allometric scaling and taxonomic variation in nutrient ...preston.kbs.msu.edu/reprints/files/edwards...
TRANSCRIPT

Allometric scaling and taxonomic variation in nutrient utilization traits and maximum
growth rate of phytoplankton
Kyle F. Edwards,a,* Mridul K. Thomas,a,b Christopher A. Klausmeier,a,c and Elena Litchman a,b
a Kellogg Biological Station, Michigan State University, Hickory Corners, MichiganbDepartment of Zoology, Michigan State University, Hickory Corners, Michiganc Department of Plant Biology, Michigan State University, Hickory Corners, Michigan
Abstract
Nutrient utilization traits can be used to link the ecophysiology of phytoplankton to population dynamicmodels and the structure of communities across environmental gradients. Here we analyze a comprehensiveliterature compilation of four traits: maximum nutrient uptake rate; the half-saturation constant for nutrientuptake; the minimum subsistence quota, measured for nitrate and phosphate; and maximum growth rate. We alsouse these traits to analyze two composite traits, uptake affinity and scaled uptake affinity. All traits tend toincrease with cell volume, except for scaled uptake affinity and maximum growth rate, which tend to decline withcell volume. Most scaling relationships are the same for freshwater and marine species, although importantdifferences exist. Most traits differ on average between major taxa, but between-taxon variation is nearly alwaysdue to between-taxon variation in volume. There is some evidence for between-trait correlations that couldconstrain trait evolution, but these correlations are difficult to disentangle from correlation driven by cell volume.These results should enhance the parameterization of models that use size or taxonomic group to structurephysiological variation in phytoplankton communities.
Nutrient utilization traits have long been used to link theecophysiology of phytoplankton to competitive interactionsand the structure of communities across environmentalgradients (Eppley et al. 1969; Tilman 1982). Such traits canbe used to parameterize models that predict populationdynamics and competitive outcomes in the laboratory(Grover 1991), implying that trait-based models may allowa mechanistic understanding of natural dynamics anddistributions (Litchman and Klausmeier 2008). Many traitshave been shown to scale allometrically with cell volume, suchas maximum nutrient uptake rate, the half-saturationconstant for uptake, subsistence quotas, and uptake affinity(Shuter 1978; Grover 1989; Litchman et al. 2007; Tambi et al.2009). Such scaling relationships allow the parameterizationof models that predict how optimal size, or community sizestructure, should vary with the nutrient supply regime. Forexample, the relative abundance of large cells is predicted toincrease with increasing nitrogen supply (Irwin et al. 2006),and nitrogen supplied in pulses may allow large cells tooutcompete small cells (Stolte and Riegman 1996; Litchmanet al. 2009). Likewise, major phytoplankton taxa often differin average trait values (Smayda 1997; Litchman et al. 2007),and this can permit a more mechanistic understanding ofpatterns in taxonomic structure and the biogeochemicaleffects of taxonomic variation. For example, annual fluctu-ation in the relative abundance of marine diatoms, cocco-lithophores, and prasinophytes can be predicted from a modelthat incorporates taxonomic differences in nutrient and lightutilization traits (Litchman et al. 2006); and predictedseasonal patterns of oceanic CO2 uptake are altered bythe inclusion of a separate functional group representingcoccolithophores (Signorini et al. unpubl.).
In order to better quantify interspecific variation innutrient utilization traits, we have comprehensively com-piled trait data from published studies. Our compilationincludes nitrate and phosphate utilization traits of bothfreshwater and marine species, as nitrogen and phosphorusare the main limiting nutrients in both environments(Hecky and Kilham 1988; Elser et al. 2007). We have alsocompiled maximum growth rates for a large number offreshwater and marine species. We use this compilationto ask a series of questions: (1) what are the power-lawexponents that describe how each trait scales with volume;(2) do the exponents differ between marine and freshwaterspecies; (3) do average trait values differ between taxa;(4) do between-taxon differences exist when controllingfor between-taxon variation in cell volume; (5) how doempirical power-law exponents compare to theoreticalpredictions; and (6) is there evidence for trade-offs amongtraits that would constrain the evolution of competitiveability. Many of these questions have been addressedpreviously, in separate analyses using smaller datasets or asubset of traits (Banse 1976; Shuter 1978; Grover 1989;Tang 1995; Smayda 1997; Litchman et al. 2007). In thisstudy we synoptically address these questions using athorough collection of currently available data.
Methods
Trait compilation—We comprehensively searched theliterature for traits related to nutrient uptake and nutrient-limited growth. Specifically, we collected parameters of theMichaelis-Menten model of nutrient uptake,
Uptake~VmaxR
KzRð1Þ
* Corresponding author: [email protected]
Limnol. Oceanogr., 57(2), 2012, 554–566
E 2012, by the Association for the Sciences of Limnology and Oceanography, Inc.doi:10.4319/lo.2012.57.2.0554
554

and the Droop model of phytoplankton growth,
Net growth~m? 1{Qmin
Q
� �{m ð2Þ
where Vmax is the maximum cell-specific nutrient uptakerate (mmol nutrient cell21 d21), K is the half-saturationconstant for nutrient uptake (mmol nutrient L21), R is theexternal nutrient concentration (mmol nutrient L21), m‘ isthe specific growth rate at infinite quota (d21), Q is theinternal nutrient concentration or quota (mmol nutrientcell21), Qmin is the minimum quota at which growth rateequals zero, and m is the specific mortality rate (d21).
Here we focused on the parameters Vmax, K, and Qmin,measured for nitrate and phosphate. We denoted thenitrate parameters as V N
max, KN, and Q Nmin, and the
phosphate parameters as V Pmax, KP, and Q P
min. Wecompared nitrate but not ammonium parameters becauseammonium parameters have been measured for manyfewer species. We used studies in which temperature was ator near 20uC, and light was not severely limiting. It ispossible that allometric relationships and taxonomicdifferences involving these parameters could vary withtemperature, but data on the temperature dependence ofthese parameters are limited, and using studies performedclose to 20uC allowed us to make consistent interspecificcomparisons. The maximum rate of uptake (Vmax) oftendeclines as cellular nutrient content increases (Morel 1987),and therefore we included only estimates of Vmax measuredunder conditions of intracellular nutrient depletion. Weincluded estimates of uptake kinetics both from studiesusing isotopic methods, and studies using the rate ofdepletion of filtrate nutrient concentration. Cell volumeswere determined from the literature, if not measured in thefocal studies. Species and traits present in the dataset arelisted in the Web Appendix (Table A1, www.aslo.org/lo/toc/vol/vol_57/issue_2/0554a.html); the dataset included 64marine species and 59 freshwater species. If the same traitwas measured on multiple occasions for a species, we usedthe mean trait value in our analysis.
