ago - greenhouse gas emissions from the australian...
TRANSCRIPT

AGO - Greenhouse gas emissions from the Australian Livestock Sector
The Australian Greenhouse Office Skip Navigation
Leading Australia's greenhouse action to achieve effective sustainable results
Greenhouse and agriculture Home > Natural resources > Agriculture > Contents
Greenhouse gas emissions from the Australian livestock sector: what do we know, what can we do?Prepared by Dr Roger Hegarty, NSW Agriculture,to contribute to the work of the Greenhouse and Agriculture Taskforce, 2001
Greenhouse Gas Emissions from the Australian Livestock Sector
What Do We Know, What Can We Do?
Contents > Next
Table of ContentsExecutive summary
1 Overview of sources of livestock emissions
2 Emissions from enteric fermentation
2.1 What information is available?
2.1.1 Emissions data
2.1.2 Detailed understanding of the process of enteric methane production
2.2 Abatement strategies for today’s Australian livestock industries
http://www.greenhouse.gov.au/agriculture/livestock/index.html (1 of 4) [5/21/2003 1:26:16 PM]

AGO - Greenhouse gas emissions from the Australian Livestock Sector
2.2.1 What features of Australian ruminant production systems will constrain the implementation of abatement strategies ?
2.2.2 Why isn’t there a clear message on nutritional management to reduce emissions?
2.2.3 What are the implications of different controllers of intake for grazing and feedlot animals on daily methane emission rates ?
2.3 Potential abatement strategies
2.3.1 Changing attributes of the animal population
2.3.2 Changing attributes of the rumen
2.4 General assessment and conclusions regarding existing information
2.4.1 Assessment of producer information
2.4.2 Assessment of scientific information
2.4.3 Conclusions about abatement through managing feed quality
2.4.4 Conclusions regarding information on other potential abatement strategies
2.5 Implications for the National Greenhouse Gas Inventory (NGGI)
2.6 Gaps in recent research activity on enteric emissions
3 Emissions from manure
3.1 Available information
3.2 Management options
4 Overview of information gaps
4.1 Information required for breeding ‘greenhouse-friendly’ ruminants
http://www.greenhouse.gov.au/agriculture/livestock/index.html (2 of 4) [5/21/2003 1:26:16 PM]

AGO - Greenhouse gas emissions from the Australian Livestock Sector
4.2 Information required to establish stable, low-methane rumen fermentations
4.3 Information required for reducing emissions through grazing management
5 Recommendations
5.1 Old things to change
5.2 New things to do
6 Glossary
7 Key personnel
8 Bibliography
Prepared by Dr. Roger Hegarty, NSW Agriculture To contribute to the work of the Greenhouse and Agriculture Taskforce
Published by the Australian Greenhouse Office, the lead Commonwealth agency on greenhouse matters.
© Commonwealth of Australia 2001
ISBN: 1 876536 69 1
This work is copyright. It may be reproduced in whole or in part for study or training purposes subject to the inclusion of an acknowledgment of the source and no commercial usage or sale. Reproduction for purposes other than those listed above requires the written permission of the Australian Greenhouse Office. Requests and inquiries concerning reproduction and rights should be addressed to:The ManagerCommunications TeamAustralian Greenhouse OfficeGPO Box 621CANBERRA ACT 2601
Copies of this document may be obtained by phoning the AGO Infoline on 1300 130 606. This document and other information on greenhouse is also available on the Internet at the following address: http://www.greenhouse.gov.au
http://www.greenhouse.gov.au/agriculture/livestock/index.html (3 of 4) [5/21/2003 1:26:16 PM]

AGO - Greenhouse gas emissions from the Australian Livestock Sector
IMPORTANT NOTICE – PLEASE READ
This document is produced for general information only and does not represent a statement of the policy of the Commonwealth of Australia or NSW Agriculture.
The Commonwealth of Australia and all persons acting for the Commonwealth in preparing this report accept no liability for the accuracy of or inferences from the material contained in this publication, or for any action as a result of any person’s or group’s interpretations, deductions, conclusions or actions in reliance on this material.
Cover photographs: © NSW Agriculture
Document designed by MA@D Communication, Canberra ACT, and printed by Canprint, Canberra ACT.
Contents > NextAGO Home | Contacts | Publications | Search | Site index | What's new | Top
Science & emissions monitoring International activities Government Business & industryCommunity & household Energy & transport Natural resources
Comments Copyright Disclaimer Notice Privacy Accessibility
http://www.greenhouse.gov.au/agriculture/livestock/index.html Last modified 17 November 2002
http://www.greenhouse.gov.au/agriculture/livestock/index.html (4 of 4) [5/21/2003 1:26:16 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Executive Summary
The Australian Greenhouse Office Skip Navigation
Leading Australia's greenhouse action to achieve effective sustainable results
Greenhouse and agriculture Home > Natural resources > Agriculture > Contents
Greenhouse gas emissions from the Australian livestock sector: what do we know, what can we do?Prepared by Dr Roger Hegarty, NSW Agriculture,to contribute to the work of the Greenhouse and Agriculture Taskforce, 2001
Executive summaryContents > Previous > Next
● Development of Australia’s capacity to understand and reduce greenhouse gas emissions from the livestock sector has not been effectively addressed in the past decade.
● Since 97% of livestock greenhouse gas emissions arise from enteric fermentation, the report focuses primarily on this source and strategies for decreasing these emissions. It does not consider sinks or net emissions associated with the sector.
● As an issue, greenhouse gas emissions have provided a negative image for the livestock sector and efforts to address it have ‘fallen through the cracks’, with no organisation committing to develop solutions to the problem in a structured way.
● Consequently, Australia has only one major abatement research project in progress, and no broadly applicable recommendations for practical abatement strategies to reduce enteric derived emissions have been put forward.
● There is a dearth of basic information on the organisms that produce methane in ruminants and this has delayed development of innovative abatement solutions.
● Despite the lack of immediate solutions, there are two distinct strategies that can be expected to deliver significant abatement and there are multiple mechanisms within each strategy by which abatement can be achieved.
● These strategies are (1) Changing attributes of the animal population and (2) the
http://www.greenhouse.gov.au/agriculture/livestock/exec.html (1 of 3) [5/21/2003 1:26:17 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Executive Summary
development of self-sustaining changes in the mixed microbial population of the rumen.
● Mechanisms for changing attributes of the animal population include selection for smaller mature size, selection for higher net-feed efficiency and selection for faster digesta kinetics.
● Mechanisms for establishing a stable, low-emission, modified rumen fermentation include: biological control directed at methanogens and associated organisms, vaccination, the establishment of effective acetogenic and bacteriocin producing populations.
● Improved pasture quality without associated reduction in grazing area and/or livestock numbers will not reduce daily methane emissions from a livestock enterprise, and needs to be considered as part of a whole farm management program.
● While manufacture of specific chemicals to inhibit enteric methane production is foreseeable, their application would be restricted by the extensive nature of the grazing industries and their threat to the ‘clean and green’ image of the Australian grazing industries is important.
● It is recommended that the impasse on knowledge and research on livestock methane emissions be overcome by creation of a jointly funded livestock emissions project team. This project would:
❍ Conduct basic research to identify and understand the organisms that produce methane in the rumen ecosystem.
❍ Increase producer understanding of the greenhouse effect, its possible consequences and the contribution of livestock.
❍ Pioneer new abatement strategies such as biological control, bacteriocins and reductive acetogenesis.
❍ Draw on overseas expertise as appropriate.
● A core program activity would be the establishment of ‘model farms’ for each livestock enterprise to determine the environmental footprint for each enterprise. This component of the project should be broader than assessment of greenhouse gas emissions alone, and would require drawing in other relevant organisations to address energy, water and nutrient fluxes of the livestock systems to ensure a
http://www.greenhouse.gov.au/agriculture/livestock/exec.html (2 of 3) [5/21/2003 1:26:17 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Executive Summary
broad and thorough assessment of the environmental credentials of the Australian livestock industries.
● A structured program as proposed is considered essential to achieve the breadth of investigation and effective delivery that is required to contain or reduce emissions in the coming decade.
Contents > Previous > NextAGO Home | Contacts | Publications | Search | Site index | What's new | Top
Science & emissions monitoring International activities Government Business & industryCommunity & household Energy & transport Natural resources
Comments Copyright Disclaimer Notice Privacy Accessibility
http://www.greenhouse.gov.au/agriculture/livestock/exec.html Last modified 17 November 2002
http://www.greenhouse.gov.au/agriculture/livestock/exec.html (3 of 3) [5/21/2003 1:26:17 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 1
The Australian Greenhouse Office Skip Navigation
Leading Australia's greenhouse action to achieve effective sustainable results
Greenhouse and agriculture Home > Natural resources > Agriculture > Contents
Greenhouse gas emissions from the Australian livestock sector: what do we know, what can we do?Prepared by Dr Roger Hegarty, NSW Agriculture,to contribute to the work of the Greenhouse and Agriculture Taskforce, 2001
1. Overview of sources of livestock emissionsContents > Previous > Next
The Australian livestock subsector released 62.6 Mt CO2-equivalents in 1999, making it the nation’s largest source of agricultural greenhouse gas emission. The principle greenhouse gas arising from agricultural livestock management is methane (99%) with a small quantity (1%) of nitrous oxide (N2O) being released from degradation of nitrogenous compounds in faeces and urine (Figure 1). The contribution of N2O is small relative to some developed countries and reflects differences in both manure management and inventory procedures for the Australian livestock industries. Extensive ruminant enterprises are the principle sources of livestock manure but emissions from these wastes are not credited to livestock in the national inventory. Only greenhouse gas emissions from the managed manures and effluent derived from the intensive livestock industries are included in the manure management portion of the livestock emissions inventory. Methane attributed to livestock is liberated from the anaerobic microbial fermentation of feedstuffs in the gut (97%) and faeces (3%) of livestock by methanogenic organisms (methanogens). While small amounts of methane are liberated from microbial digestion occurring in the digestive tract of pigs, these and other simple stomached livestock sources contribute only 0.2 % of the enteric (digestive tract) emissions from Australian livestock (Figure 2). It is ruminant animals that are the single largest source of Australia’s agricultural greenhouse gas emissions (2861Gg CO2 -e) and alone contribute 13.1% of Australia’s total national emissions (NGGI 1999).
http://www.greenhouse.gov.au/agriculture/livestock/chapter1.html (1 of 2) [5/21/2003 1:26:18 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 1
Figure 1. Contributions of enteric fermentation and manure management to the total methane emission from the livestock sector (NGGI 1999).
Figure 2. Relative importance of ruminant and non-ruminant (monogastric) animals to the total enteric methane emissions form the Australian livestock sector (NGGI 1999).
Contents > Previous > NextAGO Home | Contacts | Publications | Search | Site index | What's new | Top
Science & emissions monitoring International activities Government Business & industryCommunity & household Energy & transport Natural resources
Comments Copyright Disclaimer Notice Privacy Accessibility
http://www.greenhouse.gov.au/agriculture/livestock/chapter1.html Last modified 17 November 2002
http://www.greenhouse.gov.au/agriculture/livestock/chapter1.html (2 of 2) [5/21/2003 1:26:18 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
The Australian Greenhouse Office Skip Navigation
Leading Australia's greenhouse action to achieve effective sustainable results
Greenhouse and agriculture Home > Natural resources > Agriculture > Contents
Greenhouse gas emissions from the Australian livestock sector: what do we know, what can we do?Prepared by Dr Roger Hegarty, NSW Agriculture,to contribute to the work of the Greenhouse and Agriculture Taskforce, 2001
2. Emissions from enteric fermentationContents > Previous > Next
2.1 What information is available?
2.1.1 Emissions data
The National Greenhouse Gas Inventory (NGGI, 1999) with methodology supplements is available on-line at (http://www.greenhouse.gov.au/inventory/1999/index.html) and includes detailed breakdown tables of the sector and subsector contributions of livestock. The inventory can also be obtained free of charge on request from the Australian Greenhouse Office (telephone 1300-130606). Procedures used in deriving the inventory are routinely revised but the fundamental biological controllers of total ruminant methane emissions are:
● Number of animals of each species and class ● Feed intake, being estimated from liveweight, and in some cases from feed
availability.
Equations are then used to estimate the proportion of dietary energy lost as methane (NGGI 1998).
