adenoviral-mediated expression of human insulin-like growth factor-binding protein-3
TRANSCRIPT

AG
S*a
R
ap3rBattadBmt3ilSkpnahiIimctIct
p
8
Protein Expression and Purification 16, 202–211 (1999)Article ID prep.1999.1075, available online at http://www.idealibrary.com on
2
denoviral-Mediated Expression of Human Insulin-likerowth Factor-Binding Protein-3
ue M. Firth,*,1 Usha Ganeshprasad,* Philip Poronnik,† David I. Cook,† and Robert C. Baxter*Kolling Institute of Medical Research, Royal North Shore Hospital, St. Leonards, NSW 2065;nd †Department of Physiology, University of Sydney, Sydney, NSW 2006, Australia
eceived January 22, 1999, and in revised form March 10, 1999
iatsbflataIptaIbt(BiItiuppiraq
uow1fat
Insulin-like growth factors (IGFs) in the circulationre predominantly sequestered into ternary com-lexes comprising IGF, IGF-binding protein-3 (IGFBP-), and the acid-labile subunit (ALS). Besides its role inegulating IGF bioavailability in the circulation, IGF-P-3 has both IGF-dependent and IGF-independentctions on cell proliferation. As part of our studies intohe structure–function relationships of the multifunc-ional IGFBP-3, we have evaluated the efficiency of andenovirus-mediated expression system for rapid, me-ium-scale production of functional, glycosylated IGF-P-3. Replication-deficient adenovirus containing hu-an IGFBP-3 cDNA was generated using standard
echniques. Secreted, recombinant IGFBP-3 (IGFBP-Ad) was purified from the culture medium of virus-nfected cells by IGF-I affinity chromatography fol-owed by reverse-phase HPLC. When analyzed byDS–PAGE, IGFBP-3Ad was similar in size (43- to 45-Da glycoform doublet) to IGFBP-3Pl derived fromlasma. In addition, IGFBP-3Ad was detected by immu-oblot using an antibody specific for human IGFBP-3nd by ligand blot using radiolabeled IGF-I. IGFBP-3Ad
ad similar affinities for IGF-I and ALS and an approx-mately 25% decreased affinity for IGF-II compared toGFBP-3Pl. IGFBP-3Ad showed no significant differencen its susceptibility to an IGFBP-3 protease present in
edium conditioned by MCF-7 breast cancer cellsompared to IGFBP-3Pl, but appeared more resistanto the IGFBP-3 protease present in pregnancy serum.GFBP-3Ad also exhibited increased binding to T47Dells which may be related to the glycosylation state ofhe protein. © 1999 Academic Press
Insulin-like growth factors, IGF-I and IGF-II, areotent mitogenic peptides found in high concentrations
1 To whom correspondence should be addressed. Fax: 612-9926-
i484. E-mail: [email protected].02
n the circulation. The IGFs are sequestered into high-ffinity ternary complexes by insulin-like growth fac-or-binding protein-3 (IGFBP-3) and the acid-labileubunit (ALS); these complexes regulate both the sta-ility of the IGFs in the circulation and their egressrom the vasculature to target tissues (1). At the cellu-ar level, IGFBP-3 has been shown to modulate IGFctivity in a paradoxical fashion. It has been suggestedhat IGFBP-3 in the extracellular milieu inhibits IGFctivity by forming high-affinity binary complexes withGFs, while IGFBP-3 associated with the cell surfaceotentiates IGF action, possibly by concentrating IGFso their cognate receptors (2). In the past few years,ttention has also turned to the cellular actions ofGFBP-3 which may be independent of IGFs. It haseen shown that IGFBP-3 has growth inhibitory func-ions that are independent of the type I IGF receptor3), perhaps related to the recent discovery that IGF-P-3 can be translocated into the nuclei of cells where
t could exert a direct influence on gene expression (4).GFBP-3 function is thought to be modulated by post-ranslational events such as phosphorylation and lim-ted proteolysis (5). The diversity of functions attrib-ted to IGFBP-3 has made it a challenging prospect inrotein structure–function studies. There are few re-orts on the elucidation of structural determinantsnvolved in protein–protein or protein–cell interactionsequired for IGFBP-3 function (4, 6–8); these studiesre usually limited by the nonavailability of largeuantities of purified protein.Traditionally, heterologous protein expression is
sually achieved in bacterial cells by taking advantagef plasmid vectors, containing powerful promoters,hich are easy to manipulate in Escherichia coli (9,0). However, the limitations in using these systemsor eukaryotic protein expression include inappropri-te or lack of complex posttranslational modifications,he accumulation of insoluble aggregated products in
nclusion bodies as a result of inefficient secretion, and1046-5928/99 $30.00Copyright © 1999 by Academic Press
All rights of reproduction in any form reserved.

tspCHwd
ibetthediicirsprp
M
pmCdcAtw(
IptI
ncgwdtoaa
wt
t(MiI(mgsec
tsampatt1d1pioc
aHtAe0r(t0Heame
nspPBva
wh
203RECOMBINANT INSULIN-LIKE GROWTH FACTOR-BINDING PROTEIN-3
hus, the requirement for refolding which could lead totructure anomalies. Protein expression systems usinglasmid vectors transfected into mammalian cells likeHO or COS (11, 12) overcome these disadvantages.owever, recombinant protein yields are generally lowhich then increases costs associated with protein pro-uction.We report here the feasibility of expressing IGFBP-3
n an adenovirus-mediated expression system. It haseen shown previously that adenoviruses are highlyfficient at infecting cells, resulting in expression of theransgene in at least 95% of the target cells (13). Usinghis system, we have generated replication-deficientuman type 5 adenoviruses in which expression ofither glycosylated or nonglycosylated IGFBP-3 wasriven by the CMV promoter. Although glycosylationn IGFBP-3 has no apparent effect on the ligand-bind-ng function of IGFBP-3 (14), it is preferable to useorrectly modified proteins in structure–function stud-es. As large quantities of recombinant proteins areequired for studies on structure-function relation-hips of IGFBP-3, we have validated the functions ofurified recombinant IGFBP-3 expressed by adenovi-us (IGFBP-3Ad) and compared it to the natural proteinurified from plasma (IGFBP-3Pl).
ATERIALS AND METHODS
Materials. All radiolabeled proteins used were pre-ared as previously described (15, 16). Restriction andodifying enzymes were purchased from Promegaorp. (Madison, WI). Dulbecco’s modified Eagle’s me-ium (DMEM), RPMI medium 1640 (RPMI), and fetalalf serum were from Cytosystems (North Ryde, NSW,ustralia). All chromatography media with the excep-
ion of AffiGel-10 (Bio-Rad Laboratories, Hercules, CA)ere purchased from Amersham Pharmacia Biotech
Uppsala, Sweden).Purification of natural IGFBP-3Pl. Natural human
GFBP-3 was purified from Cohn fraction IV of humanlasma essentially as described previously (17), excepthat an IGF-I affinity column was used instead of IGF-I.
