alteration of membrane phospholipid bilayer organization...
TRANSCRIPT

Alteration of Membrane PhospholipidBilayer Organization in Human Erythrocytesduring Drug-induced Endocytosis
STANLEY L. SCHRIER, DANNYTSUN-YEE CHIU, MAGGIEYEE, KURTSIZER, andBERTRAMLUBIN, Stanford University School of Medicine, Stanford,California 94305; Bruce Lyon Memorial Research Laboratory, Children'sHospital Medical Center of Northern California, Oakland, California 94609
A B S T R A C T Our plan was to evaluate the potentiallyimportant role of phospholipids in erythrocyte shapealterations by determining if their orientation was al-tered during endocytosis. Stomatocytosis and endo-cytosis were induced in normal intact human eryth-rocytes by incubation with three agents: primaquine,vinblastine, and chlorpromazine, each of which has itsown requirements and time course for producing en-docytosis. The organization of the phospholipid bilayerwas assessed by measuring the extent of degradationof phophatidylcholine (PC), phophatidylethanolamine(PE), phosphatidylserine (PS), and sphingomyelin(SM) produced by exposure of erythrocytes to a non-penetrating protease-free phospholipase A2 alone orin combination with a purified sphingomyelinase aswell. The induction of stomatocytosis did not changethis orientation. However, correlating with the onsetof endocytosis but not its extent, there was an increaseip PE degradation, which could be detected regularlyonly by use of phospholipase A2 alone. Use of the com-bination of phospholipase A2 and sphingomyelinaseshowed that the extent and course of endocytosis wasparalleled by an apparent movement of PC and SMfrom the outer to the inner half of the lipid bilayer.Since no further PE was hydrolyzed and because noPS was ever degraded, this inward movement of PCand SM did not represent the establishment of com-plete symmetry in the membrane. By adjusting theexperimental design it was possible to implicate the
A preliminary report of this work was presented at theAnnual Meeting of the American Society of Hematology, 5-8 December 1981, and appeared in abstract form in 1981.Blood. 58(Suppl. 1) 34a. (Abstr.)
Address reprint requests to Dr. S. L. Schrier.Received for publication 29 July 1982 and in revised form
22 July 1983.
endocytic process, and not insertion of drug in themembrane, as the cause of the alterations in phospho-lipid organization seen. Our findings indicate that thephospholipid orientation is very closely involved in theendocytosis process and that specific states of phos-pholipid asymmetry may be related to identifiablemembrane events.
INTRODUCTION
The vectorial aspects of the human erythrocyte (RBC)'as well as its shape is thought to be determined, inpart, by the asymmetric distribution of proteins acrossthe membrane (1-3). Studies with both small molec-ular probes like trinitrobenzenesulfonate (4, 5) andnonpenetrating enzymes (6-8) provide generally con-sistent data showing that the phospholipids in the bi-layer are also asymmetrically arranged, with sphin-gomyelin (SM) and phosphatidylcholine (PC) beinglocated primarily in the outer leaflet while the ami-nophospholipids, phosphatidylethanolamine (PE) andphosphatidylserine (PS), being located at the innerleaflet. The negatively charged PS, in particular, seemsto be totally restricted to the inner half of the bilayer.
Wedecided to evaluate the potential role of phos-pholipids in RBCshape alterations by determining ifthe orientation of these phospholipids was altered dur-ing endocytosis. Endocytosis in intact adult RBCcanbe produced by incubation with amphipathic cationslike primaquine, chlorpromazine, and vinblastine (9).Each agent quickly produces stomatocytosis (10),
I Abbreviations used in this paper: PC, phosphatidylcho-line; PE, phosphatidylethanolamine; PS, phosphatidylserine;RBC, erythrocyte(s); SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; SM, sphingomyelin; vitB12, vitamin B12.
1698 J. Clin. Invest. © The American Society for Clinical Investigation, Inc. * 0021-9738/83/11/1698/08 $1.00Volume 72 November 1983 1698-1705

which is followed in a variable period of time by en-docytosis, a process that involves membrane invagi-nation, constriction, and fusion (11). These three drugshave very different requirements for endocytosis interms of their need for ATP and the anionic and cat-ionic contents of the incubation media (11, 12). Theentry and distribution of these agents into RBC hasalso been studied (13). By manipulating this systemand making serial measurements of endocytosis whiledetermining in parallel the accessibility of the con-stituents of the phospholipid bilayer to nonpenetratingenzymatic probes, we thought we might be able todetermine if changes in phospholipid organization ac-companied either stomatocytosis and/or endocytosis,and in particular, examine the effects of endocytosison phospholipid organization within the bilayer.
In evaluating phospholipid bilayer organization inRBC undergoing stomatocytosis and endocytosis, thecells were initially exposed to a purified protease-free,nonpenetrating bee venom phospholipase A2, whichdegrades only the glycerophospholipids located in theouter leaflet of the lipid bilayer. Usually 60% of thePC and 0-10% of the PE are hydrolyzed followingsuch treatment. More complete information aboutphospholipid orientation is provided by treating RBCfirst with phospholipase A2 and subsequently withsphingomyelinase. In addition to hydrolyzing SMthiscombined enzymatic treatment hydrolyzes all of thePC and PE located in the outer membrane leaflet.However, this attack does not degrade any PS, whichappears to be confined entirely to the inner leaflet.Based upon this combined enzymatic approach it isgenerally believed that 75% of PC, 20% of PE, and80% of SMare located in the outer leaflet of the bilayer.The results of our study indicate that significant al-terations in phospholipid organization accompany en-docytosis, but not stomatocytosis, and that particulardomains of PE and PC may be involved in the en-docytic process.
METHODSMaterials. The materials for measuring the extent of RBC
endocytosis (14), measuring [3H]chlorpromazine entry intoRBC(13), performing sodium dodecyl sulfate-polyacrylamidegel electrophoresis (SDS-PAGE) (15), and for measuringphospholipid asymmetry were obtained from previously de-scribed sources (5, 8).
