abc transporters cdr1p, cdr2p and cdr3p of a human pathogen candida albicans are general...
TRANSCRIPT

Research Article
ABC transporters Cdr1p, Cdr2p and Cdr3p ofa human pathogen Candida albicans are generalphospholipid translocators
Smriti1, S. Krishnamurthy1{, B.L. Dixit1{, C.M. Gupta2, S. Milewski3 and R. Prasad1*1Membrane Biology Laboratory, School of Life Sciences, Jawaharlal Nehru University, New Delhi-110067, India2Division of Membrane Biology, Central Drug Research Institute, Lucknow, India3Department of Pharmaceutical Technology and Biochemistry, Technical University of Gdansk, Gdansk, Poland
*Correspondence to:R. Prasad, Membrane BiologyLaboratory, School of LifeSciences, Jawaharlal NehruUniversity, New Delhi-110067,India.E-mail: [email protected]
{Current address:Institut fur Mikrobiologie,Heinrich-Heine-Universitat,Dusseldorf,Universitatsstrasse 1/Geb.26.12, D-40225 Dusseldorf,Germany
{Current address:Molecular Medicine BiotechII, Suite 309, University ofMassachusetts, Worcester,MA 01605, USA
Received: 23 April 2001
Accepted: 7 August 2001
Abstract
We have used fluorescent 7-nitrobenz-2-oxa-1,3-diazol-4-yl (NBD)-tagged phospholipid
analogues, NBD-PE (phosphatidylethanolamine), NBD-PC (phosphatidylcholine) and
NBD-PS (phosphatidylserine), to demonstrate that Cdr1p and its other homologues,
Cdr2p and Cdr3p, belonging to the ATP-binding cassette (ABC) superfamily behave as
general phospholipid translocators. Interestingly, CDR1 and CDR2, whose overexpression
leads to azole resistance in C. albicans, elicit in-to-out transbilayer phospholipid movement,
while CDR3, which is not involved in drug resistance, carries out-to-in translocation of
phospholipids between the two monolayers of plasma membrane. Cdr1p, Cdr2p and Cdr3p
could be further distinguished on the basis of their sensitivities to different inhibitors. For
example, the in-to-out activity associated with Cdr1p and Cdr2p is energy-dependent and
sensitive to sulphydryl blocking agents such as N-ethylmaleimide (NEM) and cytoskeleton
disrupting agent cytochalasin E, while Cdr3p-associated out-to-in activity is energy-
dependent but insensitive to NEM and cytochalasin E. We found that certain drugs, such
as fluconazole, cycloheximide and miconazole, to which Cdr1p confers resistance could
also affect in-to-out transbilayer movement of NBD-PE, while the same drugs had no
effect on Cdr3p-mediated out-to-in translocation of NBD-PE. The ineffectiveness of these
drugs to affect Cdr3p mediated out-to-in phospholipid translocation further confirms the
inherent difference in the directionality of phospholipid translocation between these pumps.
Notwithstanding the role of some of the Cdrps in drug resistance, this study clearly
demonstrates that these ABC transporters of C. albicans are phospholipid translocators
and this function could represent one of the physiological functions of such large family of
proteins. Copyright # 2002 John Wiley & Sons, Ltd.
Keywords: CDR1; ABC transporters; multidrug resistance; Candida; membrane
asymmetry; phospholipid translocators
Introduction
The opportunistic dimorphic fungus C. albicans isa major cause of infections in humans and ispredominantly rampant in immunocompromisedindividuals. C. albicans infections are treated withantifungal agents, particularly with the triazolederivative fluconazole. In order to combat theeffects of antifungals, Candida has evolved a varietyof mechanisms to acquire resistance to these drugs(Prasad, 1991; Prasad et al., 1996; Joseph-Horne
and Hollomon, 1997; White et al., 1998; Marichal,1999). The resistance to azoles in C. albicans, wasearlier thought to occur primarily through an alter-ation or an overexpression of the 14a-lanosteroldemethylase (P45014DM) involved in sterol bio-synthesis (Lamb et al., 1997; White, 1997). How-ever, the characterization of the first ATP-bindingcassette (ABC) protein Cdr1p (Candida drug resis-tance) by us (Dogra et al., 1999; Krishnamurthyet al., 1998a, b, c; Prasad et al., 1995a; Smriti et al.,1999) and the subsequent identification of other
Yeast
Yeast 2002; 19: 303–318.DOI: 10.1002 / yea.818
Copyright # 2002 John Wiley & Sons, Ltd.

efflux pumps (Balan et al., 1997; Sanglard et al.,1997, 1999; Calabrese et al., 2000; Franz et al.,1998) and their overexpression in azole-resistantclinical isolates has led to the suggestion that thesetransporters represent another mechanism involvedin multidrug resistance (MDR) scenario of C.albicans (Balan et al., 1997; Sanglard et al., 1997,1999; Gupta et al., 1998; Krishnamurthy et al.,1998; Smriti et al., 1999; Calabrese et al., 2000;Franz et al., 1998). In spite of the widespreadoccurrence of the MDR genes in C. albicans, themolecular mechanisms of their function and theregulation of their expression is yet to be under-stood (Prasad et al., 1995a, 1996). In view of thewidespread occurrence of a large ‘superfamily’ ofABC export pumps and their involvement infunctions other than in MDR, considerable atten-tion is being given to their physiological relevance(Higgins, 1992; Decottigines and Goffeau, 1997).Several mammalian ABC proteins are also medi-cally important, since mutations in correspondinggenes encoding ABC transporters cause geneticdiseases (Bauer et al., 1999). A few of the ABCproteins of mammalian cells, human MDR1 (mdr1aof mice) and MDR3 (mdr2 of mice) are alsoinvolved in the maintenance of membrane asymme-try by mediating phospholipid translocation betweenthe two monolayers of plasma membrane (VanHelvoort et al., 1996).
An asymmetric distribution of phospholipids iswell known across the plasma membrane ofnumerous cell types (Op den Kamp, 1979; Diazand Schroit, 1996; Bevers et al., 1999). In themajority of cell types phosphatidylethanolamine(PE) and phosphatidylserine (PS) are located inthe inner monolayer, whereas phosphatidylcholine(PC), sphingomyelin and glycolipids are located inthe outer monolayer of the plasma membrane(Diaz and Schroit, 1996; Bevers et al., 1999). Thisasymmetric distribution is very important for thecell, since loss of this distribution functions as acue for several physiological processes, such asplatelet activation, clearance of senescent red bloodcells and phagocytosis of apoptotic cells (Connoret al., 1994; Diaz and Schroit, 1996; Bevers et al.,1999).
In Saccharomyces cerevisiae aminophospholipidsare preferentially located in the cytoplasmic leafletof the plasma membrane (Balasubramanian andGupta, 1996; Kean et al., 1997; Dixit and Gupta,1998). Several mechanisms that could maintainmembrane lipid asymmetry in yeast cells have been
suggested. In one of the studies Cerbon andCalderon (1991) have shown that the surfacepotential could regulate in-to-out translocation ofphospholipids. Recently it has been observed thatthe phospholipid translocation between the twomonolayers could be protein mediated in yeast cells(Balasubramanian and Gupta, 1996; Tang et al.,1996; Kean et al., 1997; Dixit and Gupta, 1998).We had earlier shown that a drug extrusion pumpCDR1 of C. albicans could mediate aminopho-spholipid translocation (Dogra et al., 1999).
Among several homologues of CDR1, onlyCDR1 and CDR2 are involved in multidrugresistance and their expression is linked to fre-quently encountered azole resistance (Prasad et al.,1995a, b; Sanglard et al., 1997). CDR3 expression,on the other hand, is linked to phenotypic switching(white–opaque transition) of C. albicans, while noapparent function has been ascribed to CDR4(Balan et al., 1997; Sanglard et al., 1997; Franzet al., 1998). In this work we have used NBD-tagged phospholipid analogues to explore whetherCdrps other than Cdr1p could also mediate trans-bilayer movement of membrane phospholipids. Ourresults show that Cdr1p, Cdr2p and Cdr3p aregeneral phospholipid translocators. However, theydiffer in the direction of phospholipid movementbetween the two monolayers. It is observed thatCdr1p and Cdr2p are involved in the translocationof PE, PC and PS from the inner to the outer leafletof the plasma membrane, whereas Cdr3p mediatesinward directed translocation of PE, PC and PS.Both out-to-in and in-to-out activities could bedistinguished on the basis of their sensitivities todifferent inhibitors. In this study we demonstratethat these ABC transporters of C. albicans arephospholipid translocators and this function couldrepresent one of the physiological functions of suchlarge family of proteins.
