treated and non-treated cells, respectively. To evaluatethe effect of endosomal acidication inhibitor on genesilencing activities, the transfected HeLa cells were treated withDMAE–COO–PRX/siRNA polyplexes (concentration of siRNA:100 nM) in the presence of ammonium chloride (NH4Cl) (Wako)for 48 h.
2.8 Cytotoxicity assay
HeLa cells were seeded on a 96-well plate (BD Falcon) at adensity of 2.5 � 104 cells per cm2 and incubated overnight inMEM containing 10% FBS. Aer the medium was exchangedwith fresh MEM (90 mL), the DMAE–PRX/siRNA polyplexsolutions (10 mL) were added to each well. Aer incubationfor a further 48 h, Cell Counting Kit-8 reagent (DojindoLaboratories) (10 mL) was added to each well. Aer 1 h ofincubation at 37 �C, the absorbance at 450 nm was measuredon a Multiskan FC plate reader (Thermo Fisher Scientic,Waltham, MA, USA) equipped with an appropriate band-passlter. The cellular viability was calculated relative to the non-treated cells.
3538 | J. Mater. Chem. B, 2013, 1, 3535–3544
2.9 Intracellular distribution analysis by confocal laserscanning microscopy (CLSM)
HeLa cells were seeded on 35 mm glass-bottom dishes (Iwaki,Tokyo, Japan) at a density of 5� 103 cells per cm2 and incubatedovernight in MEM containing 10% FBS. Aer the medium wasexchanged with fresh MEM (900 mL), the DMAE–COO–PRX/Alexa647–siRNA polyplex (N/P 10) solutions (100 mL) were addedto the dish (concentration of Alexa647–siRNA: 100 nM). Aerincubation for 48 h, the cells were stained with LysoTrackerGreen DND-26 (Molecular Probes, Eugene OR, USA) (1 mM) for30 min at 37 �C. Then, the cells were stained with Hoechst33342 (Dojindo Laboratories) (1 mg mL�1) for 10 min at 37 �C.The CLSM observations were performed on a Fluoview FV10i(Olympus, Tokyo, Japan) equipped with a 60� water-immersionobjective lens (N/A 1.2) and a diode laser. The excitation wave-length was 405 nm (17.1 mW) for Hoechst 33342, 473 nm(11.9 mW) for LysoTracker Green, and 635 nm (9.5 mW) forAlexa647–siRNA. The colocalization ratio of Alexa647–siRNAwith endosomes/lysosomes (stained with LysoTracker Green)was quantied from the obtained images using the ImageJsoware as follows: colocalization ratio ¼ (pixels of Alexa647–siRNA)colocalized/(pixels of Alexa647–siRNA)total. The subscripts“colocalized” and “total” indicate the pixels of Alexa647–siRNAcolocalized with LysoTracker Green and the total pixels ofAlexa647–siRNA in the cells, respectively. The results areexpressed as the mean � standard deviation (S.D.) of 20 cells.
2.10 Statistical analysis
Statistical analysis was performed using a two-tail Student'st-test. A p-value of less than 0.05 was considered to indicatestatistical signicance. The values are expressed as the mean �standard deviation (S.D.).
3 Results and discussion3.1 Synthesis and pH-dependent degradation of DMAE–COO–PRX
The primary amine-terminated homobifunctional PEG bearing3-sulfanylpropionyl ester linkages (Mn¼ 4890) (PEG–COO–NH2)were utilized for the preparation of acid-degradable PRXs. Inthis study, an acid-degradable PRX with the number ofthreading a-CDs of 22.2 (coverage of CD: 43.1%) was synthe-sized (ESI Fig. S1–S5†). In our previous study, the physico-chemical stability of DMAE–PRX/siRNA polyplexes and theintracellular uptake level of siRNA are found to be improved byincreasing the number of DMAE groups modied on PRX.11
Therefore, three series of DMAE–COO–PRXs with variousnumbers of DMAE groups were synthesized (Table 1). In thisstudy, DMAE-modied reduction-degradable PRX bearingterminal disulde linkages (DMAE–SS–PRX) with the same PEGchain length and roughly comparable number of threading CDswas utilized as a control to allow the comparison with ourprevious system (Table 1).11
The time-course of hydrolysis of the DMAE–COO–PRX wasinvestigated by incubating the PRX at physiological pH (7.4) andendosomal/lysosomal pH conditions (5.5),31–33 and the
This journal is ª The Royal Society of Chemistry 2013

Table 1 Characterization of DMAE-modified PRXs used in this study
Samplecodea
Number ofthreadinga-CDsb
Number ofDMAE groupson PRXc
Number ofDMAE groupson a-CDc Mn
d
21DMAE–COO–PRX
22.2 (43.1%) 20.9 0.94 29 500
47DMAE–COO–PRX
22.2 (43.1%) 46.5 2.09 32 500
72DMAE–COO–PRX
22.2 (43.1%) 72.3 3.25 35 500
87DMAE–SS–PRX
29.2 (56.6%) 87.0 2.98 43 800
a Abbreviated as XDMAE–Y–PRX, where X and Y represent the numberof DMAE groups modied on PRX and terminal linkage of PRX,respectively. b Determined by 1H NMR in 0.1 M NaOD/D2O. Thevalues in parentheses represent the CD coverage percentage,assuming that one CD molecule includes two repeating units ofethylene glycol. c Determined by 1H NMR in D2O.
d Calculated basedon the chemical composition of DMAE–PRX determined by 1H NMR.
