a survey of fungi which kill microscopic animals in the dung of the amami rabbit
TRANSCRIPT

Volume 11, Part 3, August 1997
A SURVEY OF FUNGI WHICH KILL MICROSCOPICANIMALS IN THE DUNG OF THE AMAMI RABBIT
SALLY L. GLOCKLING & FUMIO YAMADA
Forestry and Forest Products Research Institute, P.O. Box 16, Tsukuba Norin kenkyuDanchi-Nai, Ibaraki 305, Japan. Facsimile: 81-298-73-1543
Dung of the early-evolved Amami rabbit ofJapan was surveyed for the presence of fungiwhich attack microscopic organisms. Severalspecies of predacious nematophagous fungi andspecies endoparasitic in nematodes and rotiferswere found. In addition, zygomycetous speciesattacking amoebae and tardigrades, and one fla-gellate zoosporic species holocarpic in varioushost organisms, were found.
Keywords: nematode, rotifer, amoeba, tardi-grade, nematophagous, endoparasitic, flagellate,zoospore, holocarpic.
The Amami Islands of Amami Ohshima andTokunoshima which form part of the RyukyuArchipelago are situated 127 - 1290 E and 25 - 270
N in southern Japan; together the islands areless than 1000 km'.
The Amami rabbit (Pentalagus furnessiStone), which is found only on the AmamiIslands, is an indigenous species which isbelieved to have evolved in the Pliocene period(Dawson, 1981). It differs from more recentlyevolved lagomorphs in having small eyes andears, short legs and long claws (Fig 1). The totalpopulation of the Amami rabbit on AmamiOhshima and Tokunoshima is estimated ataround 4000 (Yamada, Sugimura & Koyanagi,1995). The rabbits inhabit the forests and eatleaves of the Japanese pampas grass, Miscanthussinensis, acorns of the pasania tree, Castanopsisseibodia, bark and bamboo shoots (Sugimura,1990). They are nocturnal as their black coloura-tion indicates. The Amami rabbit was designat-ed a symbol of national heritage by thegovernment in 1921 and this status was raised in1963. Despite banning hunting of the rabbit, itsnumbers are declining due to deforestation andit is classified as endangered by the IUCN andUSDI.
The remote geographical location and smallsize of the Amami Islands combined with the
early evolution and history of the Amami rabbitmade an ecological survey of predacious fungi inthe rabbit dung an interesting area for research.
Whilst many fungi subsisting on microscopicanimals appear to be ubiquitous, others have, asyet, a very restricted reported geographical dis-tribution and many are known to exhibit a highdegree of host-specificity. Predacious and para-sitic fungi attack microscopic animals such asnematodes, rotifers, tardigrades and amoebae.Such fungal species are found in all the majormycological groups and they display a wide vari-ety of trapping and infection methods. Many arehost-specific and host-dependent whilst othersare able to exploit several host organisms andsome have a saprophytic capability. The nema-tode and rotifer species which are attacked aremainly bacteriophagous and are found in habi-tats where decomposition of organic matter istaking place. In such habitats rich in bacteria,the nematode and rotifer populations increase.At such times, fungi which subsist on these ani-mals, either host-dependently or as a supple-ment, are able to grow and propagate. Dung isusually a good source of such fungal species as itretains moisture well, is full of bacteria and har-bours a diversity of microscopicanimals.
