agronomic and physiological effects of grazing on cereal
TRANSCRIPT

Agronomic and physiological effects of grazing on cereal 1
crop growth and grain yields: a review 2 3
4 Matthew T. HarrisonA, Andrew D. MooreA and John R. EvansB 5
6 ACSIRO Plant Industry, GPO Box 1600, Canberra ACT 2601, Australia 7 8 BThe Australian National University, Research School of Biology, Canberra ACT 0200, 9
Australia 10
11
Correspondence: M.T. Harrison. Telephone: +61 2 6246 4892; Fax: +61 2 6246 5166; 12
e-mail: [email protected] 13
14
Date submitted: March 2011 15
Target journal: Crop and Pasture Science 16
Number of tables: 1 17
Number of figures: 1 18
Running title: Effects of grazing on the agronomy and physiology of cereal crops 19
Keywords: Defoliation, herbivory, leaf area, radiation, soil water, transpiration 20
21
22
23
24
25
26
27
28
29
30
31
32
33

Effects of grazing on the agronomy and physiology of cereal crops
2
List of abbreviations 34
35
Symbol Description Units
DM Dry matter g total dry matter m-2 groundDP Dual-purpose -G×E Genotype-by-environment
interaction -
GY Grain yield g grain m-2 groundHI Harvest index g grain DM g-1 SDMIPAR Intercepted PAR MJ PAR m-2 ground d-1
ΣIPAR Cumulative IPAR interception MJ IPAR m-2 ground LAI Leaf area index m2 leaf m-2 groundRUE Radiation-use efficiency g SDM MJ-1 IPARSDM Shoot dry matter g shoot dry matter m-2 groundSLA Specific leaf area m2 leaf g-1 leaf DMTE Transpiration efficiency g DM m-2 ground mm-1 transpiredWSC Water-soluble carbohydrate
concentration or content mg carbohydrate g-1 stem DM or g
carbohydrate m-2 groundWUE Water-use efficiency g DM m-2 ground mm-1 evapotranspirationYP Yield potential g grain m-2 ground

Effects of grazing on the agronomy and physiology of cereal crops
2
Abstract 1
2
The use of cereal crops for the dual purposes (DP) of animal forage and grain production has 3
been practised for over a century. DP cereal crops provide high quality forage during winter 4
when availability of other pastures is often lacking, and high-value grain at maturity. Most DP 5
crop experiments include quantification of biomass production and grain yield (GY) in 6
response to grazing. This review summarises these agronomic data and the reasons cited for 7
the differences in GYs. It then demonstrates how availability of physiological data would lead 8
to greater understanding of crop responses to grazing. 9
10
For a standardised comparison of the effects of defoliation on GY across studies, results from 11
the least intensive defoliation treatments from 33 separate experiments with DP wheat were 12
compiled and compared. Across studies the overall effect of defoliation on GY was –7 ± 25% 13
(mean ± SD, n = 276), with responses ranging from –36% to +75%. In many cases grazing 14
delayed crop phenological development. Delays increased as grazing severity increased or 15
when defoliation was conducted at later phenological stages, particularly after first node 16
appearance, but mechanistic explanations for delayed development were limited. Compared 17
with ungrazed crops, grazed crops were more prostrate, had more decumbent leaves and more 18
tillers, and were shorter at maturity. Physiological measurements of pasture grasses and grain-19
only cereal crops have shown that in general defoliation inhibits root growth, reduces N 20
uptake and carbohydrate production, promotes N remobilisation from roots to leaves, and 21
perturbs normal allocation of dry matter (DM) between roots and shoots. These changes have 22
important implications for canopy light interception, post-grazing growth rates and DM 23
accumulation, and thus assimilate available for retranslocation to grain. Physiological 24
measurements of grazed DP crops are rare but are an essential prerequisite in the development 25
of new dynamic crop-grazing models. Such models could be used to distinguish between the 26

Effects of grazing on the agronomy and physiology of cereal crops
3
effects of different grazing regimes on crop traits such as tillering and root growth, and their 1
respective implications for light interception, temporal soil water use and GY. Analysis of the 2
influence of abiotic variables on biotic variables over multiple seasons would help identify 3
genotype by environment by management interactions. Physiological data from field 4
experiments and application of crop-grazing simulation models should be conducive to (1) 5
better grazing management guidelines, (2) more appropriate trait targeting for breeding of 6
new DP cultivars and (3) better classification of the suitability of a DP cultivar to a given 7
environment. 8

Effects of grazing on the agronomy and physiology of cereal crops
4
Introduction 1
2
The use of cereal crops for the dual purposes (DP) of animal forage and grain production has 3
been practised for over a century (Shelton 1888; Georgeson et al. 1892), occurring in 4
countries such as Argentina (Arzadun et al. 2003), Australia (Kelman and Dove 2009), 5
Canada (Poysa 1985), Italy (Francia et al. 2006), Lebanon (Yau 2003), New Zealand (Scott 6
and Hines 1991), Spain (Royo 1997), Syria (Anderson 1985) and the United States (Winter 7
and Thompson 1987; Redmon et al. 1995). DP cereals include rye (e.g. Holliday 1956), oats 8
(e.g. Bortolini et al. 2005), triticale (e.g. Royo et al. 1994) and barley (e.g. Yau 2003), but the 9
most commonly DP cereal reported in the literature is winter wheat (e.g. Worrell et al. 1992; 10
Redmon et al. 1995; MacKown and Carver 2007). 11
12
DP cereal crops are typically sown in late autumn so that relatively warmer temperatures 13
promote growth and accumulation of large quantities of vegetative biomass by winter 14
(Davidson et al. 1990). Grazing may be commenced as soon as crop roots have sufficient 15
anchorage to prevent plant upheaval during defoliation, though grazing intensities and 16
durations differ from region to region. In the temperate, Mediterranean and semi-arid grazing 17
zones of Australia, Italy, West Asia and Africa, general practice is to graze crops for a single 18
continuous period of several days to weeks at relatively medium to high stocking rates 19
(Anderson 1985; Francia et al. 2006; Kelman and Dove 2009). In contrast standard practice in 20
the Central and Southern Great Plains of the United States is to graze crops for longer 21
durations at lower intensities (e.g. Khalil et al. 2002a; Holman et al. 2009). In most cases 22
grazing is terminated when vegetative development turns reproductive, a developmental stage 23
observed when the apical meristem becomes elevated above the soil surface (growth stage 30, 24
Zadoks et al. 1974). Management of grazing in this manner prevents decapitation of 25
developing spikes and potential loss of GY (Scott and Hines 1991). 26

Effects of grazing on the agronomy and physiology of cereal crops
5
DP cereal crops are a valuable resource and an attractive alternative to farmers for numerous 1
reasons. First, crop forage provides an alternative source of feed during winter when growth 2
of pastures is generally less than livestock requirements (Doole et al. 2009; Moore et al. 3
2009). Second, DP crop forage has high digestibility and crude protein content and is thus 4
conducive to high livestock weight gains (Dove et al. 2002; Dove and McMullen 2009). Third, 5
winter grazing of cereal crops in very productive environments may reduce stubble load and 6
facilitate ease of sowing in the following season (Baumhardt et al. 2009; Nuske et al. 2009). 7
8
As described by Virgona et al. (2006), commercial and scientific interest in DP wheat has 9
fluctuated with the relative profitability of grain and livestock systems and with the release of 10
new crop varieties. Grazing of wheat crops was common during the 1930-70 period (e.g. 11
Swanson 1935; Stansel et al. 1937; Hubbard and Harper 1949) but popularity of DP crops 12
waned with the development of short-season and semi-dwarf cultivars (Pugsley 1983). These 13
varieties were developed as they enabled earlier sowing and tended to lodge less than older, 14
tall varieties (Redmon et al. 1995). However these semi-dwarf cultivars also had (1) shorter 15
coleoptiles and slower growth rates, (2) decreased seedling emergence (Allan 1980), (3) 16
greater developmental rates and therefore (4) earlier stem elongation than older varieties, 17
restricting the time allowed for grazing. Release of new semi-winter DP wheat varieties at the 18
end of the 20th century renewed farmer interest since these crops had high vernalisation 19
requirements (Dove et al. 2002) and potential to meet the high protein grain-milling category 20
(Khalil et al. 2002b; Holman et al. 2009). In 2003 a national program in Australia called 21
‘Grain & Graze’ was introduced to boost the viability and encourage the adoption of mixed-22
farming systems in southern Australia (Price and Hacker 2009; Price et al. 2009).The Grain & 23
Graze program aimed to increase farm profitability by 10% by filling feed gaps arising from 24
seasonal imbalances of forage demand and supply, maintain and improve catchment health, 25
and enhance the pride and social capital of participating farmers (Grain-and-Graze 2003; 26

Effects of grazing on the agronomy and physiology of cereal crops
6
Hacker et al. 2009). By 2008, nearly 3200 farms across nine regions in southern Australia had 1
adopted at least one strategy advocated by Grain & Graze since its outset in 2003 (Price et al. 2
2009). 3
4
Previous studies of DP cereals have focussed on variation in dry matter (DM) or GY with 5
grazing intensity or frequency (Hubbard and Harper 1949; Benjamin et al. 1978; Arzadun et 6
al. 2003), cultivar (Sprague 1954; Lovett and Matheson 1974; Davidson et al. 1990), 7
availability of soil nitrogen (Dann et al. 1983; MacKown and Carver 2007), sowing date 8
(Dann et al. 1977; Yau 2003), yield components (Sharrow and Motazedian 1987) or the 9
economics of meat, wool or grain production (Doole et al. 2009; Zhang 2010). Many studies 10
also document animal performance or in vitro digestibility of grazed cereals (e.g. Dann et al. 11
1983; Kelman and Dove 2007). There is less information on changes that grazing causes to 12
crop phenology and even fewer data on the effects of abiotic factors on physiological 13
mechanisms of crop recovery. These changes are important, since they may hold the key to 14
differences in DM accumulation and GY after grazing. For example, crop regrowth and GY of 15
grazed crops depends on the rate of leaf area development (Winter and Thompson 1987), 16
which in turn is contingent upon availability of soil water (Milroy and Goyne 1995) and soil 17
nitrogen (Gastal and Lemaire 2002). 18
19
This review summarises the agronomic and known physiological effects of grazing on crop 20
growth, phenology and GY. Depending on availability of information, examples in this review 21
relate first to DP wheat crops, second to other DP cereal crops, third to pasture grasses 22
subjected to grazing (effectively grazed cereals before stem elongation) and finally to 23
defoliation experiments performed under controlled conditions. Specifically, the aims of this 24
review are to 25

