aacr centennial series cancer research aacr centennial ... · aacr centennial series aacr...
TRANSCRIPT

Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
AACR Centennial Series
CancerResearch
AACR Centennial Series: The Biology of Cancer Metastasis:Historical Perspective
James E. Talmadge1 and Isaiah J. Fidler2
Abstract
Authors' ATransplantUniversityCancer Bio
CorresponAnderson CTX 77030.mdanderso
doi: 10.115
©2010 Am
www.aacr
Downl
Metastasis resistant to therapy is the major cause of death from cancer. Despite almost 200 years of study,the process of tumor metastasis remains controversial. Stephen Paget initially identified the role of host-tumorinteractions on the basis of a review of autopsy records. His “seed and soil” hypothesis was substantiated acentury later with experimental studies, and numerous reports have confirmed these seminal observations. Animproved understanding of the metastatic process and the attributes of the cells selected by this process iscritical for the treatment of patients with systemic disease. In many patients, metastasis has occurred by thetime of diagnosis, so metastasis prevention may not be relevant. Treating systemic disease and identifyingpatients with early disease should be our goal. Revitalized research in the past three decades has focusedon new discoveries in the biology of metastasis. Even though our understanding of molecular events thatregulate metastasis has improved, the contributions and timing of molecular lesion(s) involved in metastasispathogenesis remain unclear. Review of the history of pioneering observations and discussion of current con-troversies should increase understanding of the complex and multifactorial interactions between the host andselected tumor cells that contribute to fatal metastasis and should lead to the design of successful therapy.Cancer Res; 70(14); 5649–69. ©2010 AACR.
Introduction
Tumor metastasis is a multistage process during whichmalignant cells spread from the primary tumor to discontig-uous organs (1). It involves arrest and growth in different mi-croenvironments, which are treated clinically with differentstrategies depending on the tumor histiotype and metastaticlocation. Thus, a single hepatic metastasis may be resected,whereas bone marrow metastases may be treated with bone-targeting radioisotopes. Because of cellular heterogeneity,however, therapies have varying efficacy, challenging the on-cologist and our understanding of the metastatic process (2).Nonmalignant cells, under steady-state conditions, prolifer-ate as needed to replace themselves as they age or becomeinjured. However, this process can go awry, resulting in un-controlled proliferation and tumor formation, which can bebenign or malignant. Benign tumors are generally slow grow-ing, enclosed within a fibrous capsule, noninvasive, and mor-phologically resemble their cellular precursor. If a benigntumor is not close to critical vascular or neural tissue,prompt diagnosis and treatment frequently result in a cure.
ffiliations: 1The University of Nebraska Medical Center,ation Immunology Laboratory, Omaha, Nebraska; and 2Theof Texas M.D. Anderson Cancer Center, Department oflogy, Cancer Metastasis Research Center, Houston, Texas
ding Author: Isaiah J. Fidler, The University of Texas M.D.ancer Center, 1515 Holcombe Boulevard, Unit 854, Houston,Phone: 713-792-8580; Fax: 713-745-2174; E-mail: [email protected].
8/0008-5472.CAN-10-1040
erican Association for Cancer Research.
journals.org
on June 3, 202cancerres.aacrjournals.org oaded from
In contrast, malignant tumors rarely encapsulate, grow rap-idly, invade regional tissues, have morphologic abnormalitiessuch that their tissue of origin may be unrecognizable, andmetastasize. The term “metastasis” was coined in 1829 byJean Claude Recamier (1) and is now defined as “the transferof disease from one organ or part to another not directlyconnected to it” (Fig. 1; ref. 3). Metastasis is the primary clin-ical challenge as it is unpredictable in onset and it exponen-tially increases the clinical impact to the host.In many patients, metastasis has occurred by the time of
diagnosis, although this may not be apparent clinically (4).Although tumor metastasis can occur early in tumor progres-sion when the primary tumor is small or even undetectable,most occurs later when the primary tumor is larger. This pat-tern is supported by the observation that surgical excision ofsmaller lesions is often curative and forms the basis for tu-mor, nodes, metastasis (TNM) staging. When metastasis hasalready occurred, strategies targeting tumor cell invasion andother early aspects of the metastatic process may not be rel-evant to outcome. The suggestion, based on microarray anal-ysis, that all tumor cells within a malignant tumor are equallymetastatic is likely to be incorrect (5–7). This controversialconclusion is associated with the low sensitivity of microar-ray data that support the similarity of primary and meta-static tumor expression signatures, which are similarly dueto the activation of a metastatic genetic program in earlyprogenitors. It has been suggested that the overgrowth anddominance within primary and secondary lesions by a singletumor cell population, with a uniform metastatic signature,is associated with early metastasis. An extension of this hy-pothesis is that the late emergence of metastatic clones will
5649
0. © 2010 American Association for Cancer Research.

Talmadge and Fidler
5650
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
result in divergent expression patterns between primary tu-mors and metastases, secondary to the masking of meta-static signatures in the primary tumor, by persistingnonmetastatic clones (8). However, studies of clonal cell linesderived from late-stage human carcinomas (9) have provideddirect evidence that individual cancer cells, coexisting withina tumor, differ in their metastatic capability, including onesthat are nonmetastatic, confirming the tumor heterogeneityshown in preclinical studies with murine (10–13), as well ashuman tumors (14). Furthermore, the expression signaturesof tumors derived from cloned weakly and/or nonmetastatichuman cell lines and from their isogenic metastatic counter-parts from the same patient tumors differ, although theexpression signature of metastases and that of their corre-sponding primaries are similar (15).The pathogenesis of metastasis involves a series of steps,
dependent on both the intrinsic properties of the tumor cellsand the host response (16). In 1889, the English surgeonStephen Paget (17) identified the role of host-tumor cellinteractions. He addressed the question, “What is it thatdecides what organs shall suffer in a case of disseminatedcancer?” (17). On the basis of a review of autopsy recordsfrom 735 women with fatal breast cancer, he was able to pro-pose an answer. In addition, he remarked on the discrepancybetween the blood supply and frequency of metastasis tospecific organs. This discrepancy included a high incidenceof metastasis to the liver, ovary, and specific bones, and alow incidence to the spleen. His observations contradictedthe prevailing theory of Virchow (18), that metastasis couldbe explained simply by the arrest of tumor-cell emboli in thevasculature. Paget concluded that “remote organs cannot bealtogether passive or indifferent regarding embolism” andelaborated upon the “seed and soil” principle, stating: “Whena plant goes to seed, its seeds are carried in all directions, butthey can only live and grow if they fall on congenial soil.” Healso suggested, “All reasoning from statistics is liable to many
Cancer Res; 70(14) July 15, 2010
on June 3, 202cancerres.aacrjournals.org Downloaded from
errors. But, the analogy from other diseases seems to supportwhat these records have suggested, the dependence of theseed upon the soil” (17). Forty years later, in 1928, Ewingchallenged the “seed and soil” hypothesis (19). He proposedthat mechanical forces and circulatory patterns between theprimary tumor and the secondary site accounted for organspecificity. The seminal studies by Fidler and coworkers(10, 20) conclusively showed that, although tumor cells trafficthrough the vasculature of all organs, metastases selectivelydevelop in congenial organs.Studies in the late 1970s and early 1980s (10–26) stimulat-
ed research into the pathobiology of metastasis, resulting inextensive research into the local microenvironment, or“niche,” of the primary tumor and metastatic foci. They alsoprovided new insight into biological heterogeneity, the me-tastasis phenotype, and the selection of metastatic variantsduring or by the process of metastasis (10). Thus, the studiesof Paget remain a basis for ongoing research and continue tobe cited 120 years after publication (Fig. 2). The “seed andsoil” hypothesis is now widely accepted although the “seed”may now be identified as a progenitor cell, initiating cell,cancer stem cell (CSC), or metastatic cell, and the “soil”discussed as a host factor, stroma, niche, or organ microen-vironment (27). Regardless, few currently disagree that theoutcome of metastasis is dependent on interactions betweentumor cells and host tissue. Similar to Paget's seed and soilhypothesis, Fidler's studies have withstood the test of timeand have been repeatedly verified and validated. These re-sults focused on the process of metastasis being sequentialand selective with stochastic elements, that is, the threeS's (28). This concept was initially controversial and hasprovided a lightening rod on which new concepts andhypotheses have been tested (21). Inarguably, at least to apathologist, malignancy is not a characteristic shared by allcells within a tumor. This may seem obvious; however, recentliterature includes discussions in which the term “malignant”
Figure 1. Selected events that have influenced the history and progress of cancer metastasis, 1829–1979.
Cancer Research
0. © 2010 American Association for Cancer Research.

Cancer Metastasis
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
is used as a synonym for “neoplastic” or “tumorigenic.” In-deed, given the myopia associated with historical literature,researchers continue to rediscover classic observations, fre-quently under new synonyms. Unfortunately, these “new”observations do not always incorporate the detailed under-standing of tumor biology and experimental designs criticalto research into malignancy and metastasis. Studies focusedon metastasis require both in vivo and pathologic analysesbecause malignancy and metastasis are biologic phenomenaand can only be described based on in vivo analysis. In theabsence of whole animal studies, the potential for misinter-pretation is high.
The Pathogenesis of Metastasis
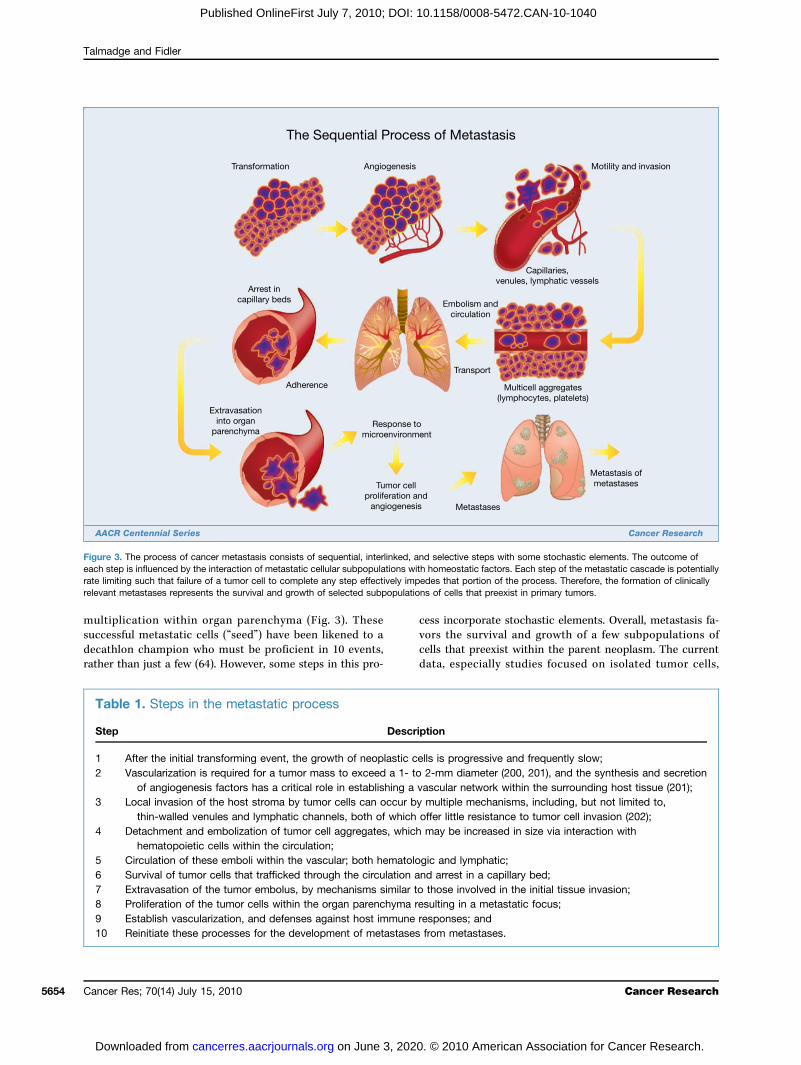
The process of cancer metastasis consists of sequential andinterrelated steps (Fig. 3), each of which can be rate limitingbecause a failure at any step may halt the process (16). Theoutcome of the process is dependent on both the intrinsicproperties of the tumor cells and the host response, such thatmetastasis is a balance of host-tumor cellular interactions thatcan vary between patients (29). In principle, the steps or eventsrequired for metastasis are the same for all tumors (Table 1).Thus, the outgrowth of a metastatic lesion requires that it de-velop a vascular network, evade the host's immune response(30), and respond to organ-specific factors that influencegrowth (31–33). Once they do, the cells can again invade thehost stroma, penetrate blood vessels, and enter the circulationto produce secondary metastases, or so-called “metastasis ofmetastases” (16, 34, 35). One critical aspect of this process isthe role of the arresting organ. As identified in the seed andsoil hypothesis, some organs can support the survival andgrowth of the tumor emboli or limit its survival. Further, bloodflow characteristics and the structure of the vascular systemcan also regulate the patterns of metastatic dissemination
www.aacrjournals.org
on June 3, 202cancerres.aacrjournals.org Downloaded from
(36, 37). Autopsy studies in breast and prostate cancer supportan increased number of bone metastases based on blood flow.Thus, some tumor-organ pairs of metastasis may have a posi-tive relationship with metastasis formation (38) and somemaybe associated with blood flow patterns (39).Only a few cells in a primary tumor are believed to be able
to give rise to a metastasis (22, 40), which is a result of theelimination of circulating tumor cells that fail to complete allthe steps in the metastatic process. The complexity of thisprogression explains, in part, why the metastatic processwas suggested to be inefficient (38). For example, the pres-ence of tumor cells in the circulation does not predict thatmetastasis will occur as most of the tumor cells that enterthe blood stream are rapidly eliminated (22). The intravenousinjection of radiolabeled B16 melanoma cells revealed that by24 hours after injection into the circulation, 0.1% or less ofthe cells were still viable, and less than 0.01% of tumor cellswithin the circulation survived to produce experimental lungmetastases (22). Observations such as this one raise the ques-tion of whether the development of metastases is the resultof fortuitous survival and growth of a few neoplastic cells, orrepresents the selective growth of a unique subpopulation ofmalignant cells with properties conducive to metastasis.That is, can all cells growing in a primary tumor produce sec-ondary lesions, or do only specific and unique cells possessthe properties required to enable them to metastasize? Be-cause this question has largely been addressed in rodents,and clinical studies have been more difficult, debate con-tinues. Recent data show, however, that human neoplasmsare biologically heterogeneous (41–43) and that the processof clinical metastasis is selective (44, 45).The presence of tumor cells or emboli distant from the pri-
mary tumor does not prove that metastasis has occurred.Circulating tumor cells are rapidly eliminated, and arrest ina capillary bed or the marrow is not indicative that a
Figure 1. Time Line. Continued.
Cancer Res; 70(14) July 15, 2010 5651
0. © 2010 American Association for Cancer Research.