Uptake affinity, or Vmax/K, is often used as an indicatorof nutrient uptake ability at limiting concentrations,because it quantifies the clearance rate of nutrients as theexternal nutrient concentration becomes close to zero(Healey 1980). In order to predict species’ relative abilitiesto grow under nutrient limitation, it may be more useful tomeasure the uptake affinity scaled by the minimum nutrientquota, or Vmax/(K 3 Qmin) (Tambi et al. 2009). Thisquantity measures the clearance rate of nutrients, relativeto the amount of nutrient needed to grow, when externalnutrient concentration becomes close to zero. Furthermore,it can be shown that under Eqs. 1, 2, as m R 0, for specieswith equal mortality rates, the winner in competition atequilibrium is the species with the greater scaled uptakeaffinity (Litchman et al. 2007; Edwards et al. 2011).Therefore, we quantified allometric relationships andbetween-taxon differences for both uptake affinity andscaled uptake affinity, for nitrate and phosphate; we willrefer to these as N affinity, P affinity, scaled N affinity, andscaled P affinity. We also tested whether scaled uptake
affinity is an effective predictor of the outcome ofcompetition by comparing the outcomes of published P-limited chemostat experiments to the outcomes predictedby those species’ scaled uptake affinities. We found thatscaled P affinity correctly predicted the winner in compe-tition in 10 out of 13 pairwise comparisons, in spite of thefact that dilution rates varied from 0.07 to 0.5 d21 (see WebAppendix, Table A1). We therefore consider scaled uptakeaffinity to be a useful proxy for equilibrium competitiveability under nutrient limitation. We refer to uptake affinityand scaled uptake affinity as ‘‘composite’’ nutrient traits,similar to R*, defined as the break-even nutrient concen-tration at which growth rate equals mortality rate (Tilman1982). We refer to Vmax, K, and Qmin as ‘‘primary’’ nutrienttraits.
We also compiled estimates of maximum growth rate,mmax, for 105 marine species and 124 freshwater species (seeWeb Appendix, Table A1). These estimates were compiledfrom studies of nutrient-limited growth, light-limitedgrowth, and maximum growth rate as a function oftemperature. We compiled maximum achievable growthrates, rather than m‘ from the Droop model (Eq. 2),because measurements of mmax were available for morespecies. If only m‘ was available, when possible weconverted to mmax using the equation mmax 5 m‘ 3 (Qmax
2 Qmin)/Qmax where Qmax is the maximum possible nutrientquota, which is obtained when m 5 mmax (Morel 1987). Allestimates of mmax were measured at or near 20uC. Marinespecies for which we have maximum growth rate data rangein volume from 4.6 3 1021 to 1.6 3 107 mm3, and thefreshwater species range in volume from 1.8 to 3.8 3104 mm3.
Statistical analyses—In order to quantify scaling rela-tionships between nutrient traits and cell volume, we usedstandardized major axis (SMA) regression (also referred toas reduced major axis regression). This method is moreappropriate than least-squares regression for estimating theline of best fit for the relationship between two variables(Warton et al. 2006). SMA slopes were estimated only if thetwo variables had a significant (at p , 0.05) Kendall rank-correlation coefficient. Analysis was performed with the‘‘smatr’’ package in R version 2.11.0 (Warton and Ormerod2007). Relationships between nutrient utilization traits andcell volume are typically linear when both variables are log-transformed, resulting in a power-law relationship on thelinear scale (Litchman et al. 2007); we therefore performedall analyses on log10-transformed values.
We tested for between-taxon trait differences usinganalysis of variance; we tested for between-taxon traitdifferences while controlling for the effect of volume byperforming analysis of variance and post hoc Tukeypairwise comparisons with the residuals from the SMAregressions. For taxonomic comparisons, species wereclassified into the following groups: diatoms, dinoflagel-lates, cyanobacteria, desmids, non-desmid chlorophytes,haptophytes, and raphidophytes. Freshwater and marinespecies were analyzed separately, because of differences intaxon representation; also, for each trait we excluded fromthe analysis those taxa for which the trait was measured on
Reviews in L&O 555
fewer than two species. Therefore, the following groupswere compared for freshwater species: cyanobacteria,diatoms, desmids, and non-desmid chlorophytes (desmidswere only present for the phosphate traits). For the marinespecies, the following groups were compared: diatoms,dinoflagellates, chlorophytes, haptophytes, and raphido-phytes. We also used t-tests to compare mean trait valuesfor freshwater and marine diatoms, because diatoms fromboth environments were relatively well represented.
We tested for correlations between the primary nutrientutilization traits, which could indicate physiological traitconstraints or correlated selection pressures. Because mosttraits are correlated with volume, we also tested for partialcorrelations that controlled for the combined effect ofvolume on each pair of traits. Correlations between thecomposite traits are the subject of a separate manuscript, inwhich we use techniques for multivariate analysis in thepresence of missing data to better estimate the correlationsbetween cell volume, uptake affinity, and scaled uptakeaffinity for N and P (Edwards et al. 2011).
Results
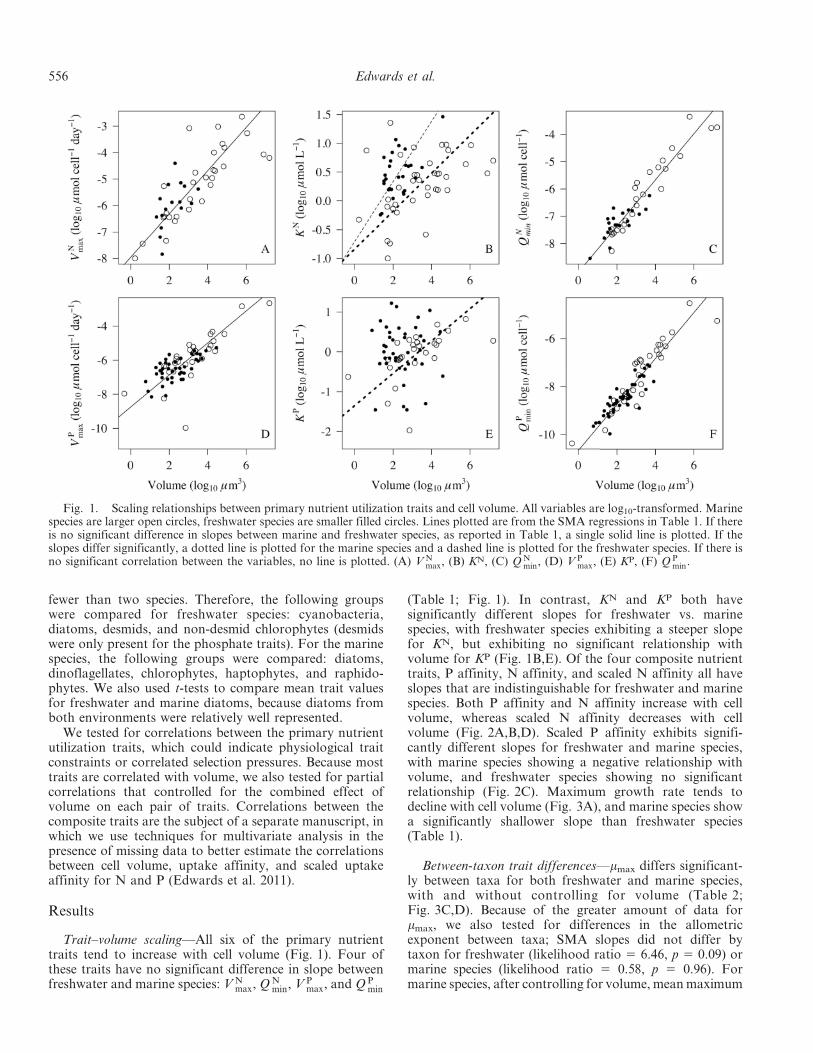
Trait–volume scaling—All six of the primary nutrienttraits tend to increase with cell volume (Fig. 1). Four ofthese traits have no significant difference in slope betweenfreshwater and marine species: V N
max, Q Nmin, V P
max, and Q Pmin
(Table 1; Fig. 1). In contrast, KN and KP both havesignificantly different slopes for freshwater vs. marinespecies, with freshwater species exhibiting a steeper slopefor KN, but exhibiting no significant relationship withvolume for KP (Fig. 1B,E). Of the four composite nutrienttraits, P affinity, N affinity, and scaled N affinity all haveslopes that are indistinguishable for freshwater and marinespecies. Both P affinity and N affinity increase with cellvolume, whereas scaled N affinity decreases with cellvolume (Fig. 2A,B,D). Scaled P affinity exhibits signifi-cantly different slopes for freshwater and marine species,with marine species showing a negative relationship withvolume, and freshwater species showing no significantrelationship (Fig. 2C). Maximum growth rate tends todecline with cell volume (Fig. 3A), and marine species showa significantly shallower slope than freshwater species(Table 1).