Methods
A total of 16 collections of dung of the Amamirabbit were made from Amami Ohshima onthree occasions in autumn, winter and summer.Collections were taken from Sumiyo Village,Yamato Village, Kawauchi and Mt Naon. Dungsamples were mixed with distilled water and sev-eral drops of the liquid were placed on to Petriplates containing corn meal agar (CMA). Moredistilled water was added to the plates after afew days to assist breeding of nematodes androtifers. Predacious hyphomycetous fungi wereroutinely isolated and obtained in pure cultureusing Raper's rings. Endoparasitic species wereisolated and cultured using methods previously
•

Volume 11, Part 3, August 1997
Fig 1 Amami rabbit (Scale bar, 40 mm). Fig 2 Passive ring and stalked knob of Dactylaria candida (Scale bar, 20 prn).Fig 3 Infection bulb inside the nematode body (Scale bar, 6 um). Fig 4 Knobs of D. candida producing assimilativehyphae in the nematode body (Scale bar, 6 llm). Fig 5 Nematode with passive rings of D. candida (Scale bar, 40 pm),Fig 6 Plectus nematode infected with Verticillium balanoides (Scale bar, 50 prn), Fig 7 Conidia of V balanoides.Conidium sticking to nematode cuticle (arrow) (Scale bar, 6 um). Fig 8 Sporulation of Harposporium cycloides fromnematode (Scale bar, 20 urn). Fig 9 Infection conidium and arthroconidium in H cycloides (Scale bar, 6 urn),
Volume 11, Part 3, August 1997
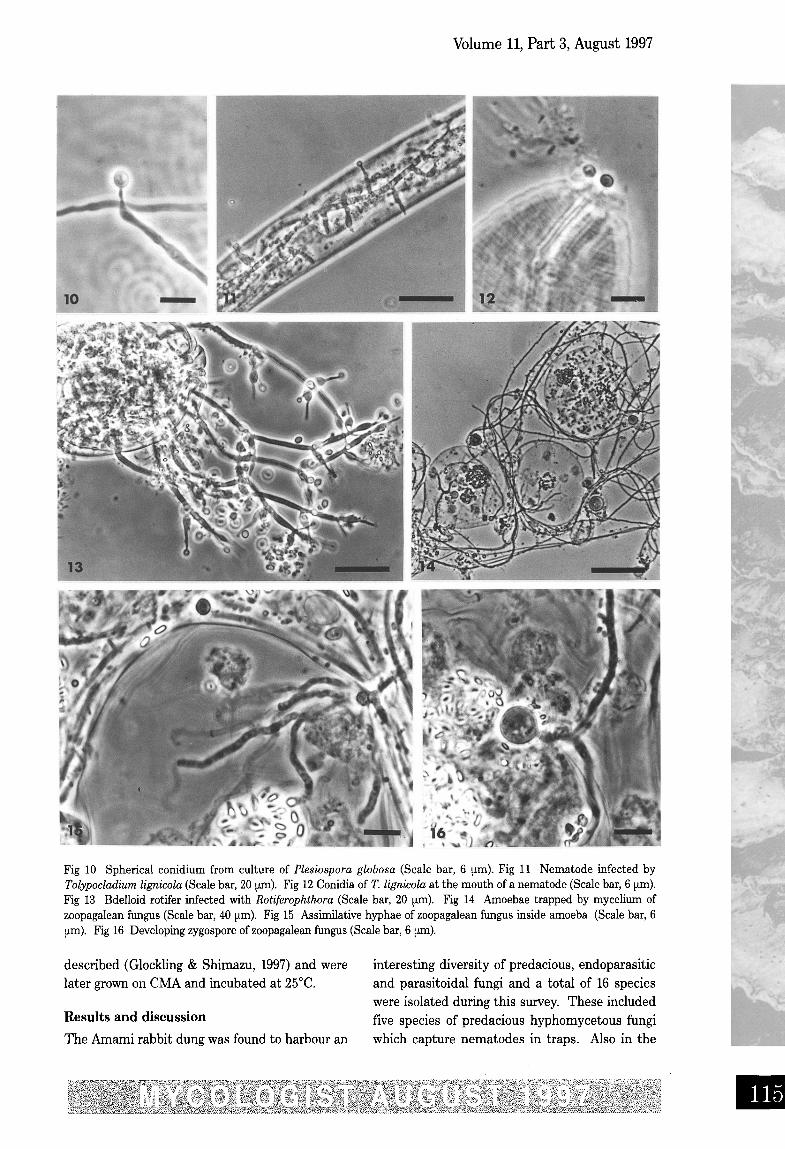
Fig 10 Spherical conidium from culture of Plesiospora globosa (Scale bar, 6 urn). Fig 11 Nematode infected byTolypocladium lignicola (Scale bar, 20 lLm). Fig 12 Conidia of T. lignicola at the mouth of a nematode (Scale bar, 6 um).Fig 13 Bdelloid rotifer infected with Rotiferophthora (Scale bar, 20 um). Fig 14 Amoebae trapped by mycelium ofzoopagalean fungus (Scale bar, 40 prn), Fig 15 Assimilative hyphae of zoopagalean fungus inside amoeba (Scale bar, 6urn), Fig 16 Developing zygospore of zoopagalean fungus (Scale bar, 6 pm),
described (Glockling & Shimazu, 1997) and werelater grown on CMA and incubated at 25°C.