Adenoviral construction and purification of recombi-ant IGFBP-3. The human IGFBP-3 cDNA was sub-loned into the adenoviral shuttle vector, pAC-CMV (aift from Dr. C. Newgard, University of Texas-South-estern Medical Center, Dallas, TX). In order to pro-uce nonglycosylated IGFBP-3 (IGFBP-3NG), a mu-ated IGFBP-3 cDNA in which the Asn residue of eachf the three N-glycosylation sites was substituted withn Ala residue by site-directed mutagenesis (14) waslso subcloned into pAC-CMV.Replication-deficient human type 5 adenovirusesith IGFBP-3 inserted into the E1-deleted region were
hen produced by homologous recombination between r
he shuttle vector and the adenoviral plasmid pJM17a gift from Dr. B. C. O’Connell, NIDR, NIH, Bethesda,
D) in 911 human embryonic retina cells (18) follow-ng calcium phosphate transfection (Life Technologiesnc., Gaithersburg, MD) using standard techniques19). The 911 cells were cultured in DMEM supple-
ented with 10% (v/v) fetal calf serum and 23 mMlutamine at 37°C in 5% CO2/95% air. Viruses wereubsequently plaque purified and single plaques werexpanded. Stocks of cell lysates containing viral parti-les were stored at 280°C and thawed prior to use.For protein production, the medium of confluent cul-
ures of 911 cells in 450-cm2 flasks was changed toerum-free DMEM supplemented with 0.1% (w/v) BSAnd the cells were incubated for an additional 24 h. Theedium was then changed to serum-free DMEM sup-
lemented with 0.1% (w/v) BSA, 20 mM glutamine,nd a cocktail of protease inhibitors (500 units/ml apro-inin, 5 mg/ml a2-macroglobulin, and 0.5 mg/ml leupep-in) and 5 ml of virus supernatant (approximately 2 3013 viral particles). After 48 h, the conditioned me-ium was collected and clarified by centrifugation at5,300g for 20 min and 0.5 mg/ml Na2EDTA was addedrior to storage at 220°C. Total protein concentrationn the conditioned medium was measured using a col-rimetric assay based on the Bradford dye-binding pro-edure (Bio-Rad Laboratories).IGFBP-3Ad and IGFBP-3NG were purified by IGF-I
ffinity chromatography followed by reverse-phasePLC, essentially as described previously (17). Briefly,
he conditioned medium was loaded onto an IGF-I–ffigel 10 column at 0.4 ml/min. The IGFBP-3 wasluted from the column with 0.5 M acetic acid, pH 3, at.5 ml/min and active fractions, as determined by aadioimmunoassay (RIA) specific for human IGFBP-315), were pooled and freeze-dried. The sample washen reconstituted in 1 ml of 15% (v/v) acetonitrile in.1% trifluoroacetic acid and applied to a 250 3 4.6-mmiPore RP-318 column (Bio-Rad Laboratories). A lin-
ar gradient of 15–60% acetonitrile in 0.1% trifluoro-cetic acid was immediately applied over 30 min at 1.5l/min. A single peak of IGFBP-3 immunoreactivity
merged at 16 min corresponding to 39% acetonitrile.SDS–PAGE and silver staining. A quantity of 100
g of IGFBP-3Ad was reconstituted in 4 ml of Laemmliample buffer, heated at 95°C for 5 min, and electro-horesed on a Phastgel 10–15% gradient gel in thehastSystem separation unit (Amersham Pharmaciaiotech). The gel was stained in the PhastSystem de-elopment unit using the PhastGel silver staining kitccording to the manufacturer’s specifications.Immuno- and ligand blotting. Each protein (50 ng)as reconstituted in 50 ml of Laemmli sample buffer,eated at 95°C for 5 min, and fractionated under non-
educing conditions on a 10% SDS–polyacrylamide gel
of(ic31tc1wiip(was
aTmp
BttmAaTfScb1PC
c5IM1bs3p
mo
t51
bInpmI(oar
scb0wcimcwovtswiC
octwespeB
admsmpc0TImIcoc
204 FIRTH ET AL.
vernight at 100 V (20). The proteins were then trans-erred to Hybond-C Extra supported nitrocelluloseAmersham Pharmacia Biotech) by electroblotting us-ng a Multiphor II Novablot unit (Amersham Pharma-ia Biotech). After transfer, the blot was incubated at7°C for 3 h in Tris-buffered saline (TBS, 10 mM Tris,50 mM NaCl, pH 7.4) containing 1% (w/v) BSA andhen probed with anti-IGFBP-3 antiserum at a finaloncentration of 1:10,000 (prepared in TBS containing% (w/v) BSA and 0.05% (v/v) Nonidet P-40). The blotas washed three times in TBS, once in TBS contain-
ng 0.05% Nonidet P-40, and an additional three timesn TBS and then incubated for 2 h at 22°C in 125I-rotein A (1 3 106 cpm/50 ml of TBS containing 1%w/v) BSA and 0.05% (v/v) Nonidet P-40). Following theash regime used above, the dried blot was placedgainst Hyperfilm MP autoradiographic film (Amer-ham Pharmacia Biotech) for 1–3 days at 70°C.The samples for ligand blotting were electrophoresed
nd transferred to nitrocellulose as described above.he blot was incubated with 125I-IGF-I (1 3 106 cpm/50l) for 16 h at 22°°C. The blot was then washed and
rocessed for autoradiography as described above.Proteolysis of IGFBP-3. Each preparation of IGF-P-3 (100 ng) was incubated with 1 ml of human third-
erm pregnancy serum in a final volume of 50 ml pro-eolysis buffer (50 mM sodium phosphate (pH 6.8), 0.5M CaCl2, 0.5 mM MgCl2) at 37°C for 0, 2, 4, and 6 h.t each time point, 15 ml of Laemmli sample buffer wasdded to the samples which were then stored at 220°C.he samples were then heated at 95°C for 5 min and
ractionated under nonreducing conditions on a 12%DS–polyacrylamide gel overnight at 100 V and pro-essed as described above for immunoblotting. Thelots were placed against storage phosphor screens for6 h and the images were scanned and analyzed on ahosphorImager SP (Molecular Dynamics, Sunnyvale,A).Medium conditioned by MCF-7 human breast cancer
ells was equilibrated with 0.1 M sodium acetate (pH.5) as described previously (21). Each preparation ofGFBP-3 (50 ng) was incubated with 50 ml of theCF-7 conditioned medium at 37°C for 0, 4, 6, 8, and
6 h. At each time-point, 15 ml of Laemmli sampleuffer was added to the samples which were thentored at 220°C. The samples were then heated at7°C for 5 min just prior to SDS–PAGE and the gel wasrocessed for immunoblotting as described above.Binding assays. Binary and ternary complex for-ation were measured essentially as described previ-