Materials and methods
Materials
1-myristoyl-2-[6-(NBD) aminocaproyl]- phospha-tidylcholine (M-C6NBD-PC), 1-myristoyl-2-[6-(NBD) aminocaproyl]- phosphatidyletanolamine(M-C6-NBD-PE), 1-myristoyl-2-[6-(NBD) amino-caproyl]- phosphatidylserine (M-C6-NBD-PS),N-rhodamine-dioleoylphosphatidylethanolamine (N-Rh-DOPE) were obtained from Avanti Polar Lipids
304 Smriti et al.
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.

Inc. (Alabaster, AL). Dipalmitoylphosphatidylcho-line (L-DPPC) and dipalmitoylphosphatidylethano-lamine (L-DPPE) were purchased from SigmaChemical Co., USA. All media components wereeither obtained from Difco, USA, or from Himedia,India. Pfizer Ltd., Sandwich, Kent, UK, kindlyprovided fluconazole. All other chemicals and drugsused were purchased from Sigma Chemical Co.,USA.
Strains and plasmids
S. cerevisiae strain AD 12345678 (Mata, pdr1-3, his,ura3, pdr5D, pdr10D, pdr11D, pdr15D, snq2D, yor1D,ycf1D), used in this study was a kind gift from A.Goffeau and A. Decottignies, Universite Catholiquede Louvain, Belgium. The strain AD 12345678(hereafter called the AD strain) was grown asdescribed earlier (Krishnamurthy et al., 1998b;Dogra et al., 1999). AD strain is deleted in sevenABC transporter genes, i.e. PDR5, PDR10, PDR11,PDR15 (pleiotropic drug resistance), SNQ2 (sensi-tivity to nitroquinoline oxide resistance), YCF1(yeast cadmium factor gene) and YOR1 (yeast
oligomycin resistance). AD strain was transformedwith plasmid containing CDR1, CDR2 and CDR3genes by the method described elsewhere (Prasadet al., 1995a). Hereafter, transformants of ADstrain having these genes will be described asAD-CDR1, AD-CDR2 and AD-CDR3. It is perti-nent to mention that the CDR1 gene was on acentromeric vector pYEura3 (Prasad et al., 1995a);the CDR2 gene was under the control of its ownpromoter in a multicopy plasmid Yep24 and wasa kind gift from Dominique Sanglard, Institutde Microbiologie, Centre Hospitalier UniversitaireVaudois, Lausanne, Switzerland (Sanglard et al.,1997); while the CDR3 gene was under the controlof ADH promoter (alcohol dehydrogenase 1 fromADH gene) in plasmid YPB–ADH, which was akind gift from Martine Raymond, Institut deRecherches Cliniques de Montreal, Quebec, CanadaH2W 1R7 (Balan et al., 1997). All the transfor-mants were grown in 0.67% yeast nitrogen base(YNB) medium without amino acids, containing2% glucose with required supplements. Disruptantof CDR3 (CIB4) (cdr3D::hisG/cdr3D::hisG) andits parent strain CAI4 (Dura3::imm434/Dura3::imm434) were kind gifts from Martine Raymondand were grown in yeast extract peptone dex-trose (YEPD) media. The disruptant of CDR1(DSY449) (Dcdr1::hisG/Dcdr1::hisG), the double
disruptant of CDR1/CDR2 (DSY1025) (Dcdr1::hisG/Dcdr1::hisG, Dcdr2::hisG/cdr2::hisG) and theirparent strain CAF2-1 (Dura3::imm434/URA3) werekind gifts from Dominique Sanglard and weregrown in YEPD media.
Drug Resistance Assays
For filter disc assay, cells were grown in YNB mediawith required supplements. 6r105 yeast cells/mlwere mixed with molten YNB agar (y40uC) andpoured into a Petri plate. After solidifying, the filterdiscs were kept on the plate and drugs were spottedon to the discs. The following amounts of drugs(in mg) were applied on disc in 1–5 ml volume(solvents used are given in parentheses): cyclo-heximide (water), 1.0; fluconazole (methanol), 100;crystal violet (water), 5. The plates were incubatedat 30uC and after 48 h, when clear circular zonesappeared, the plates were scored for drug sensitivityby measuring the diameter of the zone of inhibitionaround the filter discs. Such filter disc assays witheach drug were repeated at least four or five times.The solvents used to solublize different drugs werealso tested and there was no inhibition of growthdue to the solvents used.
Measurement of ATPase activity
ATP hydrolysis was measured by incubating theplasma membranes prepared from AD, CDR1-,CDR2- and CDR3-expressing cells, as describedpreviously (Krishnamurthy et al., 1998a). 10 mgmembrane protein was incubated with 5 mM
Mg-ATP, 7 mM MgSO4, 10 mM NaN3 and 60 mM
Tris–HCl, pH 7.5, at 30uC. After 10 min thereaction was stopped by the addition of 1 ml stopmix containing 0.5% SDS, 2% H2SO4 and 0.5%ammonium molybdate. The released Pi was mea-sured by adding 10% ascorbic acid after a 15 minincubation at 30uC and the blue colour developedwas measured spectrophotometrically by readingabsorbance at 750 nm.
Accumulation of rhodamine 123
The accumulation of rhodamine 123 was determinedby a method described previously (Krishnamurthyet al., 1998a). Briefly, cells of mid-log phase werecentrifuged at 3000 rpm for 3 min and resuspendedin 10 mM phosphate buffer saline (PBS), pH 7.4, asa 2% cell suspension. The cells (1 ml) were incu-bated in a shaking water bath at 150 rpm and 30uC
Phospholipid translocators of Candida albicans 305
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.

and 10 mM rhodamine 123 was added. The cells wereincubated for 20 min and a 0.5 ml aliquot wastaken, centrifuged and washed twice with PBS. Thewashed cells were resuspended in 2 ml fresh PBSand fluorescence was measured spectrofluorimetri-cally (Shimadzu, RF-540, Japan) with excitationand emission wavelengths of 485 nm and 538 nm,respectively. The accumulation was expressed asarbitrary fluorescence units.
Accumulation of [3H]-b-oestradiol
The accumulation of steroid hormone [3H]-b-oestradiol was determined by a method as describedpreviously (Krishnamurthy et al., 1998b). Briefly,mid-exponential phase cells were harvested andthen cells were resuspended in fresh medium as10% suspension. An aliquot of cells was incubatedwith 2 nM b-oestradiol (3.37 TBq/mmol). Aliquotsof 0.5 ml were withdrawn after 60 min and filteredrapidly, followed by three washes with buffer (PBS,pH 7.4, containing 2% glucose). The radioactivityon the filter was measured. The results presented asnet accumulation of steroids.
Preparation of lipid vesicles
All the phospholipids were stored at x20uC andperiodically checked for their purity by thin layerchromatography. Phospholipid concentrations werequantitated by lipid phosphorus assay, as describedpreviously (Dogra et al., 1999). Donor vesiclescontaining fluorescent phospholipids were preparedby mixing desired proportions of various phospho-lipids, as described by Kean et al. (1997). Thetypical proportion used was 40 mol% NBD-labelledphospholipid, 2 mol% N-Rh-DOPE and 58 mol%L-DPPC. After mixing all these phospholipids,chloroform was evaporated under the stream ofnitrogen to make a thin film and kept overnightunder vacuum desiccation to remove moisture andsolvent traces. The desiccated phospholipid film wasthen hydrated by adding YNB without glucose withcontinuous vortexing. The hydrated membrane filmwas sonicated for 30 min under a N2 atmosphere,using a waterbath sonicator to produce vesicles.The total lipid concentration in the stock of vesicleswas 1 mM. Acceptor vesicles were prepared in asimilar manner; however, instead of NBD fluores-cent phospholipid analogues, 40 mol% L-DPPE and60 mol% L-DPPC were used (Kean et al., 1997).
Labelling of yeast cells with fluorescent lipids
Labelling of yeast cells with fluorescent lipids wasessentially carried out as described by Kean et al.(1997) with minor modifications. Briefly, mid-logphase grown S. cerevisiae cells were harvested bycentrifugation at 3000 rpm for 5 min, washed twicewith YNB and resuspended in the same (1r1010
cells/ml). YEPD-grown Candida cells were harvestedand washed twice with fresh YEPD media. Thewashed cells were resuspended in YEPD (1r1010
cells/ml). Donor vesicles to the final concentrationof 50 mM were then added to the 500 ml cellsuspension and incubated for 90 min at 0uC. Atthe indicated time intervals, the fluorescence of thecell suspension was measured spectrofluorimeteri-cally (RF-540, Shimadzu) (excitation wavelength475 nm, emission wavelength 525 nm). After 90 min,S. cerevisiae cells were washed twice with ice-coldYNB and resuspended in YNB (1r1010 cells/ml)(Candida cells were washed and resuspended inYEPD). The acceptor vesicles (50 mM) were thenadded to the 500 ml labelled cell suspension at30uC, to allow back-exchange of NBD phospholi-pid analogues from the outer monolayer of plasmamembrane to acceptor vesicles. At indicated timeintervals, aliquots of cell suspension were takenout and fluorescence was measured in the super-natant. To measure stable fluorescence, 0.2% ofTriton X-100 was added to the supernatant. Thefluorescence observed in the supernatant was con-sidered to be the index of fluorescence due to backexchange of NBD phospholipid molecules fromcells to the acceptor vesicles. For a controlexperiment, labelled cell suspension was incubatedwithout acceptor vesicles, where no increase influorescence was observed.
