abcc drug efflux pumps and organic anion uptake ... · abcc drug efflux pumps and organic anion...
TRANSCRIPT

ABCC Drug Efflux Pumps and Organic Anion Uptake Transporters
in Human Gliomas and the Blood-Tumor Barrier
Holger Bronger,1Jorg Konig,
1Kathrin Kopplow,
1Hans-Herbert Steiner,
2Rezvan Ahmadi,
2
Christel Herold-Mende,2,3Dietrich Keppler,
1and Anne T. Nies
1
1Division of Tumor Biochemistry, German Cancer Research Center; 2Molecular Biology Laboratory, Neurosurgery Hospital, University ofHeidelberg; and 3Department of Head and Neck Surgery, Molecular Cell Biology Group, University of Heidelberg, Heidelberg, Germany
Abstract
Delivery of therapeutic agents to the brain and its neoplasmsdepends on the presence of membrane transport proteins inthe blood-brain barrier and in the target cells. The cellularand subcellular localization of these membrane transportersdetermines the drug accessibility to the brain and its tumors.We therefore analyzed the expression and localization ofsix members of the multidrug resistance protein family ofATP-dependent efflux pumps (ABCC1-ABCC6, formerlyMRP1-MRP6) and of six organic anion uptake transporters(OATP1A2, OATP1B1, OATP1B3, OATP1C1, OATP2B1, andOATP4A1) in 61 human glioma specimens of differenthistologic subtypes. Real-time PCRs indicated expressions ofABCC1, ABCC3, ABCC4 , and ABCC5 . In addition, we detectedexpressions of the OATP uptake transporter genes SLCO1A2,SLCO1C1, SLCO2B1 , and SLCO4A1 . At the protein level,however, only OATP1A2 and OATP2B1 were detectable byimmunofluorescence microscopy in the luminal membrane ofendothelial cells forming the blood-brain barrier and theblood-tumor barrier, but not in the glioma cells. ABCC4 andABCC5 proteins were the major ABCC subfamily members ingliomas, localized both at the luminal side of the endothelialcells and in the glioma cells of astrocytic tumors and inthe astrocytic portions of oligoastrocytomas. These resultsindicate that expression of ABCC4 and ABCC5 is associatedwith an astrocytic phenotype, in accordance with theirexpression in astrocytes and with the higher chemoresistanceof astrocytic tumors as compared with oligodendrogliomas.Our data provide a basis for the assessment of the role ofuptake transporters and efflux pumps in the accessibilityof human gliomas for chemotherapeutic agents. (Cancer Res2005; 65(24): 11419-28)
Introduction
The therapy of gliomas, the most common brain neoplasms ofadults, is often impeded by their high resistance to chemothera-peutic agents although some entities such as malignant oligoden-drogliomas and oligoastrocytomas exhibit response rates of up to70% (1). Many different mechanisms may account for thischemoresistance, including reduced blood supply to the tumor,up-regulation of antiapoptotic pathways, enhanced DNA repairmechanisms, and increased metabolic inactivation and subsequent
elimination of the applied drugs (2). Moreover, in recent years, theexpression and function of ATP-driven export pumps of the ATP-binding cassette (ABC) protein superfamily have been extensivelystudied in cancer cell lines, in in vitro models, and in tissues.Besides transporting a variety of physiologic substrates, several ofthese transporters also confer resistance to cytotoxic and antiviraldrugs (3). Entry of many hydrophobic cationic drugs into thebrain is decisively limited by multidrug resistance 1 (MDR1)P-glycoprotein (symbol ABCB1) in the blood-brain barrier (4). Thepresence of other ABC family members in the blood-brain barrierof human brain, including breast cancer resistance protein, ABCG2(5), and efflux proteins of the ABCC subfamily (6), which have othersubstrate specificities than MDR1 P-glycoprotein (7), may add tothe resistance of the brain to cytotoxic and antiviral drugs,especially of organic anions conjugated with sulfate, glucuronate,or glutathione, and of nucleoside analogues.In gliomas, mainly MDR1 P-glycoprotein and ABCC1 have been
studied thus far. Both were localized to the tumor vasculature andpartially also to the tumor cells, however, with varying results(8–12). In glioma cell lines, MDR1 P-glycoprotein and ABCC1 wereshown to confer resistance to various anticancer drugs (9, 12).Because several other ABCC subfamily members confer resistanceto cytotoxic drugs as well, such as thioguanine and 6-mercaptopu-rine in case of ABCC5 (13) or methotrexate and topotecan in case ofABCC4 (14, 15), we were interested in their expression patterns inhuman gliomas, particularly with respect to the histologic subtype.Recently, we localized ABCC4 and ABCC5 in endothelial cells andastrocytes, but not in oligodendrocytes, of perilesional human brainsamples (6). Given the possibility that gliomas originate bydedifferentiation of mature glial cells (16), the question ariseswhether the tumor cells retain the ABCC expression pattern of thecell type they derive from while undergoing malignant progression,which could account for the differential chemoresistance of thedifferent glioma entities. Another possible mechanism causinginsufficient drug delivery to the brain and its neoplasms is a reducedexpression of uptake transporters in the blood-brain barrier orblood-tumor barrier or in the glioma cells. Members of the family oforganic anion transporting polypeptides (OATP) transport chemo-therapeutic agents, such as methotrexate (17) and SN-38, the activemetabolite of irinotecan (18). OATPs are sodium-independentuptake transporters for various amphiphilic anionic compounds,including bile salts, prostanoids, steroid conjugates, thyroidhormones, anionic peptides, and xenobiotics (19). Several Oatpshave been identified in the blood-brain barrier of rat brain (20).Because many rat Oatps do not have a human orthologue (19),it is of considerable interest to identify OATPs in the humanbrain. Thus far, only OATP1A2 protein has been localized in humanbrain (21).To explore the possible role of ABCC export pumps and OATP
uptake transporters in the resistance of gliomas, we investigated
Requests for reprints: Anne Nies, Division of Tumor Biochemistry, DeutschesKrebsforschungszentrum, Im Neuenheimer Feld 280, D-69120 Heidelberg, Germany.Phone: 49-62-21-422-403; Fax: 49-62-21-422-402; E-mail: [email protected].
I2005 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-05-1271
www.aacrjournals.org 11419 Cancer Res 2005; 65: (24). December 15, 2005
Research Article
Research. on June 5, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
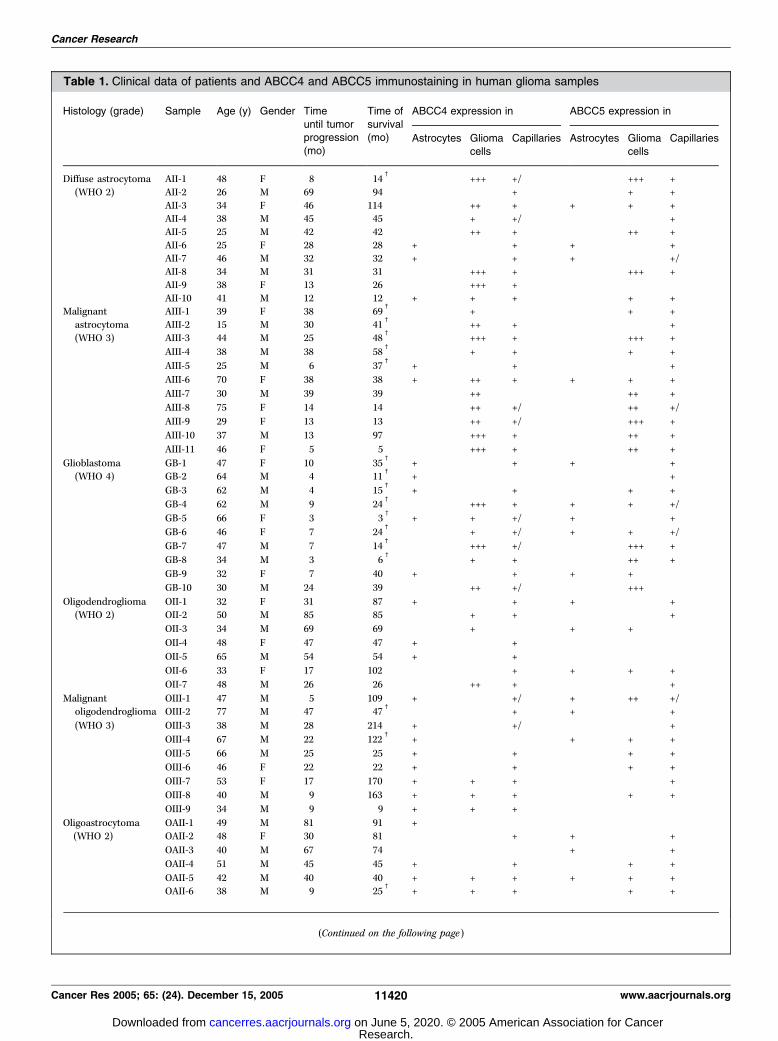
Table 1. Clinical data of patients and ABCC4 and ABCC5 immunostaining in human glioma samples
Histology (grade) Sample Age (y) Gender Time Time of ABCC4 expression in ABCC5 expression in
until tumorprogression
(mo)
survival(mo) Astrocytes Glioma
cells
Capillaries Astrocytes Glioma
cells
Capillaries
Diffuse astrocytoma AII-1 48 F 8 14c
+++ +/� +++ +
(WHO 2) AII-2 26 M 69 94 � + + +
AII-3 34 F 46 114 ++ + + + +AII-4 38 M 45 45 + +/� � +
AII-5 25 M 42 42 ++ + ++ +
