a zebrafish transgenic model of ewing’s sarcoma reveals...
TRANSCRIPT

INTRODUCTIONChromosomal translocations that create fusion oncogenes havelong been recognized as a cause of human cancers such as leukemiasand sarcomas (Mitelman et al., 2007), and recently have beendescribed in prostate (Tomlins et al., 2005; Brenner and Chinnaiyan,2009) and lung cancer (Soda et al., 2007). Drugs that targetoncogenic fusions have also led to dramatic improvements in thetreatment of certain cancers (Druker, 2009). In Ewing’s sarcoma, amalignant bone tumor that most commonly occurs in adolescentsand young adults, the discovery of a characteristic chromosomaltranslocation t(11;22)(q24;q12) (Delattre et al., 1992) marked aturning point in the understanding of the biology of the disease.In the majority of Ewing’s tumors, the N-terminal portion of EWSbecomes fused to the C-terminal portion of FLI-1, which includesan ETS family DNA binding domain (Delattre et al., 1992). In some(~15%) Ewing’s tumors, alternative chromosomal translocationscreate fusions between EWS and other ETS family transcriptionfactors. These alternative fusions suggest that EWS mediates acritical change in ETS family transcription factors that allows them
to function aberrantly and promote tumor development, possiblythrough incorrect regulation of target gene expression.
Ewing’s sarcoma is classified as a small-round-blue-cell tumor(SRBCT), a group that also includes medulloblastoma,undifferentiated neuroblastoma, alveolar rhabdomyosarcoma andsome types of leukemia. The histology of tumors in this family isthat of poorly differentiated cells, scant cytoplasm and round nucleithat stain darkly with hematoxylin. Ewing’s sarcomas are frequentlyassociated with bone; however, they can also be found in othertissues, obscuring the identification of a definite cell of origin.
The discovery of EWS-ETS translocations in Ewing’s sarcomafamily tumors (ESFTs) creates an opportunity for the developmentof targeted therapy of Ewing’s sarcoma, either through directinhibition of EWS-FLI1, or through the discovery of criticaldownstream effectors that might themselves be candidates fortargeted therapy (Uren and Toretsky, 2005; Erkizan et al., 2009).EWS-FLI1 is believed to function primarily as an aberranttranscription factor. Although microarray studies have identifieda large number of potential EWS-FLI1 targets, they have alsorevealed a few overlapping genes that are consistently regulated byEWS-FLI1 across these varying cellular backgrounds (Ohali et al.,2004; Staege et al., 2004; Baird et al., 2005; Smith et al., 2006;Hancock and Lessnick, 2008). These studies have also indicatedthat neural genes are frequently induced by EWS-FLI1 (Hu-Lieskovan et al., 2005; Siligan et al., 2005). In fact, a neuronallyexpressed homeobox transcription factor, NKX2.2, is required forEWS-FLI1-induced transformation and tumorigenesis in a mousexenograft model (Smith et al., 2006).
Cell culture experiments have demonstrated that heterologousexpression of EWS-FLI1 is toxic to many cell types by inducinggrowth arrest or apoptosis (Deneen and Denny, 2001; Lessnick etal., 2002). These observations, along with the relative lack ofdifferentiation in Ewing’s tumor cells, have led to the hypothesisthat only a subset of undifferentiated precursor cell is capable of
Disease Models & Mechanisms 95
Disease Models & Mechanisms 5, 95-106 (2012) doi:10.1242/dmm.007401
1Department of Pediatrics, University of Texas Southwestern Medical Center, 5323 Harry Hines Blvd, Dallas, TX 75390-8534, USA2Department of Molecular Biology, University of Texas Southwestern MedicalCenter, 5323 Harry Hines Blvd, Dallas, TX 75390-8534, USA3Department of Pathology, University of Texas Southwestern Medical Center, 5323 Harry Hines Blvd, Dallas, TX 75390-8534, USA4Department of Internal Medicine, University of Texas Southwestern MedicalCenter, 5323 Harry Hines Blvd, Dallas, TX 75390-8534, USA*Author for correspondence ([email protected])
Received 15 December 2010; Accepted 29 August 2011
© 2012. Published by The Company of Biologists LtdThis is an Open Access article distributed under the terms of the Creative Commons AttributionNon-Commercial Share Alike License (http://creativecommons.org/licenses/by-nc-sa/3.0), whichpermits unrestricted non-commercial use, distribution and reproduction in any medium providedthat the original work is properly cited and all further distributions of the work or adaptation aresubject to the same Creative Commons License terms.
SUMMARY
Ewing’s sarcoma, a malignant bone tumor of children and young adults, is a member of the small-round-blue-cell tumor family. Ewing’s sarcomafamily tumors (ESFTs), which include peripheral primitive neuroectodermal tumors (PNETs), are characterized by chromosomal translocations thatgenerate fusions between the EWS gene and ETS-family transcription factors, most commonly FLI1. The EWS-FLI1 fusion oncoprotein represents anattractive therapeutic target for treatment of Ewing’s sarcoma. The cell of origin of ESFT and the molecular mechanisms by which EWS-FLI1 mediatestumorigenesis remain unknown, and few animal models of Ewing’s sarcoma exist. Here, we report the use of zebrafish as a vertebrate model ofEWS-FLI1 function and tumorigenesis. Mosaic expression of the human EWS-FLI1 fusion protein in zebrafish caused the development of tumorswith histology strongly resembling that of human Ewing’s sarcoma. The incidence of tumors increased in a p53 mutant background, suggestingthat the p53 pathway suppresses EWS-FLI1-driven tumorigenesis. Gene expression profiling of the zebrafish tumors defined a set of genes thatmight be regulated by EWS-FLI1, including the zebrafish ortholog of a crucial EWS-FLI1 target gene in humans. Stable zebrafish transgenic linesexpressing EWS-FLI1 under the control of the heat-shock promoter exhibit altered embryonic development and defective convergence and extension,suggesting that EWS-FLI1 interacts with conserved developmental pathways. These results indicate that functional targets of EWS-FLI1 that mediatetumorigenesis are conserved from zebrafish to human and provide a novel context in which to study the function of this fusion oncogene.
A zebrafish transgenic model of Ewing’s sarcoma revealsconserved mediators of EWS-FLI1 tumorigenesisStefanie W. Leacock1,2, Audrey N. Basse1,2, Garvin L. Chandler1,2, Anne M. Kirk1,2, Dinesh Rakheja3 and James F. Amatruda1,2,4,*
RESEARCH ARTICLED
iseas
e M
odel
s & M
echa
nism
s
DM
M

responding to EWS-FLI1 to generate Ewing’s sarcoma. Mousemesenchymal progenitor cells are one type of cell that toleratesEWS-FLI1 expression and generate Ewing’s-like tumors whentransplanted into mice (Riggi et al., 2005). Although this result isa promising step toward understanding the possible origins ofEwing’s sarcoma, mesenchymal progenitor cells remain relativelyuncharacterized at a molecular level, therefore the cellular contextrequired for EWS-FLI1 to exert its function remains unknown.Thus, the molecular characteristics that are required for EWS-FLI1responsiveness, as well as the downstream mechanisms by whichEWS-FLI1 gives rise to tumors, remain to be elucidated.
Several mouse models of EWS-FLI1 transgenic expression havebeen reported. Expression of EWS-FLI1 in hematopoietic tissuesin an Mx1-Cre conditional mouse model induced myeloid orerythroid leukemia in transgenic mice (Torchia et al., 2007).Recently, EWS-FLI1 was conditionally expressed in mice under thecontrol of a primitive mesenchymal cell promoter, resulting in limbdefects and accelerated development of undifferentiated sarcomawhen introduced into the p53 mutant background (Lin et al., 2008).Further studies in which EWS-FLI1 is expressed in a different celltype or at a specific developmental stage are required to more fullyrecapitulate Ewing’s sarcoma in the mouse.
Models of several types of cancers have recently been developedin the zebrafish Danio rerio, including melanoma, leukemia andrhabdomyosarcoma (Amatruda and Patton, 2008). Here, we reportthat expression of human EWS-FLI1 in zebrafish causes tumorsstrongly resembling human Ewing’s sarcoma. We identify an EWS-FLI1 gene expression signature in the zebrafish tumors, whichreveals striking conservation of key EWS-FLI1 targets between fishand human Ewing’s tumors. Additionally, we show that EWS-FLI1exerts powerful effects on embryonic development, indicating thatEWS-FLI1 functions through conserved developmental signalingpathways.
RESULTSExpression of a human EWS-FLI1 transgene causes small-round-blue-cell tumors in zebrafishThe molecular basis of Ewing’s sarcoma is unknown despite thestrong association with the EWS-FLI1 oncogene. To develop ananimal model in which to study the function of EWS-FLI1, wecreated transgenic zebrafish carrying the human EWS-FLI1 fusionprotein, using the Tol2 transposon system (Kawakami et al., 2004).Three different transgenes were generated, two of which use thezebrafish heat-shock (hsp70) promoter, and another using the -actin promoter, to drive expression of a FLAG-tagged human EWS-FLI1 (Fig. 1A).
From over 300 wild-type embryos injected with the threedifferent EWS-FLI1 transgenes and raised to adulthood, weidentified two fish with tumors. One of the fish was identified bythe appearance of outgrowths covering the head at 15 months ofage. Sectioning and staining revealed the presence of diffuse tumorcells covering the surface of the head, as well as within the head,gills, and body cavity (Table 1; Fig. 1C,D). This histology resemblesthat of existing zebrafish models of leukemia (Langenau et al., 2005;Chen et al., 2007). The other tumor identified in a wild-type fishwas a solid tumor that arose in proximity to the eye at 12 monthsof age (Table 1; Fig. 1E,F). Both of these histologies are consistentwith the SRBCT morphological category. These tumor types rarely
occur spontaneously in zebrafish (Amsterdam et al., 2009). Theidentification of zebrafish tumors that histologically resembleEwing’s family tumors was encouraging, but the low incidence oftumors precluded the analysis of a large number of tumors.