usly (22). Briefly, reactions containing 125I-IGF-I or125I-IGF-II (10,000 cpm) and IGFBP-3 over the concen-ration range of 0–10 ng in a total volume of 0.3 ml of0 mM sodium phosphate buffer, pH 6.5, containing
% (w/v) BSA, were incubated at 22°C for 2 h. The winary complexes were immunoprecipitated with 0.5 mlGFBP-3 antibody and 2.5 ml of goat anti-rabbit immu-oglobulin in the presence of 4% final concentration ofolyethylene glycol. Ternary complex formation waseasured by incubating 125I-ALS (10,000 cpm) with
GF-I (50 ng) and varying concentrations of IGFBP-3over the range of 0–20 ng) in a total volume of 0.3 mlf 50 mM sodium phosphate containing 1% (w/v) BSAt 22°C for 2 h. Ternary complexes were then sepa-ated from unbound tracer as described above.The affinity of IGF binding to IGFBP-3 was mea-
ured essentially as described above except that theoncentration of IGFBP-3 was held constant. Unla-eled IGF was added over the concentration range of.0025–1 ng in a total volume of 0.3 ml. Complexesere separated from unbound tracer by immunopre-
ipitation as described above. The affinity of ALS bind-ng to the IGFBP-3–IGF-I binary complex was deter-
ined as described previously (16) except that theoncentrations of IGFBP-3 (1 ng) and IGF-I (50 ng)ere held constant while unlabeled ALS was addedver the concentration range of 2.5–200 ng in a totalolume of 0.3 ml. Bound tracer was separated from freeracer, as described above. Nonspecific binding wasubtracted from each data point prior to curve fittingith the Marquardt algorithm for one-site ligand bind-
ng using the software package Ultrafit 3.0.5 (Biosoft,ambridge, UK).Heparin binding. Four to 5 mg of each preparation
f IGFBP-3 was applied to 1 ml heparin–Sepharoseolumns (HiTrap Heparin, Amersham Pharmacia Bio-ech) at 0.4 ml/min at 4°C. Following extensive washesith 50 mM sodium phosphate (pH 6.5), IGFBP-3 wasluted by applying a step gradient from 0.4 to 1.0 Modium chloride (made in 10 mM sodium phosphate,H 6.5). Five fractions of 2 ml each were collected atach step of the elution gradient and assayed for IGF-P-3 by RIA.Detection of cell-associated IGFBP-3. Cell-surface
ssociation of IGFBP-3 was measured by a previouslyescribed immunological assay (23). Briefly, T47D hu-an breast cancer cells were plated in RPMI medium
upplemented with 10% (v/v) fetal calf serum, 2.92g/ml glutamine, and 10 mg/ml insulin at 2 3 104 cells
er well in 24-well plates for 48 h. Cultures werehanged to serum-free medium supplemented with.1% (w/v) BSA and incubated for an additional 24 h.he cells were then treated with each preparation ofGFBP-3 in 0.5 ml of medium for 4 h at 22°C. The cellonolayers were washed and incubated with either
GFBP-3 antibody (R-100) or normal rabbit serum (asontrol for nonspecific effects) diluted 1:5000 in 0.5 mlf medium. Following a 16-h incubation at 22°C, theell monolayers were washed again before incubation
125
ith I-protein A (40,000 cpm/ml medium) for 2 h.
Uwwgm((
rBute
R
BBupaAwt2cI0Wias
(phiNc
tmgnstmItnbIep
t
I
I
w
up
FTtplIgbRc
205RECOMBINANT INSULIN-LIKE GROWTH FACTOR-BINDING PROTEIN-3
nbound tracer was removed by washing and the cellsere solubilized with 0.5% (w/v) SDS. The cell lysatesere collected and radioactivity was determined in aamma counter. Total DNA in the cell lysates waseasured by fluorometry using Hoechst 33258 dye
Calbiochem, La Jolla, CA), as described previously24).
Statistical analysis. Statistical analysis was car-ied out using StatView 4.02 (Abacus Concepts Inc.,erkeley, CA). Differences between groups were eval-ated by Fisher’s protected least significant differenceest after analysis of variance, and a significant differ-nce was defined as P , 0.05.
ESULTS
Expression and purification of recombinant IGF-P-3. The expression of recombinant IGFBP-3 (IGF-P-3Ad) by the adenovirus-mediated system is outlinednder Materials and Methods. Table 1 summarizes theurification procedures for recombinant IGFBP-3Ad
nd natural IGFBP-3 from human plasma (IGFBP-3Pl).lthough the level of adenovirus-mediated expressionas modest (0.3 mg/ml) in this study, later prepara-
ions were consistently obtained at approximately 1.5–.0 mg/ml for medium collected 24 h postinfection. Inomparison, the best level of expression achieved byGFBP-3-transfected Chinese hamster ovary cells was.1 mg/ml for medium conditioned for 72–96 h (8).hen analyzed by SDS–PAGE followed by silver stain-
ng, the preparation of purified IGFBP-3Ad appeared asdoublet of approximately 43 and 45 kDa (Fig. 1a),
imilar to that shown previously for IGFBP-3Pl (17).Immunoblotting of the two preparations of IGFBP-3
IGFBP-3Ad and IGFBP-3Pl) resulted in a pattern of tworoteins of approximately 43 and 45 kDa (Fig. 1b). Weave previously shown that the characteristic doublet
s the result of variable glycosylation at one of the three-glycosylation sites in IGFBP-3 (14). The larger gly-
Pl
TAB
Purification
IGFBP-3Volume
(ml)Total protein
(mg)Total IGFBP-
(mg)
GFBP-3Pl 1760 82,250 44,400a
GFBP-3Ad 650 780c 200
a These values were taken from (17); the original IGFBP-3 RIA wasith the values obtained in the current RIA for IGFBP-3Ad.b This is an intermediate step for removing bound IGFs prior to Ic The recombinant IGFBP-3 was collected in cell culture medium s
oform of IGFBP-3 was relatively more abundant g
han the smaller glycoform. In contrast, there wasore of the 43-kDa glycoform relative to the 45-kDa
lycoform in IGFBP-3Ad; this pattern is consistent withatural IGFBP-3 that is secreted by human cell linesuch as fibroblasts and prostate, breast, and endome-rial cancer cells (25–28). An IGFBP-3 proteolytic frag-ent, of approximately 30 kDa, is present in both
GFBP-3Pl and IGFBP-3Ad (Fig. 1b). This immunoreac-ive fragment is frequently seen in preparations ofatural IGFBP-3 derived from plasma (17) and recom-inant IGFBP-3 (8), following prolonged storage. TheGF-binding function of the IGFBP-3 preparations wasxamined by ligand blotting using 125I-IGF-I as therobe (Fig. 1c). Both preparations were able to bind
125I-IGF-I and consistent with previous studies, onlyhe intact protein doublet has IGF-I binding ability.