Results and discussion
The Amami rabbit dung was found to harbour an
interesting diversity of predacious, endoparasiticand parasitoidal fungi and a total of 16 specieswere isolated during this survey. These includedfive species of predacious hyphomycetous fungiwhich capture nematodes in traps. Also in the
•

•
Volume 11, Part 3, August 1997
Fungi Imperfecti, six species endoparasitic innematodes and one species endoparasitic inrotifers were found. Within the Zygomycotina,one member of the Zoopagales capturing amoe-bae was found, and an entomophthoraceous fun-gus which was parasitic in tardigrades. Onespecies of zoosporic oomycetous fungi holocarpicin nematodes was found in the rabbit dung. Onesurprising result from the survey was the pres-ence of Dactylella leptospora Drech. (Drechsler,1937) and Dactylaria candida (Nees .) Sacco(Saccardo, 1886) which capture nematodes witha combination of both passive rings (rings whichdo not constrict), and adhesive stalked knobs(Fig 2). There are only three species knownwhich produce this combination of trappingapparatus. After capture of a nematode by anadhesive knob or a passive ring, the fungus formsa large infection bulb inside the body whicheffectively severs the internal organs and killsthe nematode (Fig 3). The adhesive knobs areeasily detached from the hyphae after contactwith a nematode and they penetrate into thenematode body and produce assimilative hyphae(Fig 4). The passive rings are also commonlydetached as they are held on narrow hyphaewhich are easily broken if a strong nematode iscaptured. The nematode swims away with therings around its body (Fig 5) and is later infected.The ease with which the captured nematodesbreak the hyphal supports of the trappingdevices ensures dispersal of the fungus awayfrom the mycelium. Another fungus producingonly stalked knobs, Dactylella ellipsospora Grove(Grove, 1885) was also isolated from the rabbitdung. Modern techniques (Borrebaek et al.,1984) have shown that nematode-trapping struc-tures produce carbohydrate-binding proteinswhich bind to N-acetyl-D-galactosamine on thenematode surface. In this way, the stalked knobsare host-specific for nematodes which secretesubstances which bind them to the knobs, andare unable to capture nematodes which do notsecrete the chemicals needed for adhesion.When Drechsler described Dactylella leptospora(Drechsler, 1937) he reported that the stalkedknobs in his cultures were ineffective and thatonly the passive rings were able to capture nema-todes. The existence of the chemical bondbetween the nematode hosts and the adhesiveknobs was not known at that time and it is nowevident that the nematodes in Drechsler 's cul-
tures were not compatible with D. leptospora.Friman (1993) developed a method for isolatingthe adhesive knobs of Dactylaria candida so thatfuture studies of the trap-specific microbodieswill be possible. Conidia of both Dactylella lep-tospora and Dactylaria candida showed an abili-ty to germinate to produce adhesive knobsdirectly. Species which display this ability forearly trap development were thought to exhibitless saprophytic ability and more host-depen-dence but, as previously stated (Cooke, 1963), nostudy to assess the extent to which these nema-tode-trapping species exist in a saprophyticcapacity in the wild has ever been made, and allare able to grow with varying growth rates onartificial media.
The two Arthrobotrys species isolated duringthis survey, A. cladodes Drech. (Drechsler, 1937)and A . sup erba Corda (Corda , 1839) are fairlywell-known species which are commonly report-ed from a variety of habitats. Neither of thesespecies produced chlamydospores in culture andboth produced conidia with a single septum justbelowor near the centre. The isolate of A. clado-des Drech. agreed well with that of the originaldescription. The fungus identified as A. superbaCorda had both proliferating and branching coni-diophores but differed slightly from Drechsler'sisolate in having slightly broader conidia .Conidia ranged from a short, squat form to acylindrical, elongate form. Although both typeswere produced on one conidiophore, they wereborne in separate whorls. A closer identificationfor this isolate could not be found. Both thesefungi and Dactylaria thaumasia Drech .(Drechsler, 1937) construct three-dimensionalnetworks of adhesive hyphal loops in whichnematodes become entangled.
The endoparasitic hyphomycetes foundincluded three species of Harposporium, onespecies of Rotiferophthora, and one species ofVerticillium. Also, a rare fungus belonging to amonotypic genus, Plesiospora, was found infect-ing just two nematodes. In addition, aTolypocladium species originally described frombdelloid rotifers and producing small disc-shapedconidia, was isolated from a single nematodehost. Harposporium and Rotiferophthora speciesusually infect the host with infection conidiawhich are ingested and which germinate insidethe host . All the nematodes infected byVerticillium balanoides (Drech.) Dowsett et al.
