a truncated version of an adp-glucose … · the plant cell, vol. 6, 601-612, may 1994 0 1994...
TRANSCRIPT

The Plant Cell, Vol. 6, 601-612, May 1994 0 1994 American Society of Plant Physiologists
A Truncated Version of an ADP-Glucose Pyrophosphorylase Promoter from Potato Specifies Guard Cell-Selective Expression in Transgenic Plants
-
Bernd Müller-Rober,' Ursula La Cognata, Uwe Sonnewald,* and Lothar Willmitzer lnstitut für Genbiologische Forschung Berlin GmbH, lhnestrasse 63, 14195 Berlin, Germany
ADP-glucose pyrophosphorylase (AGPase) is a key regulatory enzyme in starch biosynthesis in higher plants. A 3.2-kb promoter of the large subunit gene of the AGPase from potato has been isolated and its activity analyzed in transgenic potato and tobacco plants using a promoter-P-glucuronidase fusion system. The promoter was active in various starch- containing cells, including guard cells, tuber parenchyma cells, and the starch sheath layer of stems and petioles. No expression was observed in mesophyll cells. Analysis of various promoter derivatives showed that with respect to expres- sion in petioles and stems, essential elements must be located in the 5' dista1 region of the promoter, whereas elements important for expression in tuber parenchyma cells are located in an interna1 fragment comprising nucleotides from posi- tions - 500 to -1200. Finally, a 0.3-kb 5' proximal promoter fragment was identified that was sufficient to obtain exclusive expression in guard cells of transgenic potato and tobacco plants. The implications of our observations are discussed with respect to starch synthesis in various tissues and the use of the newly identified promoter as a tool for stomatal biology.
INTRODUCTION
ADP-glucose pyrophosphorylase (ATP: a-glucose-1-phosphate adenylyl transferase, EC 2.7.7.27; AGPase) plays a central role in the biosynthesis of starch in both photosynthetic and non- photosynthetic plant tissues (for review, see Preiss, 1991). Within chloroplasts and amyloplasts, the enzyme catalyzes the synthesis of ADP-glucose and PPi from glucose-l-phos- phate and ATP. ADP-glucose functions as the glucosyl donor for a-glucan synthesis by various starch synthases (Preiss,
The importance of AGPase in starch biosynthesis has been shown by mutants deficient in AGPase activity in storage organs and leaves (e.g., Tsai and Nelson, 1966; Lin et al., 1988a, 1988b), by kinetic models (Pettersson and Ryde-Pettersson, 1989), by control analysis of photosynthate partitioning (Neuhaus and Stitt, 1990), and by the use of transgenic plants in which AGPase activity has either been decreased by an antisense RNA approach (leading to reduced starch levels; Müller-Rober et al., 1992) or increased by ectopic expression of a bacterial AGPase (leading to higher starch levels; Stark et al., 1992).
Plant AGPase is supposed to be a tetrameric protein com- posed of two different but similar polypeptides (e.g., see Morell et al., 1987; Lin et al., 1988b). The size of the two small subunits ranges from 50 to 55 kD, and the size of the two large sub- units is from 51 to 60 kD, depending on the plant species (Kleczkowski et al., 1991). cDNA clones encoding the differ- ent subunit polypeptides have been isolated from a number
2 Current address: lnstitute for Plant Genetics and Crop Research, Corrensstrasse 3, 06466 Gatersleben, Germany.
1991).
To whom correspondence should be addressed.
of monocot and dicot plant species (e.g., see Anderson et al., 1989; Olive et al., 1989; Bae et al., 1990; Bhave et al., 1990; Müller-Rober et al., 1990; du Jardin and Berhin, 1991; Nakata et al., 1991; Villand et al., 1992a, 1992b; Ainsworth et al., 1993). By comparing the primary structures of 11 plant AGPase proteins, Smith-White and Preiss (1992) recently pointed out the existence of at least three types of AGPase polypeptides: (1) small subunit, that is present in photosynthetic and non- photosynthetic tissues; (2) large subunit, that is found in nonphotosynthetic tissues; and (3) large subunit, that is pres- ent in photosynthetic cells.
The complexity of the plant's AGPase system is further un- derscored by the observation of multiple AGPase polypeptides in potato tubers (Okita et al., 1990) and the rice and pea en- dosperm (Hylton and Smith, 1992; Nakamura and Kawaguchi, 1992). Developmental (coarse) control of AGPase mRNA ex- pression has been observed in several plant species (Krishnan et al., 1986; Reeves et al., 1986; Müller-Rober et al., 1990; Anderson et al., 1991; Villand et al., 1992a). In potato, the small @e., AGPase 6) subunit was strongly expressed in a variety of tissues tested, including leaves and tubers. By contrast, ex- pression of the large (i.e., AGPase S) subunit was high in tubers and generally weak to moderate in leaves (Müller-Rober et al., 1990). lncubation of excised leaves in sucrose led to a strong accumulation of the AGPase S transcript (Müller-Rober et al., 1990). 6y contrast, expression of the AGPase B mRNA was only slightly influenced by the presence of externa1 sucrose.
Starch synthesis takes place in several different cells and organs of the plant and is of major importance for carbon

602 The Plant Cell
partitioning. One of the key regulatory mechanisms is the al- Iosteric activation of the AGPase by 3-phosphoglycerate and the inhibition by Pi (see Preiss, 1991). This manner of regula- tion has been found in a wide range of AGPases from photosynthetic and nonphotosynthetic tissues, including guard cells (Robinson et al., 1983; Outlaw and Tarczynski, 1984; Preiss, 1988). Recently, Kleczkowski et al. (1993) showed the insensitivity of the barley endosperm AGPase to 3-phospho- glycerate and Pi regulation, demonstrating heterogeneity within the allosteric regulatory properties of the plant enzyme.
Understanding regulation of carbon partitioning and starch biosynthesis in higher plants not only requires knowledge about biochemical characteristics of the enzymes involved, but also of the cellular and organ-specific expression of the genes en- coding these enzymes. Here, we report the isolation of a genomic DNA fragment containing regulatory elements of the large subunit of the potato AGPase. The promoter was fused to the 13-glucuronidase (gus) reporter gene, and its expression pattern was analyzed in transgenic potato and tobacco plants.
RESULTS
Isolation of a Potato ADP-Glucose Pyrophosphorylase Large Subunit Promoter
A genomic clone homologous to the large subunit cDNA of the potato AGPase (AGPase S; M011er-R~ber et al., 1990) was isolated from a genomic library prepared from the monohaploid potato line AM 80•5793 (Liu et al., 1991). DNA sequences were obtained for several subclones and compared with the 5' end of the full-length AGPase S cDNA of plasmid S9-D (see
5" p H 6 - 1 3" HhldIIl* Nde] HindiiI Bglll* PstI* Hincli
H~cII Nsil N¢o] ~ | a l ~B~]ff~]a S E NdI|PL ,~ iEco = Hmdm I I
0,5 kb TATA-Box i i m
A G P a s S P ~ cDNA I e - r o m o t e r | [ ~
HinclI 3.2 kb BgllI
Figure 1. Schematic Alignment of the 5.2-kb AGPaso S Genomic Frag- ment with the Corresponding cDNA.