Fluorescamine labelling
Fluorescamine labelling of C. albicans cells wasdone essentially as described by Dogra et al. (1999).Cells were harvested in mid-log phase by centri-fuging at 3000 rpm for 5 min at 4uC and washedtwice or three times with buffer A (100 mM
potassium phosphate 5 mM EDTA, pH 7.5). Theharvested cells (0.6 g wet wt), were suspended in3 ml buffer B (100 mM potassium phosphate and600 mM KCl, pH 8.2) and kept at 4uC with gen-tle swirling. 15.6 mM fluorescamine in dehydratedDMSO were added dropwise to the cell suspensionwith constant gentle swirling. After 30 s the reac-tion was stopped by adding an equal volume of
306 Smriti et al.
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.

1 M ammonia in 600 mM KCl. The cells were cen-trifuged and washed at 4uC three or four timesuntil the colour of the dye disappeared from thesupernatant.
Lipid extraction and thin layer chromatography
Lipids were extracted as described previously(Ibrahim and Ghannoum, 1996; Koul and Prasad,1996). Solvents used in the first and second dimen-sions were chloroform :methanol : 30% ammoniumhydroxide (65 : 25 : 6 v/v) and chloroform :water :methanol : glacial acetic acid : acetone (45 : 4 : 8 : 9 :16 v/v), respectively. Resolved phospholipids andderivatized PE were scraped off the plates and theirphosphate content was estimated.
Treatment of cells with various inhibitors
The cells were labelled with donor vesicles havingNBD-PE as fluorescent phospholipid analogue,as described above. After labelling, the indicatedcells were washed and incubated with the indicatedconcentrations of inhibitors, sodium azide (azide)(10 mM); sodium orthovanadate (vanadate) (20 mM);N-ethylmaleimide (NEM) (10 mM) and cytochalasinE (40 mg/ml) for 20 min. The cells were thenincubated with acceptor vesicles and back-exchangeactivity in the supernatant having acceptor vesicleswas measured.
Effect of drugs on phospholipid translocation
The cells, labelled with NBD-PE fluorescent analo-gue as described above, were washed and incubatedwith 200 nM concentration of various drugs alongwith acceptor vesicles. After 90 min back-exchangedfluorescence was recorded in the supernatant havingacceptor vesicles, as described above.
Statistical analysis
All results were expressed as mean valuetstandarddeviation (SD). The significance of differencebetween the data pairs was calculated by analysisof variance (ANOVA) followed by Mann–Whitneytest. A p value of <0.05 was considered to reflectstatistically significant differences. A p value of>0.05 was considered not statistically significant.
Results
Transformation of CDR1, CDR2 and CDR3 in ahypersensitive S. cerevisiae host
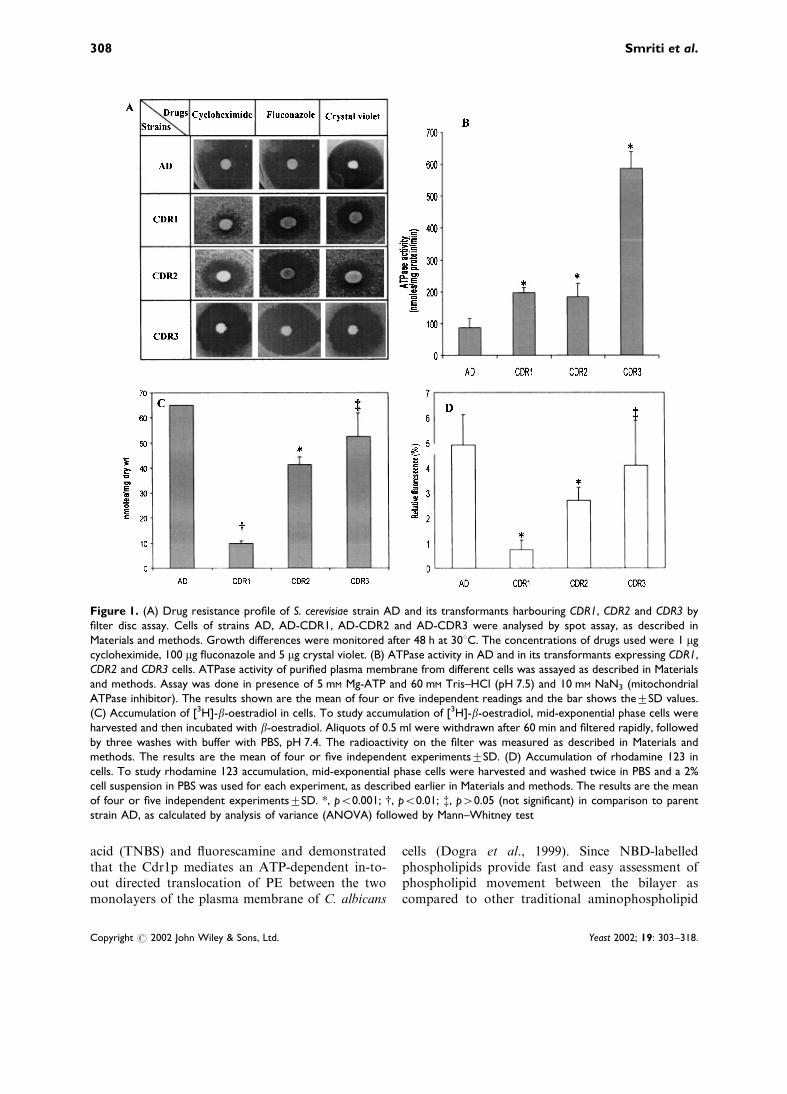
As mentioned in Materials and methods, AD cellswere transformed with genes CDR1, CDR2 andCDR3 carrying plasmids which were expressedunder different promoters. Since the objective ofthe present study was to examine the phospholipidtranslocase activities associated with such proteins,the variation in the level of activities due to variableplasmid copy number, if any, was not considered.That these proteins are functionally expressed wasapparent from the fact that CDR1- and CDR2-expressing cells conferred distinct resistance tocycloheximide, fluconazole and crystal violet ascompared to the host AD (Figure 1A). CDR3, onthe other hand, did not elicit multidrug resistancephenomena, which is consistent with earlier reportsthat it is not a drug transporter. It is evident fromFigure 1A that the inhibition zones of differentdrugs in CDR3-expressing cells were similar to itshost AD cells. In addition, the characteristicATPase activity associated with the plasma mem-brane of Cdr1p-, Cdr2p- and Cdr3p-expressing cellswas measured. As can be seen from Figure 1B,ATPase activity was much higher in the plasmamembrane fractions of cells expressing these ABCproteins. The highest activity in CDR3-expressingcells was most noteworthy (Figure 1B). Since Cdr1phas earlier been shown to transport b-oestradioland rhodamine 123 (Krishnamurthy et al., 1998a,b), the transport of these substrates was examinedto further confirm the functionality of these pro-teins. Cdr1p- and Cdr2p-expressing cells showedmuch less accumulation of b-oestradiol and rho-damine 123 as compared to parent strain AD(Figure 1C, D). In contrast, Cdr3p, which is nota multidrug transporter, also does not appear totransport b-oestradiol and rhodamine 123. Theaccumulation of these substrates by Cdr3p wasfound to be comparable to host strain AD. Takentogether, these results confirmed the functionalityof these ABC transporters when expressed in hostS. cerevisiae.
