adenosine triphosphate (atp)fac.ksu.edu.sa/sites/default/files/4-_adenosine_triphosphate.pdf ·...
TRANSCRIPT

Adenosine triphosphate (ATP)
1
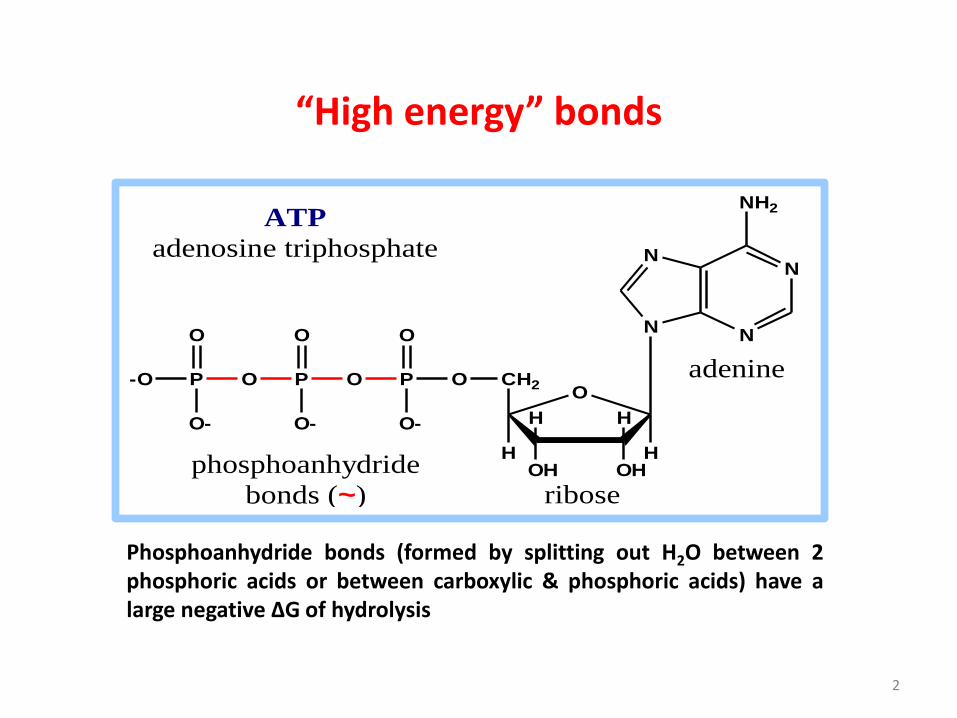
“High energy” bonds
2
N
NN
N
NH2
O
OHOH
HH
H
CH2
H
OPOPOP-O
O
O- O-
O O
O-
adenine
ribose
ATP
adenosine triphosphate
phosphoanhydride
bonds (~)
Phosphoanhydride bonds (formed by splitting out H2O between 2phosphoric acids or between carboxylic & phosphoric acids) have alarge negative ΔG of hydrolysis

• Phosphoanhydride linkages are said to be "high energy"
bonds. Bond energy is not high, just ΔG of hydrolysis. "High
energy" bonds are represented by the "~" symbol. ~P
represents a phosphate group with a large negative ΔG of
hydrolysis.
• Compounds with “high energy bonds” are said to have high
group transfer potential. For example, Pi may be
spontaneously cleaved from ATP for transfer to another
compound (e.g., to a hydroxyl group on glucose).
3

Potentially, 2 ~P bonds can be cleaved, as 2 phosphates are
released by hydrolysis from ATP.
AMP~P~P AMP~P + Pi (ATP ADP + Pi)
AMP~P AMP + Pi (ADP AMP + Pi)
Alternatively:
AMP~P~P AMP + P~P (ATP AMP + PPi)
P~P 2 Pi (PPi 2Pi)

• ATP often serves as an energy source.
Hydrolytic cleavage of one or both of the "high energy"bonds of ATP is coupled to an energy-requiring reaction.AMP functions as an energy sensor & regulator ofmetabolism.
When ATP production does not keep up with needs, ahigher portion of a cell's adenine nucleotide pool is AMP.
AMP stimulates metabolic pathways that produce ATP.
• Some examples of this role involve direct allostericactivation of pathway enzymes by AMP.
• Some regulatory effects of AMP are mediated by theenzyme AMP-Activated Protein Kinase.