For subcloning procedures, see Methods. The identity of the insert of pH6-1 was verified by sequencing an ~2.3-kb region between the Hindlll and Pstl sites that are marked with asterisks. Comparison with the 5' end of the full-length cDNA S9-D (see Methods) allowed the identification of exon and.intron sequences in the genomic clone. The sequence between the Hindlll and Bglll sites marked with asterisks is shown in Figure 2A. The transcriptional start point was tentatively assigned by comparing the sequence of the genomic DNA fragment with the sequence of the cDNA. The location of the cDNA relative to the genomic fragment and the position of the AGPase S promoter used for fusion to the gus reporter gene are shown as thick lines. The posi- tion of the start codon of the AGPase lying at the beginning of the second exon is shown by '~.TG" El, E2, and E3, exons 1, 2, and 3; I1 and 12, introns 1 and 2; PL, polylinker of the pUC19 vector.
Methods). A 5.2-kb Hincll fragment was found to comprise a 3.2-kb promoter fragment and part of the AGPase coding re- gion. A schematic alignment of the cDNA and the genomic fragment is shown in Figure 1. With the exception of two in- trons (corresponding to positions 190/191 and 426•427 of the cDNA), complete identity was found between nucleotides 1 to 484 of the cDNA and the genomic clone. (No effort was made to sequence further downstream regions of the genomic clone.)
The DNA sequence of the 1.2-kb HindllI-Bglll genomic seg- ment (see Figure 1) is shown in Figure 2A. Comparison with the full-length cDNA S9-D revealed a 56-bp overlap with the 5' untranslated leader of the cDNA. A TATA-box-like sequence element ("TATAAATA") is present at position - 66•- 59 upstream of the 5' end of the corresponding cDNA. Two direct repeats (each repeating sequence 50 nucleotides long, with five mis- matches) were identified (Figure 2A).
The 3.2-kb Promoter of the AGPase S Gene Is Most Active in Nodal and Internodal Stem Sections of Transgenic Potato Plants
To analyze the expression pattern of the AGPase S promoter, the 3.2-kb promoter fragment was transcriptionally fused to the gus reporter gene of the binary vector pBI101.1 (Jefferson et al., 1987). The structure of the chimeric AGPase S promoter- gus gene is shown in Figure 2B. This plasmid was used to transform potato plants using the Agrobacterium-mediated gene transfer technique. Approximately 50 independent trans- genic plants (B-GUS-1 transformants) were analyzed.
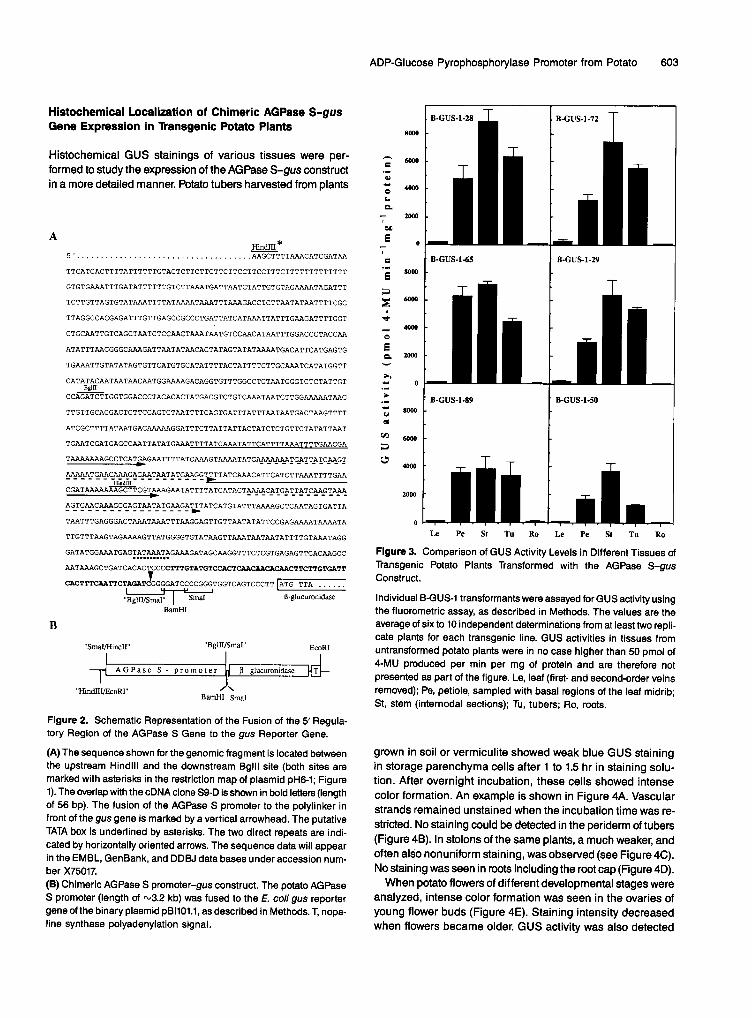
To gain insight into quantitative aspects of the expression of the AGPase S-gus construct, various tissues of transgenic potato plants were homogenized and analyzed by way of the fluorescence assay. As shown in Figure 3, the highest activi- ties were found in stem sections, reaching more than 7000 pmol of 4-methylumbelliferone (4-MU) produced per min per mg of protein in some of the transformants. The activity in nodes was higher by a factor of 1.2 to 2 compared to that in inter- nodes (data not shown). Strong GUS activity (up to about 6000 pmol of 4-MU produced per min per mg of protein) was also found in petioles (sampled with basal parts of the leaf midrib) and in tubers of all transformants tested, whereas much lower activity, which was only slightly above background (between 50 and 120 pmol of 4-MU produced per min per mg of protein as compared to 2 to 20 pmol of 4-MU produced per min per mg of protein in nontransgenic control plants), was found in leaves from which the first- (midrib) and second-order veins had been removed.
The level of expression varied over a wide (more than 10- fold) range, as shown in Tables 1 and 2; this observation has been made in the case of many other transgenes (e.g., Peach and Velten, 1991). When DNA gel blot analysis was performed on various transgenic lines differing in their expression lev- els, no correlation was observed between AGPase S-gus gene copy number and expression level, which again is in agree- ment with many other cases where this problem has been analyzed (for a review, see Weising et al., 1988).
ADP-Glucose Pyrophosphorylase Promoter from Potato 603
Histochemical Localization of Chimeric AGPase S-gus Gene Expression in Transgenic Potato Plants
Histochemical GUS stainings of various tissues were per- formed to study the expression of the AGPase S-gus construct in a more detailed manner. Potato tubers harvested from plants
A
B
HindHI* 5". . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . AAGCTTTTAAACATCGATAA
TTCATCACTTTTATTTTTTGTACTCTTCTTCTTCTTCCTTCCTTTCTTTTTTTTTTTTT
GTGTGAAATTTGATATTTTTTGTCTTAAATGATTAATCTATTGTGTAGAAAATAGATTT
TCTTGTTAGTGTATAAATTTTATAAAATAAATTTAAAGACCTCTTAATATAATTTTCGC
TTAGGCCACGAGATTTGTTGAGCCGCCCTGATTATCATAAATTATTTGAAGATTTTGGT
CTGCAATTGTCAGCTAATCTCCAACTAAATAATGTCCAACATAATTTGGACCCTACCAA
ATATTTAACGGGCAAAGATTAATATAACACTATAGTATATAAAATGACATTCATGAGTG
TGAAATTGTATATAGTGTTCATGTGCATATTTTACTATTTTCTTGCAAATCATATGGTT
CATATACAATAATAACAATGGAAAAGACAGGTGTTTGGCCTGTAATGGGTCTCTATTGT BgIH
CCAGATCTTGGTGGACCCTACACACTATGACGTCTGTCAAATAATCTTGGAAAAATAAC
TTGTTGCACGACTCTTCGAGTCTAATTTTCAGTGATTTATTTAATAATGACTAAGTTTT
ATCGCTTTTATAATGACAAAAAGGATTTCTTATTATTACTATCTCTGTTCTATATTAAT
TGAATCGATGAGCCAATTATATGAAATTTTATCAAATATTCATTTTAAATTTTGAACGA
TAAAAAAAGCCTCATGGAGAATTTTATCAAAGTAAKATATGAAAAAAATGATTATCAAGT
AAAAATGAACAAAGAGAATAATATGAAGG [T~TTTATCAAACATTCATCTTAAATTTTGAA . . . . . . - . ~ d ~ . . . . . . . . . . P" CGATAAAAAAAGCTTCGTAAAGAATATTTTATCATAGTAAAACATGATTATCAAGTAAA IB ..............