2.1.2 Detailed understanding of the process of enteric methane production
Organisms: The advent of molecular probes to differentiate between distinct microbial populations has indicated a previously unsuspected diversity in methanogenic Archaea (methanogens) within the rumen. Gene sequencing (Whitford et al., 1997) indicates
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (1 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
that only some 10% of existing rumen organisms may have been cultured, and this is in keeping with the viable count of rumen organisms being substantially less than a total cell count determined microscopically. There is no reason to think rumen methanogens will be any less genetically diverse than the rumen bacteria as a whole. Until recent years it was thought only three genera of methanogens were present in the rumen (Methanobrevibacter, Methanomicrobium and Methanosarcina). Since the total known genera of methanogenic Archaea known has increased from seven to twenty-two (Wright and Baker 1999), it seems likely that more genera and especially more subspecies of rumen methanogens will be identified as 16S-rRNA analysis investigations proceed.
Direct biological control agents, including vaccines are very likely to exhibit high host-specificity, meaning they will be effective on some methanogens but not others. Commercial development of a vaccine, or pathogens for methanogens without knowing the exact identity of the target organism would be ineffective. The lack of information on the predominant species and types of rumen methanogens in Australian ruminants places a serious constraint on developing biological approaches to emissions control.
The lack of information on the predominant species and types of rumen methanogens in Australian ruminants places a serious constraint on developing biological approaches to emissions control.
Biochemistry: The biochemical pathways (and therefore the avenues for chemical intervention) in methanogenesis are well defined and easily accessed, being found in microbial biochemistry textbooks (eg. Gottschalk 1986). In particular the enzyme systems which enable methanogens to generate adenosine triphosphate (ATP) from carbon dioxide and hydrogen have been intensively studied. It is considered that information on biochemical pathways for methanogenesis is not currently limiting development of emission reduction procedures or products. Improved understanding of the coat chemistry of methanogens, particularly the glyco-protein S-layers may be advantageous to vaccine development or designing methanogen specific antibiotics. This information will best be obtained in association with the isolation (if possible), and identification of the dominant methanogen species in the rumens of Australian livestock as highlighted previously.
Ecological inter-relationships: As plugging up a car exhaust pipe to reduce exhaust fumes causes the car to stop, so poisoning or otherwise eliminating rumen methanogens will seriously reduce the performance of the rumen and of the animal. The process of methanogenesis serves as the exhaust system of rumen fermentation in which the <1% of rumen organisms which produce methane, eliminate hydrogen gas which would otherwise inhibit the activity of the bulk of other organisms that do not
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (2 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
produce methane.
Hydrogen has been referred to as the "currency" of rumen fermentation, in recognition of the fact that transfer of hydrogen between organisms and eventual disposal is fundamental to freeing-up the primary fermenting organisms to ferment fresh feed. Hydrogen concentration affects not only the process of fermentation, but also the spatial location and associations of organisms, including methanogens. The principle sources of hydrogen are those organisms (bacteria and protozoa) which produce acetic acid (Hegarty & Gerdes 1999). It is not surprising then, that ruminants maintaining a high protozoal population or those consuming an acetate-producing feed, generally have the highest methane emission rates.
Elimination of the rumen methanogen populations without establisheing alternate hydrogen users will cause the accumulation of hydrogen and have catastrophic effects on inter-related organisms, stopping fermentation and causing livestock to stop eating.
While one study has reported 37% of rumen methanogenesis is due to methanogens living in or on rumen protozoa (Finlay et al., 1994), there has been no comprehensive assessment of the species of protozoa nor methanogens involved. This is in spite of protozoa-free ruminants having consistently lower methane emission than normal (faunated) ruminants (Kreuzer 1986). Consideration of methane production within the context of other organisms which either provide hydrogen to methanogens, or compete for hydrogen with methanogens, is likely to significantly increase the array of target organisms to consider, and should improve the stability of emissions reduction achieved. Studies of the ecology of methanogens in the rumen are only now possible since the development of molecular probes for individual Archaeal, bacterial and protozoal species.
The current understanding of the geographical and chemical associations between methanogens and other rumen organisms is inadequate to allow reliable indirect reduction of emissions through control or elimination of other organisms. Significant new options for reducing emissions are likely to be developed by improving understanding of the associations between methanogens and other rumen organisms. This understanding will permit control of the associated organisms and establishment of a new microbial equilibrium. Specific opportunities for ecological control by rebalancing the rumen microbial population are discussed in section 2.3.
The current understanding of the geographical and chemical associations between methanogens and other rumen organisms is inadequate to allow reliable indirect reduction of emissions through control or elimination of other organisms.
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (3 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
2.2 Abatement strategies for today’s Australian livestock industries
There is an abundance of reviews on rumen methanogenesis and particularly dietary and chemical factors that affect it (references provided at start of bibliography). For all the volume of scientific reporting, there are still extremely few recipes or recommendations which livestock managers can immediately implement to reduce the annual livestock emissions from a farm. More importantly, there is some encouragement to adopt allegedly methane-reducing feeding practices that may in fact increase rather than decrease total annual emissions from a farm. The reasons for the paucity of existing recommendations will be explored in the following two sections.
2.2.1 What features of Australian ruminant production systems will constrain the implementation of abatement strategies ?
The Australian sheep and cattle industries are characterised by features which largely constrain (a) the scale of livestock emissions, (b) the approaches that should work in principle and (c) the practicality of these approaches. It is appropriate to table these constraints before considering why Australia has until now failed to deliver either take-home management messages or technological solutions for abatement of livestock methane emissions.
A pasture based industry
The Australian sheep industry is almost exclusively reliant upon pasture. Far less than 10% of crossbred lambs are feedlot finished, and this only for the final 6 week period in their lives of 7-11 months. Few Merino lambs and only approximately 13,000 superfine (Sharlea) Merino sheep are supplemented for any purpose other than management during pregnancy or survival during drought.
The cattle industry is also pasture based, with all the breeding herd (14.8 million head) consuming predominantly pasture, but up to 39% of slaughter stock pass through a feedlot nationally, for between 70 and 300 days of their life (Graham 2001).
Pastures that are seasonal and predominantly of low quality
The significance of strongly pasture-based breeding and growing sectors for ruminant livestock is that the average energy density of feeds consumed is low. The weighted average digestibility of Australian sheep and cattle pastures are 68 and 63% respectively (Table 1; after NGGI 1994). These digestibilities translate into energy densities of 9.1 and 8.3 MJ ME/kg DM respectively (SCA 1990; eq 1.13b), where ME is the abbreviation for metabolisable energy. These are substantially lower digestibilities
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (4 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
than occur in the concentrate-based diets used to feed ruminants in more heavily populated developed countries and will affect the appropriateness of nutritional manipulation in Australia versus these countries.
Digestibility of Australian pasture diets is substantially lower than for the concentrate-based diets used to feed ruminants in heavily populated developed countries. This will affect the appropriateness of nutritional manipulation.
Table 1. Average seasonal digestibility (% DM) of pasture consumed by Australian sheep and cattle. Values presented are weighted according to the feed digestibility in each state in each season and the proportion of the national herd/flock consuming in that state in that season (after NGGI 1994; tables 1.1b, 1.4 .1b, 4.3)
Spring Summer Autumn Winter Mean
Sheep 74 60 66 72 68
Cattle 63 62 64 62 63
Minimal labour input
The large extensive cattle and sheep properties which characterise the semi-arid, tropical and sub-tropical pastoral zones of Australia present a major challenge to emissions abatement. Livestock production on these properties is typically constrained by inadequate feed availability and low feed digestibility. More importantly, livestock are handled as few times a year as possible. The implication of this is that to be practical, abatement strategies must be long-acting, preferably being effective for at least 6 months from application.
Graziers in the low or extremely seasonal rainfall zones are also limited in their ability to use grazing management to control the digestibility of feed on offer to the animal. The climatic constraint to pasture growth has associated effects which also impact on the farm’s own methane emissions budget, such as causing low reproductive efficiency and slow growth rates. These lead to a high ‘unit-cost’ of production (methane/kg product sold) compared to more intensive enterprises.
The infrequent handling of animals on extensive properties necessitates that any abatement strategy (be it directed at the feed or the animal), be effective for at least 6 months from a single application
There is the possibility of regular effective delivery of chemical or biological agents to stock in the extensive pastoral zone through water medication, if access to water is
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (5 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
restricted to a reticulated supply. While this opportunity exists, there are as yet no suitable chemical or biological control agents to deliver for this purpose.
2.2.2 Why isn’t there a clear message on nutritional management to reduce emissions?
Historical
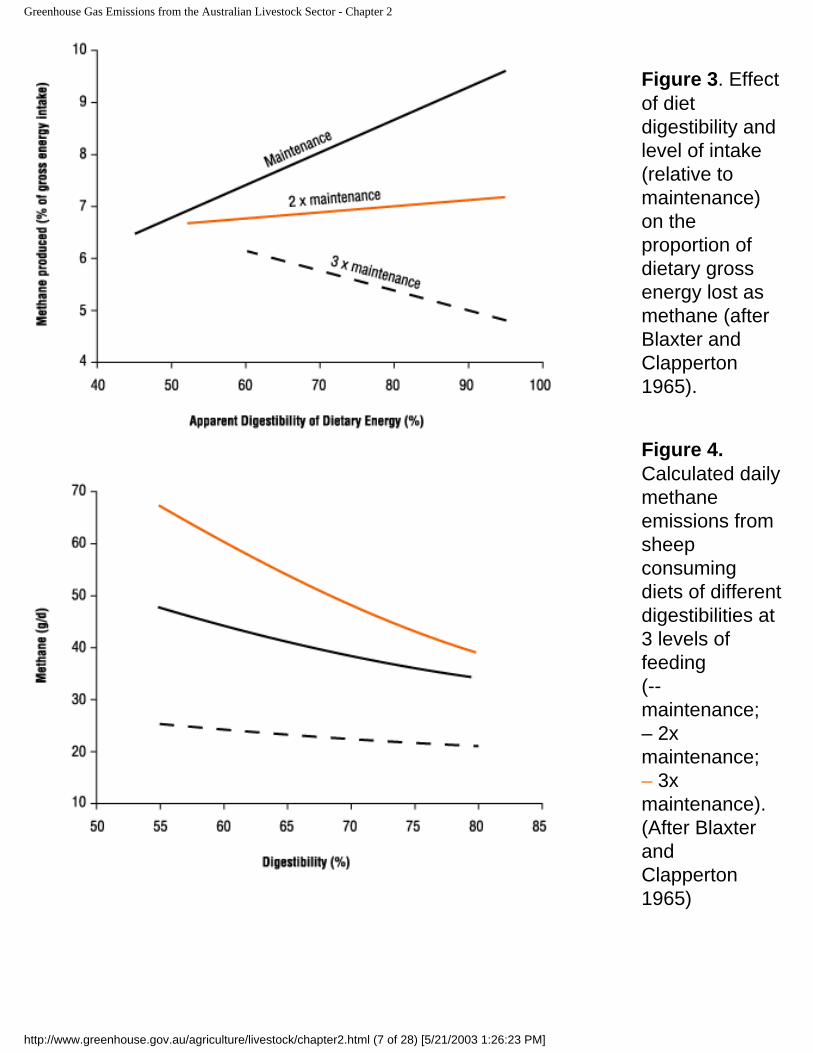
In order to understand the complexity and possible confusion in this area, it is beneficial to revisit history. In 1965, Blaxter and Clapperton published a then benchmark review of dietary effects on methane emissions. At the time, the principal research focus was on understanding energy partitioning by ruminants, and methane was merely one component in the energy budget. With this focus, the authors reported emissions as the "% of gross dietary energy" lost as methane and generated the memorable graph (Figure 3) showing the proportion of dietary energy lost as methane decreases as digestibility increases, once animals are consuming high levels of feed (eg. Three times the amount simply required to maintain their liveweight). Further calculations show that providing a fixed level of energy through a high digestibility diet will indeed cause a slight reduction in the quantity (g/d) of methane emitted per head (Figure 4). This message can easily be misconstrued to mean that simply improving the intake and digestibility of pastures will reduce daily methane emissions.
For grazing animals however, pasture quality and intake are innately linked in a positive association (Figure 6). Blaxter and Clapperton’s models completely ignore the relationship between feed digestibility and feed intake which naturally exists for most forage fed ruminants. It requires knowledge of both digestibility and diet intake to predict methane emissions using Blaxter and Clapperton’s methodology. What these authors conclude about the emissions from ruminants eating a set amount of feed of specified digestibility is largely correct, (although other reviewers conclude their findings are skewed (Benchaar et al., 1998; Pelchen & Peters 1998). However their failure to account for digestibility/intake relationships in forages means this relationship must be sought elsewhere in order to predict daily emissions of free ranging ruminants. It is the association between feed characteristics and feed intake that is pivotal to the effects of general nutrition on methane emissions. In appreciation of this critical association, the effects of feed characteristics on feed intake are considered below, with diets being partitioned into pasture and feedlot types to highlight differences.