A reaction important for equilibrating ~P among adenine nucleotides
within a cell is that catalyzed by Adenylate Kinase:
ATP + AMP 2 ADP
The Adenylate Kinase reaction is also important because the substrate for
ATP synthesis, e.g., by mitochondrial ATP Synthase, is ADP, while some
cellular reactions dephosphorylate ATP all the way to AMP.
The enzyme Nucleoside Diphosphate Kinase equilibrates ~P among the
various nucleotides that are needed, e.g., for synthesis of DNA & RNA.
NuDiKi catalyzes reversible reactions such as:
ATP + GDP ADP + GTP,
ATP + UDP ADP + UTP, etc.

Inorganic polyphosphate
Many organisms store energy as inorganic polyphosphate, a chain of
many phosphate residues linked by phosphoanhydride bonds:
P~P~P~P~P...
Hydrolysis of Pi residues from polyphosphate may be coupled to energy-
dependent reactions.
Depending on the organism or cell type, inorganic polyphosphate may
have additional functions.
E.g., it may serve as a reservoir for Pi, a chelator of metal ions, a buffer,
or a regulator.

Why do phosphoanhydride linkages have a high ΔG of
hydrolysis? Contributing factors for ATP & PPi include:
• Resonance stabilization of products of hydrolysis exceeds
resonance stabilization of the compound itself.
• Electrostatic repulsion between negatively charged
phosphate oxygen atoms favors separation of the
phosphates.

Phosphoryl group transfer and ATP
• Living cells obtain free energy in a chemical form
by the catabolism of nutrient molecules
• They use that energy to make ATP from ADP and
Pi .
• ATP donates some of its chemical energy to
1. Endergonic processes such as the synthesis of
metabolic intermediates and macromolecules from
smaller precursors
9

2. The transport of substances across membranes
against concentration gradients
3. Mechanical motion (muscle contraction)
This donation of energy from ATP can occur in the two
form
A) ATP ADP+ Pi or
B) ATP AMP+ 2 Pi
10
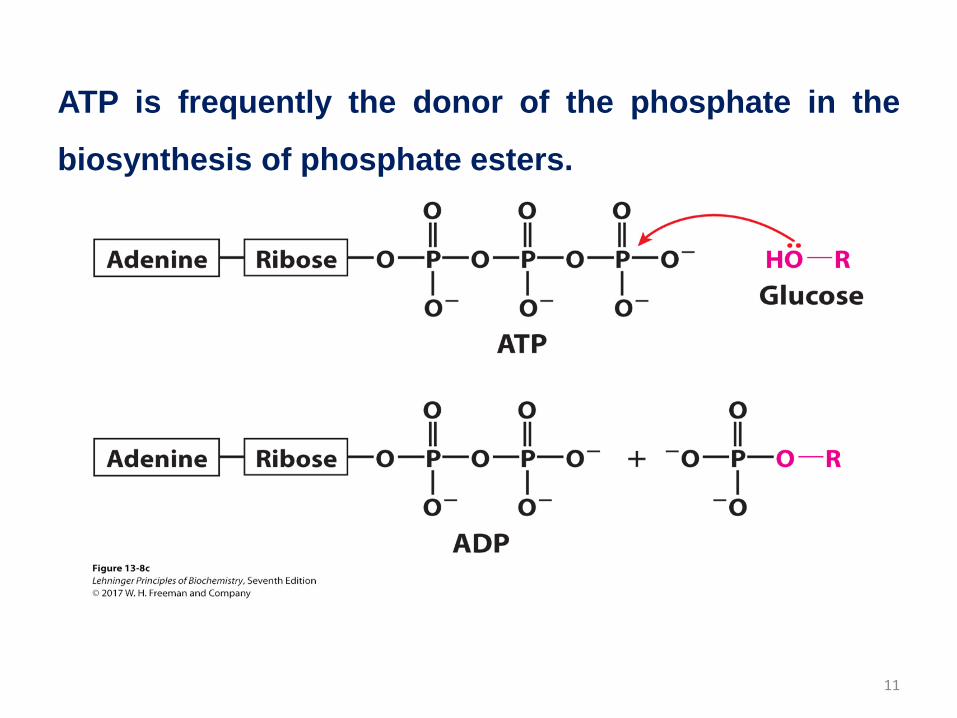
ATP is frequently the donor of the phosphate in the
biosynthesis of phosphate esters.
11

The free energy change for ATP hydrolysis is large and
negative
12

Chemical basis for the large free-energy change
associated with ATP hydrolysis.
1. The charge separation that results from hydrolysis
relieves electrostatic repulsion among the four negative
charges on ATP.
2. The product inorganic phosphate (Pi) is stabilized by
formation of a resonance hybrid, in which each of the
four phosphorus–oxygen bonds has the same degree
of double-bond character and the hydrogen ion is not
permanently associated with any one of the oxygen's.
13