AG!GA:%C'~_GgG_AGT_:~TATG~kGAT~TATCATGTATTTA.z~AGCTCAATAGTGATTA TAATTTGAGGGACTAAATAAATTTAAGGAGTTGTTAATATATTCCGAGAAAATAAAATA
TTGTTTAAGTAGAAAAGTTATGGGGTGTATAAGTTAAATAATAATATTTTGTAAATAGG
GATATGGAAATGAG~A~,.A~**~AGAAAGATAGCAAGGTTTCTCGTGAGAGTTCACAAGCC
AATAAAGCTGATCACACTCCCCTTTGTATGTCCACTCAACAAC/%CA%ACTTCTTGTGATT
CACTTTCAATTCT T T A AGACGGGG TCCCCGGGTGGTCAGTCCCTTIATG TTA I r
, ,BgllUSmar.~Sm~ ~-gluc~omdase i
BamHI
"Smal/Hincll" "BglII/Smal" EcoRI
~ AGPase S - p . . . . t e r / ~ [3-gl . . . . . idase ~ - -
"HindIIl/EcoRI" BamHI Sinai
Figure 2. Schematic Representation of the Fusion of the 5' Regula- tory Region of the AGPase S Gene to the gus Reporter Gene.
(A) The sequence shown for the genomic fragment is located between the upstream Hindlll and the downstream Bglll site (both sites are marked with asterisks in the restriction map of plasmid pH6-1; Figure 1). The overlap with the cDNA clone S9-D is shown in bold letters (length of 56 bp). The fusion of the AGPase S promoter to the polylinker in front of the gus gene is marked by a vertical arrowhead. The putative TATA box is underlined by asterisks. The two direct repeats are indi- cated by horizontally oriented arrows. The sequence data will appear in the EMBL, GenBank, and DDBJ data bases under accession num- ber X75017. (B) Chimeric AGPase S promoter-gus construct. The potato AGPase S promoter (length of ~3.2 kb) was fused to the E. coil gus reporter gene of the binary plasmid pBI101.1, as described in Methods. T, nopa- line synthase polyadenylation signal.
80@0
6000
"~ 4000 0 L.
- - 2000 I
0 i l-
° .
4 o
~ 0
B-GUS-I-28 B - G U S - 1 - 7 2
i i , 7 -
• B-GUS-I-65 B-GUS-I-29
i l l i B-GUS-1-89 B - G U S - 1 - 5 0
2000
0
Le P e S t Tu Ro Le P e S t Tu Ro
Figure 3. Comparison of GUS Activity Levels in Different Tissues of Transgenic Potato Plants Transformed with the AGPase S-gus Construct.
Individual B-GUS-1 transformants were assayed for GUS activity using the fluorometric assay, as described in Methods. The values are the average of six to 10 independent determinations from at least two repli- cate plants for each transgenic line. GUS activities in tissues from untransformed potato plants were in no case higher than 50 pmol of 4-MU produced per min per mg of protein and are therefore not presented as part of the figure. Le, leaf (first- and second-order veins removed); Pe, petiole, sampled with basal regions of the leaf midrib; St, stem (internodal sections); Tu, tubers; Ro, roots.
grown in soil or vermiculite showed weak blue GUS staining in storage parenchyma cells after 1 to 1.5 hr in staining solu- tion. After overnight incubation, these cells showed intense color formation. An example is shown in Figure 4A. Vascular strands remained unstained when the incubation time was re- stricted. No staining could be detected in the periderm of tubers (Figure 4B). In stolons of the same plants, a much weaker, and often also nonuniform staining, was observed (see Figure 4C). No staining was seen in roots including the root cap (Figure 4D).
When potato flowers of different developmental stages were analyzed, intense color formation was seen in the ovaries of young flower buds (Figure 4E). Staining intensity decreased when flowers became older. GUS activity was also detected

604 The Plant Cell
layer was very similar to the starch sheath observed in the stem (see above). An investigation for the presence of the starch
Table 1. GUS Activity Levels in Tubers of Transgenic Potato Plants
GUS Activitya (pmol 4-MU min-l mg-l protein) % of
Original sheath showed that thiscell layer is present only in the petiole and in the basal regions of major veins. No starch sheath was
Transformationb MaxC Mind Avae Activitv' detected in any of the minor veins of the leaf. The distribution
B-GUS-1 6335 f 675
S1-A1 2734 f 219 S1 -A2 1836 f 123 S1 -A4 79 f 39 S1-A13 454 f 72 S1-A14 316 f 12
211 2 54
97 f 38 28 f 1 5 f 3
32 f 2 20 f 13
"
2918
1068 267 24
129 66
Wild Type 10 f 2 2 f 1 4
1 O0
36 9 1 4 2
~ ~ ~ ~ ~ ~ ~ _ _ _ _ ~
a Ten independent transgenic lines were selected on the basis of strongest histochemical staining in guard cells and used for each anal- ysis. Tissues were harvested at around noon from plants grown in the greenhouse. At least six determinations from two to three individual plants were performed for each transgenic line. Data are presented as the mean t SE of the GUS activities.
For constructs, see Figures 28 and 6, respectively. Max, highest GUS activity measured among the transgenic lines. Min, Iowest GUS activity measured among the transgenic lines.
* Avg, average GUS activity measured in the 10 transgenic lines. 'The average GUS activity measured in the 10 B-GUS-1 lines was set at 100%.
in cross-sections of anthers (Figure 4F) and in mature pollen grains (data not shown). lmmature pollen grains remained com- pletely unstained even after more than 24 hr of incubation.
In cross-sections prepared from stems of potato 6-GUS-1 transformants, GUS staining was restricted to a single cell layer surrounding the vascular tissue. A representative example is shown in Figure 4G. This cell layer was identified as a starch sheath, which has been described for a variety of plant spe- cies, including potato and tobacco. Figure 4H shows that starch granules can easily be detected in this cell type. Dueto a higher density of vascular tissues and corresponding starch sheath cells, GUS staining was more intense in nodal than in interno- da1 stem regions (data not shown). The only other cells stained in the stem were guard cells, which frequently appeared in the stem epidermis, as shown in Figure 5A. All other cells, even parenchyma cells, which very often contained starch granules, remained completely unstained.