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (6 of 28) [5/21/2003 1:26:23 PM]
Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
Figure 3. Effect of diet digestibility and level of intake (relative to maintenance) on the proportion of dietary gross energy lost as methane (after Blaxter and Clapperton 1965).
Figure 4. Calculated daily methane emissions from sheep consuming diets of different digestibilities at 3 levels of feeding (-- maintenance; – 2x maintenance; – 3x maintenance). (After Blaxter and Clapperton 1965)
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (7 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
Figure 5. Effects of dietary energy density (MJ ME/ kg DM) on daily methane emission (g/d) – and feed intake (g/d) – of sheep (after Pelchen and Peters 1998).
Diet effects on voluntary feed intake
The regulation of feed intake by ruminants has been exhaustively researched and Australia boasts world authorities in this area, but capturing all the possible regulators of feed intake in a simple model has proved difficult. There are clearly feed-factors (eg. bulk and resistance to digestion) which limit intake, as well as animal factors (eg. weight and maturity), and factors that involve both (eg. palatability, anti-nutritive factors). By simply partitioning feeds into two types (pasture and feedlot diets) it is possible to describe nutritional effects on methane production in an applied manner. This delineation is partly artificial as diets can span a continuum from 100% roughage to 100% grain but it is a functional division selected for the following reasons and fits with the differing methodologies used in the Australian NGGI for grazing and feedlot cattle.
Discussion of the responses to management of pasture and feedlot diets differently is appropriate, considering the findings of Pelchen and Peters (1998). In their review these authors identified the effect of increasing digestibility (or energy density) of the diet on daily methane emissions changes from being positive to negative at approximately 11 MJ ME/ kg DM (Figure 5). Further reasons to place an arbitrary partition between pasture and feedlot diets when discussing dietary effects on methane emissions are outlined below.
● Pastures generally have a low to moderate digestibility with Australian sheep and
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (8 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
cattle pastures used in the NGGI (1994) having DM digestibilities of 68 and 63% respectively.
● Pastures provide long roughage; physical breakdown of this roughage in the gut is reliant upon both chewing and fermentation in order to pass from the rumen.
● In contrast, feedlot diets have the highest rate of digestion that is considered ‘safe’ for the animal. The feed is pre-processed, with grain often treated to increase digestion and the roughages chopped to facilitate rapid passage from the rumen.
These diet characteristics mean that the capacity of forage-fed ruminants to eat is limited by the size of the rumen and by the rate at which feed is cleared from the rumen. It is said that ‘rumen-fill’ limits intake of forage fed (eg. grazing) ruminants. On forages, ruminants can only eat more if something changes to increase the rate at which feed is cleared from the rumen. Increasing digestibility is the simplest way of increasing intake but more subtle (harder to measure) changes such as increasing the rate of digestion or the rate of comminution (breakdown by chewing) can have the same effect of increasing intake.
In contrast to forages consumed by grazing animals, the feedlot diet offers minimal resistance to the rumen microbes and requires little chewing before passing from the rumen to the intestines. Consequently, intake of the feedlot diet is unlikely to be limited by ‘rumen-fill’. Rather, the intake by lot-fed animals is constrained by the animal’s nutrient requirement, not the properties of the feed it consumes. This nutrient requirement is principally a function of the animal’s liveweight and state of maturity. Providing a diet of higher energy content than the 12 MJ/kg DM typically in feedlot diets is unlikely to lead to an increase in dietary dry matter or energy intake by ruminants.
2.2.3 What are the implications of different controllers of intake for grazing and feedlot animals on daily methane emission rates?
Forage fed ruminants
It is assumed from the previous argument that the quantity of energy consumed by forage fed ruminants of a given size and maturity is limited by the rate of clearance of feed from the rumen (by fermentation or passage). Intake may also be limited by the availability of feed, and increases as feed availability increases as well as forage digestibility (Figures 6 and 7).
Consider then the impact of improving pastures on the productivity and methane emissions from a 30 kg large framed Merino wether (Table 2). Simulations were conducted in GrazFeed (v 4.01) which is a feed management package based on the
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (9 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
Australian feeding standards for ruminants (SCA 1990). GrazFeed uses Blaxter and Clapperton’s (1965) equation to predict daily methane emission rate, which accounts for diet digestibility and level of intake.
Figure 6. Positive relationship between digestibility of pastures and the quantity voluntarily consumed by sheep (Freer and Jones 1984).
Figure 7. Effect of pasture availability (t DM/ha) on the voluntary feed intake of sheep (SCA 1990).
The simulations show that the proportion of energy ingested which is lost as methane declines from 6.6 to 6.0 % as feed of the higher digestibilities is provided. However, the higher digestibility pasture is more rapidly fermented and cleared from the rumen, enabling the animal to eat more of the ‘improved’ pastures. By providing unlimited access to a pasture of 65 or 75% digestibility compared to the unimproved pasture at
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (10 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
55% digestibility, feed intake has increased from 1.02 to 1.42 then 1.74 kg/head/d. Importantly, the methane emission rate (g/head/d) has increased not decreased due to providing pasture of improved digestibility.
Table 2. Feed intake, methane emission and growth rate of a 30 kg Merino wether when given free access to forages differing in digestibility but of equal protein content (15% CP). Simulations based on GrazFeed version 4.1.
Sheep parameter Digestibility
55% 65% 75%
Dry matter intake (kg DM/d) 1.09 1.42 1.74
Gross energy intake (MJ GE/d) 20.1 26.1 32.0
Digestible energy intake (MJ DE/d) 11.0 17.0 24.0
Methane emission (g methane/d) 24.5 32.4 35.4
Methane emission (% of Gross Energy) 6.6 6.7 6.0
Methane emission (% of Digested En.) 12.0 10.3 8.0
Growth Rate (g liveweight/d) 5 57 162
Grams methane emitted/g liveweight gained 4.9 0.6 0.2
Improved digestibility of pasture has however, caused a surge in animal performance, with the wether growing at up to 162 g/d instead of 5g/d. On the native (55% digestible) pasture growth the unit cost of liveweight gain was 4.9 g of methane per gram of gain whereas on the 75% digestible pasture each gram of gain was achieved for the emission of only 0.2 g of methane. Considering growth of the lamb was so slow on the unimproved pasture, it will also clearly require a lot longer to reach slaughter weight on this pasture. Consequently, the lifetime methane emission will be much higher for
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (11 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
animals grown to slaughter weights on low digestibility feeds. In previous simulations of lamb growth, it was found that to gain 10 kg of lamb liveweight on 65% digestible pastures would lead to emission of approximately 8 kg of methane, while achieving the same 10kg liveweight gain of lambs on 75% digestible pasture would result in less than 2 kg of methane being emitted (Hegarty 1998).
Providing higher digestibility forages to ruminants that are free to eat as much as they please, will increase rather than decrease daily methane emissions as a consequence of animals eating more of the higher digestibility forage
Is this a clear-cut demonstration that improving pasture quality must inevitably increase national ruminant methane emissions ?
No. In the above simulation we have only affected pasture digestibility, without making any other management decisions. The grazier has the opportunity to change the number of animals, the area they graze and the time they graze to achieve the desired animal production but with less methane released.
If livestock managers desire a specified quantity of product output (eg. t meat/year) this can be produced with less total feed eaten and less total methane produced on improved pasture but on the following conditions:
● The need to destock when the production goal is reached. ● The need to not graze (therefore waste) the unused feed for the remainder of the
year.
These constraints would be considered unrealistic by many graziers because (a) pasture improvement is very expensive and there is a need to sell more product to pay for it and (b) profitability often comes from rapid turn-over of stock (eg. finishing store animals). The only practical strategy to achieve a targeted animal production output by improved pasture quality concomitant with a reduced annual methane emission is to reduce the land area under grazing. For example, to achieve the desired production from 500 ha of improved pasture instead of on 1500 ha of unimproved pasture. Developing alternate uses for the excess (formerly grazed) land would require careful consideration.
The greater animal performance achieved on improved pastures can allow the same amount of animal product to be generated in less time and with production of less methane than if the same animal product was generated on low digestibility pastures.
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (12 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
The challenge is clearly how to avoid the possibility of running more stock and producing more product once the expense of improved pasture has been incurred.
Lot-fed ruminants
At the start of this section it was assumed that feed characteristics (digestibility, rate of digestion or DM availability) have little effect on the voluntary feed intake of lot-fed ruminants consuming diets high in cereal grains. In contrast to the forages where the extent of rumen-fill almost always constrains feed intake, the quantity of feed energy consumed by a feedlot animal is largely determined by its own energy requirements to maintain current body mass and grow its tissues at their genetic potential for growth. (This general statement has many caveats and it is recognised that climatic conditions, dust, palatability, water cleanliness health and environmental conditions all impact the potential performance and therefore energy requirement of feedlot animals). If intake is unaffected by digestibility on these diets, then what are the effects of feed characteristics on methane emissions from lot-fed ruminants?
A feedlot animal will therefore require less of a diet providing 13 MJ ME/kg DM relative to a diet providing only 12 MJ ME/kg DM. So providing diets of exceptionally high energy content will reduce daily dry matter intake in feedlots. Not only intake, but daily emissions of methane will also be reduced as feedlot diets of higher energy content are provided, in keeping with sheep data reviewed by Pelchen and Peters (1998; Figure 5). There are a number of reasons why feedlot diets with extremely high starch contents cause low enteric methane emission rates and these principally relate to the fermentation pattern and organisms in the lot-fed rumen as outlined below.
Starch is readily fermented to propionate by rumen microbes and the production of propionate uses hydrogen. Hydrogen is also required in the production of methane so if some hydrogen is used for propionate production it is not available for methane production. Correspondingly, the major source of hydrogen in the rumen is fermentation to acetate, and production of acetate is relatively less important in starch fermentation than in forage fermentation. As a result, the balance between hydrogen producing reactions (acetate production) and hydrogen using reactions (propionate production) leads to a net reduction in hydrogen availability for the methanogens in the rumen of feedlot compared to pasture finished ruminants.
Methanogens and methane production are also inhibited by acidic conditions and the rumen of animals consuming a high-starch feedlot diet is typically more acidic than that of forage fed ruminants. This effect of acidity is considered further in section 2.3.2.1. There may well be other as yet unidentified inhibitors of methanogenesis in the rumen
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (13 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
of feedlot animals. Many lactic acid bacteria also naturally produce antibiotic-like substances called bacteriocins (section 2.3.2.5) and these may possibly also contribute to the effect of higher energy feedlot diets lowering the daily enteric methane release.
In practice, there is limited practical scope for increasing the energy content of feedlot diets to reduce daily emissions using cereal grains due to the increased risk of rumen acidosis (a nutritional disease) and absence of higher energy feedstuffs. Inclusion of selected oils provides a safer way of increasing the energy content of feedlot diets which could reasonably be expected to further reduce daily methane emissions (section 2.3.2.1).
2.3 Potential abatement strategies
2.3.1 Changing attributes of the animal population
Animal number
Since the total emission of methane from Australian livestock is fundamentally calculated from the number of animals and their emissions/head/day, reducing the total number of livestock offers a direct and immediate mechanism to reduce industry emissions. The effectiveness of this strategy is apparent in the Australian sheep industry where a reduction in sheep numbers from 175 million in 1990 to a present population of 116 million, has been associated with a reduction in emissions from 1170 to 776 Gg/annum. It is, however, unlikely to find favour with cattle graziers except in association with medium term economic cycles. Although native animals may provide new enterprises in marginal areas (http://www.amonline.net.au) there are few if any realistic, financially viable, land-use alternatives than continued grazing of cattle for these semi-arid areas.
Animal size
The metabolisable energy requirements of animals increase with increasing liveweight, so use of animals with smaller mature liveweight will mean less feed is required to maintain the herd/flock and it can be assumed, less methane emissions will be produced per head per day. Reducing mature size however also brings associated changes such as slower growth of purebreds, animals with higher fat content at lower body weights and potentially more calving or lambing difficulties. Most importantly, the effectiveness of this strategy is dependent upon graziers not increasing stock numbers to ensure full pasture utilisation. After all, the stocking rate of sheep is normally higher than that of cattle on the same pasture simply because more small animals can be supported on the available feed than can big animals. If stock numbers increases as
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (14 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
animal size decreases, then the total methane release will increase.