AII-6 25 F 28 28 + � + + � +AII-7 46 M 32 32 + � + + � +/�AII-8 34 M 31 31 +++ + +++ +
AII-9 38 F 13 26 +++ + �AII-10 41 M 12 12 + + + + +
Malignant AIII-1 39 F 38 69c
+ � + +
astrocytoma AIII-2 15 M 30 41c
++ + � +
(WHO 3) AIII-3 44 M 25 48c
+++ + +++ +
AIII-4 38 M 38 58c
+ + + +
AIII-5 25 M 6 37c
+ � + � +
AIII-6 70 F 38 38 + ++ + + + +
AIII-7 30 M 39 39 ++ � ++ +
AIII-8 75 F 14 14 ++ +/� ++ +/�AIII-9 29 F 13 13 ++ +/� +++ +
AIII-10 37 M 13 97 +++ + ++ +
AIII-11 46 F 5 5 +++ + ++ +
Glioblastoma GB-1 47 F 10 35c
+ � + + � +
(WHO 4) GB-2 64 M 4 11c
+ � � +
GB-3 62 M 4 15c
+ � + + +
GB-4 62 M 9 24c
+++ + + + +/�GB-5 66 F 3 3
c+ + +/� + � +
GB-6 46 F 7 24c
+ +/� + + +/�GB-7 47 M 7 14
c+++ +/� +++ +
GB-8 34 M 3 6c
+ + ++ +
GB-9 32 F 7 40 + � + + + �GB-10 30 M 24 39 ++ +/� +++ �
Oligodendroglioma OII-1 32 F 31 87 + � + + � +
(WHO 2) OII-2 50 M 85 85 + + � +
OII-3 34 M 69 69 � + � + + �OII-4 48 F 47 47 + � + � �OII-5 65 M 54 54 + � + � �OII-6 33 F 17 102 � + + + +
OII-7 48 M 26 26 ++ + � +
Malignant OIII-1 47 M 5 109 + � +/� + ++ +/�oligodendroglioma OIII-2 77 M 47 47
c � � + + � +
(WHO 3) OIII-3 38 M 28 214 + � +/� � +
OIII-4 67 M 22 122c
+ � � + + +
OIII-5 66 M 25 25 + � + + +
OIII-6 46 F 22 22 + � + + +
OIII-7 53 F 17 170 + + + +
OIII-8 40 M 9 163 + + + + +
OIII-9 34 M 9 9 + + + � �Oligoastrocytoma OAII-1 49 M 81 91 + � �(WHO 2) OAII-2 48 F 30 81 � � + + � +
OAII-3 40 M 67 74 � � � + � +
OAII-4 51 M 45 45 + � + + +
OAII-5 42 M 40 40 + + + + + +
OAII-6 38 M 9 25c
+ + + + +
(Continued on the following page)
Cancer Research
Cancer Res 2005; 65: (24). December 15, 2005 11420 www.aacrjournals.org
Research. on June 5, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
the expression and cellular distribution of six members of theABCC subfamily (ABCC1-ABCC6) in frozen glioma samples ofdifferent histologic subtypes and of six members of the OATPfamily (OATP1A2, OATP1B1, OATP1B3, OATP1C1, OATP2B1, andOATP4A1). ABCC4 and ABCC5 were the major ABCC subfamilymembers expressed in the blood-tumor barrier and in the gliomacells, notably in astrocytic tumors. This is in good agreement withthe overall higher chemoresistance of these tumors compared witholigodendroglial and mixed gliomas. Both ABCC proteins maytherefore contribute to the MDR phenotype of gliomas. In addition,OATP1A2 and OATP2B1 proteins were detected in the blood-brainbarrier and the blood-tumor barrier.
Materials and Methods
Antibodies and materials. The monoclonal antibodies (mAb) QCRL-1and MRPr1 against ABCC1, M2III-6 and M2II-12 against ABCC2, M3II-9
against ABCC3, M5I-1 against ABCC5, M6II-31 against ABCC6, and BXP-21
against ABCG2 and the polyclonal antibody against ABCC4 (MRP4
polyclonal antibody) were purchased from Alexis (Lausen, Switzerland)and used according to the instructions of the manufacturer. Mouse mAbs
against glial fibrillary acidic protein (GFAP) and against the endothelial cell
marker CD31 were from Chemicon (Temecula, CA) and diluted 1:300 and
1:10, respectively. The use and affinity purification of the polyclonalantibodies EAG5, SNG, and AMF against human ABCC2, ABCC4, and
ABCC5, respectively, have been described elsewhere (6). The polyclonal
antibodies CKD, ENY, SPA, and LPS were raised in rabbits against theCOOH-terminal sequences of OATP1A2 (CKDIYQKSTVLKDDELKTKL;
National Center for Biotechnology Information accession no. NP_602307),
OATP1C1 (ENYTTSDHLLQPNYWPGKETQL; accession no. NP_059131),
OATP2B1 (SPAVEQQLLVSGPGKKPEDSRV; accession no. NP_009187), andOATP4A1 (LPSQSSAPDSATDSQLQSSV; accession no. BAA89288), respectively.
The peptides were synthesized and coupled to keyhole limpet hemocyanin
(Peptide Specialty Laboratories, Heidelberg, Germany). The antisera were
affinity-purified as described earlier (22) using membrane vesicles preparedfrom HEK cells expressing the respective recombinant OATP. The affinity-
purified antibodies were used at final concentrations of 0.2 to 0.4 mg/mL for
the immunofluorescence microscopy. Alexa Fluor488–conjugated goat anti-rabbit, anti-mouse, and anti-rat immunoglobulin G (IgG) were from
Molecular Probes (Eugene, OR). Cy3-conjugated anti-mouse IgG was from
Jackson ImmunoResearch (West Grove, PA). Horseradish peroxidase–
conjugated goat anti-rabbit and anti-mouse IgGs were from Bio-Rad
(Munich, Germany) and goat anti-rat IgG was from Promega (Mannheim,
Germany). All other chemicals were of analytical grade and obtained from
Merck (Darmstadt, Germany) or Sigma (St. Louis, MO).Human tissue samples. The present study was undertaken after
informed consent had been obtained from each patient, in accordance with
the regulations of the Ethics Committee, University of Heidelberg, and with
the Declaration of Helsinki. Glioma samples obtained at the NeurosurgeryHospital, University of Heidelberg (Heidelberg, Germany) were 10 diffuse
astrocytomas (WHO grade 2), 11 malignant astrocytomas (WHO grade 3),
10 glioblastomas (WHO grade 4), 7 oligodendrogliomas (WHO grade 2),9 malignant oligodendrogliomas (WHO grade 3), 6 oligoastrocytomas (WHO
grade 2), and 8 malignant oligoastrocytomas (WHO grade 3). The tumors
were classified by a neuropathologist according to the WHO classification of
brain tumors. A clinical follow-up was assessed for all patients. None of thepatients had received radiotherapy and four had received chemotherapy
before surgery (Table 1). After resection, each sample was immediately
snap-frozen in liquid nitrogen and stored at �80jC until further use.
In addition to immunofluorescence microscopy, material from someof the samples was used for mRNA analysis or homogenized and used
for immunoblot analysis. Perilesional brain samples from the temporal
lobe were from patients undergoing surgical resection of gliomas orundergoing neurosurgery because of cerebral hemorrhage (6). The
perilesional tissue was removed and immediately frozen in liquid
nitrogen before use. None of the patients had received chemotherapy
before neurosurgery.Immunofluorescence microscopy. Cryostat sections (5 Am) were
prepared, air-dried for 16 hours at room temperature, and fixed for 10
minutes in acetone (�20jC), or in phosphate-buffered formaldehyde (10%,
room temperature) for SPA antibody only. Sections were then incubated with
the primary antibodies diluted in PBS for 60 minutes at room temperature,
washed thoroughly in PBS, and incubated with the respective secondary
antibody (1:300) for 60minutes. After washing in PBS, sections weremounted
in Moviol (Calbiochem, La Jolla, CA). Fluorescence micrographs were taken
with a confocal LSM510 laser scanning microscope (Carl Zeiss, Jena,
Germany). For peptide competition experiments, affinity-purified antibodies
were incubated at 4jC for 16 hours with the respective peptide (100-200
Amol/L) used to generate the antibody, and then applied to the cryosections.
In some experiments, nuclei were stainedwith propidium iodide added to the
secondary antibody solution at a final concentration of 2 Ag/mL.