Paper Journal of Materials Chemistry B
Publ
ishe
d on
21
May
201
3. D
ownl
oade
d by
Uni
vers
ity o
f R
egin
a on
27/
10/2
014
01:2
4:19
. View Article Online
remaining percentage of the DMAE–COO–PRX was evaluated bysize exclusion chromatography (Fig. 2A). At pH 7.4, negligiblehydrolysis of the DMAE–COO–PRX was observed even aer 48 hof incubation. In contrast, time-dependent reduction of thepeak area of the DMAE–COO–PRX was conrmed at pH 5.5 (ESIFig. S6†), and approximately 50% of the DMAE–COO–PRX wasdegraded aer 24 h of incubation. This is assumed to be thereason for the hydrolysis of terminal esters of DMAE–COO–PRX.31–33
The complexation ability of DMAE–COO–PRX with siRNAwas evaluated by gel electrophoresis aer 6 h incubation at pH7.4 or 5.5 at 37 �C (Fig. 2B). At pH 7.4, the band of free siRNA wasgradually diminished with the increasing N/P ratio and thor-oughly disappeared at the N/P ratio of 1 (Fig. 2C). In sharpcontrast, the band of free siRNA was still observed at the N/Pratio of 10 at pH 5.5. Under the acidic pH, the complexationcapability between siRNA and PRX is thought to be superior dueto the protonation of the DMAE groups.10 However, the amountof uncomplexed siRNA at pH 5.5 was greater than pH 7.4 at
Fig. 2 (A) Hydrolysis kinetics of 72DMAE–COO–PRX at pH 7.4 (squares) and 5.5 (circelectrophoresis images of 72DMAE–COO–PRX/siRNA at various N/P ratios after 6 h inCOO–PRX at pH 7.4 (squares) and 5.5 (circles) at 37 �C, determined from the band
This journal is ª The Royal Society of Chemistry 2013
various N/P ratios (Fig. 2C). This result indicates that theDMAE–COO–PRX degrades under acidic pH conditions, leadingto the weakened multivalent electrostatic interaction betweenDMAE–COO–PRX and siRNA. Eventually, selective degradationof PRX under acidic conditions was achieved by DMAE–COO–PRX bearing terminal 3-sulfanylpropionyl esters.
3.2 pH-dependent membrane destabilizing activity ofDMAE–COO–PRX
Accompanied by the degradation of DMAE–COO–PRX underacidic pH conditions, an abundance of threaded a-CDs shouldbe released from the PRX. Although the cavity size of a-CD issmaller than b- and g-CDs, it can remove phospholipids fromthe membrane to induce hemolysis of erythrocytes.23 Therefore,the hemolytic activity of DMAE–COO–PRXs is thought to beincreased by the release of CDs in acidic pH. In this regard,hemolytic activities of DMAE–COO–PRXs with the variousnumbers of DMAE groups, 87DMAE–SS–PRX, and L-PEI (i.e.,commercially available transfection reagent, ExGen 500) as acontrol were evaluated aer the incubation with rat erythrocytesfor 6 h at pH 7.4 and 5.5 at 37 �C (Fig. 3). In this experiment, theconcentration of amino groups was adjusted to the same valueamong PRXs, because polyplex formation and gene silencingwere particularly conducted by adjusting the concentration ofamino groups (i.e., N/P ratio). The DMAE–COO–PRXs and the87DMAEA–SS–PRX showed negligible hemolysis of erythrocytesat pH 7.4 (less than 5%) regardless of the number of modiedDMAE groups and concentration, while the L-PEI showed asignicantly high level of hemolysis. This result suggests that thepolycation-derived membrane disruptive activity of L-PEI issignicant to induce cellular damage leading to cytotoxicity atphysiological pH, while the polycation-derived membranedisruptive activity of DMAE-modied PRXs was negligible.Additionally, the hemolytic activity of DMAE–COO–PRXs derivedfrom a-CD is also masked by the inclusion of the PEG chain intoits cavity. Therefore, it is considered that the DMAE–COO–PRXsare inherently non-invasive to the cellular membrane. In sharpcontrast, 21DMAE– and 46DMAE–COO–PRXs showed signicant
les) at 37 �C as measured by aqueous-phase size exclusion chromatography. (B) Gelcubation at pH 7.4 or 5.5 at 37 �C. (C) Amount of uncomplexed siRNA in 72DMAE–intensity of gel images.