Between-taxon trait differences—mmax differs significant-ly between taxa for both freshwater and marine species,with and without controlling for volume (Table 2;Fig. 3C,D). Because of the greater amount of data formmax, we also tested for differences in the allometricexponent between taxa; SMA slopes did not differ bytaxon for freshwater (likelihood ratio 5 6.46, p 5 0.09) ormarine species (likelihood ratio 5 0.58, p 5 0.96). Formarine species, after controlling for volume, mean maximum
Fig. 1. Scaling relationships between primary nutrient utilization traits and cell volume. All variables are log10-transformed. Marinespecies are larger open circles, freshwater species are smaller filled circles. Lines plotted are from the SMA regressions in Table 1. If thereis no significant difference in slopes between marine and freshwater species, as reported in Table 1, a single solid line is plotted. If theslopes differ significantly, a dotted line is plotted for the marine species and a dashed line is plotted for the freshwater species. If there isno significant correlation between the variables, no line is plotted. (A) V N
max, (B) KN, (C) Q Nmin, (D) V P
max, (E) KP, (F) Q Pmin.
556 Edwards et al.

growth rates for diatoms are greater than those ofchlorophytes, dinoflagellates, and cyanobacteria (Tukeypairwise comparison p , 0.001 for all tests). This meansthat at a given cell size, diatoms grow faster than othertaxonomic groups. In addition, dinoflagellates and hapto-phytes have greater growth rates than cyanobacteria whencontrolling for volume (p 5 0.0081 and p 5 0.0078,respectively). For freshwater species, cyanobacteria havelower growth rates than diatoms, desmids, non-desmidchlorophytes, and cryptomonads when controlling forvolume (p , 0.001 for all tests).
Most nutrient traits differ significantly between taxawhen cell volume is not included as a covariate (Table 2;Figs. 4–7). However, few traits differ between taxa whenthe effect of volume is accounted for (Table 2; Figs. 4–7).For freshwater species, values of primary nutrient traitstend to be greatest for the desmids and smallest for thecyanobacteria, with diatoms and non-desmid chloro-phytes having intermediate values (Fig. 4). For marinespecies, dinoflagellates and raphidophytes tend to havethe largest trait values, whereas diatoms have intermedi-ate values and haptophytes and chlorophytes have thesmallest values (Fig. 6). Because few between-taxondifferences remain when cell volume is controlled for,these differences are evidently driven by differencesbetween taxa in typical cell size. For freshwater species,the exceptions are KP, P affinity, and scaled N affinity, allof which differ between taxa while controlling for volume(Table 2; Figs. 4, 5). Tukey tests show that for KP,desmids have significantly greater values than the otherthree taxa, and non-desmid chlorophytes have significantgreater values than cyanobacteria (p , 0.01 for all tests).For P affinity, desmids have significantly greater valuesthan cyanobacteria (p 5 0.034). For scaled N affinity,diatoms and non-desmid chlorophytes both have signif-icantly greater values than cyanobacteria (p 5 0.036 and0.026, respectively). For marine species, the traits withsignificant between-taxon variation when controlling forvolume are KN and scaled N affinity (Table 2; Figs. 6, 7).For KN, chlorophytes have greater values than diatoms (p, 0.001), dinoflagellates (p , 0.001), haptophytes (p ,0.001), and raphidophytes (p , 0.037). For scaled Naffinity, although there is significant between-taxonheterogeneity, no pairwise comparisons are significantlydifferent at p , 0.05.
Diatoms—We compared log-transformed mean traitvalues for freshwater and marine diatoms using those traitsmeasured on at least five species from each environment(mmax, KN, Q N
min, V Pmax, KP, Q P
min, P affinity, and scaled Paffinity). Marine diatoms have higher mmax than freshwaterdiatoms (back-transformed means of 1.0 vs. 0.28, respec-tively; p 5 0.001). Marine diatoms have lower KN (back-transformed means of 1.6 vs. 3.5; p 5 0.018), higher V P
max(back-transformed means of 2.8 3 1026 vs. 2.5 3 1027; p 50.029), and higher KP (back-transformed means of 1.1 vs.0.23; p 5 0.035). Marine diatoms are also larger on average(means of 2090 vs. 436 m3; p 5 0.009), but the othersignificant trait differences were unchanged in analyses ofcovariance that included volume as a covariate.
Table
1.
Co
effi
cien
tsfo
rS
MA
reg
ress
ion
of
nu
trie
nt
uti
liza
tio
ntr
ait
sa
nd
mm
ax
wit
hce
llv
olu
me.
Inte
rcep
ts(i
nt.
)a
nd
slo
pes
are
list
edfo
rse
pa
rate
reg
ress
ion
so
ffr
esh
wa
ter
an
dm
ari
ne
spec
ies,
an
dth
eco
mm
on
inte
rcep
tsa
nd
slo
pes
are
list
edfo
rre
gre
ssio
ns
tha
tco
mb
ine
fres
hw
ate
ra
nd
ma
rin
esp
ecie
s.9
5%
con
fid
ence
inte
rva
lsfo
rea
chp
ara
met
era
rein
pa
ren
thes
es.
ns
isli
sted
for
tra
its
no
tsi
gn
ific
an
tly
corr
ela
ted
wit
hv
olu
me.
Th
efi
na
lco
lum
nli
sts
the
pv
alu
efo
ra
test
of
wh
eth
erth
efr
esh
wa
ter
an
dm
ari
ne
spec
ies
ha
ve
dif
fere
nt
slo
pes
.p
va
lues
,0
.05
are
ma
rked
wit
ha
na
ster
isk
.