ABCC immunostaining of tumor cells was assessed semiquantitativelybased on the percentage of tumor cells showing specific immunoreactivity
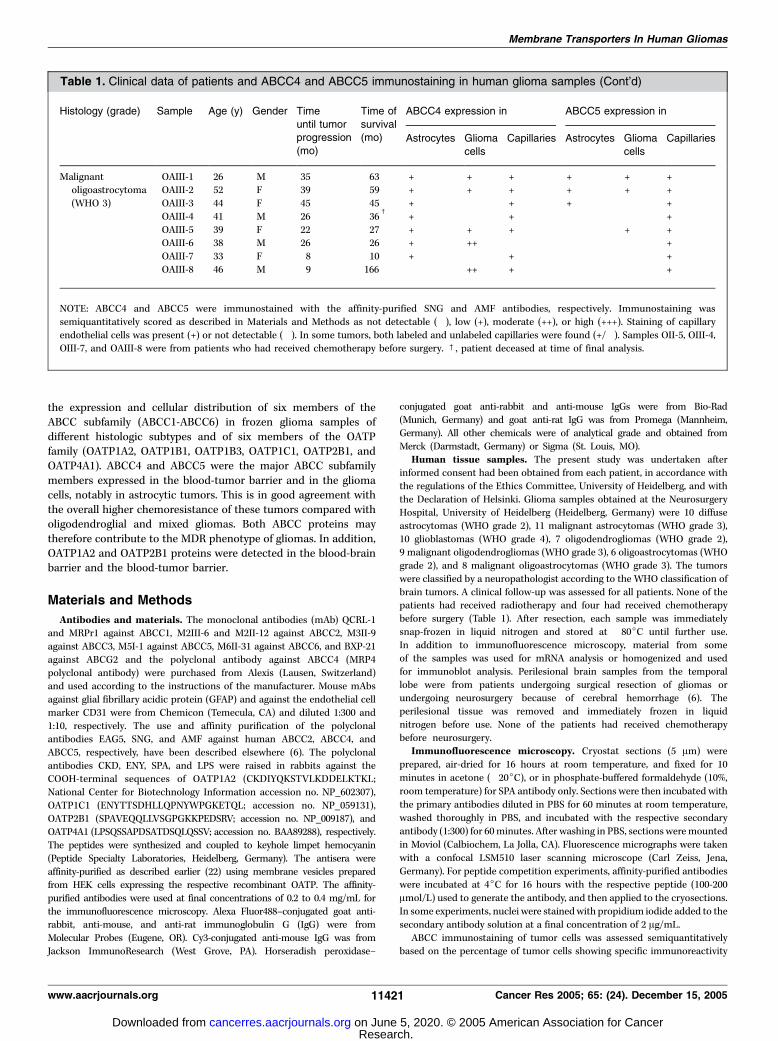
Malignant OAIII-1 26 M 35 63 + + + + + +oligoastrocytoma OAIII-2 52 F 39 59 + + + + + +
(WHO 3) OAIII-3 44 F 45 45 + � + + � +
OAIII-4 41 M 26 36c
+ � + � +
OAIII-5 39 F 22 27 + + + + +OAIII-6 38 M 26 26 + ++ � � +
OAIII-7 33 F 8 10 + � + � +
OAIII-8 46 M 9 166 ++ + � +
NOTE: ABCC4 and ABCC5 were immunostained with the affinity-purified SNG and AMF antibodies, respectively. Immunostaining was
semiquantitatively scored as described in Materials and Methods as not detectable (�), low (+), moderate (++), or high (+++). Staining of capillary
endothelial cells was present (+) or not detectable (�). In some tumors, both labeled and unlabeled capillaries were found (+/�). Samples OII-5, OIII-4,OIII-7, and OAIII-8 were from patients who had received chemotherapy before surgery. c, patient deceased at time of final analysis.
Table 1. Clinical data of patients and ABCC4 and ABCC5 immunostaining in human glioma samples (Cont’d)
Histology (grade) Sample Age (y) Gender Time Time of ABCC4 expression in ABCC5 expression in
until tumorprogression
(mo)
survival(mo) Astrocytes Glioma
cells
Capillaries Astrocytes Glioma
cells
Capillaries
Membrane Transporters In Human Gliomas
www.aacrjournals.org 11421 Cancer Res 2005; 65: (24). December 15, 2005
Research. on June 5, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
and was scored as follows: �, no positive cells detectable; +, 1% to 25% oftumor cells positive; ++, 26% to 75% of tumor cells positive; +++, >75% of
tumor cells positive. ABCC immunostaining of vessels was scored as either
positive (+) or negative (�).
Preparation of crude membranes, immunoblot analysis, anddeglycosylation. Human glioma tissue (f100 mg) was homogenized
during thawing in 1 mL of lysis buffer (10 mmol/L Tris pH 7.4, 250 mmol/L
sucrose, supplemented with 0.1 mmol/L phenylmethylsulfonyl fluoride,
1 Amol/L pepstatin). Membrane fractions were collected by ultracentrifu-gation (100,000 � g , 4jC, 60 minutes) and suspended in 10 mmol/L Tris
buffer (pH 7.4). Crude membrane fractions were diluted with sample buffer
and incubated at 37jC for 30 minutes. Proteins were separated by SDS-
PAGE (7.5% separating gels) and blotted onto nitrocellulose membranesusing a tank blotting system (Bio-Rad). For detection of ABCC4, OATP1A2,
and OATP1C1, antisera were diluted in PBS/Tween 20 (1%) containing 5%
milk powder (1:1,000, 1:5,000, and 1:8,000, respectively). For detection ofOATP2B1, the affinity-purified SPA antibody was used in the same manner
at a dilution of 1:60. For peptide competition experiments, the antisera were
incubated at 4jC for 16 hours with 200 Amol/L of the synthetic peptide
used to generate the respective antiserum and then added to thenitrocellulose membrane. Horseradish peroxidase–conjugated secondary
antibodies were used at a dilution of 1:1,000 in PBS/Tween 20 (1%)
containing 5% milk powder. Deglycosylation was done as described (23).
RNA isolation and quantification of ABCC and SLCO mRNAs. Toinitially assess which transporter genes may be expressed in human
gliomas, total RNA was isolated from glioma samples and reverse
transcribed with an oligo(dT)18 primer. ABCC and SLCO mRNAs werequantified using the LightCycler system (Roche Diagnostics, Mannheim,
Germany) and ABCC primer pairs as described (6). The primer pair used for
b-actin mRNA amplification was from Stratagene (La Jolla, CA). SLCO sense
and antisense primer pairs were as follows: 5V-TGCCATACCTGGATA-TATGGTT-3Vand 5V-CAATTTAGTTTTCAATTCATCATCTT-3Vfor SLCO1A2 ,5V-CACCTCACATGTCATGCTGATT-3V and 5V-AACAATGTGTTTCAC-TATCTGCC-3V for SLCO1B1 , 5V-TCATAAACTCTTTGTTCTCTGCAA-3V and
5V-GTTGGCAGCAGCATTGTCTTG-3V for SLCO1B3 , 5V-CCTGGATACATAT-TACTTCTGAG-3V and 5V-CATGTTTCTAAAGTTGAGTTTCCT-3V for
SLCO1C1 , 5V-CGACTCAACGTGCAGCCATC-3V and 5V-CCGACACTAG-CAATTGCTGCT-3V for SLCO2B1 , and 5V-GAGACTGTAGCTGTATCCCTC-3Vand 5V-GCGGTGGTCAGACGCTGCT-3V for SLCO4A1 . The amount of therespective cDNA was determined by serial plasmid dilution (human ABCC2
cDNA or SLCO1A2 cDNA in the expression vector pcDNA3.1, from 1 � 106
to 1 � 102 fg), and ABCC and SLCO mRNA quantities were given as
percentage of the b-actin mRNA amount in the respective glioma sample.Finally, amplified PCR products were separated by agarose gel electropho-
resis, stained with ethidium bromide, and visualized under UV light.
Plasmids containing the respective ABCC and SLCO cDNA served as
positive controls.