Labelling of plasma membrane of AD and itstransformants with NBD phospholipids
Earlier, we had used specific aminophospholipidlabelling reagents such as trinitrobenzenesulphonic
Phospholipid translocators of Candida albicans 307
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.
acid (TNBS) and fluorescamine and demonstratedthat the Cdr1p mediates an ATP-dependent in-to-out directed translocation of PE between the twomonolayers of the plasma membrane of C. albicans
cells (Dogra et al., 1999). Since NBD-labelledphospholipids provide fast and easy assessment ofphospholipid movement between the bilayer ascompared to other traditional aminophospholipid
Figure 1. (A) Drug resistance profile of S. cerevisiae strain AD and its transformants harbouring CDR1, CDR2 and CDR3 byfilter disc assay. Cells of strains AD, AD-CDR1, AD-CDR2 and AD-CDR3 were analysed by spot assay, as described inMaterials and methods. Growth differences were monitored after 48 h at 30uC. The concentrations of drugs used were 1 mgcycloheximide, 100 mg fluconazole and 5 mg crystal violet. (B) ATPase activity in AD and in its transformants expressing CDR1,CDR2 and CDR3 cells. ATPase activity of purified plasma membrane from different cells was assayed as described in Materialsand methods. Assay was done in presence of 5 mM Mg-ATP and 60 mM Tris–HCl (pH 7.5) and 10 mM NaN3 (mitochondrialATPase inhibitor). The results shown are the mean of four or five independent readings and the bar shows thetSD values.(C) Accumulation of [3H]-b-oestradiol in cells. To study accumulation of [3H]-b-oestradiol, mid-exponential phase cells wereharvested and then incubated with b-oestradiol. Aliquots of 0.5 ml were withdrawn after 60 min and filtered rapidly, followedby three washes with buffer with PBS, pH 7.4. The radioactivity on the filter was measured as described in Materials andmethods. The results are the mean of four or five independent experimentstSD. (D) Accumulation of rhodamine 123 incells. To study rhodamine 123 accumulation, mid-exponential phase cells were harvested and washed twice in PBS and a 2%cell suspension in PBS was used for each experiment, as described earlier in Materials and methods. The results are the meanof four or five independent experimentstSD. *, p<0.001; {, p<0.01; {, p>0.05 (not significant) in comparison to parentstrain AD, as calculated by analysis of variance (ANOVA) followed by Mann–Whitney test
308 Smriti et al.
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.

labelling reagents, in the present study we haveemployed NBD fluorescent analogues of PE, PCand PS to demonstrate the specificity and charac-teristics of phospholipid translocation mediated byCdr1p and its other homologues. The fluorescence(excitation wavelength 475 nm, emission wavelength525 nm) due to the NBD group present in NBD-phospholipids has made it possible not only toascertain the asymmetric nature of most of thephospholipids, but also to study the kineticsof phospholipid translocation (Mukherjee et al.,1994; Williamson et al., 1995; Chattopadhyay andLondon, 1988). Although NBD-phospholipids havebeen successfully used to measure out-to-in andin-to-out directed membrane phospholipid trans-location in a variety of higher eukaryotic cellmembranes or organelles (Diaz and Schroit, 1996;Haest et al., 1997; Angeletti and Nichols, 1998;Bevers et al., 1999), their use in yeast cells has beenrather limited (Kean et al., 1997; Siegmund et al.,1998; Marx et al., 1999).
The labelling of S. cerevisiae cells with NBD-phospholipids has been well established byKean et al. (1997; Siegmund et al., 1998). Figure 2describes a typical protocol used to determinethe translocase activity in S. cerevisiae cells, asmentioned in Materials and methods. Mid-exponentially grown cells of AD or its transfor-mants, expressing CDR1, CDR2 or CDR3, whenincubated with donor vesicles containing eitherM-C6NBD-PE, M-C6NBD-PS or M-C6NBD-PC,led to transfer of rapidly exchangeable NBD lipidanalogues from the quenched environment of thedonor vesicles to the non-quenched environment ofthe cells (Figure 2, steps A, B). NBD analogues didnot show any fluorescence in donor vesicles alone,due to presence of a phospholipid (N-Rh-DOPE)containing a different fluorophore, which has ahigher excitation wavelength, thus quenching NBDfluorescence by a resonance energy transfermechanism (Hrafnsdottir et al., 1997). N-Rh-DOPE, being a ‘non-exchangeable phospholipid’,does not participate in the donor-vesicle-to-cell-lipid exchange process due to its bulky fluorophore,covalently attached to the polar head moiety of thelipid molecule (Struck and Pagano, 1980). Whenmaximum labelling of cells with NBD analogueswas attained, they were washed to remove donorvesicles and then incubated at 30uC with theacceptor vesicles containing L-DPPC and L-DPPE(Figure 2, step C). When the unlabelled acceptorvesicles come into contact with the labelled cells, the
NBD molecules present in the outer monolayer ofthe plasma membrane are exchanged onto theacceptor vesicles, which results in an increasedfluorescence in the supernatant. Because of thecontinuos flip-flop activity of translocases in theplasma membrane, some of the NBD molecules
Figure 2. Schematic representation of the protocol usedfor determining the specificity of different transporters fordifferent phospholipids. (A) Incubation of S. cerevisiae AD,AD-CDR1, AD-CDR2 and AD-CDR3 cells with donorvesicles having L-DPPC, N-Rh-DOPE and either of M-C6NBD-PC, M-C6NBD-PE or M-C6NBD-PS. (B) Transfer offluorescent NBD phospholipid analogues to cells by theexchange process between the phospholipids of cells andNBD molecules of the donor vesicles. (C) Incubation ofthe washed NBD-labelled cells with acceptor vesicles.(D) Transfer of NBD phospholipid analogues from the cell’sexterior monolayer to the acceptor vesicles by back-exchange activity
Phospholipid translocators of Candida albicans 309
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.

present in the inner monolayer are transferred tothe outer monolayer, which then exchange phos-pholipids with the acceptor vesicles, resulting infurther change in the fluorescence with time. Thesteady-state increase/decrease in fluorescence in theacceptor vesicles has been considered to be an indexof translocase activity (Figure 2, step D) (Keanet al., 1997). For routine experiments, a 90 min timepoint, which showed maximum change in fluores-cence in the acceptor vesicles, was selected forcomparison of phospholipid translocase activity. Itshould be pointed out that NBD-tagged phospho-lipids do not degrade during the course of theexperiments. This was routinely checked by extract-ing NBD lipids from labelled cells and analysingthem on TLC.
Cdrps mediate transbilayer movement ofphospholipids
Donor vesicles containing one of the NBDanalogues (M-C6NBD-PE, M-C6NBD-PS orM-C6NBD-PC), N-Rh-DOPE and L-DPPC wereincubated with host strain AD and its transfor-mants expressing CDR1, CDR2 or CDR3. Asshown in Figure 3A, B, C, the cells showed a time-dependent increase in fluorescence, which plateausat ca. 90 min. These NBD-labelled cells were usedfor subsequent experiments.
Figure 4A depicts an increase in back-exchangedfluorescence of M-C6-NBD-PE in the supernatant,of CDR1- and CDR2-expressing cells as comparedto their common host strain AD at 90 min timeinterval. This suggests that both CDR1 and CDR2mediate outwardly directed M-C6-NBD-PE translo-case activity, which results in an increase in thefluorescence of the acceptor vesicles. Interestingly,a reverse situation was observed in the case ofCDR3-expressing cells, where fluorescence of M-C6-NBD-PE in the acceptor vesicles was decreased(Figure 4A).
In order to check the specificity of phospholipidtranslocation mediated by these ABC transporters,we studied back-exchange fluorescence using M-C6NBD-PC and M-C6NBD-PS phospholipid ana-logues. The strains expressing CDR1 and CDR2showed an increased fluorescence for M-C6NBD-PC and M-C6NBD-PS (Figure 4B, C) in theacceptor vesicles as compared to the parent strainAD. These results indicate that both CDR1 andCDR2 could translocate M-C6NBD-PC or M-C6NBD-PS from the inner to the outer monolayer
Figure 3. Labelling of yeast cells with M-C6NBD-PE, M-C6-NBD-PC, M-C6-NBD-PS phospholipid analogues. AD and itstransformants expressing CDR1, CDR2 and CDR3 cells wereincubated with donor vesicles having 58 mol% L-DPPC,2 mol% N-Rh-DPPE and 40 mol% of M-C6NBD-PE, M-C6-NBD-PC, M-C6-NBD-PS up to 90 min at 0uC. The figuredepicts the time-dependent labelling of strains AD,AD-CDR1, AD-CDR2 and AD-CDR3 with donor vesicles.The results shown are the mean of more than six indepen-dent experiments. —%—, AD; —&—, AD-CDR1; —+—,AD-CDR2; —$— AD-CDR3
310 Smriti et al.
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.