Because we observed a low incidence of tumors, we suspectedthat cellular tumor suppressive mechanisms might be inhibitingtumor development in the wild-type zebrafish. We identified thep53 pathway as a likely candidate for such tumor suppression.Mutations in tp53 occur in a subset of primary Ewing’s tumors(Hamelin et al., 1994; Huang et al., 2005) and research suggeststhat EWS-FLI1 can induce a p53-dependent growth arrest (Lessnicket al., 2002). Additionally, mutation of tp53 has been shown to bea sensitized background for the development of other tumors inthe zebrafish, including rhabdomyosarcoma and melanoma models(Patton et al., 2005; Langenau et al., 2007), as well as in a mousemodel of EWS-FLI1-driven tumor development (Lin et al., 2008).Zebrafish with a mutation in tp53 are predisposed to developmalignant peripheral nerve sheath tumors (MPNSTs) (Berghmanset al., 2005), composed of spindle cells with abundant cytoplasm.Based on the histology of the two tumors we identified in wild-type EWS-FLI1 transgenic fish, we anticipated that the histologyof a tumor driven by EWS-FLI1 would be distinct from this tumortype. Therefore, we injected the EWS-FLI1 transposons into p53-deficient zebrafish embryos and raised these fish to adulthood.
From 6 to 19 months of age, we identified 48 tumors fromapproximately 150 tp53 mutant zebrafish injected with one of thethree EWS-FLI1 transposons (Table 1). Of these, 33 were classifiedbased on their histology as malignant peripheral nerve sheathtumors (MPNSTs). Diffuse, leukemia-like tumors were identifiedin four fish (Fig. 1G). The remaining ten tumors exhibited SRBCThistology, but were not diffuse and instead presented as solid tumormasses (Fig. 1H). The identification of both solid and diffuse tumorssuggests that EWS-FLI1 can induce the formation of at least twotumor types in the zebrafish. Both MPNSTs and SRBCTs wereidentified at similar ages of onset (average age at sacrifice 13.1months each). The small round blue cell morphology was notidentified in any tumors occurring in uninjected tp53 mutants.Therefore, this histological phenotype is associated with thepresence of EWS-FLI1 and is consistent with the pathology ofhuman Ewing’s sarcoma.
To address the similarity of the diffuse tumors to leukemias, weperformed quantitative RT-PCR for two genes known to be highlyexpressed in other zebrafish models of leukemia, rag2 and lck(Langenau et al., 2005). Both genes were more highly expressed inRNA from samples classified as leukemia-like tumors(supplementary material Fig. S2). The tumors classified as zebrafishsolid SRBCTs did show slightly increased expression of rag2 andlck compared with levels in the wild type but were substantiallylower than expression in the leukemia-like tumors. Thus, theexpression of two conserved markers for leukemia is consistent withthe histological characterization of the leukemia-like tumorsidentified in zebrafish.
SRBCTs are transplantableTo show that the zebrafish Ewing’s sarcomas have malignantpotential, we performed transplantation experiments. Followingdissection and manual dissociation of two SRBCTs derived from tp53mutants, approximately 1 million cells were injected intraperitoneally
dmm.biologists.org96
Zebrafish model of Ewing’s sarcomaRESEARCH ARTICLED
iseas
e M
odel
s & M
echa
nism
s
DM
M

into irradiated wild-type adult zebrafish. Recently, a number oftransplantation experiments have been performed in zebrafish tumormodels with transplanted tumors typically developing within 2 to 8weeks post-transplant [reviewed by Taylor and Zon (Taylor and Zon,2009)]. Both of our primary transplants formed new tumors within3-5 weeks, consistent with these previously reported results. Thesetumors were then subjected to further rounds of transplantation(supplementary material Table S1). Even when the number ofinjected tumor cells was reduced more than 10 times (see numberof cells transplanted in 1° vs 4° or 8° transplant in supplementarymaterial Table S1), the tumors continued to regenerate. One of the
two secondary transplants continued to generate new tumors in eachhost and was serially transplanted nine times. These successfultransplantation experiments indicate that the tumor cells continueto proliferate. Although one tumor (D85) appeared to havediminished capacity to regenerate with subsequent transplants, thesecond tumor (D86) maintained an average tumor development timeof 17 days over eight rounds of transplantation. Both primarytumors were initially categorized as solid SRBCTs based on theirhistology. The subsequent tumors continued to exhibit similarhistology, although some tumor variation was observed (Fig. 2). Afterseveral rounds of transplantation, tumors appeared to have increased
Disease Models & Mechanisms 97
Zebrafish model of Ewing’s sarcoma RESEARCH ARTICLE
Fig. 1. EWS-FLI1 induces SRBCTs in zebrafish.(A)Schematic of Tol2 transposons including thehsp70 or the -actin promoter, FLAG-tag (blackrectangle), human EWS-FLI1 coding sequence, IRES-GFP sequence (gray rectangle), flanked by Tol2recombination sites (triangles). (B-H)H&E staining ofsagittal sections of adult zebrafish. (B)Control adultzebrafish. Wild-type zebrafish injected with EWS-FLI1 developed an invasive leukemia like-tumor (Cmagnified in D) or a solid small round blue celltumor of the eye (arrow in E) (E magnified in F). tp53mutants injected with EWS-FLI1 transposons alsodevelop leukemia (G) or solid tumors (H) with similarhistology. Scale bars: 200mm (B,C,E), 50mm (D,F-H).
Dise
ase
Mod
els &
Mec
hani
sms
D
MM

cytoplasm in some regions (see 6° and 8° in Fig. 2B). Both primarytumors were derived from tp53 mutant fish, but formed new tumorsin the recipient wild-type hosts. From these transplantationexperiments, we conclude that, although p53 might serve as a barrierto transformation by EWS-FLI1, this effect is probably autonomousto the tumor cell and not dependent on the p53 status of thesurrounding normal tissue.
Zebrafish SRBCTs express the EWS-FLI1 transgeneTo determine whether the tumors express the EWS-FLI1 mRNA,we tested for the presence of EWS-FLI1 using RT-PCR. EWS-FLI1mRNA was present at detectable levels in four of six of the SRBCTprimary or transplanted tumors analyzed (supplementary materialFig. S1 and data not shown). By contrast, only one of ten MPNSTtumors exhibited EWS-FLI1 expression (supplementary materialFig. S1 and data not shown). We conclude that the SRBCTs werecaused by the presence of the transgene, but that the MPNSTs wereattributable to the mutation in tp53. Those tumors for which wewere unable to detect EWS-FLI1 mRNA might have undetectablelevels of expression, normal tissue co-isolated with the tumor tissue
that obscures detection or might have expressed EWS-FLI1 duringtumor initiation but lost expression in later stages of tumor growth.Together, the pathology of these tumors and expression of EWS-FLI1 mRNA in the majority of tumor samples indicate that thesetumors are a model of Ewing’s sarcoma in the zebrafish.
Gene expression profiling of zebrafish SRBCTsThe histology of the zebrafish tumors strongly resembles that ofEwing’s sarcoma and other human SRBCTs. Based on the modelthat EWS-FLI1 acts as a transcription factor, we hypothesizedthat this similarity in cellular morphology was caused by changesin gene expression driven by EWS-FLI1. We isolated tissue fromthree solid SRBCTs and hybridized the RNA to Affymetrixzebrafish gene expression arrays. The most highly expressed genesin these zebrafish tumors were ribosomal proteins andcomponents of the electron transport chain, consistent with ageneral requirement for protein translation and energy productionin proliferating cells.
Using a mutation in the conserved tumor suppressor p53 as asensitized genetic background that is likely to affect the expression
dmm.biologists.org98
Zebrafish model of Ewing’s sarcomaRESEARCH ARTICLE
Table 1. Tumor spectrum of EWS-FLI1 transgenic zebrafish
SRBCT
Transgene MPNST Solid tumor Leukemia-like Other n*
p53+/+ hsp70:EWS-FLI1 0 0 0 0 100
hsp70:EWS-FLI1:IRES-GFP 0 1 0 0 120
-actin:EWS-FLI1:IRES-GFP 0 0 1 0 120
p53–/– hsp70:EWS-FLI1 10 4 1 0 50
hsp70:EWS-FLI1:IRES-GFP 14 2 2 0 70
-actin:EWS-FLI1:IRES-GFP 11 2 1 1 60
*Approximate number of injected embryos that were raised to adulthood and monitored for tumor development.
Fig. 2. H&E staining of selected seriallytransplanted tumors originating from twodifferent zebrafish SRBCTs. (A) Tumor D85; (B)tumor D86 (see supplementary material Table S1).Numbers indicate serial transplants (primary,secondary, tertiary etc.).D
iseas
e M
odel
s & M
echa
nism
s
DM
M

of many genes that contribute to tumor initiation or development.Although the precise relationship between EWS-FLI1 and p53remains unknown, we wanted to eliminate those genes withexpression affected by the loss of p53. Therefore, to determine thosegenes that might be regulated more specifically by EWS-FLI1 andnot generally overexpressed in tumors, we compared the geneexpression profile with three of the MPNSTs that occurred in thetp53 mutants.
To examine total variation within these samples, we determinedthe correlation coefficients among the biological replicates(supplementary material Table S2). The MPNST samples were wellcorrelated (mean R0.95). The SRBCT samples had a lower averagecorrelation (mean R0.90), suggesting more variation amongsamples. Increased variation might be expected from mosaicintegration of the transgene and differences in the tissue of origin,because two tumors presented in the abdominal cavity and onetumor presented in the eye.