Results
ABCC4 is predominantly expressed in endothelial andastrocytic glioma cells. Real-time PCR yielded ABCC4 mRNAquantities between 0.1% and 1.0% of b-actin mRNA (Table 2). ABCC4protein expression was analyzed in cryosections of the surgicalspecimens by immunostaining with the affinity-purified SNGantibody (Table 1; Fig. 1). ABCC4 was present in three differentcell types: in capillary endothelial cells, in astrocytes, and in gliomacells. In 53 samples (87%), ABCC4 was detectable in brain capillaries.This localization was confirmed by costaining with an antibodyspecific for the endothelial marker protein CD31 as shown in therepresentative images of Fig. 1A to C . Besides a cytoplasmic stainingin the endothelial cells, ABCC4 was localized to the luminalmembrane, which was identified by costaining for ABCG2 known tobe expressed on the luminal side of brain tumor endothelial cells(ref. 5; Fig. 1D). In a minority of tumors (8 of 61; 13%), braincapillaries did not label for ABCC4, which was particularly observed
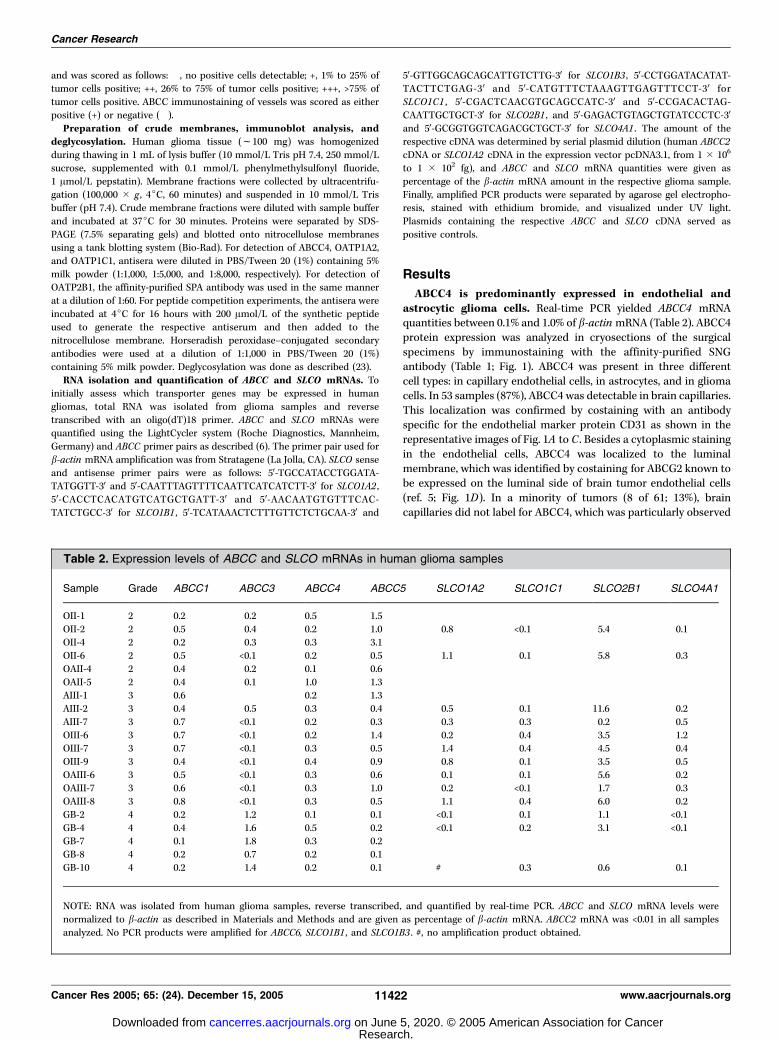
Table 2. Expression levels of ABCC and SLCO mRNAs in human glioma samples
Sample Grade ABCC1 ABCC3 ABCC4 ABCC5 SLCO1A2 SLCO1C1 SLCO2B1 SLCO4A1
OII-1 2 0.2 0.2 0.5 1.5
OII-2 2 0.5 0.4 0.2 1.0 0.8 <0.1 5.4 0.1OII-4 2 0.2 0.3 0.3 3.1
OII-6 2 0.5 <0.1 0.2 0.5 1.1 0.1 5.8 0.3
OAII-4 2 0.4 0.2 0.1 0.6
OAII-5 2 0.4 0.1 1.0 1.3AIII-1 3 0.6 0.2 1.3
AIII-2 3 0.4 0.5 0.3 0.4 0.5 0.1 11.6 0.2
AIII-7 3 0.7 <0.1 0.2 0.3 0.3 0.3 0.2 0.5
OIII-6 3 0.7 <0.1 0.2 1.4 0.2 0.4 3.5 1.2OIII-7 3 0.7 <0.1 0.3 0.5 1.4 0.4 4.5 0.4
OIII-9 3 0.4 <0.1 0.4 0.9 0.8 0.1 3.5 0.5
OAIII-6 3 0.5 <0.1 0.3 0.6 0.1 0.1 5.6 0.2OAIII-7 3 0.6 <0.1 0.3 1.0 0.2 <0.1 1.7 0.3
OAIII-8 3 0.8 <0.1 0.3 0.5 1.1 0.4 6.0 0.2
GB-2 4 0.2 1.2 0.1 0.1 <0.1 0.1 1.1 <0.1
GB-4 4 0.4 1.6 0.5 0.2 <0.1 0.2 3.1 <0.1GB-7 4 0.1 1.8 0.3 0.2
GB-8 4 0.2 0.7 0.2 0.1
GB-10 4 0.2 1.4 0.2 0.1 # 0.3 0.6 0.1
NOTE: RNA was isolated from human glioma samples, reverse transcribed, and quantified by real-time PCR. ABCC and SLCO mRNA levels were
normalized to b-actin as described in Materials and Methods and are given as percentage of b-actin mRNA. ABCC2 mRNA was <0.01 in all samples
analyzed. No PCR products were amplified for ABCC6, SLCO1B1 , and SLCO1B3 . #, no amplification product obtained.
Cancer Research
Cancer Res 2005; 65: (24). December 15, 2005 11422 www.aacrjournals.org
Research. on June 5, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
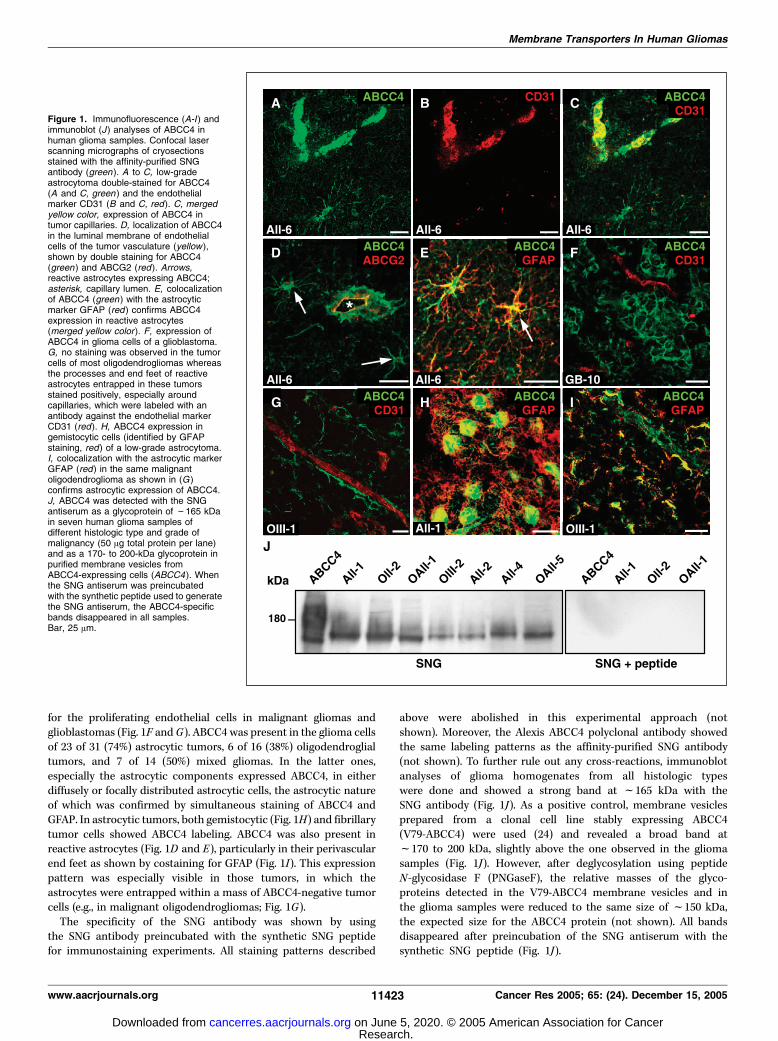
for the proliferating endothelial cells in malignant gliomas andglioblastomas (Fig. 1F and G). ABCC4 was present in the glioma cellsof 23 of 31 (74%) astrocytic tumors, 6 of 16 (38%) oligodendroglialtumors, and 7 of 14 (50%) mixed gliomas. In the latter ones,especially the astrocytic components expressed ABCC4, in eitherdiffusely or focally distributed astrocytic cells, the astrocytic natureof which was confirmed by simultaneous staining of ABCC4 andGFAP. In astrocytic tumors, both gemistocytic (Fig. 1H) and fibrillarytumor cells showed ABCC4 labeling. ABCC4 was also present inreactive astrocytes (Fig. 1D and E), particularly in their perivascularend feet as shown by costaining for GFAP (Fig. 1I). This expressionpattern was especially visible in those tumors, in which theastrocytes were entrapped within a mass of ABCC4-negative tumorcells (e.g., in malignant oligodendrogliomas; Fig. 1G).The specificity of the SNG antibody was shown by using
the SNG antibody preincubated with the synthetic SNG peptidefor immunostaining experiments. All staining patterns described
above were abolished in this experimental approach (notshown). Moreover, the Alexis ABCC4 polyclonal antibody showedthe same labeling patterns as the affinity-purified SNG antibody(not shown). To further rule out any cross-reactions, immunoblotanalyses of glioma homogenates from all histologic typeswere done and showed a strong band at f165 kDa with theSNG antibody (Fig. 1J). As a positive control, membrane vesiclesprepared from a clonal cell line stably expressing ABCC4(V79-ABCC4) were used (24) and revealed a broad band atf170 to 200 kDa, slightly above the one observed in the gliomasamples (Fig. 1J). However, after deglycosylation using peptideN-glycosidase F (PNGaseF), the relative masses of the glyco-proteins detected in the V79-ABCC4 membrane vesicles and inthe glioma samples were reduced to the same size of f150 kDa,the expected size for the ABCC4 protein (not shown). All bandsdisappeared after preincubation of the SNG antiserum with thesynthetic SNG peptide (Fig. 1J).
Figure 1. Immunofluorescence (A-I ) andimmunoblot (J ) analyses of ABCC4 inhuman glioma samples. Confocal laserscanning micrographs of cryosectionsstained with the affinity-purified SNGantibody (green ). A to C, low-gradeastrocytoma double-stained for ABCC4(A and C, green ) and the endothelialmarker CD31 (B and C, red). C, mergedyellow color, expression of ABCC4 intumor capillaries. D, localization of ABCC4in the luminal membrane of endothelialcells of the tumor vasculature (yellow ),shown by double staining for ABCC4(green ) and ABCG2 (red ). Arrows,reactive astrocytes expressing ABCC4;asterisk, capillary lumen. E, colocalizationof ABCC4 (green ) with the astrocyticmarker GFAP (red ) confirms ABCC4expression in reactive astrocytes(merged yellow color). F, expression ofABCC4 in glioma cells of a glioblastoma.G, no staining was observed in the tumorcells of most oligodendrogliomas whereasthe processes and end feet of reactiveastrocytes entrapped in these tumorsstained positively, especially aroundcapillaries, which were labeled with anantibody against the endothelial markerCD31 (red). H, ABCC4 expression ingemistocytic cells (identified by GFAPstaining, red) of a low-grade astrocytoma.I, colocalization with the astrocytic markerGFAP (red) in the same malignantoligodendroglioma as shown in (G )confirms astrocytic expression of ABCC4.J, ABCC4 was detected with the SNGantiserum as a glycoprotein of f165 kDain seven human glioma samples ofdifferent histologic type and grade ofmalignancy (50 Ag total protein per lane)and as a 170- to 200-kDa glycoprotein inpurified membrane vesicles fromABCC4-expressing cells (ABCC4). Whenthe SNG antiserum was preincubatedwith the synthetic peptide used to generatethe SNG antiserum, the ABCC4-specificbands disappeared in all samples.Bar, 25 Am.