Feed-use efficiency
Australian and overseas research has identified that within populations of cattle and pigs there are some individuals that consume more feed than expected to achieve a specified rate of growth (low net feed efficiency), while others achieve this growth while consuming much less feed than the population average (high net feed efficiency). This genetically determined variation provides an opportunity to reduce feed consumption (and assumedly daily methane emission) without altering animal size or growth performance. Commercial test facilities to determine the net feed efficiency (NFE) of leading industry sires are currently being established in southern Australia and estimated breeding values for this trait in industry sires are just becoming available. At present there is little information or research to determine what physiological differences exist between high and low feed net feed efficiency animals, so it is not possible to state conclusively that high net feed efficiency animals will produce less methane than average efficiency animals.
Selection of ruminants with a high net feed efficiency offers an opportunity to reduce daily methane emissions without reducing livestock numbers or reducing product output or quality.
In contrast to selection for net feed efficiency, simple selection of animals for feed efficiency (g feed/g of gain) or feed conversion ratio (FCR; g gain/g of feed), will have the undesirable adverse effect of increasing feed intake and presumably methane emission. (Genetic correlation between FCR and feed intake in cattle = 0.29; Mrode et al., 1990).
Differences in digestive physiology
Individual ruminants with very low methane emissions have been identified in both Australia and New Zealand, and may contain differences in either gut physiology or gut microbiology which could have application across the broader population. In particular, the mean retention time (or rate of passage) of material from the rumen dramatically affects methane production with faster passage greatly reducing daily methane emissions (Okine et al., 1989). Differences between sheep in the mean retention time of digesta are known to occur and mean retention time of digesta is a highly repeatable trait in sheep (Smuts et al., 1995; repeatability = 0.45-0.6). There is therefore strong opportunity for genetic selection of superior animals for this trait which may be expected to produce less methane.
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (15 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
While not an instant solution, there is opportunity to reduce total emissions from the livestock sector by either reducing the livestock population or changing the genetic base of the animals. Genetic changes which would be beneficial are 3 fold and include moving to smaller mature-weight animals, animals of higher net feed efficiency, and those whose digesta kinetics and/or microbial population leads to lower daily methane emissions.
2.3.2 Changing attributes of the rumen
Novel new approaches to alter the microbial ecology of the rumen for the purpose of reducing daily methane production, without requiring a decline in feed intake, are described in this section. Strategies are grouped for discussion by their primary mode of action but the divisions are descriptive not absolute. It is recognised that manipulating any rumen organism will induce an ecological shift in the remaining population. Extensive reviews of the chemical inhibition of methanogens are available. In one such review, Van Nevel and Demeyer (1996) conclude that they "are rather pessimistic concerning decreasing methane emissions by the use of additives". In recognition of this, and the extensive reviews by other authors, only a brief coverage of chemical control agents is provided below.
As mentioned earlier, a caution in reducing ruminal methane production is that methanogenesis is normally central to continued fermentative digestion of feed. The ruminant depends on fermentation for other microbial products on which the animal itself lives, such as microbial protein and volatile fatty acids. Methane producing organisms normally remove hydrogen gas from the rumen by converting it to methane. Hydrogen is a waste product of fermentation and if hydrogen accumulates, rumen digestion is suppressed (Wolin et al., 1997). The goal in addressing livestock methane emissions then, is not simply stopping methane emissions, but rather, redirecting hydrogen into more beneficial pathways.
The goal in addressing livestock methane emissions is not simply stopping methane emissions, but rather, redirecting rumen hydrogen into more beneficial end products.
2.3.2.1 Compounds directly toxic to methanogens
Analogues and inhibitors
Following the discovery that simple halogenated methane analogues inhibited ruminal methane production (Bauchop 1967), the efficacy of an array of targeted inhibitors was evaluated through the 1970s (Czerkawski and Breckenridge 1975). Most of these
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (16 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
inhibitors have been found to suffer reduced efficacy after sustained or repeated administration (Czerkawski 1986; Van Nevel and Demeyer 1995). Recent studies to extend the period of effectiveness of bromochloromethane acid in the rumen by complexing to an cyclodextrin matrix have shown promise in reducing total emissions without reducing live weight gain (McCrabb et al., 1997).
There are two challenges that necessitate careful thought before proceeding with further development of specific chemicals for use as feed additives to reduce livestock methane emissions. The first is accessibility of animals. While ever-increasing proportions of beef steers are feedlot-finished, up to 89% of the feed consumed by cattle is consumed by the maternal herd (Thompson and Barlow 1986; Faichney 1995). In extensive herds, regular feeding or treatment of animals to reduce methane emissions will be impractical especially when water supply is not reticulated. Secondly, there is growing awareness of residues as a key aspect of the safety of livestock products (O’Keeffe and Kennedy 1998). Voluntary inclusion of ‘designer’ anti-microbials in slaughter or milk-animals for reasons other than disease control may jeopardise current consumer confidence in the safety of livestock products.
Dietary fats
Unsaponified or otherwise unprotected fats or oils may have a general suppressing effect on rumen fermentation when included at 7% or above in most diets (Kowalczyk et al., 1977). Specific suppressive effects of oils on methane production were first observed by Blaxter and Czerkawski (1966) and the toxic effects of fatty acids on methanogens were confirmed by Henderson (1973). More recently, comparison of the efficacy of a range of oils has identified coconut oil as being most effective (Dong et al., 1997; Machmuller et al., 1998; Machmuller and Kreuzer 1999). It is the medium chain fatty acids (C10 – C21), which cause the greatest reduction in methane production (Dohme et al., 2000) and also the methanogen population (Dong et al., 1997). A reduced effect of medium chain fatty acids on high-fibre content diets (Dong et al., 1997; Dohme 2000) indicates the practical application of this may be limited to feedlots.
Hydrogen ion concentration
The lower proportion of gross energy lost as methane on concentrate diets relative to roughage diets is widely reported (Blaxter and Clapperton 1965) and is typically associated with a lower rumen pH in concentrate-fed ruminants. These facts are consistent with methanogenesis being inhibited at low pH in-vitro (Slyter 1979) and the requirement for all cells to maintain an electrochemical gradient to survive. Russell and Wilson (1996) demonstrated that the ability to lower intracellular pH is a major
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (17 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
influence on whether microbes may grow at low pH. Although intracellular pH has not been studied in methanogenic Archaea, Lana et al., (1998) demonstrated a near linear decline in methanogenesis by rumen fluid when cultured in media of pH ranging from 6.5 down to 5.7 (Figure 8). Reduced methanogenesis down to pH 5.7 may simply be a stoichiometric consequence of increased propionate production, with these authors explaining 78% of variation in methane production by the in-vivo acetate:propionate ratio, and only 50% by the in-vivo pH. At pH below 5.3 however, hydrogen gas accumulation may occur (Russell 1998) being evidence of a direct acid toxicity for methanogens. High hydrogen ion concentration may therefore partly explain the decline in methane emission of ruminants fed diets of energy densities exceeding 10.5 MJ/kg DM (Pelchen and Peters 1998). It is also possible that associated changes in acidic conditions, such as bacteriocin production, may be important and this is discussed subsequently.
Figure 8. Effect of pH on methanogenesis in mixed rumen fluid bacteria obtained from cattle fed hay ( ) or 90% concentrate rations ( ). All bacteria were isolated by centrifugation then resuspended in a common clarified rumen fluid (Lana et al., 1998).
Monensin
Early reports of monensin reducing methane production from livestock were attributed to (1) a reduction in voluntary feed intake causing reduced fermentation (Goodrich et al., 1984); (2) selectively reducing acetate (and therefore H2) production (Slyter 1979) and (3); inhibiting the release of H2 from formate (Van Nevel and Demeyer 1977). The methane suppressing effect of monensin is typically not maintained for long periods.
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (18 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
While its action has been shown to continue for 35 days in sheep (Mbanzamihigo et al., 1996), cattle studies have shown loss of methane suppressing activity with prolonged (Johnson et al., 1994; McCaughey et al., 1997) or repeated application (Sauer et al., 1998).
The poor persistence of monensin’s methane suppressing activity may be due to the development of resistance to the antibiotic as observed for gram positive fermentative bacteria (Newbold et al., 1993). Methanogens have not been screened for development of resistance to monensin. Strains of rumen methanogens do however differ in susceptibility to monensin (Chen and Wolin 1979) and prolonged use of the antibiotic may simply select for non-susceptible strains.
2.3.2.2 Compounds that deprive methanogens of hydrogen
Understanding of hydrogen production, transfer and distribution within the rumen has been recently reviewed (Hegarty and Gerdes 1999), as has the scope for reducing methane production by chemical and microbiological means (Hegarty 1999b). A brief summary is provided below.
While reduction of sulfate and nitrate are both thermodynamically more favourable than is the reduction of CO2 to methane (Kristjansson et al., 1982; McAllister et al., 1996), the toxicity of their reduced forms prevent these being practical means of causing a substantial reduction in rumen methane emissions. Stoichiometric calculations show that reducing methane emissions of a sheep by 50% would require ingestion of 0.75 moles of sulphate or nitrate per day. Since sulphate and nitrate are toxic to sheep at approximately 0.1 moles/d and 0.25 moles/d respectively, they cannot safely be fed at levels appropriate to significantly reduce methane emissions (Hegarty 1999b).
Malate and fumarate, as direct metabolic precursors of propionate, reduce methane production when fed in a pure form or in high malate forages (Martin 1998). Calculations by Lopez et al., (1999) suggest the relationship between fumarate supply and methane production is in keeping with stoichiometry for the repartitioning of hydrogen from FADH into succinate, rather than into methane. Asanuma et al., (1999) have further showed a number of rumen bacteria utilise formate as an electron donor for fumarate reduction and showed that this reaction proceeds at the expense of methanogenesis when these organisms are co-cultured with methanogens. Malate has similar effects on fermentation as fumarate (Martin and Streeter 1995), reflecting that malate is rapidly metabolised to fumarate in the succinate-propionate pathway.
A previously unconsidered role of fumarate is as an electron sink for acetate formation by reductive acetogens, as demonstrated in Clostridium formicoaceticum by Dorn et
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (19 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
al., (1978). Additionally, while the mechanism is not clear, malate stimulates lactate uptake by Selenomonas ruminantium (Martin 1998) and this may contribute to the higher pH frequently observed in organic-acid treated incubations (Martin and Streeter 1995; Callaway and Martin 1996).
It may be concluded that, with the possible exception of utilising malate rich forages, provision of alternate hydrogen acceptors in the diet for the purposes of competitively reducing rumen methanogenesis is unlikely to be a broadly applicable emission control strategy.
2.3.2.3 Oxidation of methane in the rumen
Oxidation of methane is a normal aerobic reaction in soils (Schnell and King 1995), but has only recently been evaluated in the anaerobic environment of the rumen using introduced organisms. Kajikawa and Newbold (2000) used stable carbon isotopes to determine that 0.3% of methane was naturally oxidised in short-term in-vitro incubations of rumen fluid. Methane oxidation was inhibited by 0.5% O2 in the headspace gas and this inhibition was linked with sulphate reduction. Small but significant decreases in methane production have also being achieved by addition of an exogenous aerobic methane oxidiser (Brevibacillus parabrevis) to short and long-term incubations of rumen fluid (Valdés et al., 1997; Nelson et al., 2000). While the added organisms did not reduce the methanogen population, Nelson et al., (2000) postulated that uptake of dissolved oxygen by B. parabrevis may have been responsible for the observed increase in total culturable bacteria. Redox potentials were not reported.
2.3.2.4 Direct biological control of methanogens
Rumen fluid contains up to 109 bacteriophage/ml (Klieve and Swain 1993) and phages have been identified in many rumen bacterial species (Klieve et al., 1989). Despite viruses having been identified in non-ruminal methanogenic Archaea, there have been no studies of natural phage infection in ruminal methanogens. Rumen bactieriophage are in some cases strain specific (Klieve et al., 1991) and if Archaeal viruses share this trait, then development of a broadly effective virus-based control for rumen methanogens could be difficult (Klieve and Hegarty 1999). The heterogeneity identified in methanogens by 16S rRNA sequencing (Wright and Baker 1999) has been taken to indicate methanogens colonised the gastrointestinal tract on a number of occasions. Consequently, it is reasonable to expect multiple strains of Archaea will be present in the rumen and this could limit the efficacy of biological control.
2.3.2.5 Indirect ways of altering rumen ecology to reduce methane production
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (20 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
Immunisation
Vaccines provide a means of avoiding livestock production losses due to pathogenic infectious bacteria (Raadsma 1995) and may also be effective against external and internal parasites (James 1990; Newton 1995; Bain 1999). Gnanasampanthan et al., (1994) reported protozoal predation of E. coli was reduced in the presence of antiprotozoal antiserum, raising the prospect of selective control of rumen organisms by vaccination. This was demonstrated for Streptococcus bovis by Shu et al., (1999). Natural development of antibodies by lambs to mixed methanogens has recently been shown (Holloway and Baker 2000) and development of a vaccination strategy to reduce rumen methanogensis is in progress (Baker 1999).