Comparing these two tumor types revealed 421 probes that weredifferentially expressed between the SRBCTs and the malignantperipheral nerve sheath tumors (Fig. 3A,B) (supplementary materialTable S3). These probes were annotated to 376 zebrafish genes,which we will refer to as the zebrafish SRBCT up- anddownregulated genes. The majority (243 genes) were
downregulated in SRBCTs, suggesting repression by EWS-FLI1.Although EWS-FLI1 is predicted to be a transcriptional activator,our finding of a larger proportion of genes with reduced expressionin EWS-FLI1-driven tumors is in agreement with gene expressionstudies in human and mouse systems. These downregulated genesprobably reflect indirect effects on gene expression. In fact, theSRBCT upregulated gene set is significantly enriched for genes withpredicted roles in transcription: at least 17 genes involved in theregulation of transcription (2.44-fold enrichment, P0.0026) arecontained within this set. Therefore, many of the gene expressionchanges, particularly those genes that are repressed, might beindirect targets of EWS-FLI1 through these transcription factors.
Although a specific cell of origin of Ewing’s sarcoma has not yetbeen identified, multiple studies have found that Ewing’s sarcomacells often express genes associated with neural development(Staege et al., 2004; Hu-Lieskovan et al., 2005; Siligan et al., 2005).Indeed, the zebrafish SRBCT upregulated gene list is enriched forgenes involved in nervous system development and neurogenesis(3.87-fold, P0.0024), including nkx2.2a, olig2, sox3 and asc1a/b(supplementary material Table S3). We performed quantitative RT-PCR to validate higher expression of nkx2.2 and olig2 in thezebrafish SRBCTs (Fig. 3C). The downregulated gene list does notexhibit enrichment in this category, indicating that a neural-associated gene expression profile is correlated with the presenceof EWS-FLI1.
To compare these zebrafish tumors to human studies of EWS-FLI1-regulated gene expression, the differentially expressedzebrafish genes were mapped to their human homologs. Thezebrafish probes were mapped to 97 human homologs (upregulatedin SRBCTs) and 162 human homologs (downregulated). Tocompare these genes with the combined results from severalhuman studies, we used a gene set generated by a meta-analysis ofmultiple microarray experiments that identified a core EWS-FLI1gene expression signature of 503 EWS-FLI1 upregulated genes and293 EWS-FLI1 downregulated genes (Hancock and Lessnick,2008). Comparing the genes from the human upregulated genesignature with the zebrafish SRBCT-enriched genes revealed anoverlap of six genes: NKX2-2, MYC, MAPT, SALL2, PADI2 andPOU3F1. The number of common genes reflects a slight, althoughnot statistically significant, overlap (1.7-fold) between the two genelists. However, the identity of the overlapping genes suggests thatthey are biologically relevant. NKX2-2 is one of very few genes thathave been shown to be a critically important downstream targetof EWS-FLI1 in tumorigenesis (Smith et al., 2006), and mediatestranscriptional repression downstream of EWS-FLI1 (Owen et al.,2008). The oncogene MYC has also been previously identified tobe upregulated by EWS-FLI1 (Dauphinot et al., 2001; Zwerner etal., 2008). The microtubule associated protein tau (MAPT) andNKX2-2 were in a group of only ten genes identified in an analysiscomparing Ewing’s family tumors and cell lines with peripheralblood (Cheung et al., 2007).
Comparing gene lists directly might fail to identify those geneswith expression that does not meet a certain criteria in someexperiments, but do exhibit the same pattern of change in geneexpression. Published analyses of expression profiling of EWS-FLI1models have produced gene sets with very limited overlap. Analternative way to compare expression profiling experiments is touse gene set enrichment analysis (GSEA), which provides a more
Disease Models & Mechanisms 99
Zebrafish model of Ewing’s sarcoma RESEARCH ARTICLE
Fig. 3. Gene expression in zebrafish SRBCTs is distinct from MPNSTs andsimilar to human EWS-FLI1 gene expression data. (A)Histology ofrepresentative zebrafish MPNST (left) and SRBCT (right) that were used formicroarray analysis. Scale bars: 50mm. (B)Heat map of 421 probes thatexhibited differential expression between MPNSTs and SRBCTs. Yellow, highexpression; blue; low expression. (C)Quantitative RT-PCR of selected SRBCTupregulated genes. (D,E) GSEA using human homologs of zebrafish genesshown in B compared with human gene expression data from (D) stableknockdown of EWS-FLI1 (red, higher expression in A673 Ewing’s cell line; blue,higher expression following EWS-FLI1 knockdown in A673 cells) or (E)inducible expression of EWS-FLI1 in a bone-marrow-derived stromal cell line(red, higher expression in EWS-FLI1 expressing cells; blue, higher expression incontrol cells).
Dise
ase
Mod
els &
Mec
hani
sms
D
MM
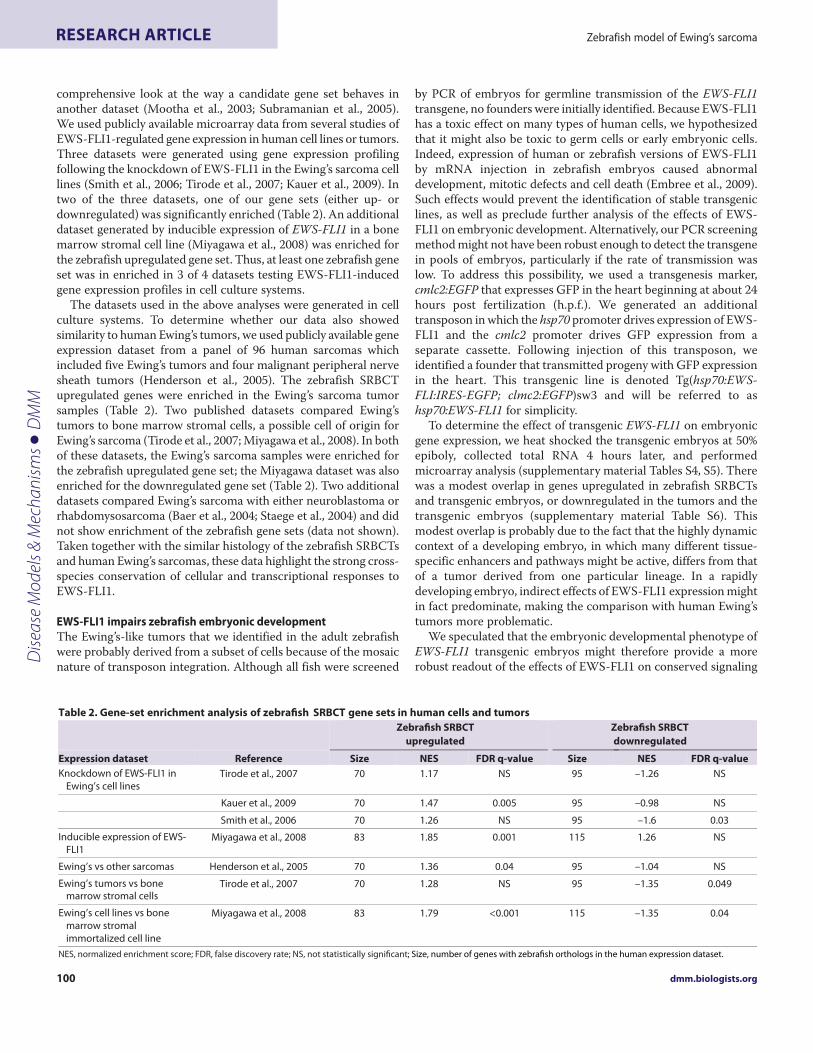
comprehensive look at the way a candidate gene set behaves inanother dataset (Mootha et al., 2003; Subramanian et al., 2005).We used publicly available microarray data from several studies ofEWS-FLI1-regulated gene expression in human cell lines or tumors.Three datasets were generated using gene expression profilingfollowing the knockdown of EWS-FLI1 in the Ewing’s sarcoma celllines (Smith et al., 2006; Tirode et al., 2007; Kauer et al., 2009). Intwo of the three datasets, one of our gene sets (either up- ordownregulated) was significantly enriched (Table 2). An additionaldataset generated by inducible expression of EWS-FLI1 in a bonemarrow stromal cell line (Miyagawa et al., 2008) was enriched forthe zebrafish upregulated gene set. Thus, at least one zebrafish geneset was in enriched in 3 of 4 datasets testing EWS-FLI1-inducedgene expression profiles in cell culture systems.
The datasets used in the above analyses were generated in cellculture systems. To determine whether our data also showedsimilarity to human Ewing’s tumors, we used publicly available geneexpression dataset from a panel of 96 human sarcomas whichincluded five Ewing’s tumors and four malignant peripheral nervesheath tumors (Henderson et al., 2005). The zebrafish SRBCTupregulated genes were enriched in the Ewing’s sarcoma tumorsamples (Table 2). Two published datasets compared Ewing’stumors to bone marrow stromal cells, a possible cell of origin forEwing’s sarcoma (Tirode et al., 2007; Miyagawa et al., 2008). In bothof these datasets, the Ewing’s sarcoma samples were enriched forthe zebrafish upregulated gene set; the Miyagawa dataset was alsoenriched for the downregulated gene set (Table 2). Two additionaldatasets compared Ewing’s sarcoma with either neuroblastoma orrhabdomysosarcoma (Baer et al., 2004; Staege et al., 2004) and didnot show enrichment of the zebrafish gene sets (data not shown).Taken together with the similar histology of the zebrafish SRBCTsand human Ewing’s sarcomas, these data highlight the strong cross-species conservation of cellular and transcriptional responses toEWS-FLI1.