J. Mater. Chem. B, 2013, 1, 3535–3544 | 3539
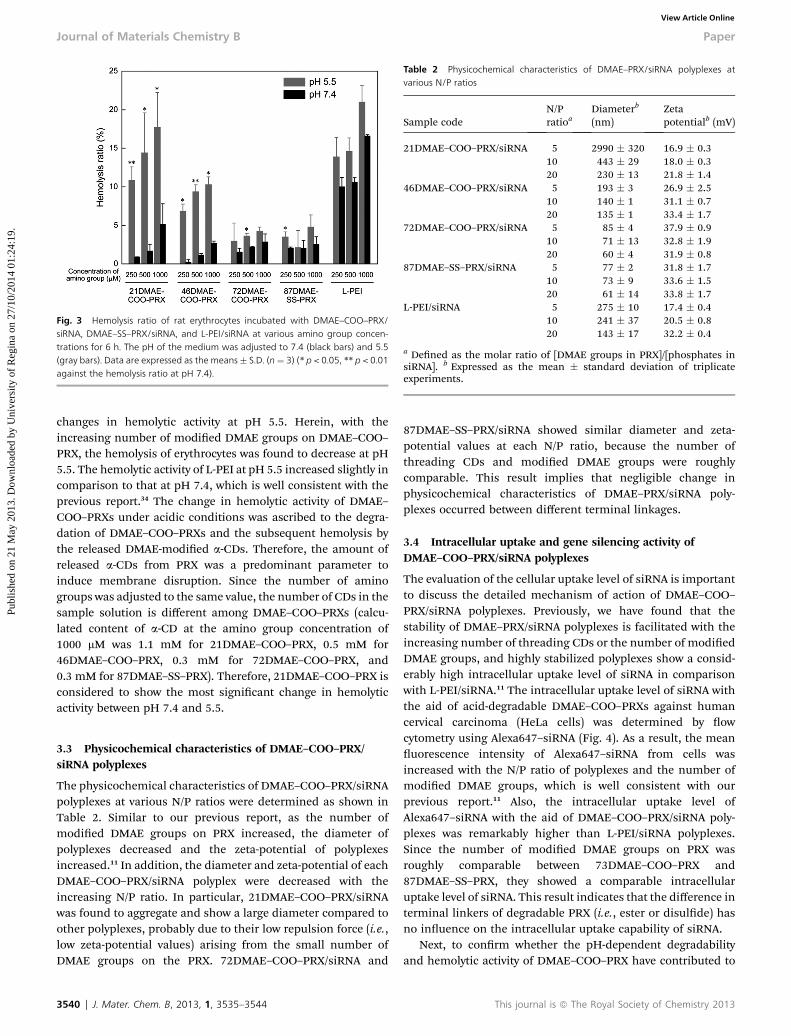
Fig. 3 Hemolysis ratio of rat erythrocytes incubated with DMAE–COO–PRX/siRNA, DMAE–SS–PRX/siRNA, and L-PEI/siRNA at various amino group concen-trations for 6 h. The pH of the medium was adjusted to 7.4 (black bars) and 5.5(gray bars). Data are expressed as the means� S.D. (n ¼ 3) (* p < 0.05, ** p < 0.01against the hemolysis ratio at pH 7.4).
Table 2 Physicochemical characteristics of DMAE–PRX/siRNA polyplexes atvarious N/P ratios
Sample codeN/Pratioa
Diameterb
(nm)Zetapotentialb (mV)
21DMAE–COO–PRX/siRNA 5 2990 � 320 16.9 � 0.310 443 � 29 18.0 � 0.320 230 � 13 21.8 � 1.4
46DMAE–COO–PRX/siRNA 5 193 � 3 26.9 � 2.510 140 � 1 31.1 � 0.720 135 � 1 33.4 � 1.7
72DMAE–COO–PRX/siRNA 5 85 � 4 37.9 � 0.910 71 � 13 32.8 � 1.920 60 � 4 31.9 � 0.8
87DMAE–SS–PRX/siRNA 5 77 � 2 31.8 � 1.710 73 � 9 33.6 � 1.520 61 � 14 33.8 � 1.7
L-PEI/siRNA 5 275 � 10 17.4 � 0.410 241 � 37 20.5 � 0.820 143 � 17 32.2 � 0.4
a Dened as the molar ratio of [DMAE groups in PRX]/[phosphates insiRNA]. b Expressed as the mean � standard deviation of triplicateexperiments.