When leaves were incubated for 15 to 24 hr in staining s o b tion, a strong blue staining was visible along the petiole and the midrib (first-order vein), with the intensity decreasing from the basal to tip region of the leaf. In addition, second-order veins and often also third-order veins showed staining, again with an acropetal decrease in intensity. A representative ex- ample is shown in Figure 56. No GUS staining was seen in the tip region of the major veins or in any of the smaller leaf veins even after prolonged (up to 48 hr) incubation. Figure 5C shows that in cross-sections strong GUS staining appeared in a single cell layer adjacent to the vascular tissues. This cell
of the starch sheath therefore corresponded well to the de- crease of GUS staining intensity toward the tip region of the veins and to the absence of GUS staining in small @e., higher order) veins. Guard cells distributed within the epidermis of the major veins were easily stained. No GUS activity was seen in any other cell type of the petiole (and midrib), including pa- renchyma cells.
Mesophyll cells that accumulated massive amounts of tran- sitory starch were completely devoid of any GUS staining, irrespective of the developmental stage of the leaf or length of incubation (see Figure 5D for an example). Also, leaf trichomes never showed any GUS staining (Figure 5E). Most interestingly, stomatal guard cells of both the abaxial and the adaxial leaf surface were stained after incubation overnight. An example is shown in Figure 5F. The fact that guard cells are the only cell type (apart from the major veins) that is stained in leaves is in agreement with the very low levels of GUS ac- tivity determined by the fluorescence assay (see above). Guard cells are estimated to represent only about 1% of the cellular population of a leaf (Hsiao, 1976).
Among the transgenic plants analyzed in about 10 to 20% of the cases, GUS staining observed in leaf guard cells was classified as strong or medium, whereas in about 30 to 40% of the plants harboring the full-length AGPase S promoter, stain- ing in guard cells was classified as weak. In the remaining 30 to 40% of the plants, no staining was found in guard cells after incubation overnight.
Expression or nonexpression in guard cells correlated in a strict manner with expressionlnonexpression in other tissues (such as tubers or petioles), suggesting that this is a typical position effect. In the few cases where quantitative numbers are given with respect to nonexpressing transgenic plants, a similar proportion of plants was identified as nonexpressers
~~~ ~
Table 2. GUS Activity Levels in Petioles of Transgenic Potato Plants
% of Original
Transformationb MaxC Mind Avge Activity'
GUS Activitya (pmol 4-MU min-l mg-l protein)
B-GUS-1 6322 f 1358 437 f 145 2668 100
S1-A1 289 f 32 7 f 2 73 3 S1-A2 160 f 49 15 f 4 69 3 S1-A4 320 f 57 12 f 4 67 3 S1-A13 8104 f 1212 104 f 28 2671 100 Sl-AI4 7003 f 1550 162 f 49 2682 101
Wild Type 40 f 10 2 f 1 10
Table 1 for descriptions.

ADP-Glucose Pyrophosphorylase Promoter from Potato 605
St
vs
an
Figure 4. Histochemical Localization of GUS Activity in Potato Plants Transformed with the AGPase S-gus Construct (B-GUS-1 Transformants).
(A) Transverse sections through potato tubers. The unstained tuber slice is from a wild-type plant.(B) Tuber section stained for 3 hr.(C) Stolons of an untransformed plant (top) and a transgenic plant (other stolons).(D) Root tip stained for 24 hr.(E) Longitudinal section through a flower bud (length of 13 mm).(F) Cross-section through an anther.(G) Cross-section through a stem. Staining was performed for 2 hr.(H) Cross-section through a stem. The tissue was stained with Lugol's solution to show starch granules within the starch sheath.Unless otherwise indicated, GUS staining was performed for 15 hr.In (B), bar = 0.5 cm; in (D), (G), and (H), bars = 200 urn; in (F), bar = 0.5 mm. an, anther; fi, filament; lo, locule; ov, ovary; p, periderm; pa,parenchyma cells; ss, starch sheath; st, stolon; vs, vascular strand; xy, xylem.

606 The Plant Cell
\
tr
Figure 5. Localization of GUS Activity in Potato Plants Transformed with the AGPase S-gus Construct.
(A) Guard cell in the stem epidermis.(B) Close-up of a leaf.(C) Cross-section through the leaf midrib (central region). Staining was performed for 30 min.(D) Cross-section through the upper part of a leaf.(E) Trichome.(F) Leaf guard cells in an epidermal strip.Unless otherwise indicated, staining was performed for 15 hr. In (A), (D), and (F), bars = 20 \im; in (B), bar = 0.5 cm; in (C), bar = 200 urn;in (E), bar = 50 urn. e, epidermis; gc, guard cell; m, mesophyll; mr, midrib; pa, parenchyma cells; ss, starch sheath; sv, second-order vein; tr,trichome; xy, xylem.

ADP-Glucose Pyrophosphorylase Promoter from Potato 607
(20 to 50%; cf., e.g., Sanders et al., 1987; Keil et al., 1990; Peach and Velten, 1991).
A Truncated Derivative of the AGPase S Promoter Leads to a Guard Cell-Specific Expression in Transgenic Potato Plants
As described above, the AGPase S promoter is able to induce strong gus expression in various starch-producing cells and tissues of transgenic potato plants, including the guard cells. To determine whether promoter regions important for the ex- pression in the petiole, stems, and tubers can be separated from the expression in guard cells, a series of 5’ and internal deletions was generated in the 3.2-kb promoter fragment. These deletions were fused to the gus reporter gene (see Methods) and were transformed in potato plants. A schematic representation of the different deletion constructs is given in Figure 6. With the exception of Sl-A5 transformants, weak GUS activity was observed in leaves (sampled without first- and second-order veins) of many transformants. Construct S1-A5, which does not contain the TATA box and the basal promoter, led to background levels of GUS activity. The results for tubers are shown in Table 1. Highest GUS activity was observed in plants containing the 3.2-kb promoter fragment. This activity decreased by -60% when 1.2 kb of the most dista1 5‘end was deleted (construct Sl-Al). Further 5‘ deletions down to -1.2
HincII TATA-Box
A 1
A 2
A 4
A 5
A 13
A 14
Figure 6. Schematic Representation of Promoter Deletion Constructs.
The end points of the deletions are indicated below the relevant re- striction sites and are numbered from the Bglll site used for the subcloning of the promoter fragments (see Methods). The 1.2-kb pro- moter fragment between the Hincll and Ncol sites in contructs B-GUS-l, Sl-Al3 (A13), and 3-814 (A14) is not shown to scale (indicated by black bars). The horizontal arrows shown in construct B-GUS-1 mark the positions of the two direct repeats (for sequences, see Figure 2A). L, 56-bp untranslated 5’leader sequence; gus, pglucuronidase gene.
kb (construct S1-A2) led to a decrease of -90%. Finally, a 5’ deletion down to position -0.3 kb (construct Sl-A4) resulted in a complete loss of GUS activity in tubers.
The fluorometric data were supported by histochemical anal- ysis. Whereas clear blue staining was observed in tubers of S1-A1 transformants, a much weaker staining could be seen in Sl-A2 transgenic plants. No staining was observed in S1- A4, Sl-Al3, and S1-A14 transgenic tubers. (This conclusion is based on the analysis of three to five tubers from each trans- genic plant; each test included 10 independent transgenic lines.)