Effects of grazing on the agronomy and physiology of cereal crops
7
(1) Summarise the reported variation in the effects of grazing on GY and the abiotic reasons 1
ascribed for such variation; 2
(2) Show how a greater understanding of the physiological factors underlying the recovery of 3
DP cereals after grazing would facilitate design of future grazing regimes, breeding of DP 4
cereals and provide a better basis for comparison against ungrazed crops, and 5
(3) Underscore the need for and utility of crop simulation models in DP crop experiments. 6
7
Effects of grazing on grain yield 8
9
Overall effects 10
11
In general GYs decrease with increasing grazing intensities, longer grazing durations 12
(Holliday 1956; Miller et al. 1993) or later end dates of grazing (Stansel et al. 1937; Royo 13
1999). The GY penalty tends to be less if defoliation is completed before stem elongation or 14
appearance of first hollow stem (Swanson 1935; Redmon et al. 1996; Taylor et al. 2010). 15
Phenologically, these crop developmental stages coincide with Zadoks stage 30 (Zadoks et al. 16
1974) or Feekes stage 5.0 (Large 1954). With these factors in mind, the effects of defoliation 17
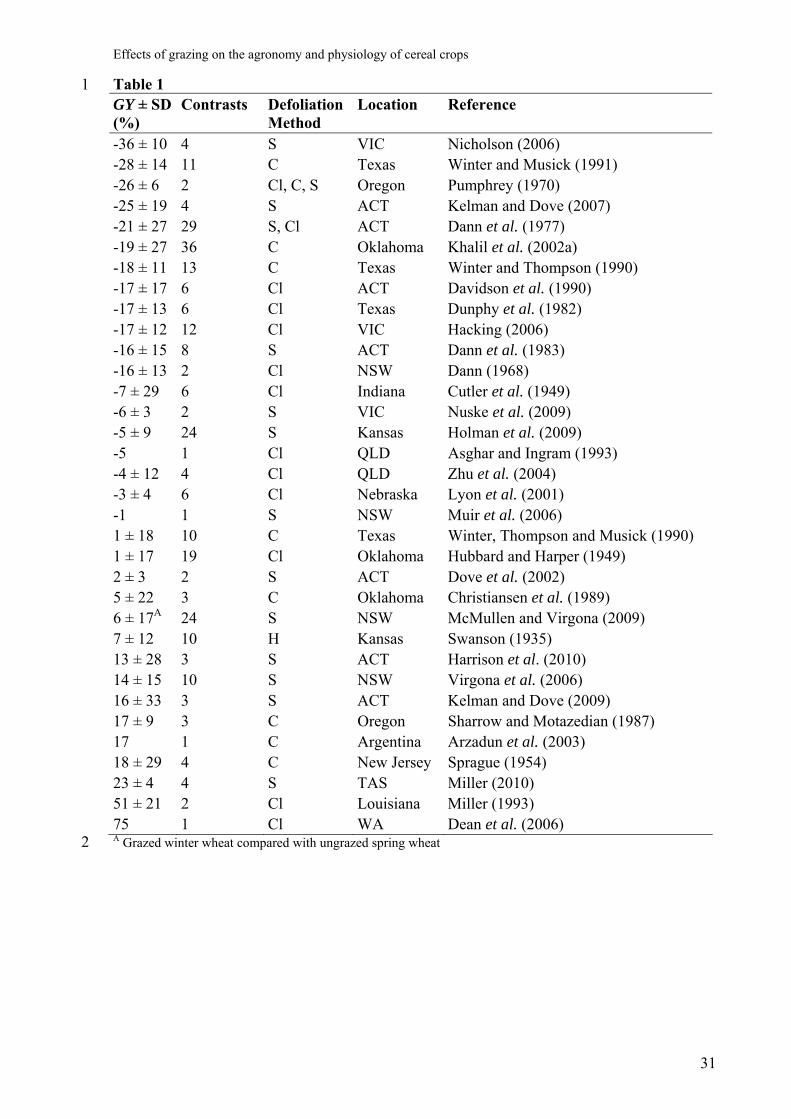
on GY in 34 experiments from a range of locations were reviewed and are shown in Table 1. 18
Studies were selected from published journal papers and agronomy-extension results, and 19
were only included in the survey if experiments (1) were conducted in the field in replicated 20
designs, (2) included an ungrazed control, (3) included valid statistical analyses for treatment 21
differences and (4) provided detailed site conditions including weather data. For consistency 22
across studies, values reported here are from the least intensive or earliest completed 23
treatments of DP wheat crops (n = 276). 24
25

Effects of grazing on the agronomy and physiology of cereal crops
8
Insert Table 1 near here 1
2
Overall, defoliation reduced GY by an average of 7%, with a standard deviation of 25%. 3
Results varied widely both within and across studies, ranging from -36 ± 11% after sheep 4
grazing in Victoria (Hacking 2006) to +75% following clipping near Perth (Dean et al. 2006). 5
6
Differences between the effects of defoliation in Table 1 arise because GY represents the 7
integration of crop processes over the entire growing season, from light interception, 8
radiation-use efficiency (RUE) and DM partitioning to grain filling. Possible approaches to 9
understanding effects of grazing on GY would be to examine each of these components 10
individually, or to use a dynamic model designed to simulate crop grazing. The first of these 11
methods is discussed below, and the second is examined later in this review. 12
13
Mode and rate of defoliation 14
15
Although there were no consistent differences between GYs of treatments subjected to 16
different defoliation methods in Table 1, 12 of the 15 reports of increased yield involved 17
livestock grazing as opposed to clipping. It may be that selective grazing by livestock avoided 18
removal of apical meristems (Virgona et al. 2006), in contrast to clipping treatments, which 19
were generally applied to a uniform crop height and may have removed apical meristems. On 20
the other hand many experiments designed to specifically compare differences between 21
grazing or clipping have shown otherwise. Francia et al. (2006: 163) comment that simulation 22
of grazing by cutting plots was highly comparable for crop growth and production with that 23
of the grazing of sheep. The experiments conducted by Francia et al. (2006) were carried out 24
under ‘crash’ conditions (342 sheep grazed plots over 1 hr), but other grazing studies 25

Effects of grazing on the agronomy and physiology of cereal crops
9
performed for longer periods have also found similar results (e.g. Dann 1968; Pumphrey 1970; 1
Dann et al. 1977; Dann et al. 1983). 2
Primary abiotic and biotic factors influencing the recovery of grazed crops 3
4
Causes ascribed by authors for the effects of grazing on GY in Table 1 are shown in Fig. 1. 5
Soil water content was the most common factor attributed to yield variation, having both 6
positive (e.g. Arzadun et al. 2003) and negative effects (e.g. Dann et al. 1983). Air 7
temperature was cited in 20% of studies and also had beneficial and detrimental effects on GY 8
(Cutler et al. 1949). For example, yield penalties were observed when temperatures were 9
either sub-optimal during winter months, limiting crop growth rates (e.g. Lyon et al. 2001), or 10
supra-optimal at the end of the growing season, inducing premature ripening, damage during 11
pollination or a truncation of the kernel filling duration (Royo and Tribo 1997; Kelman and 12
Dove 2009). Around 7% of studies in Table 1 ascribed yield penalties to crops experiencing 13
frost damage during regrowth or at anthesis (e.g. Davidson et al. 1990). In summary, almost 14
60% of studies in Table 1 attributed yield differences after grazing to soil water availability or 15
air temperature. 16
17
Insert Fig. 1 near here 18
19
In many cases in Table 1, GY of grazed crops increased compared with controls due to 20
reduced lodging (Fig. 1). Lodging was primarily reported in American studies (e.g. Swanson 21
1935; Christiansen et al. 1989) and generally occurred as a result of prolific biomass 22
accumulation in early development. Compared with ungrazed crops, grazed crops generally 23
have less shoot dry matter (SDM) and reduced height at maturity (Winter et al. 1990). These 24
characteristics reduce plant mass and the magnitude of over-turning moments generated by 25
wind (Berry et al. 2003; Sylvester-Bradley and Riffkin 2008), decreasing the likelihood of 26

Effects of grazing on the agronomy and physiology of cereal crops
10
lodging and enabling more spikes to be gathered by machine harvesters at maturity (Winter et 1
al. 1990). 2
3
Mitigation of disease was another reason attributed to increased GY after grazing in the 4
studies reported in Table 1. Dann et al. (1977) observed that grazing reduced the incidence of 5
septoria leaf spot (Septoria tritici), leaf and stem rust (Puccinia triticina, P. graminis 6
respectively), but did not explain why. Miller et al. (1993) commented that damage to 7
unclipped triticale by bacterial streak (Xanthomonas campestris), septoria glume blotch 8
(Stagonospora nodorum) and barley yellow dwarf virus (BYDV, Luteovirus spp.) was rife in 9
seasons with high rainfall. They noted that defoliation reduced leaf damage due to (1) 10
repeated removal of disease inoculum and (2) delayed spike development. It is worth noting 11
here that the chances of disease transmission (e.g. wheat-streak mosaic virus and BYDV) 12
from volunteer cereals and weeds (Hawkes and Jones 2005; Coutts et al. 2008) may be 13
enhanced in DP cereals (compared with grain-only cereals) due to their requirement for 14
earlier sowing. 15
16
Other factors implicated with differences in GY of grazed crops in Table 1 included livestock 17
trampling (Christiansen et al. 1989), smothering of weeds (Davidson et al. 1990), hailstone 18
damage (Lyon et al. 2001), and sub-optimal photoperiod (Royo et al. 1994). Many of these 19
factors were associated with delays in phenological development (see following section). 20
Effects of leaf area and light interception on regrowth and GY are discussed later in this article. 21
22
Effects of defoliation on grain yield components 23
24
Over the 276 treatments in Table 1 there were no consistent effects of defoliation on yield 25
components. Of the studies reporting positive effects on GY, 78% were due to increased 26

Effects of grazing on the agronomy and physiology of cereal crops
11
kernels per unit ground area and the remainder were caused by increased kernel weight. Only 1
the study by Sprague (1954) reported increased GY due to increases in both yield components. 2
Reports of decreased GY after grazing in Table 1 were almost equally attributed to changes in 3
kernel weight (52%) and kernels per unit ground area (48%). 4
5
Effects of defoliation on the number of fertile tillers or spikes per unit ground area at maturity 6
were variable. Del Moral (1992) observed that yields were mainly decreased after cutting due 7
to reduced number of spikes, while El-Shatnawi et al. (2004) found that grazing increased the 8
number of fertile tillers per unit ground area. Many studies also noted that defoliation did not 9
significantly affect the number of fertile spikes per unit ground area or that this yield 10
component was the least affected by defoliation (e.g. Dunphy et al. 1982; Royo 1997; 11
Virgona et al. 2006). 12
13
Yield components are determined temporally, with tiller number first, followed by spikelets 14
per spike, kernels per spikelet and finally kernel weight (Evans et al. 1975). Since the 15
developmental stage of crops during defoliation and the weather conditions following 16
defoliation in the studies above were diverse, it is not surprising that defoliation acted on 17
different yield components, even when the overall effects of defoliation on GY were similar. 18
19
Phenological delays associated with grazing 20
21
Flowering time has been suggested as the single most important factor influencing GY in rain-22
fed environments (Richards 1991), so understanding the effects of grazing on crop 23
development and the timing of flowering will be crucial if yield penalties are to be mitigated. 24
In most cases in Table 1, delayed development after grazing or clipping was associated with 25
decreased GY or reduced grain quality. These effects were particularly evident in experiments 26

Effects of grazing on the agronomy and physiology of cereal crops
12
that were rain-fed or experienced dry finishes to the growing season (Hacking 2006; Virgona 1
et al. 2006). 2
In general later maturity delays harvest and prolongs grain exposure to the environment, 3
increasing chances of weather damage to grain. Amjad et al. (2006) found that grain quality 4
traits such as screenings, fungal staining and falling numbers were a major problem in long-5
season winter wheats when their development was delayed. 6
7
Although the effects of delayed development on GY were primarily negative in studies 8
reported in Table 1, there were some cases when the delay was beneficial. A case in point is 9
that anthesis of grazed crops were less likely to experience late frosts (Dann et al. 1977). 10
Dean et al. (2006) reported large yield losses of unclipped Mackellar wheat due to a frost 11
event that occurred during late spike emergence; in contrast the development of clipped 12
Mackellar was sufficiently delayed so that the effect of frost on GY was relatively small. The 13
outcome of the study by Dean et al. (2006) was that clipped crops yielded around 75% more 14
than those of controls (114 cf. 65 g m-2 respectively). Frost during grain filling may damage 15
developing florets, causing yield penalty and loss of grain quality (Boer et al. 1993; Cromey 16
et al. 1998). Grazed crops were also more capable of using late rainfall (Holliday 1956), and 17
later ripening lowered the susceptibility of pre-harvest grain sprouting (Holman et al. 2009). 18
19
Causes of delayed development 20
21
The question of how defoliation delays crop development in a mechanistic sense remains to 22
be conclusively answered. Some authors postulate that decreased photosynthetic activity 23
reduces the supply of assimilate to the developing apex (Davidson et al. 1990; Virgona et al. 24
2006). Evans (1960) suggested that laminae area remaining after defoliation may be 25
insufficient to generate the long-day stimulus required to accelerate development. Royo et al. 26