1
f IGFBP-3
Purification stepsFinal yield of IGFBP-3
(% of total amount)
SP–Sephadex Extractionb 18,000 mg (40%)a
IGF-I affinity chromatography 3,000 mg (7%)a
Reverse-phase HPLC 2,000 mg (5%)a
IGF-I affinity chromatography 142 mg (71%)Reverse-phase HPLC 75 mg (38%)
orrectly calibrated and these values should be halved for comparison
affinity chromatography.plemented with 1 mg/ml BSA which is the bulk of the total protein.
IG. 1. 10–15% gradient SDS–PAGE of purified IGFBP-3Ad (a).he size of the molecular mass standards (lane 1) are indicated onhe left and 100 ng of purified IGFBP-3Ad was loaded in lane 2. Therotein bands were detected by silver staining. Immuno- (b) andigand (c) blots of IGFBP-3. 100 ng each of IGFBP-3Pl (lane 1) andGFBP-3Ad (lane 2) was electrophoresed on 10% SDS–polyacrylamideel, electroblotted onto nitrocellulose, and probed with either anti-ody against IGFBP-3 followed by 125I-protein A (b) or 125I-IGF-I (c).elative migration distances of molecular mass standards are indi-ated on the left of each panel. The width 3 length dimensions of the
LE
o
3
inc
GF
els are 43 3 50 mm (a) and 16 3 16 cm (b and c).

TI
IlwIpcitb(AAc(
pftfIIempcta(faa
Ft1
2ad
FoIfoI
206 FIRTH ET AL.
he 30-kDa proteolyzed form of IGFBP-3 has no IGF--binding function when analyzed in this way.
Ligand-binding functions of recombinant IGFBP-3.n order to examine the dose–response relationship forigand binding by IGFBP-3, solution binding assaysere performed in which fixed concentrations of 125I-
GF-I (Fig. 2a), 125I-IGF-II (Fig. 2b), or 125I-ALS in theresence of IGF-I (Fig. 2c) were incubated with in-reasing concentrations of each IGFBP-3. Essentiallydentical curves for IGF-II binding were obtained withhe two preparations of IGFBP-3 (Fig. 2b), whereas theinding of IGF-I by IGFBP-3Ad was significantly lowerP 5 0.008) compared to that by IGFBP-3Pl (Fig. 2a).lthough IGFBP-3Ad showed apparent lower binding toLS compared to IGFBP-3Pl (Fig. 2c), the binding
urves were not statistically different from each otherP 5 0.13).
IG. 2. Relationship between increasing concentration of IGFBP-3he presence of 50 ng of IGF-I (c). Increasing amounts of IGFBP-3Pl
25I-IGF-II (b), or 125I-ALS in the presence of 50 ng of unlabeled IGF-2°C for 2 h. Bound tracer was separated from unbound tracer by imnti-rabbit immunoglobulin in the presence of a 4% final concentrauplicate measurements in three independent experiments.
IG. 3. Nonlinear saturation curves of IGFBP-3 binding to increasif 50 ng of IGF-I (c). Increasing amounts of 125I-IGF-I (a), 125I-IGF-II (GFBP-3Pl (h) and IGFBP-3Ad (‚) in a final volume of 0.3 ml of 50 mMrom unbound tracer by immunoprecipitation with 0.5 ml of IGFBP-3f a 4% final concentration of polyethylene glycol. The binding curve
GFBP-3.In order to obtain the relative affinities of the tworeparations of IGFBP-3 for IGF-I or IGF-II, we per-ormed saturation kinetics analyses which measuredhe displacement of either 125I-IGF-I or 125I-IGF-II,rom binary complexes formed in the presence of eachGFBP-3, by increasing concentrations of unlabeledGF-I or IGF-II, respectively. Similarly, the affinity ofach IGFBP-3 for ALS was determined by the displace-ent of 125I-ALS, from ternary complexes formed in the
resence of each IGFBP-3 and 50 ng of IGF-I, by in-reasing concentrations of unlabeled ALS. Representa-ive saturation binding curves for the two preparationsre shown for IGF-I (Fig. 3a), IGF-II (Fig. 3b), and ALSFig. 3c) binding. The association constants derivedrom these data are summarized in Table 2. Both prep-rations of IGFBP-3 have similar affinities for IGF-Ind ALS. However, IGFBP-3Ad has a statistically sig-
the specific binding of 125I-IGF-I (a), 125I-IGF-II (b), and 125I-ALS inand IGFBP-3Ad (‚) are incubated with 10,000 cpm of 125I-IGF-I (a),
) in a final volume of 0.3 ml of 50 mM sodium phosphate (pH 6.5) atnoprecipitation with 0.5 ml of IGFBP-3 antibody and 2.5 ml of goat
n of polyethylene glycol. Data points shown are means 6 SEM of
amounts of 125I-IGF-I (a), 125I-IGF-II (b), and 125I-ALS in the presenceand 125I-ALS in the presence of 50 ng of IGF-I (c) are incubated withium phosphate (pH 6.5) at 22°C for 2 h. Bound tracer was separatedibody and 2.5 ml of goat anti-rabbit immunoglobulin in the presenceown are representatives of three independent experiments for each
and(h)I (cmutio
ngb),sodant
s sh

n0
tltrt4spBeseMtp(Btnln
shispsocplto
ttfMrcinead(1pdwpta
BoachsIpnls
Fmhppst
II
ecIIad
207RECOMBINANT INSULIN-LIKE GROWTH FACTOR-BINDING PROTEIN-3
ificant decrease (25%) in its affinity for IGF-II (P 5.03) compared to that of IGFBP-3Pl.Heparin appears to bind to IGFBP-3 at a site distal
o the IGF-binding site since heparin has relativelyittle effect on IGF binding (29). As a separate test forhe structural and functional integrity of the adenovi-al-expressed protein, the binding and elution profile ofhe two IGFBP-3 to heparin was determined. Between
and 5 mg of each purified protein was loaded ontoeparate heparin–Sepharose columns and similar pro-ortions of each bound to the column (87.0% for IGF-P-3Pl and 83.8% for IGFBP-3Ad). The sodium chloridelution profile of each was similar in that both proteinstarted eluting with 0.5 M sodium chloride and thelutions were completed by the second fraction of 0.75
sodium chloride. The recovery of each preparation inhis high-salt peak was between 23 and 26% of boundrotein (Fig. 4). There was a first major elution peak26% of bound protein) with 0.4 M NaCl in the IGF-P-3Pl profile, which was subsequently identified as
he 30-kDa proteolyzed IGFBP-3 fragment by immu-oblotting (data not shown). Interestingly, the proteo-
yzed fragment from the adenoviral preparation wasot recovered in corresponding fractions.Susceptibility of recombinant IGFBP-3 to IGFBP-3-
pecific proteases. Specific proteases for IGFBP-3ave been identified, although not fully characterized,
n culture media conditioned by certain cell lines and inerum. Carbohydrate moieties on proteins have theotential to influence the stability of glycoproteins andince IGFBP-3Ad appears to have different proportionsf the glycoforms compared to IGFBP-3Pl (Fig. 1b), aomparison of their susceptibility to specific IGFBP-3roteases was carried out. Pregnancy serum was uti-ized as the source of a cation-dependent serine pro-ease of IGFBP-3 which has been previously studied in
TABLE 2
Binding Parameters for Binary and Ternary ComplexFormation among IGF-I, IGF-II, and ALS
and Each IGFBP-3
IGFBP-3
Association constant (Ka), liters/nmol
IGF-I IGF-II ALS
GFBP-3Pl 13.2 6 1.6 47.8 6 3.3 6.9 6 0.4GFBP-3Ad 14.2 6 1.4 36.3 6 1.0* 5.4 6 0.7
Note. Binary complex formation was determined by the binding ofither 125I-IGF-I or 125I-IGF-II to each IGFBP-3 whereas ternaryomplex formation was determined by the binding of 125I-ALS to eachGFBP-3 in the presence of 50 ng of IGF-I. The molecular weights ofGF-I, IGF-II, IGFBP-3, and ALS were taken as 7650, 7470, 43,000,nd 85,000, respectively. Values are means 6 SE for three indepen-ent measurements. *P 5 0.03.