EWS-FLI1 impairs zebrafish embryonic developmentThe Ewing’s-like tumors that we identified in the adult zebrafishwere probably derived from a subset of cells because of the mosaicnature of transposon integration. Although all fish were screened
by PCR of embryos for germline transmission of the EWS-FLI1transgene, no founders were initially identified. Because EWS-FLI1has a toxic effect on many types of human cells, we hypothesizedthat it might also be toxic to germ cells or early embryonic cells.Indeed, expression of human or zebrafish versions of EWS-FLI1by mRNA injection in zebrafish embryos caused abnormaldevelopment, mitotic defects and cell death (Embree et al., 2009).Such effects would prevent the identification of stable transgeniclines, as well as preclude further analysis of the effects of EWS-FLI1 on embryonic development. Alternatively, our PCR screeningmethod might not have been robust enough to detect the transgenein pools of embryos, particularly if the rate of transmission waslow. To address this possibility, we used a transgenesis marker,cmlc2:EGFP that expresses GFP in the heart beginning at about 24hours post fertilization (h.p.f.). We generated an additionaltransposon in which the hsp70 promoter drives expression of EWS-FLI1 and the cmlc2 promoter drives GFP expression from aseparate cassette. Following injection of this transposon, weidentified a founder that transmitted progeny with GFP expressionin the heart. This transgenic line is denoted Tg(hsp70:EWS-FLI:IRES-EGFP; clmc2:EGFP)sw3 and will be referred to ashsp70:EWS-FLI1 for simplicity.
To determine the effect of transgenic EWS-FLI1 on embryonicgene expression, we heat shocked the transgenic embryos at 50%epiboly, collected total RNA 4 hours later, and performedmicroarray analysis (supplementary material Tables S4, S5). Therewas a modest overlap in genes upregulated in zebrafish SRBCTsand transgenic embryos, or downregulated in the tumors and thetransgenic embryos (supplementary material Table S6). Thismodest overlap is probably due to the fact that the highly dynamiccontext of a developing embryo, in which many different tissue-specific enhancers and pathways might be active, differs from thatof a tumor derived from one particular lineage. In a rapidlydeveloping embryo, indirect effects of EWS-FLI1 expression mightin fact predominate, making the comparison with human Ewing’stumors more problematic.
We speculated that the embryonic developmental phenotype ofEWS-FLI1 transgenic embryos might therefore provide a morerobust readout of the effects of EWS-FLI1 on conserved signaling
dmm.biologists.org100
Zebrafish model of Ewing’s sarcomaRESEARCH ARTICLE
Table 2. Gene-set enrichment analysis of zebrafish SRBCT gene sets in human cells and tumors
Zebrafish SRBCT
upregulated
Zebrafish SRBCT
downregulated
Expression dataset Reference Size NES FDR q-value Size NES FDR q-value
Knockdown of EWS-FLI1 in
Ewing’s cell linesTirode et al., 2007 70 1.17 NS 95 –1.26 NS
Kauer et al., 2009 70 1.47 0.005 95 –0.98 NS
Smith et al., 2006 70 1.26 NS 95 –1.6 0.03
Inducible expression of EWS-
FLI1Miyagawa et al., 2008 83 1.85 0.001 115 1.26 NS
Ewing’s vs other sarcomas Henderson et al., 2005 70 1.36 0.04 95 –1.04 NS
Ewing’s tumors vs bonemarrow stromal cells
Tirode et al., 2007 70 1.28 NS 95 –1.35 0.049
Ewing’s cell lines vs bone
marrow stromal
immortalized cell line
Miyagawa et al., 2008 83 1.79 <0.001 115 –1.35 0.04
NES, normalized enrichment score; FDR, false discovery rate; NS, not statistically significant; Size, number of genes with zebrafish orthologs in the human expression dataset.
Dise
ase
Mod
els &
Mec
hani
sms
D
MM
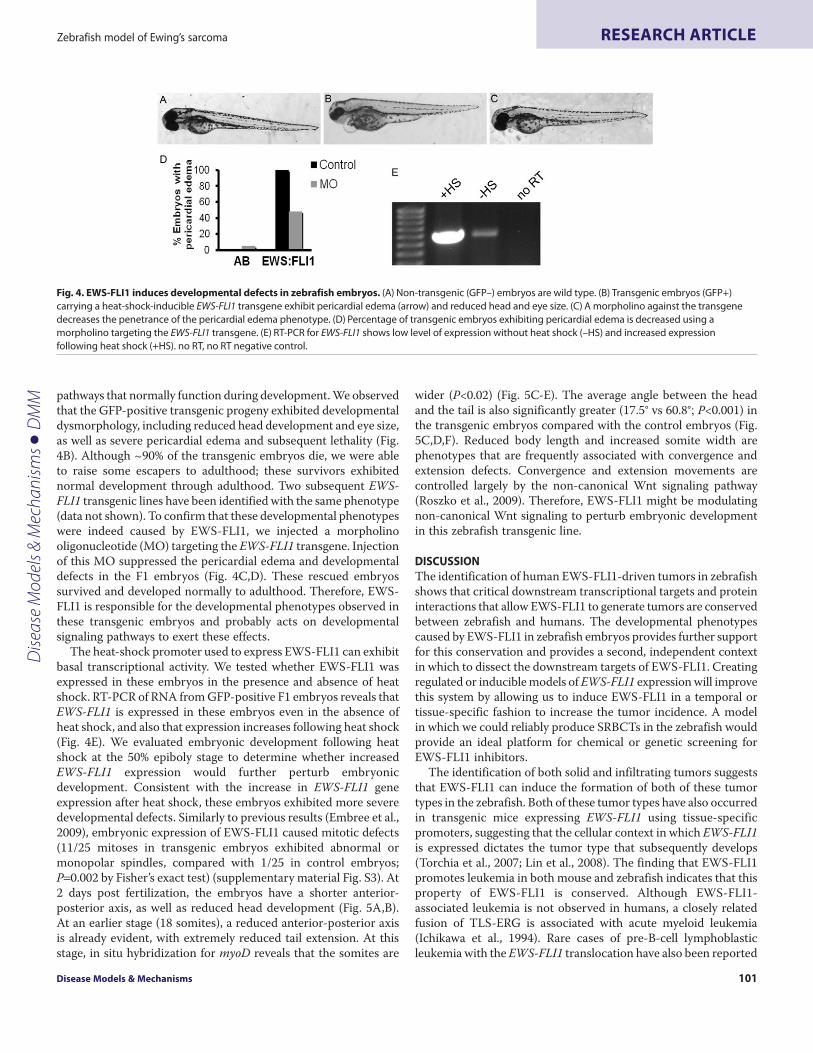
pathways that normally function during development. We observedthat the GFP-positive transgenic progeny exhibited developmentaldysmorphology, including reduced head development and eye size,as well as severe pericardial edema and subsequent lethality (Fig.4B). Although ~90% of the transgenic embryos die, we were ableto raise some escapers to adulthood; these survivors exhibitednormal development through adulthood. Two subsequent EWS-FLI1 transgenic lines have been identified with the same phenotype(data not shown). To confirm that these developmental phenotypeswere indeed caused by EWS-FLI1, we injected a morpholinooligonucleotide (MO) targeting the EWS-FLI1 transgene. Injectionof this MO suppressed the pericardial edema and developmentaldefects in the F1 embryos (Fig. 4C,D). These rescued embryossurvived and developed normally to adulthood. Therefore, EWS-FLI1 is responsible for the developmental phenotypes observed inthese transgenic embryos and probably acts on developmentalsignaling pathways to exert these effects.
The heat-shock promoter used to express EWS-FLI1 can exhibitbasal transcriptional activity. We tested whether EWS-FLI1 wasexpressed in these embryos in the presence and absence of heatshock. RT-PCR of RNA from GFP-positive F1 embryos reveals thatEWS-FLI1 is expressed in these embryos even in the absence ofheat shock, and also that expression increases following heat shock(Fig. 4E). We evaluated embryonic development following heatshock at the 50% epiboly stage to determine whether increasedEWS-FLI1 expression would further perturb embryonicdevelopment. Consistent with the increase in EWS-FLI1 geneexpression after heat shock, these embryos exhibited more severedevelopmental defects. Similarly to previous results (Embree et al.,2009), embryonic expression of EWS-FLI1 caused mitotic defects(11/25 mitoses in transgenic embryos exhibited abnormal ormonopolar spindles, compared with 1/25 in control embryos;P0.002 by Fisher’s exact test) (supplementary material Fig. S3). At2 days post fertilization, the embryos have a shorter anterior-posterior axis, as well as reduced head development (Fig. 5A,B).At an earlier stage (18 somites), a reduced anterior-posterior axisis already evident, with extremely reduced tail extension. At thisstage, in situ hybridization for myoD reveals that the somites are
wider (P<0.02) (Fig. 5C-E). The average angle between the headand the tail is also significantly greater (17.5° vs 60.8°; P<0.001) inthe transgenic embryos compared with the control embryos (Fig.5C,D,F). Reduced body length and increased somite width arephenotypes that are frequently associated with convergence andextension defects. Convergence and extension movements arecontrolled largely by the non-canonical Wnt signaling pathway(Roszko et al., 2009). Therefore, EWS-FLI1 might be modulatingnon-canonical Wnt signaling to perturb embryonic developmentin this zebrafish transgenic line.
DISCUSSIONThe identification of human EWS-FLI1-driven tumors in zebrafishshows that critical downstream transcriptional targets and proteininteractions that allow EWS-FLI1 to generate tumors are conservedbetween zebrafish and humans. The developmental phenotypescaused by EWS-FLI1 in zebrafish embryos provides further supportfor this conservation and provides a second, independent contextin which to dissect the downstream targets of EWS-FLI1. Creatingregulated or inducible models of EWS-FLI1 expression will improvethis system by allowing us to induce EWS-FLI1 in a temporal ortissue-specific fashion to increase the tumor incidence. A modelin which we could reliably produce SRBCTs in the zebrafish wouldprovide an ideal platform for chemical or genetic screening forEWS-FLI1 inhibitors.