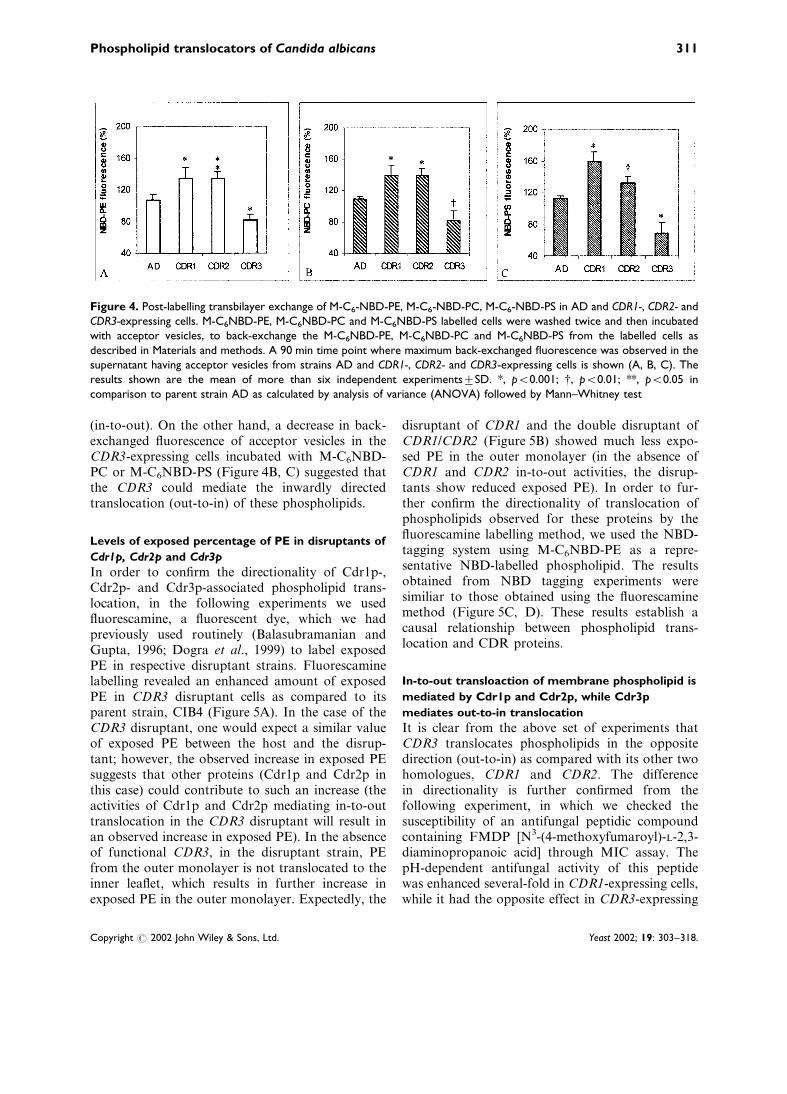
(in-to-out). On the other hand, a decrease in back-exchanged fluorescence of acceptor vesicles in theCDR3-expressing cells incubated with M-C6NBD-PC or M-C6NBD-PS (Figure 4B, C) suggested thatthe CDR3 could mediate the inwardly directedtranslocation (out-to-in) of these phospholipids.
Levels of exposed percentage of PE in disruptants of
Cdr1p, Cdr2p and Cdr3p
In order to confirm the directionality of Cdr1p-,Cdr2p- and Cdr3p-associated phospholipid trans-location, in the following experiments we usedfluorescamine, a fluorescent dye, which we hadpreviously used routinely (Balasubramanian andGupta, 1996; Dogra et al., 1999) to label exposedPE in respective disruptant strains. Fluorescaminelabelling revealed an enhanced amount of exposedPE in CDR3 disruptant cells as compared to itsparent strain, CIB4 (Figure 5A). In the case of theCDR3 disruptant, one would expect a similar valueof exposed PE between the host and the disrup-tant; however, the observed increase in exposed PEsuggests that other proteins (Cdr1p and Cdr2p inthis case) could contribute to such an increase (theactivities of Cdr1p and Cdr2p mediating in-to-outtranslocation in the CDR3 disruptant will result inan observed increase in exposed PE). In the absenceof functional CDR3, in the disruptant strain, PEfrom the outer monolayer is not translocated to theinner leaflet, which results in further increase inexposed PE in the outer monolayer. Expectedly, the
disruptant of CDR1 and the double disruptant ofCDR1/CDR2 (Figure 5B) showed much less expo-sed PE in the outer monolayer (in the absence ofCDR1 and CDR2 in-to-out activities, the disrup-tants show reduced exposed PE). In order to fur-ther confirm the directionality of translocation ofphospholipids observed for these proteins by thefluorescamine labelling method, we used the NBD-tagging system using M-C6NBD-PE as a repre-sentative NBD-labelled phospholipid. The resultsobtained from NBD tagging experiments weresimiliar to those obtained using the fluorescaminemethod (Figure 5C, D). These results establish acausal relationship between phospholipid trans-location and CDR proteins.
In-to-out transloaction of membrane phospholipid is
mediated by Cdr1p and Cdr2p, while Cdr3p
mediates out-to-in translocation
It is clear from the above set of experiments thatCDR3 translocates phospholipids in the oppositedirection (out-to-in) as compared with its other twohomologues, CDR1 and CDR2. The differencein directionality is further confirmed from thefollowing experiment, in which we checked thesusceptibility of an antifungal peptidic compoundcontaining FMDP [N3-(4-methoxyfumaroyl)-L-2,3-diaminopropanoic acid] through MIC assay. ThepH-dependent antifungal activity of this peptidewas enhanced several-fold in CDR1-expressing cells,while it had the opposite effect in CDR3-expressing
Figure 4. Post-labelling transbilayer exchange of M-C6-NBD-PE, M-C6-NBD-PC, M-C6-NBD-PS in AD and CDR1-, CDR2- andCDR3-expressing cells. M-C6NBD-PE, M-C6NBD-PC and M-C6NBD-PS labelled cells were washed twice and then incubatedwith acceptor vesicles, to back-exchange the M-C6NBD-PE, M-C6NBD-PC and M-C6NBD-PS from the labelled cells asdescribed in Materials and methods. A 90 min time point where maximum back-exchanged fluorescence was observed in thesupernatant having acceptor vesicles from strains AD and CDR1-, CDR2- and CDR3-expressing cells is shown (A, B, C). Theresults shown are the mean of more than six independent experimentstSD. *, p<0.001; {, p<0.01; **, p<0.05 incomparison to parent strain AD as calculated by analysis of variance (ANOVA) followed by Mann–Whitney test
Phospholipid translocators of Candida albicans 311
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.

cells (Table 1). The antifungal activity of Nva–FMDP is linked to the proton pumping activityof Cdrps. We have previously shown that CDR1extrudes protons, while CDR3, because of its
opposite directionality, does not extrude protons(Fritz et al., 1999; Milewski et al., 2001). SinceCDR3-expressing cells cannot lower the externalpH (because of the difference in directionality)and thus do not promote Nva–FMDP entry, theybecome resistant to it.
CDR1-, CDR2- and CDR3-mediated phospholipid
translocase activities differ in their sensitivity to
inhibitors
In order to ascertain whether or not phospholipidtranslocation mediated by Cdrps is energy-dependent, the effects of ATP-depleting agents wasstudied for the back-exchange activity of CDR1-,CDR2- and CDR3-expressing cells, using M-C6NBD-PE as a representative phospholipid ana-logue. The energy dependence of phospholipidtranslocation mediated by Cdr1p, Cdr2p andCdr3p was confirmed by using sodium azide andsodium orthovanadate, which significantly blockedthe back-exchange activity of Cdrps (Figure 6A).The presence of inhibitors led to a decreasedexposure of M-C6NBD-PE in cells expressingCDR1 and CDR2 (reduced back-exchange). Thepercentage extraction of M-C6NBD-PE in cellsexpressing CDR3 was, however, found to be greaterwhen treated with sodium azide and sodium ortho-vanadate, as compared to host AD. The increasein back-exchanged M-C6NBD-PE in the presenceof inhibitors suggests that with the inhibition ofCDR3, other transporters involved in translocatingM-C6NBD-PE from the inner to the outer mono-layer of the cells could also contribute to such ascenario (Figure 6A).