Volume 11, Part 3, August 1997
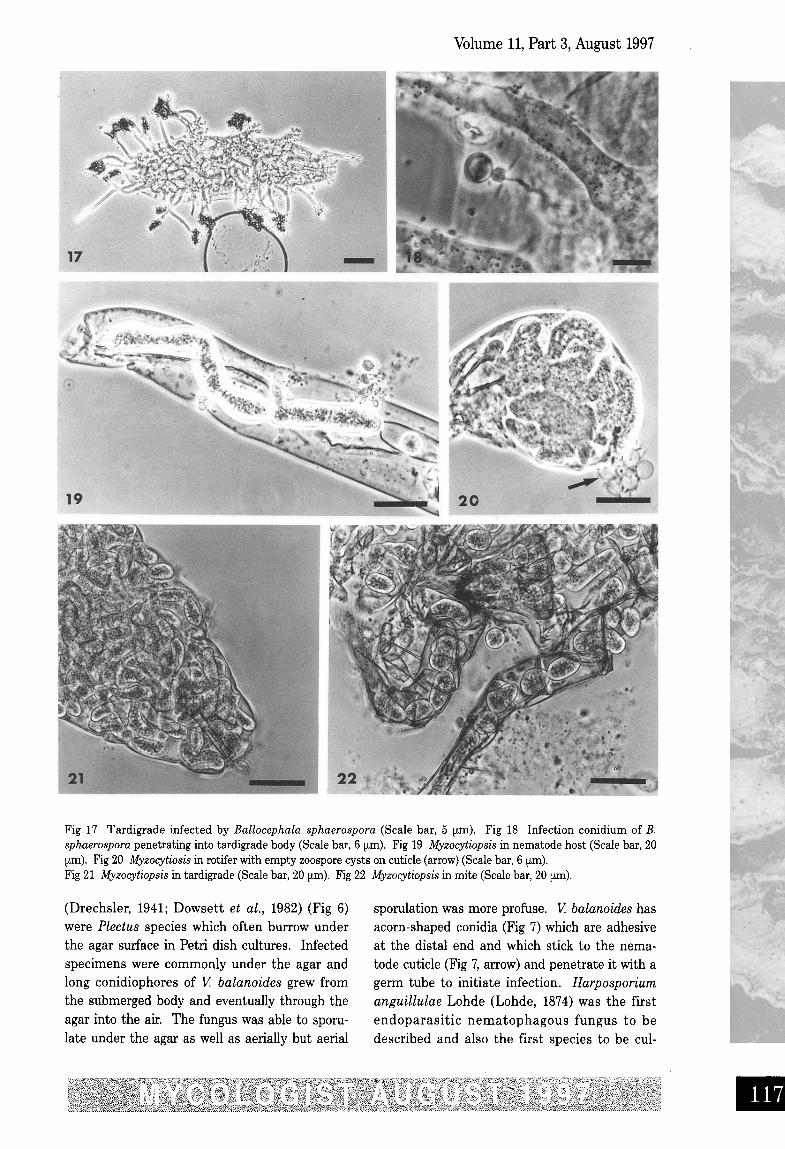
Fig 17 Tardigrade infected by Ballocephala sphaerospora (Scale bar, 5 um). Fig 18 Infection conidium of B.sphaerospora penetrating into tardigrade body (Scale bar, 6 um), Fig 19 Myzocytiopsis in nematode host (Scale bar, 20prn). Fig 20 Myzocytiosis in rotifer with empty zoospore cysts on cuticle (arrow) (Scale bar, 6 um),Fig 21 Myzocytiopsis in tardigrade (Scale bar, 20 um). Fig 22 Myzocytiopsis in mite (Scale bar, 20 urn).