Membrane Transporters In Human Gliomas
www.aacrjournals.org 11423 Cancer Res 2005; 65: (24). December 15, 2005
Research. on June 5, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
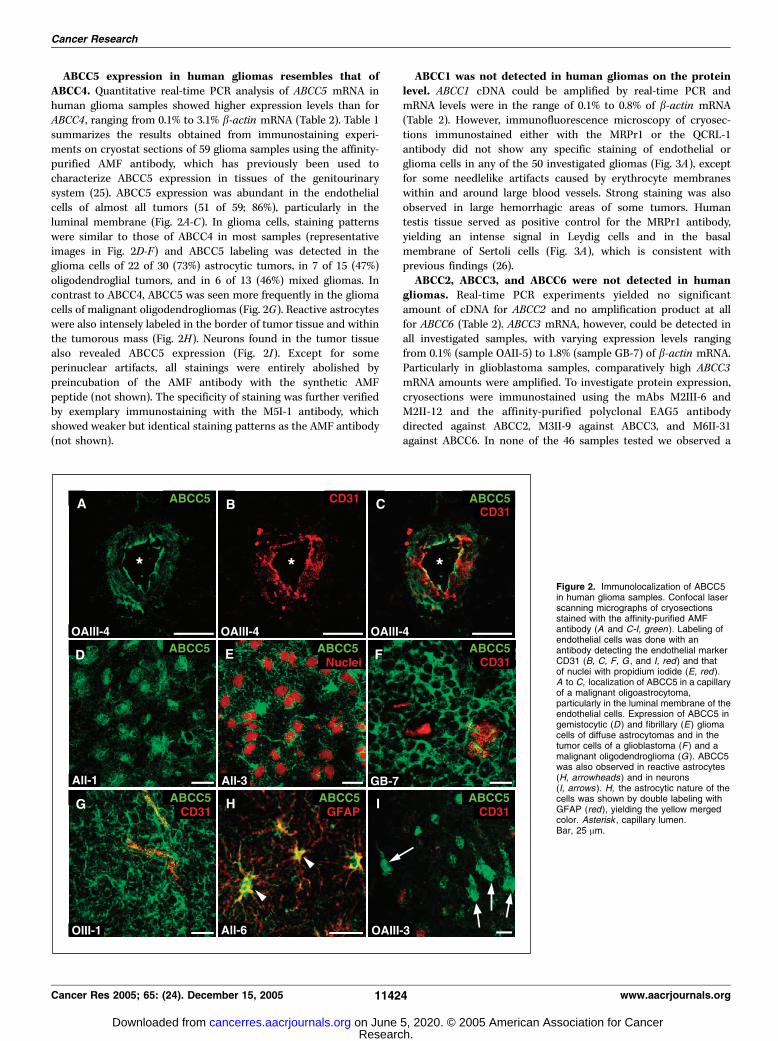
ABCC5 expression in human gliomas resembles that ofABCC4. Quantitative real-time PCR analysis of ABCC5 mRNA inhuman glioma samples showed higher expression levels than forABCC4 , ranging from 0.1% to 3.1% b-actin mRNA (Table 2). Table 1summarizes the results obtained from immunostaining experi-ments on cryostat sections of 59 glioma samples using the affinity-purified AMF antibody, which has previously been used tocharacterize ABCC5 expression in tissues of the genitourinarysystem (25). ABCC5 expression was abundant in the endothelialcells of almost all tumors (51 of 59; 86%), particularly in theluminal membrane (Fig. 2A-C). In glioma cells, staining patternswere similar to those of ABCC4 in most samples (representativeimages in Fig. 2D-F) and ABCC5 labeling was detected in theglioma cells of 22 of 30 (73%) astrocytic tumors, in 7 of 15 (47%)oligodendroglial tumors, and in 6 of 13 (46%) mixed gliomas. Incontrast to ABCC4, ABCC5 was seen more frequently in the gliomacells of malignant oligodendrogliomas (Fig. 2G). Reactive astrocyteswere also intensely labeled in the border of tumor tissue and withinthe tumorous mass (Fig. 2H). Neurons found in the tumor tissuealso revealed ABCC5 expression (Fig. 2I). Except for someperinuclear artifacts, all stainings were entirely abolished bypreincubation of the AMF antibody with the synthetic AMFpeptide (not shown). The specificity of staining was further verifiedby exemplary immunostaining with the M5I-1 antibody, whichshowed weaker but identical staining patterns as the AMF antibody(not shown).
ABCC1 was not detected in human gliomas on the proteinlevel. ABCC1 cDNA could be amplified by real-time PCR andmRNA levels were in the range of 0.1% to 0.8% of b-actin mRNA(Table 2). However, immunofluorescence microscopy of cryosec-tions immunostained either with the MRPr1 or the QCRL-1antibody did not show any specific staining of endothelial orglioma cells in any of the 50 investigated gliomas (Fig. 3A), exceptfor some needlelike artifacts caused by erythrocyte membraneswithin and around large blood vessels. Strong staining was alsoobserved in large hemorrhagic areas of some tumors. Humantestis tissue served as positive control for the MRPr1 antibody,yielding an intense signal in Leydig cells and in the basalmembrane of Sertoli cells (Fig. 3A), which is consistent withprevious findings (26).ABCC2, ABCC3, and ABCC6 were not detected in human
gliomas. Real-time PCR experiments yielded no significantamount of cDNA for ABCC2 and no amplification product at allfor ABCC6 (Table 2). ABCC3 mRNA, however, could be detected inall investigated samples, with varying expression levels rangingfrom 0.1% (sample OAII-5) to 1.8% (sample GB-7) of b-actin mRNA.Particularly in glioblastoma samples, comparatively high ABCC3mRNA amounts were amplified. To investigate protein expression,cryosections were immunostained using the mAbs M2III-6 andM2II-12 and the affinity-purified polyclonal EAG5 antibodydirected against ABCC2, M3II-9 against ABCC3, and M6II-31against ABCC6. In none of the 46 samples tested we observed a
Figure 2. Immunolocalization of ABCC5in human glioma samples. Confocal laserscanning micrographs of cryosectionsstained with the affinity-purified AMFantibody (A and C-I, green ). Labeling ofendothelial cells was done with anantibody detecting the endothelial markerCD31 (B, C, F, G , and I, red) and thatof nuclei with propidium iodide (E, red ).A to C, localization of ABCC5 in a capillaryof a malignant oligoastrocytoma,particularly in the luminal membrane of theendothelial cells. Expression of ABCC5 ingemistocytic (D ) and fibrillary (E) gliomacells of diffuse astrocytomas and in thetumor cells of a glioblastoma (F ) and amalignant oligodendroglioma (G). ABCC5was also observed in reactive astrocytes(H, arrowheads) and in neurons(I, arrows ). H, the astrocytic nature of thecells was shown by double labeling withGFAP (red), yielding the yellow mergedcolor. Asterisk , capillary lumen.Bar, 25 Am.