Journal of Materials Chemistry B Paper
Publ
ishe
d on
21
May
201
3. D
ownl
oade
d by
Uni
vers
ity o
f R
egin
a on
27/
10/2
014
01:2
4:19
. View Article Online
changes in hemolytic activity at pH 5.5. Herein, with theincreasing number of modied DMAE groups on DMAE–COO–PRX, the hemolysis of erythrocytes was found to decrease at pH5.5. The hemolytic activity of L-PEI at pH 5.5 increased slightly incomparison to that at pH 7.4, which is well consistent with theprevious report.34 The change in hemolytic activity of DMAE–COO–PRXs under acidic conditions was ascribed to the degra-dation of DMAE–COO–PRXs and the subsequent hemolysis bythe released DMAE-modied a-CDs. Therefore, the amount ofreleased a-CDs from PRX was a predominant parameter toinduce membrane disruption. Since the number of aminogroups was adjusted to the same value, the number of CDs in thesample solution is different among DMAE–COO–PRXs (calcu-lated content of a-CD at the amino group concentration of1000 mM was 1.1 mM for 21DMAE–COO–PRX, 0.5 mM for46DMAE–COO–PRX, 0.3 mM for 72DMAE–COO–PRX, and0.3 mM for 87DMAE–SS–PRX). Therefore, 21DMAE–COO–PRX isconsidered to show the most signicant change in hemolyticactivity between pH 7.4 and 5.5.
3.3 Physicochemical characteristics of DMAE–COO–PRX/siRNA polyplexes
The physicochemical characteristics of DMAE–COO–PRX/siRNApolyplexes at various N/P ratios were determined as shown inTable 2. Similar to our previous report, as the number ofmodied DMAE groups on PRX increased, the diameter ofpolyplexes decreased and the zeta-potential of polyplexesincreased.11 In addition, the diameter and zeta-potential of eachDMAE–COO–PRX/siRNA polyplex were decreased with theincreasing N/P ratio. In particular, 21DMAE–COO–PRX/siRNAwas found to aggregate and show a large diameter compared toother polyplexes, probably due to their low repulsion force (i.e.,low zeta-potential values) arising from the small number ofDMAE groups on the PRX. 72DMAE–COO–PRX/siRNA and
3540 | J. Mater. Chem. B, 2013, 1, 3535–3544
87DMAE–SS–PRX/siRNA showed similar diameter and zeta-potential values at each N/P ratio, because the number ofthreading CDs and modied DMAE groups were roughlycomparable. This result implies that negligible change inphysicochemical characteristics of DMAE–PRX/siRNA poly-plexes occurred between different terminal linkages.
3.4 Intracellular uptake and gene silencing activity ofDMAE–COO–PRX/siRNA polyplexes
The evaluation of the cellular uptake level of siRNA is importantto discuss the detailed mechanism of action of DMAE–COO–PRX/siRNA polyplexes. Previously, we have found that thestability of DMAE–PRX/siRNA polyplexes is facilitated with theincreasing number of threading CDs or the number of modiedDMAE groups, and highly stabilized polyplexes show a consid-erably high intracellular uptake level of siRNA in comparisonwith L-PEI/siRNA.11 The intracellular uptake level of siRNA withthe aid of acid-degradable DMAE–COO–PRXs against humancervical carcinoma (HeLa cells) was determined by owcytometry using Alexa647–siRNA (Fig. 4). As a result, the meanuorescence intensity of Alexa647–siRNA from cells wasincreased with the N/P ratio of polyplexes and the number ofmodied DMAE groups, which is well consistent with ourprevious report.11 Also, the intracellular uptake level ofAlexa647–siRNA with the aid of DMAE–COO–PRX/siRNA poly-plexes was remarkably higher than L-PEI/siRNA polyplexes.Since the number of modied DMAE groups on PRX wasroughly comparable between 73DMAE–COO–PRX and87DMAE–SS–PRX, they showed a comparable intracellularuptake level of siRNA. This result indicates that the difference interminal linkers of degradable PRX (i.e., ester or disulde) hasno inuence on the intracellular uptake capability of siRNA.
Next, to conrm whether the pH-dependent degradabilityand hemolytic activity of DMAE–COO–PRX have contributed to
This journal is ª The Royal Society of Chemistry 2013

Fig. 4 Mean fluorescence intensities of HeLa cells incubated with DMAE–COO–PRX/siRNA, DMAE–SS–PRX/siRNA, and L-PEI/siRNA at various N/P ratios for 48 h,as determined by flow cytometry. The concentration of Alexa647–siRNA in themedium was adjusted to 100 nM. Data are expressed as the means � S.D. (n ¼ 3)(* p < 0.05, ** p < 0.01, *** p < 0.005, **** p < 0.001, NS indicates not significant).
Paper Journal of Materials Chemistry B
Publ
ishe
d on
21
May
201
3. D
ownl
oade
d by
Uni
vers
ity o
f R
egin
a on
27/
10/2
014
01:2
4:19
. View Article Online
the gene silencing activity, the dual luciferase reporter assay ofDMAE–COO–PRX/siRNA polyplexes was conducted against HeLacells (Fig. 5). The rey luciferase gene expression level wasmoderately reduced for 46DMAE– and 72DMAE–COO–PRX/siRNA in comparison with 87DMAE–SS–PRX/siRNA and L-PEI/siRNA polyplexes at each N/P ratio. In particular, 46DMAE–COO–PRX/siRNA showed the lowest gene expression level of all thepolyplexes, although the cellular uptake level of 46DMAE–COO–PRX was lower than 72DMAE–COO–PRX and 87DMAE–SS–PRX.