ur laboratory and elsewhere. Each IGFBP-3 prepara- B
ion was incubated with pregnancy serum over a 6-hime course, separated on SDS–PAGE and processedor immunoblotting as described under Materials and
ethods. Digestion of IGFBP-3 with pregnancy serumesulted in the appearance of a 30-kDa fragment con-urrent with the disappearance of the 43- to 45-kDantact protein in both preparations of IGFBP-3 (dataot shown). The density of the intact protein signal atach time point was quantified by the PhosphorIm-ger, adding the densities of the doublet bands, and theata obtained were used to generate proteolysis plotsFig. 5a). Proteolysis of 50% of IGFBP-3Pl occurred by.5–2 h whereas 3.5 h of incubation was required toroteolyze 50% of IGFBP-3Ad. In contrast, when me-ium conditioned by MCF-7 human breast cancer cellsas used as the source of a cathepsin-like cysteinerotease for IGFBP-3 (21), there was no difference inhe susceptibility of each IGFBP-3 to this protease over16 h time course (Fig. 5b).Cell-binding function of recombinant IGFBP-3. IGF-P-3 has been shown to associate with the cell surfacesf fibroblasts (23) and T47D breast cancer cells (4). Wessessed the ability of each IGFBP-3 to bind to T47Dells and found that IGFBP-3Ad showed 1.5- to 2-foldigher cell binding than IGFBP-3Pl (Fig. 6). Since theource and relative proportion of the glycoforms inGFBP-3Ad is different from IGFBP-3Pl, we also com-ared the cell-binding function of IGFBP-3Ad with aonglycosylated form of IGFBP-3 (IGFBP-3NG) simi-
arly produced by an adenovirus-mediated expressionystem. When applied to monolayers of T47D breast
IG. 4. Elution profiles of IGFBP-3 from heparin–Sepharose chro-atography. IGFBP-3Pl (h) and IGFBP-3Ad (‚) were applied to 1 mleparin–Sepharose columns preequilibrated in 50 mM sodium phos-hate (pH 6.5) buffer. The column was washed with 50 mM sodiumhosphate (pH 6.5) buffer and adherent proteins were eluted in atepwise fashion with the same buffer containing increasing concen-rations of sodium chloride (E). The fractions were assayed for IGF-
P-3 in the RIA.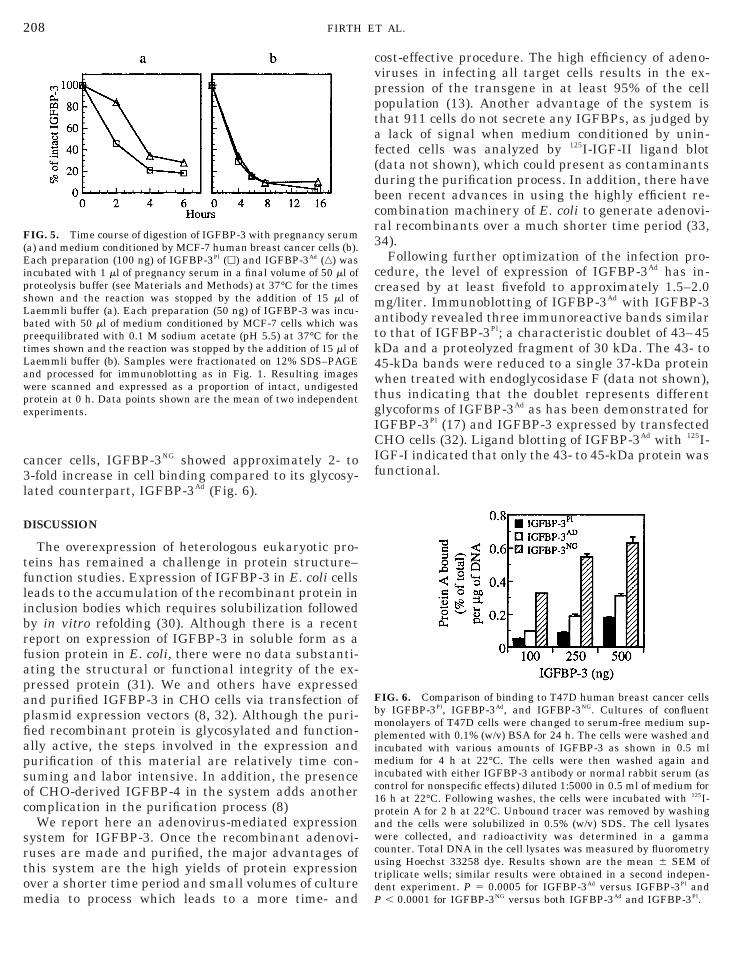
c3l
D
tflibrfapapfiapsoc
srtom
cvpptaf(dbcr3
ccmatk4wtgICIf
Fbmpimic1pawcutd
F(EipsLbptLawpe
208 FIRTH ET AL.
ancer cells, IGFBP-3NG showed approximately 2- to-fold increase in cell binding compared to its glycosy-ated counterpart, IGFBP-3Ad (Fig. 6).