The identification of both solid and infiltrating tumors suggeststhat EWS-FLI1 can induce the formation of both of these tumortypes in the zebrafish. Both of these tumor types have also occurredin transgenic mice expressing EWS-FLI1 using tissue-specificpromoters, suggesting that the cellular context in which EWS-FLI1is expressed dictates the tumor type that subsequently develops(Torchia et al., 2007; Lin et al., 2008). The finding that EWS-FLI1promotes leukemia in both mouse and zebrafish indicates that thisproperty of EWS-FLI1 is conserved. Although EWS-FLI1-associated leukemia is not observed in humans, a closely relatedfusion of TLS-ERG is associated with acute myeloid leukemia(Ichikawa et al., 1994). Rare cases of pre-B-cell lymphoblasticleukemia with the EWS-FLI1 translocation have also been reported
Disease Models & Mechanisms 101
Zebrafish model of Ewing’s sarcoma RESEARCH ARTICLE
Fig. 4. EWS-FLI1 induces developmental defects in zebrafish embryos. (A)Non-transgenic (GFP–) embryos are wild type. (B)Transgenic embryos (GFP+)carrying a heat-shock-inducible EWS-FLI1 transgene exhibit pericardial edema (arrow) and reduced head and eye size. (C)A morpholino against the transgenedecreases the penetrance of the pericardial edema phenotype. (D)Percentage of transgenic embryos exhibiting pericardial edema is decreased using amorpholino targeting the EWS-FLI1 transgene. (E)RT-PCR for EWS-FLI1 shows low level of expression without heat shock (–HS) and increased expressionfollowing heat shock (+HS). no RT, no RT negative control.
Dise
ase
Mod
els &
Mec
hani
sms
D
MM

(Ozdemirli et al., 1998; Jakovljevic et al., 2010). These cases suggesta connection between EWS translocation proteins and cancers ofthe hematopoietic lineage. Differences in cellular context, cellularniche, expression level, or temporal expression in animal modelsmay account for the lack of EWS-FLI1-associated leukemia inhumans.
Serial transplantation of two zebrafish tumors demonstrated theindependent growth of the tumor cells. The histological variationobserved following several rounds of transplantation could becaused by tumor evolution, stochastic differences in subpopulationsof cells transplanted, or differences in signals or environment withinthe host tissue. Although some rounds of transplantation requireda longer time for tumor development, the number of transplantrecipients was not large enough to confirm that this difference isstatistically significant. Presently, serial transplantation studiesperformed in zebrafish are becoming a powerful tool to studycancer in this model organism [reviewed by Taylor and Zon(Taylor and Zon, 2009)]. Most zebrafish cancer models to date havebeen shown to be serially transplantable; however, some differencesare becoming evident; a KRAS-induced MPD was unable to beserially transplanted (Le et al., 2007) and a T-ALL line showedincreasing malignancy with subsequent transplants (Frazer et al.,2009). Understanding the mechanisms underlying engraftment andproliferation in zebrafish cancer models might shed light on tumorprogression and metastasis in human cancer.
In the zebrafish model, we identified tumors in animals injectedwith a hsp70:EWS-FLI1 transgene, even in the absence of heatshock. In these tumors, the heat shock promoter is probably actingas a minimal promoter that allows EWS-FLI1 expression to becontrolled by local enhancers at the genomic site of transgeneintegration. Under this model, the two different histologies of
SRBCTs that we identified might arise because of trapping ofdifferent types of tissue-specific enhancers by the hsp70:EWS-FLI1transgene. Identification of these enhancers, and the tissue typethey define, could perhaps help clarify the Ewing’s sarcoma cell oforigin.
Mutation of tp53 provided a sensitized background for theformation of SRBCTs in zebrafish. Although mutations in tp53occur in only a subset of Ewing’s sarcomas (Huang et al., 2005),the p53 pathway might also be inhibited by other mechanisms (Banet al., 2008; Li et al., 2010). Interestingly, the solid SRBCTs that weidentified most frequently arose in proximity to the eye andabdomen, the locations in which MPNSTs also develop in tp53mutant zebrafish (Berghmans et al., 2005). This cell type might bepredisposed to tumor development in the zebrafish, or the MPNSTsand SRBCTS might both be able to arise from the same type ofnerve sheath cell. An alternative possibility is that the tumorsoriginate as a result of the loss of p53, but that the presence ofEWS-FLI1 drives the tumor cells to a less differentiated, moreprimitive state, which results in the SRBCT histology. Thesepossibilities cannot be distinguished from this study.
The microarray analysis provides firm support for the validityof this system as a model of human Ewing’s sarcoma. Theidentification of relevant target genes and the enrichment of thezebrafish gene set in both Ewing’s sarcoma cell lines and primarytumors reveal that EWS-FLI1 produces similar gene expressionchanges in the zebrafish tumors. By comparing the geneexpression profiles to MPNSTs, we eliminated those genes thatare generally affected in tumors and also those genes that mightbe changed because of the absence of p53. Therefore, additionalgene expression changes caused by EWS-FLI1 might also exist ifthey share these properties. Our gene set is therefore a
dmm.biologists.org102
Zebrafish model of Ewing’s sarcomaRESEARCH ARTICLE
Fig. 5. EWS-FLI1 affectsconvergence and extension inzebrafish embryos. Heat-shockedcontrol (A) and EWS-FLI1 transgenic(B) embryos at 2 d.p.f. reveals thattransgenic embryos are shorter withreduced head development. At the18 somite stage, in situ hybridizationfor myoD in heat-shocked control (C)and EWS-FLI1 transgenic (D) reveals awider somite width (E) and a shorteranterior-posterior axis as measuredby the angle between the head andtail (F) in transgenic embryos(*P<0.02, **P<0.001; error barsindicate s.e.m.).
Dise
ase
Mod
els &
Mec
hani
sms
D
MM

conservative estimate of the transcriptional changes caused byEWS-FLI1, but is also more likely to include specific targets andfeatures of Ewing’s sarcoma.
We identified some genes that were upregulated in zebrafishSRBCTs but downregulated in human EWS-FLI1-dependentexpression datasets, or vice versa. Differences in regulatoryelements, protein-protein interactions, cellular niche and growthfactors, and many other differences might exist between animalmodels and humans. Of course, animal models will almost nevercompletely recapitulate the human conditions. In this context, thesimilarity of these tumors suggests that these differences are notcrucial to tumorigenesis. Therefore, the gene expression profile wefound is likely to include relevant biological targets downstreamof EWS-FLI1. These gene sets also show enrichment in multiple,independent datasets generated from experiments in human celllines. Even among the various microarray analyses performed fromhuman cell lines, the differences between experiments are muchgreater than the similarities. A meta-analysis used to generate aEWS-FLI1 core-expression signature concluded that the core wasmore similar to primary Ewing’s sarcoma, than any of the individualexperiments (Hancock and Lessnick, 2008). Therefore, eachadditional model of Ewing’s sarcoma provides new informationtoward understanding the transcriptional consequences of EWS-FLI1 and, by extension, facilitates the development of improved,targeted therapies of Ewing’s sarcoma.
The stable EWS-FLI1-expressing transgenic line that weestablished represents a valuable new tool for the study of EWS-FLI1 cellular function. Many signaling pathways that function inzebrafish embryonic development have been dissected. Thephenotypes observed in the EWS-FLI1 embryos are consistent withdefects in convergence and extension during gastrulation. The non-canonical Wnt-PCP and BMP signaling pathways as well as othergenes, including those encoding Stat3, heterotrimeric G proteinsand cell adhesion molecules contribute to proper convergence andextension in zebrafish embryos (Roszko et al., 2009). Theconvergence and extension defect we observed in transgenic EWS-FLI1 embryos suggests that EWS-FLI1 directly interferes with genesthat function during this process; a finding made possible by theexpression of the transgene in the context of a developing embryo,in which the interplay of multiple signaling pathways determinesembryo morphology. Future experiments will be required to fullydissect the relationship between EWS-FLI1 and thesemorphological defects.
The phenotype we observed in the hsp70:EWS-FLI1 line was notidentical to that previously reported following mRNA injection(Embree et al., 2009). Similarly to our analysis, embryos with severetrunk defects, including a shorter axis, were observed, as well asabnormal mitoses. We speculate that mRNA injection results in ahigher level of EWS-FLI1 expression as well as an earlier onset ofexpression that might account for some of these differences.
This animal model of EWS-FLI1 function is amenable to futurepathway and genetic analysis and provides a new approach todissecting the role of EWS-FLI1. Genes that contribute to cancerfrequently have crucial roles in embryonic development; therefore,elucidating the mechanism by which EWS-FLI1 perturbsdevelopment will also shed light on its contribution totumorigenesis. Importantly, these zebrafish models represent thefirst vertebrate model for EWS-FLI1 function that is amenable to
genetic and chemical screening. A system to perform suppressorscreens for EWS-FLI1-induced phenotypes in a whole animalcontext has until now, been elusive. Future genetic and chemicalscreens for EWS-FLI1 suppressors in the zebrafish should revealnovel downstream effectors and therapeutics for the treatment ofEwing’s sarcoma.
METHODSZebrafish care, maintenance and mutantsZebrafish were raised according to standard protocols (Westerfield,2000). For mutation of tp53, the allele TP53zdf1 was used(Berghmans et al., 2005). A morpholino targeting the 5� end of theFLAG-EWS-FLI1 mRNA (5�-TGTCATCGTCGTCCTTGTAG -TCCAT-3�) was purchased from GeneTools (Philomath, OR). Heatshock of transgenic embryos was performed by incubating embryosat 39°C for 30 minutes at 50% epiboly.