Fig. 5 Gene silencing efficiency of firefly luciferase expressed in HeLa cellsincubated with DMAE–COO–PRX/siRNA, DMAE–SS–PRX/siRNA, and L-PEI/siRNAat various N/P ratios for 48 h. The black and gray bars represent the siRNAsequence against firefly luciferase and a scrambled sequence, respectively. Theconcentration of siRNA in the medium was adjusted to 100 nM. Data areexpressed as the means � S.D. (n ¼ 3) (* p < 0.05, ** p < 0.01 against L-PEI withthe corresponding N/P ratios).
This journal is ª The Royal Society of Chemistry 2013
In the case of the DMAE–SS–PRX system, the intracellular uptakelevel of siRNA is a predominant factor to determine the genesilencing efficiency, and the gene silencing activity of DMAE–SS–PRX/siRNA increased with the number of modied DMAEgroups.11 Considering these facts, the specic membranedestabilizing activity of DMAE–COO–PRXs under acidic condi-tions might contribute to enhancing the gene silencing activity,because 46DMAE–COO–PRX showed pronounced membranedestabilizing activity at acidic pH (Fig. 3). However, the21DMAE–COO–PRX/siRNA polyplex showed no gene silencingactivity, although 21DMAE–COO–PRX showed the most signi-cant change in hemolytic activity at pH 5.5 (Fig. 3). This might bedue to the aggregation of the 21DMAE–COO–PRX/siRNA poly-plex arisen from their low surface charge (Table 2). Indeed, theaggregation of the 21DMAE–COO–PRX/siRNA polyplex in themediumwas conrmed and the aggregates were found to adsorbon the surface of cells (ESI Fig. S7†). Therefore, the 21DMAE–COO–PRX/siRNA polyplex showed lower intracellular uptake ofsiRNA and failed to induce gene silencing. Taking these resultsinto account, both the pH-dependent membrane destabilizingactivity and the capability of efficient cellular internalization ofsiRNA are required for PRX to ensure the feasibility of effectivegene silencing.
When the scrambled siRNA was used, all the DMAE–COO–PRX/siRNA polyplexes resulted in negligible gene silencing ateach N/P ratio (Fig. 5), indicating that the gene silencing ofDMAE–COO–PRX/siRNA polyplexes was due to the sequence-specic effect. In the case of the L-PEI/siRNA, the rey lucif-erase gene expression level decreased with the increasing N/Pratio even with the scrambled siRNA sequence. Additionally, thecellular viability was decreased by L-PEI/siRNA polyplexes,whereas negligible cytotoxicity was observed for DMAE–COO–PRX/siRNA polyplexes (Fig. 6). As shown in Fig. 3, L-PEI showedsignicant hemolysis at pH 7.4. This membrane disruptiveactivity of L-PEI is thought to induce cytotoxicity and the non-specic gene silencing. In contrast, the DMAE–COO–PRXs werethoroughly non-invasive to cells. In our previous study, thecytotoxicity of intracellularly degradable DMAE–PRX is revealedto be lower than that of non-degradable DMAE–PRX.8 Accord-ingly, it is indicated that the intracellular dissociation ofDMAE–COO–PRXs into its constituent low molecular weightDMAE-modied a-CDs contributes to reducing the cytotoxicityand non-specic silencing effect.
To further ascertain the availability of the 46DMAE–COO–PRX/siRNA polyplex, siRNA concentration-dependency of genesilencing activity was evaluated (Fig. 7). When the concentrationof siRNA was reduced, the intracellular uptake level of siRNAshould be decreased. However, the 46DMAE–COO–PRX/siRNApolyplex (N/P 10) showed signicantly high gene silencing activityin comparison with the 87DMAE–COO–PRX/siRNA (N/P 10) andthe L-PEI/siRNA (N/P 10) polyplexes even at low siRNA concen-tration (10 nM). The 50% inhibitory concentration (IC50) of siRNAfor the 46DMAE–COO–PRX/siRNA was determined to be 14.5 nM,which was considerably lower than the 87DMAE–SS–PRX/siRNA(52.0 nM) and the L-PEI/siRNA (57.9 nM). Therefore, the acid-degradable capability of DMAE–COO–PRXs is effective to enhancethe gene silencing activity especially at low siRNA concentrations.
