adenovirus dna binding protein inhibits srcap-activated cbp and creb-mediated transcription
TRANSCRIPT

Adenovirus DNA binding protein inhibits SrCap-activated CBP andCREB-mediated transcription
Xiequn Xu,a Vera Tarakanova,a John Chrivia,b and Peter Yaciuka,*a Department of Molecular Microbiology and Immunology, Saint Louis University Health Sciences Center,
1402 South Grand Boulevard, Saint Louis, MO 63104, USAb Department of Pharmacological and Physiological Sciences, Saint Louis University Health Sciences Center,
1402 South Grand Boulevard, Saint Louis, MO 63104, USA
Received 3 March 2003; returned to author for revision 1 April 2003; accepted 21 April 2003
Abstract
The SNF2-related CBP activator protein (SrCap) is a potent activator of transcription mediated by CBP and CREB. We have previouslydemonstrated that the Adenovirus 2 DNA Binding Protein (DBP) binds to SrCap and inhibits the transcription mediated by thecarboxyl-terminal region of SrCap (amino acids 1275–2971). We report here that DBP inhibits the ability of full-length SrCap (1–2971) toactivate transcription mediated by Gal-CREB and Gal-CBP. In addition, DBP also inhibits the ability of SrCap to enhance Protein KinaseA (PKA) activated transcription of the enkaphalin promoter. DBP was found to dramatically inhibit transcription of a mammaliantwo-hybrid system that was dependent on the interaction of SrCap and CBP binding domains. We also found that DBP has no effect ontranscription mediated by a transcriptional activator that is not related to SrCap, indicating that our reported transcriptional inhibition isspecific for SrCap and not due to nonspecific effects of DBP’s DNA binding activity on the CAT reporter plasmid. Taken together, theseresults suggest a model in which DBP inhibits cellular transcription mediated by the interaction between SrCap and CBP.© 2003 Elsevier Science (USA). All rights reserved.
Introduction
The cyclic-AMP responsive element-binding protein(CREB) regulates the transcription of several genes (Danielet al., 1998; Montminy, 1997). CREB targets these genes bybinding to the sequence-specific cyclic-AMP responsiveelement (CRE) found in many promoters. An increasedlevel of cAMP stimulates gene expression from CRE-con-taining promoters. The increase in transcription is largelydue to phosphorylation of CREB at serine 133 by cAMP-activated Protein Kinase A (PKA) (Shaywitz and Green-berg, 1999). Phosphorylation of CREB by PKA facilitatesthe association of CREB with the CREB-binding protein,CBP (Chrivia et al., 1993; Kwok et al., 1994). CBP iscritical for CREB-mediated transcriptional activation (Kor-zus et al., 1998). Studies to determine what regions withinCBP were important for it to function as a CREB coactiva-
tor led to the identification of a transcriptional activationdomain defined within amino acids 227–460 (Swope et al.,1996), shown in Fig. 1A. This region overlaps a region ofCBP (amino acids 1–450) that has been shown to interactwith the Adenovirus (Ad) E1A proteins (Kurokawa et al.,1998) and is distinct from the CREB binding domain withinCBP located within amino acids 586–666 (Radhakrishnanet al., 1997).
Our previous studies utilizing CBP amino acids 227–460as bait in a yeast two-hybrid screen led to the identificationof a novel gene product that binds to CBP. This proteinshares structural features with proteins in the SNF2 proteinfamily, most notably seven regions that collectively com-prise a functional ATPase domain (Johnston et al., 1999)(see Fig. 1B). The SNF2 protein family are involved inchromatin remodeling, DNA repair, and transcriptional reg-ulation (Sudarsanam and Winston, 2000). Since we havedemonstrated that this protein can enhance the ability ofCBP to activate transcription, we have called it SrCap(SNF2-related CBP activator protein). In a more recent
* Corresponding author. Fax:�1-314-773-3403.E-mail address: [email protected] (P. Yaciuk).
R
Available online at www.sciencedirect.com
Virology 313 (2003) 615–621 www.elsevier.com/locate/yviro
0042-6822/03/$ – see front matter © 2003 Elsevier Science (USA). All rights reserved.doi:10.1016/S0042-6822(03)00386-6

study, we have demonstrated that SrCap serves as a coac-tivator for the transcription factor CREB, which is known tointeract with CBP (Johnston et al., 1999; Monroy et al.,2001).
Among a wide variety of transcription factors that phys-ically interact with CBP at several separate and definedregions (Daniel et al., 1998; Goldman et al., 1997; Mont-miny, 1997), both SrCap and the E1A proteins were shownto bind to overlapping regions on CBP (see Fig. 1A)(Johnston et al., 1999; Kurokawa et al., 1998). The bindingof the E1A proteins to CBP and to the closely relatedprotein family member, p300, are both strongly implicatedin the ability of E1A to activate cell growth and to repressdifferentiation and tissue-specific transcription (Moran,1994). We have demonstrated that wild-type E1A, but notan E1A mutant unable to bind CBP/p300, inhibits transcrip-tion in a mammalian two-hybrid CAT assay that is depen-dent on the interaction between the SrCap and CBP bindingregions. The apparent competitive inhibition of the bindinginteraction between SrCap and CBP by E1A in this systemrepresents a possible mechanism that may contribute toE1A-mediated transcriptional repression (Johnston et al.,1999).