Talmadge and Fidler
5652
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
metastatic focus will form or has formed. Indeed, cellular ar-rest during the metastatic process frequently results in cellu-lar apoptosis, dormancy, and, more rarely, a clinicallydetectable metastasis. Thus, the process of metastasis is in-efficient, seldom exceeding 0.01% (22), whereas the entry oftumor cells into the circulation is common and more than amillion cells per gram of tumor can be shed daily (46). Tarinand coworkers documented the low frequency of secondaryfoci using peritoneovenous shunts to reduce ascites in ovar-ian cancer (47). Although millions of tumor cells were direct-ly deposited into the vena cava every 24 hours by the shunt,patients rarely developed secondary tumors. Similarly, thefinding of epithelial cells within the marrow of patients withbreast cancer has prognostic indications (48). Not all cellswithin the marrow form metastatic nodules owing, in part,to host responses against the tumor cells. Thus, antitumorhost response can contribute to a soil incapable of support-ing tumor cell growth, or associated with the development oftumor dormancy, or both.The process of tumor initiation, progression, and metasta-
sis is not rapid, and tumor growth rates are generally slow.Mammography (49, 50) has shown that primary breast tu-mors have an average doubling time of 157 days (32), varyingfrom 44 to more than 1,800 days during exponential growth(51). Thus, the growth of a tumor from initiation to a size of1 cm (52), which is the limitation of most current imagingtools, requires an average of 12 years. This finding is signifi-cant because a 1-cm tumor has 109 cells and has undergoneat least 30 doublings from tumor initiation to diagnosis (4).On the basis of these parameters and the observation that atumor burden of approximately 1,000 cm3 is generally lethal(53), the time from diagnosis to mortality represents 10 dou-bling times from a 1-cm tumor and a shorter time frame.Thus, three quarters of a tumor's life history has occurredprior to diagnosis, and metastasis can occur prior to diagno-
Cancer Res; 70(14) July 15, 2010
on June 3, 202cancerres.aacrjournals.org Downloaded from
sis. Similarly, a tumor embolus associated with a surgicalshower would require 10 to 12 years to grow to a 1-cmmetastasis. These timelines are supported by registry data,which suggest that the median time from tumor resectionto a diagnosis of metastasis for patients with a T1 (<2 cm)tumor is 35 months versus 19.9 months for patients with aT3 (>5 cm) tumor (54). These data support the suggestionthat metastasis can occur years prior to diagnosis. The timerequired for the development of a malignant tumor andprogression from an adenoma to a malignant carcinomamay be longer than the progression of an adenoma to ametastasis, which may be because more mutations andclonal expansion(s) are required for progression to malig-nancy as compared with the number required to form ametastasis (55). The time between the appearance of a smalladenoma and the diagnosis ofmalignancymay be as long as 20to 25 years based on studies in patients with familial adeno-matous polyposis (56), and confirmed by serial studies of spo-radic colorectal cancer that revealed the transition from alarge adenoma to carcinoma takes about 15 years (57). Theseestimates are consistent with the doubling times of tumors,determined by serial radiologic studies and serial measure-ments of the carcinoembryonic antigen (CEA) serum biomark-er (58). This period differs from the observed mean doublingtimes of a metastasis, which may be 2 to 4 months (55).
The Organ Microenvironment
Clinical observations of cancer patients and studies inrodent models of cancer have revealed that some tumorphenotypes tend to metastasize to specific organs, inde-pendent of vascular anatomy, rate of blood flow, and num-ber of tumor cells delivered to an organ. Experimentaldata supporting Paget's “seed and soil” hypothesis havebeen derived from studies on the invasion and growth of
Figure 2. Selected events that have influenced the history and progress of cancer metastasis, 1980–Present.
Cancer Research
0. © 2010 American Association for Cancer Research.

Cancer Metastasis
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
B16 melanoma metastases in specific organs (20). In thesestudies, melanoma cells were injected intravenously intosyngeneic mice with tumor foci development in the lungsand in fragments of pulmonary or ovarian tissue implantedintramuscularly. In contrast, metastatic lesions did not de-velop in implanted renal tissue, or at the site of surgicaltrauma, suggesting that metastatic sites are not solelydetermined by neoplastic cell characteristics, or host mi-croenvironment (20). Ethical considerations rule out exper-imental analysis of cancer metastasis in humans, but theuse of peritoneovenous shunts for the palliation of ascitesin women with progressive ovarian cancer has provided anopportunity to study factors that affect metastatic spread(47). Human ovarian cancer cells can grow in the perito-neal cavity, either in the ascites fluid or by attaching tothe surface of peritoneal organs. These malignant cells,however, do not metastasize to other visceral organs.One (incorrect) explanation for the lack of visceral metas-tases is that tumor cells cannot gain entrance into thesystemic circulation. Studies by Tarin and colleagues ofmetastasis by ovarian tumors in patients whose asciticfluid was drained into the venous circulation addressedthis question (47). Clinically, this procedure resulted inpalliation with minimal complications; however, it allowedthe entry of viable cancer cells into the jugular vein. Au-topsy findings from 15 patients substantiated the clinicalobservations that the shunts did not significantly increasethe risk of metastasis to organs outside the peritonealcavity. Indeed, despite continuous entry of millions of tu-mor cells into the circulation, metastases to the lung, thefirst capillary bed encountered, were rare (47). These re-sults provide compelling verification of the “seed and soil”hypothesis. Organ-specific metastases have also beenshown in studies of cerebral metastasis after injection ofsyngeneic tumor cells into the internal carotid artery ofmice. Differences between two mouse melanomas were
www.aacrjournals.org
on June 3, 202cancerres.aacrjournals.org Downloaded from
found on the basis of patterns of brain metastasis suchthat the K-1735 melanoma produced lesions in the brainparenchyma (59) and the B16 melanomas produced men-ingeal metastases (60). Thus, different sites of tumorgrowth within an organ include differential interactionsbetween metastatic cells and the organ environment, pos-sibly in terms of specific binding to endothelial cells andresponses to local growth factors (61).
Seed and Soil
Since the 1980s, many investigators have contributed to ourunderstanding of cancer pathogenesis andmetastasis, and thedependency of these processes on the interaction betweencancer and homeostatic factors (62). Our concept of the “seedand soil” hypothesis consists of three principles. The first prin-ciple is that primary neoplasms (and metastases) consist ofboth tumor cells and host cells. Host cells include, but arenot limited to, epithelial cells, fibroblasts, endothelial cells,and infiltrating leukocytes. Moreover, neoplasms are biologi-cally heterogeneous and contain genotypically and phenotyp-ically diverse subpopulations of tumor cells, each of which hasthe potential to complete some of the steps in the metastaticprocess, but not all. Studies using in situ hybridization and im-munohistochemical staining have shown that the expressionof genes and/or proteins associated with proliferation, angio-genesis, cohesion, motility, and invasion vary among differentregions of neoplasms (62, 63). Because of the heterogeneity oftumors and the tumor infiltration by “normal” host cells, thesearch for genes and/or proteins that are associated withmetastasis cannot be conducted by the indiscriminate andnonselective analysis of tumor tissues.The second principle is that a focused analysis into the me-
tastatic process is needed to reveal the selection of cells thatsucceed in invasion, embolization, survival in the circulation,arrest in a distant capillary bed, and extravasation into and
Figure 2. Time Line. Continued.
Cancer Res; 70(14) July 15, 2010 5653
0. © 2010 American Association for Cancer Research.
Talmadge and Fidler
5654
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
multiplication within organ parenchyma (Fig. 3). Thesesuccessful metastatic cells (“seed”) have been likened to adecathlon champion who must be proficient in 10 events,rather than just a few (64). However, some steps in this pro-
Cancer Res; 70(14) July 15, 2010
on June 3, 202cancerres.aacrjournals.org Downloaded from
cess incorporate stochastic elements. Overall, metastasis fa-vors the survival and growth of a few subpopulations ofcells that preexist within the parent neoplasm. The currentdata, especially studies focused on isolated tumor cells,
Table 1. Steps in the metastatic process
Step
Description1
After the initial transforming event, the growth of neoplastic cells is progressive and frequently slow; 2 Vascularization is required for a tumor mass to exceed a 1- to 2-mm diameter (200, 201), and the synthesis and secretionof angiogenesis factors has a critical role in establishing a vascular network within the surrounding host tissue (201);
3 Local invasion of the host stroma by tumor cells can occur by multiple mechanisms, including, but not limited to,thin-walled venules and lymphatic channels, both of which offer little resistance to tumor cell invasion (202);
4 Detachment and embolization of tumor cell aggregates, which may be increased in size via interaction withhematopoietic cells within the circulation;
5 Circulation of these emboli within the vascular; both hematologic and lymphatic; 6 Survival of tumor cells that trafficked through the circulation and arrest in a capillary bed; 7 Extravasation of the tumor embolus, by mechanisms similar to those involved in the initial tissue invasion; 8 Proliferation of the tumor cells within the organ parenchyma resulting in a metastatic focus; 9 Establish vascularization, and defenses against host immune responses; and 10 Reinitiate these processes for the development of metastases from metastases.Figure 3. The process of cancer metastasis consists of sequential, interlinked, and selective steps with some stochastic elements. The outcome ofeach step is influenced by the interaction of metastatic cellular subpopulations with homeostatic factors. Each step of the metastatic cascade is potentiallyrate limiting such that failure of a tumor cell to complete any step effectively impedes that portion of the process. Therefore, the formation of clinicallyrelevant metastases represents the survival and growth of selected subpopulations of cells that preexist in primary tumors.
Cancer Research
0. © 2010 American Association for Cancer Research.