NEM is known to react with the thiol groupsof proteins. Any alteration followed by NEMtreatment in membrane lipid organization would
Figure 5. (A, B) Percentage labelling of PE in the plasmamembrane of Candida albicans cells with fluorescamine.Disruptant cells of CDR1, CDR2, CDR3 and their respectiveparent cells were labelled with fluorescamine as described inMaterials and methods. After labelling, the cells were washedand lipids were extracted. The percentage of derivatized PEwas determined as described in Materials and methods.(C, D) Postlabelling transbilayer exchange of M-C6-NBD-PEin parent and their disruptant cells. Cells labelled with M-C6NBD-PE were washed twice and then incubated withacceptor vesicles to back-exchange the M-C6NBD-PE fromthe labelled cells as described in Materials and methods. A90 min time point where maximum back-exchanged fluores-cence was observed in the supernatant having acceptorvesicles from parent strains CAF2-1, CAI4 and theirdisruptants, DSY449, DSY1025 and CIB4. The resultsshown are the mean of more than 10 independentexperimentstSD. {, p<0.01; *, p<0.001 in comparison torespective parent strain as calculated by analysis of variance(ANOVA) followed by Mann–Whitney test
Table 1. Nva–FMDP susceptibility test of AD and itstransformants expressing different CDRs
Strains Nva–FMDP [MIC (mg/ml)]
AD 3.1CDR1 0.8 (S)
CDR2 1.6 (S)
CDR3 3.1 (R)
MIC values for FMDP were determined by microdilution assay (Talibiand Raymond, 1999). The MIC test end point was defined as the
lowest drug concentration which gave>80% inhibition of growth.
S, sensitive to peptide; R, resistant to peptide.
312 Smriti et al.
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.

therefore lead to inhibition in protein-mediatedprocesses. This approach has commonly been usedto determine the mechanism of transbilayer move-ment of phospholipids in eukaryotic cells (Keanet al., 1997; Balasubramanian and Gupta, 1996;Bevers et al., 1999). When labelled cells expressingCDR1 or CDR2 were incubated with NEM,significant reduction was observed in the back-exchange of M-C6-NBD-PE (30–50% inhibition) toacceptor vesicles. Interestingly, M-C6-NBD-PEtranslocation in CDR3-expressing cells was insensi-tive to NEM (Figure 6B).
Disruption of the membrane cytoskeleton affects
phospholipid translocation mediated by Cdr1p and
Cdr2p but not by Cdr3p
The role of the membrane cytoskeleton in main-taining membrane asymmetry has been implicatedfor a variety of eukaryotic membranes (Dixit andGupta, 1998). Cytochalasins have been widelyused to disrupt the actin cytoskeleton in a varietyof eukaryotic cells, including yeast cells (Dixit andGupta, 1998). These compounds not only inducethe depolymerization of actin filaments but alsoprevent their growth. The cells expressing CDR1and CDR2, when treated with cytochalasin E,showed decreased back-exchange of M-C6-NBD-PE (Figure 6B); however, it had no effect on Cdr3p-mediated translocase activity. The selective effectof cytochalasin E on in-to-out translocase actvityand not on out-to-in activity, is consistent witha previously proposed model in yeast (Dixit andGupta, 1998), discussed below.
Antifungals and other drugs affect the transbilayer
movement of phospholipids mediated by Cdr1p
Cdr1p mediates multiple functions, includingdrug and human steroid hormone transport andphospholipid translocation (Krishnamurthy et al.,1998a, b). In order to determine any functionaloverlapping between drug extrusion and phospho-lipid binding site(s), we studied the transbilayerexchange of M-C6-NBD-PE in the presence ofdrugs to which Cdr1p confers resistance. Theresults, depicted in Figure 7, showed that in-to-outmovement of M-C6-NBD-PE mediated by Cdr1pwas inhibited if either cycloheximide, fluconazoleor miconazole drugs were present. This was evidentfrom reduced back-exchange of NBD-PE analoguefrom labelled cells to acceptor vesicles. On theother hand, Cdr3p-mediated translocation remainedunaffected in the presence of these drugs.
Discussion
The completion of the budding yeast, S. cerevisiae,genome sequence revealed the presence of at least31 ABC transporters. The accumulated data fromthe Candida genome sequence also suggests thata similar number of ABC transporters may exist inC. albicans. However, the physiological relevanceof these proteins is not so apparent. Only someof them have been shown to have a role in themultidrug resistance phenomenon in yeasts (Prasad
Figure 6. Effect of different inhibitors on translocase activityof CDR1-, CDR2- and CDR3-expressing cells. (A) CDR1-,CDR2- and CDR3-expressing cells were labelled with donorvesicles having M-C6-NBD-PE fluorescent analogue, werewashed and then incubated with the indicated concentra-tions of inhibitors sodium azide (azide), sodium orthovana-date (vanadate), N-ethylmaleimide (NEM) and cytochalasin Efor 20 min before incubating them with the acceptor vesicles.A 90 min time point where maximum change in back-exchanged fluorescence was taken to compare the percen-tage fluorescence in the supernatant having acceptorvesicles. The figure depicts percentage change in fluores-cence with and without inhibitors. The results are the meanof four or five independent experimentstSD. *, p<0.001;{, p<0.01; **, p<0.05; {, p>0.05 (not significant) in com-parison to respective control strain as calculated by analysisof variance (ANOVA) followed by Mann–Whitney test
Phospholipid translocators of Candida albicans 313
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.

et al., 1995a; Sanglard et al., 1997). Using NBDfluorescent analogues of phospholipids, we havefor the first time shown in this study that ABCtransporters of C. albicans, irrespective of theirinvolvement in drug transport, are capable oftransbilayer exchange of membrane phospholipids.These translocase activities could represent one ofthe normal physiological functions of such proteins.This is not to suggest that all pumps involved inmultidrug resistance are phospholipid translocators.For example, CaMDR1, which is a major facili-tator (MFS) protein whose expression is linked toazole resistance, shows no detectable phospholipidexchange activity (Dogra et al., 1999).
In contrast to Cdr3p, Cdr1p and Cdr2p have aestablished role in conferring azole resistance inC. albicans (Prasad et al., 1995a; Balan et al., 1997;Sanglard et al., 1997). However, these proteins actas phospholipid translocases, although they differin the directionality of membrane transbilayerexchange. Cdr1p and Cdr2p mediate in-to-outmovement of phospholipids, whereas Cdr3p directsout-to-in movement of phospholipids (Figure 8).
Furthermore, the two activities also differ in termsof their sensitivities to various inhibitors, suchas NEM and cytochalasin E. Cdr1p- and Cdr2p-associated phospholipid translocation was sensi-tive to NEM, while Cdr3p-mediated transbilayerexchange was insensitive to NEM. CytochalasinE induces alterations in the cytoskeleton, whichparticularly disrupts the actin organization in a
Figure 7. Effect of drugs on phospholipid translocase acti-vity of CDR1- and CDR3-expressing cells. AD-CDR1andAD-CDR3 cells were labelled with donor vesicles havingM-C6-NBD-PE. After labelling these cells were washed andthen incubated with 200 nM of each drug with the acceptorvesicles as described in Material and methods. A 90 min timepoint where maximum change in back-exchanged fluores-cence was observed was taken to compare the percentagefluorescence in the supernatant having acceptor vesicles. Thefigure depicts percentage change in fluorescence with andwithout drugs. The results are the mean of three indepen-dent experimentstSD. *, p<0.001; {, p<0.01; {, p>0.05(not significant) in comparison to respective control strain ascalculated by analysis of variance (ANOVA) followed byMann–Whitney test
Figure 8. A cartoon model depicting directionality ofphospholipid translocases of C. albicans. Translocase activitiesmediated by CDR1, CDR2 and CDR3
314 Smriti et al.
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.

variety of eukaryotic cells (Dixit and Gupta, 1998).These compounds not only induce depolymeriza-tion of actin filaments but also prevent their growth(Dixit and Gupta, 1998). Dixit and co-workers hadpreviously shown that actin cytoskeleton modifica-tion affects the PE translocation in yeast cells(Dixit and Gupta, 1998). They had proposed thatthe steady-state PE levels in the yeast cell plasmamembrane could be maintained by ATP-dependentout-to-in (pump I) and in-to-out (pump II) pumps.While pump I was envisaged to be high-affinity,ATP-dependent and NEM-insensitive, pump II wasconsidered to be low-affinity, ATP-dependent andNEM-sensitive. In the light of this model, it wassuggested that cytocholasin E-induced actin cytos-keleton changes should selectively affect pump II(Dixit and Gupta, 1998). In this study we confirmthe proposed model (Balasubramanian and Gupta,1996) and demonstrate that, indeed, only Cdr1p-and Cdr2p-mediated NEM-sensitive (in-to-out)translocase activity (pump II) is affected, whileCdr3p-mediated NEM-insensitive (out-to-in) acti-vity (pump I) is not affected by the disruption ofthe actin cytoskeleton.