Effects of grazing on the agronomy and physiology of cereal crops
13
(1994) attributed delays in anthesis to lower radiation interception and sub-optimal 1
photoperiod. Insufficient day length has also been associated with delayed development in 2
ungrazed crops (Slafer and Rawson 1994) and pasture grasses (Evans 1960). Virgona et al. 3
(2006) commented that the delay may be caused by cooler temperatures surrounding the 4
developing apex, though other studies have revealed that grazing tends to elevate soil 5
temperature maxima (Fahnestock and Knapp 1994) which increases development rates (Rice 6
and Parenti 1978), particularly when apical meristems are close to the soil surface (Stone et al. 7
1999). Various authors have suggested that an increased ratio of red to far-red (R-FR) light at 8
the base of the canopy after defoliation may inhibit the onset of stem elongation (Hofstede et 9
al. 1995; Gautier et al. 1999), but others have observed that exposure of young tissues to 10
increased R-FR light ratios after defoliation tends to advance inflorescence development 11
(Wan and Sosebee 1998). 12
13
Such information indicates that the defoliation-induced phenological delay may be governed 14
by a suite of abiotic variables and that effects of defoliation on crop development cannot 15
simply be generalised across studies. To adequately predict the delay in development caused 16
by grazing and possible consequences for flowering time, further research needs to be carried 17
out on each of the mechanisms discussed above, preferably first as single variates under 18
controlled conditions, then as multivariate systems in the field. 19
20
Effects of grazing on crop water-use efficiency and transpiration efficiency 21
22
Knowledge of grazing regimes that allow crops to better utilise soil water would be 23
advantageous since most DP crops are grown under rain-fed conditions (e.g. Bonachela et al. 24
1995a; Doole et al. 2009). Two indices commonly used to assess crop productivity per unit 25
water used are water-use efficiency (WUE) and transpiration efficiency (TE) (Richards 1991), 26

Effects of grazing on the agronomy and physiology of cereal crops
14
calculated as crop DM produced per unit evapotranspiration or transpiration, respectively 1
(Fischer 1979). The upper limit of WUE for grain of well-managed disease-free water-limited 2
cereal crops is 2.0-2.2 g grain/m2.ground mm (French and Schultz 1984; Sadras and Angus 3
2006). However WUE for grain is often much lower due to stresses caused by weeds, diseases, 4
poor nutrition or inhospitable soils (Passioura 2006), with typical values for rain-fed wheat 5
ranging from 0.53 g/m2.mm in the US Great Plains to 1.0 g/m2.mm in southern Australia 6
(Sadras and Angus 2006). Given the body of literature on WUE and TE in rain-fed crops 7
grown only for grain, computation of these indices for grazed crops would allow useful 8
comparisons between the effectiveness of grain-only and DP systems in terms of biomass 9
production per unit water used. 10
11
Despite their utility, however, published values of WUE and TE of grazed crops are rare, and 12
conclusions from available reports have been unclear. Bonachela et al. (1995b) contrasted the 13
effects of forage removal on the water-use of barley and triticale crops and found no 14
significant differences in evapotranspiration or soil water depletion at maturity between 15
genotypes, sowing date or growing season. In the first of three experiments they showed that 16
clipping increased WUE for DM and for grain by 10 and 26% respectively, yet in the 17
remaining two seasons clipping reduced WUE for DM and for grain by an average of 19 and 18
26% respectively. They attributed differences in WUE to weather conditions and crop water-19
use between seasons. In the first growing season they suggested that a prolonged drought 20
period between the end of clipping and anthesis heightened water stress of unclipped crops. 21
Forage removal conserved soil water and decreased crop water stress. Prolific biomass 22
accumulation at the beginning of a growing season followed by a lack of rainfall leading to 23
premature leaf area senescence generally results in lower GYs for a given vegetative biomass 24
(van Herwaarden 1996). 25
26

Effects of grazing on the agronomy and physiology of cereal crops
15
A generally accepted notion for the conservation of soil water caused by crop grazing is that 1
leaf area removal reduces transpiration and decreases the rate of soil water use (Bonachela et 2
al. 1995b; Virgona et al. 2006; Kelman and Dove 2009). However, other than the study of 3
Bonachela et al. (1995b) there are few reports that provide numerical evidence of concurrent 4
changes to leaf area, SDM and soil water content during and after defoliation. Since the 5
effects of crop defoliation on soil water content in past experiments are inconsistent, ranging 6
from a conservation of soil water (Zhu et al. 2004; Virgona et al. 2006) to using more soil 7
water than undefoliated plots (Kelman and Dove 2009), the notion that grazing conserves soil 8
water should receive further attention. 9
10
In temperate rain-fed environments the rate at which soil water is depleted over the growing 11
season is fundamental to the effects of grazing on WUE and TE. Management of grazing to 12
manipulate the timing of soil water depletion poses an intriguing dilemma, however. On the 13
one hand, WUE is increased the earlier full canopy closure is established (Passioura 2006). 14
Rapid canopy closure reduces water lost to soil evaporation and allows more growth during 15
winter when temperatures are cool and vapour pressure deficits are low (Richards et al. 2002; 16
Dunin and Passioura 2006). On the other hand, conserving soil water before anthesis so that 17
more water is available during grain filling can increase DM partitioned to kernels and 18
ultimately GY (Richards et al. 2002). Studies of temporal soil water extraction by wheat crops 19
have demonstrated that conversion efficiencies of water into grain are high after anthesis, with 20
TE values as high as 5.9 g grain/m2.mm being recorded (Condon et al. 1993; Kirkegaard et al. 21
2007). High TE values after anthesis occur because most post-anthesis assimilate is 22
partitioned directly to grain (Fischer 1979) and pre-anthesis stem assimilates are remobilised 23
to grain (Ehdaie et al. 2006), even though high late season vapour pressure deficits can reduce 24
photosynthesis through stomatal closure, leaf rolling and senescence (Kemanian et al. 2005). 25
26

Effects of grazing on the agronomy and physiology of cereal crops
16
Grazing reduces canopy cover and increases evaporation of water from the soil surface 1
(Murphy et al. 2004). However, since grazing generally delays crop growth (phenological 2
development section) and reduces transpiration, it is possible that grazing may lead to more 3
water available for use at, and after, anthesis. This could enhance GY, but there appears to be 4
only one study that has numerically confirmed this phenomenon (see Bonachela et al. 1995b). 5
Removal of shoot biomass also reduces assimilate supplied to roots (see water-soluble 6
carbohydrate section below), resulting in reduced root biomass and root length density (Trent 7
et al. 1988), and potentially reduced soil water uptake (Garwood and Sinclair 1979). 8
Assimilate translocated to roots increases as plant available water decreases, but decreases 9
with increasing defoliation severity, so defoliation and soil water stress have opposing effects 10
on root growth (Sosebee and Wiebe 1971). In summary, rates of soil water use by grazed 11
crops will depend on (1) rainfall distribution and soil water availability, (2) the severity of 12
defoliation and its affect on assimilate partitioned to roots, and (3) the rate of canopy closure 13
after grazing. It is therefore difficult to predict whether grazing will decrease or increase WUE. 14
Further work is necessary to help identify the trade-offs between soil water conservation and 15
reduced interception of sunlight on growth for different distributions of growing-season 16
rainfall. 17
18
Effects of grazing on canopy light interception, radiation-use efficiency and 19
DM partitioning 20
21
The yield potential (YP) formula (Eqn. 1) can be used to better understand the effects of 22
grazing on crop physiology and its consequences for GY. YP is often defined as the attainable 23
GY of a cultivar grown in environments to which it is adapted, with nutrients and water non-24
limiting, and with pests, diseases, weeds, lodging and other stresses effectively controlled 25
(Evans and Fischer 1999). YP is often expressed as the product of cumulative intercepted 26

Effects of grazing on the agronomy and physiology of cereal crops
17
photosynthetically active radiation, (ΣIPAR, MJ IPAR.m2 ground), radiation-use efficiency 1
(RUE, g DM/MJ IPAR), and harvest index (HI, the ratio of grain DM to SDM at crop 2
maturity), such that 3
4
YP = ΣIPAR × RUE × HI (1) 5
6
Increasing any term on the right-hand side of the identity in Eqn. 1 should increase YP, 7
assuming the other terms remain constant. Eqn. 1 provides a framework of readily measured 8
parameters that can be used to describe how the relative contributions from light interception, 9
RUE and DM partitioning affect YP or GY. The following sub-sections review how changes in 10
crop physiology and morphology affect each component on the right-hand side of Eqn. 1, the 11
effects of grazing on these attributes, and thus the pathways that grazing is likely to influence 12
GY. 13
14
Leaf area indices, canopy light interception and crop growth rates 15
16
Leaf area index (LAI) and the light extinction coefficient of the canopy, k, govern intercepted 17
photosynthetically active radiation (IPAR) (Simpson and Culvenor 1987). LAI and IPAR 18
represent two of the most important physiological parameters underlying crop responses to 19
grazing since they determine crop growth rate (Atwell et al. 1999) and the rate of recovery 20
after defoliation. Around 14% of studies in Table 1 attributed differences in GY of grazed 21
crops to sub-optimal LAI (and by inference IPAR) in the grazed crops (e.g. Swanson 1935; 22
Royo and Pares 1996), as shown in Fig. 1. Winter and Thompson (1987) found that later 23
grazing of wheat limited regrowth of leaf area, biomass accumulation in spring and 24
consequently GY. Ramos et al. (1996) similarly concluded that spring triticale needed to be 25

Effects of grazing on the agronomy and physiology of cereal crops
18
capable of regenerating leaf area rapidly after cutting to prevent GY penalty. Dunphy et al. 1
(1984) also suggested that reduced leaf area of wheat crops at anthesis was strongly correlated 2
with reduced GY. Finally, Winter and Musick (1991) observed that green leaf area removal 3
beyond certain limits could cause abortion of primary culms, reduced growth rate, delayed 4
morphological development and reduced kernel filling duration. In summary these studies 5
have indicated that later and/or more severe defoliation limits leaf area recovery and crop 6
development, and in many cases growth rates following defoliation and GYs. 7
8
The crucial knowledge gap for nearly all DP crop studies is quantification of IPAR. 9
Measurement of IPAR would allow better linkage between changes in LAI and resultant 10
growth rates. Such analyses are already well documented for pasture grasses and have 11
provided good guidelines to grazing management. Simpson and Culvenor (1987) defined the 12
LAI at which maximum growth rate was obtained as the ‘critical LAI’. For best management 13
practice of pasture grasses, Matches (1992) stipulated that (1) LAIs should not be allowed to 14
develop beyond the critical value, (2) LAIs should not be reduced excessively by defoliation, 15
and (3) grazing intensities should be regulated so as to maintain leaf area for maximum 16
growth rates throughout the season (though they conceded that in practice such management 17
may be difficult to achieve). 18
19
Effects of grazing on crop traits that influence cumulative radiation interception (ΣIPAR) 20
21
Specific leaf area 22
23
Many grassland experiments have shown that grazed swards have greater specific leaf area 24
(SLA, leaf area per unit leaf dry mass) compared with ungrazed swards (Rotundo and Aguiar 25
2008; Zhao et al. 2009). Since the product of SLA and green leaf biomass determines LAI and 26

Effects of grazing on the agronomy and physiology of cereal crops
19
ultimately IPAR, it is possible that increased SLA over an extended duration after defoliation 1
may increase ΣIPAR. Richards (2000) indicated that SLA was a primary determinant of early 2
vigour in young wheat plants, as crops with high SLA generally have higher IPAR prior to 3
canopy closure. However, grazing reduces LAI, so the trade-off between decreased IPAR due 4
to leaf area removal and increased IPAR due to greater SLA would largely depend on the rate 5
of canopy closure after grazing and the duration over which SLA was perturbed. 6
7
Canopy architecture 8
9
IPAR is also influenced by light distribution within canopies (Hammer and Wright 1994; 10
Whaley et al. 2000) which depends on leaf angle distribution (Duncan 1971; Campbell 1986), 11
canopy height (Miralles and Slafer 1997; Muurinen and Peltonen-Sainio 2006) and tiller 12
number (Duggan et al. 2005). Grazing can alter any or all of these morphological traits. First, 13
leaf angles may become more decumbent after grazing (Hofstede et al. 1995; de Mello and 14
Pedreira 2004) depending on grazing frequency (Smith 1998). Second, defoliation often 15
reduces canopy height at maturity (Dann 1968; Winter and Thompson 1987) by decreasing 16
the length of the main stem (Royo 1999), resulting in more prostrate canopies (MacKown and 17
Carver 2005). Third, the number of tillers per unit ground area may increase or decrease after 18
grazing (Royo 1999; Kelman and Harrison 2010). 19
20
Green area duration 21
22
Extending the duration with which SDM remains green (the ‘green area duration’) may also 23
increase ΣIPAR. Jenner and Rathjen (1975) and later Borrell et al. (2000) showed that DM 24
accumulation of wheat and sorghum, respectively, were improved by maintaining IPAR 25
during later stages of kernel filling, leading to greater ΣIPAR. Since grazing typically delays 26