Cancer Research
Cancer Res 2005; 65: (24). December 15, 2005 11424 www.aacrjournals.org
Research. on June 5, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
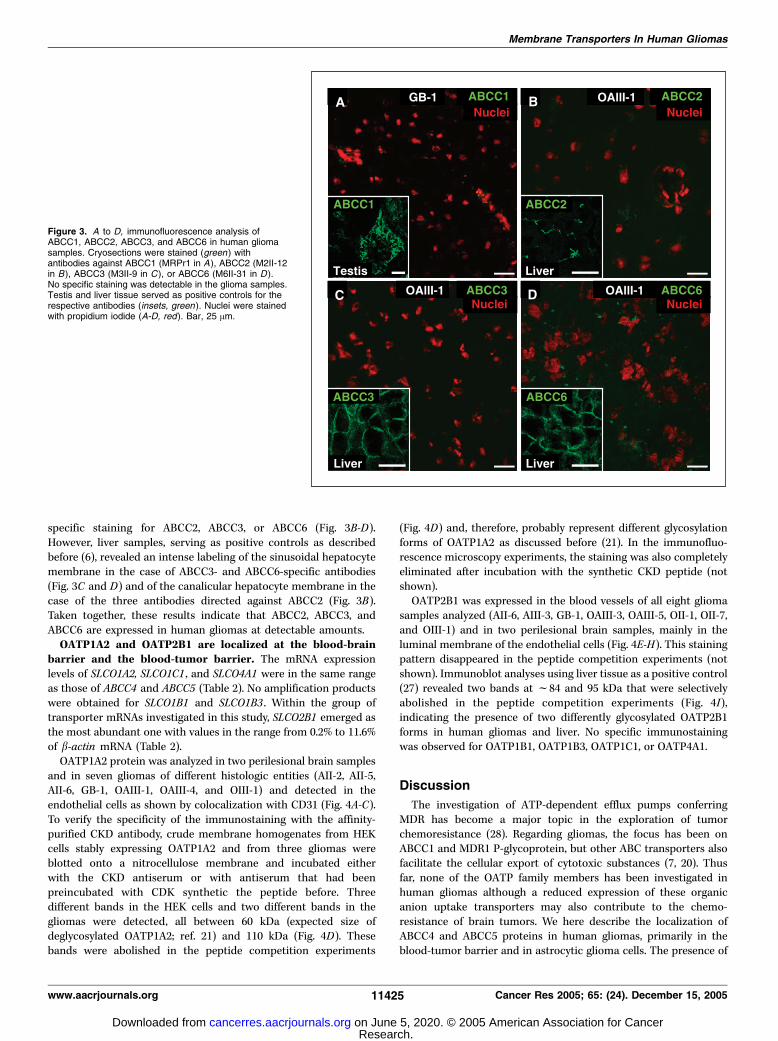
specific staining for ABCC2, ABCC3, or ABCC6 (Fig. 3B-D).However, liver samples, serving as positive controls as describedbefore (6), revealed an intense labeling of the sinusoidal hepatocytemembrane in the case of ABCC3- and ABCC6-specific antibodies(Fig. 3C and D) and of the canalicular hepatocyte membrane in thecase of the three antibodies directed against ABCC2 (Fig. 3B).Taken together, these results indicate that ABCC2, ABCC3, andABCC6 are expressed in human gliomas at detectable amounts.OATP1A2 and OATP2B1 are localized at the blood-brain
barrier and the blood-tumor barrier. The mRNA expressionlevels of SLCO1A2, SLCO1C1 , and SLCO4A1 were in the same rangeas those of ABCC4 and ABCC5 (Table 2). No amplification productswere obtained for SLCO1B1 and SLCO1B3 . Within the group oftransporter mRNAs investigated in this study, SLCO2B1 emerged asthe most abundant one with values in the range from 0.2% to 11.6%of b-actin mRNA (Table 2).OATP1A2 protein was analyzed in two perilesional brain samples
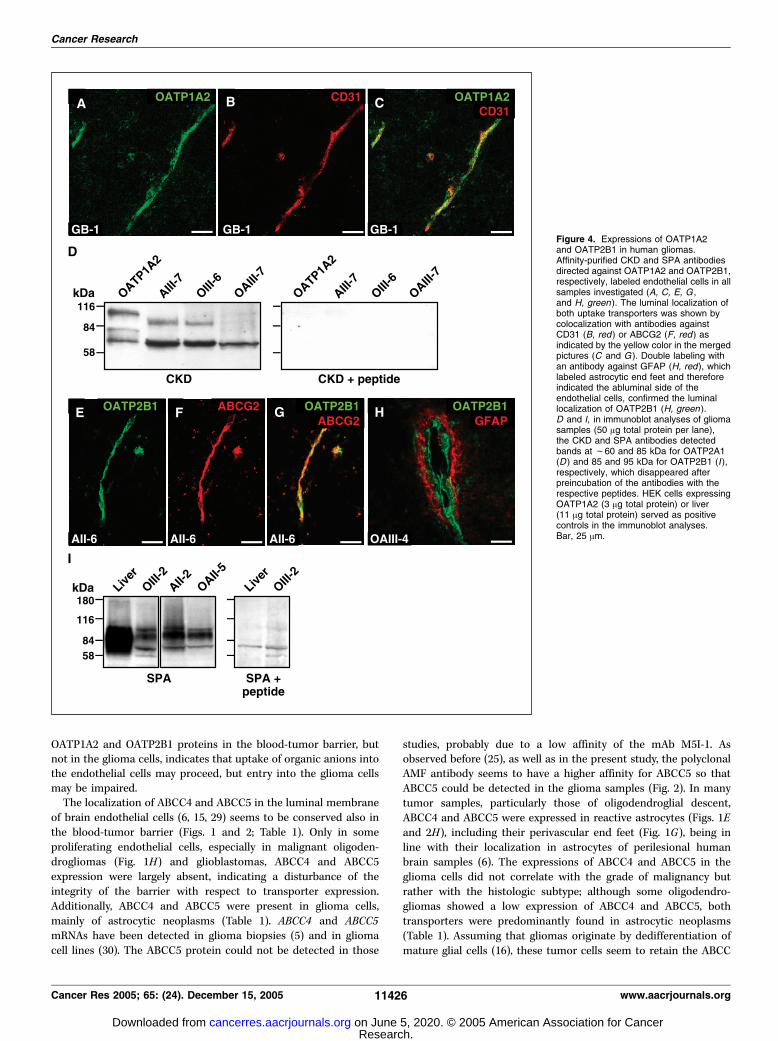
and in seven gliomas of different histologic entities (AII-2, AII-5,AII-6, GB-1, OAIII-1, OAIII-4, and OIII-1) and detected in theendothelial cells as shown by colocalization with CD31 (Fig. 4A-C).To verify the specificity of the immunostaining with the affinity-purified CKD antibody, crude membrane homogenates from HEKcells stably expressing OATP1A2 and from three gliomas wereblotted onto a nitrocellulose membrane and incubated eitherwith the CKD antiserum or with antiserum that had beenpreincubated with CDK synthetic the peptide before. Threedifferent bands in the HEK cells and two different bands in thegliomas were detected, all between 60 kDa (expected size ofdeglycosylated OATP1A2; ref. 21) and 110 kDa (Fig. 4D). Thesebands were abolished in the peptide competition experiments
(Fig. 4D) and, therefore, probably represent different glycosylationforms of OATP1A2 as discussed before (21). In the immunofluo-rescence microscopy experiments, the staining was also completelyeliminated after incubation with the synthetic CKD peptide (notshown).OATP2B1 was expressed in the blood vessels of all eight glioma
samples analyzed (AII-6, AIII-3, GB-1, OAIII-3, OAIII-5, OII-1, OII-7,and OIII-1) and in two perilesional brain samples, mainly in theluminal membrane of the endothelial cells (Fig. 4E-H). This stainingpattern disappeared in the peptide competition experiments (notshown). Immunoblot analyses using liver tissue as a positive control(27) revealed two bands at f84 and 95 kDa that were selectivelyabolished in the peptide competition experiments (Fig. 4I),indicating the presence of two differently glycosylated OATP2B1forms in human gliomas and liver. No specific immunostainingwas observed for OATP1B1, OATP1B3, OATP1C1, or OATP4A1.
Discussion
The investigation of ATP-dependent efflux pumps conferringMDR has become a major topic in the exploration of tumorchemoresistance (28). Regarding gliomas, the focus has been onABCC1 and MDR1 P-glycoprotein, but other ABC transporters alsofacilitate the cellular export of cytotoxic substances (7, 20). Thusfar, none of the OATP family members has been investigated inhuman gliomas although a reduced expression of these organicanion uptake transporters may also contribute to the chemo-resistance of brain tumors. We here describe the localization ofABCC4 and ABCC5 proteins in human gliomas, primarily in theblood-tumor barrier and in astrocytic glioma cells. The presence of
Figure 3. A to D, immunofluorescence analysis ofABCC1, ABCC2, ABCC3, and ABCC6 in human gliomasamples. Cryosections were stained (green ) withantibodies against ABCC1 (MRPr1 in A), ABCC2 (M2II-12in B), ABCC3 (M3II-9 in C ), or ABCC6 (M6II-31 in D ).No specific staining was detectable in the glioma samples.Testis and liver tissue served as positive controls for therespective antibodies (insets, green ). Nuclei were stainedwith propidium iodide (A-D, red). Bar, 25 Am.
Membrane Transporters In Human Gliomas
www.aacrjournals.org 11425 Cancer Res 2005; 65: (24). December 15, 2005
Research. on June 5, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
OATP1A2 and OATP2B1 proteins in the blood-tumor barrier, butnot in the glioma cells, indicates that uptake of organic anions intothe endothelial cells may proceed, but entry into the glioma cellsmay be impaired.The localization of ABCC4 and ABCC5 in the luminal membrane
of brain endothelial cells (6, 15, 29) seems to be conserved also inthe blood-tumor barrier (Figs. 1 and 2; Table 1). Only in someproliferating endothelial cells, especially in malignant oligoden-drogliomas (Fig. 1H) and glioblastomas, ABCC4 and ABCC5expression were largely absent, indicating a disturbance of theintegrity of the barrier with respect to transporter expression.Additionally, ABCC4 and ABCC5 were present in glioma cells,mainly of astrocytic neoplasms (Table 1). ABCC4 and ABCC5mRNAs have been detected in glioma biopsies (5) and in gliomacell lines (30). The ABCC5 protein could not be detected in those
studies, probably due to a low affinity of the mAb M5I-1. Asobserved before (25), as well as in the present study, the polyclonalAMF antibody seems to have a higher affinity for ABCC5 so thatABCC5 could be detected in the glioma samples (Fig. 2). In manytumor samples, particularly those of oligodendroglial descent,ABCC4 and ABCC5 were expressed in reactive astrocytes (Figs. 1Eand 2H), including their perivascular end feet (Fig. 1G), being inline with their localization in astrocytes of perilesional humanbrain samples (6). The expressions of ABCC4 and ABCC5 in theglioma cells did not correlate with the grade of malignancy butrather with the histologic subtype; although some oligodendro-gliomas showed a low expression of ABCC4 and ABCC5, bothtransporters were predominantly found in astrocytic neoplasms(Table 1). Assuming that gliomas originate by dedifferentiation ofmature glial cells (16), these tumor cells seem to retain the ABCC
Figure 4. Expressions of OATP1A2and OATP2B1 in human gliomas.Affinity-purified CKD and SPA antibodiesdirected against OATP1A2 and OATP2B1,respectively, labeled endothelial cells in allsamples investigated (A, C, E, G ,and H, green). The luminal localization ofboth uptake transporters was shown bycolocalization with antibodies againstCD31 (B, red ) or ABCG2 (F, red ) asindicated by the yellow color in the mergedpictures (C and G ). Double labeling withan antibody against GFAP (H, red), whichlabeled astrocytic end feet and thereforeindicated the abluminal side of theendothelial cells, confirmed the luminallocalization of OATP2B1 (H, green ).D and I, in immunoblot analyses of gliomasamples (50 Ag total protein per lane),the CKD and SPA antibodies detectedbands at f60 and 85 kDa for OATP2A1(D ) and 85 and 95 kDa for OATP2B1 (I ),respectively, which disappeared afterpreincubation of the antibodies with therespective peptides. HEK cells expressingOATP1A2 (3 Ag total protein) or liver(11 Ag total protein) served as positivecontrols in the immunoblot analyses.Bar, 25 Am.