Procedures. All experiments were performed on freshlydrawn heparinized venious blood provided by normal vol-unteer donors under protocols established and approved bythe Stanford Committee on Human Experimentation and bythe Oakland Children's lHospital Medical Center Committee.The plasma and buffy coat were removed and the plasmawas retained for subsequent use in the endocytosis experi-ments. The RBC were then washed, once at 4°C in 0.145M NaCl and three times with Hanks' balanced salt solution(HBSS), also at 4°C.
Measurement of stomatocytosis and endocytosis was per-formed as previously described (14). Briefly, 1 vol of washed,packed RBC in HBSS (hematocrit -75%) was mixed with1 vol of HBSScontaining the endocytosis-inducing agent and1 vol of autologous plasma containing [57Co]vitamin (vit) B12.Incubation was carried out at 37°C, for varying periods oftime. The trapping of the [57Co]vit B12-vit B12-binding pro-tein complex with endocytic vacuoles provided a quantita-tive measurement of endocytosis (14). Confirmation of en-docytosis and evaluation of shape change was routinely doneby phase-microscopic examination of coded samples. Whenexperiments were performed in isotonic sodium lactate orisotonic sodium glutamate, all washes were done with eitherisotonic sodium lactate or isotonic glutamate buffered to pH7.4. In such experiments, the autologous plasma had beenpreviously extensively dialyzed against the same isotonic lac-tate or glutamate supplemented with 5 mMCa2' and 2 mgglucose/ml. The appropriate lactate or glutamate solutionthen replaced the HBSS regularly used. The reaction mix-tures were shaken for 5 min at 37°C before the endocytosis-inducing drugs were added. Upon addition of drug, sampleswere immediately withdrawn and centrifuged for 2 min inthe Brinkmann 3200 centrifuge (Brinkmann Instruments,Inc., Westbury, NY) for initial evaluation of RBC shape,endocytosis, and phospholipid bilayer organization. Approx-imately 5 min were required for isolation of this initial sam-ple, which is called the "0 time sample." Measurement of[3H]chlorpromazine entry into RBCwas performed as pre-viously described (13).
Assessment of RBCmembrane phospholipid organizationwas performed by using phospholipase A2 from bee venomand sphingomyelinase C from Staphylococcus aureus (8). Inbrief, the 0.25-ml aliquot of drug treated or control RBCwas suspended in 5 ml of 5 mMphosphate-buffered saline(PBS) (290 mosM) containing 5 mMKCI, 1 mMMgSO4, and0.5 mMCaCl2, pH 7.4. For the single enzyme treatmentexperiments, 30 IU of phospholipase A2 from bee venom or10 IU of sphingomyelinase from S. aureus was added toappropriate samples and incubated at 37°C for 1 or 2 h. Forthe double-enzyme treatment experiments, appropriateerythrocyte samples were first incubated with 30 IU of phos-pholipase A2 from bee venom for 1 h, followed by incubationwith 10 IU of sphingomyelinase C for an additional hour.The degradation of phospholipid by phospholipase A2 or thecombination of phospholipase A2 and sphingomyelinase wasterminated by washing the RBC three times with PBS con-taining 5 mMEDTA. The extent of hemolysis was determinedat the end of each incubation before the EDTA wash bycomparing the hemoglobin content in the supernate of eachsample to that of a 100% hemolyzed control. Hemolysis wasfound to be <7% in all experiments reported in this paper.To determine the extent of phospholipid degradation, phos-pholipase-treated and control RBCsamples were washed withPBS and subjected to lipid extraction by the method of Roseand Oklander (16). Lipid extracts from each sample wereevaporated to dryness under nitrogen and redissolved in asmall volume (100-200 jl) of 2:1 chloroform/methanol mix-ture. Individual phospholipids were separated by the two-dimensional thin-layer chromatographic technique describedby Roelofsen and Zwaal (17). The individual lipid componentswere examined by staining with iodine vapor. All spots werescraped from the plate and transferred to test tubes, and thequantity of phospholipid was determined by measuring theamount of phosphorus in each spot, using the method ofBottcher et al. (18). The percentage of phospholipid hydro-lyzed after treatment of RBC with phospholipase A2 wasdetermined by measuring the ratio of remaining diacylglyc-
Phospholipid Bilayer Organization in Human RBC 1699
erophospholipid to the corresponding lyso derivative. Thespecificity of phospholipase A2 toward pure lipid substratesin an isolated system was not altered in the presence of eitherprimaquine, chlorpromazine, or vinblastine in the reactionmixture. For the determination of SMdegradation by sphin-gomyelinase, the absolute and relative quantity of SM re-covered from the sample was compared with the absoluteand relative quantity of SM recovered from the nontreatedcontrol sample. Since neither of these enzymes penetrate thelipid bilayer (17), hydrolysis of phospholipids can be inter-preted to indicate location in the outer leaflet of the bilayer.