The results obtained for the fluorometric GUS determina- tion in petioles are summarized in Table 2. Highest GUS activity was maintained in plants transformed with the internal dele- tion constructs Sl-Al3 and S1-A14. On the other hand, deletion of the 5’-most 1.2-kb fragment (Sl-Al) resulted in a complete loss of GUS activity in petioles. When histochemical stainings were performed, no difference was seen between B-GUS-1 and 3 - 8 1 3 or %A14 transformants, that is, dark blue stain- ing occurred in cells of the starch sheath (data not shown). The same was true for stem sections of these transformants (data not shown). Stems, petioles, and veins from the other transgenic lines remained completely unstained, even when the incubation time was extended to 48 hr.
With respect to expression in leaves, none of the %A5 trans- formants showed any GUS staining in guard cells. In all of the other transformants, a similar distribution of plants showing strong, middle, weak, orno expression in leaf guard cells was observed for the different constructs, as described above for the full-length construct B-GUS-1. This distribution was also found for Sl-A4 transgenic plants that did not show staining in tubers and petioles (see above).
Figure 7A shows GUS staining in a leaf of one of the S1-A4 transformants at a higher magnification. The blue staining seen in the figure is dueto staining of guard cells. We have not de- tected GUS activity in other cells, including the vascular tissue. Therefore, by shortening the AGPase S promoter to -0.3 kb, we have been able to identify a promoter fragment that gives rise to exclusive expression in guard cells of transgenic potato plants.
Transgenic Tobacco Plants Harboring the B-GUS-1 or the Si-A4 Constructs Display Expression Patterns Similar to the Ones Observed in Potato
To analyze whether the expression profile conferred by the AGPase S promoter is maintained in other species, two chi- meric genes, that is, the full-length promoter construct B-GUS-1 and the truncated promoter construct S l d 4 resulting in a guard cell-selective expression, were introduced into tobacco plants. As shown in Figures 78 and 7C, GUS activity was observed in the major veins of leaves and within the petiole. Cross- sections showed strong staining in the starch sheath surround- ing the vascular tissue. The same was true for the stem (data not shown). In addition, guard cells showed an intense staining

608 The Plant Cell
B
sv
mr
i
pa
\
gc
~ss
Figure 7. Histochemical Localization of GUS Activity in Tissues of Transgenic Potato and Tobacco Plants.
(A) Close-up of a leaf of potato transformant S1-A4-43.(B) Leaves of tobacco B-GUS-1 transformants.(C) Cross-section through a leaf petiole of a tobacco B-GUS-1 transformant.(D) Leaf guard cell of a tobacco B-GUS-1 transformant. The leaf was not bleached before photography.Staining time in (A), (B), and (D) was 15 hr; in (C), staining was performed for 30 min. In (A), bar = 1 mm; in (C), bar = 0.5 mm; in (D), bar= 50 urn. Abbreviations are as given in Figure 5.
(Figure 7D), thus being in complete agreement with the dataobtained for potato. The expression pattern in flowers was verysimilar to the one observed for potato. Furthermore, histochem-ical analysis did not reveal any activity in roots, including theroot cap. When tobacco plants harboring the deletion deriva-tive S1-A4 were analyzed, GUS staining was restrictedexclusively to guard cells. Thus, the elements necessary andsufficient to give rise to a guard cell-selective expression inpotato also function in transgenic tobacco plants.
DISCUSSION
AGPase is one of the major regulatory enzymes in starch bio-synthesis in higher plants. We isolated genomic DNA fragmentsrepresenting the large subunit of the AGPase from potato. The5' upstream regulatory region of one of the fragments was fusedto the gus reporter gene and transferred into potato plants.
The AGPase is a plastidic enzyme and, hence, contains tar-geting signals at its N-terminal end. To avoid interference ofplastidial targeting information with GUS activity, we decidedto perform transcriptional instead of translational gus fusions.Expression of the AGPase-gus construct in leaves and stemsof transgenic plants was restricted to stomatal guard cells andto a specialized cell layer, the starch sheath. In addition, strongexpression was observed in mature pollen grains and develop-ing tubers. Thus, the AGPase S promoter is active in manybut not all starch-containing cells. Most surprising in this re-spect is the observation that no expression was observed inthe cells representing the major starch reservoirs in leaves (i.e.,mesophyll cells). Also, no staining was detected in parenchy-matous cells of the petiole and stem.
Currently, there is no explanation for the inactivity of thispromoter in leaf mesophyll cells. However, because thegenomic clone described here, with the exception of the twointrons, is completely homologous to a cDNA recently isolatedby us, it is likely that this promoter originates from an expressed

ADP-Glucose Pyrophosphorylase Promoter from Potato 609
gene. Thus, with respect to the clear requirement for a pro- moter driving AGPase S gene expression in mesophyll cells, two possibilities remain: (1) the promoter fragment isolated and used in this study is devoid of some regulatory elements (i.e., expression in mesophyll cells requires elements located in the coding or 3' untranslated region), or (2) there is a second AGPase S gene, with a different promoter, driving expression in mesophyll cells. We cannot distinguish between the two possibilities.
Recently, restriction fragment length polymorphism map- ping experiments revealed two loci in the potato genome homologous to the AGPase S cDNA (Gebhardt et al., 1993). This observation is compatible with the putative existence of a second gene with a different promoter specificity. The latter interpretation would also offer an explanation for the fact that the expression of the gus gene driven by the promoter de- scribed here is not inducible by sucrose (data not shown). This observation is seemingly in contrast to our previous results. In this study, we observed a sucrose-dependent accumula- tion of AGPase S homologous mRNA in detached leaves (Müller-Rober et al., 1990). The exact number of genes en- coding AGPase polypeptides in any plant is not known. In potato, at least four different AGPase genes are expressed (Müller-Rober et al., 1990; U. La Cognata, L. Willmitzer, and B. Müller-Rober, manuscript in preparation).
Starch biosynthesis occurs in many different cell types. With respect to photoassimilates, these can be divided into exporters and importers. It is interesting to note that the expression pat- tern displayed by the AGPase promoter used in this study is restricted to various cell types that are all net importers of pho- toassimilates, such as tubers, the starch sheath cells of the stem and petioles, pollen grains, ovaries, and guard cells. No activity was observed in the main exporters of photoassimi- lates @e., the mesophyll cells of leaves). It is conceivable, although not certain, that starch biosynthesis is regulated in a different manner in photoassimilate importers versus ex- porters. One way to achieve such regulations could occur by way of the fusion of different promoters to the key enzyme of starch biosynthesis (i.e., the AGPase). We are currently pur- suing this possibility by isolating other genomic clones homologous to the AGPase S cDNA.
By analyzing the expression pattern of the gus gene fused to various derivatives of the AGPase S promoter in transgenic potato plants, different regions responsible for the expression in the petioles/stems, the tubers, and the guard cells were dis- tinguished. With respect to expression in petioles and stems, essential elements must be located in the distal 5' upstream region of this promoter, because a 5' deletion devoid of only the most distal 1.2 kb was completely inactive in the starch sheath of the vascular tissue. In tubers, a more gradual de- crease in activity was seen upon deleting 5' regions of the promoter. However, when a 0.7-kb-long interna1 fragment en- compassing nucleotides from position -0.5 to -1.2 kb was removed (construct Sl-A13), a more than 90% decrease in GUS activity occurred, indicating that a sequence element essen- tia1 for expression in tubers is present in the corresponding
region of the AGPase promoter. Finally, the most proxima10.3 kb of the promoter are necessary and sufficient to give full expression in stomatal guard cells. In addition, this expres- sion is highly specific because no other tissue showed activity upon histochemical analysis. To the best of our knowledge, this represents the first functional identification of a guard cell-specific promoter.