ISCUSSION
The overexpression of heterologous eukaryotic pro-eins has remained a challenge in protein structure–unction studies. Expression of IGFBP-3 in E. coli cellseads to the accumulation of the recombinant protein innclusion bodies which requires solubilization followedy in vitro refolding (30). Although there is a recenteport on expression of IGFBP-3 in soluble form as ausion protein in E. coli, there were no data substanti-ting the structural or functional integrity of the ex-ressed protein (31). We and others have expressednd purified IGFBP-3 in CHO cells via transfection oflasmid expression vectors (8, 32). Although the puri-ed recombinant protein is glycosylated and function-lly active, the steps involved in the expression andurification of this material are relatively time con-uming and labor intensive. In addition, the presencef CHO-derived IGFBP-4 in the system adds anotheromplication in the purification process (8)
We report here an adenovirus-mediated expressionystem for IGFBP-3. Once the recombinant adenovi-uses are made and purified, the major advantages ofhis system are the high yields of protein expressionver a shorter time period and small volumes of culture
IG. 5. Time course of digestion of IGFBP-3 with pregnancy seruma) and medium conditioned by MCF-7 human breast cancer cells (b).ach preparation (100 ng) of IGFBP-3Pl (h) and IGFBP-3Ad (‚) was
ncubated with 1 ml of pregnancy serum in a final volume of 50 ml ofroteolysis buffer (see Materials and Methods) at 37°C for the timeshown and the reaction was stopped by the addition of 15 ml ofaemmli buffer (a). Each preparation (50 ng) of IGFBP-3 was incu-ated with 50 ml of medium conditioned by MCF-7 cells which wasreequilibrated with 0.1 M sodium acetate (pH 5.5) at 37°C for theimes shown and the reaction was stopped by the addition of 15 ml ofaemmli buffer (b). Samples were fractionated on 12% SDS–PAGEnd processed for immunoblotting as in Fig. 1. Resulting imagesere scanned and expressed as a proportion of intact, undigestedrotein at 0 h. Data points shown are the mean of two independentxperiments.
edia to process which leads to a more time- and P
ost-effective procedure. The high efficiency of adeno-iruses in infecting all target cells results in the ex-ression of the transgene in at least 95% of the cellopulation (13). Another advantage of the system ishat 911 cells do not secrete any IGFBPs, as judged by
lack of signal when medium conditioned by unin-ected cells was analyzed by 125I-IGF-II ligand blotdata not shown), which could present as contaminantsuring the purification process. In addition, there haveeen recent advances in using the highly efficient re-ombination machinery of E. coli to generate adenovi-al recombinants over a much shorter time period (33,4).Following further optimization of the infection pro-
edure, the level of expression of IGFBP-3Ad has in-reased by at least fivefold to approximately 1.5–2.0g/liter. Immunoblotting of IGFBP-3Ad with IGFBP-3
ntibody revealed three immunoreactive bands similaro that of IGFBP-3Pl; a characteristic doublet of 43–45Da and a proteolyzed fragment of 30 kDa. The 43- to5-kDa bands were reduced to a single 37-kDa proteinhen treated with endoglycosidase F (data not shown),
hus indicating that the doublet represents differentlycoforms of IGFBP-3Ad as has been demonstrated forGFBP-3Pl (17) and IGFBP-3 expressed by transfectedHO cells (32). Ligand blotting of IGFBP-3Ad with 125I-
GF-I indicated that only the 43- to 45-kDa protein wasunctional.
IG. 6. Comparison of binding to T47D human breast cancer cellsy IGFBP-3Pl, IGFBP-3Ad, and IGFBP-3NG. Cultures of confluentonolayers of T47D cells were changed to serum-free medium sup-
lemented with 0.1% (w/v) BSA for 24 h. The cells were washed andncubated with various amounts of IGFBP-3 as shown in 0.5 ml
edium for 4 h at 22°C. The cells were then washed again andncubated with either IGFBP-3 antibody or normal rabbit serum (asontrol for nonspecific effects) diluted 1:5000 in 0.5 ml of medium for6 h at 22°C. Following washes, the cells were incubated with 125I-rotein A for 2 h at 22°C. Unbound tracer was removed by washingnd the cells were solubilized in 0.5% (w/v) SDS. The cell lysatesere collected, and radioactivity was determined in a gamma
ounter. Total DNA in the cell lysates was measured by fluorometrysing Hoechst 33258 dye. Results shown are the mean 6 SEM ofriplicate wells; similar results were obtained in a second indepen-ent experiment. P 5 0.0005 for IGFBP-3Ad versus IGFBP-3Pl and
NG Ad Pl
, 0.0001 for IGFBP-3 versus both IGFBP-3 and IGFBP-3 .
ftA3wcbmcddccmoAhiTa
pltSfecbatfptaItiwcpcsar
iv(hIoaiM
lfiwpspn
aBiBoBmmsrhsstmBocraiicmefI
eeectwrtettascInfd
209RECOMBINANT INSULIN-LIKE GROWTH FACTOR-BINDING PROTEIN-3
Solution-based ligand-binding assays were per-ormed to compare the relative binding affinities of thewo IGFBP-3 preparations. The affinities for IGF-I andLS were not significantly different between IGFBP-Ad and IGFBP-3Pl. In contrast, the affinity for IGF-IIas decreased by approximately 25% in IGFBP-3Ad
ompared to IGFBP-3Pl. This difference in affinity maye due to the different sources of the proteins whichay result in differences in posttranslational modifi-
ations. The relative proportions of the 43- to 45-kDaoublet in the two preparations presumably reflect theifference in glycosylation processes by the sourceells. Although there are no data available about theoncentration and composition of the carbohydrateoieties in each IGFBP-3, glycosylation has been dem-
nstrated to have no significant effects on the IGF- andLS-binding functions of IGFBP-3 (14). On the otherand, in vitro phosphorylation of IGFBP-3 decreases
ts affinity for ALS binding as well as cell binding (5).he phosphorylation state of each of the IGFBP-3 prep-rations is yet to be established.It is interesting to note that although the 30-kDa
roteolyzed fragment of IGFBP-3Pl was eluted at aower molarity of sodium chloride (0.4 M) compared tohe intact protein (0.5–0.75 M) from the heparin–epharose column, the apparently similar proteolyzed
ragment in IGFBP-3Ad was not recovered in any of thelutions presumably because it had not bound to theolumn. Regardless of the differences in the heparininding ability of the proteolyzed fragments, it wouldppear that heparin affinity chromatography is a po-ential method for separating the 30-kDa proteolyzedragment from intact IGFBP-3. The two consensus he-arin-binding sites in IGFBP-3 are located in the C-erminal region and in the central region of the aminocid sequence. Sequencing of the 30-kDa fragment ofGFBP-3Ad revealed that it contains the same aminoerminal sequence as the intact protein, thus indicat-ng that it is probably truncated C-terminally. Thisould suggest that the C-terminal region of IGFBP-3
ontains the higher affinity heparin-binding site. Arevious study has shown that a synthetic peptideontaining the C-terminal consensus heparin-bindingite of IGFBP-3 bound heparin with fourfold higherffinity than a synthetic peptide containing the hepa-in-binding site of the central region (35).