RESULTS
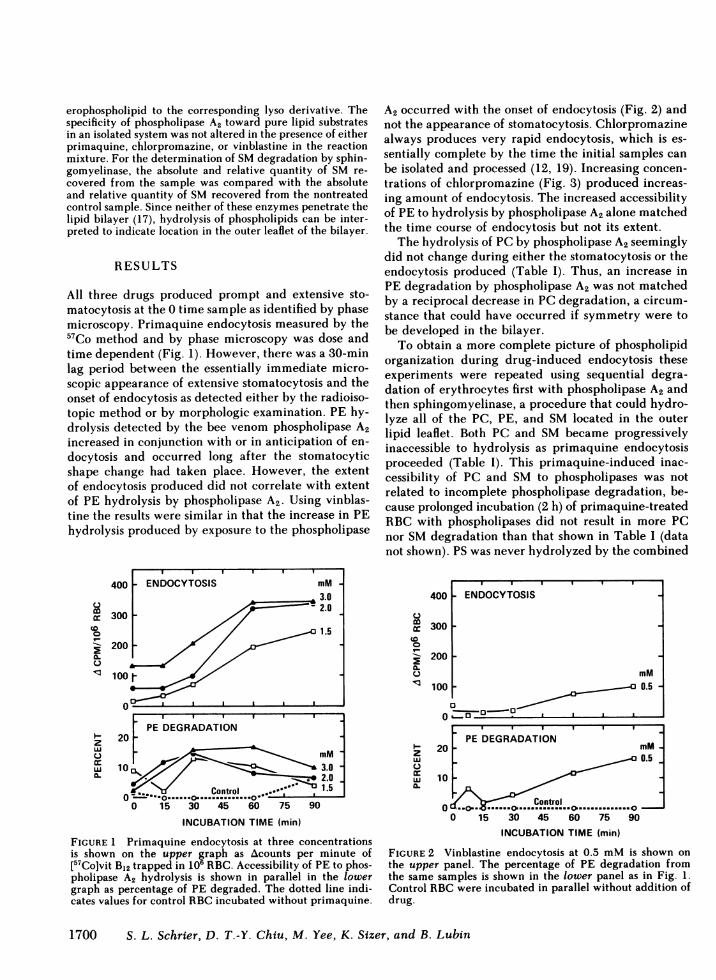
All three drugs produced prompt and extensive sto-matocytosis at the 0 time sample as identified by phasemicroscopy. Primaquine endocytosis measured by the57Co method and by phase microscopy was dose andtime dependent (Fig. 1). However, there was a 30-minlag period between the essentially immediate micro-scopic appearance of extensive stomatocytosis and theonset of endocytosis as detected either by the radioiso-topic method or by morphologic examination. PE hy-drolysis detected by the bee venom phospholipase A2increased in conjunction with or in anticipation of en-docytosis and occurred long after the stomatocyticshape change had taken place. However, the extentof endocytosis produced did not correlate with extentof PE hydrolysis by phospholipase A2. Using vinblas-tine the results were similar in that the increase in PEhydrolysis produced by exposure to the phospholipase
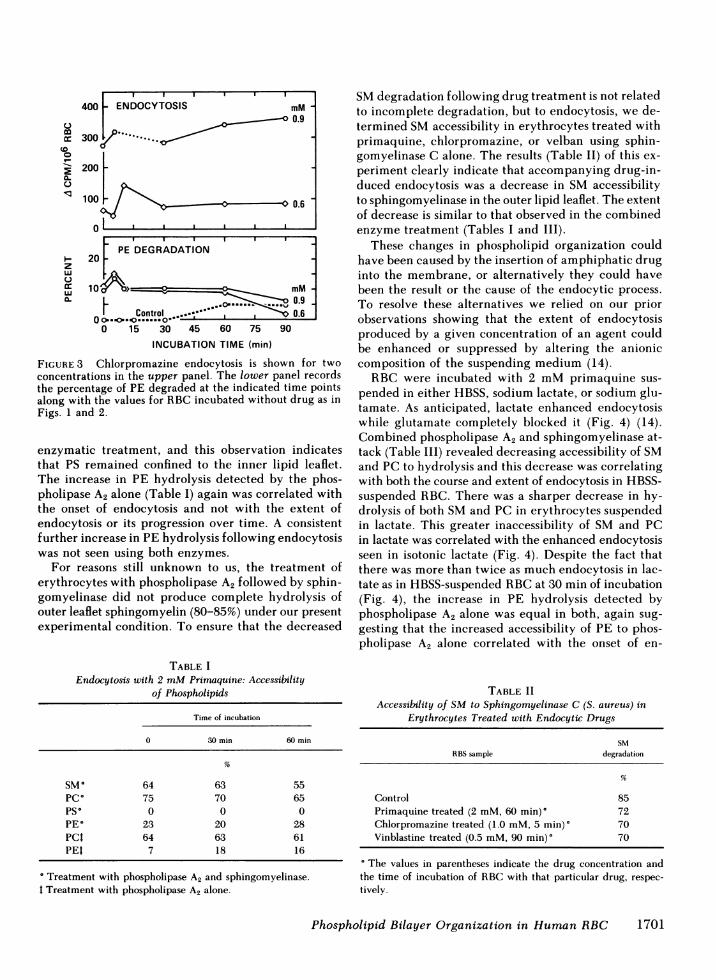
A2 occurred with the onset of endocytosis (Fig. 2) andnot the appearance of stomatocytosis. Chlorpromazinealways produces very rapid endocytosis, which is es-sentially complete by the time the initial samples canbe isolated and processed (12, 19). Increasing concen-trations of chlorpromazine (Fig. 3) produced increas-ing amount of endocytosis. The increased accessibilityof PE to hydrolysis by phospholipase A2 alone matchedthe time course of endocytosis but not its extent.
The hydrolysis of PCby phospholipase A2 seeminglydid not change during either the stomatocytosis or theendocytosis produced (Table I). Thus, an increase inPE degradation by phospholipase A2 was not matchedby a reciprocal decrease in PC degradation, a circum-stance that could have occurred if symmetry were tobe developed in the bilayer.
To obtain a more complete picture of phospholipidorganization during drug-induced endocytosis theseexperiments were repeated using sequential degra-dation of erythrocytes first with phospholipase A2 andthen sphingomyelinase, a procedure that could hydro-lyze all of the PC, PE, and SM located in the outerlipid leaflet. Both PC and SM became progressivelyinaccessible to hydrolysis as primaquine endocytosisproceeded (Table I). This primaquine-induced inac-cessibility of PC and SM to phospholipases was notrelated to incomplete phospholipase degradation, be-cause prolonged incubation (2 h) of primaquine-treatedRBC with phospholipases did not result in more PCnor SMdegradation than that shown in Table I (datanot shown). PS was never hydrolyzed by the combined
PE DEGRADATION20
10 3.0 -
Control ..0.0 .