In the course of these studies on the effect of adenovirusgene expression on SrCap, we found that the Ad 2 DNABinding Protein (DBP) binds to SrCap and strongly inhibitsSrCap-mediated transcriptional activity (Xu et al., 2001).DBP is a 529 amino acid protein that binds to both single-and double-stranded DNA and RNA (Cleghon and Klessig,1986; Neale and Kitchingman, 1990). In vitro studies haveshown that DBP can function in the initiation and elonga-tion phases of DNA replication, suggesting an in vivo modelthat DBP functions to unwind double-strand viral DNA bymultimerizing along the displaced single-stranded viral
DNA (Shenk, 1996). DBP is also tightly involved in otheressential functions important in the adenovirus life cycle,including viral RNA stability (Cleghon et al., 1989), virusassembly (Nicolas et al., 1983), determination of virus hostrange (Anderson et al., 1983), and transformation (Rice etal., 1987).
In addition, DBP is implicated in transcriptional regula-tion. With the exception of the adenovirus E4 promoter,which is slightly inhibited by DBP, DBP generally activatestranscription from viral promoters. These include the ade-novirus E1A, E2A, and major late promoters, and the ad-eno-associated virus P5 promoter (Chang and Shenk, 1990).In the major late promoter and P5 promoter, DBP increasestranscription by enhancing the binding affinity of a tran-scription factor (termed upstream stimulatory factor 1(USF1), and also known as the major late transcriptionfactor (MLTF)), to its associated binding site present in bothpromoters (Zijderveld et al., 1994). DBP also enhances theability of the transcription/replication factor, NF1, to bind toits associated DNA recognition site in the adenovirus originof replication (Stuiver and van der Vliet, 1990). This abilityof DBP to enhance the affinity of transcription factors fortheir cognate recognition sites may represent a mechanismof DBP-mediated activation of transcription. While DBPhas been shown to activate several viral promoters, theeffects of DBP on cellular promoters have not been re-ported. This is an interesting question because, similar to theE1A’s abilities to both activate and repress gene expression(Moran and Mathews, 1987), DBP may also contribute to areduction of at least a subset of host cell gene expressionwhile activating viral gene expression.
Given our previous observation that DBP binds to SrCapand strongly inhibits SrCap-mediated transcription (Xu etal., 2001), and considering that SrCap is a coactivator ofCBP (Johnston et al., 1999; Monroy et al., 2001), we testedwhether DBP is able to influence transcription of the CBPand CREB-responsive cellular enkephalin promoter.
Results and discussion
DBP inhibits SrCap-dependent activation of thePKA/CREB-activated enkephalin promoter
To test whether DBP has any effect on PKA-activatedCREB-mediated transcription, we transfected CHO cells asdescribed previously (Xu et al., 2001) with a CAT reporterconstruct containing the enkephalin promoter, pEnk12-CAT. This promoter contains two cyclic-AMP responsiveelements and has been used by others to study CREB-mediated transcription (Huggenvik et al., 1991). Also trans-fected were one or more of the following plasmids: pRc/RSV-CREB, a plasmid that expresses the CREB protein(Walton et al., 1992); pPKA, a plasmid that expresses thecatalytic subunit of protein kinase; pSrCap(1–2971); andpcDNA-DBP. As shown in Fig. 2, cotransfection of plas-
Fig. 1. Diagram of SrCap, E1A, and CREB protein binding regions on CBP(A), and CBP and DBP protein binding regions on SrCap (B). Numbersindicate the amino acid boundaries of the indicated protein binding regions.Filled boxes indicate the regions of SrCap that are homologous to the SNF2regions that comprise the ATPase domains. Additional functional andprotein binding regions for CBP can be found in Goodman and Smolik(2000) and for SrCap in Johnson et al. (1999).