Different classes of IGFBP-3 proteases are presentn vivo in serum and other body fluids as well as initro in culture media conditioned by various cell types36). A cation-dependent serine protease of IGFBP-3as been identified in pregnancy serum (37) and anGF-I-inhibitable, cation-dependent cysteine proteasef IGFBP-3 secreted by MCF-7 breast cancer cells haslso been characterized (21). Although the susceptibil-ty of the two IGFBP-3 toward the protease secreted by
CF-7 cells was identical, it appears that the proteo- f
ysis kinetics of the pregnancy serum protease are dif-erent for IGFBP-3Ad. There appears to be an initial lagn the proteolysis of IGFBP-3Ad compared to IGFBP-3Pl
hich is overcome by 4 h. This initial resistance toroteolysis is not due to glycosylation differences sinceimilar studies with IGFBP-3NG resulted in identicalroteolysis susceptibility curves as IGFBP-3Ad (dataot shown).The most interesting finding in this study is the
pparent increased binding to T47D cells by IGF-P-3Ad. Compared to IGFBP-3Pl, there is a 1.5- to 2-fold
ncrease in binding by IGFBP-3Ad at each of the IGF-P-3 concentrations used. The binding to cells is an-ther 2-fold higher by IGFBP-3NG compared to IGF-P-3Ad which would suggest that the carbohydrateoieties on the protein, which constitute 30% of itsolecular mass, may partially obscure the cell-binding
ite on IGFBP-3. We have previously shown that theatio of cell-associated IGFBP-3 to secreted IGFBP-3 isigher in transfected CHO cells expressing nonglyco-ylated IGFBP-3 compared to those expressing glyco-ylated IGFBP-3 (38). The immunological method usedo detect cell-associated IGFBP-3 is unable to deter-ine if the higher cell binding by IGFBP-3Ad and IGF-P-3NG in this study reflects increased binding affinityr increase in the number of molecules bound. Theell-binding site of IGFBP-3 has been localized to aegion in the C-terminus which is rich in basic aminocids (8) and we have also implicated the same regionn the translocation of IGFBP-3 to cell nuclei (4) wheret may exert influences on gene expression. It is con-eivable that the extent of glycosylation on IGFBP-3ay influence the distribution of IGFBP-3 between the
xtracellular milieu and cell surfaces and will there-ore have direct consequences on the cellular actions ofGFBP-3.
The IGFBP-3 recombinant type 5 adenoviruses gen-rated in this study are infectious but nonreplicativexcept in permissive cell lines which express the ad-noviral E1 gene in trans. Although this study hasoncentrated solely on the overexpression of IGFBP-3,he availability of IGFBP-3-expressing adenovirusesill enable the infection of nonpermissive cell lines
esulting in a highly efficient transient expression sys-em. Cell function can be altered by the expression ofxogenous genes introduced by the infection of replica-ion-deficient, recombinant adenoviruses, for example,he expression of aquaporin-5 in epithelial cells (39)nd the expression of a dominant negative G proteinubunit in salivary gland cells (13). Such a systemould be exploited for studies into the cellular actions ofGFBP-3. In summary, IGFBP-3 expressed by an ade-ovirus-mediated system essentially performs all theunctions of natural IGFBP-3. The relative ease of pro-ucing and purifying recombinant protein will enable
urther studies to elucidate the ligand-binding deter-
ms
A
RCddt
R
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
210 FIRTH ET AL.
inants, the extra- and intracellular actions, and, pos-ibly, the structure of IGFBP-3.
CKNOWLEDGMENTS
This work was supported by Project Grant 940447 (awarded to.C.B. and S.M.F.) from the National Health and Medical Researchouncil of Australia. P.P. is an Australian Research Council Post-octoral Fellow. D.I.C. and R.C.B. are Fellows of the Medical Foun-ation of the University of Sydney, Australia. We thank S. M. Pat-ison for help with SDS–PAGE.
EFERENCES
1. Rajaram, S., Baylink, D. J., and Mohan, S. (1997) Insulin-likegrowth factor-binding proteins in serum and other biologicalfluids: Regulation and functions. Endocrine Rev. 18, 801–831.
2. Jones, J. I., and Clemmons, D. R. (1995) Insulin-like growthfactors and their binding proteins: Biological actions. EndocrineRev. 16, 3–34.
3. Valentinis, B., Bhala, A., DeAngelis, T., Baserga, R., and CohenP. (1995) The human insulin-like growth factor (IGF) bindingprotein-3 inhibits the growth of fibroblasts with a targeted dis-ruption of the IGF-I receptor gene. Mol. Endocrinol. 9, 361–367.
4. Schedlich, L. J., Young, T. F., Firth, S. M., and Baxter, R. C.(1998) Insulin-like growth factor-binding protein (IGFBP)-3 andIGFBP-5 share a common nuclear transport pathway in T47Dhuman breast carcinoma cells. J. Biol. Chem. 273, 18347–18352.
5. Baxter, R. C. (1997) Molecular aspects of insulin-like growthfactor binding proteins. Adv. Mol. Cell. Endocrinol. 1, 123–159.
6. Vorwerk, P., Yamanaka, Y., Spagnoli, A., Oh, Y. M., and Rosen-feld, R. G. (1998) Insulin and IGF binding by IGFBP-3 fragmentsderived from proteolysis, baculovirus expression and normal hu-man urine. J. Clin. Endocrinol. Metab. 83, 1392–1395.
7. Hashimoto, R., Ono, M., Fujiwara, H., Higashihashi, N., Yo-shida, M., Enjoh-Kimura, T., and Sakano, K. (1997) Binding sitesand binding properties of binary and ternary complexes of insu-lin-like growth factor-II (IGF-II), IGF-binding protein-3, andacid-labile subunit. J. Biol. Chem. 272, 27936–27942.
8. Firth, S. M., Ganeshprasad, U., and Baxter, R. C. (1998) Struc-tural determinants of ligand and cell surface binding of insulin-like growth factor binding protein-3. J. Biol. Chem. 273, 2631–2638.
9. Olins, P. O., and Lee, S. C. (1993) Recent advances in heterolo-gous gene expression in Escherichia coli. Curr. Opin. Biotechnol.4, 520–525.
0. Gold, L. (1990) Expression of heterologous proteins in Esche-richia coli. Methods Enzymol. 185, 11–14.
1. Levinson, A. D. (1990) Expression of heterologous genes in mam-malian cells. Methods Enzymol. 185, 485–487.
2. Goeddel, D. V. (1990) Systems for heterologous gene expression.Methods Enzymol. 185, 3–7.
3. Poronnik, P., O’Mullane, L. M., Harding, E. A., Greger, R., andCook, D. I. (1998) Use of replication deficient adenoviruses toinvestigate the role of G proteins in Ca21 signalling in epithelialcells. Cell Calcium 24, 97–103.