J. Mater. Chem. B, 2013, 1, 3535–3544 | 3541

Fig. 6 Viability of HeLa cells incubated with 21DMAE–COO–PRX/siRNA,46DMAE–COO–PRX/siRNA, 72DMAE–COO–PRX/siRNA, 87DMAE–SS–PRX/siRNA,and L-PEI/siRNA at various N/P ratios for 48 h. The concentration of siRNA wasadjusted to 100 nM. Data are expressed as the means � S.D. (n ¼ 6) (* p < 0.05,**** p < 0.001).
Fig. 7 Gene silencing efficiency of firefly luciferase expressed in HeLa cellsincubated with 46DMAE–COO–PRX/siRNA (N/P 10, diamonds), 87DMAE–SS–PRX/siRNA (N/P 10, triangles), and L-PEI/siRNA (N/P 10, squares) at varioussiRNA concentrations for 48 h. Data are expressed as the means � S.D. (n ¼ 3)(* p < 0.05, ** p < 0.01, *** p < 0.005 against L-PEI with the corresponding siRNAconcentrations).
Fig. 8 Gene silencing efficiency of firefly luciferase expressed in HeLa cellsincubated with 46DMAE–COO–PRX/siRNA (N/P 10, diamonds) and 87DMAE–SS–PRX/siRNA (N/P 10, triangles) in the presence of ammonium chloride with variousconcentrations for 48 h. The concentration of siRNA in the medium was adjustedto 100 nM. Data are expressed as the means� S.D. (n¼ 3) (* p < 0.05, ** p < 0.01,*** p < 0.005 against the samples without ammonium chloride treatment).
Journal of Materials Chemistry B Paper
Publ
ishe
d on
21
May
201
3. D
ownl
oade
d by
Uni
vers
ity o
f R
egin
a on
27/
10/2
014
01:2
4:19
. View Article Online
To validate the contribution of the acid-degradable characterof 46DMAE–COO–PRX in the gene silencing activity, the genesilencing activity was determined in the presence of ammoniumchloride (NH4Cl) (Fig. 8). The treatment with NH4Cl mightinhibit the degradation of DMAE–COO–PRXs and the release ofCDs in endosomes/lysosomes, because NH4Cl is known to elicitthe neutralization of intraendosomal/intralysosomal pH.15,35
With the increasing concentration of NH4Cl in the medium,the rey luciferase gene expression level increased forthe 46DMAE–COO–PRX/siRNA in a concentration-dependentmanner, whereas no signicant change was observed for the87DMAE–SS–PRX/siRNA. This result strongly indicates that the
3542 | J. Mater. Chem. B, 2013, 1, 3535–3544
acid-degradable capability of the 46DMAE–COO–PRX plays acrucial role in enhancing the gene silencing activity of siRNA.
3.5 Endosomal escape efficiency of DMAE–COO–PRX/siRNApolyplexes
We hypothesized that the mechanism of action of 46DMAE–COO–PRX/siRNA is as follows: the 46DMAE–COO–PRX/siRNAwas internalized in cells through the endocytosis pathway andmost part of them was accumulated in endosomes and lyso-somes. Wherein, the ester terminal linkages of DMAE–COO–PRXs were hydrolyzed, resulting in the dissociation of PRXs andpolyplexes. Subsequently, the released CDs are thought todisturb the integrity of endosomal membranes, resulting in thepromotion of endosomal escape of siRNA. To elucidate thishypothesis, intracellular localization of Alexa647–siRNA wasobserved by confocal laser scanning microscopy (CLSM)(Fig. 9A–C). In this experiment, acidic endosomes and lyso-somes were stained with LysoTracker Green for visualizing thelocalization of siRNA. Since L-PEI is known to show endosomalescape through the proton sponge effect,16,17 the red spots ofAlexa647–siRNA were observed by separation from endosomesand lysosomes (green) (Fig. 9C). However, the number of redspots was obviously lower than PRXs, again conrming that theintracellular uptake level of L-PEI/siRNA was lower than that ofPRX/siRNA polyplexes. In the case of the 87DMAE–SS–PRX/siRNA polyplex, most of the spots of Alexa647–siRNA wereobserved as yellow, indicating that the siRNA was localized inendosomes and lysosomes (Fig. 9B). In contrast, some portionof Alexa647–siRNA was observed by separation from endosomesand lysosomes for the 46DMAE–COO–PRX/siRNA (Fig. 9A). Toenable a quantitative comparison of these images, the colocal-ization ratio of Alexa647–siRNA to endosomes/lysosomes wasanalyzed from images (Fig. 9D).18,34,36 Consistent with the CLSMimages, the colocalization ratio of the 87DMAE–SS–PRX/siRNA
This journal is ª The Royal Society of Chemistry 2013

Fig. 9 (A–C) CLSM images of HeLa cells incubated with 46DMAE–COO–PRX/siRNA (N/P 10) (A), 87DMAE–SS–PRX/siRNA (N/P 10) (B), and L-PEI/siRNA (N/P 10) (C) for6 h, followed by 42 h incubation without samples (scale bars: 20 mm). The red spots represent Alexa647–siRNA. The cell nuclei and late endosomes/lysosomes werestained with Hoechst 33342 (blue) and LysoTracker Green (green), respectively. The concentration of Alexa647–siRNA in the medium was adjusted to 100 nM. (D) Thecolocalization ratio of Alexa647–siRNA to late endosomes/lysosomes as determined from CLSM images. The bars represent the means � S.D. of 20 cells.