0 ". ....o .......nto...o... 0L- ----**- L.50 15 30 45 60 75 90
INCUBATION TIME (min)
FIGURE 1 Primaquine endocytosis at three concentrationsis shown on the upper graph as Acounts per minute of[57Co]vit B12 trapped in 106 RBC. Accessibility of PE to phos-pholipase A2 hydrolysis is shown in parallel in the lowergraph as percentage of PE degraded. The dotted line indi-cates values for control RBCincubated without primaquine.
400
0
mccCo0
I-
o
uJ
zwU0)
cc
0.I
300
200
100
I I I I I
PE DEGRADATION20 - M
-~~~~~~~~~~0.5 _
10
Control0 15 30 45 60 75 90
INCUBATION TIME (min)
FIGURE 2 Vinblastine endocytosis at 0.5 mMis shown on
the upper panel. The percentage of PE degradation fromthe same samples is shown in the lower panel as in Fig. 1.Control RBCwere incubated in parallel without addition ofdrug.
1700 S. L. Schrier, D. T.-Y. Chiu, M. Yee, K. Sizer, and B. Lubin
m
o
ccCD
0
w.
02
- ENDOCYTOSIS
mMa 0.5 -
- n0 O
400
0.
U-
300
200 [
100
0
zw
u
w
0L
20PE DEGRADATION
10<̂6> ~~~mM-I
Control0-<- ...o
. 0.6___
0 15 30 45 60 75 90
INCUBATION TIME (min)
FIGURE 3 Chlorpromazine endocytosis is shown for twoconcentrations in the upper panel. The lower panel recordsthe percentage of PE degraded at the indicated time pointsalong with the values for RBC incubated without drug as inFigs. 1 and 2.
enzymatic treatment, and this observation indicatesthat PS remained confined to the inner lipid leaflet.The increase in PE hydrolysis detected by the phos-pholipase A2 alone (Table I) again was correlated withthe onset of endocytosis and not with the extent ofendocytosis or its progression over time. A consistentfurther increase in PE hydrolysis following endocytosiswas not seen using both enzymes.
For reasons still unknown to us, the treatment oferythrocytes with phospholipase A2 followed by sphin-gomyelinase did not produce complete hydrolysis ofouter leaflet sphingomyelin (80-85%) under our presentexperimental condition. To ensure that the decreased
TABLE IEndocytosis with 2 mMPrinaquine: Accessibility
of Phospholipids
Time of incubation
0 30 min 60 min
SM° 64 63 55PC° 75 70 65PS* 0 0 0PE* 23 20 28PCI 64 63 61PEt 7 18 16
Treatment with phospholipase A2 and sphingomyelinase.I Treatment with phospholipase A2 alone.
SMdegradation following drug treatment is not relatedto incomplete degradation, but to endocytosis, we de-termined SMaccessibility in erythrocytes treated withprimaquine, chlorpromazine, or velban using sphin-gomyelinase C alone. The results (Table II) of this ex-periment clearly indicate that accompanying drug-in-duced endocytosis was a decrease in SM accessibilityto sphingomyelinase in the outer lipid leaflet. The extentof decrease is similar to that observed in the combinedenzyme treatment (Tables I and III).
These changes in phospholipid organization couldhave been caused by the insertion of amphiphatic druginto the membrane, or alternatively they could havebeen the result or the cause of the endocytic process.To resolve these alternatives we relied on our priorobservations showing that the extent of endocytosisproduced by a given concentration of an agent couldbe enhanced or suppressed by altering the anioniccomposition of the suspending medium (14).
RBC were incubated with 2 mMprimaquine sus-pended in either HBSS, sodium lactate, or sodium glu-tamate. As anticipated, lactate enhanced endocytosiswhile glutamate completely blocked it (Fig. 4) (14).Combined phospholipase A2 and sphingomyelinase at-tack (Table III) revealed decreasing accessibility of SMand PC to hydrolysis and this decrease was correlatingwith both the course and extent of endocytosis in HBSS-suspended RBC. There was a sharper decrease in hy-drolysis of both SMand PC in erythrocytes suspendedin lactate. This greater inaccessibility of SM and PCin lactate was correlated with the enhanced endocytosisseen in isotonic lactate (Fig. 4). Despite the fact thatthere was more than twice as much endocytosis in lac-tate as in HBSS-suspended RBCat 30 min of incubation(Fig. 4), the increase in PE hydrolysis detected byphospholipase A2 alone was equal in both, again sug-gesting that the increased accessibility of PE to phos-pholipase A2 alone correlated with the onset of en-
TABLE IIAccessibility of SM to Sphingomyelinase C (S. aureus) in
Erythrocytes Treated with Endocytic Drugs
SMRBSsample degradation
Control 85Primaquine treated (2 mM, 60 min)' 72Chlorpromazine treated (1.0 mM, 5 min)' 70Vinblastine treated (0.5 mM, 90 min)' 70
The values in parentheses indicate the drug concentration andthe time of incubation of RBCwith that particular drug, respec-tively.