(Drechsler, 1941; Dowsett et al., 1982) (Fig 6)were Plectus species which often burrow underthe agar surface in Petri dish cultures. Infectedspecimens were commonly under the agar andlong conidiophores of V balanoides grew fromthe submerged body and eventually through theagar into the air. The fungus was able to sporu-late under the agar as well as aerially but aerial
sporulation was more profuse. V balanoides hasacorn-shaped conidia (Fig 7) which are adhesiveat the distal end and which stick to the nema-tode cuticle (Fig 7, arrow) and penetrate it with agerm tube to initiate infection. Harposporiumanguillulae Lohde (Lohde, 1874) was the firstendoparasitic nematophagous fungus to bedescribed and also the first species to be cul-
•

•
Volume 11, Part 3, August 1997
tured artificially on agar medium (Aschner &Kahn, 1958). H. anguillulae and H. leptospira arecommon endoparasites found in a variety of sub-strates. These fungi both produce curved infec-tion conidia and also form chlamydosporesinside the nematode body. The otherHarposporium species found, H. cycloides Drech.(Drechsler, 1968), produced curved conidia fromthe infected nematode (Fig 8). In pure culture, inaddition to sharply pointed curved conidia, thisstrain of H. cycloides also produced arthroconidia(Fig 9) although these have not been observed inprevious studies (Glockling & Shimazu, 1997).Plesiospora globosa Drech . (Drechsler, 1970) isan unusual and rare endoparasitic fungus whichparasitises a species of slender, slow-movingnematode. It is unusual because it does not pro-duce any external conidiophores and the narrownecks of the conidiogenous cells protrudethrough the nematode cuticle and small spheri-cal conidia are formed externally. There havebeen few, if any, reports of this species since itsdiscovery but it was found on one occasion inrabbit dung (Oryctolagus cuniculi L.) in Englandinfecting a single nematode (Glockling, unpub-lished thesis data). On the previous occasion, P.globosa was not isolated into culture but thistime a pure culture was successfully obtainedfrom one of the two infected specimens. This isthe first time that this fungus has been grown onartificial media. In culture on CMA, P. globosaproduced small spherical conidia from flask-shaped and elongate conidiogenous cells (Fig 10).Conidia were produced in abundance and oftenformed in long chains in a similar mode to thatof Verticillium coccosporum (Drechsler) Gams(Drechsler 1941; Gams, 1988) which also infectsnematodes. Reinfection experiments to decipherthe mode of infection for P. globosa were not car-ried out at this time as the host nematode forthis fungus has not yet been identified. Theother nematophagous endoparasitic species iso-lated, Tolypocladium lignicola Barron (Barron,1983), also appears to be an uncommon fungus.It was initially isolated from rotten wood usingbaiting techniques with rotifers although, duringreinfection experiments, Barron noted thatrotifers ingested, digested and excreted largenumbers of T lignicola conidia, apparently usingthem as a food source. Only the occasional coni-dium lodged in the rotifer mastax region and ger-minated to produce infection. It may be that
species of nematodes, not rotifers, are in fact thetarget host organisms for this fungus. It was alsoisolated from nematodes in a sample of muntjac(Muntiaco) dung on a previous occasion inEngland (Glockling, unpublished thesis data)when nematodes were repeatedly infected inmixed culture containing bdelloid rotifers overseveral weeks. On CMA large numbers of disc-shaped conidia were produced and introductionof conidia into Rhabditis nematode culturesresulted in infection of several nematodesafter about 4 days (Fig 13). Infected nematodes(Fig 11), during the early stages of infection, hadone or several conidia adhering to the mouthwhich appeared to be penetrating via a germtube into the nematode body (Fig 12). If infec-tion of T lignicola is in fact by spore adhesion,this would be in accordance with several specieswhich are pathogenic to insects and mites .However, T parasiticum Barron (Barron , 1980),which is endoparasitic in rotifers , infects byspore ingestion. Another rotifer parasite, T.trigonosporum Barron (Barron, 1981), had such alow infection rate in trials that the infectionmethod could not be ascertained. For specieswith a capability of infecting both nematodesand rotifers , the infection method maydiffer according to the host organism. TheRotiferophthora species (Fig 13) infected rotifersin samples collected at the same time in summerfrom both Naon and Kawauchi. Many bdelloidrotifers succumbed to the infection which pro-duced an abundance of small infection conidiafrom flask-shaped conidiogenous cells and alsofrom aphanophialides . Multi-septate restingspores (dictyochlamydospores) were produced inthis species but they were very rare until theinfection, which persisted over several weeks,began to decline. Specimens infected during theend of the infection duration produced up tothree dictyochlamydospores which became largeand bullate at maturity. The Rotiierophihorasuperficially resembled R. turbinaspora Barron(Barron, 1985) and R. barronii Glockling(Glockling & Dick, 1994) but differed from thesespecies in the size and shape of the conidio-genous cells and conidia. Consequently, thisspecies is being described as new (Glockling, inpreparation).