While this manuscript was in preparation, the promoter of the Arabidopsis rhal gene (encoding a small GTP binding pro- tein) was reported to be active in developing guard cells and severa1 other tissues (including flowers and root tips) of Arabi- dopsis plants (Terryn et al., 1993). No deletion analysis was performed on the rhal promoter. Therefore, it is currently not known whether expression in guard cells is conferred by a pro- moter element different from the ones driving expression in the other cell types.
Stomatal guard cells of higher plants play an important role in gas exchange and transpiration in the leaves of higher plants. Opening and closing of stomates are under the control of a large variety of plant endogenous and exogenous (environ- mental) factors, including light intensity, COn concentration, humidity, and hormones (Zeiger et al., 1987). Stomatal open- ing is caused by the accumulation of both inorganic and organic compounds in guard cells (for a recent review, see Tallman, 1992).
Thus far, no mutants have been described that are specifi- cally changed with respect to guard cell metabolism or signal transduction. We believe that the truncated AGPase S promoter from potato will be avaluable tool in creating transgenic plants that are altered in various biochemical pathways, specifically those in guard cells. These plants will allow us to test in vivo the importance of certain biochemical pathways or signal trans- duction steps for stomatal functioning.
METHODS
Plants, Bacterial Strains, and Growth Conditlons
Potato (Solanum tubemsum L.) cv Desiree was obtained through Saat- zucht Fritz Lange KG (Bad Schwartau,. Germany). Plants in tissue culture were maintained under a 16-hr lighV8-hr dark regime on Murashige and Skoog medium (Murashige and Skoog, 1962) containing 2% (whr) sucrose. Plants in the greenhouse were cultivated in soil or vermiculite under a regime of -16 hr of light and 8 hr of darkness. Plants were grown in individual pots (200 cm2, 15 cm deep) and were watered daily.
Escherichia coli DH5a (Bethesda Research Laboratories) was cul- tured using standard techniques (Sambmk et al., 1989). &tuba&rium tumefaciens C58C1 containing plasmid pGV2260 (Deblaere et al., 1985) was cultured in YEB medium'(Verv1iet et al., 1975).
Enzymes and Chemicals
DNA restriction and modification enzymes were purchased from Boehringer Mannheim and New England Biolabs (Beverly, MA).

610 The Plant Cell
5Bromo-4chloro-3-indolyl P-D-glucuronide was obtained from Biosynth AG (Staad, Switzerland). Other chemicals were purchased from Sigma or Merck.
DNA Manlpulations
DNA manipulations were performed essentially as described by Sambrook et al. (1989). DNA sequences were determined by the dideoxy method (Sanger et al., 1977) using supercoiled plasmids (Chen and Seeburg, 1985). Either commercial sequencing primers (Pharmacia) or specifically synthesized oligonucleotides (DNA Synthesizer 380A; Applied Biosystems, Foster City, CA) were used for sequencing with a T7 DNA polymerase sequencing kit (Pharmacia).
lsolatlon of the Potato ADP-Glucose Pyrophosphorylase Large Subunit Promoter
To isolate the ADP-glucose pyrophosphorylase large subunit (AGPase S) promoter, a genomic library prepared from the monohaploid potato line AM 80/5793 (Liu et al., 1991) was screened with a partia1 AGPase S cDNA (EcoRI insert of plasmid S25-1; Müller-Rober et al., 1990). Severa1 hybridizing phage clones were obtained. The >12-kb-long Sall- insert of clone gSF-6 was subcloned into pUC19. The resulting plas- mid SF-6 was analyzed by restriction analysis and DNA gel blot hybridization using an N200-bp-lOng 5' fragment of the AGPase S cDNA. A 5.2-kb Hincll subfragment strongly hybridizing to the probe was subcloned into pUC19, resulting in plasmid pH6-1 (see Figure 1). DNA sequences obtained for severa1 subfragments were compared with the full-length potato AGPase S cDNA S9-D. This cDNA was ob- tained from a tuber-specific cDNA library (provided by J. KoRmann, lnstitut für Genbiologische Forschung, Berlin, Germany) using the pre- viously isolated cDNA S25-1 (Müller-Rober et al., 1990) as a probe.
Construction of Promoter-gus Fusions
Binary plasmids containing chimeric AGPase-P-glucuronidase (gus) gene fusions were constructed as follows. In all cases, the binary vec- tor pBllOl.l (Jefferson et al., 1987) was used to obtain plasmids for the transformation of potato and tobacco plants.
AGPase S-gus: Plasmid pH6-1 (see Figure 1) was cleaved with Hincll, partially digested with Bglll, and blunt-ended by a fill-in reac- tion with T4 polymerase. The resulting 3.2-kb promoter fragment was cloned into the Smal site of pUCl9 (5' end oriented to the EcoRl site of the polylinker), resulting in plasmid pPSA. The AGPase S promoter was subsequently introduced as an EcoRl (blunt-ended)-BamHI frag- ment in front of the gos gene of the binary vector pBllOl.l (Jefferson et al., 1987), which was previously cut with Hindlll (blunt-ended) and BamHI. The resulting plasmid is shown in Figure 28.
To create SI-A1, plasmid pPSA was cut with Ncol, blunt-ended, and recut with BamHI. The resulting 2-kb promoter fragment was ligated to pBI101.1, cut with Hindlll, blunt-ended, and recut with BamHI.
To construct SI-A2, the 1.2-kb-long Hindlll-BamHI promoter frag- ment of plasmid pPSA was inserted into vector pBI101.1, which was previously cleaved with Hindlll and BamHI.
To create SI-A4, the 0.35-kb-long Hindlll-BamHI promoter fragment of plasmid pPSA was inserted into vector pBllOl.l, which was previ- ously cut with Hindlll and BamHI.
To construct Sl-A5, the 0.9-kb-long internal Hindlll promoter frag- ment of plasmid pPSA was inserted into vector pBI1O1.l, which was previously cleaved with Hindlll.
,
Creating SI-AI3 required an internal 0.7-kb promoter fragment to be deleted by cleaving plasmid pPsA with Clal and subsequent reli- gation of the remaining vector. The resulting plasmid pPSA-Al3 was cut with EcoRI, blunt-ended with T4 polymerase, and cut with BamHI. The 2.5-kb-long promoter fragment was subsequently cloned into pBI101.1, which was previouslycleaved with Hindlll, blunt-ended, and recut with BamHI.
To construct Si-A14, a 1.5-kb internal fragment of the AGPase pro- moter needed to be deleted. To this end, plasmid pPSA was cut with Ncol and Clal, blunt-ended, and religated. The 1.7-kb-long promoter fragment from the resulting plasmid pPSA-A14 was inserted as an EcoRl (blunt-ended)-BamHI fragment into pBI101.1, which was previously cleaved with Hindlll, blunt-ended, and recut with BamHI.