(Some degree of resonance stabilization also occurs in
phosphates involved in ester or anhydride linkages,
but fewer resonance forms are possible than for Pi)
3. A third factor that favors ATP hydrolysis is the
greater degree of solvation (hydration) of the products
Pi and ADP relative to ATP, which further stabilizes the
products relative to the reactants.
14

• Although the hydrolysis of ATP is highly exergonic
(ΔG°´ = -30,5 kJ/mol), the ATP is stable at pH 7,
because the activation energy for ATP hydrolysis is
relatively high. Rapid hydrolysis of ATP occurs only
when catalyzed by an enzyme.
• The free energy change for ATP hydrolysis is -30,5
kJ/mol under standard conditions but the actual free
energy change (ΔG) of ATP hydrolysis in living cells
is very different.
15

• The cellular concentrations of ATP, ADP and Pi are
not same and are much lower than the 1 M standard
conditions.
• In addition, Mg2+ in the cytosol binds to ATP and ADP
and for most enzymatic reactions that involve ATP as
phosphoryl group donor, the true substrate is
MgATP-2. The relevant ΔG°´ is therefore that for
MgATP-2 hydrolysis.
16
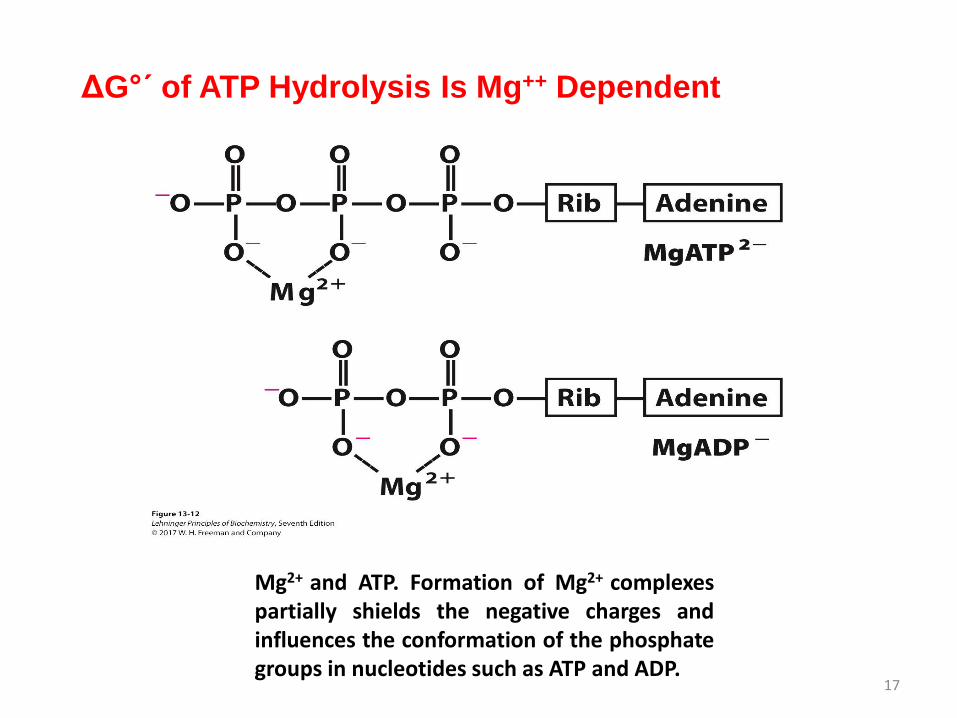
ΔG°´ of ATP Hydrolysis Is Mg++ Dependent
17
Mg2+ and ATP. Formation of Mg2+ complexespartially shields the negative charges andinfluences the conformation of the phosphategroups in nucleotides such as ATP and ADP.