Cancer Research
Cancer Res 2005; 65: (24). December 15, 2005 11426 www.aacrjournals.org
Research. on June 5, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

expression pattern of the original cell. Otherwise, if gliomas emergefrom the transformation of an immature precursor (31), theexpression of ABCC4 and ABCC5 may accompany a differentiationtowards an astrocytic phenotype. This may also account for theirexpression primarily in the astrocytic portions of mixed oligoas-trocytomas, of which oligodendroglial and astrocytic portions arethought to derive from a single common progenitor cell (32). Thelow expressions of ABCC4 and ABCC5 also in some oligodendrog-lial tumor cells (Table 1) may then be explained as an earlier stageof differentiation of these cells, as shown for the expression of otherastrocytic proteins in oligodendroglioma cells (33).ABCC4 and ABCC5 are the major ABCC subfamily members in
human gliomas identified in this study (Figs. 1 and 2; Table 1). Theirpresence in the blood-tumor barrier and in glioma cells rendersthem candidate transporters that could contribute to the intrinsicMDR phenotype (13, 34, 35) of gliomas. Methotrexate transport byABCC4 and ABCC4-mediated resistance to ganciclovir and top-otecan has been shown (15, 36, 37). A recent study (15) stronglysupports the importance of Abcc4 in the blood-brain barrier as wellas in the blood-cerebrospinal fluid barrier by showing an enhancedaccumulation of topotecan in the brain and the cerebrospinal fluidof Abcc4-deficient mice. In contrast, ABCC1, ABCC2, ABCC3, andABCC6 are apparently not expressed in relevant amounts (Fig. 3) sothat their role in the intrinsic chemoresistance of gliomas may belimited. We did not observe any specific ABCC1 immunoreactivityin gliomas, being in line with other studies (5, 38). However, somestudies showed ABCC1 expression in human gliomas with a varyingnumber of positive tumor cells, few of them also in the blood-tumorbarrier (9, 11, 12, 39–41). Several other studies did not detect ABCC1in brain at all (5, 10, 41, 42). Our recent demonstration of ABCC1 inendothelial cells of perilesional human brain (6), but not in theblood-tumor barrier (Fig. 3), indicates a lower expression of ABCC1in the blood-tumor barrier than in the blood-brain barrier. This maybe due to the loss of modulating expression factors released fromastrocytes or from other cells of non-neoplastic brain. By usingthree different antibodies, we could not detect ABCC2 in the humanglioma samples. Our results are in agreement with previous studiesin which ABCC2 mRNA or ABCC2 protein was not detected insignificant amounts in normal or perilesional brain tissue (6, 10) incultured endothelial or parenchymal cells of different species (29,43, 44) or in glioma tissue and glioma cells (45, 46). Similarly, ABCC3was not detectable in any of the samples, which is in accordancewith earlier studies (5, 38). Presence of ABCC3 in glioma cell lines(30, 46) indicates a discrepancy between protein detectability inglioma biopsies and glioma cell lines, which might be caused bylong-term culturing conditions as proposed for ABCC1 (38).Besides ABCC4 and ABCC5 (Figs. 1 and 2), other ABC
transporters such as MDR1 P-glycoprotein (47) and ABCG2 (5)
are present in the blood-tumor barrier. Although not systematicallyexamined in the present study, we also observed staining forABCG2 exclusively in the blood-tumor barrier, but not in theglioma cells, and therefore we used it as a marker for the luminalmembrane of endothelial cells (Figs. 1 and 4). Most recently,Aronica et al. (48) reported similar findings. Presence of ABCG2 inthe blood-tumor barrier may also contribute to the chemo-resistance of gliomas.In addition to ATP-dependent export pumps, the blood-brain
barrier and the blood-tumor barrier comprise the uptaketransporters OATP1A2 and OATP2B1 (Fig. 4) whereas OATP1C1,OATP1B1, OATP1B3, and OATP4A1 were not detectable byimmunofluorescence microscopy. Only the localization ofOATP1A2 in the blood-brain barrier had been described before(21). The observed expression pattern, with OATP1A2 andOATP2B1 present in the blood-tumor barrier and all of the sixinvestigated OATP family members absent in the gliomacells, does not fully rule out the existence of other path-ways of transporter-mediated drug uptake [e.g., by additionalmembers of the OATP family (19) or by members of the SLC22family (49)].In conclusion, localization of OATP1A2 and OATP2B1 in the
blood-tumor barrier indicates that organic anions can be taken upinto the endothelial cells, however, other transporters for theuptake of organic anions into endothelial and glioma cells mayexist. The presence of ABCC4 and ABCC5 proteins in theendothelial and in the astrocytic glioma cells suggests thatexpression of both proteins is a characteristic of the astrocyticphenotype in human gliomas and that both could participate in thedrug resistance of these tumors. In view of the divergent resultsbetween the in vivo and in vitro expressions of drug transporters,this study provides a basis for the assessment of results previouslyobtained from cell culture experiments in comparison with ourresults from frozen human glioma tissue.
Acknowledgments
Received 4/13/2005; revised 8/2/2005; accepted 9/30/2005.Grant support: Tumorzentrum Heidelberg/Mannheim (A.T. Nies and H.-H.
Steiner), Wilhelm-Sander-Stiftung Munchen (A.T. Nies, H.-H. Steiner, and C. Herold-Mende), research collaboration between the German Cancer Research Center andPfizer Research Laboratories, Groton, Connecticut (D. Keppler), Deutsche Forschungs-gemeinschaft (J. Konig and D. Keppler).
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Dr. Horst-Peter Schmitt (Institute of Pathology, University of Heidelberg)for helpful discussions, Dr. Herbert Spring (Division of Cell Biology, German CancerResearch Center) for expert help in confocal laser scanning microscopy, Dr. Maria Rius(Division of Tumor Biochemistry, German Cancer Research Center) for providingABCC4 -transfected V79 cells, and Elke Herrmann and Jessica Longin for excellenttechnical assistance.
References1. Carpentier AF. Neuro-oncology: the growing role ofchemotherapy in glioma. Lancet Neurol 2005;4:4–5.
2. Bredel M. Anticancer drug resistance in primaryhuman brain tumors. Brain Res Brain Res Rev 2001;35:161–204.
3. Dean M, Rzhetsky A, Allikmets R. The human ATP-binding cassette (ABC) transporter superfamily. GenomeRes 2001;11:1156–66.
4. Schinkel AH. P-glycoprotein, a gatekeeper in theblood-brain barrier. Adv Drug Deliv Rev 1999;36:179–94.
5. Cooray HC, Blackmore CG, Maskell L, Barrand MA.
Localisation of breast cancer resistance protein in micro-vessel endothelium of human brain. Neuroreport 2002;13:2059–63.
6. Nies AT, Jedlitschky G, Konig J, et al. Expression andimmunolocalization of the multidrug resistance pro-teins, MRP1-MRP6 (ABCC1-ABCC6), in human brain.Neuroscience 2004;129:349–60.
7. Haimeur A, Conseil G, Deeley RG, Cole SP. The MRP-related and BCRP/ABCG2 multidrug resistance proteins:biology, substrate specificity and regulation. Curr DrugMetab 2004;5:21–53.
8. Sawada T, Kato Y, Sakayori N, Takekawa Y, KobayashiM. Expression of the multidrug-resistance P-glycoprotein
(Pgp, MDR-1) by endothelial cells of the neovasculaturein central nervous system tumors. Brain Tumor Pathol1999;16:23–7.
9. Spiegl-Kreinecker S, Buchroithner J, Elbling L, et al.Expression and functional activity of the ABC-trans-porter proteins P-glycoprotein and multidrug-resistanceprotein 1 in human brain tumor cells and astrocytes.J Neurooncol 2002;57:27–36.
10. Aronica E, Gorter JA, Jansen GH, et al. Expressionand cellular distribution of multidrug transporterproteins in two major causes of medically intractableepilepsy: focal cortical dysplasia and glioneuronaltumors. Neuroscience 2003;118:417–29.