Fre
shw
ate
rin
t.F
resh
wa
ter
slo
pe
Ma
rin
ein
t.M
ari
ne
slo
pe
Co
mm
on
int.
Co
mm
on
slo
pe
Slo
pe
p
VN m
ax
28
.8(2
10
,2
7.4
)1
.3(0
.82
,2
.1)
28
.1(2
8.8
,2
7.3
)0
.82
(0.6
5,
1)
28
.0(2
8.5
,2
7.4
)0
.82
(0.6
8,
0.9
8)
0.0
69
KN
20
.71
(21
.1,
20
.28
)0
.52
(0.3
7,
0.7
3)
20
.84
(21
.2,
20
.44
)0
.33
(0.2
4,
0.4
5)
20
.61
(20
.88
,2
0.3
4)
0.3
3(0
.26
,0
.42
)0
.04
4*
QN m
in2
8.7
(29
.2,
28
.2)
0.6
8(0
.51
,0
.93
)2
9.2
(29
.6,
28
.7)
0.8
8(0
.77
,1
)2
9.0
(29
.3,
28
.8)
0.8
4(0
.76
,0
.94
)0
.13
VP m
ax
28
.4(2
8.9
,2
7.9
)0
.81
(0.6
4,
1)
29
.1(2
10
,2
8.2
)1
.0(0
.8,
1.3
)2
8.7
(29
.2,
28
.3)
0.9
4(0
.81
,1
.1)
0.1
7
KP
ns
ns
21
.4(2
1.8
,2
0.8
8)
0.4
1(0
.29
,0
.56
)2
1.5
(21
.9,
21
.1)
0.5
3(0
.42
,0
.67
)0
.00
97
*
QP m
in2
10
.5(2
10
.8,
21
0.2
)0
.86
(0.7
4,
1)
21
0.6
(21
1.1
,2
10
.1)
0.9
7(0
.84
,1
.1)
21
0.7
(21
0.9
,2
10
.4)
0.9
6(0
.87
,1
.1)
0.3
Pa
ffin
ity
29
(29
.7,
28
.2)
1.0
(0.7
6,
1.3
)2
8.1
(28
.8,
27
.5)
0.7
3(0
.58
,0
.94
)2
8.5
(29
,2
8.1
)0
.85
(0.7
2,
1)
0.0
89
Na
ffin
ity
28
.6(2
9.8
,2
7.3
)1
.0(0
.6,
1.7
)2
8.2
(29
,2
7.4
)0
.75
(0.5
8,
0.9
8)
28
.1(2
8.7
,2
7.6
)0
.75
(0.6
1,
0.9
2)
0.3
3P
sca
led
aff
init
yn
sn
s3
.6(2
.8,
4.3
)2
0.5
5(2
0.3
8,
20
.8)
3.6
(3.1
,4
.1)
20
.65
(20
.5,
20
.83
)0
.04
6*
Nsc
ale
da
ffin
ity
20
.96
(22
.1,
0.2
)0
.78
(0.4
3,
1.4
)2
.9(1
.9,
3.9
)2
0.6
3(2
0.4
4,
20
.91
)2
.4(1
.8,
3.0
)2
0.5
7(2
0.4
3,
20
.78
)0
.54
mm
ax
0.6
9(0
.52
,0
.86
)2
0.3
6(2
0.3
0,
20
.43
)0
.70
(0.5
4,
0.8
5)
20
.24
(20
.20
,2
0.2
9)
0.6
5(0
.53
,0
.76
)2
0.2
8(2
0.2
5,
20
.32
)0
.00
14
*
Reviews in L&O 557

Correlations between traits—Raw correlations betweenprimary nutrient utilization traits are dominated by strongpositive correlations (Table 3). For the marine species, allpairs of traits are significantly positively correlated. For thefreshwater species, most pairs of traits either are signifi-cantly positively correlated or show no significant correla-tion. The single exception to this pattern is a significantnegative correlation between KP and KN among thefreshwater species (Table 3). The dominance of positivecorrelations among traits is likely driven by the fact that alltraits tend to increase with cell volume (Fig. 1). A similarargument applies to the fact that all of the raw correlationsof nutrient traits with mmax are negative (Table 3). Wetherefore tested for partial correlations, which control forthe simultaneous effect of cell volume on each pair of traits.In this analysis, positive correlations between nutrient traitsstill dominate, as do negative correlations between nutrienttraits and mmax (Table 3). Among marine species, five pairsof traits still show significant positive correlations betweennutrient traits; among freshwater species, four pairs oftraits show significant positive correlations, whereas KP
and KN again show a negative correlation. As discussedbelow, the preponderance of positive correlations thatremain while controlling for volume may indicate that theeffect of volume is not fully removed.
Q Nmin : Q P
min ratios—We tested whether the ratio Q Nmin :
Q Pmin tended to differ between marine and freshwater
habitats. Both habitats showed broad variation in thisratio, but freshwater species had a significantly higher ratioon average (Fig. 8; freshwater median molar ratio, 24.4;marine median molar ratio, 13.6; two-sample Wilcoxontest, p 5 0.032).
Discussion
Scaling relationships and marine–freshwater differences—We find that Vmax, K, and Qmin all tend to increase as cellvolume increases, although this is not the case for KP
among freshwater species (Fig. 1; Table 1). It is noteworthythat marine and freshwater slopes are indistinguishable forthose traits that appear to be the most strongly constrainedby volume, Vmax and Qmin (Fig. 1). In contrast, the half-saturation constants for uptake appear to be less stronglyconstrained by volume, and have significantly differentslopes for freshwater and marine species (Fig. 1B,E),possibly reflecting different selective pressures from N orP limitation. If P limitation is more prevalent in freshwaterenvironments, it may select for low KP and thus decreasethis trait’s dependence on cell volume. The generallypositive effect of volume on all of these traits is consistent
Fig. 2. Scaling relationships between composite nutrient traits and cell volume. All variablesare log10-transformed. Lines plotted are from the SMA regressions in Table 1. If there is nosignificant difference in slopes between marine and freshwater species, as reported in Table 1, asingle solid line is plotted. Symbols as in Fig. 1. (A) P affinity (aff.), (B) N aff., (C) scaled P aff.,(D) scaled N aff.
558 Edwards et al.

with prior analyses (Shuter 1978; Grover 1989; Litchmanet al. 2007). These results are also largely consistent withthe idea that marine and freshwater phytoplankton havesimilar physiological ecology (Kilham and Hecky 1988),although the different environments may modify traitpatterns to some extent. For example, diatoms in marineand freshwater systems appear to occupy somewhatdifferent ecological niches and exhibit different strategies.Marine diatoms are a fast-growing group compared toother marine taxa and appear to be adapted to highnutrient conditions. In contrast, freshwater diatoms haveintermediate growth rates and low KP compared to otherfreshwater taxa. They also have a relatively high P affinityamong freshwater groups, likely being adapted to P-limitedconditions. When marine and freshwater diatoms arecompared, marine diatoms have significantly higher growthrates and KP than freshwater diatoms; conversely, marine
diatoms also tend to have lower KN than freshwaterdiatoms. These differences possibly reflect different selec-tive pressures in marine and freshwater environments.Because of the heavy silica frustules of diatoms, in marineenvironments diatoms benefit from mixing conditionsassociated with high nutrients that select for high growthrates. In freshwaters, diatoms also are associated withmixing conditions, but in shallow lakes mixed layer depthsare too small and strongly select against diatoms (Ptacniket al. 2003). Therefore, freshwater diatoms may be moreprevalent in deeper lakes that also tend to be lower in P(Dillon and Rigler 1974), causing freshwater diatoms to begood P competitors.