Cancer Metastasis
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
support a clonal origin for metastases, such that differentmetastases originate from different single cells (21, 64). Thethird principle is that metastatic development occurs inspecific organs, or microenvironments (“soil”), that are bio-logically unique. Endothelial cells in the vasculature of differ-ent organs express different cell-surface receptors (65) andgrowth factors that can support or inhibit the growth of meta-static cells (66). Thus, the outcome of metastasis depends onmultiple interactions (“cross-talk”) of metastasizing cellswith homeostatic mechanisms, such that the therapy ofmetastases can target not only the cancer cells, but alsothe homeostatic factors that promote tumor-cell growth,survival, angiogenesis, invasion, and metastasis (67).As discussed above, the analysis by Stephen Paget showed
that breast tumors preferentially spread to the liver, but notthe spleen. The bone marrow is also a preferential target forbreast cancer metastases, with some patients developing me-tastases more frequently than others. These observations (68,69) identify that the site of secondary tumors is dependent onthe tumor cell (“seed”) and the target organ (“soil”). Therefore,a metastatic focus is established only if the seed can grow inthe soil, that is, if the microenvironment of the target siteis compatible with the properties and requirements of thedisseminated tumor cell. Although this hypothesis was chal-lenged (2), it is now largely accepted that the anatomic archi-tecture of the blood flow is not sufficient to fully describe thepatterns of metastatic tumor spread. There are several molec-ular and cellular explanations that support the seed and soilhypothesis, including the observations that endothelial cellslining blood vessels in different organs express differentadhesion molecules (61); results from in vivo phage displaystudies suggest that every vascular bed may have its ownspecific molecular “address” (70); and the observation thattumor cells expressing the corresponding receptor mayuse these receptor-ligands to traffic to and arrest in specifictissues. There is also compelling evidence supporting a rolefor chemokines in the homing or chemoattraction of organ-selective metastasis (71). The CXCR4 receptor seems to havea role in the bone marrow, lymph nodes, and pulmonarytargeting of metastases from breast tumors that expressits ligand, CXCL12 (72). CXCR4 regulates chemotaxis in vitroand CXCR4-blocking antibodies have been shown to impairlung metastasis in SCID mouse xenograft experiments (71).However, in vivo, CXCR4 activation can also promote theoutgrowth of metastases in specific tissues rather than inva-sion (73). Recently, an immunohistochemical analysis ofCXCL12 expression in tumor endothelial cells was reportedto have a significant correlation with survival in patients withbrain metastases (74). Additional support for organ-specifictargeting has come from the gene-expression analysis ofbreast cancer cells selected for increased metastasis to bone(75) or lungs (76) that identified genes functionally involvedin organ-specific metastasis.
Cellular Infiltration of Tumors and Metastases
The role of tumor infiltration by leukocytes in tumor growthand development is complex. Although tumor-associated
www.aacrjournals.org
on June 3, 202cancerres.aacrjournals.org Downloaded from
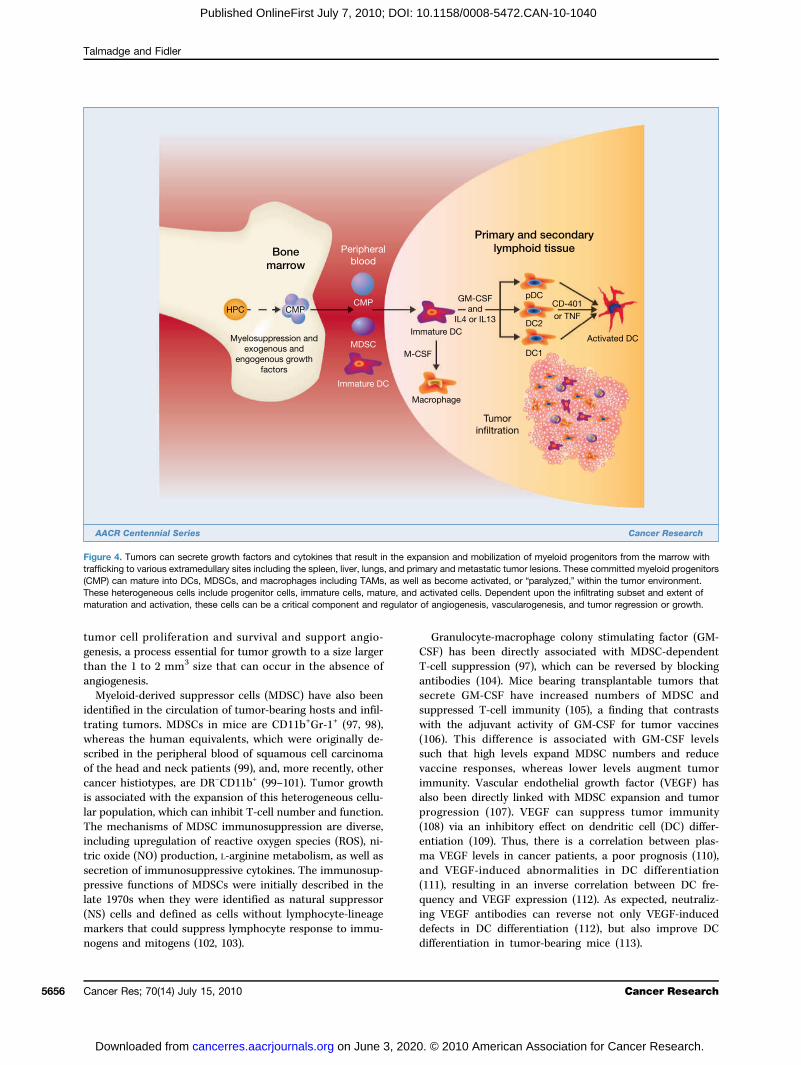
macrophages (TAM) have tumoricidal activity and can stim-ulate antitumor T cells, tumor cells can block inflammatorycell infiltration and the function of infiltrating cells (77).Tumor-derived molecules can regulate the expansion of my-eloid progenitors (Fig. 4), their mobilization from the mar-row, chemo-attraction to tumors, and activation, resultingin the promotion of tumor survival, growth, angiogenesis,and metastasis. Numerous studies have shown that followingactivation, macrophages can kill tumor cells in vitro (78).Indeed, if a macrophage-activating agent is injected priorto isolation of macrophages from metastases, they have tu-moricidal activity in vitro (78). However, tumor infiltrationby macrophages seems to have predominantly a protu-morigenic and/or metastatic phenotype; this was initiallyshown in a study in which the macrophage content ofsix carcinogen-induced fibrosarcomas was reported to di-rectly correlate with immunogenicity and, inversely, withmetastatic potential (79). Other studies using human breastcarcinomas and melanomas have been equivocal (80). Oneclinical study showed that patients with metastatic diseasehad a low macrophage infiltration (≤10%) of their primaryneoplasm, whereas 13 patients with benign tumors and 6out of 31 patients with malignant tumors and no clinical ev-idence of metastases had a TAM frequency of ≥10% (80).Thus, most (81–83), but not all studies (84), have shown thatthere is no relationship between immunogenicity, metastaticpropensity, and TAM frequency. Macrophages differentiatefrom CD34+ bone marrow progenitors following expansionand commitment and are mobilized from the marrow intothe periphery (56, 85), in which they differentiate intomonocytes and following invasion into a tissue, matureinto macrophages (86). Macrophage infiltration of primarytumors is regulated, at least in part, by cytokines, growthfactors, and enzymes secreted by the primary tumor. Theprimary tumor can also regulate the function of TAMs,including tumor cell cytotoxicity, which can be tumordependent (87).Clinical studies of macrophage infiltration of tumors have
suggested that it does not correlate with the immunogenicityand metastatic propensity (81–83, 88); rather, it is associatedwith a poor prognosis (89). Thus, a low frequency of macro-phage infiltration does not guarantee that a tumor will metas-tasize as multiple factors are critical (90). Macrophageinfiltration of tumors is regulated by tumor-associated che-moattractants such that host and/or tumor interactionsmay result in qualitative (87) or quantitative (91) regulationof macrophage infiltration, which can be obscured by thetechnologies used to assess macrophage infiltration (82, 92).Macrophages present in the extracellular matrix and capsulararea of tumors can facilitate tumor invasion and are importantduring the development of early stage lesions (93). Further,the proteolytic enzyme secreted by activated macrophagesfacilitates tumor invasion and extravasation. Indeed, in vitrococulture of macrophages with tumor cells can acceleratetumor growth (94). Macrophage infiltration of some tumormodels has been found to inversely correlate with relapse-freesurvival (RFS), microvessel density, and mitotic index (95).TAMs also express a number of factors (96) that stimulate
Cancer Res; 70(14) July 15, 2010 5655
0. © 2010 American Association for Cancer Research.
Talmadge and Fidler
5656
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
tumor cell proliferation and survival and support angio-genesis, a process essential for tumor growth to a size largerthan the 1 to 2 mm3 size that can occur in the absence ofangiogenesis.Myeloid-derived suppressor cells (MDSC) have also been
identified in the circulation of tumor-bearing hosts and infil-trating tumors. MDSCs in mice are CD11b+Gr-1+ (97, 98),whereas the human equivalents, which were originally de-scribed in the peripheral blood of squamous cell carcinomaof the head and neck patients (99), and, more recently, othercancer histiotypes, are DR−CD11b+ (99–101). Tumor growthis associated with the expansion of this heterogeneous cellu-lar population, which can inhibit T-cell number and function.The mechanisms of MDSC immunosuppression are diverse,including upregulation of reactive oxygen species (ROS), ni-tric oxide (NO) production, L-arginine metabolism, as well assecretion of immunosuppressive cytokines. The immunosup-pressive functions of MDSCs were initially described in thelate 1970s when they were identified as natural suppressor(NS) cells and defined as cells without lymphocyte-lineagemarkers that could suppress lymphocyte response to immu-nogens and mitogens (102, 103).
Cancer Res; 70(14) July 15, 2010
on June 3, 202cancerres.aacrjournals.org Downloaded from
Granulocyte-macrophage colony stimulating factor (GM-CSF) has been directly associated with MDSC-dependentT-cell suppression (97), which can be reversed by blockingantibodies (104). Mice bearing transplantable tumors thatsecrete GM-CSF have increased numbers of MDSC andsuppressed T-cell immunity (105), a finding that contrastswith the adjuvant activity of GM-CSF for tumor vaccines(106). This difference is associated with GM-CSF levelssuch that high levels expand MDSC numbers and reducevaccine responses, whereas lower levels augment tumorimmunity. Vascular endothelial growth factor (VEGF) hasalso been directly linked with MDSC expansion and tumorprogression (107). VEGF can suppress tumor immunity(108) via an inhibitory effect on dendritic cell (DC) differ-entiation (109). Thus, there is a correlation between plas-ma VEGF levels in cancer patients, a poor prognosis (110),and VEGF-induced abnormalities in DC differentiation(111), resulting in an inverse correlation between DC fre-quency and VEGF expression (112). As expected, neutraliz-ing VEGF antibodies can reverse not only VEGF-induceddefects in DC differentiation (112), but also improve DCdifferentiation in tumor-bearing mice (113).
Figure 4. Tumors can secrete growth factors and cytokines that result in the expansion and mobilization of myeloid progenitors from the marrow withtrafficking to various extramedullary sites including the spleen, liver, lungs, and primary and metastatic tumor lesions. These committed myeloid progenitors(CMP) can mature into DCs, MDSCs, and macrophages including TAMs, as well as become activated, or “paralyzed,” within the tumor environment.These heterogeneous cells include progenitor cells, immature cells, mature, and activated cells. Dependent upon the infiltrating subset and extent ofmaturation and activation, these cells can be a critical component and regulator of angiogenesis, vascularogenesis, and tumor regression or growth.
Cancer Research
0. © 2010 American Association for Cancer Research.

Cancer Metastasis
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
Recently, Finke and colleagues (114) reported that admin-istration of sunitinib, a receptor tyrosine kinase inhibitor, re-sulted in a significant decrease in MDSC within patients withmetastatic renal cell carcinoma (115). Whether this decreasewas associated with a reduction in VEGF levels and neoan-giogenesis or as an inhibition of Flt3-mediated expansion ofMDSC remains to be addressed. Regardless of this observa-tion there have been few studies that examined the infiltra-tion of tumors by MDSCs. In one recent rodent study, it wasshown that CCL2 mediated MDSC chemotaxis in vitro andthat migration or chemotaxis of MDSCs could be blockedwith neutralizing CCL2 antibodies (Abs) or by blockingCCR2. Sunitinib has also been shown to mediate reversal ofMDSC accumulation in renal cell carcinoma (115). Theseobservations suggest that the regulation of infiltrating mye-loid cells has the potential to control the growth of primarytumors and metastasis (Fig. 5).
Metastatic Heterogeneity
Cells with different metastatic properties have been iso-lated from the same parent tumor, supporting the hypoth-esis that not all the cells in a primary tumor cansuccessfully metastasize (41–43). Two approaches havebeen to show that cells within the parent neoplasm differin metastatic capacity. In the first approach, metastaticcells are selected in vivo, such that tumor cells are im-planted at a primary site or injected intravenously, metas-tases allowed to grow, individual lesions collected andexpanded in vitro, and these cells used to repeat the pro-cess. This cycle is repeated several times and the functionof the cycled cells compared with that of cells from theparent tumor to determine whether the selection processresulted in an enhanced metastatic capacity (116). Severalstudies have shown that the increase in metastatic capac-ity did not result from the adaptation of tumor cells to
www.aacrjournals.org
on June 3, 202cancerres.aacrjournals.org Downloaded from
preferential growth in a specific organ but, rather, wasdue to selection (23, 24, 117). This procedure was originallyused to isolate the B16-F10 line from the wild-type B16melanoma (116), but has also been successfully used toproduce tumor cell lines with increased metastatic capac-ity from many murine and human tumors (118, 119).In the second approach, cells are selected for an enhanced
expression of a phenotype believed to be important to a stepin the metastatic process and are then tested in vivo to de-termine whether their metastatic potential has changed. Thismethod has been used to examine properties as diverse asresistance to T lymphocytes (120), adhesive characteristics(121), invasive capacity (122, 123), lectin resistance (124),and resistance to natural killer cells (125). The finding thata characteristic, typically using a single tumor model withonly one or two variants, is associated with metastasis is of-ten considered as proof that this parameter is directly re-sponsible for metastasis. However, this can be modeldependent, associated with the selective procedure and hasresulted in conclusions and associations that are not repro-ducible. Clearly, numerous phenotypes are associated, inpart, with the metastatic process, but rarely represent allthe phenotypes critical to the final outcome. Thus, studiesof this nature, as well as comparisons of clonal subpopula-tions, lend insight into phenotypes that may have a role inmetastasis, but can lead one astray in terms of overall con-clusions into the process. This finding is also true of micro-array analyses, whereby, the primary tumor can havemultiple cellular phenotypes because of clonal heterogeneity,as well as the confounding variable of infiltrating “normal”host cells obfuscating conclusions. As such, conclusionsbased on a limited number of tumor samples and/or clones,into a process as complex as metastasis, have the potential tobe unsupportable.One criticism of studies into metastatic heterogeneity is
that isolated tumor line(s) may have arisen as a result of
Figure 5. The clonal selection model of metastasis suggests that the cell populations in the primary tumor with all of the genetic prerequisites required formetastatic capacity are the subpopulations that metastasize. Further, both the cells within the primary tumor and the metastatic lesion(s) can continueto diversify as the lesions grow.
Cancer Res; 70(14) July 15, 2010 5657
0. © 2010 American Association for Cancer Research.

Talmadge and Fidler
5658
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
an adaptive rather than a selective process, which was ad-dressed by the analysis of metastatic heterogeneity by Fidlerand Kripke in 1977 using the murine B16 melanoma (10).A modification of the fluctuation assay of Luria and Delbruck(126) showed that different tumor cell clones, each derivedfrom individual cells isolated from the parent tumor, variedin their ability to form pulmonary nodules following intrave-nous inoculation into syngeneic mice. Control subcloningprocedures showed that the observed diversity was not aconsequence of the cloning procedure (10). To exclude thepossibility that the metastatic heterogeneity found in theB16 melanoma might have been introduced as a result ofin vivo or in vitro cultivation, the biologic and metastaticheterogeneity of a mouse melanoma induced in a C3H/HeNmouse was also studied (127). This melanoma designatedK-1735 was established in culture and immediately cloned(128, 129). An experiment similar to that undertaken withthe B16 melanoma confirmed that (10) the clones differedfrom each other and the parent tumor in their ability toproduce lung metastases. In addition, the clones differed inmetastatic potential, resulting in metastases that varied insize and pigmentation. Metastases to the lungs, lymph nodes,brain, heart, liver, and skin were not melanotic, whereasthose growing in the brain were uniformly melanotic(59, 128, 129).An extension of studies into metastatic heterogeneity
assessed immunologic rejection by a normal syngeneic host(130, 131), as compared with young nude mice (132), amodel in which the immunologic barrier to metastasis isremoved and antigenic metastatic cells are able to success-fully complete the process. In these studies, cells of twoclones that did not produce metastases in normal syngeneicmice produced lung metastasis in the young nude mice.However, most of the nonmetastatic clones were nonmeta-static in both normal syngeneic and nude recipients. There-fore, the ability to metastasize in syngeneic mice wasprimarily not due to immunologic rejection, but by otherdeficiencies that prevented completion of a step in themetastatic cascade.The observation that preexisting tumor subpopulations
with metastatic properties exist in the primary tumor hasbeen confirmed by many laboratories using a wide rangeof tumors (16, 26, 30, 53, 133–135). In addition, studies us-ing young nude mice as models for metastasis of humanneoplasms have shown that human tumor cell lines andfreshly isolated human tumors, such as colon and renal car-cinomas, also contain subpopulations of cells with widelydiffering metastatic properties. Talmadge and Fidler under-took a series of studies to address the question of whetherthe cells that survive to form metastases possess a greatermetastatic capacity than cells in an unselected neoplasm(23, 25, 116). In these studies, cell lines derived from spon-taneous metastatic foci were found to produce significantlymore metastases than cells from the parent tumor. Studieswith heterogeneous, unselected tumors support the hypo-thesis that metastasis is a selective process, regulated bya number of different mechanisms. It should be noted thatmetastatic foci can be clonal in origin, yet originate from
Cancer Res; 70(14) July 15, 2010
on June 3, 202cancerres.aacrjournals.org Downloaded from
tumor emboli containing more than a single tumor cell. Tu-mor emboli composed of heterogeneous tumor cells canarise if an embolus is derived from a nonhomologous zonewithin a primary tumor (136). The zonal composition of atumor may play a role in the metastatic process, but alsohas the potential to obscure the results of microarray stud-ies. The zonal composition can result in the appearance ofminimal heterogeneity if a sample is obtained from a singlezone within a neoplastic lesion. Thus, as recommended byBachtiary and colleagues (137), microarray analyses need toconsider intratumoral heterogeneity and zonal gene expres-sion, such that multiple tumor biopsies should be studiedfrom discontiguous sites. Moroni and colleagues (138), us-ing human colorectal carcinomas, reported intratumoralheterogeneity and epidermal growth factor receptor (EGFR)gene amplification that correlated with protein levels. Ex-periments using human tumors in nude mice by Pettawayand colleagues (139) also revealed that orthotopic implemen-tation of heterogeneous human prostate cancer cell lines intonude mice resulted in rapid local growth and distant meta-static foci with an increased metastatic propensity to variousorgan sites including the gastrointestinal tract. The clonal or-igin of these metastatic foci was documented with karyotypicmarkers extending and confirming the prior studies by Fidlerand Talmadge (21).
The Clonal Origin of Metastases
Nowell (140, 141) suggested that acquired genetic vari-ability within developing clones of tumors, coupled withselection pressures, results in the emergence of new tumorcell variants that display increased malignancy. This hy-pothesis suggested that tumor progression toward malig-nancy is accompanied by heightened genetic instability ofthe progressing cells. To test this hypothesis, Cifone andFidler measured the mutation rates of paired metastaticand nonmetastatic cloned lines isolated from four differentmurine tumors (142). They found that highly metastaticcells were phenotypically less stable than their benigncounterparts. Moreover, the spontaneous mutation rate ofthe highly metastatic clones was several fold higher thanin nonmetastatic or poorly metastatic clones. These obser-vations were subsequently confirmed and extended withother murine (143) and human neoplasms (144, 145). Tak-en together, these studies suggest that metastatic tumorpopulations have a greater likelihood of cells undergoingrapid phenotypic diversification. This genetic instability,coupled with a Darwinian selection, may result in popula-tions resistant to normal homoeostatic growth controls,chemotherapeutic intervention, immune attack, and envi-ronmental restraints (146). Indeed, this process can be ex-tended by the mutagenic activity of drugs commonly usedto treat tumors (147). This biological heterogeneity isfound both within a single metastasis (intralesional hetero-geneity) and among different metastases (interlesional het-erogeneity). The heterogeneity reflects two processes: theselective nature of the metastatic process and the rapidevolution and phenotypic diversification of clonal tumor
Cancer Research
0. © 2010 American Association for Cancer Research.