Phospholipid Bilayer Organization in Human RBC
- ENDOCYTOSIS mM0.9
(P*.6.0
1701
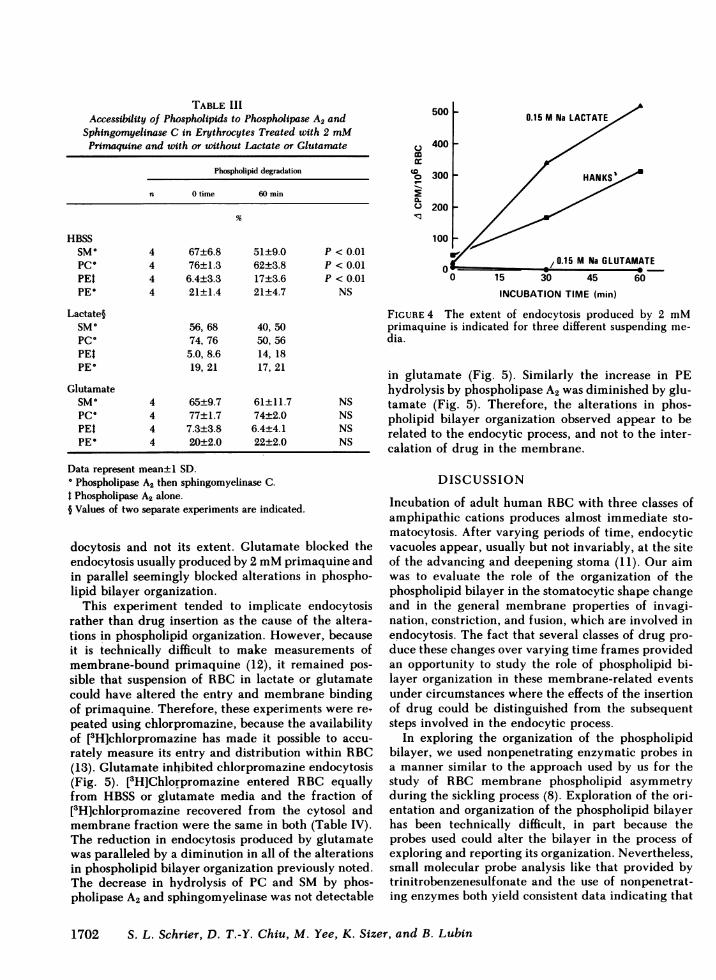
TABLE IIIAccessibility of Phospholipids to Phospholipase A2 and
Sphingomyelinase C in Erythrocytes Treated with 2 mMPrimaquine and with or without Lactate or Glutamate
Phospholipid degradation
n 0 time 60 min
HBSSSM° 4 67±6.8 51±9.0 P < 0.01PC" 4 76±1.3 62±3.8 P < 0.01PEt 4 6.4±3.3 17±3.6 P < 0.01PE* 4 21±1.4 21±4.7 NS
Lactate§SM° 56, 68 40, 50PC* 74, 76 50, 56PEt 5.0, 8.6 14, 18PE* 19, 21 17, 21
GlutamateSM° 4 65±9.7 61±11.7 NSPC* 4 77±1.7 74±2.0 NSPEt 4 7.3±3.8 6.4±4.1 NSPE 4 20±2.0 22±2.0 NS
Data represent mean±1 SD." Phospholipase A2 then sphingomyelinase C.t Phospholipase A2 alone.§ Values of two separate experiments are indicated.
docytosis and not its extent. Glutamate blocked theendocytosis usually produced by 2 mMprimaquine andin parallel seemingly blocked alterations in phospho-lipid bilayer organization.
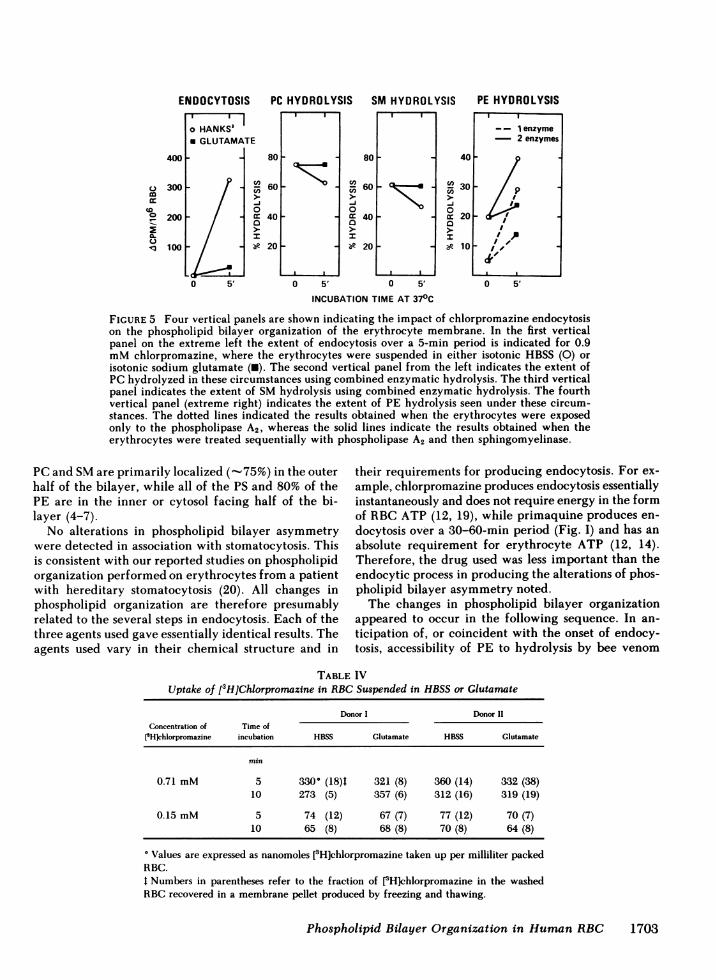
This experiment tended to implicate endocytosisrather than drug insertion as the cause of the altera-tions in phospholipid organization. However, becauseit is technically difficult to make measurements ofmembrane-bound primaquine (12), it remained pos-sible that suspension of RBC in lactate or glutamatecould have altered the entry and membrane bindingof primaquine. Therefore, these experiments were re-peated using chlorpromazine, because the availabilityof [3HJchlorpromazine has made it possible to accu-rately measure its entry and distribution within RBC(13). Glutamate inhibited chlorpromazine endocytosis(Fig. 5). [3H]Chlorpromazine entered RBC equallyfrom HBSS or glutamate media and the fraction of[3H]chlorpromazine recovered from the cytosol andmembrane fraction were the same in both (Table IV).The reduction in endocytosis produced by glutamatewas paralleled by a diminution in all of the alterationsin phospholipid bilayer organization previously noted.The decrease in hydrolysis of PC and SM by phos-pholipase A2 and sphingomyelinase was not detectable
500 0.15 M Na LACTATE
400 -
0 300 - / HANKS
200 -/
,0.15 M Na GLUTAMATE
0 15 30O 45 60
INCUBATION TIME (min)
FIGURE 4 The extent of endocytosis produced by 2 mMprimaquine is indicated for three different suspending me-dia.