Elimination of ciliate protozoa
Ciliate protozoa are an important cohabitant with methanogens in the rumen, providing both a site for methanogen attachment and a source of hydrogen for these organisms (Newbold et al., 1995). The role of protozoa in ruminal methanogenesis and the potential to reduce methanogenesis indirectly, through the elimination or suppression of ciliate protozoa have been reviewed (Hegarty 1999a). Unlike strategies which directly target methanogens, control of protozoa is unlikely to lead to hydrogen accumulation and an inhibition of general fermentation. While use of chemicals for protozoal control is no more desirable than is their use for methanogen control, the scope for biological control of rumen protozoa is promising (Klieve and Hegarty 1999).
Potential biological control agents for rumen protozoa include the Chytrid fungi, first observed in 1921 (see Kirby 1964). Both DNA and RNA viruses have also been reported in protozoa from other ecosystems but in many cases these viruses appear to have coexisted with the protozoal host (Widmir and Dooley 1995) and there are few cases where significant host lysis has been demonstrated (Garza and Suttle 1995). No viruses have been isolated from rumen protozoa and there has been no recent research to follow up early observations of fungal (Lubinsky 1955) and rickettsia-like organisms (Prins and Vorstenbosch 1975) in rumen protozoa that may have application as biological control agents.
Enhanced bacteriocin production
Bacteriocins are naturally occuring bacterial products with a bacteriocidal action. The most well understood bacteriocin is the food additive, nisin, which is effective against not just food-borne bacteria but also ruminal staphylococci and enterococci (Laukova 1995). Although the mechanism is unclear, nisin has been shown to reduce rumen methanogenesis by 36% (Callaway et al., 1997).
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (21 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
More important than exogenous bacteriocins however, may be the bacteriocins produced within the rumen itself. Kalmakoff et al., (1996) surveyed 50 strains of Butyrivibrio and found approximately half of the strains produced inhibitory (presumed bacteriocin) activity, and a large spectrum in inhibition was observed. Since many lactic-acid bacteria produce bacteriocins, it may well be that part of the reduced methane production observed at very low pH (Russell 1998) is due to bacteriocin effects on methanogens rather than a direct pH effect. Although the potential for ruminally produced bacteriocins to suppress methanogenesis is unknown, their potential as a new generation of rumen modifiers is already being considered (Teather and Forster 1998).
Enhanced acetogenesis
Reductive acetogenesis utilises hydrogen gas and CO2 to produce acetate. Acetogenic bacteria are present in the rumen at population densities which may reach that of methanogens (Leedle and Greening 1988) but despite their presence, reductive acetogenesis is extremely difficult to induce in the rumen (Immig et al., 1996).
One of the major constraints to induction of the acetogenic pathway in the rumen is thought to be that the hydrogen threshold for acetogenesis is higher than for methanogenesis as shown by Le Van et al (1998) for Acetitomaculum ruminis. In reviewing constraints to acetogenesis, Joblin (1999) lists at least 5 genera of acetogens as being present in the rumen so acetogens with low hydrogen thresholds may still be found. Recent in-vivo studies have shown a small flux of ruminal CO2 into acetate but this could have come via cycling through mammalian metabolism and is not conclusive evidence of ruminal acetogenesis (Tahir 1999). Joblin (1999) concludes that acetogenesis may find its ruminal application in ensuring hydrogen does not accumulate when methanogenesis is inhibited by other means, rather than acetogens out-competing methanogens for hydrogen in an otherwise untreated rumen.
Yeast cultures
There are limited reports indicating yeast cultures may reduce methane production (Martin et al., 1989). Other more readily measured effects of yeast extract or cultures on rumen fermentation are supportive of reduced methanogenesis based on stoichiometric principles. Observed effects which would reduce hydrogen availability to methanogens include (1) frequent shift in fermentation towards butyrate or propionate (Martin et al., 1989), (2) reduction in protozoal numbers (Newbold et al., 1998) and (3) promotion of acetogenesis as a sink for hydrogen (Chaucheyras et al., 1995). Importantly the effects of yeast culture on the general fermentation pattern are not
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (22 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
consistent across experiments or consistent even between strains of yeast (Newbold et al., 1995b). The effects are also short term, diminishing 2-4 h after feeding (Koul 1998). As such it is unrealistic to draw general conclusions on the efficacy of yeast cultures as inhibitors of rumen methanogenesis.
2.4 General assessment and conclusions regarding existing information
2.4.1 Assessment of producer information
A review of information already disseminated to livestock producers via the media and government agencies has not been conducted, however the following basic points bear consideration:
● Only 4% of Bachelor of Arts (Education) undergraduates can convincingly explain the greenhouse effect (Love 1997) and it is regularly confused with global warming due to polar holes in the ozone layer. There is no reason to believe that Australian livestock producers or the general community have any higher technical understanding than the British university students surveyed, and the frequent comments about ‘cows burning a hole in the ozone layer’ and ‘flatulent cows’, support this notion.
● Livestock producers cannot be aware of emission reduction solutions because there have been no sure and simple abatement policies recommended, even in the scientific literature.
● There has been no consideration of the environmental ramifications of current animal selection and breeding objectives.
2.4.2 Assessment of scientific information
Access
● Much of the current livestock methane research worldwide is not available in the public domain due to commercial-in-confidence agreements, but will hopefully lead to commercially useful products in time.
● This will necessitate repetition of research between laboratories. Considering the lack of basic information available and of research being conducted, this should be considered desirable rather than inefficient.
Clarity
● Scientifically, nutritionists have provided an inadequate message on the effects of feed quality and grazing system management on methane emissions.
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (23 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
● There is a need to clarify the message on feed quality effects and promote a clear unified position to producers and policymakers.
Comprehensiveness
● A stalling of rumen microbial research in the 1990s means that nationally and internationally there is a paucity of skills and tools in rumen ecology management to deliver ‘off-the-shelf’ emission abatement tools or even proven principles to the livestock industries.
● Existing knowledge of fermentation chemistry and rumen microbial ecology will need to be extended as a basis for building emission abatement strategies.
Applicability
● Scientific reviews have generally avoided providing practical recommendations on how emissions can be reduced.
● Feed additives (including dietary fats and dedicated antibiotics) likely to be developed and used overseas may only have application to the small portion of the Australian ruminant livestock industry that are intensively fed due to the extensive nature of most Australian grazing enterprises.
2.4.3 Conclusions about abatement through managing feed quality
A simple practical understanding of the effects of improved nutrition (feed digestibility) on the daily emission of methane from ruminants can be described below.
1. Historical expression of methane production as the proportion of dietary energy lost as methane has led to a false perception that better nutrition reduces daily emissions per head.
2. The studies providing this understanding did not consider the real effects of diet characteristics on voluntary feed intake.
3. The fact that improving forage digestibility increases the voluntary feed intake of grazing ruminants means that daily methane emission from a grazing ruminant will be increased by increasing the digestibility of pasture.
4. There remains scope for improving pasture digestibility and simultaneously reducing daily emissions, but this effect will only be achieved through reducing animal numbers and/or time spent grazing in association with pasture improvement.
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (24 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
5. Raising feed digestibility for animals already consuming high digestibility feeds (eg. feedlot rations) reduces rumen methane emission because of changes in the rumen conditions associated with diets rich in starch.
6. The simple partitioning of feed into forages versus feedlot diets provides one framework to account for differential effects of diet type on voluntary feed intake.
2.4.4 Conclusions regarding information on other potential abatement strategies
Despite the lack of immediate solutions, there are two distinct approaches that can be expected to deliver significant abatement, and there are multiple mechanisms within each strategy by which it can be achieved. These strategies are (1) changing attributes of the animal population and (2) the development of self-sustaining changes in the mixed microbial population of the rumen.
Breeding for ‘greenhouse-friendly’ animals is an achievable and implementable goal, albeit with a delayed pay-off period.
There are a number of opportunities for rumen modification to achieve sustained inhibition of rumen methanogenesis. These approaches will rely on initiating a stable ecological change in the rumen by some trigger. These triggers may include the suppression of methanogens by specific pathogens, vaccination, or the presence of a bacteriocin-producing organism. Alternately, these control approaches may be directed to the rumen protozoa with which the methanogens interact or may involve establishment of organisms introduced into the rumen.
Importantly, it may be concluded that development of some of these novel control strategies is essential to reduce the annual methane emission of grazing ruminants in the future.
2.5 Implications for the National Greenhouse Gas Inventory (NGGI)
At present the annual enteric emissions from the livestock sector are calculated from the number of animals and their daily emissions which are estimated from some systematically derived estimate of feed intake and the proportion of energy lost as methane. No allowance is made for carbon sinks in the livestock sector so sinks and implications for net emissions have been excluded from this report.
Of the potential abatement strategies described, the effects of some will be accounted for directly by the current inventory procedure but others will not. For example, reduction of emissions by altering pasture digestibility and stocking rate will be
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (25 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
automatically reflected by the current inventory procedure. Similarly, simple reduction in animal numbers or animal size will be accounted for by the current procedures. For abatement strategies that ‘bend the rules’ away from normal growth or digestive metabolism however, specific correction factors will need to be built into the National Greenhouse Gas Inventory (NGGI). This is the case for many of the listed strategies as indicated below.
● Selection for high NFE animals. It will not be possible to use equations that use growth rate of the animal as a basis for estimating intake without a correction factor once selection for NFE is occurring.
● Rumen manipulation. Any of the chemical or biological control tools that reduce the quantity of methane released per unit food or energy eaten, will require to be accounted for by a correction factor. This will need to be developed and applied in the NGGI to the proportion of livestock using this strategy.
In conclusion, the issue of having the abatements achieved, recognised in the NGGI is critical. Technically, abatement effects can be quantified by documenting the efficacy of the strategy (% reduction in emissions relative to untreated animals) plus the number of animals affected, then calculating an appropriate correction factor. The validity of these estimates could be easily proven but the ability to implement correction factors within the IPCC methodologies would need to be investigated.
2.6 Gaps in recent research activity on enteric emissions
In considering Australia’s current capabilities and options for future emissions abatement, it is worth reflecting on the weaker points of ruminant research pertinent to methane.
In 30 years of research, ruminant nutritionists have comprehensively demonstrated the inability of almost all known ‘methane inhibiting’ compounds to achieve a sustained inhibition of methanogenesis. We have often demonstrated that more effective direct inhibition of methane production simply leads to more serious reduction in general rumen function.
Research on rumen methane has done much to build and perpetuate the current understanding of rumen stoichiometry (chemical pathways). This stoichiometry has gone unchallenged, but is rarely able to account for all the hydrogen generated in the rumen. Few other fields of science would have tolerated the failure of rumen microbiology to refine and expand this stoichiometry in the past 15 years. We have allowed studies of methane inhibition to be reported in which 50-75% of the calculated hydrogen production is accounted for, yet have rarely questioned where the missing
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (26 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
hydrogen goes, or if it was ever produced!
The above criticisms are indicative of a fundamental failure world-wide to adequately study the physical and chemical interactions which occur between the multitude of rumen organisms. The continued failure in the past decade to define this basic ecology has left the livestock industries ill-prepared to develop ‘clever’ strategies to reduce rumen emissions. We do not even have on the public record the identity of methanogens which predominate in Australian ruminants, nor do we know whether the process of acetogenesis ever can or does, take the place of methanogenesis in rumen fermentations. Against this background, the following specific deficiencies are identified in information pertinent to reducing emissions through animal or ruminal management:
Basic animal information which is lacking
● The scope for breeding animals which produce less methane per day without any decrease in product yield (meat, milk, fibre) has not been assessed. Animals with a lower methane emission but uncompromised performance can be expected to occur due to:
eating less but maintaining a similar growth rate (high net feed efficiency; NFE)being smaller (reduced mature size) having faster digesta kinetics (less favourable to methanogens)
● Possible differences in digestive function that underlie differences in NFE have not been evaluated.
● Simple modelling exercises to show the emissions and commercial consequences of selecting for low mature liveweight breeders and use of cross breeding to attain desired progeny performance have not been undertaken. This could be one of the first practical options to be put to livestock breeders to reduce emissions.
● Producing lower emissions through selection of animals for differential gut function (specifically lower methane production) has not been researched, although low methane producing individuals have been identified.
Basic grazing system information that is lacking
● Pasture intake is a critical regulator of enteric emissions but its association with pasture and animal traits is only poorly understood.