A close homologue of MDR1, human MDR3 isnot a drug extrusion pump (Van Helvoort et al.,1996; Bevers et al., 1999); however, both ABCtransporters are able to mediate in-to-out trans-bilayer movement of phospholipids, albeit to differ-ent specificities. MDR3 is known to translocatespecifically PC, whereas MDR1 translocates a broadrange of lipid-soluble compounds, including avariety of anti-tumour drugs (Van Helvoort et al.,1996; Bevers et al., 1999). In addition, the inwardlydirected translocation of phospholipids is mediatedby a non-ABC protein, designated as amino-phospholipid translocase (Schroit and Zwaal,1991; Diaz and Schroit, 1996; Bevers et al., 1999).Scramblase is another enzyme that mediatesphospholipid transfer between the two monolayers.Scrambling is a bidirectional process and involvesall major classes of phospholipids (Bevers et al.,1999). It is the interplay of scramblase and amino-phospholipid translocase that maintains the asym-metry of the plasma membrane in mammalian cells(Bevers et al., 1999)
In earlier studies, phospholipid translocation hasbeen shown to be a protein-mediated phenomenonin yeast but the nature of such protein was notknown (Balasubramanian and Gupta, 1996). DRS2of S. cerevisiae a P-type ATPases has been shown tobe linked to phospholipid translocation; however,
such reports have been refuted (Tang et al., 1996;Siegmund et al., 1998; Marx et al., 1999). This is thefirst report in which a causal relationship betweenphospholipid translocation and ABC transportersof C. albicans has been described. Based on ourresults, it appears that the C. albicans plasma mem-brane has bidirectional energy-dependent phospho-lipid translocases to maintain high concentration ofaminophospholipids in the cytoplasmic leaflet. Thetranslocases responsible for in-to-out translocation(Cdr1p and Cdr2p) are different from its out-to-incounterpart (Cdr3p) (Figure 6). In spite of similar-ity with the mammalian MDR phospholipid trans-locators, yeast translocases differ from them indirectionality and specificity. For example, out-to-indirected aminophospholipid translocase activity ofmammalian plasma membrane is specific foraminophospholipids, while similar activity mediatedby Cdr3p in C. albicans membrane is rather uns-pecific (general phospholipid translocator).
It is interesting to note that drugs like fluocona-zole, cycloheximide and miconazole could affecttransbilayer movement of phospholipids mediatedby Cdr1p but had no effect on Cdr3p-mediatedtransbilayer exchange. These results do point outthat Cdr1p presumably has common binding sitesfor drugs and phospholipids, while the out-to-inactivity of Cdr3p is independent of drug binding.The difference in the directionality of phospholipidtransfer between Cdrps could be linked to theirability to efflux cytotoxic drugs. If the activity ofthe Cdr3p pump is inwardly directed, then itsinability to participate in drug transport can beexplained. However, comprehension of the mole-cular basis of functional differences between thesetransporters must await further experimentation.
What can be the functional relevance of phos-pholipid asymmetry in yeast membrane? It is anestablished fact that the appearance of aminopho-spholipids like PS in the outer monolayer triggersprocoagulant activity in RBCs and is a signal forapoptosis in mammalian cells (Diaz and Schroit,1996; Bevers et al., 1999). However, the implica-tions of aminophospholipids predominance in thecytoplasmic leaflet of the plasma membrane ofC. albicans cells and its movement to the exteriorcytoplasmic half of the lipid bilayer are not known.It is worth stressing that C. albicans is a pleio-morphic yeast which can exist in a variety ofmorphological forms (Odds, 1985; Soll et al.,1993). Although many environmental factors havebeen characterized that trigger the yeast–filament
Phospholipid translocators of Candida albicans 315
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.

interconversion, little is known about sensors orevents occurring at or within the cellular membraneduring the induction of the morphogenetic programof dimorphism (Hubbard et al., 1985; Stoldt et al.,1997; White and Larsen, 1997). Our earlier finding,that the availability of PE in the outer monolayer ofmembrane was considerably enhanced in the myce-lial form of C. albicans, may suggest that changes inlipid asymmetry could influence phenotypic diver-gence of this yeast (Dogra et al., 1999). In view ofthe recent findings that the membrane could affecttransversal of the stress signal and that its physicalstate is important for transcriptional activation ofcertain genes (Vigh et al., 1998; Smriti et al., 1999;Moskvina et al., 1999), membrane asymmetry andits maintenance could be of considerable impor-tance for a unicellular organism and more so for apathogenic yeast.
Acknowledgements
We are grateful to A. Decottignies, E. Balziand A. Goffeau for kindly providing the strainAD1234568. We would like to thank MartineRaymond for kindly providing the strain CIB4and CAI4. We are grateful to Dominique Sanglardfor kindly providing the strains CAF2-1, DSY449and DSY1025. We would like to thank DrVishwajeet Puri for providing us with some NBDderivatives used in this study. We are thankful toAvmeet for providing her suggestions during thepreparation of the manuscript. The work presentedin this paper has been supported in part by grantsto R.P. from the Department of Biotechnology(DBT-BT/PRO798/HRD20/8/98), the Departmentof Science and Technology (SP/SO/D57/97) andthe Council of Scientific and Industrial Research(60(0028)/98-EMR-II), India. Smriti acknowledgesthe fellowship award from the Council of Scientificand Industrial Research, India.
References
Angeletti C, Nichols JW. 1998. Dithionite quenching rate
measurement of the inside-outside membrane bilayer distribu-
tion of 7-nitrobenz-2-oxa-1,3-diazol-4-yl-labelled phospholi-
pids. Biochemistry 37: 15114–15119.
Balan I, Alarco AM, Raymond M. 1997. The Candida albicans
CDR3 gene codes for an opaque-phase ABC transporter.
J Bacteriol 179: 7210–7218.
Balasubramanian K, Gupta CM. 1996. Transbilayer phosphati-
dylethanolamine movements in the yeast plasma membrane:
evidence for a protein-mediated, energy-dependent mechanism.
Eur J Biochem 240: 798–806.
Bauer BE, Wolfger H, Kuchler K. 1999. Inventory and function
of yeats ABC proteins: about sex, stress, and pleiotropic drug
and heavy metal resistance. Biochim Biophys Acta 1461:
217–236.
Bevers EM, Confurius P, Dekkers DWC, Zwaal RFA. 1999.
Lipid translocation across the plasma membrane of mamma-
lian cells. Biochim Biophys Acta 1439: 317–330.
Calabrese D, Bille J, Sanglard D. 2000. A novel multidrug efflux
transporter gene of the major facilitator superfamily from C.
albicans (FLU1) conferring resistance to fluconazole. Micro-
biology 146: 2743–2754.
Connor J, Pak CC, Schroit AJ. 1994. Exposure of phosphati-
dylserine in the outer leaflet of human red blood cells: relation-
ship to cell density, cell stage, and clearance by mononuclear
cells. J Biol Chem 269: 2399–2404.
Cerbon J, Calderon V. 1991. Changes of the compositional
asymmetry of phopholipids associated to the increment in the
membrane surface potential. Biochim Biophys Acta 1067:
139–144.
Chattopadhyay A, London E. 1988. Spectroscopic and ionization
properties of N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)-labelled
lipids in model membranes. Biochim Biophys Acta 938: 24–34.
Decottignies A, Goffeau A. 1997. Complete inventory of the
yeast ABC proteins. Nature Genet 15: 137–145.
Diaz C, Schroit AJ. 1996. Role of translocases in the generation
of phosphatidylserine asymmetry. J Memb Biol 151: 1–9.
Dogra S, Krishnamurthy S, Gupta V, et al. 1999. Asym-
metric distribution of phosphatidylethanolamine in C. albicans:
possible mediation by CDR1, a multidrug transporter belong-
ing to ATP binding cassette (ABC) superfamily. Yeast 15:
111–121.
Dixit BL, Gupta CM. 1998. Role of the actin cytoskeleton in
regulating the outer phosphatidylethanolamine levels in yeast
plasma membrane. Eur J Biochem 254: 202–206.
Franz R, Michel S, Morschhauser J. 1998. A fourth gene from
the Candida albicans CDR family of ABC transporters. Gene
220: 91–98.
Fritz F, Howard EM, Hoffman MM, Roepe PD. 1999. Evidence
for altered ion transport in Saccharomyces cerevisiae over-
expressing human MDR1 protein. Biochemistry 38: 4214–
4226.
Gupta V, Kohli AK, Krishnamurthy S, Prasad R. 1998.
Identification of mutant alleles of CaMDR1, a major facil-
itator of Candida albicans which confers multidrug resistance
and its in vitro transcriptional activation. Curr Genet 34:
192–199.
Haest CWM, Oslender A, Kamp D. 1997. Non-mediated flip-
flop of anionic phospholipids and long-chain amphiphiles in
the erthrocyte membrane depends on membrane potential.
Biochemistry 36: 10885–10891.
Higgins CF. 1992. ABC Transporters: From Microorganisms to
Man. Ann Rev Cell Biol 8: 67–113.