616 X. Xu et al. / Virology 313 (2003) 615–621
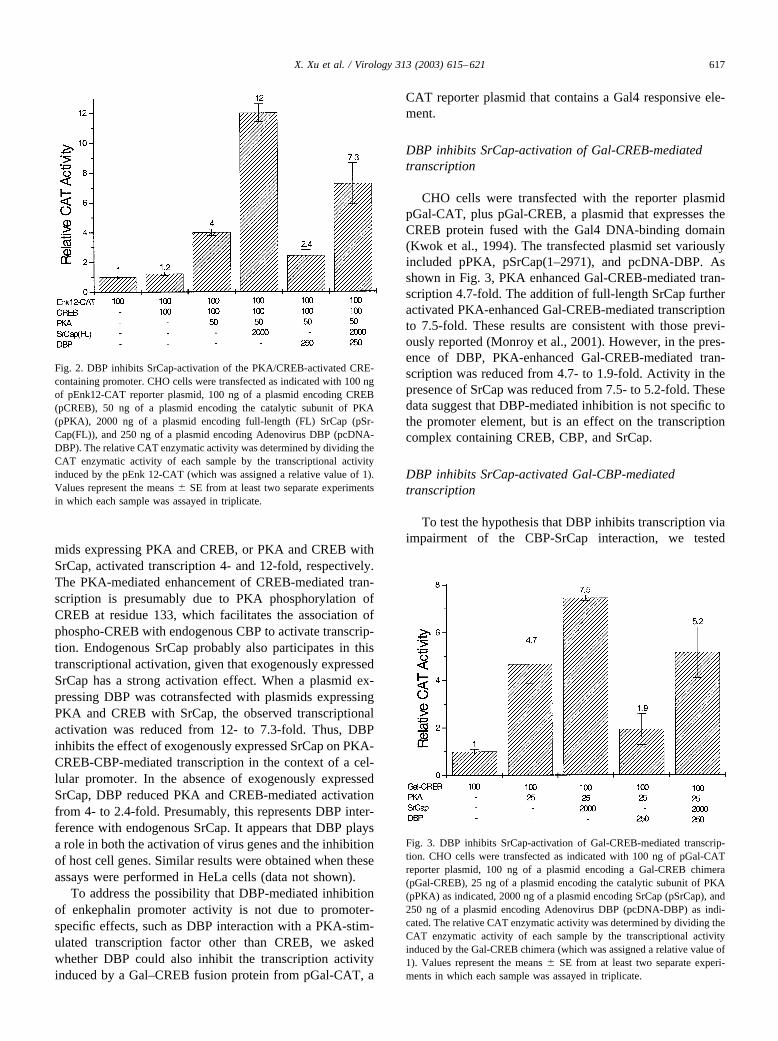
mids expressing PKA and CREB, or PKA and CREB withSrCap, activated transcription 4- and 12-fold, respectively.The PKA-mediated enhancement of CREB-mediated tran-scription is presumably due to PKA phosphorylation ofCREB at residue 133, which facilitates the association ofphospho-CREB with endogenous CBP to activate transcrip-tion. Endogenous SrCap probably also participates in thistranscriptional activation, given that exogenously expressedSrCap has a strong activation effect. When a plasmid ex-pressing DBP was cotransfected with plasmids expressingPKA and CREB with SrCap, the observed transcriptionalactivation was reduced from 12- to 7.3-fold. Thus, DBPinhibits the effect of exogenously expressed SrCap on PKA-CREB-CBP-mediated transcription in the context of a cel-lular promoter. In the absence of exogenously expressedSrCap, DBP reduced PKA and CREB-mediated activationfrom 4- to 2.4-fold. Presumably, this represents DBP inter-ference with endogenous SrCap. It appears that DBP playsa role in both the activation of virus genes and the inhibitionof host cell genes. Similar results were obtained when theseassays were performed in HeLa cells (data not shown).
To address the possibility that DBP-mediated inhibitionof enkephalin promoter activity is not due to promoter-specific effects, such as DBP interaction with a PKA-stim-ulated transcription factor other than CREB, we askedwhether DBP could also inhibit the transcription activityinduced by a Gal–CREB fusion protein from pGal-CAT, a
CAT reporter plasmid that contains a Gal4 responsive ele-ment.
DBP inhibits SrCap-activation of Gal-CREB-mediatedtranscription
CHO cells were transfected with the reporter plasmidpGal-CAT, plus pGal-CREB, a plasmid that expresses theCREB protein fused with the Gal4 DNA-binding domain(Kwok et al., 1994). The transfected plasmid set variouslyincluded pPKA, pSrCap(1–2971), and pcDNA-DBP. Asshown in Fig. 3, PKA enhanced Gal-CREB-mediated tran-scription 4.7-fold. The addition of full-length SrCap furtheractivated PKA-enhanced Gal-CREB-mediated transcriptionto 7.5-fold. These results are consistent with those previ-ously reported (Monroy et al., 2001). However, in the pres-ence of DBP, PKA-enhanced Gal-CREB-mediated tran-scription was reduced from 4.7- to 1.9-fold. Activity in thepresence of SrCap was reduced from 7.5- to 5.2-fold. Thesedata suggest that DBP-mediated inhibition is not specific tothe promoter element, but is an effect on the transcriptioncomplex containing CREB, CBP, and SrCap.
DBP inhibits SrCap-activated Gal-CBP-mediatedtranscription
To test the hypothesis that DBP inhibits transcription viaimpairment of the CBP-SrCap interaction, we tested
Fig. 3. DBP inhibits SrCap-activation of Gal-CREB-mediated transcrip-tion. CHO cells were transfected as indicated with 100 ng of pGal-CATreporter plasmid, 100 ng of a plasmid encoding a Gal-CREB chimera(pGal-CREB), 25 ng of a plasmid encoding the catalytic subunit of PKA(pPKA) as indicated, 2000 ng of a plasmid encoding SrCap (pSrCap), and250 ng of a plasmid encoding Adenovirus DBP (pcDNA-DBP) as indi-cated. The relative CAT enzymatic activity was determined by dividing theCAT enzymatic activity of each sample by the transcriptional activityinduced by the Gal-CREB chimera (which was assigned a relative value of1). Values represent the means � SE from at least two separate experi-ments in which each sample was assayed in triplicate.