Plasmid constructionThe plasmid containing a triple-FLAG-tagged human EWS-FLI1of the type 1 fusion, which fuses codon 265 of EWS to codon 219of FLI1 was provided by Chris Denny, University of California-LosAngeles, Los Angeles, CA. This plasmid was digested with SmaIand ClaI and ligated to the plasmid containing the 1.5 kb hsp70promoter digested with EcoRV and ClaI fragment (provided by JonKuwada, University of Michigan, Ann Arbor, MI). This plasmidwas then digested with SmeI and ClaI and ligated into pT2KXIGfollowing digestion with XhoI and ClaI to create pTZ109(hsp70:EWS-FLI1). pTZ109 was digested with NotI and XhoI torelease the ~3 kb hsp70:EWS-FLI1 fragment, which was then ligatedinto the 3.8 kb Tol2 transposon backbone of pT2KHG digested withXhoI and NotI (provided by Koichi Kawakami, National Instituteof Genetics, Shizuoka, Japan) to create pTZ307 (hsp70:EWS-FLI1).To create pME-EWS-FLI1, triple-FLAG-EWS-FLI1 was PCRamplified using primers with Gateway recombination sites attB1-EF1 (5�-GGGGACAAGTTTGTACAAAAAAGCAGGCTATG -GACTACAAGGACGACGATG-3�) and attB2-EF1 (5�-GGGG -ACCACTTTGTACAAGAAAGCTGGGTCTAGTAGTAGCTGC -CTAAGTGTG-3�) and recombined with pDONR221 using BPClonase II (Invitrogen, Carlsbad, CA). To generate constructs fortransposition, entry vectors p5E-bactin2 or p5E-hsp70l respectively,pME-EWS-FLI1, and p3E-IRES-EGFPpA (Kwan et al., 2007) were recombined with destination vector pDESTTol2A orpDESTTol2CG2, which includes cmlc2:EGFP, (provided by KoichiKawakami) in an LR reaction with LR Clonase II Plus (Invitrogen).Transposase RNA was synthesized from plasmid pCS2FA using theMMessage mMachine kit (Applied Biosystems/Ambion, FosterCity, CA). Plasmids were injected at a concentration of 10-100 ng/mlwith transposase RNA at 250 ng/ml.
Fixation and stainingFish were euthanized in Tricaine, dissected, fixed in 4%paraformaldehyde in 1� PBS for 2 days at 4°C., decalcified in 0.25M EDTA for 4-5 days at room temperature, embedded, sagittallysectioned and stained with hematoxylin and eosin (H and E). Forexamination of mitotic spindles, Tg(HSP:EWS-FLI1) embryos wereheat shocked at the 50% epiboly stage and fixed in 4%paraformaldehyde at 24 h.p.f. Anti-tubulin staining was performedas described (Verduzco and Amatruda, 2011). In situ hybridization
Disease Models & Mechanisms 103
Zebrafish model of Ewing’s sarcoma RESEARCH ARTICLED
iseas
e M
odel
s & M
echa
nism
s
DM
M

was performed as described (Thisse and Thisse, 2008). Embryoswere photographed and measurements of somite width and head-tail angle were performed in ImageJ.
Tumor transplantsThe tumor was removed, minced and washed in 0.9� PBS withFBS. Cells were counted and injected intraperitoneally into adultfish 2 days after irradiation (20 Gy).
RNA isolation, RT-PCR and microarray analysisRNA from tumors or embryos was isolated using Trizol followedby RNeasy purification (Qiagen). For leukemia-like tumors, onlyfixed tissue was available. The tumor-containing regions werescraped from the paraffin-embedded sections and then the RNAwas purified as described above. RT-PCR for human EWS-FLI1was performed using primers hEWS-FLi1-R: CTAGTAGT -AGCTGCCTAAGTGTG and hEWS-Fli1-Fint1: ATCCTACA -GCCAAGCTCCAAGT, which are not present in the zebrafishgenome. qPCR primers were resuspended at 500 nM each andvalidated by performing a tenfold dilution curve qPCR, startingwith 2000 ng cDNA, down to 2 ng cDNA, in duplicate. RPL13aprimers for reference were previously validated (Neumann et al.,2011). All qPCRs were performed with three technical replicateson and Applied Biosystems 7500 Real Time PCR system using iTaqFast SYBR green supermix with Rox from Bio-Rad (catalog no. 172-5101). Reactions were 20 ml total consisting of 2 ml primer mixes,10 ml SYBR supermix, and 2000 ng cDNA. The Forward (F) andReverse (R) primers used were: sox2, (F) GGAAACTTCTG -TCCGAGAGC, (R) GGCAGGGTGTACTTGTCCTT; nkx2.2a,(F) CTAGATGGCTCGCAACCAC, (R) CGGACGTGTCTT -GAGAGTTC; olig2, (F) ATGTCGATATTCCCGGTAGC, (R)CAAACACAAACACTGCACGA; rag2, (F) CAAACAGCTCTC -AGATTTCG, (R) CCAGGTCTAGTAAGGAGAAAC; lck, (F)GCCTCCAGTCAGTCAGAATTT, (R) TTGTATATGGCC -ACCACCAG; rpl13a, (F) TCTGGAGGACTGTAAGAGGTATGC,(R) AGACGCACAATCTTGAGAGCAG.
For microarray experiments, MPNST samples were taken fromtumors in three uninjected tp53 mutant adults. Solid small roundblue cell tumor samples were isolated from one primary tumor andtransplant recipient tumor from two independent primary tumors.Total RNA (2.5 mg) from tumors was reverse-transcribed using aT7-Oligo(dT) Promoter Primer, followed by RNaseH-mediatedsecond-strand cDNA synthesis. Biotinylated cRNA probe wasprepared with MessageAmp II (Ambion), fragmented andhybridized to Affymetrix GeneChip Zebrafish Genome Arraysaccording to the manufacturer’s protocol. Array data normalizedusing the RMA method and preprocessed using Chipster (FinnishIT Center for Science CSC). In accordance with the MIAMEguidelines, CEL files have been deposited at the GEO repository(Barrett and Edgar, 2006), under accession number GSE31217. Anempirical Bayes two groups test (P<0.05, Benjamini-Hochbergcorrected) was used to generate the list of 421 differentiallyexpressed probes. Human homologs of zebrafish genes werecompiled from three sources: Zebrafish Gene and MicroarrayAnnotation Project (ZGMAP, http://zfblast1.danio.tchlab.org/zgMap/Affymetrix.aspx), AILUN Cross-species mapping(http://ailun.stanford.edu/crossSpecies.php), and Database for Annotation, Visualization and Integrated Discovery
(DAVID, http://david.abcc.ncifcrf.gov/) (Huang et al., 2009a; Huang et al., 2009b). Gene set enrichment analysis (GSEA,http://www.broadinstitute.org/gsea/) was used to compare thezebrafish EWS-FLI1 up- and downregulated genes to humanmicroarray data (Subramanian et al., 2005). Analysis ofoverrepresented gene categories was performed using Gene SetAnalysis Toolkit V2 (http://bioinfo.vanderbilt.edu/webgestalt).
Publicly available datasets used for gene set enrichment analysis(GSEA)The datasets used for GSEA are available as follows. Expressionprofiling of Ewing’s sarcoma and rhabdomyosarcoma (Baer et al.,2004), GEO GSE967; expression profiling of human mesenchymaltumors (Henderson et al., 2005), Array Express: E-MEXP-353;knockdown of EWS-FLI1 in Ewing’s cell lines and expressionprofiling of Ewing’s tumors and Bone Marrow Stem Cells (Tirodeet al., 2007), GEO: GSE7007. Additional BMSC samples(Shahdadfar et al., 2005), E-MEXP-215; siRNA knockdown of EWS-FLI1 in Ewing’s cell lines (Kauer et al., 2009), Array Express: E-GEOD-14543; knockdown of EWS-FLI1 in Ewing’s cell lines(Smith et al., 2006), from http://www.huntsmancancer.org/publicweb/content/lessnick/mscSupplementalSmith2006_files/msc
dmm.biologists.org104
Zebrafish model of Ewing’s sarcomaRESEARCH ARTICLE
TRANSLATIONAL IMPACT
Clinical issueTraditionally, cancer treatment has relied on radiation and chemotherapy,which affect both the tumor as well as normal tissue. These treatments areparticularly detrimental to children and adolescents, whose developing tissuescan be severely impacted, resulting in long-term side effects even when thecancer is successfully treated. Targeted therapy in which only the cancer cellsare treated is the ideal solution to this problem. Cancers caused by fusiononcogenes resulting from chromosomal translocations (such as the EWS-FLI1protein that can cause Ewing’s sarcoma) are promising candidates for targetedtherapies, because they contain a unique protein that is not found in healthytissues. To date, few models have been successful in revealing the molecularfunction of EWS-FLI1 in cancer, which has made testing new therapies difficult.Thus, new animal models are needed to support translational studies aimed atdeveloping targeted therapies for Ewing’s sarcoma.
ResultsThe authors describe a new animal model for dissecting EWS-FLI1 functionusing the zebrafish Danio rerio. They generate the first reported lines oftransgenic zebrafish expressing human EWS-FLI1. The transgenic animalsexhibit two main phenotypes. First, adult fish expressing EWS-FLI1 in a subsetof cells develop tumors that resemble human Ewing’s sarcoma. Second,embryos that express EWS-FLI1 exhibit developmental defects, revealingspecific signaling pathways that are altered in the presence of EWS-FLI1.