4. Firth, S. M., and Baxter, R. C. (1995) The role of glycosylation inthe action of IGFBP-3. Prog. Growth Factor Res. 6, 223–229.
5. Baxter, R. C., and Martin, J. L. (1986) Radioimmunoassay ofgrowth hormone-dependent insulin-like growth factor bindingprotein in human plasma. J. Clin. Invest. 78, 1504–1512.
6. Baxter, R. C., Bayne, M. L., and Cascieri, M. A. (1992) Structural
determinants for binary and ternary complex formation betweeninsulin-like growth factor-I (IGF-I) and IGF binding protein-3.J. Biol. Chem. 267, 60–65.
7. Martin, J. L., and Baxter, R. C. (1986) Insulin-like growth factorbinding protein from human plasma: Purification and character-ization. J. Biol. Chem. 261, 8754–8760.
8. Fallaux, F. J., Kranenburg, O., Cramer, S. J., Houweling, A., vanOrmondt, H., Hoeben, R. C., and van der Eb, A. J. (1996) Char-acterization of 911: A new helper cell line for the titration andpropagation of early region 1-deleted adenoviral vectors. Hum.Gene Ther. 7, 215–222.
9. Graham, F. L., and Prevec, L. (1995) Methods for construction ofadenovirus vectors. Mol. Biotechnol. 3, 207–220.
0. Laemmli, U. K. (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227, 680–685.
1. Salahifar, H., Baxter, R. C., and Martin, J. L. (1997) Insulin-likegrowth factor binding protein (IGFBP)-3 protease activity se-creted by MCF-7 breast cancer cells—Inhibition by IGFs doesnot require IGF-IGFBP interaction. Endocrinology 138, 1683–1690.
2. Baxter, R. C., Martin, J. L., and Beniac, V. A. (1989) Highmolecular weight insulin-like growth factor binding protein com-plex: Purification and properties of the acid-labile subunit fromhuman serum. J. Biol. Chem. 264, 11843–11848.
3. Martin, J. L., Ballesteros, M., and Baxter, R. C. (1992) Insulin-like growth factor-I (IGF-I) and transforming growth factor-b1release IGF-binding protein-3 from human fibroblasts by differ-ent mechanisms. Endocrinology 131, 1703–1710.
4. Labarca, C., and Paigen, K. (1980) A simple, rapid, and sensitiveDNA assay procedure. Anal. Biochem. 102, 344–352.
5. Birnbaum, R. S., Ware, J. L., and Plymate, S. R. (1994) Insulin-like growth factor-binding protein-3 expression and secretion bycultures of human prostate epithelial cells and stromal fibro-blasts. J. Endocrinol. 141, 535–540.
6. Camacho-Hubner, C., Busby, W. H., McCusker, R. H., Wright,G., and Clemmons, D. R. (1992) Identification of the forms ofinsulin-like growth factor-binding proteins produced by humanfibroblasts and the mechanisms that regulate their secretion.J. Biol. Chem. 267, 11949–11956.
7. Clemmons, D. R., Camacho-Hubner, C., Coronado, E., and Os-borne, C. K. (1990) Insulin-like growth factor binding proteinsecretion by breast carcinoma cell lines: Correlation with estro-gen receptor status. Endocrinology 127, 2679–2686.
8. Hana, V., and Murphy, L. J. (1994) Expression of insulin-likegrowth factors and their binding proteins in the estrogen respon-sive Ishikawa human endometrial cancer cell line. Endocrinol-ogy 135, 2511–2516.
9. Baxter, R. C. (1990) Glycosaminoglycans inhibit formation of the140 kDa insulin-like growth factor-binding protein complex. Bio-chem. J. 271, 773–777.
0. Sommer, A., Maack, C. A., Spratt, S. K., Mascarenhas, D., Tres-sel, T. J., Rhodes, E. T., Lee, R., Roumas, M., Tatsuno, G. P.,Flynn, J. A., Gerber, N., Taylor, J., Cudny, H., Nanney, L., Hunt,T. K., and Spencer, E. M. (1991) Molecular genetics and actionsof recombinant insulin-like growth factor binding protein-3 in“Modern Concepts of Insulin-like Growth Factors” (Spencer,E. M., Ed.), pp. 715–728, Elsevier, San Francisco.
1. Zhang, Y., Olsen, D. R., Nguyen, K. B., Olson, P. S., Rhodes,E. T., and Mascarenhas, D. (1998) Expression of eukaryoticproteins in soluble form in Escherichia coli. Protein Express.Purif. 12, 159–165.
2. Tressel, T. J., Tatsuno, G. P., Spratt, K., and Sommer, A. (1991)Purification and characterization of human recombinant insulin-like growth factor binding protein 3 expressed in Chinese ham-
ster ovary cells. Biochem. Biophys. Res. Commun. 178, 625–633.
3
3
3
3
3
3
3
211RECOMBINANT INSULIN-LIKE GROWTH FACTOR-BINDING PROTEIN-3
3. Chartier, C., Degryse, M., Gantzer, M., Dieterle, A., Pavirani, A.,and Mehtali, M. (1996) Efficient generation of recombinant ad-enovirus vectors by homologous recombination in Escherichiacoli. J. Virol. 70, 4805–4810.
4. He, T.-C., Zhou, S., da Costa, L. T., Yu, J., Kinzler, K. W., andVogelstein, B. (1998) A simplified system for generating recom-binant adenoviruses. Proc. Natl. Acad. Sci. USA 95, 2509–2514.
5. Fowlkes, J. L., and Serra, D. M. (1996) Characterization ofglycosaminoglycan-binding domains present in insulin-likegrowth factor-binding protein-3. J. Biol. Chem. 271, 14676–14679.
6. Conover, C. A. (1998) IGFBP regulation by proteases in “Molec-
ular Mechanisms to Regulate the Activities of Insulin-LikeGrowth Factors” (Takano, K., Hizuka, N., and Takahashi, S.-I.,Ed.), pp. 107–114, Elsevier, Tokyo.
7. Suikkari, A. M., and Baxter, R. C. (1992) Insulin-like growthfactor-binding protein-3 is functionally normal in pregnancy se-rum. J. Clin. Endocrinol. Metab. 74, 177–183.
8. Firth, S. M., and Baxter, R. C. (1999) Characterisation of recom-binant glycosylation variants of insulin-like growth factor bind-ing protein-3. J. Endocrinol. 160, 379–387.
9. Delporte, C., O’Connell, B. C., He, X., Ambudkar, I. S., Agre, P.,and Baum, B. J. (1996) Adenovirus-mediated expression of aqua-
porin-5 in epithelial cells. J. Biol. Chem. 271, 22070–22075.