Phosphorylated compounds
• Phosphoenolpyruvate
• 1,3-bisphosphoglycerate
• Phosphocreatine
• Thioesters
• ATP
• AMP
• PPi
• Glucose 1-phosphate
• Fructose 6-phosphate
• Glucose 6-phosphate
18
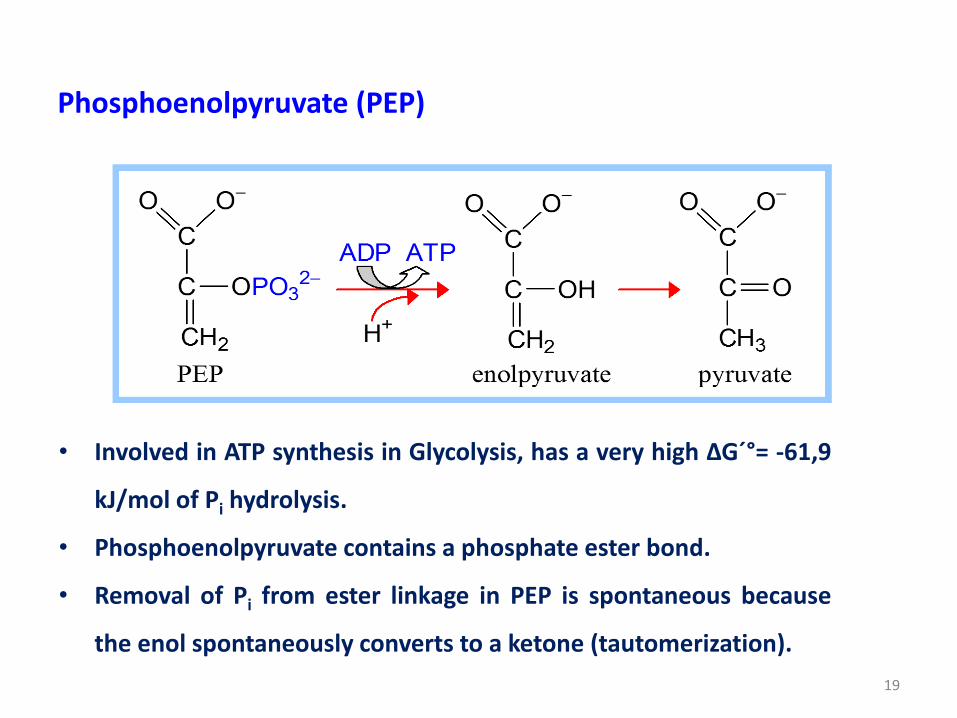
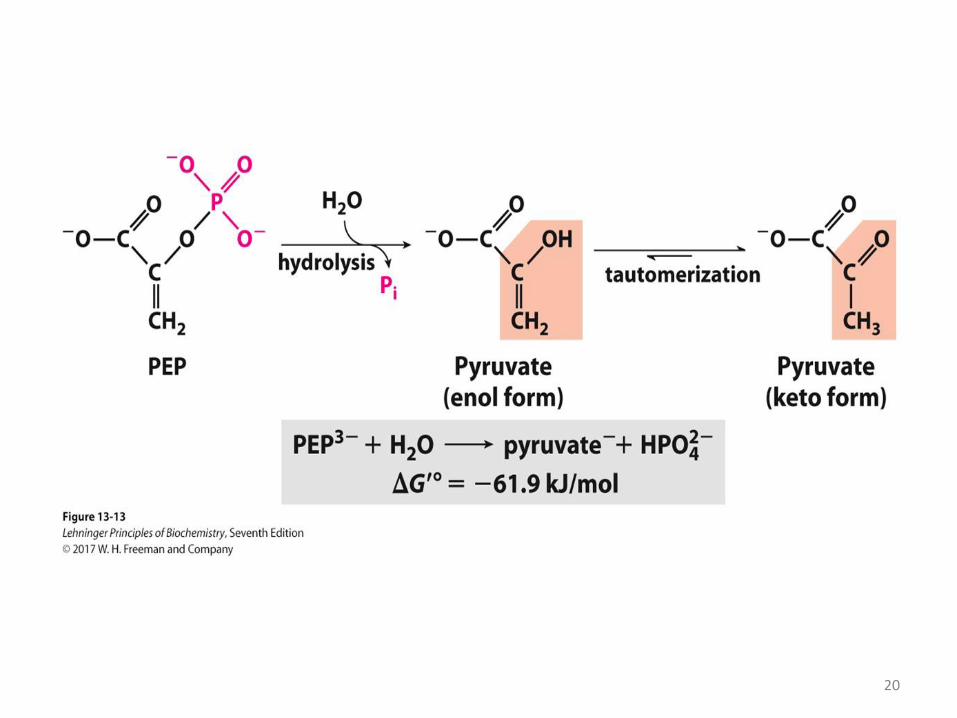
Phosphoenolpyruvate (PEP)
19
• Involved in ATP synthesis in Glycolysis, has a very high ΔG´°= -61,9
kJ/mol of Pi hydrolysis.
• Phosphoenolpyruvate contains a phosphate ester bond.
• Removal of Pi from ester linkage in PEP is spontaneous because
the enol spontaneously converts to a ketone (tautomerization).
20