Effects of grazing on the agronomy and physiology of cereal crops
20
crop development, grazing may also affect the duration of growth and green area duration in 1
late development (Royo 1997). Under these circumstances it can be postulated that grazing 2
would increase ΣIPAR provided sufficient resources were available. Whether such increases 3
were due to delayed leaf senescence or growth of more leaves and tillers in later development 4
is yet unknown. 5
6
Radiation-use efficiency 7
8
Positive linear correlations are often found between DM and ΣIPAR for crops grown under 9
favourable conditions (Sinclair and Muchow 1999). The slope of this relationship is the RUE 10
(Monteith 1977), generally expressed as g total biomass/MJ IPAR. Since RUE is linked with 11
photosynthetic activity, factors affecting photosynthesis should also alter RUE (Monteith 12
1977; Sinclair and Muchow 1999; Fischer 2007). However, studies of both photosynthesis 13
and RUE in DP cereals are limited and the effects of defoliation on the photosynthesis of 14
grass species vary (e.g. see Nowak and Caldwell 1984; Fahnestock and Detling 2000). 15
Harrison et al. (2010) indicate that there are generally three pathways by which 16
photosynthesis may increase after defoliation. These include (1) improved leaf water status, (2) 17
increased leaf nitrogen content and/or (3) increased activity of photosynthetic enzymes. In one 18
of the few studies examining the effects of grazing on the photosynthesis of DP cereals, 19
Harrison et al. (2010) showed that grazed winter wheat transiently increased photosynthesis 20
by 33-68% over a 2-4 week period after grazing ended. Harrison et al. (2010) attributed 21
changes in photosynthesis to increased Rubisco activity which in turn was associated with a 22
partial alleviation of leaf water stress. 23
24
Other stresses or adverse conditions influencing photosynthesis are also likely to affect RUE. 25
These include water-, temperature- or nitrogen-stresses (Sinclair and Horie 1989; Whaley et 26

Effects of grazing on the agronomy and physiology of cereal crops
21
al. 2000), frost, pests, diseases or senescence (Gallagher and Biscoe 1978; Siddique et al. 1
1989; Gregory et al. 1992), prolonged periods of high vapour pressure deficit (Stockle and 2
Kiniry 1990) and low proportions of diffuse to total solar radiation (Sinclair et al. 1992). 3
4
From the above it is apparent that grazing may have positive (e.g. increased SLA) or negative 5
(e.g. decreased LAI) impacts on ΣIPAR and RUE, which makes the overall effect of 6
defoliation on crop productivity difficult to predict. The physiological parameters discussed 7
above underpin crop growth rate and biomass accumulation, and their quantification in a crop 8
grazing scenario would reveal their relative importance for crop recovery and GY. The other 9
main pathway that grazing may influence GY is through HI (Eqn. 1), which is examined next. 10
11
Effects of defoliation and grazing on dry matter partitioning during reproductive 12
development 13
14
HI may be increased by allocating a greater proportion of SDM to spikes, particularly in the 15
weeks around anthesis (Richards 2000). SDM partitioned to kernels is derived from (1) 16
carbohydrate produced after anthesis and translocated directly to kernels, (2) carbohydrate 17
produced after anthesis, stored temporarily in the stem then retranslocated to kernels, or (3) 18
carbohydrate produced before anthesis, stored mainly in the stem then retranslocated during 19
kernel filling (Ehdaie et al. 2006). Mode 3 depends on the assimilate storage capacity of the 20
crop, thought to be largely a function of stem DM (Austin et al. 1980). Assimilate used during 21
kernel filling in modes 1 and 2 is mainly derived from current photosynthesis and therefore 22
depends on green area duration and activity after anthesis (Welbank et al. 1966; Garcia del 23
Moral 1992). 24
25

Effects of grazing on the agronomy and physiology of cereal crops
22
The extent to which defoliation alters the DM accumulation of individual crop organs, and by 1
implication the size of the assimilate pool available for retranslocation (mode 3), depends on 2
the severity and time of defoliation, as well as the crop species. Noy-Meir and Briske (2002) 3
showed that a single severe clipping of vegetative wild wheat (Triticum dicoccoides) reduced 4
spike and spikelet numbers and total reproductive biomass compared with unclipped wild 5
wheat. A second severe clipping reduced not only survival to reproduction, but also 6
reproductive biomass, GY and grain quality of plants that did become reproductive. 7
Defoliation of barley or triticale crops similarly caused reductions in the DM of leaves, stems 8
and spikes at anthesis relative to unclipped controls (Royo 1999). In this study the 9
contribution of pre-anthesis assimilate to kernels of clipped barley was lower than that of 10
unclipped barley, but the opposite was true for triticale (Royo 1999). 11
12
Defoliation of DP crops has resulted in (1) increased (+58%, Kelman and Dove 2009), (2) no 13
change (Royo et al. 1997) or (3) decreased (-28%, Bonachela et al. 1995a) HI relative to 14
controls. More work is needed to clarify how grazing affects HI. For example HI of grazed 15
crops may be greater than controls due to greater GY, lower SDM or both. Moreover nearly all 16
DP crop studies cite HI values that have been estimated as the ratio of kernel DM to SDM at 17
maturity, as is the convention for grain-only systems. A more informative approach would be 18
to also cite ‘apparent HI’, i.e., to include an estimation of the SDM removed by defoliation 19
since this would demonstrate how above-ground productivity was affected by grazing. 20
Greater understanding of how SDM allocated to developing spikes changes with crop species 21
and with the timing and severity of defoliation should lead to grazing management strategies 22
that mitigate the effects of grazing on GY. 23
24
25
26

Effects of grazing on the agronomy and physiology of cereal crops
23
The need to study water-soluble carbohydrates in grazed cereal crops 1
2
Water-soluble carbohydrates in grazed pastures and grain-only crops 3
4
Concentrations of water-soluble carbohydrates (WSCs) in plant tissues have been extensively 5
studied in pasture grasses (Matches 1992; Fulkerson and Donaghy 2001) and grain-only crops 6
(Shearman et al. 2005) for a number of reasons. First, WSCs relate to a plant’s ability to 7
recover from defoliation. If the duration of grazing is excessive or defoliation is too severe, 8
WSC concentrations may fall to a level that suppresses regrowth and hinders long term 9
persistence (Fulkerson and Donaghy 2001). In ryegrass (Lolium spp.), WSC reserves decline 10
after defoliation as WSCs are used to regrow new shoots, whilst root growth slows and may 11
cease completely (Morvan-Bertrand et al. 1999). It is not until about 75% of a new leaf or 12
tiller has regrown that the plant has enough photosynthetic capacity to support growth and 13
maintenance, and only then does WSC concentrations increase and root growth recommence 14
(Fulkerson and Donaghy 2001). Second, studies of WSC have provided useful guides to 15
grazing management of pasture grasses. Based on relationships between biomass removal, 16
WSCs and regrowth, Fulkerson and Donaghy (2001) suggested that successive grazings of 17
ryegrass should not occur before at least two leaves per tiller were established. If grazing was 18
conducted before the two leaf/tiller stage of regrowth, recovery was severely suppressed. 19
Third, WSCs stored in stems provide a buffer for assimilate supply to kernels when adverse 20
environmental conditions occur after anthesis, so WSCs can potentially have a large influence 21
on final GY (Richards et al. 2010; also see DM partitioning section). Genetic gains in the 22
yield potential of wheat varieties over time have also been associated with increased WSCs in 23
stems and leaf sheaths at anthesis (Shearman et al. 2005). 24
25

Effects of grazing on the agronomy and physiology of cereal crops
24
Water-soluble carbohydrates in DP crops 1
2
Despite extensive studies of WSC dynamics in grazed pastures and grain-only cereals, 3
information on WSC in grazed cereal crops is limited and appears contradictory. Some reports 4
suggest that grazing depletes WSC content (i.e. g/m2), but not WSC concentrations (i.e. g/g), 5
and that the implications of this depletion for GY may be major (Trent et al. 1988). For 6
example Trent et al. (1988) found that total WSC contents in live shoots, leaves and stem 7
bases of grazed wheat were lower than WSC contents measured in ungrazed shoots for about 8
four months after the end of grazing. By physiological maturity, total WSC contents of grazed 9
wheat were similar to those of ungrazed wheat, but the GY of grazed wheat was 16% less than 10
that of the controls. Although other studies have also shown that grazing of wheat crops 11
reduces WSC content of stems at anthesis and does not affect WSC concentrations (average 12
0.38 g g-1), they have also found that effects of grazing on GY were minimal (Muir et al. 13
2006). It is possible that GYs in the study conducted by Muir et al. (2006) were not as 14
dependent on WSCs stored in stems as GYs in the study by Trent et al. (1988). Further 15
investigation of the effect of grazing on DP crop WSCs is required to clarify such 16
discrepancies and to determine how WSC concentrations and contents change temporally, not 17
only to help understand the relationship between regrowth rates and crop persistence, but also 18
to determine how grazing affects WSC accumulation in crop stems and its implications for GY. 19
20
Soil nitrogen uptake and nitrogen metabolism in DP crops 21
22
Grazing or defoliation of pasture grasses may reduce N uptake by roots (Davidson and 23
Milthorp 1966; Thornton and Millard 1997) which in turn (1) reduces plant N supply, (2) 24
causes proteolysis and remobilisation of soluble N compounds from roots and/or stubble to 25

Effects of grazing on the agronomy and physiology of cereal crops
25
establish growth of new laminae (Thornton and Millard 1997), and (3) reduces synthesis of 1
new protein-N compounds (Ourry et al. 1988). Despite the advanced physiological 2
understanding of N metabolism in pasture grasses, knowledge of N-relations in DP crops is 3
still in its infancy. The few available reports focus mainly on N uptake and appear to be 4
inconsistent. Virgona et al. (2006) suggested that N-uptake of grazed crops did not differ from 5
that of ungrazed crops, even though grazing caused a net loss of N (38 kg N ha-1; estimate 6
does not account for recycling of N in urine or faeces). In contrast Thomason et al. (2002) 7
found that more fertiliser N was recovered by forage wheat than grain-only wheat, and Zhu et 8
al. (2006) showed that defoliating wheat at the mid to late tillering stages increased total N 9
removal and N uptake relative to controls (provided adequate fertiliser was applied and crops 10
were sown early). 11
12
Since the majority of soluble leaf N in C3 crops is invested in Rubisco, grazing-induced 13
perturbations to leaf N may affect photosynthesis and therefore have consequences for DM 14
accumulation (see RUE section). Although increases in shoot N concentrations have been 15
observed after defoliation of pasture grasses (Fulkerson and Donaghy 2001), this may reflect 16
a change in SLA or decreasing WSC rather than increasing N content (Davidson and Milthorp 17
1966). Indeed when expressed per unit leaf area, changes in leaf N content after grazing are 18
less frequently observed (Harrison et al. 2010). In one of the few studies examining the 19
changes in leaf N after grazing, Harrison et al. (2010) showed that although grazed crops had 20
significantly elevated leaf photosynthesis, there were no systematic temporal effects of 21
grazing on total leaf N content. 22
23
Because grazed DP crops tend to remain greener than ungrazed crops in later phenological 24
phases (see phenological development section), grazed crops may have increased competition 25
between leaves and grain for N. After anthesis, up to one half of grain N may be sourced from 26