Membrane Transporters In Human Gliomas
www.aacrjournals.org 11427 Cancer Res 2005; 65: (24). December 15, 2005
Research. on June 5, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

11. Andersson U, Malmer B, Bergenheim AT, BrannstromT, Henriksson R. Heterogeneity in the expression ofmarkers for drug resistance in brain tumors. ClinNeuropathol 2004;23:21–7.
12. Benyahia B, Huguet S, Decleves X, et al. Multidrugresistance-associated protein MRP1 expression in hu-man gliomas: chemosensitization to vincristine andetoposide by indomethacin in human glioma cell linesoverexpressing MRP1. J Neurooncol 2004;66:65–70.
13. Wijnholds J, Mol CA, van Deemter L, et al. Multidrug-resistance protein 5 is a multispecific organic aniontransporter able to transport nucleotide analogs. ProcNatl Acad Sci U S A 2000;97:7476–81.
14. Lee K, Klein-Szanto AJ, Kruh GD. Analysis of theMRP4 drug resistance profile in transfected NIH3T3cells. J Natl Cancer Inst 2001;92:1934–40.
15. Leggas M, Adachi M, Scheffer GL, et al. Mrp4 confersresistance to topotecan and protects the brain fromchemotherapy. Mol Cell Biol 2004;24:7612–21.
16. Sanson M, Thillet J, Hoang-Xuan K. Molecularchanges in gliomas. Curr Opin Oncol 2004;16:607–13.
17. Mikkaichi T, Suzuki T, Onogawa T, et al. Isolation andcharacterization of a digoxin transporter and its rathomologue expressed in the kidney. Proc Natl Acad SciU S A 2004;101:3569–74.
18. Nozawa T, Minami H, Sugiura S, Tsuji A, Tamai I. Roleof organic anion transporter OATP1B1 (OATP-C) inhepatic uptake of irinotecan and its active metaboliteSN-38: in vitro evidence and effect of single nucleotidepolymorphisms. Drug Metab Dispos 2005;33:434–9.
19. Hagenbuch B, Meier PJ. Organic anion transportingpolypeptides of the OATP/SLC21 family: phylogeneticclassification as OATP/SLCO superfamily, new nomen-clature and molecular/functional properties. PflugersArch 2004;447:653–65.
20. Kusuhara H, Sugiyama Y. Active efflux across theblood-brain barrier: role of the solute carrier family.NeuroRx 2005;2:73–85.
21. Gao B, Hagenbuch B, Kullak-Ublick GA, Benke D,Aguzzi A, Meier PJ. Organic anion-transportingpolypeptides mediate transport of opioid peptidesacross blood-brain barrier. J Pharmacol Exp Ther 2000;294:73–9.
22. Mayer R, Kartenbeck J, Buchler M, Jedlitschky G,Leier I, Keppler D. Expression of the MRP gene-encodedconjugate export pump in liver and its selective absencefrom the canalicular membrane in transport-deficientmutant hepatocytes. J Cell Biol 1995;131:137–50.
23. Konig J, Cui Y, Nies AT, Keppler D. A novel humanorganic anion transporting polypeptide localized to thebasolateral hepatocyte membrane. Am J Physiol Gastro-intest Liver Physiol 2000;278:G156–64.
24. Rius M, Nies AT, Hummel-Eisenbeiss J, Jedlitschky G,Keppler D. Cotransport of reduced glutathione with bilesalts by MRP4 (ABCC4) localized to the basolateralhepatocyte membrane. Hepatology 2003;38:374–84.
25. Nies AT, Spring H, Thon WF, Keppler D, JedlitschkyG. Immunolocalization of multidrug resistance protein 5(ABCC5) in human genitourinary system. J Urol 2002;167:2271–5.
26. Wijnholds J, Scheffer GL, van d V, et al. Multidrugresistance protein 1 protects the oropharyngeal mucosallayer and the testicular tubules against drug-induceddamage. J Exp Med 1998;188:797–808.
27. Kullak-Ublick GA, Ismair MG, Stieger B, et al. Organicanion-transporting polypeptide B (OATP-B) and itsfunctional comparison with three other OATPs ofhuman liver. Gastroenterology 2001;120:525–33.
28. Gottesman MM, Fojo T, Bates SE. Multidrug resis-tance in cancer: role of ATP-dependent transporters.Nat Rev Cancer 2002;2:48–58.
29. Zhang Y, Schuetz JD, Elmquist WF, Miller DW.Plasma membrane localization of multidrug resis-tance-associated protein homologs in brain capillaryendothelial cells. J Pharmacol Exp Ther 2004;311:449–55.
30. Decleves X, Fajac A, Lehmann-Che J, et al. Molecularand functional MDR1-Pgp and MRPs expression inhuman glioblastoma multiforme cell lines. Int J Cancer2002;98:173–80.
31. Noble M, Dietrich J. The complex identity of braintumors: emerging concerns regarding origin, diversityand plasticity. Trends Neurosci 2004;27:148–54.
32. Shoshan Y, Nishiyama A, Chang A, et al. Expression ofoligodendrocyte progenitor cell antigens by gliomas:implications for the histogenesis of brain tumors. ProcNatl Acad Sci U S A 1999;96:10361–6.
33. Popko B, Pearl DK, Walker DM, et al. Molecularmarkers that identify human astrocytomas and oligo-dendrogliomas. J Neuropathol Exp Neurol 2002;61:329–38.
34. Chen ZS, Lee K, Kruh GD. Transport of cyclicnucleotides and estradiol 17-h-D-glucuronide bymultidrug resistance protein 4. Resistance to 6-mercap-topurine and 6-thioguanine. J Biol Chem 2001;276:33747–54.
35. Wielinga PR, Reid G, Challa EE, et al. Thiopurinemetabolism and identification of the thiopurine metab-olites transported by MRP4 and MRP5 overexpressed inhuman embryonic kidney cells. Mol Pharmacol 2002;62:1321–31.
36. Chen ZS, Lee K, Walther S, et al. Analysis ofmethotrexate and folate transport by multidrug resis-tance protein 4 (ABCC4): MRP4 is a component of themethotrexate efflux system. Cancer Res 2002;62:3144–50.
37. Adachi M, Sampath J, Lan LB, et al. Expression ofMRP4 confers resistance to ganciclovir and compro-mises bystander cell killing. J Biol Chem 2002;277:38998–9004.
38. Bahr O, Rieger J, Duffner F, Meyermann R, WellerM, Wick W. P-glycoprotein and multidrug resistance-associated protein mediate specific patterns ofmultidrug resistance in malignant glioma cell lines,but not in primary glioma cells. Brain Pathol 2003;13:482–94.
39. Abe T, Mori T, Wakabayashi Y, et al. Expression ofmultidrug resistance protein gene in patients withglioma after chemotherapy. J Neurooncol 1998;40:11–8.
40. Mohri M, Nitta H, Yamashita J. Expression ofmultidrug resistance-associated protein (MRP) in hu-man gliomas. J Neurooncol 2000;49:105–15.
41. Tews DS, Nissen A, Kulgen C, Gaumann AK. Drugresistance-associated factors in primary and secondaryglioblastomas and their precursor tumors. J Neurooncol2000;50:227–37.
42. Sisodiya SM, Lin WR, Harding BN, Squier MV, ThomM. Drug resistance in epilepsy: expression of drugresistance proteins in common causes of refractoryepilepsy. Brain 2002;125:22–31.
43. Berezowski V, Landry C, Dehouck MP, CecchelliR, Fenart L. Contribution of glial cells and peri-cytes to the mRNA profiles of P-glycoprotein andmultidrug resistance-associated proteins in an in vitromodel of the blood-brain barrier. Brain Res 2004;1018:1–9.
44. Hirrlinger J, Konig J, Dringen R. Expression of mRNAsof multidrug resistance proteins (Mrps) in cultured ratastrocytes, oligodendrocytes, microglial cells and neuro-nes. J Neurochem 2002;82:716–9.
45. Decleves X, Regina A, Laplanche JL, et al. Functionalexpression of P-glycoprotein and multidrug resistance-associated protein (Mrp1) in primary cultures of ratastrocytes. J Neurosci Res 2000;60:594–601.
46. Haga S, Hinoshita E, Ikezaki K, et al. Involvement ofthe multidrug resistance protein 3 in drug sensitivityand its expression in human glioma. Jpn J Cancer Res2001;92:211–9.
47. Becker I, Becker KF, Meyermann R, Hollt V. Themultidrug-resistance gene MDR1 is expressed inhuman glial tumors. Acta Neuropathol (Berl) 1991;82:516–9.
48. Aronica E, Gorter JA, Redeker S, et al. Localization ofbreast cancer resistance protein (BCRP) in microvesselendothelium of human control and epileptic brain.Epilepsia 2005;46:849–57.
49. Koepsell H, Endou H. The SLC22 drug transporterfamily. Pflugers Arch 2004;447:666–76.
Cancer Research
Cancer Res 2005; 65: (24). December 15, 2005 11428 www.aacrjournals.org
Research. on June 5, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

2005;65:11419-11428. Cancer Res Holger Bronger, Jörg König, Kathrin Kopplow, et al. BarrierTransporters in Human Gliomas and the Blood-Tumor ABCC Drug Efflux Pumps and Organic Anion Uptake
Updated version
http://cancerres.aacrjournals.org/content/65/24/11419
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/65/24/11419.full#ref-list-1
This article cites 48 articles, 15 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/65/24/11419.full#related-urls
This article has been cited by 23 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://cancerres.aacrjournals.org/content/65/24/11419To request permission to re-use all or part of this article, use this link
Research. on June 5, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from