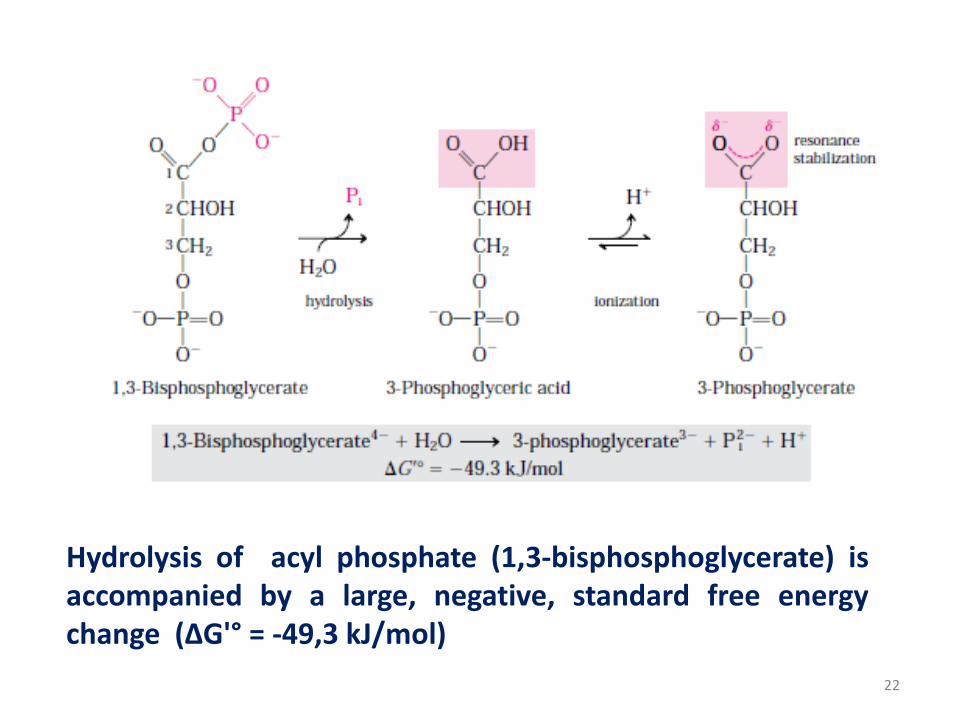
1,3-bisphosphoglycerate
• 1,3-bisphosphoglycerate contains an anhydride bond between the
carboxyl group at C-1 and phosphoric acid.
• The direct product of hydrolysis 1,3-bisphosphoglycerate is 3-
phosphoglyceric acid, with an undissociated carboxylic acid group,
but dissociation occurs immediately. This ionization and the
resonance structures it makes possible stabilize the product
relative to the reactants. Resonance stabilization of Pi further
contributes to the negative free energy change.
21
22
Hydrolysis of acyl phosphate (1,3-bisphosphoglycerate) isaccompanied by a large, negative, standard free energychange (ΔG'° = -49,3 kJ/mol)

Phosphocreatine
The P-N bond can be hydrolyzed to generate free creatine and
Pi. The release of Pi and the resonance stabilization of creatine
favor the forward reaction. The standard free energy change of
phosphocreatine is large and negative (ΔG'° = -43.0 kJ/mol).
23