Uptake affinity is often used as a proxy of the ability togrow or compete under nutrient limitation; therefore, it isimportant to note that uptake affinity on a per-cell basisincreases with cell volume (Fig. 2A,B), because Vmax scales
Fig. 3. Allometric scaling and between-taxon differences in mmax. (A) mmax vs. cell volumefor freshwater and marine species. The fitted SMA regressions a plotted for marine (dashed line)and freshwater (dotted line) species. (B) Violin plots of mmax (on a linear scale). The violin plotscombine a box plot (showing 10th, 25th, 50th, 75th, and 90th quantiles) with a density plot. (C)mmax vs. volume for freshwater species, coded by taxon. (D) mmax vs. volume for marine species,coded by taxon. Chrys, chrysophytes, crypt, cryptophytes; diat, diatoms; cyan, cyanobacteria;chlor, chlorophytes; dino, dinoflagellates; hapt, haptophytes.
Reviews in L&O 559

more steeply with volume than K. In contrast, scaleduptake affinity tends to decrease with cell volume, althoughstrong support for this relationship is present only formarine species. Among freshwater species, scaled P affinityshows no relationship with cell volume (Fig. 2C). Further-more, although the slopes for scaled N affinity cannot bedistinguished for freshwater and marine species, thefreshwater species occupy a limited range of volumes, andby themselves show no correlation with volume (Kendallrank correlation 0.03, p 5 0.91; Fig. 2D). Scaled uptakeaffinity should be a more appropriate proxy for competitiveability under nutrient limitation, because it standardizesuptake ability by the nutrient requirement for growth. Thisis supported by our comparison of scaled uptake affinitieswith the outcome of P-limited chemostat experiments (WebAppendix, Table A2). Our results therefore support theexpectation that the ability to compete for limitingnutrients tends to decline with increasing cell volume(Chisholm 1992), although this relationship may be weakeror nonexistent in freshwater habitats. These results are alsoconsistent with a study of marine species that measuredscaled P affinity directly, as the biomass-specific P turnoverrate, and found that scaled P affinity declines withincreasing equivalent spherical radius (Tambi et al. 2009).
Maximum growth rate tends to decline with increasing cellvolume, consistent with prior results (Banse 1976; Chisholm1992). To our knowledge, this is the first study comprehen-sively comparing marine and freshwater species, and we findthat the allometric slope is shallower for marine species(Table 1; Fig. 3). The consequence of this difference in slopesis that, on average, marine phytoplankton of a certain sizegrow faster than freshwater species of the same size, and thisdifference is greater as cell size increases (Fig. 3A). Theallometric slopes of mmax for freshwater and marine species are20.36 and 20.24 respectively, and the slope when all speciesare combined is 20.28 (Table 1). These slopes are steeperthan some previously reported slopes (Banse 1976; Tang1995), but this may be due to our use of SMA regression.Lines fit using ordinary least squares (OLS) have slopesbiased towards zero (Warton et al. 2006), and if we fit ourdata using OLS the slopes are 20.10 and 20.096 forfreshwater and marine species, respectively. Because allome-tric exponents are used to parameterize ecosystem models(Irwin et al. 2006), and to test hypothesized mechanisms forscaling relationships (DeLong et al. 2010), we recommend theuse of SMA regression in comparative analyses, as discussedin detail by Warton et al. (2006).
In general, there are a number of limitations to ourcomparisons of freshwater and marine species. Thefreshwater species in our dataset cover a smaller range ofcell volumes, which may limit our ability to detect scalingrelationships and therefore obscure potential effects ofvolume on scaled uptake affinity. However, freshwaterphytoplankton may have a smaller cell size range comparedto marine phytoplankton in general (Litchman et al. 2009;Kamenir et al. 2010). In addition, the species in ourcompilation are those that phytoplankton ecologists havechosen to study in the lab, rather than a random or size-structured sample from natural communities. Future workon trait–volume scaling relationships would benefit from a
Table
2.
Tes
tso
fb
etw
een
-ta
xo
ntr
ait
dif
fere
nce
s.F
or
each
tra
it,
the
Fst
ati
stic
an
dp
va
lue
are
list
edfo
ra
on
e-w
ay
an
aly
sis
of
va
ria
nce
test
ing
the
effe
cto
fta
xo
nin
iso
lati
on
(‘‘T
ax
on
on
ly’’
row
s).
Nu
mer
ato
ra
nd
den
om
ina
tor
deg
rees
of
free
do
ma
rein
pa
ren
thes
es.
Ina
dd
itio
n,
the
Wa
ld(W
)st
ati
stic
an
dp
va
lue
are
list
edfo
ra
test
of
com
mo
nin
terc
epts
acr
oss
tax
ain
an
SM
Are
gre
ssio
nw
ith
cell
vo
lum
e(‘
‘Ta
xo
nw
ith
vo
lum
e[w
/vo
l.]’
’ro
ws)
.P
va
lues
,0
.05
are
ma
rked
wit
ha
na
ster
isk
.
VN m
ax
KN
QN m
inV
P ma
xK
PQ
P min
Pa
ffin
ity
Na
ffin
ity
Psc
ale
da
ffin
ity
Nsc
ale
da
ffin
ity
mm
ax
Ma
rin
esp
ecie
s
Ta
xo
no
nly
F4
.5(4
,25
)5
.3(4
,32
)6
.3(3
,19
)0
.9(4
,22
)0
.6(4
,22
)5
.2(4
,23
)0
.81
(4,2
0)
6.5
(4,2
5)
3.5
(4,1
6)
5.8
(3,1
2)
12
.2(4
,9
1)
Ta
xo
no
nly
p0
.00
75
*0
.00
22
*0
.00
38
*0
.48
0.6
70
.00
38
*0
.53
0.0
01
*0
.03
1*
0.0
11
*5
.63
10
28*
Ta
xo
nw
/vo
l.W
1.5
8.8
0.8
31
.21
.61
.21
.63
.30
.43
0.5
13
0T
ax
on
w/v
ol.
p0
.24
7.1
03
10
25*
0.5
0.3
50
.21
0.3
50
.22
0.0
29
*0
.78
0.6
84
.53
10
26*
Fre
shw
ate
rsp
ecie
s
Ta
xo
no
nly
F9
.2(2
,13
)0
.77
(2,2
4)
3.1
(2,1
7)
5.5
(3,3
4)
5.1
(3,3
1)
14
(3,4
0)
1.7
(3,3
1)
4.2
(2,1
2)
2.8
(3,2
8)
3.3
(2,1
1)
2.3
(4,
11
2)
Ta
xo
no
nly
p0
.00
32
*0
.48
0.0
74
*0
.00
34
*0
.00
56
*2
.803
10
26*
0.1
90
.04
2*
0.0
60
.07
70
.06
1T
ax
on
w/v
ol.