Plant Transformations
Binary vectors were used to transform potato (Solanum tuberosum cv Desiree) and tobacco (Nicotiana tabacum cv Samsun NN) by way of Agrobacterium C58CI:pGV2260, as described by Rocha-Sosa et al. (1989) and Rosahl et al. (1987).
Hlstochemical Localizatlon of GUS Activlty
Histochemical staining for GUS activity was performed using 5-bromc- 4chloro-3-indolyl P-D-glucuronide as chromogenic substrate (Jefferson et al., 1987). Freshly cut plant tissues were incubated in GUS staining solution (for 30 min to 24 hr at P C , depending on staining intensity) and rinsed with water before photography. Photosynthetic tissues were bleached with ethanol. Microscopic analysis was performed using an Axiophot microscope (Zeiss, Oberkochen, Germany).
Fluorometric GUS Assay
Transgenic lines showing strongest histochemical GUS staining in leaf guard cells were used for fluorometric GUS assays. Tissue samples were homogenized in extraction buffer (50 mM sodium phosphate buffer, pH 7.0, 10 mM B-mercaptoethanol, 10 mM EDTA, 0.1% SDS, 0.1% Triton X-100) in a microcentrifuge tube and centrifuged for 10 min at 13,000 rpm in a microcentrifuge (Biofuge A; Heraeus Instruments, Hanau, Germany). Aliquots of the supernatant were taken for enzyme assay using 4-methylumbelliferyl P-D-glucuronide as substrate (Jefferson, 1987) and for protein determination (Bradford, 1976). The product of the reaction (4-methylumbelliferone [4-MU]) was measured fluorometrically (SFM 25 Fluorescence Spectrophotometer; Kontron Instruments, Hamburg, Germany).
lodine Staining
To visualize starch granules, tissues were incubated for 2 min in Lugol's solution (5 g KI, 2 g l2 in 100 mL H20), washed in H20 for 30 sec, and subsequently viewed microscopically.
ACKNOWLEDGMENTS
The excellent workof our technician Gaby Nast isgreatlyacknowledged. We thank Joseph Bergstein and Antje Voigt for photographic work, Jessyca Dietze for tissue culture work, and our gardeners Birgit Burose,

ADP-Glucose Pyrophosphorylase Promoter from Potato 61 1
Birgit Hidde, and Regina Breitfeld for taking care of greenhouse plants. We thank Jens KoEmann for providing the potato tuber cDNA library and Nicholas Provart for critically reading the manuscript.
Received February 14, 1994; accepted March 17, 1994.
REFERENCES
Ainsworth, C., Tawls, M., and Clark, J. (1993). lsolation and analy- sis of a cDNA clone encoding the small subunit of ADP-glucose pyrophosphorylase from wheat. Plant Moi. Biol. 23, 23-33.
Anderson, J.M., Hnllo, J., Raymond, L., Oklta, T.W., Morell, M., and Preiss, J. (1989). The encoded primary sequence of a rice seed ADP-glucose pyrophosphorylase subunit and its homology to the bacterial enzyme. J. Biol. Chem. 264, 12238-12242.
Anderson, J.M., Larsen, R., Laudencia, D., Kim, W. T., Morrow, D., Oklta, T.W., and Prelss, J. (1991). Molecular characterization of the gene encoding a rice endosperm-specific ADPglucose pyrophosphorylase subunit and its developmental pattern of tran- scription. Gene 97, 199-205.
Bae, J.M., Giroux, M., and Hannah, L.C. (1990). Cloning and char- acterization of the brittle-2 gene of maize. Maydica 35, 317-322.
Bhave, M.R., Lawrence, S., Barton, C., and Hannah, L.C. (1990). ldentification and molecular characterization of shrunken-2 cDNA clones of maize. Plant Cell 2, 581-588.
Bradford, M.M. (1976). Rapid and quantitative method for quantita- tion of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248-252.
Chen, E.J., and Seeburg, RH. (1985). Supercoiling sequencing: A simple method for sequencing plasmid DNA. DNA 4, 165-170.
Deblaere, R., Bytebier, B., de Greve, H., Deboeck, F., Schell, J., Van Montagu, M., and Leemans, J. (1985). Efficient octopine Ti plasmid-derived vectors for Agrobacterium-mediated gene transfer to plants. Nucl. Acids Res. 13, 4777-4788.
du Jardin, P., and Berhln, A. (1991). lsolation and sequence analysis of a cDNA clone encoding a subunit of the ADP-glucose pyrophos- phorylase of potato tuber amyloplasts. Plant MOI. Biol. 16,349-351.
Gebhardt, C., Rltter, E., and Salamini, F. (1993). RFLP map of the potato. In Advances in Cellular and Molecular Biology of Plants, Vol. I: DNA-Based Markers in Plants, I.K. Vasil and R.L. Phillips, eds (Dordrecht, The Netherlands: Kluwer Academic Publishers), in prass.
Hsiao, T.C. (1976). Stomatal ion transport. In Encyclopedia of Plant Physiology, New Series, Vol. 2, U. Lijttge and M.G. Pitman, eds (Ber- lin: Springer-Verlag), pp. 195-221.
Hylton, C., and Smlth, A.M. (1992). The rb mutation of peas causes structural and regulatory changes in ADP glucose pyrophosphory- lase from developing embryos. Plant Physiol. 99, 1626-1634.
Jefferson, R.A. (1987). Assaying chimeric genes in plants: The GUS gene fusion system. Plant MOI. Biol. Rep. 5, 387-405.
Jefferson, R.A., Kavanagh, T.A., and Bevan, M.W. (1987). GUS fu- sions: P-Glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 6, 3901-3907.
Kell, M., SBnchez-Serrano, J., Schell, J., and Willmltzer, L. (1990). Localization of elements important for the wound-inducible expres- sion of a chimeric potato proteinase inhibitor II-CAT gene in transgenic tobacco plants. Plant Cell 2, 61-70.
Kleukowski, L.A., Vllland, P., Unneborg, A., Olsen, O.A., and Liithi, E. (1991). Plant ADP-glucose pyrophosphorylase: Recent ad- vances and biotechnological perspectives. Z. Naturforsch. 46c,
Kleukowski, L.A., Vllland, R, Liithl, E., Olsen, O.A., and Prelss, J. (1993). lnsensitivity of barley endosperm ADP-glucose pyro- phosphorylase to 3-phosphoglycerate and orthophosphate regulation. Plant Physiol. 101, 179-186.
Krishnan, H.B., Reeves, C.D., and Okita, T.W. (1986). ADP glucose pyrophosphorylase is encoded bydifferent mRNA transcripts in leaf and endosperm of cereal. Plant Physiol. 81, 642-645.
Lln, T.-P., Caspar, T., Somewille, C., and Preiss, J. (1988a). Isola- tion and characterization of a starchless mutant of Arabidopsis thaliana (L.) Heynh. lacking ADPglucose pyrophosphorylase activ- ity. Plant Physiol. 86, 1131-1135.
Lin, T.-P., Caspar, T., Somerville, CR., and Preiss, J. (1988b). A starch deficient mutant of Arabidopsis thaliana with low ADPglucose pyrophosphorylase activity lacks one of the two subunits of the en- zyme. Plant Physiol. 88, 1175-1181.