in glutamate (Fig. 5). Similarly the increase in PEhydrolysis by phospholipase A2 was diminished by glu-tamate (Fig. 5). Therefore, the alterations in phos-pholipid bilayer organization observed appear to berelated to the endocytic process, and not to the inter-calation of drug in the membrane.
DISCUSSION
Incubation of adult human RBCwith three classes ofamphipathic cations produces almost immediate sto-matocytosis. After varying periods of time, endocyticvacuoles appear, usually but not invariably, at the siteof the advancing and deepening stoma (11). Our aimwas to evaluate the role of the organization of thephospholipid bilayer in the stomatocytic shape changeand in the general membrane properties of invagi-nation, constriction, and fusion, which are involved inendocytosis. The fact that several classes of drug pro-duce these changes over varying time frames providedan opportunity to study the role of phospholipid bi-layer organization in these membrane-related eventsunder circumstances where the effects of the insertionof drug could be distinguished from the subsequentsteps involved in the endocytic process.
In exploring the organization of the phospholipidbilayer, we used nonpenetrating enzymatic probes ina manner similar to the approach used by us for thestudy of RBC membrane phospholipid asymmetryduring the sickling process (8). Exploration of the ori-entation and organization of the phospholipid bilayerhas been technically difficult, in part because theprobes used could alter the bilayer in the process ofexploring and reporting its organization. Nevertheless,small molecular probe analysis like that provided bytrinitrobenzenesulfonate and the use of nonpenetrat-ing enzymes both yield consistent data indicating that
1702 S. L. Schrier, D. T.-Y. Chiu, M. Yee, K. Sizer, and B. Lubin
SM HYDROLYSIS
400
mcoCD
to0a-C-
300 _
200 _
100 _
80-
cn 6vs60-J0D 40
I* 20
IFIF
0 5'
80 FCd,
-J0
0I
60 -
40-
20 -
0 5'
401
-J0
0a
P
o-
301
20
10
- - 1 enzyme2 enzymes
II )
I/
0 .50 5'
INCUBATION TIME AT 370C
FIGURE 5 Four vertical panels are shown indicating the impact of chlorpromazine endocytosison the phospholipid bilayer organization of the erythrocyte membrane. In the first verticalpanel on the extreme left the extent of endocytosis over a 5-min period is indicated for 0.9mMchlorpromazine, where the erythrocytes were suspended in either isotonic HBSS (0) orisotonic sodium glutamate (U). The second vertical panel from the left indicates the extent ofPC hydrolyzed in these circumstances using combined enzymatic hydrolysis. The third verticalpanel indicates the extent of SMhydrolysis using combined enzymatic hydrolysis. The fourthvertical panel (extreme right) indicates the extent of PE hydrolysis seen under these circum-stances. The dotted lines indicated the results obtained when the erythrocytes were exposedonly to the phospholipase A2, whereas the solid lines indicate the results obtained when theerythrocytes were treated sequentially with phospholipase A2 and then sphingomyelinase.
PCand SMare primarily localized (-75%) in the outerhalf of the bilayer, while all of the PS and 80% of thePE are in the inner or cytosol facing half of the bi-layer (4-7).
No alterations in phospholipid bilayer asymmetrywere detected in association with stomatocytosis. Thisis consistent with our reported studies on phospholipidorganization performed on erythrocytes from a patientwith hereditary stomatocytosis (20). All changes inphospholipid organization are therefore presumablyrelated to the several steps in endocytosis. Each of thethree agents used gave essentially identical results. Theagents used vary in their chemical structure and in
their requirements for producing endocytosis. For ex-ample, chlorpromazine produces endocytosis essentiallyinstantaneously and does not require energy in the formof RBCATP (12, 19), while primaquine produces en-docytosis over a 30-60-min period (Fig. I) and has anabsolute requirement for erythrocyte ATP (12, 14).Therefore, the drug used was less important than theendocytic process in producing the alterations of phos-pholipid bilayer asymmetry noted.
The changes in phospholipid bilayer organizationappeared to occur in the following sequence. In an-ticipation of, or coincident with the onset of endocy-tosis, accessibility of PE to hydrolysis by bee venom
TABLE IVUptake of [3H]Chlorpromazine in RBCSuspended in HBSSor Glutamate
Donor I Donor 11Concentration of Time of
['H]chlorpromazine incubation HBSS Glutamate HBSS Glutamate
min
0.71 mM 5 330 (18)4 321 (8) 360 (14) 332 (38)10 273 (5) 357 (6) 312 (16) 319 (19)
0.15 mM 5 74 (12) 67 (7) 77 (12) 70 (7)10 65 (8) 68 (8) 70 (8) 64 (8)
Values are expressed as nanomoles [3H]chlorpromazine taken up per milliliter packedRBC.t Numbers in parentheses refer to the fraction of [3Hlchlorpromazine in the washedRBC recovered in a membrane pellet produced by freezing and thawing.