W3
.21
.62
2.4
13
0.5
73
.11
.11
.25
.23
5T
ax
on
w/v
ol.
p0
.07
50
.22
0.1
70
.08
81
.503
10
25*
0.6
40
.04
3*
0.3
80
.35
0.0
28
*5
.03
10
26*
560 Edwards et al.

more comprehensive sampling of natural communities,including endpoints of the size spectrum, from picoplank-ton to giant diatoms.
Between-taxon differences—We find strong differencesbetween taxa in average nutrient trait values (Figs. 3–6;Table 2). At the same time, few between-taxon differenc-es remain when controlling for cell volume (Table 2),indicating that taxonomic variation in nutrient utilizationtraits is largely driven by size variation. Considering rawtrait values (not corrected for volume), among marinespecies differences in scaled uptake affinities suggest thatthe relatively small haptophytes and prymnesiophytesshould be better competitors under P limitation, whereasthe relatively large dinoflagellates and raphidophytesshould be poorer competitors (Fig. 7C). The same trendsoccur for scaled N affinity, although there are no data forchlorophytes (Fig. 7D). Among freshwater species, thedata for scaled P affinity suggest that cyanobacteria tendto be better competitors, whereas the desmids are poorcompetitors, with diatoms and non-desmid chlorophytesin between (Fig. 5C). There are fewer data for scaledN affinity among freshwater species, but the presentpatterns suggest that cyanobacteria may be poorercompetitors for nitrate (Fig. 5D); however, only twocyanobacteria are present in this analysis, and one ofthese species can fix nitrogen, which may compensate forlow nitrate affinity.
There is significant variation in mean mmax between taxa(Table 2), with most variation due to the fact that amongmarine species, diatoms have high growth rates relativeto their cell volume, whereas among freshwater species,cyanobacteria have low growth rates relative to their cellvolume (Fig. 3C,D). However, because of the relativelysmall size of cyanobacteria cells, their raw growth rates arestill relatively high among freshwater species, on average(Fig. 3B).
Q Nmin : Q P
min ratios—Our analysis of the structural N : Pratio supports prior work showing broad variation in thisratio across phytoplankton (Rhee and Gotham 1980;Klausmeier et al. 2004). We also find that Q N
min : Q Pmin is
greater on average for freshwater species. If the evolutionof this ratio is driven by the relative availability of N and P,the difference between habitats may be due to greaterprevalence of P limitation in freshwater systems (Elser et al.2007). Alternatively, if the evolution of this ratio is drivenby allocation to resource acquisition proteins vs. ribosomes(Klausmeier et al. 2004), higher ratios among freshwaterspecies may indicate a greater tendency for nutrient or lightlimitation in freshwater systems, and a greater tendency fornear-maximal growth rates in marine systems (Goldmanet al. 1979).
Comparison of scaling exponents to theoretical predic-tions—The observed scaling exponents of nutrient utilization
Fig. 4. Between-taxon variation in primary nutrient traits for freshwater species. All variables are log10-transformed. For eachtaxon, mean trait value and mean volume are plotted, 6 1 SE. The SMA fits from Fig. 1 are plotted as dotted lines. cyan, cyanobacteria;diat, diatoms; chlor, non-desmid chlorophytes; desm, desmids. (A) V N
max, (B) KN, (C) Q Nmin, (D) V P
max, (E) KP, (F) Q Pmin.
Reviews in L&O 561

traits can be compared to those expected under amechanistic model of nutrient uptake (Aksnes and Egge1991). If Vmax is determined by the number of uptake siteson the cell surface, and if the density of uptake sites doesnot vary with cell volume, then cell-specific maximumuptake rate should be proportional to surface area. Forspherical cells, this will result in a scaling exponent of 2/3for Vmax relative to cell volume. Although a prior analysisof nitrate traits in marine species found a scaling exponentconsistent with two-thirds scaling (Litchman et al. 2007),with the current, larger dataset, both V N
max and V Pmax scale
more steeply than 2/3 (95% confidence intervals are [0.68,0.98] and [0.81, 1.1], respectively; Table 1). However, aninterspecific comparison has found that species withgreater cell volume tend to show greater deviation froma spherical shape, and as a result the surface area : volumeratio declines with increasing cell volume, but it declinesmore slowly than expected for spherical cells (Reynolds2006). Therefore, our results for the scaling of Vmax arenot inconsistent with a scaling relationship driven bysurface area.
K appears to be less constrained by cell volume thaneither Vmax or Qmin, and unlike these traits, K exhibitsdifferent scaling patterns for freshwater and marine species,with KP showing no correlation with volume for freshwater
species (Table 1; Fig. 1). Therefore, although cell volumemay affect K by altering the mass transfer coefficient(Aksnes and Egge 1991), our results suggest that otherfactors beyond cell volume strongly affect K.
We hypothesized earlier (Litchman et al. 2007) thatallometric exponents for Qmin may range from 2/3 to 1, ifwe assume that the minimum nutrient quota is contained inthe cytoplasm that occupies most of the cell volume insmall cells (isometric scaling), or is distributed along the cellwall in cells with large vacuoles (thus scales with cellsurface, 2/3 of volume). We are, however, unaware ofmechanistic predictions for whether Qmin scaling exponentsshould be different for N and P. Intriguingly, Q N
min scalesallometrically with volume (95% confidence interval [0.76,0.94]; Table 1), whereas the scaling of Q P
min is indistin-guishable from isometric (95% confidence interval [0.87,1.1]; Table 1). This also implies that the ratio Q N
min : Q Pmin
may decline as volume increases. In our dataset, this is truewhen marine and freshwater species are combined (Kendallrank correlation: 20.28, p 5 0.013), although not whenmarine and freshwater species are analyzed separately(marine correlation 20.059, p 5 0.77; freshwater correla-tion 20.25, p 5 0.14).
The allometric scaling of uptake affinities and scaleduptake affinities are consistent with the scaling of the
Fig. 5. Between-taxon variation in composite traits for freshwater species. All variables arelog10-transformed. For each taxon, mean trait value and mean volume are plotted, 6 1 SE. TheSMA fits from Fig. 2 are plotted as dotted lines. cyan, cyanobacteria; diat, diatoms; chlor, non-desmid chlorophytes; desm, desmids. (A) P affinity, (B) N affinity, (C) scaled P affinity, (D)scaled N affinity.
562 Edwards et al.

component parameters Vmax, K, and Qmin. Vmax tends toincrease more steeply with volume than K, resulting in anincrease in cell-specific uptake affinity with volume (Fig. 2).When uptake affinity is scaled by Qmin, this results in adecrease in scaled uptake affinity with increasing volume,because the scaling exponents of Vmax and Qmin tend to besimilar. The exception to this trend is scaled P affinity forfreshwater species, which does not decline with increasingvolume; this may be due to the fact that KP does not declinewith volume.