Cancer Metastasis
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
growth, resulting from the inherent genetic and phenotypicinstability of clonal tumor cell populations. Like primaryneoplasms, metastases have the potential for a unicellularor multicellular origin. To determine whether metastasesarise from the same clone, or whether different metastasesare produced from different progenitor cells, Talmadge andcolleagues designed a series of experiments based on thegamma-irradiation induction of random chromosomebreaks and rearrangements to serve as “markers” (148).They examined the metastatic phenotype of individuallyspontaneous lung metastases that arose from subcutane-ously implanted tumors (148). In 10 metastases, all thechromosomes were normal, making it impossible to es-tablish whether they were of uni- or multicellular origin.In other lesions, unique karyotypic patterns of abnormalmarker chromosomes were found, indicating that eachmetastasis originated from a single progenitor cell.Subsequent experiments showed that when heteroge-neous clumps of different melanoma cell lines wereinjected intravenously, the resulting lung metastases wereof unicellular origin (21). These studies indicated thatregardless of whether an embolus was homogeneousor heterogeneous, metastases originate from a singleproliferating cell. The clonality of metastases has alsobeen reported for numerous other tumors, includingbreast cancer and fibrosarcomas, as well as melano-ma (145).The role of clonal selection (Fig. 6) during the process
of metastasis and the emergence of successive clonal sub-populations has been supported by studies in which indi-vidual cells were tagged by unique markers allowing themto be tracked (149). This tracking has been undertakenwith radiation-induced cytogenetic (chromosomal) markers(28), which clearly showed clonal selection. This observa-tion was confirmed with the use of drug resistance mar-
www.aacrjournals.org
on June 3, 202cancerres.aacrjournals.org Downloaded from
kers (150) and X-linked isoenzyme mosaicisms (151).However, all of these approaches are limited and recentstudies have used markers on the basis of the random in-sertion of transfected plasmids, which results in the gen-eration of large numbers of uniquely marked cell clones(139, 152, 153). Clearly, clonal selection and intrahostmetastatic heterogeneity are supported by preclinicalstudies (148, 150, 151, 153–155). However, clinical confir-mation has been challenging owing to methodologicalimpediments. Metastatic tissue is generally obtainedasynchronously, relative to primary tumor tissue, and com-parisons complicated by tissue availability. Several studieshave successfully compared primary tumors to metastaticfoci within the same patient on the basis of a number ofdifferent phenotypes. Kuukasjarvei and colleagues (44) an-alyzed the genetic composition of 29 primary breast carci-nomas and paired asynchronous metastases. They foundthat 69% of the metastases had a high degree of clo-nality with the corresponding primary tumor, whereaschromosomal X inactivation patterns supported the re-maining metastatic lesions as originating from thesame clone as the primary tumor. It was concluded thatalthough all metastases were derived from the parenttumor, metastasis occurred at different times. Studiesusing microarray analyses have also addressed thisprocess and are discussed in the section below entitled“Microarray analysis of heterogeneity, clonality, andprognostic potential.”Results from the microarray and computational analy-
ses have revealed that a small number of genes are dif-ferentially expressed between tumors and metastases,supporting the conclusion that although metastases re-semble the primary tumor on a mutational basis, a fewgenes differ consistently and are of significant mechanisticimportance.
Figure 6. The immune cells infiltrating tumors can regulate their growth, progression, and metastasis. Tumor regression is associated with infiltration bymature DCs and cytotoxic T cells (CTL) and type 1 T-helper type 1 bioactivity. Contrasting with this, tumor growth is facilitated via immune-mediatedimmunosuppression and neoangiogenesis following infiltration of tumors with immature DCs, MDSCs, plasmacytoid DCs (pDC) M2 macrophages, as wellas T-regulatory (T-reg) cells and a low frequency of CD4 and CD8 effector T cells.
Cancer Res; 70(14) July 15, 2010 5659
0. © 2010 American Association for Cancer Research.

Talmadge and Fidler
5660
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
Current Controversies in Metastatic Research
Premetastatic nicheRecently, it was suggested that to form a metastatic fo-
cus, primary tumors produce factors that induce the forma-tion of an appropriate environment in the soil (organ) priorto the seeding of metastatic cells. This suggestion led to theconcept of a premetastatic niche, whereby a special, per-missive microenvironment in the secondary site is devel-oped (156). The initial study into a premetastatic nicheindicated that vascular endothelial growth factor receptor1 (VEGFR1+) hematopoietic progenitor cells contribute tometastatic spread (157). These VEGFR1+ cells were sug-gested to arrive at a distant organ site, forming a premeta-static niche of clustered bone marrow hematopoietic cells(157). They were suggested to alter the tissue microenviron-ment and promote the recruitment of tumor cells by theupregulation of integrins and cytokines. The blockade ofVEGFR1 cells in vivo has been suspected to abrogate theformation of premetastatic niches and inhibit the growthof metastatic foci. The regulation of VLA-4 and fibronectinby VEGFR1+ cells also helps dictate which organs attracthematopoietic progenitors and support their adhesion,thereby promoting premetastatic niches (157). In addition,this adhesion molecule expression may contribute to surviv-al of the metastatic tumor cells as discussed in the sectionon MDSCs. Gao and colleagues showed that angiogenesis isessential for the growth of micrometastases (158), and thatthis process was regulated by bone marrow–derived endo-thelial progenitor cells (EPC). At the micrometastatic stage,EPCs traffic to and colonize micrometastases. Acceleratingangiogenesis facilitates the growth into gross metasta-ses. The blockade of EPCs by short hairpin RNA directedagainst the Id1 gene was found to inhibit angiogenesis atthe micrometastatic stage and to impair subsequent growth(158). Furthermore, although the number of EPCs contrib-uting to angiogenesis in metastases remained low (in therange of 11%), their impact on the development of micro-metastases and rapid cell proliferation is critical for pro-gressive growth of metastasis (159).Thus, experimental evidence supports the formation of
a premetastatic niche, which can facilitate metastasis viaVEGFR1+ bone marrow–derived hematopoietic progenitorcells (157). These cells express hematopoietic markers,and chemokines and chemokine receptors (e.g., CXCL12/CXCR4), promoting either their homing to the target tis-sue or recruitment and/or attachment of tumor metastaticcells. This process includes the CXCL12/CXCR4 axis,which has a critical role in stem cell migration (160). Ac-tivation of CXCR4 induces motility, chemotactic response,adhesion, secretion of MMPs, and release of angiogenicfactors, such as VEGF-A. Thus, disseminating tumor cells,in order to survive at distant sites, need a supportive mi-croenvironment similar to that found in the primary tu-mor. One way this may occur is by the mobilization andrecruitment to a premetastatic niche of immune and/or in-flammatory cells that can stimulate growth of metastaticcells (161).
Cancer Res; 70(14) July 15, 2010
on June 3, 202cancerres.aacrjournals.org Downloaded from
Multiple cellular and molecular mechanisms, wherebytumor cells interact with immune cells, regulate theirnumber and function, and allow tumor cells to escapefrom immunologic recognition and control (Fig. 4). Theseimmunoregulatory events have important physiological andpathophysiological significance, not only for host-tumor in-teractions and clinical outcomes, but also for clinical inter-vention. Studies to date have focused on the inflammatorycell infiltration within tumors and draining lymph nodes and,more recently, subset analyses. T-lymphocytic responses totumors are generally associated with a good prognosis; how-ever, vaccine strategies have had little impact to date (162).This is due, in part, to the escape mechanisms that tumorsuse to circumvent immunologic responses. Host-tumor inter-actions limit lymphocyte infiltration such that the majority ofinfiltrating cells are from the innate immune system (163)and are typically immature or with impaired function (162).These cells can “paralyze” the induction of T-cell and DC re-sponses (164), in addition to facilitating tumor proliferationand metastasis (165) via pro-angiogenic activity. Indeed, theimmune cells infiltrating tumors can secrete immunosup-pressive enzymes, resulting in lymphocytic apoptosis andthe induction of tolerance (166). Critical questions remain,including how tumors become refractory to immune inter-vention strategies to overcome these immunosuppressivemechanisms.
Epithelial-mesenchymal transitionsMolecular technologies, including DNA, antibody, and pro-
teomic arrays, and short-hairpin RNA libraries, have beenused to identify multiple molecules that contribute to themetastatic process. This group includes growth factors, cyto-kines and chemokines, pro-angiogenic factors, extracellularmatrix-remodeling molecules, and, most recently, transcrip-tion factors that may regulate cellular changes during tumorinvasion-metastasis. These transcription factors include Slug,Snail, Goose-coid, Twist, and ZEB1, which are highly ex-pressed by metastatic cells and have been suggested to havea role in inducing epithelial-mesenchymal transitions (EMT;refs. 167–170), defined as a rapid and reversible change incellular phenotypes. This concept originated from studies in-to embryologic development, including the change in pheno-type and behavior of cells invaginating from the surface intothe interior of an embryo to form the mesoderm during gas-trulation (171), changes during renal organogenesis, and theorigin and fate of the neural crest (172). EMT studies havefocused on the invasive behavior and spindling of culturedcells (173), using explanted embryonic tissue (174). Thus,the potential relevance of EMT to tumor progression islargely based on in vitro studies (175), using transformedepithelial cells and growth factors or extracellular matrixmanipulations. Genes upregulated during tumor progres-sion and metastasis are associated with multiple processes,including embryogenesis, tissue morphogenesis, and woundhealing (176). In neoplasia, EMT is suggested to result inthe loss of epithelial properties, including cell-cell adhesionand baso-apical polarity, and the gain of mesenchymalproperties, such as an increased ability to migrate on and
Cancer Research
0. © 2010 American Association for Cancer Research.

Cancer Metastasis
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
invade through extracellular matrix proteins (176). EMT hasalso been associated with the loss of expression and/ormislocalization of proteins involved in the formation ofcell-cell junctions, such as E-cadherin, zonula occludens(ZO-1) and/or claudin, and the gain of mesenchymal pro-tein expression. Thus, a depression in E-cadherin expressionis frequently observed during cancer progression (171), andsolid cancers are associated with a loss or downregulationof E-cadherin expression (171, 177).EMT has been suggested to have a role in the progression
of benign tumor cells to invasive and metastatic cells (171).This suggestion is largely based on cell culture, microarray,and histiologic studies. EMT proponents have suggested thattumors that display malignant histopathologic featuresincluding loose, scattered cancer cells and spindle cell cytol-ogy, which are more aggressive based on grade, stage, andclinical outcome, support the concept that EMT is mecha-nistically significant (168). Inarguably, carcinomas are histo-logically heterogeneous in addition to being infiltrated bymesenchymal cell histiotypes. The histologic identificationof mesenchymal elements in human tumors, identified asfetal-type fibroblasts and malignant stroma was describedyears ago (178). Indeed, a collection of 100 dual-stainingspindle cell breast carcinomas was reported in 1989 (179).Further, antibodies against keratin 14 and smooth muscleactin have identified spindle cell elements, which may ormay not be clinically significant (180, 181). Foci with a his-tologic appearance of EMT are found in human tumors andthis anaplastic appearance, including loss of cell polarity andlineage-specific or tissue-specific cytology are common fea-tures of carcinomas. The morphologic disorder, pleomor-phism, and anaplasia as observed by an unskilled observermay be suggestive of a mesenchymal lineage. The examplescited as evidence of EMT in clinical neoplasms—includingscattered single cell infiltration by lobular carcinomas ofthe breast (171) or diffuse carcinomas of the stomach (182),spindle cell differentiation in squamous carcinomas, andblending of sheets of carcinomatous cells into a highly cellular,desmoplastic stroma without a clear line of demarcation—potentially represent inappropriate analysis of these lesions.As discussed below, diffusely infiltrating cells in lobular carci-nomas retain their epithelial identity.The data supporting EMT, on the basis of in vitro observa-
tions, have largely used two-dimensional environments thatdo not contain host stroma, such as vascular, immune,endocrine, or neurologic elements. Similarly, studies of cellsin three-dimensional matrices do not recapitulate the hostenvironment, and the identification of cells in vitro as epithe-lial or mesenchymal in origin, based on morphology, is sub-jective. Indeed, studies in which epithelial and mesenchymalcell markers are used to provide evidence for a switch fromone differentiated cell lineage to another are also suspect asthey relate a marker to a morphologic phenotype, whichitself is claimed to transmute into another cell type and thusthe identification process becomes self-fulfilling. Inappro-priate gene expression by tumors is well established, andgenetically unstable cells and infiltrating cells could contri-bute to shifts in lineage markers. Indeed, it is difficult to
www.aacrjournals.org
on June 3, 202cancerres.aacrjournals.org Downloaded from
define cells on morphologic criteria alone, and the gain orloss of markers is insufficient to conclude that a cell popu-lation has undergone whole-scale gene expression repro-gramming. Although the presence of EMT is largely arguedon the basis of evidence from in vitro experiments, the in vivodata are unclear (183). Whether the progression of a nonin-vasive tumor into a metastatic tumor involves EMT (177,183) needs further study. This controversy is extended bythe rarity of EMT-like morphological changes in primary tu-mors and the observation that metastases are histologicallyand molecularly similar to the primary tumor. Thus, the roleof EMT in tumor metastasis is difficult to assess as it is chal-lenging to obtain tumor cells that have activated in the EMTprogram for or during metastasis (171).
Microarray analysis of heterogeneity, clonality, andprognostic potentialExpression profiling has revealed “metastasis signatures”
or poor prognosis gene signatures expressed by neoplasticcells within individual carcinomas and the potential to pre-dict metastasis (5, 184). This finding has been suggested toinfer that metastatic propensity might be an intrinsic prop-erty of a primary tumor, which is developed relatively early inmultistep tumor progression and is, therefore, expressed bythe bulk of neoplastic cells within a tumor. It has also beenproposed (75, 76) that a set of genes may mediate metastasisto specific organs, such as to the bone marrow or lungs.Studies by Urquidi and colleagues (9) provided direct proofthat individual cancer cells coexist within a tumor, differ inmetastatic capability, and that metastatic primary tumorscontain tumor subpopulations with variant metastatic pro-clivities and expression profiles. Montel and colleagues(185) reported that as the metastatic proclivity of a tumorincreases, the cellular populations within the tumor developincreasingly variable expression profiles. They suggested, onthe basis of the concept of tumor progression (140), that themetastatic foci selected during clonal evolution represent thefinal stage in the development of a metastasis. However, aswe have reviewed here, tumor progression is a continuumand does not end with metastasis. It continues during thegrowth of primary tumors and metastatic nodule(s), includ-ing the development and selection of variants resistant tochemotherapy. Tumor cells within a metastasis, at least ini-tially, express all of the genetic alterations necessary for themetastatic process. However, as tumors progress, hetero-genic tumor subpopulations develop within a metastasis, in-cluding cells with different metastatic capabilities. Gancbergand colleagues (186) examined pairs of primary and meta-static tumors from 100 breast cancer patients and reportedthat 6% had discordant Her2/neu overexpression by the me-tastatic tissue as compared with the primary tumor. FISHanalysis from 68 of the patients revealed a 7% discordancebetween primary and secondary lesions, but from differentpatients than those found by immunohistochemistry. Inaddition, they examined Her2/neu overexpression frommultiple metastatic lesions in 17 patients, revealing that18% differed in Her2/neu expression. Suzuki and Tarin (8)also examined matched, primary breast tumors and their
Cancer Res; 70(14) July 15, 2010 5661
0. © 2010 American Association for Cancer Research.