24
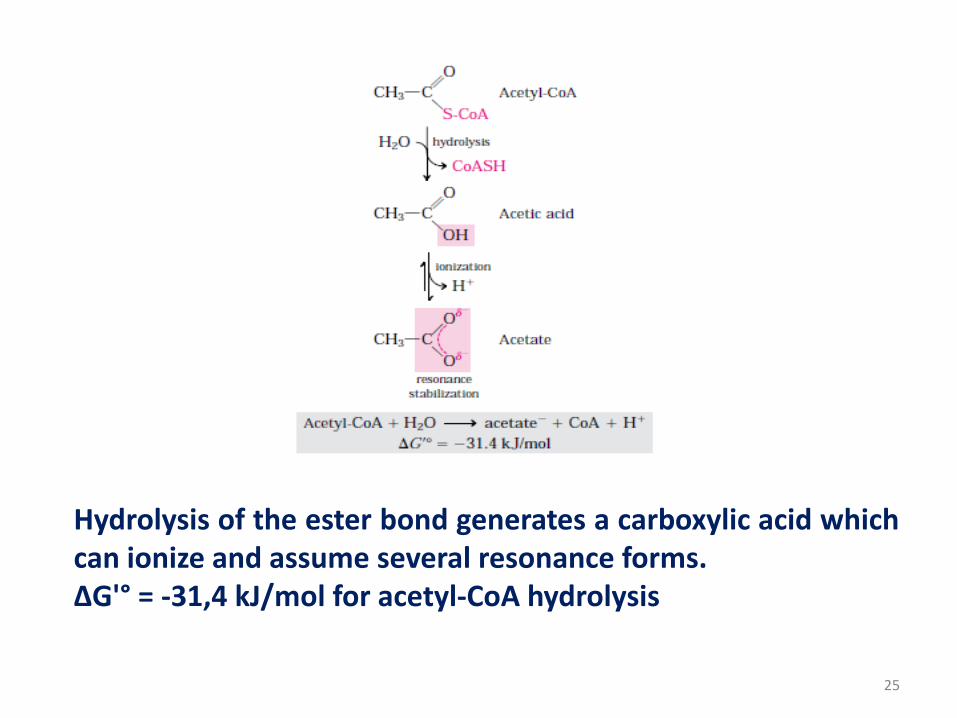
Thioesters (Acetyl COA)
• In thioesters a sulfur atom is replaced the usual oxygen in
the ester bond.
• Thioesters have large, negative standard free energy change
of hydrolysis.
• Acetyl coenzyme A is the one of important thioesters in
metabolism. The acyl group in these compounds is activated
for trans-acylation, condensation or oxidation-reduction
reactions.
25
Hydrolysis of the ester bond generates a carboxylic acid whichcan ionize and assume several resonance forms.ΔG'° = -31,4 kJ/mol for acetyl-CoA hydrolysis
26
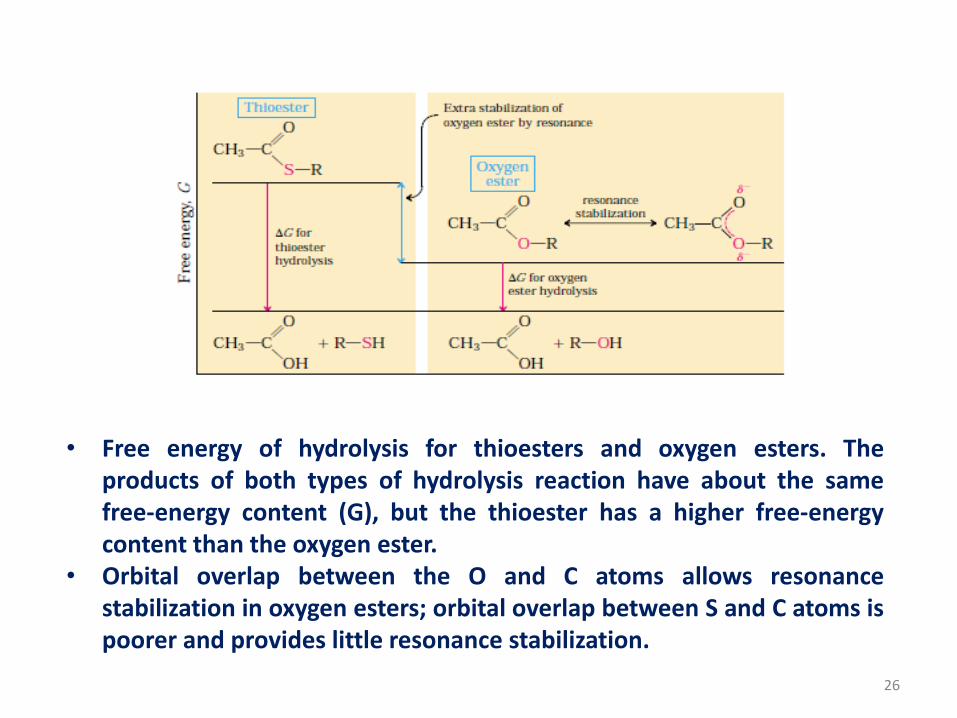
• Free energy of hydrolysis for thioesters and oxygen esters. Theproducts of both types of hydrolysis reaction have about the samefree-energy content (G), but the thioester has a higher free-energycontent than the oxygen ester.
• Orbital overlap between the O and C atoms allows resonancestabilization in oxygen esters; orbital overlap between S and C atoms ispoorer and provides little resonance stabilization.