Implications and future directionsBoth types of transgenic models reported here will be useful for the futurestudy of EWS-FLI1 function. The adult zebrafish model of Ewing’s sarcoma canbe used in forward genetic suppressor-enhancer screens to reveal the geneticfactors underlying tumor development, which might be candidates fortargeted therapies. In addition, the finding that transgenic embryos havedefects in convergence and extension suggests that EWS-FLI1 acts in part bysuppressing non-canonical Wnt signaling, offering a candidate pathway fornew therapies. Finally, because zebrafish embryos are small, transparent anddevelop externally, they are amenable to chemical screening. Therefore,transgenic embryos can be used in high-throughput chemical screens fordrugs that inhibit the effects of EWS-FLI1. Drugs identified using such screenswill be candidate therapeutics for Ewing’s sarcoma.
Dise
ase
Mod
els &
Mec
hani
sms
D
MM

Supplemental-Smith-2006.html. Inducible Expression of EWS-FLI1 in immortalized bone marrow stromal cell line and expressionprofiling of Ewing’s cell lines (Miyagawa et al., 2008), GEO: GSE8665and GSE8596. Expression profiling of Ewing’s sarcoma andNeuroblastoma (Staege et al., 2004), Array Express: E-GEOD-15757.ACKNOWLEDGEMENTSWe thank Chris Denny, Jon Kuwada, Chi-Bin Chien, Nathan Lawson and Tom Lookfor reagents.
FUNDINGThis work was supported by Kevin’s Ewing’s sarcoma Fund; Curing Kid’s Cancer; theAmon G. Carter Foundation; the American Cancer Society [grant number ACS-IRG60219604] (J.F.A.); the National Institutes of Health [grant number 1R01CA135731](J.F.A.); The Cancer Prevention and Research Institute of Texas [grant numberRP110395]; Welch Foundation [grant number I-1679] (J.F.A.); and a Post-DoctoralFellowship from the American Cancer Society [grant number PF0826701DDC](S.W.L.).
COMPETING INTERESTSThe authors declare that they do not have any competing or financial interests.
AUTHOR CONTRIBUTIONSJ.F.A. and S.W.L. conceived and designed the experiments. S.W.L., A.N.B., A.M.K. andG.L.C. performed the experiments. J.F.A., S.W.L. and D.R. analyzed the data. S.W.L.and J.F.A. wrote the paper.
SUPPLEMENTARY MATERIALSupplementary material for this article is available athttp://dmm.biologists.org/lookup/suppl/doi:10.1242/dmm.007401/-/DC1
REFERENCESAmatruda, J. F. and Patton, E. E. (2008). Genetic models of cancer in zebrafish. Int.
Rev. Cell Mol. Biol. 271, 1-34.Amsterdam, A., Lai, K., Komisarczuk, A. Z., Becker, T. S., Bronson, R. T., Hopkins, N.
and Lees, J. A. (2009). Zebrafish Hagoromo mutants up-regulate fgf8postembryonically and develop neuroblastoma. Mol. Cancer Res. 7, 841-850.
Baer, C., Nees, M., Breit, S., Selle, B., Kulozik, A. E., Schaefer, K. L., Braun, Y., Wai, D.and Poremba, C. (2004). Profiling and functional annotation of mRNA geneexpression in pediatric rhabdomyosarcoma and Ewing’s sarcoma. Int. J. Cancer 110,687-694.
Baird, K., Davis, S., Antonescu, C. R., Harper, U. L., Walker, R. L., Chen, Y., Glatfelter,A. A., Duray, P. H. and Meltzer, P. S. (2005). Gene expression profiling of humansarcomas: insights into sarcoma biology. Cancer Res. 65, 9226-9235.
Ban, J., Bennani-Baiti, I. M., Kauer, M., Schaefer, K. L., Poremba, C., Jug, G.,Schwentner, R., Smrzka, O., Muehlbacher, K., Aryee, D. N. et al. (2008). EWS-FLI1suppresses NOTCH-activated p53 in Ewing’s sarcoma. Cancer Res. 68, 7100-7109.
Barrett, T. and Edgar, R. (2006). Gene expression omnibus: microarray data storage,submission, retrieval, and analysis. Methods Enzymol. 411, 352-369.
Berghmans, S., Murphey, R. D., Wienholds, E., Neuberg, D., Kutok, J. L., Fletcher, C.D., Morris, J. P., Liu, T. X., Schulte-Merker, S., Kanki, J. P. et al. (2005). tp53 mutantzebrafish develop malignant peripheral nerve sheath tumors. Proc. Natl. Acad. Sci.USA 102, 407-412.
Brenner, J. C. and Chinnaiyan, A. M. (2009). Translocations in epithelial cancers.Biochim. Biophys. Acta 1796, 201-215.
Chen, J., Jette, C., Kanki, J. P., Aster, J. C., Look, A. T. and Griffin, J. D. (2007).NOTCH1-induced T-cell leukemia in transgenic zebrafish. Leukemia 21, 462-471.
Cheung, I. Y., Feng, Y., Danis, K., Shukla, N., Meyers, P., Ladanyi, M. and Cheung, N.K. (2007). Novel markers of subclinical disease for Ewing family tumors from geneexpression profiling. Clin. Cancer Res. 13, 6978-6983.
Dauphinot, L., De Oliveira, C., Melot, T., Sevenet, N., Thomas, V., Weissman, B. E.and Delattre, O. (2001). Analysis of the expression of cell cycle regulators in Ewingcell lines: EWS-FLI-1 modulates p57KIP2and c-Myc expression. Oncogene 20, 3258-3265.
Delattre, O., Zucman, J., Plougastel, B., Desmaze, C., Melot, T., Peter, M., Kovar, H.,Joubert, I., de Jong, P., Rouleau, G. et al. (1992). Gene fusion with an ETS DNA-binding domain caused by chromosome translocation in human tumours. Nature359, 162-165.
Deneen, B. and Denny, C. T. (2001). Loss of p16 pathways stabilizes EWS/FLI1expression and complements EWS/FLI1 mediated transformation. Oncogene 20,6731-6741.
Druker, B. J. (2009). Perspectives on the development of imatinib and the future ofcancer research. Nat. Med. 15, 1149-1152.
Embree, L. J., Azuma, M. and Hickstein, D. D. (2009). Ewing sarcoma fusion proteinEWSR1/FLI1 interacts with EWSR1 leading to mitotic defects in zebrafish embryosand human cell lines. Cancer Res. 69, 4363-4371.
Erkizan, H. V., Kong, Y., Merchant, M., Schlottmann, S., Barber-Rotenberg, J. S.,Yuan, L., Abaan, O. D., Chou, T. H., Dakshanamurthy, S., Brown, M. L. et al.(2009). A small molecule blocking oncogenic protein EWS-FLI1 interaction with RNAhelicase A inhibits growth of Ewing’s sarcoma. Nat. Med. 15, 750-756.
Frazer, J. K., Meeker, N. D., Rudner, L., Bradley, D. F., Smith, A. C., Demarest, B.,Joshi, D., Locke, E. E., Hutchinson, S. A., Tripp, S. et al. (2009). Heritable T-cellmalignancy models established in a zebrafish phenotypic screen. Leukemia 23, 1825-1835.
Hamelin, R., Zucman, J., Melot, T., Delattre, O. and Thomas, G. (1994). p53mutations in human tumors with chimeric EWS/FLI-1 genes. Int. J. Cancer 57, 336-340.
Hancock, J. D. and Lessnick, S. L. (2008). A transcriptional profiling meta-analysisreveals a core EWS-FLI gene expression signature. Cell Cycle 7, 250-256.
Henderson, S. R., Guiliano, D., Presneau, N., McLean, S., Frow, R., Vujovic, S.,Anderson, J., Sebire, N., Whelan, J., Athanasou, N. et al. (2005). A molecular mapof mesenchymal tumors. Genome Biol. 6, R76.
Hu-Lieskovan, S., Zhang, J., Wu, L., Shimada, H., Schofield, D. E. and Triche, T. J.(2005). EWS-FLI1 fusion protein up-regulates critical genes in neural crestdevelopment and is responsible for the observed phenotype of Ewing’s family oftumors. Cancer Res. 65, 4633-4644.
Huang, da W., Sherman, B. T. and Lempicki, R. A. (2009a). Systematic and integrativeanalysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 4, 44-57.
Huang, da W., Sherman, B. T. and Lempicki, R. A. (2009b). Bioinformatics enrichmenttools: paths toward the comprehensive functional analysis of large gene lists. NucleicAcids Res. 37, 1-13.
Huang, H. Y., Illei, P. B., Zhao, Z., Mazumdar, M., Huvos, A. G., Healey, J. H., Wexler,L. H., Gorlick, R., Meyers, P. and Ladanyi, M. (2005). Ewing sarcomas with p53mutation or p16/p14ARF homozygous deletion: a highly lethal subset associatedwith poor chemoresponse. J. Clin. Oncol. 23, 548-558.
Ichikawa, H., Shimizu, K., Hayashi, Y. and Ohki, M. (1994). An RNA-binding proteingene, TLS/FUS, is fused to ERG in human myeloid leukemia with t(16;21)chromosomal translocation. Cancer Res. 54, 2865-2868.
Jakovljevic, G., Nakic, M., Rogosic, S., Kardum-Skelin, I., Mrsic-Davidovic, S.,Zadro, R. and Kruslin, B. (2010). Pre-B-cell acute lymphoblastic leukemia with bulkextramedullary disease and chromosome 22 (EWSR1) rearrangement masqueradingas Ewing sarcoma. Pediatr. Blood Cancer 54, 606-609.
Kauer, M., Ban, J., Kofler, R., Walker, B., Davis, S., Meltzer, P. and Kovar, H. (2009). Amolecular function map of Ewing’s sarcoma. PLoS One 4, e5415.
Kawakami, K., Takeda, H., Kawakami, N., Kobayashi, M., Matsuda, N. and Mishina,M. (2004). A transposon-mediated gene trap approach identifies developmentallyregulated genes in zebrafish. Dev. Cell 7, 133-144.