The zoopagalean fungus which trapped amoe-bae on adhesive aseptate hyphae (Fig 14) wasrecovered from Amami rabbit dung from two col-

Table 1 Fungi isolated from Amami rabbit dung.
Species isolated Host
Volume 11, Part 3, August 1997
Trap/Infection method
Arthrobotrys superba
Arthrobotrys cladodes
Dactylaria thaumasia
Dactylaria candida
Dactylella leptospora
Dactylella ellipsospora
Harposporium anguillulae
Harposporium leptospira
Harposporium cycloides
Verticillium balanoides
Plesiospora globosa
Tolypocladium lignicola
Rotiferophthora sp.
Zoopagalean sp.
Ballocephala sphaerospora
Myzocytiopsis sp.
Nematode
Nematode
Nematode
Nematode
Nematode
Nematode
Rhabditis nematode
Rhabditis nematode
Rhabditis nematode
Plectus nematode
Nematode
Nematode
Bdelloid rotifer
Amoeba
Tardigrade sp.
Rhabditis nematode
Adhesive network
Adhesive network
Adhesive network
Passive ring/stalked knob
Passive ring/stalked knob
Stalked knob
Spore ingestion
Spore ingestion
Spore ingestion
Spore adhesion
?
Spore adhesion?
Spore ingestion
Adhesive hyphae
Spore adhesion
Zoospore encystment
lections taken in winter and summer. The fun-gus did not produce conidia on either occasion soit could not be identified to species level but themorphology of the assimilative hyphae was simi-lar to that of Stylopage areae Drech. (Drechsler,1935) and Zoopage mitospora Drech. (Drechsler,1938) although zygospores have not beendescribed for either of these species. At thepoint of attachment and penetration of the fun-gus into the amoeba host, the fungus producedseveral assimilative branches (Fig 15). Sphericalzygospores were formed inside the capturedamoebae after intertwining of two hyphae(Fig 16).
The tardigrade parasite, Ballocephalasphaerospora Drech. (Drechsler, 1951), producedshort conidiophores from the tardigrade bodyand masses of infection conidia were formedfrom short, swollen conidiogenous cells (Fig 17).Later, resting spores developed inside the body.Infection was initiated by small spherical sporeswhich attached to the host cuticle and penetrat-ed with a fine germ tube (Fig 18). The species oftardigrade infected in the Amami rabbit dungappeared similar to the Macrobiotus speciesinfected in Drechsler's original isolate. There areonly three known species of Ballocephala, allinfecting tardigrades, None of these zygomyce-
tous fungi have been cultured at this time.The Myzocytiopsis fungus, which produced
motile biflagellate zoospores, infected Rhabditis
nematodes in several cultures derived from rab-bit dung collected from Naon and Kawauchi.The fungus formed swollen cylindrical zoospo-rangia which developed evacuation tubes torelease the zoospores (Fig 19). Rotifers were notoften infected but one specimen (Fig 20) wasfound to have thalli of Myzocytiopsis in the egginside the rotifer body, and empty zoospore cystswere still attached to the rotifer cuticle (arrow).Zoospores emerged from the evacuation tubes ofthe zoosporangia initially as an amoeboid mass,but individual zoospores became apparent after aperiod of slow movement. Tardigrades were alsooccasionally infected by the fungus (Fig 21) andone mite also succumbed to infection (Fig 22).
This is the first record of a mite being attackedby Myzocytiopsis.
Conclusions
A diversity of common and rare fungi and onenew species were found in the Amami rabbitdung. The occurrence of several host-specificendoparasitic and predacious fungi as well asthose with a broad host range reflected the widediversity of host organisms which had colonized
•

Volume 11, Part 3, August 1997
the dung habitat. Rarely reported species ofendoparasitic fungi are usually associated withthe rarer nematode species which thrive within anarrow range of conditions and which are not sofecund as common species . The presence ofDactylaria candida and Rotijerophthora in cul-tures prepared from collections taken from bothKawauchi and Naon during the same collectiontrip showed that there was effective local disper-sal over 10 km. The Amami rabbit dung provedto be a good source of predacious fungi, especial-ly those capturing nematodes on adhesive knobs.
The importance of dung as a habitat for fungiwhich attack microscopic animals has not yetbeen thoroughly investigated, but it wouldappear that a rabbit pellet provides a choice tem-porary habitat for several transitory host-specificfungi as well as those with greater tolerance.
Acknowledgements
We thank the Science and Technology Agency(STA), Japan, for funding this research.