Fig. 2. DBP inhibits SrCap-activation of the PKA/CREB-activated CRE-containing promoter. CHO cells were transfected as indicated with 100 ngof pEnk12-CAT reporter plasmid, 100 ng of a plasmid encoding CREB(pCREB), 50 ng of a plasmid encoding the catalytic subunit of PKA(pPKA), 2000 ng of a plasmid encoding full-length (FL) SrCap (pSr-Cap(FL)), and 250 ng of a plasmid encoding Adenovirus DBP (pcDNA-DBP). The relative CAT enzymatic activity was determined by dividing theCAT enzymatic activity of each sample by the transcriptional activityinduced by the pEnk 12-CAT (which was assigned a relative value of 1).Values represent the means � SE from at least two separate experimentsin which each sample was assayed in triplicate.
617X. Xu et al. / Virology 313 (2003) 615–621
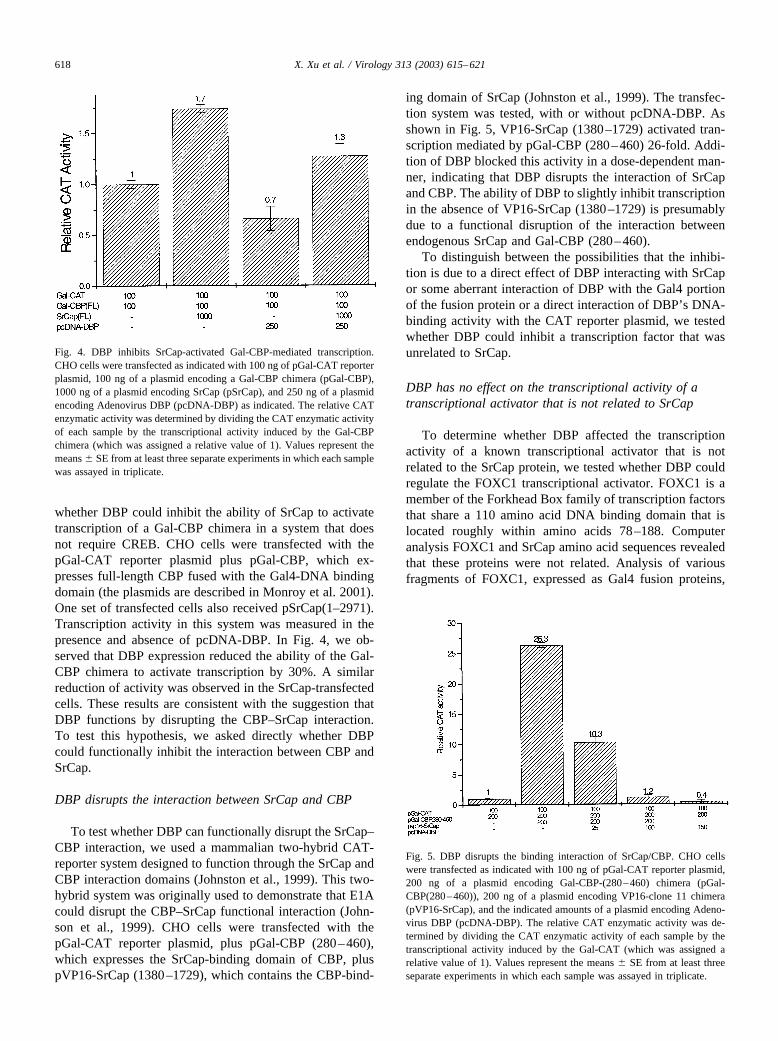
whether DBP could inhibit the ability of SrCap to activatetranscription of a Gal-CBP chimera in a system that doesnot require CREB. CHO cells were transfected with thepGal-CAT reporter plasmid plus pGal-CBP, which ex-presses full-length CBP fused with the Gal4-DNA bindingdomain (the plasmids are described in Monroy et al. 2001).One set of transfected cells also received pSrCap(1–2971).Transcription activity in this system was measured in thepresence and absence of pcDNA-DBP. In Fig. 4, we ob-served that DBP expression reduced the ability of the Gal-CBP chimera to activate transcription by 30%. A similarreduction of activity was observed in the SrCap-transfectedcells. These results are consistent with the suggestion thatDBP functions by disrupting the CBP–SrCap interaction.To test this hypothesis, we asked directly whether DBPcould functionally inhibit the interaction between CBP andSrCap.