Potential constraints among nutrient utilization traits—Araw correlation analysis reveals a preponderance of positivecorrelations among nutrient traits (Table 3). These positivecorrelations may partially constrain the evolution ofcompetitive ability, because for a given nutrient, a positivecorrelation between Vmax and K, or between Vmax andQmin, constrains the evolution of equilibrium competitiveability, whereas a positive correlation between K and Qmin
has the opposite effect (Litchman et al. 2007). However, ifthese trait correlations are driven solely by cell size, thenthe only relevant constraints for trait evolution are thosethat will constrain size evolution, i.e., the trait–volumescaling parameters (Litchman et al. 2009). Our partialcorrelation analysis, which tested for trait correlations aftercontrolling for the effect of cell volume, yielded equivocalresults. This analysis yielded only positive significantcorrelations between nutrient traits, with the exception of
KP vs. KN for freshwater species, which is also negative inthe raw correlation analysis (Table 3). Furthermore, thesignificant correlations occurred among pairs of traits thatwere strongly correlated in the raw correlation analysis(Table 3). It therefore seems possible that the partialcorrelation analysis did not succeed in fully removing thecorrelated effect of volume on nutrient traits. Measurementerror for either nutrient traits or cell volume, along withplasticity in cell volume, will tend to add noise to the trait–volume relationship. This noise will prevent the partialcorrelation from fully removing the true relationshipbetween a trait and volume. The same considerations applyto the negative correlations between mmax and nutrienttraits (Table 3).
However, some of the partial correlations may indeedrepresent constraints, or correlated selection pressures. Thenegative correlation between KP and KN among freshwaterspecies is unlikely to be driven by volume, and may con-tribute to a trade-off in competitive ability for nitrate vs.phosphate among freshwater species. Among the positivepartial correlations, nearly all would constrain the evo-lution of high scaled uptake affinity for a single nutrient(for marine species, V N
max vs. Q Nmin, V P
max vs. KP; forfreshwater species, V N
max vs. Q Nmin), or would constrain the
simultaneous evolution of high scaled uptake affinity forboth nutrients (for marine species, V N
max vs. KP, V Pmax vs. KN;
for freshwater species, V Nmax vs. V P
max, V Pmax vs. Q N
min).
Therefore, these relationships may represent eco-evolutionary
Fig. 6. Between-taxon variation in primary nutrient traits for marine species. All variables are log10-transformed. For each taxon,mean trait value and mean volume are plotted, 6 1 SE. The SMA fits from Fig. 1 are plotted as dotted lines. hapt, haptophytes; diat,diatoms; chlor, chlorophytes; dino, dinoflagellates; raph. raphidophytes. (A) V N
max, (B) KN, (C) Q Nmin, (D) V P
max, (E) KP, (F) Q Pmin.
Reviews in L&O 563

Fig. 7. Between-taxon variation in composite traits for marine species. All variables arelog10-transformed. For each taxon, mean trait value and mean volume are plotted, 6 1 SE. TheSMA fits from Fig. 2 are plotted as dotted lines. hapt, haptophytes; diat, diatoms; chlor,chlorophytes; dino, dinoflagellates; raph, raphidophytes. (A) P affinity, (B) N affinity, (C) scaledP affinity, (D) scaled N affinity.
Table 3. Correlation matrices for between-trait relationships. Entries in the upper right are for marine species, entries in the lowerleft are for freshwater species.
V Nmax KN Q N
min V Pmax KP Q P
min mmax
Raw Kendall rank correlations
V Nmax
0.28* 0.69** 0.6** 0.71** 0.54* 20.37*
KN 0.27 0.53* 0.6** 0.52* 0.56* 20.27*
Q Nmin
0.66** 0.13 0.66** 0.69** 0.84** 20.61**
V Pmax
0.43* 20.066 0.52* 0.59** 0.66** 20.22
KP 20.14 20.45* 20.3 0.19 0.64** 20.14
Q Pmin
0.7** 20.037 0.55** 0.42** 20.0034 20.35
mmax 20.029 20.16 20.22 20.2 20.032 20.2
Partial Kendall rank correlations thatcontrol for cell volume
V Nmax
0.086 0.45* 0.34 0.53* 0.072 20.12
KN 0.18 0.032 0.37* 0.19 0.078 20.091
Q Nmin
0.57* 0.065 0.087 0.13 0.53* 20.35*
V Pmax
0.39* 20.1 0.62** 0.31* 0.13 0.06
KP 0.051 20.38* 20.0095 0.19 0.20 0.12
Q Pmin
0.62* 20.18 0.3 0.21 0.059 20.23
mmax 0.024 20.12 20.25 20.2 20.027 20.042
* p , 0.05, ** p , 0.001.
564 Edwards et al.

trade-offs among nutrient utilization traits. Edwards et al.(2011) used the same dataset to test for trade-offs amongscaled N affinity, scaled P affinity, and cell volume. Theyfound evidence for a general three-way trade-off among thesethree traits, with the relationship between scaled N affinityand scaled P affinity most evident in freshwater species, andthe relationship between cell volume and the two nutrienttraits most evident in marine species.
Our extensive compilation of nutrient utilization traits inmarine and freshwater phytoplankton shows that there arefundamental scaling relationships for most traits, and theserelationships are usually shared by marine and freshwaterphytoplankton. However, there are some trait differencesthat possibly reflect different selective pressures betweenmarine and freshwater environments, such as the relativestrength of limitation by different nutrients. Major taxadiffer in their nutrient utilization traits, reflecting contrast-ing ecological strategies that are often driven by differencesin cell volume. Our data also suggest that some taxa, suchas diatoms, may have different strategies in marine vs.
freshwater environments, highlighting the interaction ofphylogeny and environmental controls.
AcknowledgmentsThis work was in part supported by National Science
Foundation grant 09-28819 to E.L. and C.A.K., 08-45932 toE.L., and a James S. McDonnell Foundation grant to C.A.K. andE.L. We thank Jonathan J. Cole and two anonymous reviewersfor helpful comments. This is Kellogg Biological Stationcontribution No. 1635.
References
AKSNES, D. L., AND J. K. EGGE. 1991. A theoretical model fornutrient uptake in phytoplankton. Mar. Ecol. Prog. Ser. 70:65–72, doi:10.3354/meps070065
BANSE, K. 1976. Rates of growth, respiration and photosynthesisof unicellular algae as related to cell size—review. J. Phycol.12: 135–140.
CHISHOLM, S. W. 1992. Phytoplankton size, p. 213–237. In P. G.Falkowski and A. D. Woodhead [eds.], Primary productivityand biogeochemical cycles in the sea. Plenum.