• For hydrolysis reactions with large, negative standard free energy
changes, the products are more stable than the reactants for one or
more of the following reasons:
1. The bond strain in reactants due to electrostatic repulsion is relieved by
charge separation, as for ATP.
2. The products are stabilized by ionization, as for ATP, acyl phosphates,
thioesters.
3. The products are stabilized by isomerization (tautomerization) as for
phosphoenolpyruvate
4. The products are stabilized by resonance as for creatine released from
phosphocreatine, carboxylate ion released from acyl phosphates and
thioesters and phosphate released from anhydride or ester linkages.
27

• The phosphate compounds found in living organisms
divided into two groups based on their standard free
energy changes of hydrolysis.
‘High-energy’ compounds have a ΔG'° of hydrolysis more
negative than -25 kJ/mol (ATP, with a ΔG'° of hydrolysis of
-30 kJ/mol).
‘Low-energy’ compounds have a less negative ΔG'°
(glucose 6-phosphate with a (ΔG'° of hydrolysis of -13,8
kJ/mol).
28
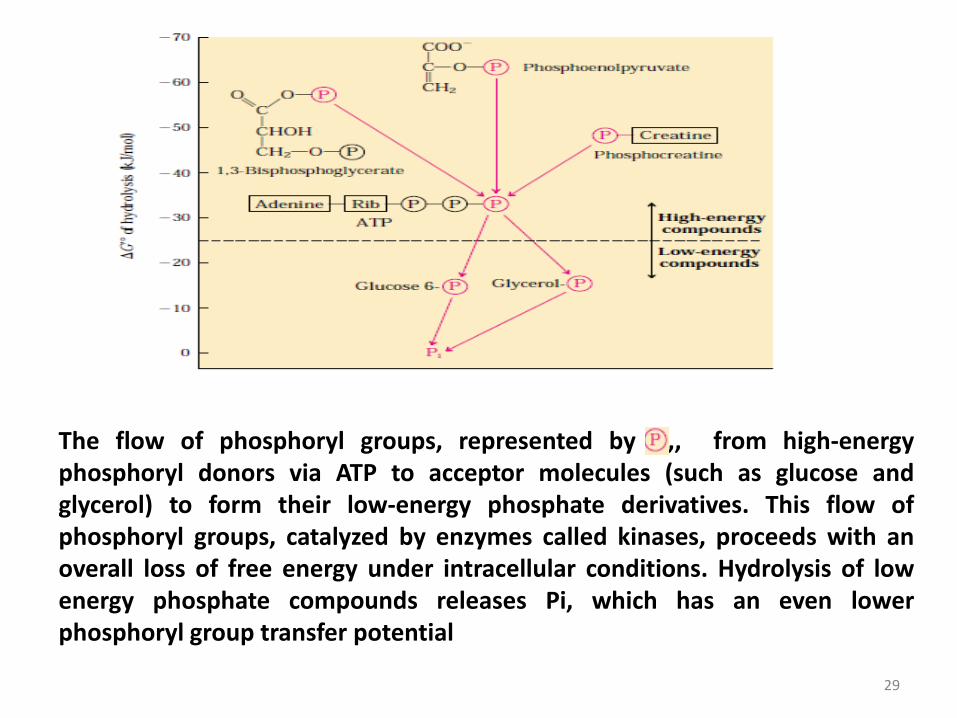
29
The flow of phosphoryl groups, represented by ,, from high-energyphosphoryl donors via ATP to acceptor molecules (such as glucose andglycerol) to form their low-energy phosphate derivatives. This flow ofphosphoryl groups, catalyzed by enzymes called kinases, proceeds with anoverall loss of free energy under intracellular conditions. Hydrolysis of lowenergy phosphate compounds releases Pi, which has an even lowerphosphoryl group transfer potential

• The breaking of all chemical bonds requires an input of
energy. The free energy released by hydrolysis of phosphate
compounds does not come from the specific bond that is
broken. It results from the products of the reaction having a
lower free energy content than the reactants.
• As is evident from the additivity of free energy changes of
sequential reactions, any phosphorylated compound can be
synthesized by coupling the synthesis to the breakdown of
another phosphorylated compound with a more negative
standard free energy change of hydrolysis.
30

PEP + H2O Pyruvate + Pi -61,9
ADP+ Pi ATP+ H2O +30,5
PEP + ADP Pyruvate + ATP -31,4
Cleavage of Pi from PEP releases more energy than is
needed to drive to condensation of Pi with ADP, the direct
donation of a phosphoryl group from PEP to ADP is
thermodynamically feasible.
31