DBP disrupts the interaction between SrCap and CBP
To test whether DBP can functionally disrupt the SrCap–CBP interaction, we used a mammalian two-hybrid CAT-reporter system designed to function through the SrCap andCBP interaction domains (Johnston et al., 1999). This two-hybrid system was originally used to demonstrate that E1Acould disrupt the CBP–SrCap functional interaction (John-son et al., 1999). CHO cells were transfected with thepGal-CAT reporter plasmid, plus pGal-CBP (280–460),which expresses the SrCap-binding domain of CBP, pluspVP16-SrCap (1380–1729), which contains the CBP-bind-
ing domain of SrCap (Johnston et al., 1999). The transfec-tion system was tested, with or without pcDNA-DBP. Asshown in Fig. 5, VP16-SrCap (1380–1729) activated tran-scription mediated by pGal-CBP (280–460) 26-fold. Addi-tion of DBP blocked this activity in a dose-dependent man-ner, indicating that DBP disrupts the interaction of SrCapand CBP. The ability of DBP to slightly inhibit transcriptionin the absence of VP16-SrCap (1380–1729) is presumablydue to a functional disruption of the interaction betweenendogenous SrCap and Gal-CBP (280–460).
To distinguish between the possibilities that the inhibi-tion is due to a direct effect of DBP interacting with SrCapor some aberrant interaction of DBP with the Gal4 portionof the fusion protein or a direct interaction of DBP’s DNA-binding activity with the CAT reporter plasmid, we testedwhether DBP could inhibit a transcription factor that wasunrelated to SrCap.
DBP has no effect on the transcriptional activity of atranscriptional activator that is not related to SrCap
To determine whether DBP affected the transcriptionactivity of a known transcriptional activator that is notrelated to the SrCap protein, we tested whether DBP couldregulate the FOXC1 transcriptional activator. FOXC1 is amember of the Forkhead Box family of transcription factorsthat share a 110 amino acid DNA binding domain that islocated roughly within amino acids 78–188. Computeranalysis FOXC1 and SrCap amino acid sequences revealedthat these proteins were not related. Analysis of variousfragments of FOXC1, expressed as Gal4 fusion proteins,
Fig. 5. DBP disrupts the binding interaction of SrCap/CBP. CHO cellswere transfected as indicated with 100 ng of pGal-CAT reporter plasmid,200 ng of a plasmid encoding Gal-CBP-(280–460) chimera (pGal-CBP(280–460)), 200 ng of a plasmid encoding VP16-clone 11 chimera(pVP16-SrCap), and the indicated amounts of a plasmid encoding Adeno-virus DBP (pcDNA-DBP). The relative CAT enzymatic activity was de-termined by dividing the CAT enzymatic activity of each sample by thetranscriptional activity induced by the Gal-CAT (which was assigned arelative value of 1). Values represent the means � SE from at least threeseparate experiments in which each sample was assayed in triplicate.
Fig. 4. DBP inhibits SrCap-activated Gal-CBP-mediated transcription.CHO cells were transfected as indicated with 100 ng of pGal-CAT reporterplasmid, 100 ng of a plasmid encoding a Gal-CBP chimera (pGal-CBP),1000 ng of a plasmid encoding SrCap (pSrCap), and 250 ng of a plasmidencoding Adenovirus DBP (pcDNA-DBP) as indicated. The relative CATenzymatic activity was determined by dividing the CAT enzymatic activityof each sample by the transcriptional activity induced by the Gal-CBPchimera (which was assigned a relative value of 1). Values represent themeans � SE from at least three separate experiments in which each samplewas assayed in triplicate.
618 X. Xu et al. / Virology 313 (2003) 615–621
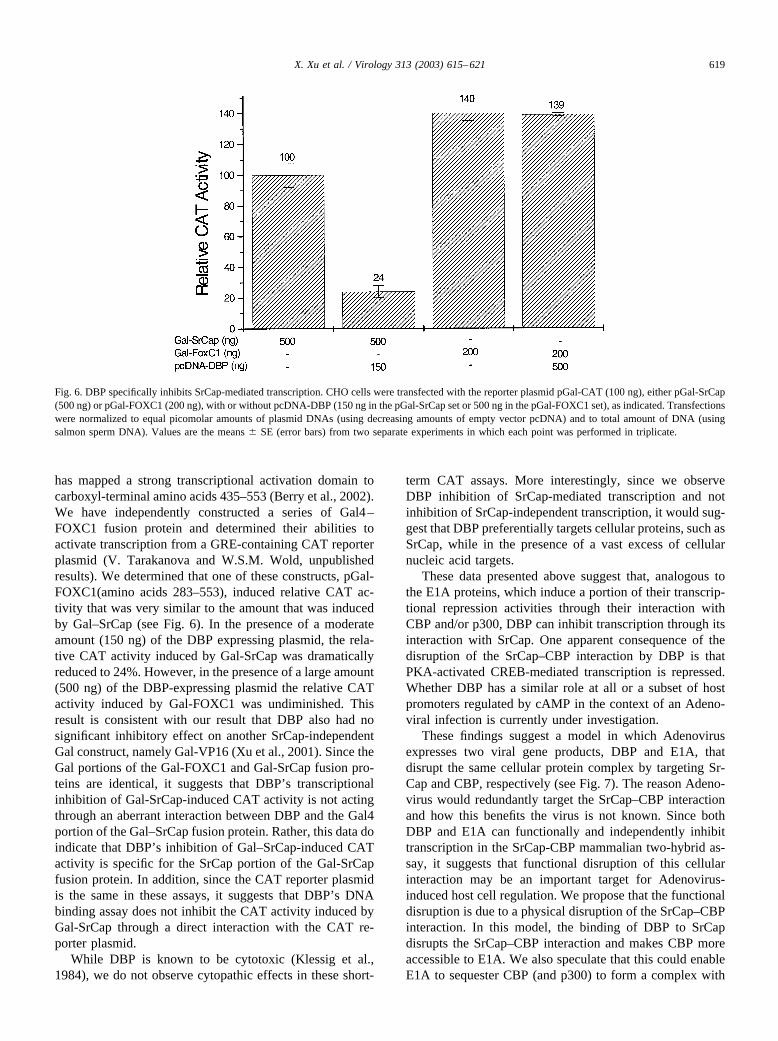
has mapped a strong transcriptional activation domain tocarboxyl-terminal amino acids 435–553 (Berry et al., 2002).We have independently constructed a series of Gal4–FOXC1 fusion protein and determined their abilities toactivate transcription from a GRE-containing CAT reporterplasmid (V. Tarakanova and W.S.M. Wold, unpublishedresults). We determined that one of these constructs, pGal-FOXC1(amino acids 283–553), induced relative CAT ac-tivity that was very similar to the amount that was inducedby Gal–SrCap (see Fig. 6). In the presence of a moderateamount (150 ng) of the DBP expressing plasmid, the rela-tive CAT activity induced by Gal-SrCap was dramaticallyreduced to 24%. However, in the presence of a large amount(500 ng) of the DBP-expressing plasmid the relative CATactivity induced by Gal-FOXC1 was undiminished. Thisresult is consistent with our result that DBP also had nosignificant inhibitory effect on another SrCap-independentGal construct, namely Gal-VP16 (Xu et al., 2001). Since theGal portions of the Gal-FOXC1 and Gal-SrCap fusion pro-teins are identical, it suggests that DBP’s transcriptionalinhibition of Gal-SrCap-induced CAT activity is not actingthrough an aberrant interaction between DBP and the Gal4portion of the Gal–SrCap fusion protein. Rather, this data doindicate that DBP’s inhibition of Gal–SrCap-induced CATactivity is specific for the SrCap portion of the Gal-SrCapfusion protein. In addition, since the CAT reporter plasmidis the same in these assays, it suggests that DBP’s DNAbinding assay does not inhibit the CAT activity induced byGal-SrCap through a direct interaction with the CAT re-porter plasmid.
While DBP is known to be cytotoxic (Klessig et al.,1984), we do not observe cytopathic effects in these short-
term CAT assays. More interestingly, since we observeDBP inhibition of SrCap-mediated transcription and notinhibition of SrCap-independent transcription, it would sug-gest that DBP preferentially targets cellular proteins, such asSrCap, while in the presence of a vast excess of cellularnucleic acid targets.
These data presented above suggest that, analogous tothe E1A proteins, which induce a portion of their transcrip-tional repression activities through their interaction withCBP and/or p300, DBP can inhibit transcription through itsinteraction with SrCap. One apparent consequence of thedisruption of the SrCap–CBP interaction by DBP is thatPKA-activated CREB-mediated transcription is repressed.Whether DBP has a similar role at all or a subset of hostpromoters regulated by cAMP in the context of an Adeno-viral infection is currently under investigation.
These findings suggest a model in which Adenovirusexpresses two viral gene products, DBP and E1A, thatdisrupt the same cellular protein complex by targeting Sr-Cap and CBP, respectively (see Fig. 7). The reason Adeno-virus would redundantly target the SrCap–CBP interactionand how this benefits the virus is not known. Since bothDBP and E1A can functionally and independently inhibittranscription in the SrCap-CBP mammalian two-hybrid as-say, it suggests that functional disruption of this cellularinteraction may be an important target for Adenovirus-induced host cell regulation. We propose that the functionaldisruption is due to a physical disruption of the SrCap–CBPinteraction. In this model, the binding of DBP to SrCapdisrupts the SrCap–CBP interaction and makes CBP moreaccessible to E1A. We also speculate that this could enableE1A to sequester CBP (and p300) to form a complex with
Fig. 6. DBP specifically inhibits SrCap-mediated transcription. CHO cells were transfected with the reporter plasmid pGal-CAT (100 ng), either pGal-SrCap(500 ng) or pGal-FOXC1 (200 ng), with or without pcDNA-DBP (150 ng in the pGal-SrCap set or 500 ng in the pGal-FOXC1 set), as indicated. Transfectionswere normalized to equal picomolar amounts of plasmid DNAs (using decreasing amounts of empty vector pcDNA) and to total amount of DNA (usingsalmon sperm DNA). Values are the means � SE (error bars) from two separate experiments in which each point was performed in triplicate.
619X. Xu et al. / Virology 313 (2003) 615–621

Rb. This resulting complex formed between E1A, CBP (orp300), and Rb has been shown to be essential for theabilities of E1A proteins to fully activate the cell cycle(Wang et al., 1995). In addition, in the E1A/CBP/p300/Rbcomplex, the intrinsic CBP/p300 acetyltransferase activitieshave been shown to acetylate Rb and regulate the ability ofRb to form a protein–protein interaction with MDM2 (Chanet al., 2001). In addition, since SrCap could no longercoactivate CBP-mediated transcription, a portion of CBP-mediated transcription of CBP-regulated cellular geneswould be repressed. Taken together, these results suggest anintriguing DBP-mediated mechanism of host cell transcrip-tional regulation.
Acknowledgments
We thank William S.M. Wold and Betty Moran forcomments, discussions, and for reading this manuscript. Wethank Elizabeth Pesek for expert technical assistance and forreading this manuscript. This work was supported by PublicHealth Service Grant CA-68066 from the National CancerInstitute.
References
Anderson, C.W., Hardy, M.M., Dunn, J.J., Klessig, D.F., 1983. Indepen-dent, spontaneous mutants of adenovirus type 2-simian virus 40 hybridAd2 � ND3 that grow efficiently in monkey cells possess identicalmutations in the adenovirus type 2 DNA-binding protein gene. J. Virol.48, 31–39.
Berry, F.B., Saleem, R.A., Walter, M.A., 2002. FOXC1 transcriptionalregulation is mediated by N- and C-terminal activation domains andcontains a phosphorylated transcriptional inhibitory domain. J. Biol.Chem. 277, 10292–10297.
Chan, H.M., Krstic-Demonacos, M., Smith, L., Demonacos, C., LaThangue, N.B., 2001. Acetylation control of the retinoblastoma tu-mour-suppressor protein. Nat. Cell Biol. 3, 667–674.
Chang, L.S., Shenk, T., 1990. The adenovirus DNA-binding protein stim-ulates the rate of transcription directed by adenovirus and adeno-associated virus promoters. J. Virol. 64, 2103–2109.
Chrivia, J.C., Kwok, R.P., Lamb, N., Hagiwara, M., Montminy, M.R.,Goodman, R.H., 1993. Phosphorylated CREB binds specifically to thenuclear protein CBP. Nature 365, 855–859.
Cleghon, V., Voelkerding, K., Morin, N., Delsert, C., Klessig, D.F., 1989.Isolation and characterization of a viable adenovirus mutant defectivein nuclear transport of the DNA-binding protein. J. Virol. 63, 2289–2299.
Cleghon, V.G., Klessig, D.F., 1986. Association of the adenovirus DNA-binding protein with RNA both in vitro and in vivo. Proc. Natl. Acad.Sci. USA 83, 8947–8951.
Daniel, P.B., Walker, W.H., Habener, J.F., 1998. Cyclic AMP signalingand gene regulation. Annu. Rev. Nutr. 18, 353–383.
Goldman, P.S., Tran, V.K., Goodman, R.H., 1997. The multifunctional roleof the co-activator CBP in transcriptional regulation. Recent Prog.Horm. Res. 52, 103–119.
Goodman, R.H., Smolik, S., 2000. CBP/p300 in cell growth, transforma-tion, and development. Genes Dev. 14, 1553–1577.
Huggenvik, J.I., Collard, M.W., Stofko, R.E., Seasholtz, A.F., Uhler, M.D.,1991. Regulation of the human enkephalin promoter by two isoforms ofthe catalytic subunit of cyclic adenosine 3�,5�-monophosphate-depen-dent protein kinase. Mol. Endocrinol. 5, 921–930.
Johnston, H., Kneer, J., Chackalaparampil, I., Yaciuk, P., Chrivia, J., 1999.Identification of a novel SNF2/SWI2 protein family member, SRCAP,which interacts with CREB-binding protein. J. Biol. Chem. 274,16370–16376.
Klessig, D.F., Brough, D.E., Cleghon, V., 1984. Introduction, stable inte-gration, and controlled expression of a chimeric adenovirus gene whoseproduct is toxic to the recipient human cell. Mol. Cell Biol. 4, 1354–1362.
Korzus, E., Torchia, J., Rose, D.W., Xu, L., Kurokawa, R., McInerney,E.M., Mullen, T.M., Glass, C.K., Rosenfeld, M.G., 1998. Transcriptionfactor-specific requirements for coactivators and their acetyltransferasefunctions. Science 279, 703–707.
Kurokawa, R., Kalafus, D., Ogliastro, M.H., Kioussi, C., Xu, L., Torchia,J., Rosenfeld, M.G., Glass, C.K., 1998. Differential use of CREBbinding protein-coactivator complexes. Science 279, 700–703.
Kwok, R.P., Lundblad, J.R., Chrivia, J.C., Richards, J.P., Bachinger, H.P.,Brennan, R.G., Roberts, S.G., Green, M.R., Goodman, R.H., 1994.Nuclear protein CBP is a coactivator for the transcription factor CREB[see comments]. Nature 370, 223–226.