Phospholipid Bilayer Organization in Human RBC
ENDOCYTOSIS
HANKS' I
PC HYDROLYSISI I I I I l I
170t3
PE HYDROLYSIS

phospholipase A2 increased. The detection of this in-crease in PE accessibility was regularly demonstratedwhen the phospholipase A2 probe was used alone undercircumstances where usually 0-10% of PE is accessible.The enhancement of PE accessibility was not regularlyseen when the combined enzymatic treatment withphospholipase A2 and sphingomyelinase was used (Ta-ble I). Under these latter circumstances 20-25% of PEis accessible to enzymatic degradation in control eryth-rocytes and this amount is not increased further as aconsequence of endocytosis. This observation sug-gested that a subclass of PE in the outer half of thebilayer became exposed coincident with endocytosisand that use of combined enzymatic attack might haveobliterated the selectivity that allowed the detectionof the PE subclass involved. The idea that there maybe subpopulations of PE within the membrane is sup-ported by a recent report (21) showing that a smallsubpopulation of PE in the outer leaflet had a muchhigher turnover rate than the remainder of PE. Theextent of PE accessibility to the phospholipase A2 wasnot paralleled by the extent of endocytosis produced.Therefore, the increase in PE accessibility appears toreflect the trigger or switch that accompanies the onsetof endocytosis. Perhaps the PE that becomes accessibleto the phospholipase A2 may be all of the PE in thatsubclass, which is made available in an all or nonemanner in association with the onset of endocytosis.
In contrast to the behavior of PE, both externallyoriented PC and SM became similarly less accessiblewith the onset of endocytosis, and the decrease in ac-cessibility paralleled the extent of endocytosis, becom-ing progressively inaccessible as endocytosis contin-ued. This reduction in PCand SMaccessibility was notcaused by drug intercalation alone, because in the faceof normal uptake and distribution of [3H]chlor-promazine, but with endocytosis blocked by glutamatethese alterations did not occur (Fig. 5 and Table IV).
The decreased accessibility of PC and SM to deg-radation could have been the result of bulk inacces-sibility of membrane phospholipids trapped in endo-cytic vacuoles, which were now in the cytosol, shieldedfrom the hydrolytic enzymes by the intact erythrocyteplasma membrane. However, "trapping" cannot ac-count for all the observations we made. Firstly, usingphospholipase A2 alone in our system did not revealany decrease in PC degradation (Table I). Secondly,although the combined enzyme treatment showed de-creased PC and SM degradation, PE degradation re-mained fairly constant (Table III). Lastly, if trappedin the endocytic vesicles, the phospholipases could stillbe active in degrading phospholipids. Taken together,these observations suggest that the decreased PC andSM degradation by combined enzymatic treatment(Table III) should not be explained as a simple con-
sequence of bulk inaccessibility of membrane phos-pholipids trapped in endocytic vacuoles.
Although the progressive inaccessibility of both SMand PC during endocytosis could reflect PC and SMmovement toward the inner half of the lipid bilayer,such movement of PC was only detected by combinedenzymatic degradation (Tables I-III) and not whenphospholipase A2 was used alone (Table I). Therefore,it is likely that endocytosis involves a subclass of PCthat interacts with SM, and there is new evidence thatthere are domains of PC in human erythrocyte mem-branes (22). However, because this progressive loss inaccessibility of SM and PC that accompanied endo-cytosis was not matched by a continuing increase inaccessibility of PE, nor by any accessibility of PS, thesechanges do not represent complete loss of bilayer asym-metry.
Furthermore, the continued inability to detect anyPS accessibility argues that the interrelationship be-tween spectrin and PS is not perturbed during endo-cytosis, in contrast to the increase in PS accessibilityseen with sickling (8) or oxidant damage (23, 24),where there probably is alteration of this relationship(23, 24). Wesearched for evidence of spectrin cross-linking using nonreducing 6%SDS-PAGEgels (23, 24)and found none (data not shown).
During the invasion of rhesus RBCby Plasmodiumknowlesi it was noted that there was a decrease inaccessibility of PC while PE became more accessibleto the phospholipase A2 used. PS remained completelyinaccessible to either chemical or enzymatic probesduring the parasitic invasion (25). These results arequite similar to ours. It is known that the process ofplasmodial invasion of RBCrequires the fixation of themalarial parasite to a specific membrane receptor site,following which there is a convulsive endocytosis ofthe entire merozoite (26). On the basis of our obser-vations we would conclude that the changes reportedin parasitized RBC reflect the endocytic process bywhich the malarial parasite entered the erythrocyte.
In summary, there are striking alterations in themembrane phospholipid bilayer organization that occurduring endocytosis but not stomatocytosis. The changesseen are not simply produced by the intercalation ofdrug into the membrane. There is an increase in ac-cessibility to bee venom phospholipase A2 of a subclassof PE that precedes endocytosis and does not correlatewith its extent or progression. Then, correlating withthe extent and progression of endocytosis, there appearsto be an inward movement of SMalong with a subclassof PC, which is probably closely associated with SM.This inward movement of SMand PCdoes not indicatecomplete loss of bilayer asymmetry, since there is nooutward movement of PS.
The importance of these observations relates to sev-
1704 S. L. Schrier, D. T.-Y. Chiu, M. Yee, K. Sizer, and B. Lubin

eral aspects of RBC membrane biology. Our resultssupport the concept, based on chemical data, (21, 22)that erythrocyte membrane PC and PE do not rep-resent single pools of phospholipid, but each is prob-ably made up of subclasses of these compounds resid-ing in specific domains. Furthermore, our studies im-ply that these putative subclasses of PE and PC havefunctional importance. Recent evidence suggests thata critical step involved in membrane fusion (an im-portant step in endocytosis) is the hexagonal phasetransition within the bilayer structure (27). Subclassesof phospholipids may be involved in this hexagonalform, and such changes may affect accessibility ofmembrane phospholipids to phospholipase degrada-tion as a consequence of involvement in the endocyticprocess. Our data also shows that membrane phospho-lipids can no longer be considered as existing only intwo organizations, asymmetrical and symmetrical. Infact, there may be several levels of asymmetry reflect-ing a variety of membrane events, and we believe wehave identified one such state and its association withendocytosis.