Talmadge and Fidler
Ca5662
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
Table 2. Metastasis timeline
Year
ncer Res;
Download
Discovery
1829
Jean Claude Recamier developed the term “metastasis” (1). 1858 Rudolf Virchow suggested that metastatic tumor dissemination was determined by mechanical factors (18). 1889 Stephen Paget proposed the “seed and soil” hypothesis to explain the organs that are afflicted withdisseminated cancer (17).
1915 Makoto Takahashi developed the first murine model of metastasis (203). 1929 James Ewing proposed that metastasis was determined by the anatomy of the vascular and lymphatic channelsthat drain the primary tumor (19).
1944 Dale Rex Coman identified the role of cellular adhesiveness to metastasis and the distribution ofembolic tumor cells (204).
1952 Irving Zeidman showed the transpulmonary passage of tumor cell emboli resulting in metastases on thearterial side (205).
1952 Balduin Lucke studied the organ specificity of tumor growth following intravenous injection of tumorcells and comparing growth in the liver and lung (206).
1955 Eva Kline showed that cells adapted to ascites growth preexist within the parent tumor and have anincreased malignant phenotype (207).
1962 Gabriel and Tatiana Gasic showed that enzymatic manipulation of cell surfaces can affect metastatic potential (208). 1965 Bernard and Edwin Fisher used 51chromium-labeled tumor cells in quantitative dissemination studies (209). 1966 Bernard and Edwin Fisher distinguished lymphatic from hematogenous metastasis (210). 1970 Isaiah Fidler reported that metastasis can result from the survival of only a few tumor cells usingIUdR-labeled tumor cells (22).
1973 Isaiah Fidler reported the in vivo selection of tumor cells for enhanced metastatic potential (116). 1973 Beppino Giovanella and coworkers showed that human tumors can metastasize in thymic-deficient nude mice (211). 1975 Irwin Bross proposed the metastatic cascade to define a number of sequential events needed fordisseminated cancer (212).
1975 Garth Nicolson proposed that the organ specificity of metastasis is determined by cell adhesion (213). 1976 Peter Nowell proposed the clonal evolution of tumor-cell population (140). 1976 Lance Liotta and Jerome Kleinerman linked invasion and metastasis to the production of proteolyticenzymes by metastatic cells (214).
1977 Isaiah Fidler and Margaret Kripke showed the metastatic heterogeneity of neoplasms (10). 1980 Lance Liotta showed metastatic potential is correlated with enzymatic degradation of basementmembrane collagen (215).
1980 Ian Hart and Isaiah Fidler showed the organ specificity of metastasis using ectopic organs (20). 1980 James Talmadge and collaborators showed the regulatory role of natural killer cells in tumor metastasis (216). 1981 George Poste and Isaiah Fidler reported that interactions between clonal subpopulations couldstabilize their metastatic properties, whereas a clonal subpopulation is unstable, which, following a short coculture,results in the emergence of metastatic variants (13).
1982
Karl Hellmann designed first antimetastasis drug clinical trial (Razoxane; 217). 1982 James Talmadge and Sandra Wolman showed that cancer metastases are clonal and that metastases originatefrom a single surviving cell (148).
1982 James Talmadge and Isaiah Fidler provided direct evidence that the metastatic process was selective usingspontaneous metastases from multiple metastatic variants (25).
1982 George Poste and Irving Zeidman showed the rapid development of metastatic heterogeneity by clonalorigin metastases (150).
1982 Garth Nicolson and Susan Custead showed that metastasis is not due to adaption to a new organ environmentbut rather is a selective process (134).
1984 David Tarin and colleagues reported the evidence of organ-specific metastasis in ovarian carcinoma patientswho were treated with peritoneovenous shunts (47).
1984 James Kozlowski and coworkers showed metastatic heterogeneity of human tumors using immuneincompetent mice (14).
1984 Avraham Raz and coworkers showed tumor heterogeneity for motility and adhesive properties and theirassociation with metastasis (218).
(Continued on the following page)
70(14) July 15, 2010 Cancer Research
on June 3, 2020. © 2010 American Association for Cancer Research. cancerres.aacrjournals.org ed from

Cancer Metastasis
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
metastases and concluded that there were limited but statis-tically significant differences between the primary tumorsand their lymphatic metastases. This study (8) also suggestedthat the genetic program for metastasis developed over time,although occasionally it occurred early during tumor pro-gression, ultimately resulting in heterogeneity both withinand between metastases.The identification of microarray profiles with clinical rel-
evance has supported the development of prognostic signa-tures for classifying tumors into risk groups and groupstargeted for differential treatment approaches or no treat-ment. In breast cancer this approach has been undertakenby multiple investigators with disparate results. The re-search strategy used a retrospective analysis of sampleswith follow-up, selection of discriminator genes, and the de-velopment of a predictive multigene classifier (187). A sta-tistical analysis from the early studies suggested flaws inthis experimental design (188). In retrospect two data setsare needed: one to develop the classifier (training set), andanother to test the classifier (test or validation set). Thesetwo sets are obtained by splitting the original data set, if it
www.aacrjournals.org
on June 3, 202cancerres.aacrjournals.org Downloaded from
is large enough. Further, a validation using a third indepen-dent set is desirable. With this approach multiple genomic-prognostic classifiers for breast cancer have been developed,although the overlap of genes in these signatures has beenlow. Nonetheless, the primary determinants of all the signa-tures are proliferation, ER-status, HER2-status, and, lessprominently, angiogenesis, invasiveness, and apoptosis.Thus, it is likely that these signatures detect the same bio-logical processes and pathways involved in metastasis.
Cancer Stem Cells
It has been more than a century since Cohnheim (189) pro-posed the “embryonic theory” of cancer that postulated thathuman tumors arise from embryonic cells that persevere in tis-sues without reachingmaturity. This theory has developed intothe CSC hypothesis, which states that cancers develop from asubset of malignant cells that possess stem cell characteristics,such that tumors have rare cells with infinite growth potential(190). It is noted that this characteristic of infinite growth po-tential is also the definition of a tumor cell. CSCs are suggested
Table 2. Metastasis timeline (Cont'd)
Year
Discovery1985
Isaiah Fidler addressed the critical role of macrophages in the process of metastasis (219). 1986 Leonard Weiss and coworkers identified the concept of metastatic inefficiency (220). 1988 Patricia Steeg and collaborators identified the first metastasis suppressor gene (221). 1990 Richard Wahl and coworkers showed the ability of 2-deoxy-2[18F]fluoro-D-glucose (FDG) to detect lymphnode metastases with positron emission tomography (PET) scans (222).
1990 Lloyd Culp and collaborators documented the utility of bacterial LacZ as a marker to detect micrometastasesduring tumor progression (223).
1991 Judah Folkman and coworkers showed a relationship between metastasis and angiogenesis in patients withbreast cancer (200).
1992 Jo Van Damme reported the role of chemokines in tumor-associated macrophage facilitation of metastasis (225). 1994 Judah Folkman showed that the removal of a malignant primary tumor in mice spurs the growth of remotetumors, or metastases (226).
1997 Robert Hoffman and colleagues demonstrated visualization of tumor cell invasion and metastasis using greenfluorescent protein expression (227).
2000 David Botstein showed diversity in the gene expression patterns by breast cancer tissues (228). 2001 Irving Weissman and coworkers suggested the role of CSCs in metastasis (229). 2002 Rene Bernards and Robert Weinberg proposed that metastatic potential is determined early in tumorigenesis,explaining the apparent metastatic molecular signature by most cells in a primary tumor (230).
2002 Jean Paul Thiery and Robert Weinberg proposed that EMT could explain metastatic progression (171, 230). 2002 Christopher Klein and collaborators reported the genetic heterogeneity of single disseminated tumors cells inminimal residual cancer (231).
2002 Laura Van 't Veer and coworkers reported that a specific gene expression profile of primary breast cancerswas associated with the development of metastasis and a poor clinical outcome (184).
2003 Yibin Kang and colleagues identified a molecular signature associated with metastasis of breast tumors to bone (75). 2003 Michael Clarke and Max Wicha suggest that the ability of breast cancer tumors to metastasize resides injust a few “breast CSCs” that are highly resistant to chemotherapy (232).
2006 Kent Hunter emphasized the role of genetic susceptibility for the metastatic process (233). 2007 Li Ma and Robert Weinberg identified the first metastasis-promoting micro-RNA (234). 2008 William Harbor and coworkers showed that micro-RNA expression patterns could predict metastatic risk (235). 2008 Joan Massagué and coworkers reported that micro-RNAs can suppress metastases (236).Cancer Res; 70(14) July 15, 2010 5663
0. © 2010 American Association for Cancer Research.

Talmadge and Fidler
5664
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
to express characteristics of both stem cells and cancer cells,and have properties of self-renewal, asymmetric cell division,resistance to apoptosis, independent growth, tumorigenicity,andmetastatic potential. The CSC theory suggests that tumorsinclude a population of asymmetrically dividing CSCs thatgive rise to rapidly expanding progenitor cells, which eventu-ally differentiate and exhaust their proliferative potential.The stem cells retain their original phenotype and proliferativecompetence, which is referred to as “self-renewal” capacity.Cell populations enriched for CSCs are hypothesized to displayincreased tumorigenic potential in serial transplantationassays, as compared with the bulk of tumor cells.The first definitive evidence of a CSC was with acute mye-
loid leukemia (AML) by Bonnet and Dick in 1997 (191). Thisstudy used the cellular hierarchy observed for hematopoieticcells as a model to describe the development of AML from aprimitive progenitor cancer cell, now known as the CSC. Thetumor initiating cells were all found to have a distinct surfacephenotype (CD34+/CD38−), irrespective of AML subtypes and/or the degree of heterogeneity in the tumor from which theywere isolated (191). A CSC origin has now been shown for arange of “liquid tumors” (192), including B- and T-acute lym-phoblastic leukemia (B-ALL and T-ALL, respectively) and ar-guably with a variety of solid tumors (193).Given the similarities between normal and CSCs it has been
speculated that CSCs originate from normal self-renewingstem cells that have accumulated oncogenic mutations.However, it is suggested that mutations in progenitor or dif-ferentiated cells induce dedifferentiation and self-renewal ca-pacity (194, 195). The self-renewal capacity and multipotencyof CSCs is suggested to provide a mechanism for tumor re-currence, frequently observed after surgical excision of theprimary tumor, but presumably not the CSCs. As discussedabove, tumors are heterogeneous such that not all tumorcells possess the same phenotype including metastatic po-tential. The CSC theory implies that if CSCs are the only sub-set of cells capable of initiating new tumor growth, thenCSCs must be involved in the metastatic process. Therefore,it is posited that as a CSC possesses tumorigenic, invasive,and migratory characteristics it will also have all of the nec-essary attributes to result in a metastasis. However, many re-ports on CSCs are contradictory, varying in how they shouldbe identified, their characteristics, and correlations betweenclinical outcome and tumor CSC status (196). These contra-dictory observations have made the CSC hypothesis highlycontentious, including the finding that CSCs are tumor cellswith metastatic properties and that the cells within the me-tastatic lesion may then lose the metastatic phenotype; thehypotheses are not well supported clinically. Further, it isnot clear whether CSCs are “true” stem cells or represent ahighly malignant cellular subpopulation. The recent publica-tion by Quintana and colleagues showed that by optimizingxenotransplantation procedures, one can increase the fre-quency of tumor-initiating human melanoma cells (197). Thisfinding suggests that, at least in some human cancers,tumor-initiating cells are frequent, and the question is raisedof whether the CSC concept applies to solid tumors. Thus,the link between CSCs and metastasis is circumstantial,
Cancer Res; 70(14) July 15, 2010
on June 3, 202cancerres.aacrjournals.org Downloaded from
based on correlations between the frequency of cells requiredto form a metastasis, cellular phenotypes, and the expressionof genes associated with “stem cells” or EMT (198). Indeed,the concept that the rare metastasis-forming cells found intumors correspond to the equally rare CSCs seems to repre-sent a new nomenclature for a concept discussed in the1970s by Fidler and coworkers. Both selected cells and CSCshave the potential to form a secondary tumor, with a similarhistological architecture as tumor cells in the primary tumor(199). Further, both stem cells and metastatic cells are criti-cally reliant on the microenvironment (“metastatic niche”)that they reside in. As Kaplan and colleagues have discussedsuch a niche is required for the formation of metastases(157), and the need to change an organ microenvironmentinto an appropriate niche may contribute to the latency(and potentially failure) of a disseminated tumor cell to growinto a gross metastasis (190).
Summary and Future Directions
Once a diagnosis of a primary cancer is established, theurgent question is whether it is localized or metastasized.Despite improvements in diagnosis, surgical techniques, gen-eral patient care, and local and systemic adjuvant therapies,most deaths from cancer result from metastases that are re-sistant to conventional therapies. The organ microenviron-ment, in addition to facilitating metastasis, can also modifythe response of metastatic tumor cells to therapy and reducethe effectiveness of anticancer agents. In addition, metastaticcells are genetically unstable with diverse karyotypes, growthrates, cell-surface properties, antigenicities, immunogenici-ties, marker enzymes, and sensitivity to various therapeuticagents resulting in biological heterogeneity. The process ofcancer metastasis is sequential and selective and incorpo-rates stochastic elements. Thus, the growth of metastases re-presents the endpoint of many events that few tumor cellssurvive. Primary tumors consist of multiple subpopulationsof cells with invasive and metastatic properties, with the po-tential to form a metastasis in a process dependent on theinterplay of tumor cells with host factors. The finding thatdifferent metastases can originate from multiple progenitorcells contributes to the biological diversity and provides ad-ditional challenges to therapeutic intervention. Further, evenwithin a solitary metastasis of clonal origin, heterogeneitycan rapidly develop. Thus, it is critical to improve our under-standing of metastatic cell characteristics that will allow usto target them for therapeutic intervention. Clearly, the path-ogenesis of metastasis depends on multiple interactions be-tween metastatic cells and host homeostatic mechanisms(Table 2). We posit that the interruption of these interactionswill inhibit or help eradicate metastasis. Clinical efforts havefocused on the inhibition or destruction of tumor cells; how-ever, strategies to treat metastatic tumor cells and modulatethe host microenvironment now offer new treatment ap-proaches. The recent advances in our understanding of themetastatic process at the cellular and molecular level provideunprecedented potential for improvement and the develop-ment of effective adjuvant therapies.
Cancer Research
0. © 2010 American Association for Cancer Research.