Hrafnsdottir S, Nichols JW, Menon AK. 1997. Transbilayer
movement of flourescent phospholipids in Bacillus megaterium
membrane vesicles. Biochemistry 36: 4969–4978.
Hubbard MJ, Sullivan PA, Shepherd MG. 1985. Morphological
studies of N-acetylglucosamine induced germ tube formation
by Candida albicans. Can J Microbiol 31: 696–701.
Ibrahim AS, Ghannoum MA. 1996. Chromatographic analysis
316 Smriti et al.
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.

of lipids. In Manual on Membrane Lipids, Prasad R (ed.).
Springer Verlag: Berlin; 52–79.
Joseph-Horne T, Hollomon DW. 1997. Molecular mechanisms
of azole resistance in fungi. FEMS Microbiol Lett 149:
1141–149.
Kean LS, Grant AM, Angeletti C, et al. 1997. Plasma membrane
translocation of fluorescent-labelled phosphatidylethanolamine
is controlled by transcription regulators, PDR1 and PDR3.
J Cell Biol 138: 255–270.
Koul A, Prasad R. 1996. Extraction of membrane lipids. In
Manual on Membrane Lipids, Prasad R (ed.). Springer Verlag:
Berlin; 37–52.
Krishnamurthy S, Chatterjee U, Gupta V, et al. 1998a. Deletion
of transmembrane domain 12 of CDR1, a multidrug trans-
porter from Candida albicans, leads to altered drug specificity:
expression of a yeast multidrug transporter in Baculovirus
expression system. Yeast 14: 535–550.
Krishnamurthy S, Gupta V, Snehlata P, Prasad R. 1998b.
Characterization of human steroid hormone transport
mediated by Cdr1p, multidrug transporter of Candida albicans,
belonging to the ATP binding cassette super family. FEMS
Microbiol Lett 158: 69–74.
Krishnamurthy S, Gupta V, Prasad R, Panwar SL, Prasad R.
1998c. Expression of CDR1, a multidrug resistance gene of
Candida albicans: In vitro transcriptional activation by heat
shock, drugs and human steroid hormones. FEMS Microbiol
Lett 160: 191–197.
Lamb DC, Kelly DE, Schunck W-H, et al. 1997. The mutation
T315A in Candida albicans sterol 14a-demethylase causes
reduced enzyme activity and fluconazole resistance through
reduced affinity. J Biol Chem 272: 5682–5688.
Marichal P. 1999. Mechanisms of resistance to azole antifungal
compounds. Curr Opin Antiinfect Invest Drugs 1: 318–333.
Marx U, Polakowski T, Pomorski T, et al. 1999. Rapid
transbilayer movement of fluorescent phospholipid analogues
in the plasma membrane of endocytosis-deficient yeast cells
does not require the DRS2 protein. Eur J Biochem 263:
254–263.
Milewski S, Mignini F, Prasad R, Borowski E. 2001. Un-
usual susceptibility of a multidrug-resistance yeast strain
to peptidic antifungals. Antimicrob Agents Chemother 45:
223–228.
Moskvina E, Imre EM, Ruis H. 1999. Stress factors acting at the
level of plasma membrane induce transcription via the stress
response element (STRE) of the yeast Saccharomyces cerevi-
siae. Mol Microbiol 32: 1263–1272.
Mukherjee S, Chattopadhyay A, Samanta A, Soujanya T.
1994. Dipole moment change of NBD group upon excita-
tion studied using solvatochromic and quantum chemical
approaches :Implications in membrane research. J Phys Chem
98: 2809–2812.
Odds FC. 1985. Morphogenesis in Candida albicans. CRC Crit
Rev Microbiol 12: 45–93.
Op den Kamp JAF. 1979. Lipid asymmetry in membranes. Ann
Rev Biochem 48: 47–71.
Prasad R. 1991. The plasma membrane of Candida albicans:
its relevance to transport phenomenon. In Candida albicans:
Cellular and Molecular Biology, Prasad R (eds). Springer
Verlag: Berlin; 108–127.
Prasad R, Worgifosse PD, Goffeau A, Balzi E. 1995a. Molecular
cloning and characterization of a novel gene of C.albicans,
CDR1, conferring multiple resistance to drugs and antifungals.
Curr Genet 27: 320–329.
Prasad R, Krishnamurthy S, Prasad R, Gupta V. 1995b.
Multiple drug resistance in Candida albicans. Acta Biochem
Polon 42: 497–504.
Prasad R, Krishnamurthy S, Prasad R, Gupta V, Lata S.
1996. Multidrug resistance: an emerging threat. Curr Sci
71: 205–213.
Prasad R, Panwar SL, Krishnamurthy S. 2000. Drug resistance
mechanisms of human pathogenic fungi. In Fungal Pathogene-
sis: Principles and Clinical Applications, Cihlar RL, Calderone
RA (eds) (in press).
Sanglard D, Ischer F, Monod M, Bille J. 1997. Cloning of
Candida albicans genes conferring resistance to azole anti-
fungal agents: characterization of CDR2, a new multidrug
ABC transporter gene. Microbiology 143: 405–416.
Sanglard D, Ischer F, Monod M, Dogra S, Prasad R, Bille J.
1999. Analysis of the ATP-binding cassette (ABC)-transporter
gene of CDR4 from Candida albicans (abstr). ASM Conference
on Candida and Candidiasis 1999; C27: 56.
Schroit AJ, Zwaal RFA. 1991. Transbilayer movement of
phospholipids in red cell and platelet membranes. Biochim
Biophys Acta 1071: 313–329.
Siegmund A, Grant A, Angeletti C, Malone L, Nichols JW,
Rudolph HK. 1998. Loss of Drs2p does not abolish trans-
fer of fluorescence-labelled phospholipids across the plasma
membrane of Saccharomyces cerevisiae. J Biol Chem 273:
34399–34405.
Smriti, Krishnamurthy S, Prasad R. 1999. Membrane fluidity
affects functions of Cdr1p, a multidrug ABC transporter of
Candida albicans. FEMS Microbiol Lett 173: 475–481.
Soll DR, Morrow B, Srikantha T. 1993. High frequency
phenotypic switching in Candida albicans. Trends Genet
9: 61–65.
Stoldt VR, Sonneborn A, Leuker CE, Ernst J. 1997. Efg1,
an essential regulator of morphogenesis of human pathogen
Candida albicans, is a member of conserved class of bHLH
regulating the morphogenetic process in fungi. EMBO J
16: 1982–1991.
Struck DK, Pagano RE. 1980. Insertion of fluorescent phospho-
lipids into the plasma membrane of a mammalian cell. J Biol
Chem 255: 5404–5410.
Tang X, Halleck MS, Schlegel RA, Williamson P. 1996. A
subfamily of P-type ATPases with aminophospholipid trans-
porting activity. Science 272: 1495–1497.
Talibi D, Raymond M. 1999. Isolation of a putative Candida
albicans transcriptional regulator involved in pleiotropic
drug resistance by functional complementation of a pdr1
pdr3 mutation in Saccharomyces cerevisiae. J Bacteriol
181: 231–240.
Van HelvoortA, SmithAJ, SprongH, et al. 1996. MDR1
P-glycoprotein is a lipid translocase of broad specifity, while
MDR3 P-glycoprotein specifically translocates phosphatidyl-
choline. Cell 87: 507–517.
Vigh L, Maresca B, Harwood JL. 1998. Does the membrane’s
physical state control the expression of heat shock and other
genes? Trends Biochem Sci 23: 369–374.
Williamson P, Bevers EM, Smeets M, et al. 1995. Continous
analysis of the mechanism of activated transbilayer lipid
movement in platelets. Biochemistry 34: 10448–10454.
Phospholipid translocators of Candida albicans 317
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.

White S, Larsen B. 1997. Candida albicans morphogenesis is
influenced by estrogen. Cell Mol Life Sci 53: 744–749.
White TC. 1997. Increased mRNA levels of ERG16, CDR,
and MDR1 correlate with increased azole resistance in
Candida albicans isolates from a patient infected with human
immunodeficiency virus. Antimicrob Agents Chemother
41: 1482–1487.
White TC, Marr KA, Bowden RA. 1998. Clinical, cellular, and
molecular factors that contribute to antifungal drug resistance.
Clin Microbiol Rev 11: 382–402.
Zhou Q, Sims PJ, Wiedmer T. 1998. Identity of a conserved
motif in phospholipid scramblase that is required for Ca2+-
accelerated transbilayer movement of membrane phospho-
lipids. Biochemistry 37: 23356–2360.
318 Smriti et al.
Copyright # 2002 John Wiley & Sons, Ltd. Yeast 2002; 19: 303–318.