Effects of grazing on the agronomy and physiology of cereal crops
26
retranslocation from leaves and stems (Harper et al. 1987) depending on plant N status and 1
water stress (Borrell and Hammer 2000). If soil N is insufficient to meet N demand of 2
developing kernels, N requirements of kernels are met first through retranslocation from 3
stems and second from leaves (van Oosterom et al. 2010). When the latter occurs, delayed 4
leaf senescence may inhibit leaf N retranslocation, potentially limiting grain protein (Triboi 5
and Triboi-Blondel 2002). Indeed since grazing removes N and delays phenological 6
development it is not surprising that grain protein is often reduced by grazing or defoliation 7
(Asghar and Ingram 1993; Virgona et al. 2006). On the contrary, others have found that 8
defoliation during early vegetative stages has little effect (Francia et al. 2006) or even 9
increases grain protein (Pumphrey 1970). Again these inconsistencies indicate that a more 10
systematic approach is required to disentangle the interactions between phenology, leaf N, N 11
retranslocation and grain protein after crop defoliation. 12
13
Use of models to enhance understanding and forecasting of crop responses 14
to grazing 15
16
Complex interactions and effects of physiological traits in DP systems could be better 17
understood using simulation models 18
19
A crop-grazing simulation model could be used to classify crop traits that are conducive to 20
both forage and grain production, since there are likely to be many traits that are beneficial to 21
one aspect of DP productivity yet detrimental to another (MacKown and Carver 2005). For 22
instance, crops with more erect leaves and vertical growth habits allow a more uniform 23
distribution of light within the canopy, potentially leading to greater GY (see light interception 24
section). However such cultivars are also likely to elevate meristematic zones in early 25

Effects of grazing on the agronomy and physiology of cereal crops
27
development (Noy-Meir and Briske 2002), making apical meristems more prone to 1
decapitation by grazing. In fact pasture grass species most tolerant of grazing are generally 2
those with delayed elevation of meristematic zones (Branson 1953). Crop cultivars with more 3
erect growth habits may also have more rapid accumulation of biomass earlier in the season 4
than more prostrate cultivars, altering the timing of forage availability (MacKown and Carver 5
2005). Effects of other traits (such as tillering and WSC contents) or aspects of DP systems 6
(such as crop soil water use or N status) on the trade-offs between forage and grain production 7
could also be examined using an appropriately designed and calibrated crop-grazing model. 8
9
Simulating grazing of DP crops over multiple seasons and nutritional levels 10
11
A key advantage of a mechanistic crop-grazing model would be the ability to simulate crop 12
responses to defoliation over multiple growing seasons and/or nutritional states. In contrast to 13
one or two years of expensive and labour intensive field experimentation, in silico 14
experiments can be conducted over decades of weather data and multiple nutritional states, to 15
formulate best management practices over the long-term. Such analyses would be beneficial 16
since many physiological processes occurring after grazing are governed by weather, abiotic 17
conditions or endogenous N levels (Fig. 1, N metabolism section, respectively). In fact, 18
abiotic factors often have greater influence on regrowth and GY in DP systems than grazing 19
treatment per se (Christiansen et al. 1989; Brelsford et al. 1998). More recent case studies 20
have suggested that use of forecast-derived decision information to predict differences in 21
forage production and cattle liveweight gains would be a valuable tool for beef producers, 22
provided seasonal forecasts were both timely and strong (Garbrecht et al. 2010). 23
24
Simulation of DP crop responses to grazing over many seasons could also help interpret crop 25
or animal genotype by environment interactions (G×E). In grain-only systems, models have 26

Effects of grazing on the agronomy and physiology of cereal crops
28
been used to investigate three main areas of crop G×E (Chapman 2008). These include (1) 1
using models as ‘environmental integrators’ to interpret G×E observed in the field, (2) model 2
simulation of G×E in multi-environment trials to understand how GY is driven by different 3
combinations of traits, and (3) interpretation of changes in G×E with actual or simulated 4
breeding programs (Chapman 2008). Published model analyses of G×E for grazed pastures 5
are more limited than those of grain-only systems, but are also more diversified. Reports have 6
provided information on, for example, expected livestock phenotypes under different 7
environments (Bryant et al. 2005) or economic outcomes under different stocking rates over 8
many growing seasons (Salmon and Donnelly 2008). Similar analyses conducted for DP 9
cereals would provide information of use to farmers (e.g. risk management associated with 10
grazing different cultivars under different conditions), crop or animal breeders (e.g. suitability 11
of a proposed trait in a target environment, for examples in grain-only systems see Hammer et 12
al. 2005) and policy-makers (e.g. long-term economic outcomes caused by introducing DP 13
crops into new regions). Currently it appears that most analyses of G×E in DP systems are 14
restricted to field experiments over a relatively small number of seasons (e.g. Krenzer et al. 15
1992). 16
17
The need for more dynamic crop-grazing simulation models 18
19
Currently there are only two published models capable of simulating DP crop grazing 20
(Rodriguez et al. 1990; Zhang et al. 2008), but these models have restricted applicability for 21
several reasons. First, both models were designed for winter wheat simulation. Different 22
cultivars and species respond differently to grazing (Redmon et al. 1995; Kelman and Dove 23
2009) so it is unlikely that models developed for winter wheat would produce realistic results 24
for other crop species. Second, both models simulate cattle grazing, but DP crops are also 25
commonly grazed by sheep (e.g. Virgona et al. 2006; Dove and McMullen 2009). Third, 26

Effects of grazing on the agronomy and physiology of cereal crops
29
neither model was calibrated with temporal measurements of LAI, and since key processes 1
such as light interception and transpiration are predominantly driven by LAI, it is crucial that 2
effects of grazing on LAI over the course of the growing season are used in model calibration. 3
Fourth, neither model included the delay in phenological development commonly observed 4
after grazing, which has important ramifications on post-grazing DM accumulation and 5
partitioning, the time-course of LAI and GY (Garcia del Moral 1992; Royo et al. 1999; also 6
see phenological development section). Finally, and most importantly, both models used 7
empirical factors directly linking grazing to observed GY reductions. Such empiricisms 8
severely restrict understanding of crop behaviour after grazing since they blur the distinction 9
between the extent to which GYs are reduced by removal of shoot biomass and inhibition of 10
carbohydrate supply or by removal of apical meristems. Clearly, development of new 11
dynamic crop-grazing simulation models is now required. 12
13
14
Conclusions 15
16
The emphasis of future crop-grazing experiments could profitably shift away from agronomic 17
effects of grazing on GYs towards phenological and physiological recovery of DP crops after 18
grazing. Questions relate to the effects of grazing on leaf angles, SLA, canopy height, tillering 19
potential and rates of canopy closure, since these traits govern light interception, post-grazing 20
growth rates and DM accumulation. It will be important to elucidate the mechanisms 21
underlying delayed phenological development, for crop ontogeny determines the time of 22
anthesis and green area duration after anthesis, both of which strongly affect allocation of 23
assimilate to kernels and ultimately GY. Radiation- and transpiration-use efficiencies are 24
useful indices for contrasting the light- and water-use of grazed crops to those of grain-only 25
crops, yet without field data the complexities implicit in these indices make the effects of 26

Effects of grazing on the agronomy and physiology of cereal crops
30
grazing difficult to project. Grazing reduces leaf area and assimilate supplied to roots, which 1
can inhibit or interrupt root growth, resulting in reduced water and N uptake. Depending on 2
the quantity and frequency of growing season rainfall this may slow soil water use and 3
conserve deep soil water, in turn mitigating terminal crop water stress and increasing the 4
duration of kernel filling. Reduced N uptake caused by grazing has several important 5
consequences in grain-only crops and pasture grasses, ranging from decreased leaf N content 6
and effects on photosynthesis and DM accumulation, changes in forage quality and dry matter 7
digestibility, and increased source-sink competition between leaves and kernels for soluble N 8
compounds after anthesis. Whether such effects occur in grazed cereal crops remains to be 9
seen. Future field experiments should be designed to determine how the severity or frequency 10
of grazing affects the aforementioned physiological variables and thus identify the thresholds 11
with which each variable becomes limiting to productivity. 12
13
Interactions between physiological variables and feedbacks from abiotic factors on post-14
grazing crop recovery are equally important since they often govern whether crop 15
productivity is increased or decreased by grazing. Documented interactions will need to be 16
obtained from field data and from simulations using appropriately calibrated DP crop-grazing 17
models. Information from field data and modelling will further understanding of each other. 18
Simulation models will be a pivotal component for progress in physiological understanding of 19
DP crops as they will allow simulation of grazing systems under multiple seasons and in 20
different environments, hopefully enabling insights into the trade-offs between (1) forage 21
removal and GY, and (2) specific crop traits on recovery rates and GY. Knowledge from field 22
experiments and simulation models would aid risk management planning by farmers, 23
facilitate future breeding of DP crops by scientists and enable better planning of changes in 24
farming enterprises by policy-makers. 25
26
Effects of grazing on the agronomy and physiology of cereal crops
31
Table 1 1 GY ± SD (%)
Contrasts Defoliation Method
Location Reference
-36 ± 10 4 S VIC Nicholson (2006) -28 ± 14 11 C Texas Winter and Musick (1991) -26 ± 6 2 Cl, C, S Oregon Pumphrey (1970) -25 ± 19 4 S ACT Kelman and Dove (2007) -21 ± 27 29 S, Cl ACT Dann et al. (1977) -19 ± 27 36 C Oklahoma Khalil et al. (2002a) -18 ± 11 13 C Texas Winter and Thompson (1990) -17 ± 17 6 Cl ACT Davidson et al. (1990) -17 ± 13 6 Cl Texas Dunphy et al. (1982) -17 ± 12 12 Cl VIC Hacking (2006) -16 ± 15 8 S ACT Dann et al. (1983) -16 ± 13 2 Cl NSW Dann (1968) -7 ± 29 6 Cl Indiana Cutler et al. (1949) -6 ± 3 2 S VIC Nuske et al. (2009) -5 ± 9 24 S Kansas Holman et al. (2009) -5 1 Cl QLD Asghar and Ingram (1993) -4 ± 12 4 Cl QLD Zhu et al. (2004) -3 ± 4 6 Cl Nebraska Lyon et al. (2001) -1 1 S NSW Muir et al. (2006) 1 ± 18 10 C Texas Winter, Thompson and Musick (1990) 1 ± 17 19 Cl Oklahoma Hubbard and Harper (1949) 2 ± 3 2 S ACT Dove et al. (2002) 5 ± 22 3 C Oklahoma Christiansen et al. (1989) 6 ± 17A 24 S NSW McMullen and Virgona (2009) 7 ± 12 10 H Kansas Swanson (1935) 13 ± 28 3 S ACT Harrison et al. (2010) 14 ± 15 10 S NSW Virgona et al. (2006) 16 ± 33 3 S ACT Kelman and Dove (2009) 17 ± 9 3 C Oregon Sharrow and Motazedian (1987) 17 1 C Argentina Arzadun et al. (2003) 18 ± 29 4 C New Jersey Sprague (1954) 23 ± 4 4 S TAS Miller (2010) 51 ± 21 2 Cl Louisiana Miller (1993) 75 1 Cl WA Dean et al. (2006) A Grazed winter wheat compared with ungrazed spring wheat 2

Effects of grazing on the agronomy and physiology of cereal crops
32
Soil watercontent31%
Airtemperature20%
Other 11%
Disease7%
Leaf area14%
Frost damage7%
Lodging11%
1
Fig. 1 2

Effects of grazing on the agronomy and physiology of cereal crops
33
Table 1 1
2
Percentage change in grain yield (GY) of wheat crops relative to undefoliated controls. 3
‘Contrasts’ column shows the number of experiments conducted in each study. Data are mean 4
± one standard deviation (SD) and were calculated for the earliest or least intensive treatments 5
(C = cattle grazing, Cl = mechanical clipping, H = horse grazing and S = sheep grazing). 6
7
Figure 1 8
9
Primary reasons ascribed to differences in grain yield after defoliation as a proportion of the 10
total number of treatments in Table 1. Note that ‘differences’ refer to both positive and 11
negative effects of defoliation (see text for details). ‘Other’ fraction includes livestock 12
trampling, competition with weeds and photoperiod. 13