Cancer Metastasis
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
The authors thank Jill Hallgren, Alice Cole, and Lola López for their expertassistance in the preparation of this manuscript.
www.aacrjournals.org
on June 3, 202cancerres.aacrjournals.org Downloaded from
Grant Support
Nebraska Research Initiative Grant “Translation of Biotechnology into theClinic” (J. E. Talmadge), Avon-National Cancer Institute Patient AwardCA036727 (J. E. Talmadge), Cancer Center Support Core Grant CA16672, andGrant 1U54CA143837 (I. J. Fidler) from the National Cancer Institute, NationalInstitutes of Health.
Received 09/29/2009; revised 04/12/2010; accepted 04/25/2010; publishedOnlineFirst 07/06/2010.
References
1. Recamier JC. Recherches sur le traitement du cancer sur la com-pression methodique simple ou combinee et sur l'histoire generalede la meme maladie. 2nd ed. 1829.
2. Fidler IJ. The pathogenesis of cancer metastasis: the ‘seed and soil’hypothesis revisited. Nat Rev Cancer 2003;3:453–8.
3. Dorland WA. Dorland's illustrated medical dictionary. 24th ed.Philadelphia (PA): Saunders Co.; 1965.
4. Del Monte U. Does the cell number 10(9) still really fit one gram oftumor tissue? Cell Cycle 2009;8:505–6.
5. van de Vijver MJ, He YD, van't Veer LJ, et al. A gene-expressionsignature as a predictor of survival in breast cancer. N EnglJ Med 2002;347:1999–2009.
6. Weigelt B, Glas AM, Wessels LF, et al. Gene expression profiles ofprimary breast tumors maintained in distant metastases. Proc NatlAcad Sci U S A 2003;100:15901–5.
7. Ramaswamy S, Ross KN, Lander ES, Golub TR. A molecular sig-nature of metastasis in primary solid tumors. Nat Genet 2003;33:49–54.
8. Suzuki M, Tarin D. Gene expression profiling of human lymph nodemetastases and matched primary breast carcinomas: clinical impli-cations. Mol Oncol 2007;1:172–80.
9. Urquidi V, Sloan D, Kawai K, et al. Contrasting expression of throm-bospondin-1 and osteopontin correlates with absence or presenceof metastatic phenotype in an isogenic model of spontaneous hu-man breast cancer metastasis. Clin Cancer Res 2002;8:61–74.
10. Fidler IJ, Kripke ML. Metastasis results from preexisting variant cellswithin a malignant tumor. Science 1977;197:893–5.
11. Kripke ML, Gruys E, Fidler IJ. Metastatic heterogeneity of cells froman ultraviolet light-induced murine fibrosarcoma of recent origin.Cancer Res 1978;38:2962–7.
12. Nicolson GL, Brunson KW, Fidler IJ. Specificity of arrest, survival,and growth of selected metastatic variant cell lines. Cancer Res1978;38:4105–11.
13. Poste G, Doll J, Fidler IJ. Interactions among clonal subpopulationsaffect stability of the metastatic phenotype in polyclonal popula-tions of B16 melanoma cells. Proc Natl Acad Sci U S A 1981;78:6226–30.
14. Kozlowski JM, Hart IR, Fidler IJ, Hanna N. A human melanoma lineheterogeneous with respect to metastatic capacity in athymic nudemice. J Natl Cancer Inst 1984;72:913–7.
15. Montel V, Huang TY, Mose E, Pestonjamasp K, Tarin D. Expressionprofiling of primary tumors and matched lymphatic and lung metas-tases in a xenogeneic breast cancer model. Am J Pathol 2005;166:1565–79.
16. Poste G, Fidler IJ. The pathogenesis of cancer metastasis. Nature1980;283:139–46.
17. Paget S. The distribution of secondary growths in cancer of thebreast. Lancet 1889;133:571–3.
18. Virchow R. Cellular pathologie, 1858. Nutr Rev 1989;47:23–5.19. Ewing J. Neoplastic diseases: a treatise on tumors. 3rd ed.
Philadelphia (PA): WB Saunders; 1928.20. Hart IR, Fidler IJ. Role of organ selectivity in the determination of me-
tastatic patterns of B16 melanoma. Cancer Res 1980;40:2281–7.21. Fidler IJ, Talmadge JE. Evidence that intravenously derived murine
pulmonary melanoma metastases can originate from the expansionof a single tumor cell. Cancer Res 1986;46:5167–71.
22. Fidler IJ. Metastasis: guantitative analysis of distribution and fate of
tumor embolilabeled with 125I-5-iodo-2'-deoxyuridine. J Natl Can-cer Inst 1970;45:773–82.
23. Talmadge JE, Fidler IJ. Cancer metastasis is selective or randomdepending on the parent tumor population. Nature 1978;27:593–4.
24. Raz A, Hanna N, Fidler IJ. In vivo isolation of a metastatic tumor cellvariant involving selective and nonadaptive processes. J Natl Can-cer Inst 1981;66:183–9.
25. Talmadge JE, Fidler IJ. Enhanced metastatic potential of tumorcells harvested from spontaneous metastases of heterogeneousmurine tumors. J Natl Cancer Inst 1982;69:975–80.
26. Fidler IJ. Tumor heterogeneity and the biology of cancer invasionand metastasis. Cancer Res 1978;38:2651–60.
27. Langley RR, Fidler IJ. Tumor cell-organ microenvironment interac-tions in the pathogenesis of cancer metastasis. Endocr Rev 2007;28:297–321.
28. Price JE, Aukerman SL, Fidler IJ. Evidence that the process of mu-rine melanoma metastasis is sequential and selective and containsstochastic elements. Cancer Res 1986;46:5172–8.
29. Willis RA. The spread of tumors in the human body. London: Butter-worths and Company; 1952. pp. 1–447.
30. Fidler IJ, Gersten DM, Hart IR. The biology of cancer invasion andmetastasis. Adv Cancer Res 1978;28:149–250.
31. Nicolson GL. Cancer metastasis: tumor cell and host organ proper-ties important in metastasis to specific secondary sites. BiochimBiophys Acta 1988;948:175–224.
32. Naito S, Giavazzi R, Fidler IJ. Correlation between the in vitro inter-action of tumor cells with an organ environment and metastatic be-havior in vivo. Invasion Metastasis 1987;7:16–29.
33. Price JE, Naito S, Fidler IJ. Growth in an organ microenvironmentas a selective process in metastasis. Clin Exp Metastasis 1988;6:91–102.
34. Sugarbaker EV. Cancer metastasis: a product of tumor-host inter-actions. Curr Probl Cancer 1979;3:1–59.
35. Weiss L. Principles of metastasis. Orlando (FL): Academic Press;1985.
36. Fidler IJ, Kripke ML. Genomic analysis of primary tumors does notaddress the prevalence of metastatic cells in the population. NatGenet 2003;34:23.
37. Chambers AF, Groom AC, MacDonald IC. Dissemination andgrowth of cancer cells in metastatic sites. Nat Rev Cancer 2002;2:563–72.
38. Weiss L. Metastatic inefficiency: causes and consequences. Can-cer Rev 1986;3:1–24.
39. Chen LL, Blumm N, Christakis NA, Barabasi AL, Deisboeck TS.Cancer metastasis networks and the prediction of progression pat-terns. Br J Cancer 2009;101:749–58.
40. Fidler IJ. Biological behavior of malignant melanoma cells correlat-ed to their survival in vivo. Cancer Res 1975;35:218–24.
41. Chiang AC, Massague J. Molecular basis of metastasis. N EnglJ Med 2008;359:2814–23.
42. Waghorne C, Thomas M, Lagarde A, Kerbel RS, Breitman ML. Ge-netic evidence for progressive selection and overgrowth of primarytumors by metastatic cell subpopulations. Cancer Res 1988;48:6109–14.
43. Wang K, Li J, Li S, Bolund L, Wiuf C. Estimation of tumor hetero-geneity using CGH array data. BMC Bioinformatics 2009;10:12.
44. Kuukasjarvi T, Karhu R, Tanner M, et al. Genetic heterogeneity and
Cancer Res; 70(14) July 15, 2010 5665
0. © 2010 American Association for Cancer Research.

Talmadge and Fidler
5666
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
clonal evolution underlying development of asynchronous metasta-sis in human breast cancer. Cancer Res 1997;57:1597–604.
45. Bissig H, Richter J, Desper R, et al. Evaluation of the clonal re-lationship between primary and metastatic renal cell carcinomaby comparative genomic hybridization. Am J Pathol 1999;155:267–74.
46. Butler TP, Gullino PM. Quantitation of cell shedding into efferentblood of mammary adenocarcinoma. Cancer Res 1975;35:512–6.
47. Tarin D, Price JE, Kettlewell MG, et al. Mechanisms of human tumormetastasis studied in patients with peritoneovenous shunts. CancerRes 1984;44:3584–92.
48. Kasimir-Bauer S. Circulating tumor cells as markers for cancer riskassessment and treatment monitoring. Mol Diagn Ther 2009;13:209–15.
49. Arnerlöv C, Emdin SO, Lundgren B, et al. Breast carcinoma growthrate described by mammographic doubling time and S-phase frac-tion. Correlations to clinical and histopathologic factors in ascreened population. Cancer 1992;70:1928–34.
50. Shackney SE, McCormack GW, Cuchural GJ, Jr. Growth rate pat-terns of solid tumors and their relation to responsiveness to thera-py: an analytical review. Ann Intern Med 1978;89:107–21.
51. von Fournier D, Weber E, Hoeffken W, et al. Growth rate of 147mammary carcinomas. Cancer 1980;45:2198–207.
52. Peer PG, van Dijck JA, Hendriks JH, Holland R, Verbeek AL.Age-dependent growth rate of primary breast cancer. Cancer1993;71:3547–51.
53. Fidler IJ, Balch CM. The biology of cancer metastasis and implica-tions for therapy. Curr Probl Surg 1987;24:129–209.
54. Engel J, Eckel R, Kerr J, et al. The process of metastasisation forbreast cancer. Eur J Cancer 2003;39:1794–806.
55. Jones S, Chen WD, Parmigiani G, et al. Comparative lesion se-quencing provides insights into tumor evolution. Proc Natl AcadSci U S A 2008;105:4283–8.
56. Robinson SN, Chavez JM, Blonder JM, et al. Hematopoietic pro-genitor cell mobilization in mice by sustained delivery of granulo-cyte colony-stimulating factor. J Interferon Cytokine Res 2005;25:490–500.
57. Muto T, Bussey HJ, Morson BC. The evolution of cancer of the co-lon and rectum. Cancer 1975;36:2251–70.
58. Staab HJ, Anderer FA, Hornung A, Stumpf E, Fischer R. Doublingtime of circulating cea and its relation to survival of patients withrecurrent colorectal cancer. Br J Cancer 1982;46:773–81.
59. Price JE, Tarin D, Fidler IJ. Influence of organ microenvironment onpigmentation of a metastatic murine melanoma. Cancer Res 1988;48:2258–64.
60. Schackert G, Fidler IJ. Site-specific metastasis of mouse melano-mas and a fibrosarcoma in the brain or meninges of syngeneic ani-mals. Cancer Res 1988;48:3478–84.
61. Nicolson GL. Organ specificity of tumor metastasis: role of prefer-ential adhesion, invasion and growth of malignant cells at specificsecondary sites. Cancer Metastasis Rev 1988;7:143–88.
62. Fidler IJ. The organ microenvironment and cancer metastasis.Differentiation 2002;70:498–505.
63. Simone NL, Bonner RF, Gillespie JW, Emmert-Buck MR, Liotta LA.Laser-capture microdissection: opening the microscopic frontier tomolecular analysis. Trends Genet 1998;14:272–6.
64. Fidler IJ. Critical factors in the biology of human cancer metastasis:twenty-eighth G.H.A. Clowes memorial award lecture. Cancer Res1990;50:6130–8.
65. Pasqualini R, Ruoslahti E. Organ targeting in vivo using phage dis-play peptide libraries. Nature 1996;380:364–6.
66. Uehara H, Kim SJ, Karashima T, et al. Effects of blocking platelet-derived growth factor-receptor signaling in a mouse model of ex-perimental prostate cancer bone metastases. J Natl Cancer Inst2003;95:458–70.
67. Fidler IJ. Critical determinants of metastasis. Semin Cancer Biol2002;12:89–96.
68. Paget S. The distribution of secondary growths in cancer of thebreast. 1889. Cancer Metastasis Rev 1989;8:98–101.
69. Fuchs E. In: Braumueller W, editor. Metastasenbildung. Wien:Sarcom des Uvealtractus; 1882, p. 197–206.
Cancer Res; 70(14) July 15, 2010
on June 3, 202cancerres.aacrjournals.org Downloaded from
70. Ruoslahti E. Vascular zip codes in angiogenesis and metastasis.Biochem Soc Trans 2004;32:397–402.
71. Muller A, Homey B, Soto H, et al. Involvement of chemokine recep-tors in breast cancer metastasis. Nature 2001;410:50–6.
72. Huang EH, Singh B, Cristofanilli M, et al. A CXCR4 antagonistCTCE-9908 inhibits primary tumor growth and metastasis of breastcancer. J Surg Res 2009;155:231–6.
73. Zeelenberg IS, Ruuls-Van SL, Roos E. The chemokine receptorCXCR4 is required for outgrowth of colon carcinoma micrometas-tases. Cancer Res 2003;63:3833–9.
74. Andrea S, Emanuela M, Chiara C, et al. CXCL12, CXCR4 andCXCR7 expression in brain metastases. Cancer Biol Ther 2009;8:1608–14.
75. Kang Y, Siegel PM, Shu W, et al. A multigenic program medi-ating breast cancer metastasis to bone. Cancer Cell 2003;3:537–49.
76. Minn AJ, Gupta GP, Siegel PM, et al. Genes that mediate breastcancer metastasis to lung. Nature 2005;436:518–24.
77. Talmadge JE, Singh RK, Fidler IJ, Raz A. Murine models to evaluatenovel and conventional therapeutic strategies for cancer. AmJ Pathol 2007;170:793–804.
78. Sone S, Fidler IJ. Activation of rat alveolar macrophages to the tu-moricidal state in the presence of progressively growing pulmonarymetastases. Cancer Res 1981;41:2401–6.
79. Eccles SA, Alexander P. Macrophage content of tumours in relationto metastatic spread and host immune reaction. Nature 1974;250:667–9.
80. Gauci CL, Alexander P. The macrophage content of some humantumors. Cancer Lett 1975;1:29–32.
81. Evans R, Lawler EM. Macrophage content and immunogenicity ofC57BL/6J and BALB/cByJ methylcholanthrene-induced sarcomas.Int J Cancer 1980;26:831–5.
82. Key M, Talmadge JE, Fidler IJ. Lack of correlation between theprogressive growth of spontaneous metastases and their con-tent of infiltrating macrophages. J Reticuloendothel Soc 1982;32:387–96.
83. Kripke ML, Fisher MS. Immunologic parameters of ultraviolet carci-nogenesis. J Natl Cancer Inst 1976;57:211–5.
84. Pross HF, Kerbel RS. An assessment of intratumor phagocytic andsurface marker-bearing cells in a series of autochthonous and earlypassaged chemically induced murine sarcomas. J Natl Cancer Inst1976;57:1157–67.
85. Watanabe T, Dave B, Heimann DG, et al. GM-CSF-mobilized pe-ripheral blood CD34+ cells differ from steady- state bone marrowCD34+ cells in adhesion molecule expression. Bone Marrow Trans-plant 1997;19:1175–81.
86. Ross J, Auger M. The biology of the macrophage. In: Burke B,Lewis C, editors. The macrophage. 2nd ed. Oxford: UniversityPress; 2002.
87. Mantovani A. Effects on in vitro tumor growth of murine macro-phages isolated from sarcoma lines differing in immunogenicityand metastasizing capacity. Int J Cancer 1978;22:741–6.
88. Steele RJ, Eremin O, Brown M, Hawkins RA. A high macrophagecontent in human breast cancer is not associated with favourableprognostic factors. Br J Surg 1984;71:456–8.
89. Sun XF, Zhang H. Clinicopathological significance of stromal vari-ables: angiogenesis, lymphangiogenesis, inflammatory infiltration,MMP and PINCH in colorectal carcinomas. Mol Cancer 2006;5:43–63.
90. Fidler IJ, Schackert G, Zhang RD, Radinsky R, Fujimaki T. The bi-ology of melanoma brain metastasis. Cancer Metastasis Rev 1999;18:387–400.
91. Alexander P, Eccles SA, Gauci CL. The significance of macro-phages in human and experimental tumors. Ann N Y Acad Sci1976;276:124–33.
92. Donkor MK, Lahue E, Hoke TA, et al. Mammary tumor heterogene-ity in the expansion of myeloid-derived suppressor cells. Int Immu-nopharmacol 2009;9:937–48.
93. Lin EY, Nguyen AV, Russell RG, Pollard JW. Colony-stimulating fac-tor 1 promotes progression of mammary tumors to malignancy.J Exp Med 2001;193:727–40.
Cancer Research
0. © 2010 American Association for Cancer Research.