Paper Journal of Materials Chemistry B
Publ
ishe
d on
21
May
201
3. D
ownl
oade
d by
Uni
vers
ity o
f R
egin
a on
27/
10/2
014
01:2
4:19
. View Article Online
was high (0.74 � 0.06), indicating that most of the siRNA werelocalized in endosomes/lysosomes. In contrast, the colocaliza-tion ratio of 46DMAE–COO–PRX/siRNA (0.56 � 0.13) wassignicantly lower than that of 87DMAE–SS–PRX/siRNA,strongly indicating that the endosomal escape efficiency ofDMAE–COO–PRX/siRNA was improved by the use of acid-degradable 46DMAE–COO–PRX. Although the colocalizationratio of the 46DMAE–COO–PRX/siRNA was still higher than thatof L-PEI/siRNA (0.47 � 0.12), the gene silencing activity of the46DMAE–COO–PRX/siRNA was denitely higher than that ofL-PEI (Fig. 5 and 7). This is presumably due to the difference inthe intracellular uptake level of siRNA (Fig. 4). Therefore, we canconclude that the remarkable gene silencing efficiency of the46DMAE–COO–PRX/siRNA was attributed to the excellence ofthe intracellular uptake of siRNA as well as the promotion ofendosomal escape by membrane destabilization of CDsreleased under acidic conditions.
4 Conclusion
Acid-degradable DMAE–COO–PRXs were developed forimproving the endosomal escape efficiency of siRNA. TheDMAE–COO–PRXs were sufficiently stable to form polyplexeswith siRNA under physiological pH conditions. In contrast, theDMAE–COO–PRXs were degraded under acidic conditions bythe hydrolysis of terminal ester linkages and threaded a-CDs
This journal is ª The Royal Society of Chemistry 2013
were released. Therefore, DMAE–COO–PRXs showed pH-dependent hemolytic activity by the membrane released desta-bilizing activity of released a-CDs. As a result, acid-degradableDMAE–COO–PRXs showed higher gene silencing activity thanreduction-degradable DMAE–SS–PRX and L-PEI, withoutmarked cytotoxicity. The confocal uorescence microscopicimages revealed that the colocalization ratio of siRNAs toendosomes and lysosomes were decreased by the use of aciddegradable DMAE–COO–PRXs, indicating that the endosomalescape of siRNA was promoted. Accordingly, the DMAE–COO–PRXs are thought to be an effective candidate as a carrier forcytoplasmic siRNA delivery. Besides the siRNA delivery, theDMAE–COO–PRXs could be applied to the cytoplasmic deliveryof diverse biomacromolecules such as proteins. We believe thatthe DMAE–COO–PRXs would be a promising candidate for boththe therapeutic and analytical applications and contributed to abroad range of biotechnological elds.
Acknowledgements
This work was nancially supported by the Grant-in-Aid forScientic Research (no. 23107004) on Innovative Areas “Nano-medicine Molecular Science” (no. 2306) from the Ministry ofEducation, Culture, Sports, Science, and Technology (MEXT) ofJapan.
J. Mater. Chem. B, 2013, 1, 3535–3544 | 3543

Journal of Materials Chemistry B Paper
Publ
ishe
d on
21
May
201
3. D
ownl
oade
d by
Uni
vers
ity o
f R
egin
a on
27/
10/2
014
01:2
4:19
. View Article Online
References
1 Y. Dorsett and T. Tuschl, Nat. Rev. Drug Discovery, 2004, 3,318–329.
2 E. Iorns, C. J. Lord, N. Turner and A. Ashworth, Nat. Rev. DrugDiscovery, 2007, 6, 556–568.
3 M. Stevenson, N. Engl. J. Med., 2004, 351, 1772–1777.4 B. L. Davidson and P. B. McCray Jr, Nat. Rev. Genet., 2011, 12,329–340.
5 K. A. Whitehead, R. Langer and D. G. Anderson, Nat. Rev.Drug Discovery, 2009, 8, 129–138.
6 J. W. Yoo, D. J. Irvine, D. E. Discher and S. Mitragotri, Nat.Rev. Drug Discovery, 2011, 10, 521–535.
7 A. Tamura and Y. Nagasaki, Nanomedicine, 2010, 5, 1089–1102.8 T. Ooya, H. S. Choi, A. Yamashita, N. Yui, Y. Sugaya, A. Kano,A. Maruyama, H. Akita, R. Ito, K. Kogure and H. Harashima,J. Am. Chem. Soc., 2006, 128, 3852–3853.