● Adequate modelling of the effects of pasture improvement on feed intake, methane emission, sustainable stocking rate and soil carbon changes associated
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (27 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapter 2
with the grazing system is required. ● A modelling framework is needed to support development of recommendations
on allocation of land within a farm to improved pasture to achieve increased livestock output while reducing methane and net carbon emissions.
Basic rumen ecology information which is lacking
● Publicly available information on the species and strains of methanogens in Australian livestock is currently inadequate to allow development of control measures targeted at predominant methanogens.
● Chemical elimination of rumen protozoa has conclusively demonstrated this treatment achieves reduced methane emissions with no adverse and often positive livestock performance consequences. Commercial means of eliminating rumen protozoa need to be developed.
● Potential biological control (pathogenic) organisms targeting rumen methanogens or the protozoa with which they associate, have not been researched.
● The adaptation of methanogens to chemical inhibitors is not understood, and monitoring and assessment of adaptation will be required for novel control measures such as vaccines or biological controls.
● The effect of natural bacteriocins in inhibiting rumen methanogenesis looks encouraging but is not being evaluated in Australia.
● Information on the range, activity and hydrogen thresholds of naturally occurring, gut-dwelling reductive acetogens in Australian livestock is not available.
Contents > Previous > NextAGO Home | Contacts | Publications | Search | Site index | What's new | Top
Science & emissions monitoring International activities Government Business & industryCommunity & household Energy & transport Natural resources
Comments Copyright Disclaimer Notice Privacy Accessibility
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html Last modified 17 November 2002
http://www.greenhouse.gov.au/agriculture/livestock/chapter2.html (28 of 28) [5/21/2003 1:26:23 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
The Australian Greenhouse Office Skip Navigation
Leading Australia's greenhouse action to achieve effective sustainable results
Greenhouse and agriculture Home > Natural resources > Agriculture > Contents
Greenhouse gas emissions from the Australian livestock sector: what do we know, what can we do?Prepared by Dr Roger Hegarty, NSW Agriculture,to contribute to the work of the Greenhouse and Agriculture Taskforce, 2001
3. Emissions from manureContents > Previous
3.1 Available information
Microbial degradation of manures can release the greenhouse gases carbon dioxide, methane and nitrous oxide.
In microbial and chemical degradation of excreta, a trade-off exists between disposing of wastes by aerobic treatments (leading to carbon dioxide and nitrous oxide emissions) compared with anaerobic treatment (which promotes methane release). The 1999 NGGI inventory shows that approximately 98% of greenhouse gas emissions from livestock manure occur as methane and only approximately 2% occur as nitrous oxide and these two products will be addressed separately.
Although 98% of manure emissions are methane, this contributes only 3% of total methane emissions from the livestock sector, with almost all of the methane being released from collected, ponded or composted manures from pigs, poultry and dairy cattle, rather than the ‘free range’ manure that characterises the extensive livestock industries. Manure therefore constitutes a very small source of greenhouse gas emissions relative to enteric fermentation and therefore benefits to national emissions from targeting these emissions can only be small.
Since methanogens are easily killed by oxygen, it would be expected that aeration of slurries or turning of manure piles, would be effective in stopping methane production. In practice, the opposite is true and this probably reflects the effect of stirring or turning in improving substrate availability to the microbes associated with fermentation, that in
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (1 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
turn provide hydrogen to the methanogens. For effluent ponds, the retention of an unbroken crust will ensure lower emissions than regular decrusting of the pond (Farran et al., 2000). Similarly, turning of manure piles increases methane production for 3 to 4 days. This effect of turning is associated with increased stack temperature and ammonia release (Kuroda et al., 1996).
One of the features of manure slurries is the extremely high oxygen demand they possess. This means that aerating/oxygenating these slurries to avoid methane release is almost a technological and economic impossibility due to the scale of agitation and aeration required.
Nitrous oxide from manures is only <0.1% of the total greenhouse gas emission from the livestock sector and estimation of its release relies on description of energy and protein and digestibility characteristics of the feed. Under different climatic conditions, Oenema et al., (1997) suggested 2% of excreted nitrogen is emitted as N2O. Use of this value has recently increased the UK inventory (Chadwick et al., 1999).
3.2 Management options
Manure is simply undigested feed which was too resilient to be digested by gut microbes in the time available, and has subsequently been enriched with nitrogen and minerals through mixing with urine. The continuation of further microbial digestion of this undigested feed, now enriched with urinary nutrients, is inevitable.
The simplest abatement approach is to minimise the manure (DM) production and loss of nitrogen in urine. Manure can be minimised by ensuring the energy requirements of the animals are met from the highest digestibility feed available, fed only at levels required for the desired animal performance. This is easily done in pig and poultry industries and to some extent in dairies.
The same approach of feeding to dietary requirements also holds true for limiting nitrous oxide emissions. In particular, formulating complete diets for their specific amino acid content rather than formulating on crude protein content can reduce the oversupply of non-essential amino acids provided in most protein-rich feedstuffs, so reducing nitrogen excretion. This is a growing practice in the Australian pig and poultry industries for economic rather than environmental reasons. Reduced nitrogen excretion without compromising animal performance has been demonstrated for this approach (Hobbs et al., 1996) and reduced N2O emissions resulted from this slurried manure being spread on grassland.
For poultry and pigs, inclusion of enzymes in the diet to digest soluble non-starch
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (2 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
polysaccahrides is becoming an industry standard procedure. It enables increased utilisation of feed by the animal, reducing excretion of energy in the faeces (Angkanaporn et al. 1994; Hughes et al. 2000).
Aside from using dietary design to minimise emissions, manure storage and disposal also provide opportunities to reduce emissions. Farran et al., (2000) reviewed the options for Australian piggery waste disposal and concluded "the use of direct land application of wastes offers the pig industry a reduction in greenhouse gas emissions of about 44% over the traditional anaerobic lagoon systems of waste management. The risks of nutrient run-off/seepage and legal constraints for waste disposal need to be considered, together with the area of land required."
Two other options for inhibiting emissions from manures include acidification and antibiotics, particularly those antibiotics included prophylactically in pig and poultry diets that are not absorbed or degraded in the gut. Both nitric and lactic acid have been considered for acidification, but the costs and corrosion risks are high. Use of prophylactic antibiotics is becoming less acceptable and use of antibiotics directly for manure management is unlikely to be acceptable to any registration authority for general use.
4 Overview of information gaps
There are 2 avenues by which emissions can be reduced without reducing livestock numbers. These are (1) the correct selection of animals and (2) the development of self-sustaining changes in the microbial population of the rumen. Additionally, reducing methane emissions in association with changing stock numbers and attributes of the grazing system needs to be considered.
4.1 Information required for breeding ‘greenhouse-friendly’ ruminants
● Budgets of feed use, greenhouse gas emissions and product output from enterprises with animals of smaller mature size.
● Methane emission measurements from ruminants selected for high net feed efficiency.
● Identification of low-methane producing ruminants for more intensive study. ● Assessment of the genetic differences in rumen kinetics among Australian
ruminants. ● The above information will likely result in selection of animals which use less feed
and produce less methane than average to achieve a given level of performance.
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (3 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
● If producers simply increase stocking rate to utilise this ‘spare feed’, total emissions will increase. A strategy is therefore needed to avoid subsequent increases in total stock numbers. Reducing grazing area may be one means to this end, and planting of trees for carbon sequestration, wildlife or forestry benefits would be one alternate land use for the area released from grazing.
4.2 Information required to establish stable, low-methane rumen fermentations
● Assessment, description and public access to the species and strains of methanogens in Australian ruminants.
● Identification of organisms which suppress or are pathogenic to the predominant rumen methanogens and associated rumen protozoa.
● Assessment of the susceptibility of ruminal methanogens to naturally occurring bacteriocins.
4.3 Information required for reducing emissions through grazing management
It is the author’s understanding that for Australia’s pasture based industries, pasture management or improvement to increase digestibility without a reduction in livestock numbers will increase the total methane emissions from livestock. There remains, however, a need for an unquestionable declaration of how pasture quality affects total methane emissions for the purpose of developing management recommendations. This statement will necessitate thorough modelling of the effect of pasture digestibility on (1) feed intake (2) metabolisability of digested energy, (3) daily emissions and (4) emissions per unit output. These studies would also include soil carbon reserves to enable net emissions for the enterprise to be calculated. Both Australian and overseas pasture intake models should be used to ensure the conclusion is robust and widely accepted.
5. Recommendations
5.1 Old things to change
● Get more irons in the fire. At present, only one potential abatement strategy for the Australian ruminant livestock industries is under development, this being a vaccine to target rumen methanogens. Placing total reliance on one potential strategy 8-12 years out from the first accounting period is naive. Should it fail,
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (4 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
there will be insufficient time to develop alternate emission reduction strategies before 2008. It is therefore recommended that Australia broaden research into abatement strategies to include other approaches listed in this report.
● Ignorance is not bliss. While we are unable to recommend practical emission abatement strategies for livestock producers, improving their understanding of the greenhouse effect and consequences of global warming for regional agriculture would be beneficial. It is therefore recommended that a public awareness program be developed for rural Australia, and this be an extension of a research and development program to develop abatement strategies.
● Bearing and sharing of responsibility. In the past decade, no single organisation has been willing to take ownership of livestock methane abatement. It has been perceived as a ‘bad-news’ agricultural story, with no obvious solutions. In contrast, tree planting has been a simple message, easily marketed with ecological, timber and possibly carbon-credit value, for those who participate. As a result only one major Australian R&D project on livestock emissions abatement has been funded compared to multiple funding avenues being available for extensive tree planting programs. It is recommended AGO, AFFA, livestock industry R&D corporations and departments of natural resources and environment, contribute resources to develop a significant research, development and extension (R,D &E) program to improve Australia’s understanding of emissions, and develop new technologies to reduce them. Let livestock emissions be faced and addressed rather than being left to the vaccine, or to somebody else.
5.2 New things to do
In previous pages, an array of potential abatement tools and specific deficiencies in information has been identified. Implementation of the following recommendations will ensure each of these abatement options is explored and developed, and the information deficiencies are overcome.
It is recommended:
1. That development of a livestock emissions R,D & E program be supported jointly by AGO, AFFA, Livestock Industry R&D corporations and departments of natural resources or environment. The program should make use of the research and advisory skills available in all relevant organisations.
2. That the program generate information for national good, with ownership of the patents and intellectual property arising, reflecting this purpose.
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (5 of 15) [5/21/2003 1:26:26 PM]
Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
3. That the program be managed in at least 3 sections: ❍ Rumen modification (identification of organisms, modifying ecological
balance) ❍ Information delivery (raised rural awareness, dissemination of research
results) ❍ Livestock management (selection, stocking rate, feed quality)
4. That the program establish model farms (farmletts) for the purpose of quantifying the environmental footprint of Australia’s livestock industries. While this extends beyond the charter of reducing greenhouse gas emissions, these model farms should be established to deliver budgets for energy, carbon, methane, nutrients, economics and water (including deep drainage and run-off). A thorough detailing of the full environmental impact of livestock enterprises will inevitably be required within the next decade. The greenhouse gas emission issue provides an incentive for this large, broad-based benchmarking exercise to be set-up. It will also provide opportunity to demonstrate the effectiveness of recommended stock and feed management practices arising from the R&D program.
6. Glossary
Anaerobic Without oxygen. Methane producing organisms are strictly anaerobic and easily killed through contact with oxygen.Archaea Primitive organisms of simpler biology than bacteria. Some Archaea are able to produce methane and these are referred to as the methanogenic
Archaea All methane produced from livestock is generated by methanogenic Archaea.
Defaunated Animals are said to be defaunated when their rumen has been treated to remove all ciliate protozoa.
Digestibility The percentage of feed dry matter consumed which disappears during passage through the digestive tract ((feed DM – faecal DM)/feed DM).
DM Dry matter.
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (6 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
Faunated Animals with rumen protozoa.
Fermentation Chemical digestion of feedstuffs by microorganisms in the absence of oxygen.
Maintenance The amount of energy required by an animal to simply maintain itself without either net gain or loss of energy. For domestic livestock the maintenance energy requirement approximates the amount of energy required to maintain a constant body weight.
Metabolisable energy (ME)
Energy obtained from the diet which is able to be utilised by the animal for maintenance or growth.
Methanogen Methane producing organism. Methanogens are primitive organisms and are not true bacteria, but rather are Archaea.
Protozoa Complex rumen organisms which are larger than bacteria and ferment dietary sugars and protein. The ciliate protozoa are important source do hydrogen for rumen methanogens and many methanogens live on and in the protozoa in a symbiotic relationship.
Rumen The large stomach of ruminants in which feed is fermented by micro-organisms. The fermentation vessel is often considered to include the 2nd stomach or reticulum.