Kwan, K. M., Fujimoto, E., Grabher, C., Mangum, B. D., Hardy, M. E., Campbell, D.S., Parant, J. M., Yost, H. J., Kanki, J. P. and Chien, C. B. (2007). The Tol2kit: amultisite gateway-based construction kit for Tol2 transposon transgenesisconstructs. Dev. Dyn. 236, 3088-3099.
Langenau, D. M., Feng, H., Berghmans, S., Kanki, J. P., Kutok, J. L. and Look, A. T.(2005). Cre/lox-regulated transgenic zebrafish model with conditional myc-induced Tcell acute lymphoblastic leukemia. Proc. Natl. Acad. Sci. USA 102, 6068-6073.
Langenau, D. M., Keefe, M. D., Storer, N. Y., Guyon, J. R., Kutok, J. L., Le, X.,Goessling, W., Neuberg, D. S., Kunkel, L. M. and Zon, L. I. (2007). Effects of RAS onthe genesis of embryonal rhabdomyosarcoma. Genes Dev. 21, 1382-1395.
Le, X., Langenau, D. M., Keefe, M. D., Kutok, J. L., Neuberg, D. S. and Zon, L. I.(2007). Heat shock-inducible Cre/Lox approaches to induce diverse types of tumorsand hyperplasia in transgenic zebrafish. Proc. Natl. Acad. Sci. USA 104, 9410-9415.
Lessnick, S. L., Dacwag, C. S. and Golub, T. R. (2002). The Ewing’s sarcomaoncoprotein EWS/FLI induces a p53-dependent growth arrest in primary humanfibroblasts. Cancer Cell 1, 393-401.
Li, Y., Tanaka, K., Fan, X., Nakatani, F., Li, X., Nakamura, T., Takasaki, M.,Yamamoto, S. and Iwamoto, Y. (2010). Inhibition of the transcriptional function ofp53 by EWS-Fli1 chimeric protein in Ewing Family Tumors. Cancer Lett. 294, 57-65.
Lin, P. P., Pandey, M. K., Jin, F., Xiong, S., Deavers, M., Parant, J. M. and Lozano, G.(2008). EWS-FLI1 induces developmental abnormalities and accelerates sarcomaformation in a transgenic mouse model. Cancer Res. 68, 8968-8975.
Mitelman, F., Johansson, B. and Mertens, F. (2007). The impact of translocations andgene fusions on cancer causation. Nat. Rev. Cancer 7, 233-245.
Miyagawa, Y., Okita, H., Nakaijima, H., Horiuchi, Y., Sato, B., Taguchi, T., Toyoda,M., Katagiri, Y. U., Fujimoto, J., Hata, J. et al. (2008). Inducible expression ofchimeric EWS/ETS proteins confers Ewing’s family tumor-like phenotypes to humanmesenchymal progenitor cells. Mol. Cell. Biol. 28, 2125-2137.
Disease Models & Mechanisms 105
Zebrafish model of Ewing’s sarcoma RESEARCH ARTICLED
iseas
e M
odel
s & M
echa
nism
s
DM
M

Mootha, V. K., Lindgren, C. M., Eriksson, K. F., Subramanian, A., Sihag, S., Lehar, J.,Puigserver, P., Carlsson, E., Ridderstrale, M., Laurila, E. et al. (2003). PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinatelydownregulated in human diabetes. Nat. Genet. 34, 267-273.
Neumann, J. C., Chandler, G. L., Damoulis, V. A., Fustino, N. J., Lillard, K.,Looijenga, L., Margraf, L., Rakheja, D. and Amatruda, J. F. (2011). Mutation in thetype IB bone morphogenetic protein receptor alk6b impairs germ-cell differentiationand causes germ-cell tumors in zebrafish. Proc. Natl. Acad. Sci. USA 108, 13153-13158.
Ohali, A., Avigad, S., Zaizov, R., Ophir, R., Horn-Saban, S., Cohen, I. J., Meller, I.,Kollender, Y., Issakov, J. and Yaniv, I. (2004). Prediction of high risk Ewing’ssarcoma by gene expression profiling. Oncogene 23, 8997-9006.
Owen, L. A., Kowalewski, A. A. and Lessnick, S. L. (2008). EWS/FLI mediatestranscriptional repression via NKX2.2 during oncogenic transformation in Ewing’ssarcoma. PLoS One 3, e1965.
Ozdemirli, M., Fanburg-Smith, J. C., Hartmann, D. P., Shad, A. T., Lage, J. M.,Magrath, I. T., Azumi, N., Harris, N. L., Cossman, J. and Jaffe, E. S. (1998).Precursor B-Lymphoblastic lymphoma presenting as a solitary bone tumor andmimicking Ewing’s sarcoma: a report of four cases and review of the literature. Am. J.Surg. Pathol. 22, 795-804.
Patton, E. E., Widlund, H. R., Kutok, J. L., Kopani, K. R., Amatruda, J. F., Murphey, R.D., Berghmans, S., Mayhall, E. A., Traver, D., Fletcher, C. D. et al. (2005). BRAFmutations are sufficient to promote nevi formation and cooperate with p53 in thegenesis of melanoma. Curr. Biol. 15, 249-254.
Riggi, N., Cironi, L., Provero, P., Suva, M. L., Kaloulis, K., Garcia-Echeverria, C.,Hoffmann, F., Trumpp, A. and Stamenkovic, I. (2005). Development of Ewing’ssarcoma from primary bone marrow-derived mesenchymal progenitor cells. CancerRes. 65, 11459-11468.
Roszko, I., Sawada, A. and Solnica-Krezel, L. (2009). Regulation of convergence andextension movements during vertebrate gastrulation by the Wnt/PCP pathway.Semin. Cell Dev. Biol. 20, 986-997.
Shahdadfar, A., Fronsdal, K., Haug, T., Reinholt, F. P. and Brinchmann, J. E. (2005).In vitro expansion of human mesenchymal stem cells: choice of serum is adeterminant of cell proliferation, differentiation, gene expression, and transcriptomestability. Stem Cells 23, 1357-1366.
Siligan, C., Ban, J., Bachmaier, R., Spahn, L., Kreppel, M., Schaefer, K. L., Poremba,C., Aryee, D. N. and Kovar, H. (2005). EWS-FLI1 target genes recovered from Ewing’ssarcoma chromatin. Oncogene 24, 2512-2524.
Smith, R., Owen, L. A., Trem, D. J., Wong, J. S., Whangbo, J. S., Golub, T. R. andLessnick, S. L. (2006). Expression profiling of EWS/FLI identifies NKX2.2 as a criticaltarget gene in Ewing’s sarcoma. Cancer Cell 9, 405-416.
Soda, M., Choi, Y. L., Enomoto, M., Takada, S., Yamashita, Y., Ishikawa, S., Fujiwara,S., Watanabe, H., Kurashina, K., Hatanaka, H. et al. (2007). Identification of thetransforming EML4-ALK fusion gene in non-small-cell lung cancer. Nature 448, 561-566.
Staege, M. S., Hutter, C., Neumann, I., Foja, S., Hattenhorst, U. E., Hansen, G., Afar,D. and Burdach, S. E. (2004). DNA microarrays reveal relationship of Ewing familytumors to both endothelial and fetal neural crest-derived cells and define noveltargets. Cancer Res. 64, 8213-8221.
Subramanian, A., Tamayo, P., Mootha, V. K., Mukherjee, S., Ebert, B. L., Gillette, M.A., Paulovich, A., Pomeroy, S. L., Golub, T. R., Lander, E. S. et al. (2005). Gene setenrichment analysis: a knowledge-based approach for interpreting genome-wideexpression profiles. Proc. Natl. Acad. Sci. USA 102, 15545-15550.
Taylor, A. M. and Zon, L. I. (2009). Zebrafish tumor assays: the state of transplantation.Zebrafish 6, 339-346.
Thisse, C. and Thisse, B. (2008). High-resolution in situ hybridization to whole-mountzebrafish embryos. Nat. Protoc. 3, 59-69.
Tirode, F., Laud-Duval, K., Prieur, A., Delorme, B., Charbord, P. and Delattre, O.(2007). Mesenchymal stem cell features of Ewing tumors. Cancer Cell 11, 421-429.
Tomlins, S. A., Rhodes, D. R., Perner, S., Dhanasekaran, S. M., Mehra, R., Sun, X. W.,Varambally, S., Cao, X., Tchinda, J., Kuefer, R. et al. (2005). Recurrent fusion ofTMPRSS2 and ETS transcription factor genes in prostate cancer. Science 310, 644-648.
Torchia, E. C., Boyd, K., Rehg, J. E., Qu, C. and Baker, S. J. (2007). EWS/FLI-1 inducesrapid onset of myeloid/erythroid leukemia in mice. Mol. Cell. Biol. 27, 7918-7934.
Uren, A. and Toretsky, J. A. (2005). Ewing’s sarcoma oncoprotein EWS-FLI1: theperfect target without a therapeutic agent. Future Oncol. 1, 521-528.
Verduzco, D. and Amatruda, J. F. (2011). Analysis of cell proliferation, senescence, andcell death in zebrafish embryos. Methods Cell Biol. 101, 19-38.
Westerfield, M. (2000). The zebrafish book. A guide for the laboratory use of zebrafish(Danio rerio). Eugene, Oregon: University of Oregon Press.
Zwerner, J. P., Joo, J., Warner, K. L., Christensen, L., Hu-Lieskovan, S., Triche, T. J.and May, W. A. (2008). The EWS/FLI1 oncogenic transcription factor deregulatesGLI1. Oncogene 27, 3282-3291.
dmm.biologists.org106
Zebrafish model of Ewing’s sarcomaRESEARCH ARTICLED
iseas
e M
odel
s & M
echa
nism
s
DM
M