9 A. Yamashita, N. Yui, T. Ooya, A. Kano, A. Maruyama,H. Akita, K. Kogure and H. Harashima, Nat. Protoc., 2006,1, 2861–2869.
10 A. Yamashita, D. Kanda, R. Katoono, N. Yui, T. Ooya,A. Maruyama, H. Akita, K. Kogure and H. Harashima,J. Controlled Release, 2008, 131, 137–144.
11 A. Tamura and N. Yui, Biomaterials, 2013, 34, 2480–2491.12 C. D. Austin, X. Wen, L. Gazzard, C. Nelson, R. H. Scheller
and S. J. Scales, Proc. Natl. Acad. Sci. U. S. A., 2005, 102,17987–17992.
13 I. Mellman, R. Fuchs and A. Helenius, Annu. Rev. Biochem.,1986, 55, 663–700.
14 S. Mukherjee, R. N. Ghosh and F. R. Maxeld, Physiol. Rev.,1997, 77, 759–803.
15 S. Ohkuma and B. Poole, Proc. Natl. Acad. Sci. U. S. A., 1978,75, 3327–3331.
16 N. D. Sonawane, F. C. Szoka Jr and A. S. Verkman, J. Biol.Chem., 2003, 278, 44826–44831.
17 O. Boussif, F. Lezoualc'h, M. A. Zanta, M. D. Mergny,D. Scherman, B. Demeneix and J. P. Behr, Proc. Natl. Acad.Sci. U. S. A., 1995, 92, 7297–7301.
18 K. Miyata, M. Oba, M. Nakanishi, S. Fukushima,Y. Yamasaki, H. Koyama, N. Nishiyama and K. Kataoka,J. Am. Chem. Soc., 2008, 130, 16287–16294.
3544 | J. Mater. Chem. B, 2013, 1, 3535–3544
19 R. A. Jones, C. Y. Cheung, F. E. Black, J. K. Zia, P. S. Stayton,A. S. Hoffman and M. R. Wilson, Biochem. J., 2003, 372,65–75.
20 N. Sakaguchi, C. Kojima, A. Harada and K. Kono,Bioconjugate Chem., 2008, 19, 1040–1048.
21 Y. Ohtani, T. Irie, K. Uekama, K. Fukunaga and J. Pitha, Eur.J. Biochem., 1989, 186, 17–22.
22 T. Irie and K. Uekama, J. Pharm. Sci., 1997, 86, 147–162.
23 F. Fauvelle, J. C. Debouzy, S. Crouzy, M. Goschl andY. Chapron, J. Pharm. Sci., 1997, 86, 935–943.
24 E. P. Kilsdonk, P. G. Yancey, G. W. Stoudt, F. W. Bangerter,W. J. Johnson, M. C. Phillips and G. H. Rothblat, J. Biol.Chem., 1995, 270, 17250–17256.
25 H. Arima, F. Kihara, F. Hirayama and K. Uekama.,Bioconjugate Chem., 2001, 12, 476–484.
26 T. Tsutsumi, F. Hirayama, K. Uekama and H. Arima,J. Controlled Release, 2007, 119, 349–359.
27 H. S. Choi, A. Yamashita, T. Ooya, N. Yui, H. Akita,K. Kogure, R. Ito and H. Harashima, ChemBioChem, 2005,6, 1986–1990.
28 M. E. Davis, J. E. Zuckerman, C. H. J. Choi, D. Seligson,A. Tolcher, C. A. Alabi, Y. Yen, J. D. Heidel and A. Ribas,Nature, 2010, 464, 1067–1070.
29 K. Chaturvedi, K. Ganguly, A. R. Kulkarni, V. H. Kulkarni,M. N. Nadagouda, W. E. Rudzinski and T. M. Aminabhavi,Expert Opin. Drug Delivery, 2011, 8, 1455–1468.
30 C. O. Mellet, J. M. Garcıa Fernandez and J. M. Benito, Chem.Soc. Rev., 2011, 40, 1586–1608.
31 R. G. Schoenmakers, P. van de Wetering, D. L. Elbert andJ. A. Hubbell, J. Controlled Release, 2004, 95, 291–300.
32 M. Oishi, S. Sasaki, Y. Nagasaki and K. Kataoka,Biomacromolecules, 2003, 4, 1426–1432.
33 K. Dan, R. Pan and S. Ghosh, Langmuir, 2011, 27, 612–617.
34 H. Uchida, K. Miyata, M. Oba, T. Ishii, T. Suma, K. Itaka,N. Nishiyama and K. Kataoka, J. Am. Chem. Soc., 2011, 133,15524–15532.
35 B. Poole and S. Ohkuma, J. Cell Biol., 1981, 90, 665–669.36 A. Tamura, M. Oishi and Y. Nagasaki, J. Controlled Release,
2010, 146, 378–387.
This journal is ª The Royal Society of Chemistry 2013