Effects of grazing on the agronomy and physiology of cereal crops
34
References 1 2 Allan RE (1980) Influence of semi-dwarfism and genetic background on stand establishment 3 of wheat. Crop Science 20(5), 634-638. 4 5 Amjad M, Sharma DL, Curtis B, Anderson WK Dual-purpose long season winter wheats to 6 improve productivity in Western Australia In 'Proceedings of the 13th Australian society of 7 Agronomy Conference', September 2006, Perth. (Eds N Turner and T Acuna), 8 9 Anderson WK (1985) Production of green feed and grain from grazed barley in northern Syria. 10 Field Crops Research 10(1), 57-75. 11 12 Arzadun MJ, Arroquy JI, Laborde HE, Brevedan RE (2003) Grazing pressure on beef and 13 grain production of dual-purpose wheat in Argentina. Agronomy Journal 95(5), 1157-1162. 14 15 Asghar M, Ingram BF (1993) Effects of defoliation on dryland wheat production in central 16 Queensland. Australian Journal of Experimental Agriculture 33(3), 349-351. 17 18 Atwell BJ, Kriedemann PE, Turnbull CGN (1999) Growth analysis: a quantitative approach. 19 In 'Plants in action: adaptation in nature, performance in cultivation.' (Eds BJ Atwell, PE 20 Kriedemann and CGN Turnbull) pp. 186-220. (Macmillan Education Australia: Melbourne) 21 22 Austin RB, Bingham J, Blackwell RD, Evans LT, Ford MA, Morgan CL, Taylor M (1980) 23 Genetic improvements in winter wheat yields since 1900 and associated physiological 24 changes. Journal of Agricultural Science 94(3), 675-689. 25 26 Baumhardt RL, Schwartz RC, Greene LW, MacDonald JC (2009) Cattle gain and crop yield 27 for a dryland wheat-sorghum-fallow rotation. Agronomy Journal 101(1), 150-158. 28 29 Benjamin RW, Chen M, Seligman NG, Wallach D, Hadad MJA (1978) Primary production of 30 grazed annual natural pasture and of grazed wheat in a semi-arid region of Israel. Agricultural 31 Systems 3(3), 205-220. 32 33 Berry PM, Sterling M, Baker CJ, Spink J, Sparkes DL (2003) A calibrated model of wheat 34 lodging compared with field measurements. Agricultural and Forest Meteorology 119(3-4), 35 167-180. 36 37 Boer R, Campbell LC, Fletcher DJ (1993) Characteristics of frost in a major wheat growing 38 region of Australia. Australian Journal of Agricultural Research 44(8), 1731-1743. 39 40 Bonachela S, Orgaz F, Fereres E (1995a) Winter cereals grown for grain and for the dual 41 purpose of forage plus grain. 1. Production. Field Crops Research 44(1), 1-11. 42 43 Bonachela S, Orgaz F, Fereres E (1995b) Winter cereals grown for grain and for the dual 44 purpose of forage plus grain. 2. Water use and water-use efficiency. Field Crops Research 45 44(1), 13-24. 46 47 Borrell AK, Hammer GL (2000) Nitrogen dynamics and the physiological basis of stay-green 48 in sorghum. Crop Science 40(5), 1295-1307. 49 50

Effects of grazing on the agronomy and physiology of cereal crops
35
Bortolini PC, de Moraes A, Carvalho PCD (2005) Forage and grain yield of white oat under 1 grazing. Revista Brasileira De Zootecnia-Brazilian Journal of Animal Science 34(6), 2192-2 2199. 3 4 Branson FA (1953) Two new factors affecting resistance of grasses to grazing. Journal Range 5 Management 6(3), 165-171. 6 7 Brelsford MJ, Peek JM, Murray GA (1998) Effects of grazing by wapiti on winter wheat in 8 northern Idaho. Wildlife Society Bulletin 26(2), 203-208. 9 10 Bryant J, Lopez-Villalobos N, Holmes C, Pryce J (2005) Simulation modelling of dairy cattle 11 performance based on knowledge of genotype, environment and genotype by environment 12 interactions: current status. Agricultural Systems 86(2), 121-143. 13 14 Campbell GS (1986) Extinction coefficients for radiation in plant canopies calculated using 15 an ellipsoidal inclination angle distribution. Agricultural and Forest Meteorology 36(4), 317-16 321. 17 18 Chapman SC (2008) Use of crop models to understand genotype by environment interactions 19 for drought in real-world and simulated plant breeding trials. Euphytica 161(1-2), 195-208. 20 21 Christiansen S, Svejcar T, Phillips WA (1989) Spring and fall cattle grazing effects on 22 components and total grain yield of winter wheat. Agronomy Journal 81(2), 145-150. 23 24 Condon AG, Richards RA, Farquhar GD (1993) Relationships between carbon-isotope 25 discrimination, water-use efficiency and transpiration efficiency for dryland wheat. Australian 26 Journal of Agricultural Research 44(8), 1693-1711. 27 28 Coutts BA, Strickland GR, Kehoe MA, Severtson DL, Jones RAC (2008) The epidemiology 29 of wheat streak mosaic virus in Australia: case histories, gradients, mite vectors, and 30 alternative hosts. Australian Journal of Agricultural Research 59(9), 844-853. 31 32 Cromey MG, Wright DSC, Boddington HJ (1998) Effects of frost during grain filling on 33 wheat yield and grain structure. New Zealand Journal of Crop and Horticultural Science 34 26(4), 279-290. 35 36 Cutler GH, Pavez DS, Mulvey RR (1949) The effect of clipping to simulate pasturing winter 37 wheat on the growth, yield and quality of the crop. Agronomy Journal 41, 169-173. 38 39 Dann PR (1968) Effect of clipping on yield of wheat. Australian Journal of Experimental 40 Agriculture 8(35), 731-735. 41 42 Dann PR, Axelsen A, Dear BS, Williams ER, Edwards CBH (1983) Herbage, grain and 43 animal production from winter grazed cereal crops. Journal of Experimental Agriculture and 44 Animal Husbandry 23(121), 154-161. 45 46 Dann PR, Axelsen A, Edwards CBH (1977) The grain yield of winter grazed crops. 47 Australian Journal of Experimental Agriculture 17(86), 452-461. 48 49 Davidson JL, Jones DB, Christian KR (1990) Winter feed production and grain yield in 50 mixtures of spring and winter wheats. Australian Journal of Agricultural Research 41(1), 1-51 18. 52

Effects of grazing on the agronomy and physiology of cereal crops
36
1 Davidson JL, Milthorp FL (1966) Leaf growth in Dactylis glomerata following defoliation. 2 Annals of Botany 30(118), 173-184. 3 4 de Mello ACL, Pedreira CGS (2004) Morphological responses of irrigated Tanzaniagrass 5 (Panicum maximum jacq. cv. Tanzania-1) to grazing intensity under rotational stocking. 6 Revista Brasileira De Zootecnia-Brazilian Journal of Animal Science 33(2), 282-289. 7 8 Dean G, Davey B, Munford S (2006) 'Effect of fodder cuts on grain yield of early sown wheat 9 and triticale - Perth, Tas. Southern Farming Systems. Available at 10 http://www.sfs.org.au/G&G/2006TrialResults/GG6.htm. [Accessed 12 November 2007].' 11 (Newtown) 12 13 Doole GJ, Bathgate AD, Robertson MJ (2009) Economic value of grazing vegetative wheat 14 (Triticum aestivum L.) crops in mixed-farming systems of Western Australia. Animal 15 Production Science 49(9/10), 807-815. 16 17 Dove H, Holst PJ, Stanley DF, Flint PW (2002) Grazing value of dual-purpose wheats for 18 young sheep. Animal Production in Australia 24, 53-56. 19 20 Dove H, McMullen KG (2009) Diet selection, herbage intake and liveweight gain in young 21 sheep grazing dual-purpose wheats and sheep responses to mineral supplements. Animal 22 Production Science 49(9-10), 749-758. 23 24 Duggan BL, Richards RA, van Herwaarden AF (2005) Agronomic evaluation of a tiller 25 inhibition gene (tin) in wheat. II. Growth and partitioning of assimilate. Australian Journal of 26 Agricultural Research 56(2), 179-186. 27 28 Duncan WG (1971) Leaf angles, leaf area, and canopy photosynthesis. Crop Science 11(4), 29 482-&. 30 31 Dunin F, Passioura J (2006) Prologue: amending agricultural water use to maintain 32 production while affording environmental protection through control of outflow. Australian 33 Journal of Agricultural Research 57(3), 251-255. 34 35 Dunphy DJ, Holt EC, McDaniel ME (1984) Leaf area and dry matter accumulation of wheat 36 following forage removal. Agronomy Journal 76(6), 871-874. 37 38 Dunphy DJ, McDaniel ME, Holt EC (1982) Effect of forage utilization on wheat grain yield. 39 Crop Science 22(1), 106-109. 40 41 Ehdaie B, Alloush GA, Madore MA, Waines JG (2006) Genotypic variation for stem reserves 42 and mobilization in wheat: I. Postanthesis changes in internode dry matter. Crop Science 43 46(2), 735-746. 44 45 El-Shatnawi MKJ, Al-Qurran LZ, Ereifej KI, Saoub HM (2004) Management optimization of 46 dual-purpose barley (Hordeum spontaneum C. Koch) for forage and seed yield. Journal of 47 Range Management 57(2), 197-202. 48 49 Evans LT (1960) Inflorescence initiation in Lolium temulentum L. 1. Effect of plant age and 50 leaf area on sensitivity to photo-periodic induction. Australian Journal of Biological Sciences 51 13(2), 123-31. 52

Effects of grazing on the agronomy and physiology of cereal crops
37
1 Evans LT, Fischer RA (1999) Yield potential: its definition, measurement, and significance. 2 Crop Science 39(6), 1544-1551. 3 4 Evans LT, Wardlaw IF, Fischer RA (1975) Wheat. In 'Crop physiology: some case histories.' 5 Ed. LT Evans) pp. 101-150. (Cambridge University Press: London) 6 7 Fahnestock JT, Detling JK (2000) Morphological and physiological responses of perennial 8 grasses to long-term grazing in the Pryor Mountains, Montana. American Midland Naturalist 9 143(2), 312-320. 10 11 Fahnestock JT, Knapp AK (1994) Plant responses to selective grazing by bison - interactions 12 between light, herbivory and water-stress. Vegetatio 115(2), 123-131. 13 14 Fischer RA (1979) Growth and water limitation to dryland wheat yield in Australia - a 15 physiological framework. Journal of the Australian Institute of Agricultural Science 45(2), 16 83-94. 17 18 Fischer RA (2007) Understanding the physiological basis of yield potential in wheat. Journal 19 of Agricultural Science 145, 99-113. 20 21 Francia E, Pecchioni N, Nicosia OLD, Paoletta G, Taibi L, Franco V, Odoardi M, Stanca AM, 22 Delogu G (2006) Dual-purpose barley and oat in a Mediterranean environment. Field Crops 23 Research 99(2-3), 158-166. 24 25 French RJ, Schultz JE (1984) Water-use efficiency of wheat in a Mediterranean-type 26 environment. 1. The relation between yield, water-use and climate. Australian Journal of 27 Agricultural Research 35(6), 743-764. 28 29 Fulkerson WJ, Donaghy DJ (2001) Plant-soluble carbohydrate reserves and senescence - key 30 criteria for developing an effective grazing management system for ryegrass-based pastures: a 31 review. Australian Journal of Experimental Agriculture 41(2), 261-275. 32 33 Gallagher JN, Biscoe PV (1978) Radiation absorption, growth and yield of cereals. Journal of 34 Agricultural Science 91(AUG), 47-60. 35 36 Garbrecht JD, Zhang XC, Schneider JM, Steiner JL (2010) Utility of seasonal climate 37 forecasts in management of winter-wheat grazing. Applied Engineering in Agriculture 26(5), 38 855-866. 39 40 Garcia del Moral LF (1992) Leaf area, grain yield and yield components following forage 41 removal in triticale. Journal of Agronomy and Crop Science-Zeitschrift Fur Acker Und 42 Pflanzenbau 168(2), 100-107. 43 44 Garwood EA, Sinclair J (1979) Use of water by six grass species. 2. Root distribution and use 45 of soil-water. Journal of Agricultural Science 93(AUG), 25-35. 46 47 Gastal F, Lemaire G (2002) N uptake and distribution in crops: an agronomical and 48 ecophysiological perspective. Journal of Experimental Botany 53(370), 789-799. 49 50