Liu, X.Y., Rocha-Sosa, M., Hummel, S., Wlllmitzer, L., and Frommer, W.B. (1991). A detailed study of the regulation and evolution of the two classes of patatin genes in Solanum tuberosum L. Plant MOI. Biol. 17, 1139-1154.
Morell, M.K., Bloom, M., Knowles, V., and Prelss, J. (1987). Subunit structure of spinach leaf ADPglucose pyrophosphorylase. Plant Phys- iol. 85, 182-187.
Miiller-Rober, B., KoBmann, J., Hannah, L.C., Wlllmitzer, L., and Sonnewald, U. (1990). One of two different ADP-glucose pyrophosphorylase genes from potato responds strongly to elevated levels of sucrose. MOI. Gen. Genet. 224, 136-146.
Müller-Rober, B., Sonnewald, U., and Wlllmitzer, L. (1992). Inhibi- tion of the ADP-glucose pyrophosphorylase in transgenic potatoes leads to sugar-storing tubers and influences tuber formation and expression of tuberstorage protein genes. EMBO J. 11,1229-1238.
Murashige, T., and SWg, F. (1962). A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant. 15, 493-497.
Nakamura, Y., and Kawaguchi, K. (1992). Multiple forms of ADPgIu- cose pyrophosphorylase of rice endosperm. Physiol. Plant. 84,
Nakata, P.A., Greene, T.W., Anderson, J.M., Smith-Whlte, B.J., Oklta, T.W., and Prelss, J. (1991). Comparison of the primary sequences of two potato tuber ADP-glucose pyrophosphorylase subunits. Plant MOI. Biol. 17, 1089-1093.
Neuhaus, H.E., and Stitt, M. (1990). Control analysis of photosyn- thate partitioning. lmpact of reduced activity of ADP-glucose pyrophosphorylase or plastid phosphoglucomutase on the fluxes of starch and sucrose in Arabidopsis thaliana (L.) Heynh. Planta 182, 445-454.
Oklta, T.W., Nakata, P.A., Anderson, J.M., Sowokinos, J., Morell, M., and Preiss, J. (1990). The subunit structure of potato tuber ADP- glucose pyrophosphorylase. Plant Physiol. 93, 785-790.
Olive, M.R., Ellis, R.J., and Schuch, W.W. (1989). lsolation and nucleotide sequences of cDNA clones encoding ADP-glucose pyrophosphorylase polypeptides from wheat leaf and endosperm. Plant Moi. Biol. 12, 525-538.
Outlaw, W.H., Jr., and Tarczynski, M.C. (1984). Guard cell starch bio- synthesis regulated by effectors of ADP-glucose pyrophosphorylase. Plant Physiol. 74, 424-429.
605-612.
336-342.

612 The Plant Cell
Peach, C., and Velten, J. (1991). Transgene expression variability (po- sition effect) of CAT and GUS reporter genes driven by linked divergent T-DNA promoters. Plant Moi. Biol. 17, 49-60.
Pettersson, G., and Ryde-Pettersson, U. (1989). Metabolic control- ling the rate of starch synthesis in the chloroplast of C3 plants. Eur. J. Biochem. 179, 169-172.
Preiss, J. (1988). Biosynthesis of starch and its regulation. In The Bio- chemistryof Plants, Vol. 14, J. Preiss, ed (New York: Academic Press),
Preiss, J. (1991). Biology and molecular biology of starch synthesis and its regulation. In Oxford Surveys of Plant Molecular and Cell Biology, Vol. 7,B.J. Miflin and H.F. Miflin, eds (Oxford: Oxford Univer- sity Press), pp. 59-114.
Reeves, C.D., Krishnan, H.B., and Okita, T.W. (1986). Gene expres- sion in developing wheat endosperm. Plant Physiol. 82, 34-40.
Robinson, N.L., Zeiger, E., and Preiss, J. (1983). Regulation of ADP- glucose synthesis in guard cells of Commelina communis. Plant Physiol. 73, 862-864.
Rocha-Sosa, M., Sonnewald, U., Frommer, W., Stratmann, M., Schell, J., and WillmRzer. L. (1989). Bdh developmental and meta- bolic signals activate the promoter of a class l patatin gene. EMBO
Rosahl, S., Schell, J., and Willmitzer, L. (1987). Expression of a tuber- specific storage protein in transgenic tobacco plants: Demonstra- tion of an esterase activity. EMBO J. 6, 1155-1159.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). Molecular Clon- ing: A Laboratory Manual, 2nd ed. (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press).
Sanders, P.R., Winter, J.A., Barnason, A.R., Rogers, S.G., and Fraley, R.T. (1987). Comparison of cauliflower mosaic virus 355 and nopaline synthase promoters in transgenic plants. Nucl. Acids Res. 15, 1543-1558.
pp. 181-254.
J. 8, 23-29.
Sanger, F., Nicklen, S., and Coulson, A.R. (1977). DNAsequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA. 74,
Smlth-White, B.J., and Preiss, J. (1992). Comparison of proteins of ADP-glucose pyrophosphorylase from diverse sources. J. MOI. Evol.
Stark, D.M., Timmermann, K.P., Barry, G.F., Preiss, J., and Klshore, G.M. (1992). Regulation of the amount of starch in plant tissues by ADP glucose pyrophosphorylase. Science 258, 287-292.
Tallman, G. (1992). The chemiosmotic model of stomatal opening revisited. Crit. Rev. Plant Sci. 11, 35-57.
Terryn, N., Arias, M.B., Engler, G., Ti&, C., Vlllarroel, R., Van Montagu, M., and Inz6, D. (1993). rhal, a gene encoding a small GTP binding protein from Arabidopsis, is expressed primarily in de- veloping guard cells. Plant Cell 5, 1761-1769.
Tsai, C.Y., and Nelson, O.E. (1966). Starch-deficient maize mutant lacking adenosine diphosphate glucose pyrophosphorylase activ- ity. Science 151, 341-343.
Venrliet, G., Holsters, M., Teuchy, H., Van Montagu, M., and Schell, J. (1975). Characterization of different plaque-forming and defec- tive temperate phages in Agrobacterium strains. J. Gen. Virol. 26,
Villand, P., Aalen, R., Olsen, O.A., Wthi, E., Unneborg, A., and Kleczkowski, L.A. (1992a). PCR amplification and sequences of cDNA clones for the small and large subunits of ADP-glucose pyrophosphorylase from barley tissues. Plant MOI. Biol. 19,381-389.
Villand, P., Olsen, O.A., Kilian, A., and Kleczkowski, L.A. (1992b). ADP-glucose pyrophosphorylase large subunit cDNA from barley endosperm. Plant Physiol. 100, 1617-1618.
Weising, K., Schell, J., and Kahl, O. (1988). Foreign genes in plants: Transfer, structure, expression and applications. Annu. Rev. Genet.
Zeiger, E., Farquhar, G.D., and Cowan, IA., eds (1987). Stomatal
5463-5467.
34, 449-464.
33-40.
22, 421-477.
Function. (Stanford, CA: Stanford University Press).

DOI 10.1105/tpc.6.5.601 1994;6;601-612Plant Cell
B Müller-Röber, U La Cognata, U Sonnewald and L Willmitzerguard cell-selective expression in transgenic plants.
A truncated version of an ADP-glucose pyrophosphorylase promoter from potato specifies
This information is current as of September 12, 2018
Permissions 98X
https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw15322
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists