adenovirus type 2 assembly analyzed reversible …jvi.asm.org/content/26/2/357.full.pdf · crose...
TRANSCRIPT

JOURNAL OF VIROLOGY, May 1978, p. 357-363 Vol. 26, No. 20022-538X/78/0026-0357$02.00/0Copyright © 1978 American Society for Microbiology Printed in U.S.A.
Adenovirus Type 2 Assembly Analyzed by Reversible Cross-Linking of Labile Intermediates
JEAN-CLAUDE D'HALLUIN, GUY R. MARTIN, G1RRARD TORPIER, ANDPIERRE A. BOULANGER*
Unite de Recherches de Virologie no. 102 de l'Institut National de la Sante et de la Recherche Medicale,Lille, France
Received for publication 13 October 1977
Dimethyl-4,4'-dithiobisbutyrimidate dihydrochloride was used as a cleavablecross-linking reagent to maintain the structure of labile intermediates in adeno-virus type 2 assembly. Analysis on sucrose gradients of nuclear adenovirusparticles revealed two size classes, with sedimentation rates of 750 and 600S. Afterreversible fixation with diimido ester, the different classes were further separatedon CsCl gradients and characterized with regard to their buoyant density, DNAcontent, and polypeptide composition. The 750S particles banded at 1.345 g/cm3in CsCl, contained a DNA with a sedimentation coefficient of 34S in alkalinesucrose gradients, and had a polypeptide composition similar to that of youngvirions. The 600S population consisted of two types of particles with buoyantdensities of 1.315 and 1.37 g/cm3. The 1.315-g/cm3 particles contained a DNAfragment of 7-11S and lacked the core proteins V and VII. In their place werefound precursors P VI and P VIII and two nonvirion proteins with molecularweights of 50,000 (50K) and 39,000 (39K). 34S DNA was present in the 1.37-g/cm3particles, which also lacked core proteins V and VII, as well as the 50K and 39K.Pulse-chase labeling kinetics suggested that the 1.315-g/cm3 particles were ante-rior to the 1.37-g/cm3 particles, themselves preceding the 1.345-g/cm3 youngvirions, and that the release of both 50K and 39K, possible scaffolding proteins,was required for entry of viral DNA.
Incomplete virus particles that are copro- labile adenovirus type 2 (Ad2) wild-type inter-duced with mature adenovirus during produc- mediate particles in CsCl gradients and to ana-tive infection have been identified as interme- lyze the DNA content and polypeptide compo-diates in adenovirus assembly (4, 9, 12). They sition of each class. Through the use of pulse-differ from mature virions in DNA content and chase labeling kinetics, identification of somepolypeptide composition. Centrifugation in CsCl steps in the adenovirus assembly pathway be-gradients has been, therefore, widely used for came possible.isolation and characterization of the differentclasses of particles, taking advantage of their MATERIALS AND METHODSdifferences in buoyant density.
However, it has been recently demonstrated Virus and celis. Human Ad2, ori inally suppliedthat some of the adenovirus particle classes can by J.F. Williams (Carnegie-Mellon Institute, Pitts-be artifactually generated in the course of cell burgh, Pa.), was grown on KB cells maintained inferactioactio andeioatediona tha frae iner- suspension culture with Joklik-modified medium F 13mdioates candisieateo and ec frapleinvesa- (Grand Island Biological Co., Grand Island, N.Y.) sup-mediates can disintegrate and escape investiga- plemented with 5% horse serum.tion (4). The necessity of using a mild isolation Infection and labeling conditions. KB cells wereprocedure and glutaraldehyde fixation before infected at a multiplicity of infection of 25 PFU perCsCl gradient centrifugation has been therefore cell. The cells were centrifuged at low speed at differ-clearly established (4). ent times postinfection and suspended at a density ofThe major drawback of this glutaraldehyde lo' cells per ml in culture medium containing 2.5% of
fixation resides in its irreversibility, which does the normal methionine concentration. Proteins werenot permit further analysis of the different labeled with [3S]methionine (20 jACi/ml), and DNAclasses of fixed particles isolated by CsCl gra- was labeled with [3H]thymidine (20 ,uCi/ml).dientcentrifugation.Usin amildproceduRadioisotopes and counting method. [3S]me-dient centrifugation. Using a mild procedure of thionine (600 to 700 Ci/mmol) was purchased from
cell extraction and a reversible cross-linking of the Radiochemical Centre (Amersham, England), andvirus particles by a cleavable diimido ester (13), [3H]thymidine (30 Ci/mmol) was purchased from theit was possible to isolate and characterize some Comn 'ssatiat a l'Energie Atomique (Saclay, France).
357
on Septem
ber 16, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

358 D'HALLUIN ET AL. J. VIROL.
The radioactive samples were precipitated with cold coil (Pharmacia Fine Chemicals AB, Uppsala, Swe-10% trichloroacetic acid and filtered on Whatman GF- den) and sucrose (E. Merck A.G., Darmstadt, WestC filters. The filters were washed successively with Germany) gradients were used for the isolation ofcold 5% trichloroacetic acid and cold ethanol and air- virus and assembly intermediate particles from celldried. They were counted in a toluene scintillation extracts. The gradients were made of 5 to 12.5% Ficollfluid [5 g of 2,5-diphenyloxazole-0.3 g of 1,4-bis-(5- in 20 mM sodium borate, pH 8.0-150 mM NaCI-10phenyloxazolyl)benzene in 1 liter of toluene, scintilla- mM sodium EDTA or 25 to 40% sucrose in 20 mMtion grade] with an Intertechnique liquid spectrome- sodium borate, pH 8.0-200 mM NaCl-10 mM sodiumter. EDTA and centrifuged at 85,000 x g for 105 min at
Virus production and purification. Cells were 40C in an SW 27 rotor (4). A cushion of CsCl (1.43concentrated 100-fold in hypotonic buffer (50 mM g/ml) was placed at the bottom of each gradient. TheTris-hydrochloride, pH 8.0-50 mM NaCl-1 mM so- gradients were collected dropwise from the bottoms ofdium EDTA) and subjected to three cycles of freezing the tubes, and the fractions were assayed for trichlo-and thawing, and the cell lysate was extracted with an roacetic acid-precipitable radioactivity. The differentequal volume of Freon 113, as previously described populations of particles thus isolated were fixed before(1). The aqueous phase of the Freon step was saved, further electron microscopic and biochemical analyses.layered on top of a CsCl cushion (1.43 g/cm3), and Reversible fixation of virus particles. Samplescentrifuged for 1 h at 20,000 rpm and 4°C in an SW 27 from Ficoll or sucrose gradients containing completerotor. The opalescent band of virus on top of the CsCl or incomplete virus particles were reacted with 3 mgcushion was collected and purified by equilibrium cen- of methyl-4-mercaptobutyrimidate hydrochloridetrifugation in a self-generating CsCl gradient (1.34 (Pierce Chemical Co., Rockford, Ill.) per ml for 30 ming/cm3) at 30,000 rpm and 40C for 16 h in an SW 50 at 4°C in a medium devoid of primary amine (20 mMrotor. After dilution in 3 volumes of 50 mM Tris- sodium borate buffer, pH 8.0-200 mM NaCl-10 mMhydrochloride, pH 8.0-1 mM sodium EDTA, the virus sodium EDTA). After amidination of proteins, disul-was layered on top of a linear gradient (9 ml; 25 to fide bridge formation was induced by dialysis against40%, wt/vol) made in 50 mM Tris-hydrochloride, pH phosphate-buffered saline containing 100 mM H202.8.0-200 mM NaCl-10 mM sodium EDTA-0.25% Tri- The reversibly fixed virus particles could then beton X-100, placed over two cushions of 1 ml each of further analyzed by centrifugation on a self-generatingCsCl solutions (1.43 and 1.31 g/cm3), and centrifuged CsCl gradient (1.34 g/ml, containing 100 mM H202),for 2 h at 25,000 rpm and 4°C in an SW 41 rotor. The by sodium dodecyl sulfate (SDS)-polyacrylamide gelvirus band concentrated between the two cushions electrophoresis, and by electron microscopy, and thewas collected, and an equal volume of glycerol was nature of their DNA could be determined. The in-added to this virus suspension. The virus was stored duced disulfide cross-bridges could be cleaved by re-in 1-ml portions at -20°C. ducing agents, such as 2-mercaptoethanol or dithio-
Infectivity was assayed by the fluorescent focus threitol, as described in the following section.assay technique (11) or the plaque method on HeLa Analytical SDS-polyacrylamide gel electro-cells grown as monolayers in petri dishes. The titer phoresis. The different diimido ester-fixed virus pop-was expressed as fluorescent focus units or PFU. ulations separated in sucrose or Ficoll gradients and
Cell fractionation and extraction of assembly subsequently in CsCl gradients were precipitated over-intermediates. Infected cells were washed with cold night at 4°C with 10% trichloroacetic acid. The precip-phosphate-buffered saline, suspended at 4 x 107 cells itate was centrifuged and washed successively withper ml in reticulocyte standard buffer (10 mM Tris- cold 5% and 0.5% trichloroacetic acid and then dis-hydrochloride, pH 7.4-10 mM NaCl-1.5 mM MgCl2), solved in SDS denaturing mix (62.5 mM Tris-hydro-and left for swelling at 0°C for 10 min. Triton X-100 chloride, pH 6.8-4% SDS-10% 2-mercaptoethanol-6was added up to a final concentration of 0.5%, and the M urea) and heated for 2 min at 100'C. The polypep-cells were disrupted by 10 strokes in a tight-fitting tides were analyzed in a 17.5% acrylamide-0.08% bis-Dounce homogenizer. NaCl was added up to 100 mM, acrylamide slab gel overlayered by a 5% acrylamide-and the cell lysate was centrifuged at 1,000 x g for 5 0.13% bisacrylamide spacer gel in the discontinuousmin. The supernatant, referred to as the cytoplasmic buffer system of Laemmli (10). Electrophoresis wasfraction, was adjusted to 20 mM sodium EDTA to carried out for 16 h at 30 V (constant voltage) with adissociate the polysomes and centrifuged at 16,000 x Bio-Rad model 220 electrophoresis unit. The gels wereg for 10 min before analysis on sucrose or Ficoll fixed and stained with 0.3% Coomassie brilliant bluegradients as described below. R-250 in 12% acetic acid-30% methanol, destained inThe nuclei in the 1,000 x g pellet were lysed by the the same fluid, dried under vacuum, and autoradi-
technique described by Wallace and Kates (15). They ographed on Kodak Kodirex film.were suspended in 50 mM Tris-hydrochloride, pH Analysis of virus DNA. The DNA content of8.0-10 mM sodium EDTA (TE buffer) at 0°C, adjusted reversibly fixed mature virions and intermediates iso-to 0.3 M (NH4)2SO4, homogenized in a tight-fitting lated in sucrose or Ficoll gradients and further sepa-Dounce homogenizer (three strokes), and immediately rated on self-generating CsCl gradients was analyzeddiluted with 2 volumes of TE buffer. The nuclear according to a modification of the procedure describedlysate was then centrifuged over a 30% (wt/vol) su- by Doerfler (3). The virus suspension was adjusted tocrose cushion at 16,000 x g for 10 min. The nuclear 0.5 M NaOH-50 mM sodium EDTA-10% 2-mercap-supernatant was further analyzed on a sucrose or a toethanol and layered on top of a 5 to 20% (wt/vol)Ficoll gradient. alkaline sucrose gradient made in 0.3 M NaOH-0.7 MSucrose and Ficoll gradient centrifugation. Fi- NaCl-1 mM sodium EDTA. After standing for 16 h at
on Septem
ber 16, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

VOL. 26, 1978 Ad2 ASSEMBLY INTERMEDIATES 359
4°C, the gradient was centrifuged for 5 hat 35,000 rpm ible at 1.5 h of chase and marked at 2.5 h ofand 4°C in an SW 41 rotor. Fractions were collected chase. A second peak (fractions 21 to 25), rela-dropwise from the bottom and assayed for trichloroa- tively homogeneous in shape, contained viruscetic acid-precipitable radioactivity.. . .Eeactpromicroscopy.cthedifferenttclas. of particles sedimenting at about 600S, with a ratioElectron microscopy. The different classes of DAt rti oe hnta fanradiimido ester-fixed virus particles obtained from su- of DNA to protei lower than that of a normalcrose or Ficoll gradients and further isolated in self- vron. This peak was more important after agenerating CsCl gradients were examined in a Hitachi 1.5-h chase than after 2.5 h; it corresponded toHU-12 electron microscope after staining with 1% Ad2 assembly intermediates (4). In mock-in-potassium phosphotungstate, pH 7.2. fected cell nucleus extracts obtained in the same
way, no peak of nucleoproteinic material wasRESULTS found in that gradient zone (not shown). Similar
Isolation and reversible fixation of Ad2 results were obtained with Ficoll gradients (notmature and intermediate particles. KB cells shown).in suspension culture were infected with wild- The fractions corresponding to each peak weretype Ad2, concentrated to a density of 4 x 107 pooled, and the virus particles were cross-linkedcells per ml, pulse-labeled for 15 min at 16 h with the cleavable diimido ester dimethyl-4,4'-postinfection with [3S]methionine and [3H]thy- dithiobisbutyrimidate dihydrochloride, formedmidine, and chased for 1.5 and 2.5 h. The cells by oxidation of neighboring thiol groups carriedwere fractionated into cytoplasm and nucleus. by proteins previously amidinated with methyl-Mock-infected cells were treated in the same 4-mercaptobutyrimidate (8, 13).way. The nuclei from Ad2-infected and mock- Buoyant density analysis of reversiblyinfected cells were lysed with 0.3 M (NH4)2S04 fixed Ad2 particles. The different populationsin TE buffer, and the nuclear lysate was ana- of fixed virus particles were then analyzed onlyzed on a sucrose gradient. Two distinct peaks self-generating CsCl gradients (mean density,of virus particles were visible on the sucrose 1.34 g/cm3). The virus particles sedimenting atgradient pattern of nuclear extract (Fig. 1). A 750S in sucrose gradients were found banding asfirst peak of virus particles sedimented in the a sharp peak at 1.345 g/cm3, the normal buoyantposition of mature adenovirion marker at 750S density for mature and young adenovirions (Fig.(fractions 14 to 17). This peak was scarcely vis- 2a). The material sedimenting at 600S was found
to consist of two populations of virus particles,with densities of 1.37 and 1.315 g/cm3. The latter
a b constituted the major class and had a lower ratioofDNA to protein label than did the 1.37-g/cm3
60- Rspecies (Fig. 2b).DNA analysis of Ad2 particles. Each pop-
40 Q 8s ulation was analyzed with regard to its DNA20- A't t content. DNA extracted under mfld conditions
- was analyzed in alkaline sucrose gradients. Both10 ~ O l t ° 1.345- and 1.37-g/cm3 particles contained a DNA
6 sedimenting at 34S, the expected value for de-natured complete Ad2 DNA. The 1.315-g/cm3particles contained a DNA fragment sediment-
5 ° n °° t o ing as a broad peak between 7 and llS (not
Polypeptide composition of the differentclasses of Ad2 particles. The protein compo-
10 20 30 4.10 20 30 40 sition of the different classes of particles was10 2'0 30 410 10 2'0 310 40 analyzed on SDS-polyacrylamide slab gels afterfraction number cleavage of the disulfide cross-bridges with SDS
FIG. 1. Velocity gradient analysis ofwild- typeAd2 denaturing mix. Figure 3 shows the polypeptidenuclear particles. Ad2-infected KB-cell cultures were pattern of each class of particles, compared withpulse-labeled with I:HJthymidine and [3S]methio- that of a mature adenovirion isolated at the endnine for 15 min at 16 h p.i., and the label was chased of a productive cycle by conventional techniquesfor 1.5 h (a) and 2.5 h (b). The nuclear virus particles (6). Some differences could be discerned be-were extracted with (NH,22SO4 as in the text andanalyzed on a 25 to 40% sucrose gradient with a CsCl tween the nuclear 1.345-g/cm3 particles (Fig. 3e)cushion at the bottom (1.43 g/cm3). The arrow indi- and the mature adenovirions (Fig. 3f). The pre-cates the position of mature adenovirus marker, sed- cursor proteins (P VI, P VIII, and P VII) wereimenting with an apparent sedimentation coefficient present, along with their cleavage products (vir-of 750S. Symbols: 0, 3S; 0, 3H. Bottom is at the left. ion proteins VI, VIII, and VII), in the 1.345-
on Septem
ber 16, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

360 D'HALLUIN ET AL. J. VIROL.
a b V a d e
50-
340 i ]
12001,38 a !. -K0) 5
a. 50K13f1r34ction VbrV
10 -1.30
1 *t. Pv1II10 20 30 10 20 30
fraction number
FIG. 2. Buoyant density analysis of diimido ester-fixed adenovirus particles in CsCl gradients. The _ PVIIfractions corresponding to each virus particle peak vii m Wobtained in sucrose gradients, as in Fig. lb, were Wpooled, fixed with the bifunctional reagent dimethyl-4,4'-dithiobisbutyrimidate dihydrochloride, and fur-ther separated in a self-generating CsCl gradient(mean density: 1.34 g/cm3). (a) Lower peak of 750Sparticles; (b) upper peak of 60OS particles. Symbols: V Vi0, I3Slmethionine; 0, ['H]thymidine. Bottom is at xthe left.
g/cm3 particles, thus suggesting that they wereyoung virions (4, 9). The 1.315-g/cm3 particles FIG. 3. SDS-polyacrylamide gel electrophoretic(Fig. 3c) totally lacked core proteins V and VII analysis of Ad2 particles isolated on sucrose andbut possessed the precursors P VI and P VIII. CsCl gradients after reversible fixation with a cleav-In addition they contained two nonvirion pro- able diimido ester. Virus particles extracted fromteins, with molecular weights of 50,000 (50K) infected cell nuclei after a 15-min pulse with [5S]-and 39,000 (39K). Traces ofP VII were occasion- methionine at 16 h p.i., followed by a 2.5-h chase,ally visible. P VII were found in the intermediate were first isolated in sucrose gradients, reversiblypeak of the sucrose gradient before fixation (Fig. fixed, and further separated in CsCl gradients. (a)3a) and seemed to disappear almost completely 600S intermediates isolated in a sucrose gradient,from the intermnediates after fixation and further before fixation, as in Fig. Ib; (b) nonfixed 750S virusfromthenterediaes ater ixaton ad futherparticles from a sucrose gradient; (c) reversibly fixedpurification in a CsCl gradient (Fig. 3c). P VII intermediates of 1.315 g/cm3 of buoyant density, ob-was, in fact, present in all sucrose gradient frac- tained from a CsCl gradient, as in Fig. 2b; (d) revers-tions (not shown) and, most likely, represented ibly fixed intermediates banding at 1.37 g/cm3 incontaminating protein bound to particulate CsCl; (e) reversibly fixed 750S particles banding atstructures and sedimenting with a broad S range. 1.345 g/cm3 in CsCl, as in Fig. 2a; (v) control adeno-The absence of P VII in the purified 1.315-g/cm3 virus. Anode is at the bottom.particles did not result from an incomplete cleav-age of cross-linked P VII, since the same reduc- and 50K were absent from the 1.37-g/cm3 spe-ing conditions on 1.345-g/cm3 particles released cies, and the V was present only in minuteP VII monomer (Fig. 3e). amounts. Both 1.315- and 1.37-g/cm3 particlesThe 1.37-g/cm3 particles also contained pre- lacked core structure sedimenting at 180-200S
cursors P VI and P VIII and, occasionally, traces (7) after 0.5% deoxycholate treatment at 56°Cof P VII but totally lacked core protein VII (Fig. (not shown). The 1.345-, 1.315-, and 1.37-g/cm33d). In contrast to 1.315-g/cm3 particles the 39K particles occasionally contained a protein, 100K,
on Septem
ber 16, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
VOL. 26, 1978 Ad2 ASSEMBLY INTERMEDIATES 361
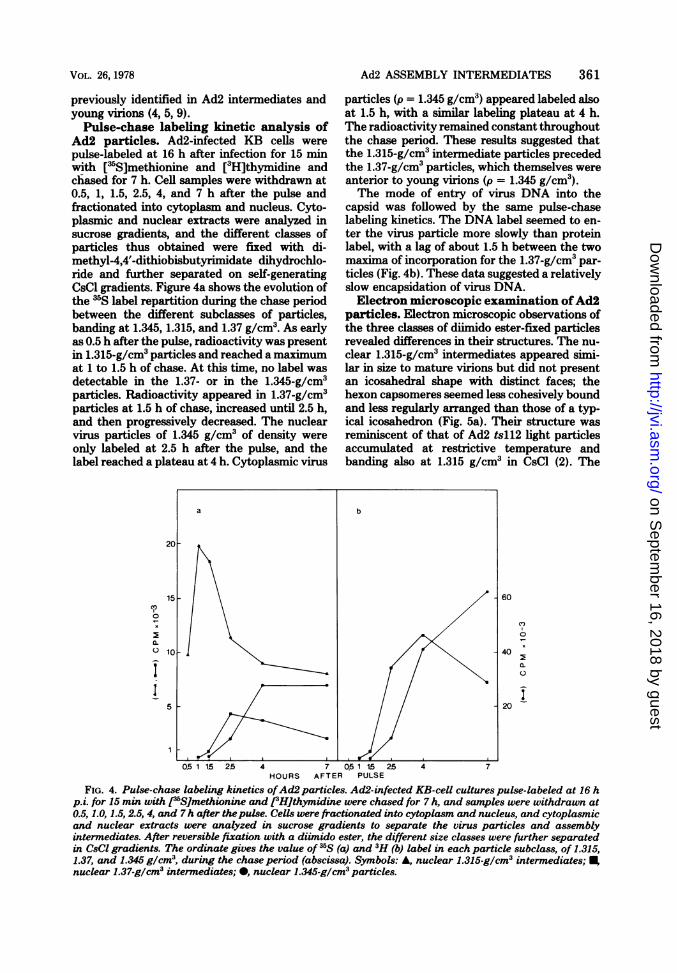
previously identified in Ad2 intermediates and particles (p = 1.345 g/cm3) appeared labeled alsoyoung virions (4, 5, 9). at 1.5 h, with a similar labeling plateau at 4 h.Pulse-chase labeling kinetic analysis of The radioactivity remained constant throughout
Ad2 particles. Ad2-infected KB cells were the chase period. These results suggested thatpulse-labeled at 16 h after infection for 15 min the 1.315-g/cm3 intermediate particles precededwith [3S]methionine and [3H]thymidine and the 1.37-g/cm3particles, which themselves werechased for 7 h. Cell samples were withdrawn at anterior to young virions (p = 1.345 g/cm3).0.5, 1, 1.5, 2.5, 4, and 7 h after the pulse and The mode of entry of virus DNA into thefractionated into cytoplasm and nucleus. Cyto- capsid was followed by the same pulse-chaseplasmic and nuclear extracts were analyzed in labeling kinetics. The DNA label seemed to en-sucrose gradients, and the different classes of ter the virus particle more slowly than proteinparticles thus obtained were fixed with di- label, with a lag of about 1.5 h between the twomethyl-4,4'-dithiobisbutyrimidate dihydrochlo- maxima of incorporation for the 1.37-g/cm3 par-ride and further separated on self-generating ticles (Fig. 4b). These data suggested a relativelyCsCl gradients. Figure 4a shows the evolution of slow encapsidation of virus DNA.the 3S label repartition during the chase period Electron microscopic examination ofAd2between the different subclasses of particles, particles. Electron microscopic observations ofbanding at 1.345, 1.315, and 1.37 g/cm3. As early the three classes of diimido ester-fixed particlesas 0.5 h after the pulse, radioactivity was present revealed differences in their structures. The nu-in 1.315-g/cm3 particles and reached a maximum clear 1.315-g/cm3 intermediates appeared simi-at 1 to 1.5 h of chase. At this time, no label was lar in size to mature virions but did not presentdetectable in the 1.37- or in the 1.345-g/cm3 an icosahedral shape with distinct faces; theparticles. Radioactivity appeared in 1.37-g/cm3 hexon capsomeres seemed less cohesively boundparticles at 1.5 h of chase, increased until 2.5 h, and less regularly arranged than those of a typ-and then progressively decreased. The nuclear ical icosahedron (Fig. 5a). Their structure wasvirus particles of 1.345 g/cm3 of density were reminiscent of that of Ad2 tsll2 light particlesonly labeled at 2.5 h after the pulse, and the accumulated at restrictive temperature andlabel reached a plateau at 4 h. Cytoplasmic virus banding also at 1.315 g/cm3 in CsCl (2). The
a b
20-
15 60C,,
0 4CL x~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~,
010- 40
5 20
0.51 15 2.5 4 7 0.51 15 25 4 7HOURS AFTER PULSE
FIG. 4. Pulse-chase labeling kinetics ofAd2 particles. Ad2-infected KB-cell cultures pulse-labeled at 16 hp.i. for 15 min with [j5SJmethionine and e:Hithymidine were chased for 7 h, and samples were withdrawn at0.5,1.0,1.5,2.5,4, and 7h after thepulse. Cells were fractionated into cytoplasm and nucleus, and cytoplasmicand nuclear extracts were analyzed in sucrose gradients to separate the virus particles and assemblyintermediates. After reversible fixation with a diimido ester, the different size classes were further separatedin CsCI gradients. The ordinate gives the value of IS (a) and 3H (b) label in each particle subclass, of 1.315,1.37, and 1.345 g/cm3, during the chase period (abscissa). Symbols: A, nuclear 1.315-g/cm3 intermediates; ,nuclear 1.37-g/cm3 intermediates; 0, nuclear 1.345-glcm3 particles.
on Septem
ber 16, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

362 D'HALLUIN ET AL. J. VIROL.
nuclear intermediates of 1.37 g/cm3 of buoyant parently, less damaging for adenovirus interme-density appeared rounder and less polyhedral in diates, as controlled by electron microscopy andshape than mature adenovirions, with vertexes sucrose or Ficoll gradients. In addition, lighthardly distinguishable (Fig. 5b). In contrast, no intermediate particles similar to those observedmorphological difference could be observed be- in wild-type Ad2 are also found accumulated intween the 1.345-g/cm3 nuclear particles and the an Ad2 temperature-sensitive mutant, tsll2, atmature virions: their icosahedral contour was restrictive temperature (2), and pulse-chase la-clearly visible (Fig. 5c). beling kinetics performed on wild-type Ad2 sug-
DISCUSSIONgest a precursor-product relationship betweenthe different classes of intermediate particles.
The question raised by the occurrence of in- All these data favor the idea that the differentcomplete particles in adenovirus infection is classes of Ad2 particles isolated here constitutewhether these particles are true preexisting in- true intermediates in the adenovirus assemblytermediates in the assembly pathway or artifac- pathway.tual particles generated by the isolation proce- As previously described (4), two size classes ofdure. Minimization of such artifacts is therefore Ad2 particles were separated by velocity gra-the major requirement of any extraction proce- dient centrifugation, one sedimenting as infec-dure for virus assembly study. Purification and tious mature or young adenovirions, at 750S,characterization of adenovirus intermediates by and the other one sedimenting as assembly in-means of CsCl density gradient centrifugation termediates, at about 600S. After reversible fix-have been shown to disrupt fragile adenovirus ation, the intermediate class could be furtherintermediates, and glutaraldehyde fixation has separated into two subclasses of particles, withbeen recommended before analysis in CsCl (4). buoyant densities of 1.315 and 1.37 g/cm3. TheUnfortunately, neither DNA nor protein com- 750S material consisted of a unique species ofposition can be analyzed on such irreversibly particles banding at 1.345 g/cm3 in CsCl. Bothfixed particles. 1.345- and 1.37-g/cm3 particles contained a DNAThis major drawback has led us to use di- with a sedimentation coefficient of 34S in alka-
methyl-4,4'-dithiobisbutyrimidate dihydrochlo- line sucrose gradients. The 1.315-g/cm3 particlesride, a diimido ester cleavable under mild reduc- contained a DNA fragment of 7-11S, whiching conditions, which has been successfully used might represent the beginning of the moleculein neighbor analysis of proteins (13) and in the entering the capsid to form the 1.37-g/cm3 inter-cross-linking of adenovirus with its cell receptor mediate and broken during isolation. In this(8). The reversibly fixed virus particles can be case, the light particles containing a piece ofisolated by density gradient centrifugation in DNA would be at a stage intermediate in pack-CsCl and characterized with regard to their aging, posterior to empty capsids.buoyant density, DNA content, and protein After reduction of the natural and artificialcomposition. disulfide bridges and SDS denaturation, theThe diimido ester fixation was carried out on three classes of particles were analyzed on SDS-
virus particles extracted from isolated nuclei polyacrylamide gels. The 1.345-g/cm3 particleswith 0.3 M (NH4)2SO4 and isolated on sucrose resembled in polypeptide composition and elec-gradients. A comparison between the conditions tron microscopic aspect the young virions al-used here and other techniques, such as sonic ready characterized (4, 9), with some residue of(4) or Freon (1) treatment of nuclei, was made precursor P VII of core protein VII. The 1.315-with respect to their effects on the integrity of g/cm3 particles totally lacked core proteins Vlabile intermediates. The ammonium sulfate and VII but contained precursors P VI and Pprocedure appeared more reproducible and, ap- VIII and two extra proteins (50K and 39K) not
a b cFIG. 5. Electron microscopy of the three classes ofAd2 particles. (a) 1.315-g/cm3 intermediates; (b) 1.37-
g/cm3 intermediates; (c) 1.345-g/cm3 virus particles. x240,000.
on Septem
ber 16, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

VOL. 26, 1978 Ad2 ASSEMBLY INTERMEDIATES 363
found in mature virions. Both 50K and 39K were Pasteur of Lille.absent from the 1.37-g/cm3 particles, which pos- We thank M. Milleville and C. Cousin for skillful andsessed a full length ofDNA without appreciable patient technical assistance and J. Croquette for secretarialaid. We are indebted to E. Kelmenson for the help with theamounts of core proteins V and P VII-VII. The English manuscript.1.37-g/cm3 particles also contained precursors PVI and P VIII. A 100K protein was visible in the LITERATURE CITEDthree classes of particles. It is unlikely that the 1. Boulanger, P. A., and F. Puvion. 1973. Large-scaleammonium sulfate extraction of nuclei had re- preparation of adenovirus soluble antigens in a highlymoved some DNA-bound proteins, since the purified form. Eur. J. Biochem. 39:37-42.2. D'Halluin, J.-C., M. Milleville, P. A. Boulanger, and72K and 45K (14) remain attached to DNA G. R. Martin. 1978. Temperature-sensitive mutant ofduring this extraction (personal observation), adenovirus type 2 blocked in virion assembly: accumu-and since adenovirus cores can be subjected to lation of light intermediate particles. J. Virol.2M NaCl without detectable loss ofDNA-bound 26:344-356.3. Doerfler, W. 1969. Nonproductive infection of baby ham-protein material (7). Moreover, this extraction ster kidney cells (BHK21) with adenovirus type 12.procedure maintains the RNA polymerases ca- Virology 38:587-606.pable of elongating nascent RNA chains on 4- Edvardsson, B., E. Everitt, H. Jornvall, L. Prage,DNA molecules (15). and L. Philipson. 1976. Intermediates in adenovirus
Pulse-chaselabelingkinetics revealed that assembly. J. Virol. 19:533-547.Pulse-chase labeling ketics revealed that 5. Everitt, E., S. A. Meador, and A. J. Levine. 1977.
1.315-g/cm3 particles preceded the 1.37-g/cm3 Synthesis and processing of the precursor to the majorspecies, which themselves preceded 1.345-g/cm3 core protein of adenovirus type 2. J. Virol. 21:199-214.particles. This suggests the following sequence 6. Green, M., and M. Pina. 1963. Biochemical studies onparticles: capsid components--..i. adenovirus multiplication. IV. Isolation, purificationof events: capsid components -> light interme- and chemical analysis of adenoviruses. Virologydiates (1.315 g/cm3; 7-1S DNA) -- heavy in- 20:199-208.termediates (1.37 g/cm3; 34S DNA) -. young 7. Harpst, J. A., J. F. Ennever, and W. C. Russell. 1977.virions (1.345 g/cm3; 34S DNA) -* mature vimi- Physical properties of nucleoprotein cores from adeno-ons(1.345 g/CM3; 34S DNA). In this hypothetical
virus type 5. Nucleic Acids Res. 4:477-490.ons1.34g/c3;35 DN). I thi hypthetcal 8. Hennache, B., and P. Boulanger. 1977. Biochemicalpathway, the 50K and 39K, which are possibly study of KB cell receptor for adenovirus. Biochem. J.scaffolding proteins, are released from the 1.315- 166:237-247.g/cm3 paticles upon entry of 34S DNA to form 9. Ishibashi, M., and J. V. Maizel, Jr. 1974. The polypep-
partiCl3 upondenr tides of adenovirus. V. Young virions, structural inter-1.37-g/cm3 particles devoid of core protein V and mediates between top components and aged virions.P VII-VII. It is possible that other intermediate Virology 57:409-424.steps take place between free capsid components 10. Laemmli, U. K. 1970. Cleavage of structural proteinsand light intermediates. during the assembly of the head of bacteriophage T4.Inantherstudy(2),we decrib a tepera 11.Nature (London) 227:680-68.
In another study (2), we describe a tempera- 11. Philipson, L., K. Lonberg-Holm, and U. Pettersson.ture-sensitive mutant of Ad2 blocked at the 1968. Virus-receptor interaction in an adenovirus sys-stage of 1.315-g/cm3 particles. Our results con- tem. J. Virol. 2:1064-1075.firm recent findings (5) and suggest that virus 12. Rosenwirth, B., S. Tjia, M. Westphal, and W. Doer-DNAenterstheadenovirus pasiclebefore core
fler. 1974. Incomplete particles of adenovirus. II. Ki-DNA enters the adenovirus particle before core netics of formation and polypeptide composition ofprotein V and core precursor P VII and that adenovirus type 2. Virology 60:431-437.DNA is not incorporated into the virion as a 13. Traut, R. R., A. Bollen, T. T. Sun, J. W. B. Hershey,core structure. It remains to be elucidated J. Sundberg, and L. R. Pierce. 1973. Methyl 4-mer-
captobutyrimidate as a cleavable cross-linking reagentwhether the 7-11S DNA fragment present in and its application to the Escherichia coli 30S ribo-the light intermediates corresponds to the same somes. Biochemistry 12:3266-3273.genome piece. 14. Van der Vliet, P. C., and A. J. Levine. 1970. DNA-
binding proteins specific for cells infected by adenovi-ACKNOWLEDGMENTS rus. Nature (London) New Biol. 246:170-174.
15. Wallace, R. D., and J. Kates. 1972. State of adenovirusThis work was supported by the Centre National de la 2 deoxyribonucleic acid in the nucleus and its mode of
Recherche Scientifique (ERA-225), the Institut National de transcription: studies with isolated viral deoxyribonu-la Sante et de la Recherche Medicale (ATP 77-82), the Univ- cleic acid-protein complexes and isolated nuclei. J. Vi-ersite du Droit et de la Sante (UER-III), and the Institut rol. 9:627-635.
on Septem
ber 16, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from