Ruminant Herbivore with an expanded fore-gut consisting of several stomach compartments located prior to the intestine. Domestic ruminants (sheep, cattle, goats) have a 4-compartment rumen.
7. Key personnel
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (7 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
Research staff
Name Association Contact detailsDr. Sue Baker CSIRO Livestock Industries [email protected]. Graeme Faichney University of Sydney 02-93514508Dr. Roger Hegarty NSW Agriculture [email protected]
Dr. Bob Hunter CSIRO Livestock Industries [email protected]
Dr. Keith Joblin Agresearch, New Zealand [email protected]
Dr. Athol Klieve QDPI, Yeerongpilly [email protected]
Prof. John Nolan University of New England [email protected]
8. Bibliography
Reviews
Crutzen, P.J., Aselmann, I., Seiler, W. 1986. Methane Production by Domestic Animals, Wild Ruminants, Other Herbivorous Fauna and Humans. Tellus, 38B, 271-284.
Czerkawski, J.W. 1969. Methane Production in Ruminants and its Significance. World Review of Nutrition and Dietetics, 11, 240-282.
Joblin, K.N. 1996. Options for reducing methane emissions from ruminants in New Zealand and Australia. In Coping with Climate Change. (Eds. W. J. Bouma, G.I. Pearman, M.R. Manning) pp 437-449. CSIRO Publishing, Collingwood, Australia.
Johnson, D.E., Ward, G. 1996. Estimates of Animal Methane Emissions. Environmental Monitoring and Assessment, 42, 133-141.
Johnson, D.E., Hill, T.M., Carmean, B.R., Branine, M.E., Lodman, D.W., Ward, G. 1991. New Perspectives on Ruminant Methane Emissions. In Proceedings of the XII Symposium on Energy Metabolism of Farm Animals. pp376-379. Kartause Ittingen.
Jouany, J.P. 1994. Methane Production from Ruminants: Its Contribution to the Greenhouse Effect and how to Control Emission. International Conference on Feed Production, 1-17.
Kirchgessner, M., Windisch, W., and Muller, H.L. 1995. Nutritional factors for the quantification of methane production. In Ruminant Physiology: Digestion, Metabolism, Growth and Reproduction: Proceedings of the 8th International Symposium on Ruminant Physiology (Eds. W. v Engelhardt, S. Leonhard-Marek, G. Breves, D. Giesecke). pp 333-348. Ferdinand Enke Verlag, Stuttgart.
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (8 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
Kurihara, M., Shibata, M., Nishida, T., Purnomoadi, A., Terada, F. 1997. Methane Production and its Dietary Manipulation in Ruminants. In Ruminal Microbes and Digestive Physiology in Ruminants (Eds. R. Onodera et al.,) pp 199-208. Japan Scientific Societies Press, Tokyo.
McCrabb, G.J., Kurihara, M., Hunter, R.A. 1998. The Effect of Finishing Strategy on Lifetime Methane Production for Beef Cattle in Northern Australia. Proceedings of the Nutrition Society of Australia 22, 55.
Moss, A.R. (1993). Methane: Global Warming and Production by Animals. Chalcombe Publications, Kent.
Ulyatt, M.J., Lassey, K.R., Martin, R.J., Walker, C.F., Shelton, I.D. 1997. Methane Emission from Grazing Sheep and Cattle. Proceedings of the New Zealand Society of Animal Production, 57, 130-133.
Van Nevel, C.J., Demeyer, D.I. 1995. Feed Additives and Other Interventions for Decreasing Methane Emissions. In. Biotechnology and Animal Feeds and Feeding (Eds. R.J. Wallace, A. Chesson) pp 329-349. VCH Publishers, New York.
Van Nevel, C.J., Demeyer, D.I. 1996. Control of Rumen Methanogenesis. Environmental Monitoring and Assessment, 42, 73-97.
Full reference list
Angkanaporn, K., Choct, M., Bryden, W.L., Annison, E.F., Annison, G. 1994. Effects of wheat pentosans on endogenous amino acid losses in chickens. J. Sci. Food Agric. 66, 399-404.
Asanuma, N., Iwamoto, M., Hino, T. 1999. Effect of the addition of fumarate on methane production by ruminal microoganisms in vitro. J. Dairy Sci. 4, 780-787.
Bain, R.K. 1999. Irradiated vaccines for helminth control in livestock. Int. J. Parasit. 29, 185-191.
Baker, S.K. 1999. Biology of rumen methanogens and stimulation of animal immunity. In. Meeting the Kyoto Protocol: Implications for the Australian Livestock Industries. (Eds. P.J. Reyenga, S.M. Howden). pp29-37. Bureau of Rural Sciences, Canberra.
Bauchop, T. 1967. Inhibition of rumen methanogenesis by methane analogues. J. Bact. 94,171-175.
Benchaar, C., Rivest, J., Pomar, C., Chiquette, J. 1998. Prediction of methane production from dairy cows using existing mechanistic models and regression equations. J. Anim. Sci. 76, 617-627.
Blaxter, K.L., Clapperton, J.L. 1965. Prediction of the amount of methane produced by ruminants. Brit. J. Nutr. 19, 511-522.
Blaxter, K.L., Czerkawski, J. 1966. Modifications of the methane production of the sheep by supplementation of its diet. J. Sci. Food Agric. 17, 417-421.
Callaway, T.R., Martin, S.A. 1996. Effects of organic acid and monensin treatment on in vitro mixed ruminal microorganism fermentation of cracked corn. J. Anim. Sci. 74, 1982-1989.
Callaway, T.R., Carneiro De Melo, A.M.S., Russell, J.B. 1997. The effect of nisin and monensin on ruminal fermentations in vitro. Curr. Micro. 35, 90-96.
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (9 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
Chadwick, D.R., Sneath, R.W., Phillips, V.R., Pain, B.F. 1999. A UK inventory of nitrous oxide emissions fom farmed livestock. Atmos. Env. 33, 3345-3354.
Chaucheyras, F., Fonty, G., Bertin, G., Salmon, Gouet, P. 1995. In-vitro H2 utilisation by a ruminal acetogenic bacterium cultivated alone or in association with an Archea methanogen is stimulated by a probiotic strain of Saccharomyces cerevisiae. Appl. Env. Micro. 61, 3466-3467.
Chen, M., Wolin, M.J. 1979. Effect of monensin and lasalocid-sodium on the growth of methanogenic and rumen saccharolytic bacteria. Appl. Env. Micro. 38, 72-77.
Czerkawski, J.W. 1986. An introduction to rumen studies. Pergamon Press Ltd. Oxford.
Czerkawski, J.W., Breckenridge, G. 1975. New inhibitors of methane production by rumen microorganisms. Development and testing of inhibitors in vitro. Brit. J. Nutr. 34, 429-445.
Dohme, F. 2000. Reasons and conditions for the methane-suppressing effect of medium-chain saturated fatty acids as investigated with rumen simulation technique. Ph.D. Thesis. Swiss Federal Institute of Technology, Zurich.
Dohme, F., Machmuller, A., Wasserfallen, Kreuzer, M. 2000. Comparative efficiency of various fats rich in medium chain fatty acids to suppress ruminal methanogenesis as measured with RUSITEC. Can. J. Anim. Sci. 80, 473-482.
Dong. Y., Bae, H.D., McAllister, T.A., Mathison, G.W., Cheng, K.G. 1997. Lipid induced depression of methane production and digestibility in the artificial rumen system (RUSITEC). Can. J. Anim. Sci. 77, 269-278.
Dorn, M., Andreesen, J.R., Gottschalk, G. 1978. Fermentation of fumarate, and L-malate by Clostridium formicoaceticum. J. Bact. 133, 26-32.
Faichney, G.J. 1995. Contribution of the breeding herd to methane emissions during the production of beef from pasture. Proc. Nutr. Soc. Aust. 19, 195.
Farran, I., Maul, C., Scott, C. 2000. Greenhouse gas emissions: Waste management options for the pig industry. Project Report DPI 3/1156. Pig Research and Development Corporation and the Department of Agriculture, Fisheries and Forestry Australia.
Finlay, B.J., Esteban, G., Clarke, K.G., Williams, A.G., Embley, T.M., Hirt, R.P. 1994. Some rumen ciliates have endo-symbiotic methanogens. FEMS Micro. Lett. 117, 157-162.
Freer, M., Jones, D.B. 1984. Feeding value of subterranean clover, lucerne, phalaris and Wimmera ryegrass for lambs. Aust. J. Exp. Agric. Anim. Husb. 24, 156-164.
Garza, D.R., Suttle, C.A. 1995. Large double-stranded DNA viruses which cause the lysis of a marine heterotrophic nanoflagellate (Bodo sp.) occur in natural marine viral communities. Aquatic Microb. Ecol. 9, 203-210.
Gnanasampanthan, G., Hynd, P.I., Mayrhoffer, G. 1994. Effect of sheep antibodies on engulfment of bacteria by rumen protozoa. Proc. Aust. Soc. Anim. Prod. 20, 389.
Goodrich, R.D., Garrett, J.E., Gast, D.R., Kirick M.A., Larson, D.A., Meiske, J.C. 1984. Influence of monensin on the performance of cattle. J. Anim. Sci. 58, 1484-1498.
Gottschalk, G. 1986. Bacterial Metabolism (2nd ed.). Springer-Verlag, New York.
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (10 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
Graham, J. 2001. The significance of the feedlot sector of the beef industry. In. The Beef Program – Challenges and future directions. Proceedings of the NSW Agriculture Beef Products Conference, Dubbo, May 2001. pp145-146. NSW Agriculture, Orange, Australia.
Hegarty, R.S. 1998. Practical methods for reducing methane emissions from Australian livestock. In Meeting the Kyoto Protocol: Implications for the Australian Livestock Industries. (Eds. P.J. Reyenga, S.M. Howden) pp87-94. Bureau of Rural Sciences, Canberra.
Hegarty, R.S. 1999a. Reducing rumen methane emissions through elimination of rumen protozoa. Aust. J. Agric. Res. 50, 1321-1327.
Hegarty, R.S. 1999b. Mechanisms for competitively reducing ruminal methanogenesis. Aust. J. Agric. Res. 50, 1299-1305.
Hegarty, R., Gerdes, R. 1998. Hydrogen production and transfer in the rumen. Rec. Adv. Anim. Nutr. 12, 37-44.
Henderson, C. 1973. The effects of fatty acids on pure cultures of rumen bacteria. J. Agric. Sci. 81, 107-112.
Hobbs, P.J., Pain., B.F. , Kay, R.M., Lee, P.A. 1996. Reduction of odorous compounds in fresh pig slurry by dietary control of crude protein. J. Sci. Food Agric. 71, 508-514.
Holloway, P.E., Baker, S.K., 2000. Development of antibodies to rumen methanogens in sheep. Proc. Austr. Soc. Anim. Prod.
Hughes, R.J., Choct, M., Kocher, A., Van-Barneveld, R.J. 2000. Effect of food enzymes on AME and composition of digesta from broiler chickens fed on diets containing non-starch polysaccharides isolated from lupin kernel. Brit. Poult. Sci. 41, 318-323.
Immig, I., Demeyer, D.I., Fiedler, D., Van Devel, C.J., Mbanzamihigo, L. 1996. Attempts to induce acetogenesis into a sheep rumen. Arch. of Anim. Nutr. Berlin 49, 363-370.
James, P.J. 1990. Strategic use of, and alternatives to, chemicals for the control of arthropod pests of livestock. Proc. Aust. Soc. Anim. Prod. 18, 37-52.
Joblin, K.N. 1999. Ruminal acetogens and their potential to lower ruminant methane emissions. Aust. J. Agric. Res. 50, 1307-1313.
Johnson, D.E., Abo-Omar, J.S., Saa, C.F., Carmean, B.R. 1994. Persistence of methane suppression by propionate enhancers in cattle diets. In Proceedings of XIII Symposium on Energy Metabolism of Farm Animals. (Ed. J.F. Aguilera) pp339-342. EAAP Publ. 76 CSIS, Spain.
Kajikawa, H. and Newbold, C.J. 2000. Methane oxidation in the rumen. J. Anim. Sci. 78 (Suppl.), 291.
Kalmakoff, M.L., Bartlett, F., Teather, R.M. 1996. Are ruminal bacteria armed with bacteriocins. J. Dairy Sci. 79, 2297-2306.
Kirby, H. Jr. 1964. Organisms living on and in protozoa. In Protozoa in Biological Research. (Eds. G.N. Calkins, F.M. Summers.) pp1009-1114. Hafner Publishing Co. Inc., New York.