ReferencesAschner, M. & Kohn , S. (1958) The biology of
Harposporium anguillulae. Journal of GeneralMicrobiology 19: 182- 189.
Barron, G. L. (1980) Fungal parasites of rotifers : a newTolypocladium with underwater conidiation.Canadian Journal of Botany 58: 439 - 442.
Barron, G. L. (1981) Two fungal parasites of rotifers.Canadian Journal of Botany 59: 1449- 1455.
Barron, G. L. (1983) Structure and biology of a newTolypocladium species attacking bdelloid rotifers.Canadian Journal of Botany 61: 2566 - 2569.
Barron, G. L. (1985) Fungal parasites of rotifers :Diheterospora. Canadian Journal of Botany 63: 211 -222.
Borrebaek, C. A. K, Mattiasson, B. & Nordbring-Hertz,B. (1984) Isolation and partial characterization of acarbohydrate-binding protein from a nematode-trap-ping fungus. Journal of Bacteriology 159:53 -56.
Cooke, R. C. (1963) Ecological characteristics of nema-tode-trapping hyphomycetes. Annals of AppliedBiology 52: 431 - 437.
Corda, A. C. J . (1839) Pracr-flora EuropaeischerSchimmelbildungen. Leipzigund Dresden .
Dawson, M. R. (1981) Evolution of th e modernLagomorphs. In: Meyers, K. & Macinnes, C. P. (eds.),Protection of the world Lagomorph Conference,Guelph University press, Guelph, Ontario, 1 - 8.
Dowsett, J . A., Reid , J . & Hopkin, A. (1982) OnCephalosporium balanoides Drechsler . Mycologia 74:687 - 690.
Drechsler, C. (1935) Some non-catenulate conidial phy-comycetes preying on terricolous amoebae. Mycologia27: 176- 203.
Drechsler, C. (1937) Some hyphomycetes that prey onfree-living nematodes. Mycologia 29: 447 - 550.
Drechsler, C. (1938) New Zoopagaceae capturing andconsuming soil amoebae. Mycologia 30: 137- 157.
Drechsler, C. (1941) Some hyphomycetes parasitic onfree-living terricolous nematodes. Phytopathology 31:773- 801.
Drechsler, C. (1951) An entomophthoraceous tardigradeparasite producing small conidia on propulsive cells inspicate heads. Bulletin of the Torrey Botanical Club78: 183- 200.
Drechsler, C. (1968) A new nematode-destroyingHarposporium. American Journal of Botany 55: 1251-1253.
Drechsler, C. (1970) A nematode-destroying parasitebearing lageniform conidiiferous branches on endozoichyphae. Sydowia 24: 173- 176.
Friman, E. (1993) Isolation of trap cells from thenematode-trapping fungus Dactylaria candida.Experimental Mycology 17: 368 - 370.
Gams, W. (1988) A contribution to the knowledge ofnematophagous species of Verticillium. NetherlandsJournal of Plant Pathology 94: 123- 148.
Glockling, S. L. Three new species of Rotiferophthoraattacking bdelloid rotifers in Japan. MycologicalResearch (in preparation).
GIockiing, S. L. & Dick, M. W. (1994) Five new species ofRotijerophthora isolated from dung. MycologicalResearch 98: 833 - 844.
GIockiing, S. L. & Shimazu, M. (1997) Culturing of threespecies of endoparasitic fungi infecting nematodes.MycologicalResearch 101:55 - 60.
Grove, W. B. (1885) New or noteworthy fungi. Journal ofBotany 22, 129.
Lohde, G. (1874) Einige neue parasitische Pilze.Tageblatt der 47. Versammlung deutscherNaturforscher und Artze in Breslau . p 203.
Saccardo, P. A. (1886) Sylloge Fungorum 4, 195.Saikawa, M. (1985) Ultrastructural features of the non-
constricting-ring trap in Dactylella leptospora .Transactions of the Mycological Society of Japan 26:209 - 213.
Sugiyama, K. (1990) The Arnami rabbit Pentalagus fur-nessi. In : Rabbits, hares and pikas. Chapter 11, 140 -142. mCN, Gland, Switzerland.
Yamada, F., Sugimura, K. & Koyanagi, H. (1995). Statusof Arnami rabbit Pentalagus [urnessi and requirementof ecological engineering for its conservation .Conference newsletter. International conference onhabitat fragmentation, infrastructure and the role ofecological engineering.
•