• The overall reaction above is represented as the algebraic
sum of first two reactions, the overall reaction (third) does
not involve Pi ; PEP donates a phosphoryl group directly to
ADP.
• We can describe phosphorylated compounds as having a
high or low phosphoryl group transfer potential, on the basis
of their standard free energy changes of hydrolysis
32

• Much of catabolism is directed toward the synthesis of
high-energy phosphate compounds, but their formation
is not an end in itself; they are the means of activating a
wide variety of compounds for further chemical
transformation.
• The transfer of a phosphoryl group to a compound
effectively puts free energy into that compound, so that
it has more free energy to give up during subsequent
metabolic transformations.
33

• Because of its intermediate position on the scale of
group transfer potential, ATP can carry energy from
high-energy phosphate compounds produced by
catabolism to compounds such as glucose, converting
them into more reactive species.
• ATP serves as the universal energy currency in all living
cells
34

• One more chemical feature of ATP is crucial to its role in
metabolism: although in aqueous solution ATP is
thermo-dynamically unstable and is therefore a good
phosphoryl group donor, it is kinetically stable.
• Because of high activation energies required for
uncatalyzed reaction ATP does not spontaneously
donate phosphoryl groups to water or to the other
potential acceptors in the cell.
35

• ATP hydrolysis occurs only when specific enzymes which
lower the energy of activation are present
• The cell is therefore able to regulate the disposition of the
energy carried by ATP by regulating the various enzymes that
act on ATP
• Each of the three phosphates of ATP is susceptible to
nucleophilic attack and each position of attack yields a
different type of product
36
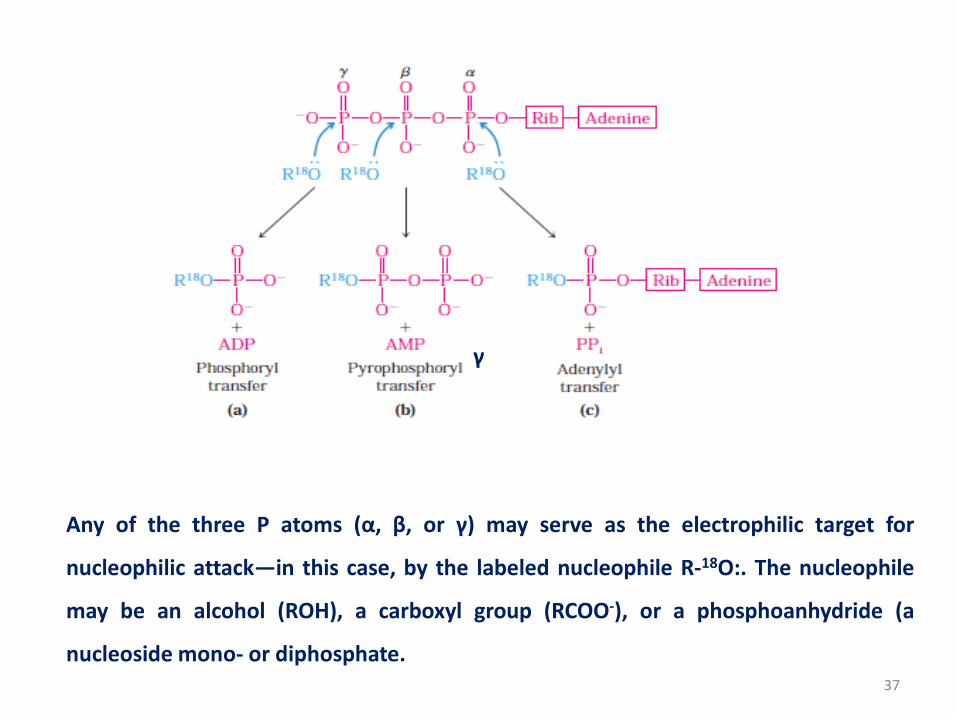
37
Any of the three P atoms (α, β, or γ) may serve as the electrophilic target for
nucleophilic attack—in this case, by the labeled nucleophile R-18O:. The nucleophile
may be an alcohol (ROH), a carboxyl group (RCOO-), or a phosphoanhydride (a
nucleoside mono- or diphosphate.

• For example (a) When the oxygen of the nucleophile attacks
the γ position, the bridge oxygen of the product is labeled,
indicating that the group transferred from ATP is a
phosphoryl (-PO32-), not a phosphate (-OPO3
2-).
(b) Attack on the β position displaces AMP and leads to the
transfer of a pyrophosphoryl (not pyrophosphate) group to
the nucleophile.
(c) Attack on the α position displaces PPi and transfers the
adenylyl group to the nucleophile.
38