Klieve, A.V., Swain, R.A. 1993. Estimating ruminal bacteriophage numbers using pulsed field gel electrophoresis and laser densitometry. Appl. Env. Micro. 59, 2299-2303.
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (11 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
Klieve, A.V., Hudman J.F., Bauchop, T. 1989. Inducible bacteriophages from ruminal bacteria. Appl. Env. Micro. 55, 1630-1634.
Klieve, A.V., Gregg, K., Bauchop, T. 1991. Isolation and characteristics of lytic phages from Bacteroides ruminicola ss brevis. Curr. Micro. 23, 183-187.
Klieve, A., Hegarty, R.S. 1999. Opportunities for biological control of methanogensis. Aust. J. Agric. Res. 50, 1315-1319.
Koul, V. 1998. Mode of action of yeast culture (YEA-SACC 1026) for stimulation of rumen fermentation in buffalo calves. J. Sci. Food Agric. 77, 407-413.
Kowalczyk, J., Orskov, E.R., Robinson, J.J., Stewart, C.S. 1977. Effect of fat supplementation on voluntary feed intake and rumen metabolism in sheep. Brit. J. Nutr. 37, 251-257.
Kreuzer, M. 1986. Methodik und Anwendung der Defaunierung bein wachsenden Wiederkauer. J. Vet. Med. (A) 33, 721-745.
Kristjansson, J., Schonheit, P., Thauer, R. 1982. Different K values for hydrogen of methanogenic bacteria and sulfate reducing bacteria: An explanation for the apparent inhibition of methanogenesis by sulfate. Arch. Microbiol. 131, 278-282.
Kuroda, K., Osada, T., Yonaga, M., Kanematu, A., Nitta, T., Mouri, S., Kojima, T. 1996. Emissions of malodorous compounds and greenhouse gases from composting swine feces. Bioresource Tech. 56, 265-271.
Lana, R.P., Russell, J.B., Van Amburgh, M.E. 1998. The role of pH in regulating ruminal methane and ammonia production. J. Anim. Sci. 76, 2190-2196.
Laukova, A. 1995. Inhibition of Ruminal Staphylococci and Enterococci by Nisin In-Vitro. Lett. Appl. Micro. 20, 24-36.
Leedle, J.A.Z., Greening, R.C. 1988. Postprandial changes in methanogenic and acidogenic bacteria in the rumens of steers fed high- or low-forage diets once daily. Appl. Env. Micro. 54, 502-506.
Le Van, T.D., Robinson, J.A., Ralph, J., Greening, R.C., Smolenski, W.J., Leedle, J.A.Z., Schaefer, D.M. 1998. Assessment of reductive acetogenesis with indigenous ruminal bacterium populations and Acetitomacumum ruminis. Appl. Env. Micro. 64, 3429-3436.
Lopez, S., Newbold, C.J., Bochi-Brum, O., Moss, A.R., Wallace, R.J. 1999. Propionate precursors and other metabolic intermediates as possible alternative electron acceptors to methanogenesis in ruminal fermentation in vitro. S Afr. J. Anim. Sci. 29, 106-107.
Love, J. 1997. Student teacher understanding of the greenhouse effect, ozone layer depletion and acid rain. Env. Educ. Res. 2, 89-100.
Lubinsky, G. 1955. On some parasites of parasitic protozoa. 1. Sphaerita hoari sp. N. - A chytrid parasitising Eremoplastron bovis. Can. J. Micro. 1, 440-450.
Machmuller, A. and Kreuzer, M. 1999. Methane suppression by coconut oil and associated effects on nutrient and energy balance in sheep. Can. J. Anim. Sci. 79, 65-72.
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (12 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
Machmuller, A., Ossowski, D.A., Wanner, W., Kreuzer, M. 1998. Potential of various fatty feeds to reduce methane release from rumen fermentation in vitro (rusitec). Anim. Feed Sci. Tech. 71, 117-130.
Martin S.A. 1998. Manipulation of ruminal fermentation with organic acids: a review. J. Anim. Sci. 76, 3123-3132.
Martin, S.A., Streeter, M.N. 1995. Effect of malate on in vitro mixed ruminal microorganism fermentation. J. Anim. Sci. 7, 2141-2145.
Martin, S.A., Nisbet, D.J., Dean R.G. 1989. Influence of a commercial yeast supplement on the in-vitro ruminal fermentation. Nutr. Rep. Int. 40, 395-403.
Mbanzamihigo, L., Van Nevel, C.J., Demeyer, D.I. 1996. Lasting effects of monensin on rumen and caecal fermentation in sheep fed a high grain diet. Anim. Feed Sci. Tech. 62, 215-228.
McAllister, T.A., Okine, E.K., Mathison, G.W., Cheng, K.N. 1996. Dietary, environmental and microbiological aspects of methane production in ruminants. Can. J. Anim. Sci. 76, 231-243
McCaughey, W.P., Wittenburg, K.M., Corrigan, D. 1997. Methane production by steers on pasture. Can. J. Anim. Sci. 519-524.
McCrabb, G.J., Berger, K.T., Magner, T., May, C., Hunter, R.A. 1997. Inhibiting methane production in Brahman cattle by dietary supplementation with a novel compound and the effects on growth. Aust. J. Agric. Res. 48, 323-329.
Mrode, R.A., Smith, C., Thompson, R. 1990. Selection for rate and efficiency of lean gain in Hereford cattle. 2. Evaluation of correlated responses. Anim. Prod. 51, 35-46.
NGGI 1994. Agriculture: Workbook 6.0 for Livestock. Department of the Environment, Sport and Territories, Canberra.
NGGI 1998. Agriculture: Workbook 6.1 for Livestock. Australian Greenhouse Office, Canberra.
NGGI 1999. National Greenhouse Gas Inventory 1999 with methodology supplements. Australian Greenhouse Office, Canberra.
Nelson, N., Valdes, C., Hillman, K., McEwan, N.R., Wallace, R.J., Newbold, C.J. 2000. Effect of a methane oxidizing bacterium isolated from the gut of piglets on methane production in Rusitec. Reprod. Nutr. Devel. 40, 212.
Newbold, C.J. Wallace, R.J. Walker, N. 1993. The effect of Tetronasin and Monensin on fermentation, microbial numbers and the development of ionophore-resistant bacteria in the rumen. J. Appl. Bact. 75, 129-134.
Newbold, C.J., Lassalas, B., Jouany, J.P. 1995. The importance of methanogens associated with ciliate protozoa in ruminal methane production in-vitro. Lett. Appl. Micro. 21, 230-234.
Newbold. C.J., Wallace, R.J., Chen, X.B., McIntosh, F.M. 1995b. Different strains of Saccharomyces cerevisiae differ in their effects on ruminal bacterial numbers in-vitro and in sheep. J. Anim. Sci. 73, 1811-1818.
Newbold, C.I., McIntosh, F.M., Wallace, R.J. 1998. Changes in the microbial population of a rumen simulating fermenter in response to yeast culture. Can. J. Anim. Sci. 78, 241-244.
Newton, S.E. 1995. Progress on vaccination against Haemonchus contortus. Int. J. Parasitology. 25, 1281-1289.
Oenema, O., Velthof, G.L., Yamulki, S., Jarvis, S.C. 1997. Nitrous oxide emissions from grazed grassland. Soil Use
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (13 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
Manage. 13, 288-295.
O’Keeffe, M., Kennedy, O. 1998. Residues – A food safety problem? J. Food Safety 18, 297-319.
Okine, E.K., Mathison, G.W., and R.T. Hardin 1989. Effects of changes in frequency of reticular contractions on fluid and particulate passage rates in cattle. J. Anim. Sci. 67, 3388-3396.
Pelchen, A., Peters, K.J. 1998. Methane emissions from sheep. Small Ruminant Res. 27, 137-150.
Prins, R.A., van den Vorstenbosch, C.J.A.H.V. 1975. Interrelationships between rumen microorganisms. Physiology of Digestion. Miscellaneous Papers. (Landbouwhogeschool Wageningen.) 11, 18-23.
Raadsma, H. W. 1995. Genetic variation in resistance to bacteria. In. Breeding for resistance to infectious diseases in small ruminants. (Eds. G.D. Gray, R.R. Woolaston, B.T. Eaton) pp.15-42. Australian Centre for International Agricultural Research, Canberra.
Russell, J.B. 1998. The importance of pH in the regulation of ruminal acetate to propionate ratio and methane production in vitro. J. Dairy Sci. 81, 3222-3230.
Russell, J.B., Wilson, D.B. 1996. Why are cellulolytic bacteria unable to digest cellulose at low pH? J. Dairy Sci. 79, 1503-1509.
Sauer, F.D., Fellner, V., Kinsman, R., Kramer, J.K.G., Jackson, H.A., Lee, A.J., Chen, S. 1998. Methane output and lactation response in Holstein cattle with monensin or unsaturated fat added in the diet. J. Anim. Sci. 76, 906-914.
SCA 1990. Feeding standards for livestock; ruminants. Standing Committee on Agriculture, Ruminants Sub-Committee. CSIRO, Melbourne, Australia.
Schnell, S., King, G.M. 1995. Stability of methane oxidation capacity to variations in methane and nutrient concentrations. FEMS. Microbiol. Ecol. 4, 285-294.
Shu, Q., Gill, H.S., Hennessy, D.W., Leng, R.A., Bird, S.H., Rowe, J.B. 1999. Immunisation against lactic acidosis in cattle. Res. Vet. Sci. 67, 65-71.
Smuts, M., Meissner, H.H. and P.B. Cronje 1995. Retention time of digesta in the rumen: its repeatability and relationship with wool production of Merino rams. J. Anim. Sci. 73, 206-210.
Slyter, L.L. 1979. Monensin and dichloroacetamide influences on methane and volatile fatty acid production by rumen bacteria in vitro. Appl. Env. Micro. 37, 283-288.
Tahir, I. 1999. Dietary and protozoal effects on the rate and pathways of rumen methanogenesis. Ph.D. Thesis. University of New England, Armidale, Australia.
Teather, R.M., Forster, R J. 1998. Manipulating the rumen microflora with bacteriocins to improve ruminant production. Can. J. Anim. Sci. 78, 57-69.
Thompson, J.M., Barlow, R. 1986. The relationship between feeding and growth parameters and biological efficiency in cattle and sheep. Proc. 3rd World Congress Genet. Appl. Livest. Prod. 2, 271-282.
Valdés, C., Newbold, C.J., Hillman, K., Wallace, R.J. 1997. Los microorganismos metanotrofos como agentes modificadores de la fermentacion ruminal ITEA 18, 157-159
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (14 of 15) [5/21/2003 1:26:26 PM]

Greenhouse Gas Emissions from the Australian Livestock Sector - Chapters 3 to 8
Van Nevel C., Demeyer, D.I. 1977. Effect of monensin on rumen metabolism in-vitro. Appl Env. Micro. 34, 257-257.
Van Nevel, C.J., Demeyer, D.I. 1995. Feed Additives and Other Interventions for Decreasing Methane Emissions. In. Biotechnology and Animal Feeds and Feeding (Eds. R.J. Wallace, A. Chesson) pp 329-349. VCH Publishers., New York.
Van Nevel, C., Demeyer, D.I. 1996. Control of rumen methanogenesis. Environ. Monit. Assess. 42, 73-97.
Widmir, G., Dooley, S. 1995. Phylogenetic analysis of Leishmania RNA virus and Leishmania suggests ancient virus-parasite association. Nucl. Acid. Res.. 23, 2300-2304.
Whitford, M.E., Forster, R.J., Beard, C.E., Gong, J., Teather, R.M. 1997. Phylogenetic analysis of rumen bacteria by comparitive sequence analysis of cloned 16S-rRNA genes. Anaerobe 4, 153-163.
Wolin, M.J., Miller, T.L., Stewart, C.S. 1997. Microbe-microbe interactions. In. The rumen microbial ecosystem, 2nd ed (Eds. P.N. Hobson, C.S. Stewart). pp. 475-491. Blackie Academic and Professional.
Wright, A.D.G., Baker, S.K. 1999. Molecular evolution of the methanogenic archaea inferred from 16S rRNA gene sequences. S. Afr. J. Anim. Sci., 29, 132-134.
Contents > PreviousAGO Home | Contacts | Publications | Search | Site index | What's new | Top
Science & emissions monitoring International activities Government Business & industryCommunity & household Energy & transport Natural resources
Comments Copyright Disclaimer Notice Privacy Accessibility
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html Last modified 17 November 2002
http://www.greenhouse.gov.au/agriculture/livestock/chapters3_8.html (15 of 15) [5/21/2003 1:26:26 PM]