Effects of grazing on the agronomy and physiology of cereal crops
38
Gautier H, Varlet-Grancher C, Hazard L (1999) Tillering responses to the light environment 1 and to defoliation in populations of perennial ryegrass (Lolium perenne L.) selected for 2 contrasting leaf length. Annals of Botany 83(4), 423-429. 3 4 Georgeson CC, Burtis FC, Shelton W (1892) Experiments with wheat. Kansas Agricultural 5 Experiment Station Bulletin 33, 1-50. 6 7 Grain-and-Graze (2003) Grain and graze: profit through knowledge. Australian Government 8 - Land and Water Australia. Available at http://lwa.gov.au/products/pb030542. [Accessed 11 9 July 2010]. 10 11 Gregory PJ, Tennant D, Belford RK (1992) Root and shoot growth, and water and light use 12 efficiency of barley and wheat crops grown on a shallow duplex soil in a Mediterranean-type 13 environment. Australian Journal of Agricultural Research 43(3), 555-573. 14 15 Hacker RB, Robertson MJ, Price RJ, Bowman AM (2009) Evolution of mixed farming 16 systems for the delivery of triple bottom line outcomes: a synthesis of the Grain & Graze 17 program. Animal Production Science 49(9-10), 966-974. 18 19 Hacking C (2006) Dual purpose cereal variety evaluation - Bairnsdale, Vic. Available at 20 http://www.sfs.org.au/G&G/2006TrialResults/GG5.htm [Accessed 14 November 2007]. In. ' 21 (Southern Farming Systems: Newtown) 22 23 Hammer GL, Chapman S, van Oosterom E, Podlich DW (2005) Trait physiology and crop 24 modelling as a framework to link phenotypic complexity to underlying genetic systems. 25 Australian Journal of Agricultural Research 56(9), 947-960. 26 27 Hammer GL, Wright GC (1994) A theoretical analysis of nitrogen and radiation effects on 28 radiation use efficiency in peanut. Australian Journal of Agricultural Research 45(3), 575-29 589. 30 31 Harper LA, Sharpe RR, Langdale GW, Giddens JE (1987) Nitrogen cycling in a wheat crop - 32 soil, plant, and aerial nitrogen transport. Agronomy Journal 79(6), 965-973. 33 34 Harrison MT, Kelman WM, Moore AD, Evans JR (2010) Grazing winter wheat relieves plant 35 water stress and transiently enhances photosynthesis. Functional Plant Biology 37, 726-736. 36 37 Hawkes JR, Jones RAC (2005) Incidence and distribution of Barley yellow dwarf virus and 38 Cereal yellow dwarf virus in over-summering grasses in a Mediterranean-type environment. 39 Australian Journal of Agricultural Research 56(3), 257-270. 40 41 Hofstede RGM, Chilito EJ, Sandovals EM (1995) Vegetative structure, microclimate, and leaf 42 growth of a paramo tussock grass species, in undisturbed, burned and grazed conditions. 43 Vegetatio 119(1), 53-65. 44 45 Holliday R (1956) Fodder production from winter-sown cereal and its effect upon grain yields. 46 Field Crops Abstracts 9, 129-135. 47 48 Holman JD, Thompson CR, Hale RL, Schlegel AJ (2009) Grazing effects on yield and quality 49 of hard red and hard white winter wheat. Agronomy Journal 101(4), 775-788. 50 51

Effects of grazing on the agronomy and physiology of cereal crops
39
Hubbard VC, Harper HJ (1949) Effect of clipping small grains on composition and yield of 1 forage and grain. Agronomy Journal 41(2), 85-92. 2 3 Jenner CF, Rathjen AJ (1975) Factors regulating the accumulation of starch in ripening wheat 4 grain. Australian Journal of Plant Physiology 2(3), 311-322. 5 6 Kelman WM, Dove H (2007) Effects of a spring-sown brassica crop on lamb performance 7 and on subsequent establishment and grain yield of dual-purpose winter wheat and oat crops. 8 Australian Journal of Experimental Agriculture 47(7), 815-824. 9 10 Kelman WM, Dove H (2009) Growth and phenology of winter wheat and oats in a dual-11 purpose management system. Crop & Pasture Science 60(10), 921-932. 12 13 Kelman WM, Harrison MT Interactions between plant density and grazing in cereals under 14 dual-purpose management. In 'Proceedings of the 15th Australian Society of Agronomy 15 Conference', 15-19 November 2010, Lincoln University, New Zealand, 16 17 Kemanian AR, Stockle CO, Huggins DR (2005) Transpiration-use efficiency of barley. 18 Agricultural and Forest Meteorology 130, 1-11. 19 20 Khalil IH, Carver BF, Krenzer EG, MacKown CT, Horn GW (2002a) Genetic trends in winter 21 wheat yield and test weight under dual-purpose and grain-only management systems. Crop 22 Science 42(3), 710-715. 23 24 Khalil IH, Carver BF, Krenzer EG, MacKown CT, Horn GW, Rayas-Duarte P (2002b) 25 Genetic trends in winter wheat grain quality with dual-purpose and grain-only management 26 systems. Crop Science 42(4), 1112-1116. 27 28 Kirkegaard JA, Lilley JM, Howe GN, Graham JM (2007) Impact of subsoil water use on 29 wheat yield. Australian Journal of Agricultural Research 58(4), 303-315. 30 31 Krenzer EG, Thompson JD, Carver BF (1992) Partitioning of genotype x environment 32 interactions of winter wheat forage yield. Crop Science 32(5), 1143-1147. 33 34 Large EC (1954) Growth stages in cereals - illustration of the Feekes scale. Plant Pathology 3, 35 128-129. 36 37 Lovett JV, Matheson EM (1974) Cereals for winter grazing on the northern tablelands of New 38 South Wales. Australian Journal of Experimental Agriculture and Animal Husbandry 14, 39 790-95. 40 41 Lyon DJ, Baltensperger DD, Siles M (2001) Wheat grain and forage yields are affected by 42 planting and harvest dates in the Central Great Plains. Crop Science 41(2), 488-492. 43 44 MacKown CT, Carver BF (2005) Fall forage biomass and nitrogen composition of winter 45 wheat populations selected from grain-only and dual-purpose environments. Crop Science 46 45(1), 322-328. 47 48 MacKown CT, Carver BF (2007) Nitrogen use and biomass distribution in culms of winter 49 wheat populations selected from grain-only and dual-purpose systems. Crop Science 47(1), 50 350-358. 51 52

Effects of grazing on the agronomy and physiology of cereal crops
40
Matches AG (1992) Plant-response to grazing - a review. Journal of Production Agriculture 1 5(1), 1-7. 2 3 McMullen KG, Virgona JM (2009) Dry matter production and grain yield from grazed wheat 4 in southern New South Wales. Animal Production Science 49(9-10), 769-776. 5 6 Miller DR, Dean GJ, Ball PD (2010) Influence of end-grazing forage residual and grazing 7 management on lamb growth performance and crop yield from irrigated dual-purpose winter 8 wheat. Animal Production Science 50, 508-512. 9 10 Miller GL, Joost RE, Harrison SA (1993) Forage and grain yields of wheat and triticale as 11 affected by forage management practices. Crop Science 33(5), 1070-1075. 12 13 Milroy SP, Goyne PJ (1995) Leaf area development in barley - model construction and 14 response to soil moisture status. Australian Journal of Agricultural Research 46(5), 845-860. 15 16 Miralles DJ, Slafer GA (1997) Radiation interception and radiation use efficiency of near-17 isogenic wheat lines with different height. Euphytica 97(2), 201-208. 18 19 Monteith JL (1977) Climate and efficiency of crop production in Britain. Philosophical 20 Transactions of the Royal Society of London Series B-Biological Sciences 281(980), 277-294. 21 22 Moore AD, Bell LW, Revell DK (2009) Feed-gaps in mixed-farming systems: insights from 23 the Grain & Graze program. Animal Production Science 49(10), 736-748. 24 25 Morvan-Bertrand A, Boucaud J, Prud'homme MP (1999) Influence of initial levels of 26 carbohydrates, fructans, nitrogen, and soluble proteins on regrowth of Lolium perenne L-cv. 27 Bravo following defoliation. Journal of Experimental Botany 50(341), 1817-1826. 28 29 Muir CE, Virgona JM, Angus JF Grazing effects on the retranslocation of assimilates during 30 the grain filling of wheat. Available at 31 www.regional.org.au/au/asa/2006/poster/pests/4672_muirc.htm [Accessed 18 November 32 2007]. In 'Proceedings of the 13th Australian Agronomy Conference', 2006, Perth. (Eds NC 33 Turner, T Acuna and RC Johnson), 34 35 Murphy SR, Lodge GM, Harden S (2004) Surface soil water dynamics in pastures in northern 36 New South Wales. 3. Evapotranspiration. Australian Journal of Experimental Agriculture 37 44(6), 571-583. 38 39 Muurinen S, Peltonen-Sainio P (2006) Radiation-use efficiency of modern and old spring 40 cereal cultivars and its response to nitrogen in northern growing conditions. Field Crops 41 Research 96(2-3), 363-373. 42 43 Nicholson C (2006) Effect of grazing on the grain yield and quality of seven cereals - 44 Inverleigh, Vic. In. ' (Southern Farming Systems, Australia: Newtown) 45 46 Nowak RS, Caldwell MM (1984) A test of compensatory photosynthesis in the field - 47 implications for herbivory tolerance. Oecologia 61(3), 311-318. 48 49 Noy-Meir I, Briske DD (2002) Response of wild wheat populations to grazing in 50 Mediterranean grasslands: the relative influence of defoliation, competition, mulch and 51 genotype. Journal of Applied Ecology 39(2), 259-278. 52

Effects of grazing on the agronomy and physiology of cereal crops
41
1 Nuske K, Hunt J, Best F (2009) Grazing cereals. In 'BCG 2009 season research results.' pp. 2 46-51. (Birchip Cropping Group: Melbourne) 3 4 Ourry A, Boucaud J, Salette J (1988) Nitrogen mobilization from stubble and roots during re-5 growth of defoliated perennial ryegrass. Journal of Experimental Botany 39(203), 803-809. 6 7 Passioura J (2006) Increasing crop productivity when water is scarce - from breeding to field 8 management. Agricultural Water Management 80(1-3), 176-196. 9 10 Poysa VW (1985) Effect of forage harvest on grain yield and agronomic performance of 11 winter triticale, wheat and rye. Canadian Journal of Plant Science 65(4), 879-888. 12 13 Price RJ, Hacker RB (2009) Special Issue: Grain & Graze: an innovative triple bottom line 14 approach to collaborative and multidisciplinary mixed-farming systems research, 15 development and extension. Animal Production Science 49(9/10), 729-974. 16 17 Price RJ, Nicholson C, McGuckian N (2009) Contesting targets as a measurement of success 18 in agricultural extension: a case study of the Grain & Graze Change-on-farm strategy. Animal 19 Production Science 49(9/10), 941-955. 20 21 Pugsley AT (1983) The impact of plant physiology on Australian wheat breeding. Euphytica 22 32(3), 743-748. 23 24 Pumphrey FV (1970) Semidwarf winter wheat response to early spring clipping and grazing. 25 Agronomy Journal 62(5), 641-643. 26 27 Ramos JM, Garcia del Moral LF, Boujenna A, Serra J, Insa JA, Royo C (1996) Grain yield, 28 biomass and leaf area of triticale in response to sowing date and cutting stage in three 29 contrasting Mediterranean environments. Journal of Agricultural Science 126, 253-258. 30 31 Redmon LA, Horn GW, Krenzer EG, Bernardo DJ (1995) A review of livestock grazing and 32 wheat grain yield - boom or bust. Agronomy Journal 87(2), 137-147. 33 34 Redmon LA, Krenzer EG, Bernardo DJ, Horn GW (1996) Effect of wheat morphological 35 stage at grazing termination on economic return. Agronomy Journal 88(1), 94-97. 36 37 Rice EL, Parenti RL (1978) Causes of decreases in productivity in undisturbed tall grass 38 prairie. American Journal of Botany 65(10), 1091-1097. 39 40 Richards RA (1991) Crop improvement for temperate Australia - future opportunities. Field 41 Crops Research 26(2), 141-169. 42 43 Richards RA (2000) Selectable traits to increase crop photosynthesis and yield of grain crops. 44 Journal of Experimental Botany 51, 447-458. 45 46 Richards RA, Rebetzke GJ, Condon AG, van Herwaarden AF (2002) Breeding opportunities 47 for increasing the efficiency of water use and crop yield in temperate cereals. Crop Science 48 42(1), 111-121. 49 50