DELONG, J. P., J. G. OKIE, M. E. MOSES, R. M. SIBLY, AND J. H.BROWN. 2010. Shifts in metabolic scaling, production, andefficiency across major evolutionary transitions. Proc. Natl.Acad. Sci. USA 107: 12941–12945, doi:10.1073/pnas.1007783107
DILLON, P. J., AND F. H. RIGLER. 1974. Test of a simple nutrientbudget model predicting phosphorus concentration in lake water.J. Fish. Res. Board Can. 31: 1771–1778, doi:10.1139/f74-225
EDWARDS, K. F., C. A. KLAUSMEIER, AND E. LITCHMAN. 2011.Evidence for a three-way tradeoff between nitrogen andphosphorus competitive abilities and cell size in phytoplank-ton. Ecology 92: 1085–1095.
ELSER, J. J., AND OTHERS. 2007. Global analysis of nitrogen andphosphorus limitation of primary producers in freshwater,marine and terrestrial ecosystems. Ecol. Lett. 10: 1135–1142,doi:10.1111/j.1461-0248.2007.01113.x
EPPLEY, R. W., J. N. ROGERS, AND J. J. MCCARTHY. 1969. Half-saturation constants for uptake of nitrate and ammonium bymarine phytoplankton. Limnol. Oceanogr. 14: 912–920,doi:10.4319/lo.1969.14.6.0912
GOLDMAN, J. C., J. J. MCCARTHY, AND D. G. PEAVEY. 1979.Growth influence on the chemical composition of phyto-plankton in oceanic waters. Nature 279: 210–215, doi:10.1038/279210a0
GROVER, J. P. 1989. Influence of cell shape and size on algalcompetitive ability. J. Phycol. 25: 402–405, doi:10.1111/j.1529-8817.1989.tb00138.x
———. 1991. Dynamics of competition among microalgae invariable environments: Experimental tests of alternativemodels. Oikos 62: 231–243, doi:10.2307/3545269
HEALEY, F. P. 1980. Slope of the monod equation as an indicatorof advantage in nutrient competition. Microb. Ecol. 5:281–286, doi:10.1007/BF02020335
HECKY, R. E., AND P. KILHAM. 1988. Nutrient limitation ofphytoplankton in freshwater and marine environments: Areview of recent evidence on the effects of enrichment. Limnol.Oceanogr. 33: 796–822, doi:10.4319/lo.1988.33.4_part_2.0796
IRWIN, A. J., Z. V. FINKEL, O. M. E. SCHOFIELD, AND P. G.FALKOWSKI. 2006. Scaling-up from nutrient physiology to thesize-structure of phytoplankton communities. J. PlanktonRes. 28: 459–471, doi:10.1093/plankt/fbi148
KAMENIR, Y., Z. DUBINSKY, AND R. HARRIS. 2010. Taxonomic sizestructure consistency of the English Channel phytoplankton. J. Exp.Mar. Biol. Ecol. 383: 105–110, doi:10.1016/j.jembe.2009.12.009
Fig. 8. Distribution of Q Nmin:Q P
min molar ratios. X-axis is ona log scale. (A) Freshwater species, (B) marine species.
Reviews in L&O 565

KILHAM, P., AND R. E. HECKY. 1988. Comparative ecology ofmarine and freshwater phytoplankton. Limnol. Oceanogr. 33:776–795, doi:10.4319/lo.1988.33.4_part_2.0776
KLAUSMEIER, C. A., E. LITCHMAN, T. DAUFRESNE, AND S. A. LEVIN.2004. Optimal nitrogen-to-phosphorus stoichiometry of phy-toplankton. Nature 429: 171–174, doi:10.1038/nature02454
LITCHMAN, E., AND C. A. KLAUSMEIER. 2008. Trait-based commu-nity ecology of phytoplankton. Annu. Rev. Ecol. Evol. Syst.39: 615–639, doi:10.1146/annurev.ecolsys.39.110707.173549
———, ———, J. R. MILLER, O. M. SCHOFIELD, AND P. G.FALKOWSKI. 2006. Multi-nutrient, multi-group model ofpresent and future oceanic phytoplankton communities.Biogeosciences 3: 585–606, doi:10.5194/bg-3-585-2006
———, ———, O. M. SCHOFIELD, AND P. G. FALKOWSKI. 2007. Therole of functional traits and trade-offs in structuring phyto-plankton communities: Scaling from cellular to ecosystem level.Ecol. Lett. 10: 1170–1181, doi:10.1111/j.1461-0248.2007.01117.x
———, ———, AND K. YOSHIYAMA. 2009. Contrasting sizeevolution in marine and freshwater diatoms. Proc. Natl. Acad.Sci. USA 106: 2665–2670, doi:10.1073/pnas.0810891106
MOREL, F. M. M. 1987. Kinetics of nutrient uptake and growth inphytoplankton. J. Phycol. 23: 137–150.
PTACNIK, R., S. DIEHL, AND S. BERGER. 2003. Performance ofsinking and nonsinking phytoplankton taxa in a gradient ofmixing depths. Limnol. Oceanogr. 48: 1903–1912, doi:10.4319/lo.2003.48.5.1903
REYNOLDS, C. S. 2006. Ecology of phytoplankton. CambridgeUniv. Press.
RHEE, G. Y., AND I. J. GOTHAM. 1980. Optimum N : P ratios andcoexistence of planktonic algae. J. Phycol. 16: 486–489,doi:10.1111/j.1529-8817.1980.tb03065.x
SHUTER, B. J. 1978. Size-dependence of phosphorus and nitrogensubsistence quotas in unicellular micro-organisms. Limnol.Oceanogr. 23: 1248–1255, doi:10.4319/lo.1978.23.6.1248
SMAYDA, T. J. 1997. Harmful algal blooms: Their ecophysiology andgeneral relevance to phytoplankton blooms in the sea. Limnol.Oceanogr. 42: 1137–1153, doi:10.4319/lo.1997.42.5_part_2.1137
STERNER, R., AND J. J. ELSER. 2002. Ecological stoichiometry.Princeton Univ. Press.
STOLTE, W., AND R. RIEGMAN. 1996. A model approach for size-selective competition of marine phytoplankton for fluctuatingnitrate and ammonium. J. Phycol. 32: 732–740, doi:10.1111/j.0022-3646.1996.00732.x
TAMBI, H., G. A. F. FLATEN, J. K. EGGE, G. BODTKER, A.JACOBSEN, AND T. F. THINGSTAD. 2009. Relationship betweenphosphate affinities and cell size and shape in various bacteriaand phytoplankton. Aquat. Microb. Ecol. 57: 311–320,doi:10.3354/ame01369
TANG, E. P. Y. 1995. The allometry of algal growth rates. J.Plankton Res. 17: 1325–1335, doi:10.1093/plankt/17.6.1325
TILMAN, D. 1982. Resource competition and community structure.Princeton Univ. Press.
WARTON, D. I., AND J. ORMEROD. 2007. smatr: (Standardised)major axis estimation and testing routines. R packageversion 2.1. Available from http://web.maths.unsw.edu.au/,dwarton
———, I. J. WRIGHT, D. S. FALSTER, AND M. WESTOBY. 2006.Bivariate line-fitting methods for allometry. Biol. Rev. 81:259–291, doi:10.1017/S1464793106007007
Associate editor: Jonathan J. Cole
Received: 04 October 2011Accepted: 03 January 2012Amended: 20 January 2012
566 Edwards et al.