Cancer Metastasis
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
94. Hagemann T, Robinson SC, Schulz M, et al. Enhanced invasivenessof breast cancer cell lines upon co-cultivation with macrophages isdue to TNF-α dependent up-regulation of matrix metalloproteases.Carcinogenesis 2004;25:1543–9.
95. Hamada I, Kato M, Yamasaki T, et al. Clinical effects of tumor-associated macrophages and dendritic cells on renal cell carcino-ma. Anticancer Res 2002;22:4281–4.
96. Lewis C, Murdoch C. Macrophage responses to hypoxia: implica-tions for tumor progression and anti-cancer therapies. Am J Pathol2005;167:627–35.
97. Bronte V, Wang M, Overwijk WW, et al. Apoptotic death of CD8+T lymphocytes after immunization: induction of a suppressive pop-ulation of Mac-1+/Gr-1+ cells. J Immunol 1998;161:5313–20.
98. Kusmartsev S, Gabrilovich DI. STAT1 signaling regulates tumor-associated macrophage-mediated T cell deletion. J Immunol2005;174:4880–91.
99. Mirza N, Fishman M, Fricke I, et al. All-trans-retinoic acid improvesdifferentiation of myeloid cells and immune response in cancer pa-tients. Cancer Res 2006;66:9299–307.
100. Almand B, Clark JI, Nikitina E, et al. Increased production of imma-ture myeloid cells in cancer patients: a mechanism of immunosup-pression in cancer. J Immunol 2001;166:678–89.
101. Almand B, Resser JR, Lindman B, et al. Clinical significance of de-fective dendritic cell differentiation in cancer. Clin Cancer Res 2000;6:1755–66.
102. Strober S. Natural suppressor (NS) cells, neonatal tolerance, andtotal lymphoid irradiation: exploring obscure relationships. AnnuRev Immunol 1984;2:219–37.
103. Badger AM, King AG, Talmadge JE, et al. Induction of non-specificsuppressor cells in normal Lewis rats by a novel azaspirane SK&F105685. J Autoimmun 1990;3:485–500.
104. Bronte V, Apolloni E, Cabrelle A, et al. Identification of a CD11b(+)/Gr-1(+)/CD31(+) myeloid progenitor capable of activating or sup-pressing CD8(+) T cells. Blood 2000;96:3838–46.
105. Young MR, Lathers DM. Myeloid progenitor cells mediate immunesuppression in patients with head and neck cancers. Int J Immuno-pharmacol 1999;21:241–52.
106. Dranoff G. GM-CSF-secreting melanoma vaccines. Oncogene2003;22:3188–92.
107. Ohm JE, Carbone DP. VEGF as a mediator of tumor-associated im-munodeficiency. Immunol Res 2001;23:263–72.
108. Gabrilovich D, Ishida T, Oyama T, et al. Vascular endothelial growthfactor inhibits the development of dendritic cells and dramaticallyaffects the differentiation of multiple hematopoietic lineages in vivo.Blood 1998;92:4150–66.
109. Ellis LM, Takahashi Y, Liu W, Shaheen RM. Vascular endothelialgrowth factor in human colon cancer: biology and therapeutic im-plications. Oncologist 2000;5[Suppl 1]:11–5.
110. Toi M, Kondo S, Suzuki H, et al. Quantitative analysis of vascularendothelial growth factor in primary breast cancer. Cancer 1996;77:1101–6.
111. Gabrilovich DI, Chen HL, Girgis KR, et al. Production of vascularendothelial growth factor by human tumors inhibits the functionalmaturation of dendritic cells. Nat Med 1996;2:1096–103.
112. Saito H, Tsujitani S, Ikeguchi M, Maeta M, Kaibara N. Relationshipbetween the expression of vascular endothelial growth factorand the density of dendritic cells in gastric adenocarcinoma tissue.Br J Cancer 1998;78:1573–7.
113. Gabrilovich DI, Ishida T, Nadaf S, Ohm JE, Carbone DP. Antibodiesto vascular endothelial growth factor enhance the efficacy of cancerimmunotherapy by improving endogenous dendritic cell function.Clin Cancer Res 1999;5:2963–70.
114. Finke JH, Rini B, Ireland J, et al. Sunitinib reverses type-1 immunesuppression and decreases T-regulatory cells in renal cell carcino-ma patients. Clin Cancer Res 2008;14:6674–82.
115. Ko JS, Zea AH, Rini BI, et al. Sunitinib mediates reversal of myeloid-derived suppressor cell accumulation in renal cell carcinoma pa-tients. Clin Cancer Res 2009;15:2148–57.
116. Fidler IJ. Selection of successive tumour lines for metastasis. NatNew Biol 1973;242:148–9.
117. Nicolson GL, Dulski KM. Organ specificity of metastatic tumor
www.aacrjournals.org
on June 3, 202cancerres.aacrjournals.org Downloaded from
colonization is related to organ-selective growth properties of ma-lignant cells. Int J Cancer 1986;38:289–94.
118. Fidler IJ. General considerations for studies of experimental cancermetastasis. In: Busch H, editor. Methods of cancer research, Vol.XV. New York: New Academic Press; 1978, p 399–439.
119. Brunson KW, Nicolson GL. Selection and biologic properties of ma-lignant variants of a murine lymphosarcoma. J Natl Cancer Inst1978;61:1499–503.
120. Fidler IJ, Gersten DM, Budmen MB. Characterization in vivo andin vitro of tumor cells selected for resistance to syngeneic lym-phocyte-mediated cytotoxicity. Cancer Res 1976;36:3160–5.
121. Briles EB, Kornfeld S. Isolation and metastatic properties of detach-ment variants of B16 melanoma cells. J Natl Cancer Inst 1978;60:1217–22.
122. Hart IR. The selection and characterization of an invasive variant ofthe B16 melanoma. Am J Pathol 1979;97:587–600.
123. Poste G, Doll J, Hart IR, Fidler IJ. In vitro selection of murine B16melanoma variants with enhanced tissue-invasive properties. Can-cer Res 1980;40:1636–44.
124. Kerbel RS. Immunologic studies of membrane mutants of a highlymetastatic murine tumor. Am J Pathol 1979;97:609–22.
125. Hanna N. Role of natural killer cells in control of cancer metastasis.Cancer Metastasis Rev 1982;1:45–64.
126. Luria SE, Delbruck M. Mutations of bacteria from virus sensitivity tovirus resistance. Genetics 1943;28:491–511.
127. Kripke ML. Speculations on the role of ultraviolet radiation in thedevelopment of malignant melanoma. J Natl Cancer Inst 1979;63:541–8.
128. Fidler IJ, Kripke ML. Metastatic heterogeneity of cells from the K-1735 melanoma. In: Grundmann E, editor. Metastatic tumor growth.Stuttgart: Gustav Fischer Verlag; 1980, p. 71–81.
129. Fidler IJ, Gruys E, Cifone MA, Barnes Z, Bucana C. Demonstrationof multiple phenotypic diversity in a murine melanoma of recent or-igin. J Natl Cancer Inst 1981;67:947–56.
130. Fidler IJ, Gersten DM, Kripke ML. Influence of immune status on themetastasis of three murine fibrosarcomas of different immunogeni-cities. Cancer Res 1979;39:3816–21.
131. Fidler IJ, Kripke ML. Tumor cell antigenicity, host immunity and can-cer metastasis. Cancer Immunol Immunother 1980;7:201–5.
132. Aukerman SL, Price JE, Fidler IJ. Different deficiencies in the pre-vention of tumorigenic-low-metastatic murine K-1735b melanomacells from producing metastases. J Natl Cancer Inst 1986;77:915–24.
133. Heppner GH. Tumor heterogeneity. Cancer Res 1984;44:2259–65.134. Nicolson GL, Custead SE. Tumor metastasis is not due to adap-
tation of cells to a new organ environment. Science 1982;215:176–8.
135. Poste G. Pathogenesis of metastatic disease: implications for cur-rent therapy and for the development of new therapeutic strategies.Cancer Treat Rep 1986;70:183–99.
136. Nakamura T, Kuwai T, Kitadai Y, et al. Zonal heterogeneity for geneexpression in human pancreatic carcinoma. Cancer Res 2007;67:7597–604.
137. Bachtiary B, Boutros PC, Pintilie M, et al. Gene expression profilingin cervical cancer: an exploration of intratumor heterogeneity. ClinCancer Res 2006;12:5632–40.
138. Moroni M, Veronese S, Benvenuti S, et al. Gene copy number forepidermal growth factor receptor (EGFR) and clinical response toantiEGFR treatment in colorectal cancer: a cohort study. LancetOncol 2005;6:279–86.
139. Pettaway CA, Pathak S, Greene G, et al. Selection of highly me-tastatic variants of different human prostatic carcinomas usingorthotopic implantation in nude mice. Clin Cancer Res 1996;2:1627–36.
140. Nowell PC. The clonal evolution of tumor cell populations. Science1976;194:23–8.
141. Nowell PC. Chromosomal and molecular clues to tumor progres-sion. Semin Oncol 1989;16:116–27.
142. Cifone MA, Fidler IJ. Increasing metastatic potential is associatedwith increasing genetic instability of clones isolated from murineneoplasms. Proc Natl Acad Sci U S A 1981;78:6949–52.
Cancer Res; 70(14) July 15, 2010 5667
0. © 2010 American Association for Cancer Research.