ACKNOWLEDGMENTS
This work was supported in part by grants AM-13682, HL-27059, Bio-Medicine Research support grant RR-05467, Na-tional Institutes of Health, Department of Health, Educa-tion, and Welfare, and March of Dimes grant 1-768.
REFERENCES1. Bennett, V. 1982. The molecular basis for membrane-
cytoskeleton association in human erythrocytes. J. Cell.Biochem. 18:49-65.
2. Lux, S. E. 1979. Spectrin-actin membrane skeleton ofnormal and abnormal red blood cells. Semin. Hematol.16:21-51.
3. Palek, J., and S. C. Liu. 1979. Dependence of spectrinorganization in red blood cell membranes on cell me-tabolism: implications for control of red cell shape, de-formability, and surface area. Semin. Hematol. 16:75-93.
4. Gordesky, S. E., G. V. Marinetti, and R. Love. 1975. Thereaction of chemical probes with the erythrocyte mem-brane. J. Membr. Biol. 20:111-132.
5. Chiu, D., B. Lubin, and S. Shohet. 1979. Erythrocytemembrane lipid reorganization during the sickling pro-cess. Br. J. Haematol. 41:223-234.
6. Zwaal, R. F. A., B. Roelofsen, P. Comfurius, andL. L. M. Van Deenen. 1975. Organization of phospho-lipids in human red cell membranes as detected by ac-tion of various purified phospholipases. Biochim. Bio-phys. Acta. 406:83-86.
7. Martin, J. K., M. G. Luthra, M. A. Wells, R. P. Watts,and D. J. Hanahan. 1975. Phospholipase A2 as a probeof phospholipid distribution in erythroid membrane.Factors influencing the apparent specificity of the re-action. Biochemistry. 14:5400-5408.
8. Lubin, B., D. Chiu, J. Bastacky, B. Roelofsen, andL. L. M. Van Deenen. 1981. Abnormalities in membrane
phospholipid organization in sickled erythrocytes. J.Clin. Invest. 67:1643-1649.
9. Ben-Bassat, I., K. G. Bensch, and S. L. Schrier. 1972.Drug-induced erythrocyte membrane internalization. J.Clin. Invest. 51:1833-1844.
10. Deuticke, B. 1968. Transformation and restoration ofbiconcave shape of human erythrocytes induced by am-phiphilic agents and changes of ionic environment.Biochim. Biophys. Acta. 163:494-500.
11. Schrier, S. L., B. Hardy, and K. G. Bensch. 1979. En-docytosis in erythrocytes and their ghosts. Normal andabnormal red cell membranes. Alan R. Liss, Inc., NewYork. 437-499.
12. Schrier, S. L., I. Junga, J. Krueger, and M. Johnson. 1978.Requirements of drug-induced endocytosis by intacthuman erythrocytes. Blood Cells. 4:339-353.
13. Schrier, S. L., and K. Junga. 1981. Entry and distributionof chlorpromazine and vinblastine into human eryth-rocytes during endocytosis. Proc. Soc. Exp. Biol. Med.168:159-167.
14. Schrier, S. L., I. Junga, and M. Seeger. 1974. The mech-anism of drug-induced erythrocyte vacuole formation.J. Lab. Clin. Med. 83:215-227.
15. Schrier, S. L., B. Hardy, I. Junga, and L. Ma. 1981.Actin-activated ATPase in human red cell membranes.Blood. 58:953-962.
16. Rose, H. G., and M. Oklander. 1965. Improved proce-dures for the extraction of lipids from human erythro-cytes. J. Lipid Res. 6:428-431.
17. Roelofsen, B., and R. F. A. Zwaal. 1976. The use ofphospholipases in the determination of asymmetricphospholipid distribution in membrane. Methods Membr.Biol. 7:147-177.
18. Bottcher, C. J. F., C. M. van Gent, and C. Pries. 1961.A rapid and sensitive sub-micro-phosphorus determi-nation. Anal. Chim. Acta. 24:203-208.
19. Feo, C., and N. Mohandas. 1977. Clarification of roleof ATP in red cell morphology and function. Nature(Lond.). 265:165-166.
20. Lubin, B., and D. Chiu. 1982. Membrane phospholipidorganization in pathologic human erythrocytes. Prog.Clin. Biol. Res. 97:137-150.
21. Marinetti, G. V., and K. Cattieu. 1982. Asymmetricmetabolism of phosphatidylethanolamine in the humanred cell membrane. J. Biol. Chem. 257:245-248.
22. Shukla, S. D., and D. J. Hanahan. 1982. Identificationof domains of phosphatidylcholine in human erythro-cyte plasma membranes. J. Biol. Chem. 257:2908-2911.
23. Haest, C. W. M., and B. Deuticke. 1976. Possible rela-tionship between membrane proteins and phospholipidasymmetry in the human erythrocyte membrane.Biochim. Biophys. Acta. 436:353-365.
24. Haest, C. W. M., G. Plasa, D. Kamp, and B. Deuticke.1978. Spectrin as a stabilizer of the phospholipid asym-metry in the human erythrocyte membrane. Biochim.Biophysica Acta. 509:21-32.
25. Gupta, C. M., and G. C. Mishra. 1981. Transbilayerphospholipid asymmetry in Plasmodium knowlesi-in-fected host cell membrane. Science (Wash. DC).212:1047-1049.
26. Miller, L. H., F. M. McAuliffe, and J. G. Johnson. 1979.Invasion of erythrocytes by malaria merozoites. Normaland abnormal red cell membrane. Alan R. Liss, Inc.,New York. 497-502.
27. Cullis, P. R., and M. H. Hope. 1978. Effects of fusogenicagents on membrane structure of erythrocyte ghosts andthe mechanism of membrane fusion. Nature (Lond.).276:672-674.
Phospholipid Bilayer Organization in Human RBC 1705