Fig. 7. Diagram of the functional inhibition of the interaction between SrCap and CBP by adenoviral proteins E1A and DBP. The left side shows an activepromoter containing a cyclic AMP responsive element binding protein (CREB) bound to a cyclic AMP responsive element (CRE). CREB has beenphosphorylated by Protein Kinase A (PKA) and recruits the CREB binding protein (CBP) to the promoter site. The association of SrCap with CBP furtheractivates transcription from this promoter, as indicated by the rightward arrow at the transcriptional start site. The introduction of adenoviral proteins E1Aand/or DBP inhibits transcription, as indicated by the “X” on the rightward arrow at the transcriptional start site. The model on the right indicates thatfunctional inhibition of the SrCap-CBP complex is by physical disruption of the complex by E1A and/or DBP.
620 X. Xu et al. / Virology 313 (2003) 615–621

Monroy, M.A., Ruhl, D.D., Xu, X., Granner, D.K., Yaciuk, P., Chrivia,J.C., 2001. Regulation of cAMP-responsive element-binding protein-mediated transcription by the SNF2/SWI-related protein, SRCAP.J. Biol. Chem. 276, 40721–40726.
Montminy, M., 1997. Transcriptional regulation by cyclic AMP. Annu.Rev. Biochem. 66, 807–822.
Moran, E., 1994. Mammalian cell growth control reflected through proteininteractions with the adenovirus E1A gene products. Sem. Virol. 5,327–340.
Moran, E., Mathews, M.B., 1987. Multiple functional domains in theadenovirus E1A gene. Cell 48, 177–178.
Neale, G.A., Kitchingman, G.R., 1990. Conserved region 3 of the adeno-virus type 5 DNA-binding protein is important for interaction withsingle-stranded DNA. J. Virol. 64, 630–638.
Nicolas, J.C., Sarnow, P., Girard, M., Levine, A.J., 1983. Host rangetemperature-conditional mutants in the adenovirus DNA binding pro-tein are defective in the assembly of infectious virus. Virology 126,228–239.
Radhakrishnan, I., Perez-Alvarado, G.C., Parker, D., Dyson, H.J., Mont-miny, M.R., Wright, P.E., 1997. Solution structure of the KIX domainof CBP bound to the transactivation domain of CREB: a model foractivator:coactivator interactions. Cell 91, 741–752.
Rice, S.A., Klessig, D.F., Williams, J., 1987. Multiple effects of the72-kDa, adenovirus-specified DNA binding protein on the efficiency ofcellular transformation. Virology 156, 366–376.
Shaywitz, A.J., Greenberg, M.E., 1999. CREB: a stimulus-induced tran-scription factor activated by a diverse array of extracellular signals.Annu. Rev. Biochem. 68, 821–861.
Shenk, T., 1996. Adenoviridae: the viruses and their replication. in: Field,B.N., Knipe, D.M., Howley, P.M. (Eds.), Fields Virology, Vol. 2.Lippincott-Raven, Philadelphia, pp. 2111–2148.
Stuiver, M.H., van der Vliet, P.C., 1990. Adenovirus DNA-binding proteinforms a multimeric protein complex with double-stranded DNA andenhances binding of nuclear factor I. J. Virol. 64, 379–386.
Sudarsanam, P., Winston, F., 2000. The Swi/Snf family nucleosome-remodeling complexes and transcriptional control. Trends Genet. 16,345–351.
Swope, D.L., Mueller, C.L., Chrivia, J.C., 1996. CREB-binding proteinactivates transcription through multiple domains. J. Biol. Chem. 271,28138–28145.
Walton, K.M., Rehfuss, R.P., Chrivia, J.C., Lochner, J.E., Goodman, R.H.,1992. A dominant repressor of cyclic adenosine 3�,5�-monophosphate(cAMP)-regulated enhancer-binding protein activity inhibits thecAMP-mediated induction of the somatostatin promoter in vivo. Mol.Endocrinol. 6, 647–655.
Wang, H.G., Moran, E., Yaciuk, P., 1995. E1A promotes associationbetween p300 and pRB in multimeric complexes required for normalbiological activity. J. Virol. 69, 7917–7924.
Xu, X., Chackalaparampil, I., Monroy, M.A., Cannella, M.T., Pesek, E.,Chrivia, J., Yaciuk, P., 2001. Adenovirus DNA binding protein inter-acts with the SNF2-related CBP activator protein (SrCap) and inhibitsSrCap-mediated transcription. J. Virol. 75, 10033–10040.
Zijderveld, D.C., d’Adda, d.F., Giacca, M., Timmers, H.T., van der Vliet,P.C., 1994. Stimulation of the adenovirus major late promoter in vitroby transcription factor USF is enhanced by the adenovirus DNA bind-ing protein. J. Virol. 68, 8288–8295.
621X. Xu et al. / Virology 313 (2003) 615–621