Effects of grazing on the agronomy and physiology of cereal crops
42
Richards RA, Rebetzke GJ, Watt M, Condon AG, Spielmeyer W, Dolferus R (2010) Breeding 1 for improved water productivity in temperate cereals: phenotyping, quantitative trait loci, 2 markers and the selection environment. Functional Plant Biology 37(2), 85-97. 3 4 Rodriguez A, Trapp JN, Walker OL, Bernardo DJ (1990) A wheat grazing systems model for 5 the United States southern plains. 1. Model description and performance. Agricultural 6 Systems 33(1), 41-59. 7 8 Rotundo JL, Aguiar MR (2008) Herbivory resistance traits in populations of Poa ligularis 9 subjected to historically different sheep grazing pressure in Patagonia. Plant Ecology 194, 10 121-133. 11 12 Royo C (1997) Grain yield and yield components as affected by forage removal in winter and 13 spring triticale. Grass and Forage Science 52(1), 63-72. 14 15 Royo C (1999) Plant recovery and grain yield formation in barley and triticale following 16 forage removal at two cutting stages. Journal of Agronomy and Crop Science 182(3), 175-183. 17 18 Royo C, Insa JA, Boujenna A, Ramos JM, Montesinos E, Garcia del Moral LF (1994) Yield 19 and quality of spring triticale used for forage and grain as influenced by sowing date and 20 cutting stage. Field Crops Research 37(3), 161-168. 21 22 Royo C, Lopez A, Serra J, Tribo F (1997) Effect of sowing date and cutting stage on yield 23 and quality of irrigated barley and triticale used for forage and grain. Journal of Agronomy 24 and Crop Science 179(4), 227-234. 25 26 Royo C, Pares D (1996) Yield and quality of winter and spring triticales for forage and grain. 27 Grass and Forage Science 51(4), 449-455. 28 29 Royo C, Tribo F (1997) Triticale and barley for grain and for dual-purpose (forage plus grain) 30 in a Mediterranean-type environment. 2. Yield, yield components, and quality. Australian 31 Journal of Agricultural Research 48(4), 423-432. 32 33 Royo C, Voltas J, Romagosa I (1999) Remobilization of pre-anthesis assimilates to the grain 34 for grain only and dual-purpose (forage and grain) triticale. Agronomy Journal 91(2), 312-316. 35 36 Sadras VO, Angus JF (2006) Benchmarking water-use efficiency of rainfed wheat in dry 37 environments. Australian Journal of Agricultural Research 57(8), 847-856. 38 39 Salmon L, Donnelly JR (2008) Using grazing systems models to evaluate business options for 40 fattening dairy bulls in a region with a highly variable feed supply. Animal Feed Science and 41 Technology 143(1-4), 296-313. 42 43 Scott WR, Hines SE (1991) Effects of grazing on grain yield of winter barley and triticale - 44 the position of the apical dome relative to the soil surface. New Zealand Journal of 45 Agricultural Research 34(2), 177-184. 46 47 Sharrow SH, Motazedian I (1987) Spring grazing effects on components of winter-wheat 48 yield. Agronomy Journal 79(3), 502-504. 49 50 Shearman VJ, Sylvester-Bradley R, Scott RK, Foulkes MJ (2005) Physiological processes 51 associated with wheat yield progress in the UK. Crop Science 45(1), 175-185. 52

Effects of grazing on the agronomy and physiology of cereal crops
43
1 Shelton EM (1888) Experiments with wheat. Kansas Agricultural Experiment Station Bulletin 2 4, 15-26. 3 4 Siddique KHM, Belford RK, Perry MW, Tennant D (1989) Growth, development and light 5 interception of old and modern wheat cultivars in a Mediterranean-type environment. 6 Australian Journal of Agricultural Research 40(3), 473-487. 7 8 Simpson RJ, Culvenor RA (1987) Photosynthesis, carbon partitioning and herbage yield. In 9 'Temperate pastures - their production, use and management.' (Eds JL Wheeler, CJ Pearson 10 and GE Robards) pp. 103-118. (Australian Wool Corporation Technical Publication: 11 Melbourne) 12 13 Sinclair TR, Horie T (1989) Leaf nitrogen, photosynthesis, and crop radiation use efficiency - 14 a review. Crop Science 29(1), 90-98. 15 16 Sinclair TR, Muchow RC (1999) Radiation use efficiency. Advances in Agronomy, Vol 65 65, 17 215-265. 18 19 Sinclair TR, Shiraiwa T, Hammer GL (1992) Variation in crop radiation-use efficiency with 20 increased diffuse radiation. Crop Science 32(5), 1281-1284. 21 22 Slafer GA, Rawson HM (1994) Sensitivity of wheat phasic development to major 23 environmental factors - a reexamination of some assumptions made by physiologists and 24 modelers. Australian Journal of Plant Physiology 21(4), 393-426. 25 26 Smith SE (1998) Variation in response to defoliation between populations of Bouteloua 27 curtipendula var. caespitosa (Poaceae) with different livestock grazing histories. American 28 Journal of Botany 85(9), 1266-1272. 29 30 Sosebee RE, Wiebe HH (1971) Effect of water stress and clipping on photosynthate 31 translocation in two grasses. Agronomy Journal 63(1), 14-19. 32 33 Sprague MA (1954) The effect of grazing management on forage and grain production from 34 rye, wheat and oats. Agronomy Journal 46(1), 29-33. 35 36 Stansel RH, Dunkle PB, Jones DL (1937) Small grains and rye grass for winter pasture. Texas 37 Agricultural Experiment Station Bulletin 539, 1-38. 38 39 Stockle CO, Kiniry JR (1990) Variability in crop radiation-use efficiency associated with 40 vapor-pressure deficit. Field Crops Research 25(3-4), 171-181. 41 42 Stone PJ, Sorensen IB, Jamieson PD (1999) Effect of soil temperature on phenology, canopy 43 development, biomass and yield of maize in a cool-temperate climate. Field Crops Research 44 63(2), 169-178. 45 46 Swanson AF (1935) Pasturing winter wheat in Kansas. Kansas Agricultural Experiment 47 Station Bulletin 271. 48 49 Sylvester-Bradley R, Riffkin PA (2008) Designing resource-efficient ideotypes for new 50 cropping conditions: wheat in the UK and Australasia. Aspects of Applied Biology(88), 127-51 133. 52

Effects of grazing on the agronomy and physiology of cereal crops
44
1 Taylor KW, Epplin FM, Brorsen BW, Fieser BG, Horn GW (2010) Optimal grazing 2 termination date for dual-purpose winter wheat production. Journal of Agricultural and 3 Applied Economics 42(1), 87-103. 4 5 Thomason WE, Raun WR, Johnson GV, Freeman KW, Wynn KJ, Mullen RW (2002) 6 Production system techniques to increase nitrogen use efficiency in winter wheat. Journal of 7 Plant Nutrition 25(10), 2261-2283. 8 9 Thornton B, Millard P (1997) Increased defoliation frequency depletes remobilization of 10 nitrogen for leaf growth in grasses. Annals of Botany 80(1), 89-95. 11 12 Trent JD, Wallace LL, Svejcar TJ, Christiansen S (1988) Effect of grazing on growth, 13 carbohydrate pools, and mycorrhizae in winter wheat. Canadian Journal of Plant Science 14 68(1), 115-120. 15 16 Triboi E, Triboi-Blondel AM (2002) Productivity and grain or seed composition: a new 17 approach to an old problem - invited paper. European Journal of Agronomy 16(3), 163-186. 18 19 van Herwaarden AF (1996) Haying-off in wheat: enduring myth or current problem? 20 Proceedings of the 8th Australian Agronomy Conference, Toowoomba, Queensland, Australia, 21 30 January-2 February, 1996., 566-569. 22 23 van Oosterom EJ, Chapman SC, Borrell AK, Broad IJ, Hammer GL (2010) Functional 24 dynamics of the nitrogen balance of sorghum. II. Grain filling period. Field Crops Research 25 115(1), 29-38. 26 27 Virgona JM, Gummer FAJ, Angus JF (2006) Effects of grazing on wheat growth, yield, 28 development, water use, and nitrogen use. Australian Journal of Agricultural Research 57(12), 29 1307-1319. 30 31 Wan CG, Sosebee RE (1998) Tillering responses to red: far-red light ratio during different 32 phenological stages in Eragrostis curvula. Environmental and Experimental Botany 40(3), 33 247-254. 34 35 Welbank PJ, French SAW, Witts KJ (1966) Dependence of yields of wheat varieties on their 36 leaf area durations. Annals of Botany 30(118), 291-299. 37 38 Whaley JM, Sparkes DL, Foulkes MJ, Spink JH, Semere T, Scott RK (2000) The 39 physiological response of winter wheat to reductions in plant density. Annals of Applied 40 Biology 137(2), 165-177. 41 42 Winter SR, Musick JT (1991) Grazed wheat grain yield relationships. Agronomy Journal 43 83(1), 130-135. 44 45 Winter SR, Thompson EK (1987) Grazing duration effects on wheat growth and grain yield. 46 Agronomy Journal 79(1), 110-114. 47 48 Winter SR, Thompson EK (1990) Grazing winter wheat. I. Response of semidwarf cultivars 49 to grain and grazed production systems. Agronomy Journal 82(1), 33-37. 50 51

Effects of grazing on the agronomy and physiology of cereal crops
45
Winter SR, Thompson EK, Musick JT (1990) Grazing winter wheat. II. Height effects on 1 response to production systems. Agronomy Journal 82(1), 37-41. 2 3 Worrell MA, Undersander DJ, Khalilian A (1992) Grazing wheat to different morphological 4 stages for effects on grain yield and soil compaction. Journal of Production Agriculture 5(1), 5 81-85. 6 7 Yau SK (2003) Yields of early planted barley after clipping or grazing in a semiarid area. 8 Agronomy Journal 95(4), 821-827. 9 10 Zadoks JC, Chang TT, Konzak CF (1974) Decimal code for growth stages of cereals. Weed 11 Research 14(6), 415-421. 12 13 Zhang XC (2010) Optimizing stocking rate for maximum return to a wheat-cattle enterprise 14 using model simulation and economics. Agronomy Journal 102(1), 197-209. 15 16 Zhang XC, Phillips WA, Garbrecht JD, Steiner JL, Hunt LA (2008) A wheat grazing model 17 for simulating grain and beef production: Part I - model development. Agronomy Journal 18 100(5), 1242-1247. 19 20 Zhao W, Chen SP, Han XG, Lin GH (2009) Effects of long-term grazing on the 21 morphological and functional traits of Leymus chinensis in the semiarid grassland of Inner 22 Mongolia, China. Ecological Research 24(1), 99-108. 23 24 Zhu GX, Midmore DJ, Radford BJ, Yule DF (2004) Effect of timing of defoliation on wheat 25 (Triticum aestivum) in central Queensland. 1. Crop response and yield. Field Crops Research 26 88(2-3), 211-226. 27 28 Zhu GX, Midmore DJ, Yule DF, Radford BJ (2006) Effect of timing of defoliation on wheat 29 (Triticum aestivum) in central Queensland. 2. N uptake and relative N use efficiency. Field 30 Crops Research 96(1), 160-167. 31 32 33 34