Talmadge and Fidler
5668
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
143. Hill RP, Chambers AF, Ling V, Harris JF. Dynamic heterogeneity:rapid generation of metastatic variants in mouse B16 melanomacells. Science 1984;224:998–1001.
144. Masramon L, Vendrell E, Tarafa G, et al. Genetic instability and di-vergence of clonal populations in colon cancer cells in vitro. J CellSci 2006;119:1477–82.
145. Jones TD, Carr MD, Eble JN, et al. Clonal origin of lymph node me-tastases in bladder carcinoma. Cancer 2005;104:1901–10.
146. Welch DR, Tomasovic SP. Implications of tumor progression onclinical oncology. Clin Exp Metastasis 1985;3:151–88.
147. Poste G, Greig R. On the genesis and regulation of cellular hetero-geneity in malignant tumors. Invasion Metastasis 1982;2:137–76.
148. Talmadge JE, Wolman SR, Fidler IJ. Evidence for the clonal origin ofspontaneous metastases. Science 1982;217:361–3.
149. Talmadge JE, Zbar B. Clonality of pulmonary metastases from thebladder 6 subline of the B16 melanoma studied by southern hybrid-ization. J Natl Cancer Inst 1987;78:315–20.
150. Poste G, Tzeng J, Doll J, et al. Evolution of tumor cell heterogeneityduring progressive growth of individual lung metastases. Proc NatlAcad Sci U S A 1982;79:6574–8.
151. Ootsuyama A, Tanaka K, Tanooka H. Evidence by cellular mosai-cism for monoclonal metastasis of spontaneous mouse mammarytumors. J Natl Cancer Inst 1987;78:1223–7.
152. Talmadge C, Tanio Y, Meeker A, Talmadge J, Zbar B. Tumor cellstransfected with the neomycin resistance gene (neo) contain uniquegenetic markers useful for identification of tumor recurrence andmetastasis. Invasion Metastasis 1987;7:197–207.
153. Korczak B, Robson IB, Lamarche C, Bernstein A, Kerbel RS. Genet-ic tagging of tumor cells with retrovirus vectors: clonal analysis oftumor growth and metastasis in vivo. Mol Cell Biol 1988;8:3143–9.
154. Talmadge JE, Benedict K, Madsen J, Fidler IJ. Development of bi-ological diversity and susceptibility to chemotherapy in murine can-cer metastases. Cancer Res 1984;44:3801–5.
155. Wolman SR, McMorrow LE, Fidler IJ, Talmadge JE. Developmentand progression of karyotypic variability in melanoma K1735 follow-ing X-irradiation. Cancer Res 1985;45:1839–44.
156. Kaplan RN, Rafii S, Lyden D. Preparing the “soil”:the premetastaticniche. Cancer Res 2006;66:11089–93.
157. Kaplan RN, Riba RD, Zacharoulis S, et al. VEGFR1-positive haema-topoietic bone marrow progenitors initiate the pre-metastatic niche.Nature 2005;438:820–7.
158. Gao D, Nolan DJ, Mellick AS, et al. Endothelial progenitor cells con-trol the angiogenic switch in mouse lung metastasis. Science 2008;319:195–8.
159. Talmadge JE, Key M, Fidler IJ. Macrophage content of metastaticand nonmetastatic rodent neoplasms. J Immunol 1981;126:2245–8.
160. Petit I, Jin D, Rafii S. The SDF-1-CXCR4 signaling pathway: a mo-lecular hub modulating neo-angiogenesis. Trends Immunol 2007;28:299–307.
161. DeNardo DG, Johansson M, Coussens LM. Immune cells as media-tors of solid tumor metastasis. Cancer Metastasis Rev 2008;27:11–8.
162. Brigati C, Noonan DM, Albini A, Benelli R. Tumors and inflammatoryinfiltrates: friends or foes? Clin Exp Metastasis 2002;19:247–58.
163. ChenQ,WangWC, EvansSS. Tumormicrovasculature as a barrier toantitumor immunity. Cancer Immunol Immunother 2003;52:670–9.
164. Dvorak HF. Tumors: wounds that do not heal. Similarities betweentumor stroma generation and wound healing. N Engl J Med 1986;315:1650–9.
165. Sica A, Schioppa T, Mantovani A, Allavena P. Tumour-associatedmacrophages are a distinct M2 polarised population promoting tu-mour progression: potential targets of anti-cancer therapy. EurJ Cancer 2006;42:717–27.
166. Malmberg KJ. Effective immunotherapy against cancer: a questionof overcoming immune suppression and immune escape? CancerImmunol Immunother 2004;53:879–92.
167. Batlle E, Sancho E, Franci C, et al. The transcription factor snail is arepressor of E-cadherin gene expression in epithelial tumour cells.Nat Cell Biol 2000;2:84–9.
168. Yang J, Mani SA, Donaher JL, et al. Twist, a master regulator ofmorphogenesis, plays an essential role in tumor metastasis. Cell2004;117:927–39.
Cancer Res; 70(14) July 15, 2010
on June 3, 202cancerres.aacrjournals.org Downloaded from
169. Hartwell KA, Muir B, Reinhardt F, et al. The Spemann organizergene, Goosecoid, promotes tumor metastasis. Proc Natl AcadSci U S A 2006;103:18969–74.
170. Spaderna S, Schmalhofer O, Wahlbuhl M, et al. The transcriptionalrepressor ZEB1 promotes metastasis and loss of cell polarity incancer. Cancer Res 2008;68:537–44.
171. Thiery JP. Epithelial-mesenchymal transitions in tumour progres-sion. Nat Rev Cancer 2002;2:442–54.
172. Duband JL, Monier F, Delannet M, Newgreen D. Epithelium-mesenchyme transition during neural crest development. Acta Anat(Basel) 1995;154:63–78.
173. Stoker M, Gherardi E, Perryman M, Gray J. Scatter factor is a fibro-blast-derived modulator of epithelial cell mobility. Nature 1987;327:239–42.
174. Hay ED. An overview of epithelio-mesenchymal transformation.Acta Anat (Basel) 1995;154:8–20.
175. Greenburg G, Hay ED. Cytodifferentiation and tissue phenotypechange during transformation of embryonic lens epithelium tomesenchyme-like cells in vitro. Dev Biol 1986;115:363–79.
176. Thiery JP, Sleeman JP. Complex networks orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell Biol 2006;7:131–42.
177. Thompson EW, Newgreen DF, Tarin D. Carcinoma invasion andmetastasis: a role for epithelial-mesenchymal transition? CancerRes 2005;65:5991–5.
178. Delinassios JG, Kottaridis SD, Garas J. Uncontrolled growth oftumour stromal fibroblasts in vitro. Exp Cell Biol 1983;51:201–9.
179. Wargotz ES, Deos PH, Norris HJ. Metaplastic carcinomas of thebreast. II. Spindle cell carcinoma. Hum Pathol 1989;20:732–40.
180. Dairkee SH, Blayney C, Smith HS, Hackett AJ. Monoclonal anti-body that defines human myoepithelium. Proc Natl Acad Sci U S A1985;82:7409–13.
181. Dairkee SH, Mayall BH, Smith HS, Hackett AJ. Monoclonalmarker that predicts early recurrence of breast cancer. Lancet1987;1:514.
182. Rosivatz E, Becker I, Specht K, et al. Differential expression of theepithelial-mesenchymal transition regulators snail, SIP1, and twistin gastric cancer. Am J Pathol 2002;161:1881–91.
183. Tarin D, Thompson EW, Newgreen DF. The fallacy of epithelialmesenchymal transition in neoplasia. Cancer Res 2005;65:5996–6000.
184. van 't Veer LJ, Dai H, van de Vijver MJ, et al. Gene expression pro-filing predicts clinical outcome of breast cancer. Nature 2002;415:530–6.
185. Montel V, Mose ES, Tarin D. Tumor-stromal interactions reciprocal-ly modulate gene expression patterns during carcinogenesis andmetastasis. Int J Cancer 2006;119:251–63.
186. Gancberg D, Di LA, Cardoso F, et al. Comparison of HER-2 statusbetween primary breast cancer and corresponding distant meta-static sites. Ann Oncol 2002;13:1036–43.
187. Pfeffer U, Romeo F, Noonan DM, Albini A. Prediction of breast can-cer metastasis by genomic profiling: where do we stand? Clin ExpMetastasis 2009;26:547–58.
188. Dupuy A, Simon RM. Critical review of published microarray studiesfor cancer outcome and guidelines on statistical analysis and re-porting. J Natl Cancer Inst 2007;99:147–57.
189. Cohnheim J. Ueber Entzundung und Eiterung. Path Anat PhysioKlin Med 1867;40:1–79.
190. Li F, Tiede B, Massagué J, Kang Y. Beyond tumorigenesis: cancerstem cells in metastasis. Cell Res 2007;17:3–14.
191. Bonnet D, Dick JE. Human acute myeloid leukemia is organized asa hierarchy that originates from a primitive hematopoietic cell. NatMed 1997;3:730–7.
192. Cox CV, Martin HM, Kearns PR, et al. Characterization of a progen-itor cell population in childhood T-cell acute lymphoblastic leuke-mia. Blood 2007;109:674–82.
193. Hermann PC, Huber SL, Herrler T, et al. Distinct populations of can-cer stem cells determine tumor growth and metastatic activity inhuman pancreatic cancer. Cell Stem Cell 2007;1:313–23.
194. Pardal R, Clarke MF, Morrison SJ. Applying the principles of stem-cell biology to cancer. Nat Rev Cancer 2003;3:895–902.
195. Bachoo RM, Maher EA, Ligon KL, et al. Epidermal growth factor
Cancer Research
0. © 2010 American Association for Cancer Research.

Cancer Metastasis
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040
receptor and Ink4a/Arf: convergent mechanisms governing terminaldifferentiation and transformation along the neural stem cell to as-trocyte axis. Cancer Cell 2002;1:269–77.
196. Kelly PN, Dakic A, Adams JM, Nutt SL, Strasser A. Tumor growthneed not be driven by rare cancer stem cells. Science 2007;317:337.
197. Quintana E, Shackleton M, Sabel MS, et al. Efficient tumour forma-tion by single human melanoma cells. Nature 2008;456:593–8.
198. Geiger TR, Peeper DS. Metastasis mechanisms. Biochim BiophysActa 2009;1796:293–308.
199. Brabletz T, Jung A, Spaderna S, Hlubek F, Kirchner T. Opinion: mi-grating cancer stem cells - an integrated concept of malignant tu-mour progression. Nat Rev Cancer 2005;5:744–9.
200. Folkman J. Angiogenesis: initiation and modulation. In: NicholsonGL, Milas L, editors. Cancer invasion and metastasis: biologicand therapeutic aspects. New York: Raven Press; 1984, p. 201–9.
201. Folkman J, Klagsbrun M. Angiogenic factors. Science 1987;235:442–7.
202. Liotta LA. Tumor invasion and metastases-role of the extracellularmatrix: Rhoads Memorial Award lecture. Cancer Res 1986;46:1–7.
203. Takahashi M. An experimental study of metastasis. J Pathol Bacter-iol 1915;20:1–13.
204. Coman DR. Decreased mutual adhesiveness, a property of cellsfrom squamous cell carcinomas. Cancer Res 1944;4:625–9.
205. Zeidman I. BUSS JM. Transpulmonary passage of tumor cell em-boli. Cancer Res 1952;12:731–3.
206. Lucke B, Breedis C, Woo ZP, Berwick L, Nowell P. Differentialgrowth of metastatic tumors in liver and lung: experiments with rab-bit V2 carcinoma. Cancer Res 1952;12:734–8.
207. Klein E. Immediate transformation of solid into ascites tumors;stud-ies on a mammary carcinoma of an inbred mouse strain. Exp CellRes 1955;8:213–25.
208. Gasic G, Gasic T. Removal of sialic acid from the cell coat in tumorcells and vascular endothelium, and its effects on metastasis. ProcNatl Acad Sci U S A 1962;48:1172–7.
209. Fisher B, Fisher ER. The organ distribution of disseminated 51Cr-labeled tumor cells. Cancer Res 1967;27:412–20.
210. Fisher B, Fisher ER. The interrelationship of hematogenous andlymphatic tumor cell dissemination. Surg Gynecol Obstet 1966;122:791–8.
211. Giovanella BC, Yim SO, Morgan AC, Stehlin JS, Williams LJ, Jr.Brief communication: metastases of human melanomas trans-planted in “nude” mice. J Natl Cancer Inst 1973;50:1051–3.
212. Bross ID, Viadana E, Pickren JW. The metastatic spread of myelo-ma and leukemias in men. Virchows Arch A Pathol Anat Histol 1975;365:91–101.
213. Nicolson GL, Winkelhake JL. Organ specificity of blood-borne tu-mour metastasis determined by cell adhesion? Nature 1975;255:230–2.
214. Liotta LA, Kleinerman J, Catanzaro P, Rynbrandt D. Degradation ofbasement membrane by murine tumor cells. J Natl Cancer Inst1977;58:1427–31.
215. Liotta LA, Tryggvason K, Garbisa S, et al. Metastatic potential cor-relates with enzymatic degradation of basement membrane colla-gen. Nature 1980;284:67–8.
216. Talmadge JE, Meyers KM, Prieur DJ, Starkey JR. Role of NK cells in
www.aacrjournals.org
on June 3, 202cancerres.aacrjournals.org Downloaded from
tumour growth and metastasis in beige mice. Nature 1980;284:622–4.
217. Herman EH, Witiak DT, Hellmann K, Waravdekar VS. Biologicalproperties of ICRF-159 and related bis(dioxopiperazine) com-pounds. Adv Pharmacol Chemother 1982;19:249–90.
218. Volk T, Geiger B, Raz A. Motility and adhesive properties of high-and low-metastatic murine neoplastic cells. Cancer Res 1984;44:811–24.
219. Fidler IJ. Macrophages and metastasis-a biological approach tocancer therapy. Cancer Res 1985;45:4714–26.
220. Weiss L, Mayhew E, Rapp DG, Holmes JC. Metastatic inefficiencyin mice bearing B16 melanomas. Br J Cancer 1982;45:44–53.
221. Steeg PS, Bevilacqua G, Kopper L, et al. Evidence for a novel geneassociated with low tumor metastatic potential. J Natl Cancer Inst1988;80:200–4.
222. Wahl RL, Kaminski MS, Ethier SP, Hutchins GD. The potential of2-deoxy-2[18F]fluoro-D-glucose (FDG) for the detection of tumor in-volvement in lymph nodes. J Nucl Med 1990;31:1831–5.
223. Lin WC, Pretlow TP, Pretlow TG, Culp LA. Bacterial lacZ gene as ahighly sensitive marker to detect micrometastasis formation duringtumor progression. Cancer Res 1990;50:2808–17.
224. Weidner N, Semple JP, Welch WR, Folkman J. Tumor angiogenesisand metastasis-correlation in invasive breast carcinoma. N EnglJ Med 1991;324:1–8.
225. Opdenakker G, Van DJ. Chemotactic factors, passive invasion andmetastasis of cancer cells. Immunol Today 1992;13:463–4.
226. O'Reilly MS, Holmgren L, Shing Y, et al. Angiostatin: a novel angio-genesis inhibitor that mediates the suppression of metastases by aLewis lung carcinoma. Cell 1994;79:315–28.
227. Chishima T, Miyagi Y, Wang X, et al. Cancer invasion and microme-tastasis visualized in live tissue by green fluorescent protein expres-sion. Cancer Res 1997;57:2042–7.
228. Perou CM, Sorlie T, Eisen MB, et al. Molecular portraits of humanbreast tumours. Nature 2000;406:747–52.
229. Reya T, Morrison SJ, Clarke MF, Weissman IL. Stem cells, cancer,and cancer stem cells. Nature 2001;414:105–11.
230. Bernards R, Weinberg RA. A progression puzzle. Nature 2002;418:823.
231. Klein CA, Blankenstein TJ, Schmidt-Kittler O, et al. Genetic hetero-geneity of single disseminated tumour cells in minimal residual can-cer. Lancet 2002;360:683–9.
232. Dontu G, Al-Hajj M, Abdallah WM, Clarke MF, Wicha MS. Stem cellsin normal breast development and breast cancer. Cell Prolif 2003;36[Suppl 1]:59–72.
233. Hunter K. Host genetics influence tumour metastasis. Nat Rev Can-cer 2006;6:141–6.
234. Ma L, Teruya-Feldstein J, Weinberg RA. Tumour invasion and me-tastasis initiated by microRNA-10b in breast cancer. Nature 2007;449:682–8.
235. Worley LA, Long MD, Onken MD, Harbour JW. Micro-RNAs asso-ciated with metastasis in uveal melanoma identified by multiplexedmicroarray profiling. Melanoma Res 2008;18:184–90.
236. Tavazoie SF, Alarcon C, Oskarsson T, et al. Endogenous humanmicroRNAs that suppress breast cancer metastasis. Nature 2008;451:147–52.
Cancer Res; 70(14) July 15, 2010 5669
0. © 2010 American Association for Cancer Research.

2010;70:5649-5669. Published OnlineFirst July 7, 2010.Cancer Res James E. Talmadge and Isaiah J. Fidler Historical PerspectiveAACR Centennial Series: The Biology of Cancer Metastasis:
Updated version
10.1158/0008-5472.CAN-10-1040doi:
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/70/14/5649.full#ref-list-1
This article cites 226 articles, 72 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/70/14/5649.full#related-urls
This article has been cited by 63 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/70/14/5649To request permission to re-use all or part of this article, use this link
on June 3, 2020. © 2010 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 7, 2010; DOI: 10.1158/0008-5472.CAN-10-1040