altered cell tropism and cytopathicity of feline immunodeficiency
TRANSCRIPT

JOURNAL OF VIROLOGY, June 1992, p. 3893-38980022-538X/92/063893-06$02.00/0Copyright © 1992, American Society for Microbiology
Altered Cell Tropism and Cytopathicity of Feline ImmunodeficiencyViruses in Two Different Feline CD4-Positive,
CD8-Negative Cell LinesKENZO TOKUNAGA,"2 YOSHII NISHINO,' HIROSHI OIKAWA,3 CHIAKI ISHIHARA,2
TAKESHI MIKAMI,4 AND KAZUYOSHI IKUTAl*
Section of Serology, Institute of Immunological Science, Hokkaido University, Kita-ku, Sapporo 060,1Department of Laboratory Animal Research, School of Veterinary Medicine, Rakuno GakuenUniversity, Ebetsu 069,2 Aburahi Laboratories, Shionogi & Co., Ltd., Koka, Shiga 520-34,
and Department of Veterinary Microbiology, Faculty ofAgriculture,University of Tokyo, Bunkyo-ku, Tokyo 113,4 Japan
Received 18 November 1991/Accepted 10 March 1992
Two interleukin-2-dependent feline CD4-positive and CD8-negative cell lines, MYA-1 and the newlyestablished FeL-039, were used as host cells for infection with feline immunodeficiency virus (FIV). All FIVstrains used, the Petaluma strain and several new isolates, were highly cytopathic to MYA-1. In contrast, thekinetics of FIN replication in FeL-039 differed greatly depending on the strain tested, i.e., noninfectious strain,highly cytopathic strain, and less cytopathic strain producing a persistent state for a long period. It appears,
therefore, that cell tropism for FIV differed with each FIV strain tested even in T-cell lines showing similar cellsurface phenotypes. Cytopathicity of FIV is evidently due to both the FIV strain and the host T cell.
Feline immunodeficiency virus (FIV), a recently isolatedvirus from domestic cats (12), is more closely related tohuman irnmunodeficiency virus (HIV) and simian immuno-deficiency virus of the lentiviruses than to oncoviruses (12,23) but is antigenically distant from any primate lentivirus(16, 23) and is highly T lymphotropic. Replication of FIV inestablished cell lines has been achieved by using non-T4 celllines, such as Crandell feline kidney cells and feline lympho-blastoid (FL74) cells chronically infected with feline leuke-mia virus (FeLV) (12, 22). These infections are generally notcytopathic (23). In contrast, a continuous feline cell line, T4thymic lymphoma 3201, was highly susceptible to the cyto-pathic effect of FIV (19). Similarly, an interleukin-2-depen-dent feline lymphoblastoid cell line, designated MYA-1,which was positive for feline CD4 (fCD4) and negative forfCD8 (8), was also highly sensitive to replication of FIV (7,10). Interleukin-2-independent feline T-cell lines chronicallyinfected with FIV have been developed by selecting cellsfrom interleukin-2-dependent cultures of mixed peripheralblood lymphocytes infected with FIV (21). In addition, itwas recently shown that two subsets of fCD4+ and fCD8+ Tcells were sensitive for the productive infection of thePetaluma strain, although the reverse transcriptase (RT)production after infection with a new FIV isolate was 4- to11-fold higher in supernatants from the CD4+ clone than inthose from the CD8+ clone (2). Thus, the determinants ofcell tropism and cytopathicity for FIV are still not clear. Inthe present study, we compared the kinetics of FIV replica-tion by using two fCD4+ and fCD8- T-cell lines infectedwith several FIV strains.A new cell line, designated FeL-039, was established by
culturing the peripheral blood mononuclear cells from a malecat by the method of Miyazawa et al. (10). The FeL-039 cellline originated from a cat that was Western immunoblotantibody negative against FIV-Petaluma and FeLV-FL74
* Corresponding author.
(12, 17) (data not shown). The peripheral blood mononuclearcell fraction prepared by centrifugation through Ficoll-Paquewas treated with 5 ,ug of phytohemagglutinin per ml for 2days and cultured in complete medium (RPMI 1640 mediumcontaining 10% fetal bovine serum, 50 ,uM 2-mercaptoetha-nol, 2 ,ug of Polybrene per ml, and 100 U of recombinanthuman interleukin-2 per ml) in a humidified atmosphere of5% CO2 in air. The cells were of the lymphoblastic type andcontinuously proliferated at a slightly higher rate thanMYA-1 cells. Flow cytometry with murine monoclonalantibodies (MAbs) to fCD4, fCD8, and feline Pan T Cell(Southern Biotechnology Associates, Inc., Birmingham,Ala.) showed that most of the FeL-039 cells expressed fCD4and Pan T, but not fCD8, antigens (Fig. 1A). A similarphenotype was also observed in MYA-1 cells (Fig. 1B) as
reported previously (8). Cell smears of FeL-039, which werefixed with cold acetone, did not show any specific reactionsby the indirect immunofluorescence (IF) test with rabbitanti-FeLV, rabbit anti-feline syncytium-forming virus(FeSFV), the serum from an FIV-seropositive cat, and MAbto FIV p241ag (F17) (lla). F17 MAb, the subclass of whichis an immunoglobulin G2b, was obtained by using FIVp5Jgas precursor protein expressed by baculovirus vector(11) as an immunogen.
Several Japanese FIV strains were newly isolated bycoculturing peripheral blood mononuclear cells from catsseropositive for FIV but seronegative for FeLV with MYA-1cells by the method described previously (9). Virus isolationwas determined by the appearance of RT activity in theculture fluid of the coculture. FIV-specific RT activity was
measured by using a reaction mixture containing poly(rA)oligo(dT), [32P]dTTP (800 Ci/mmol; New England Nuclear),and 4.95 mM MgCl2, as described previously (20). A total ofsix isolates (Gyu, 006, 048, 126, 127, and 148) were obtainedfrom 15 FIV-seropositive cats. The MYA-1 cells infectedwith these isolates were smeared, fixed with cold acetone,and then subjected to IF tests. The serum from an FIV-seropositive cat and F17 MAb gave strong reactions,
3893
Vol. 66, No. 6
on April 14, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
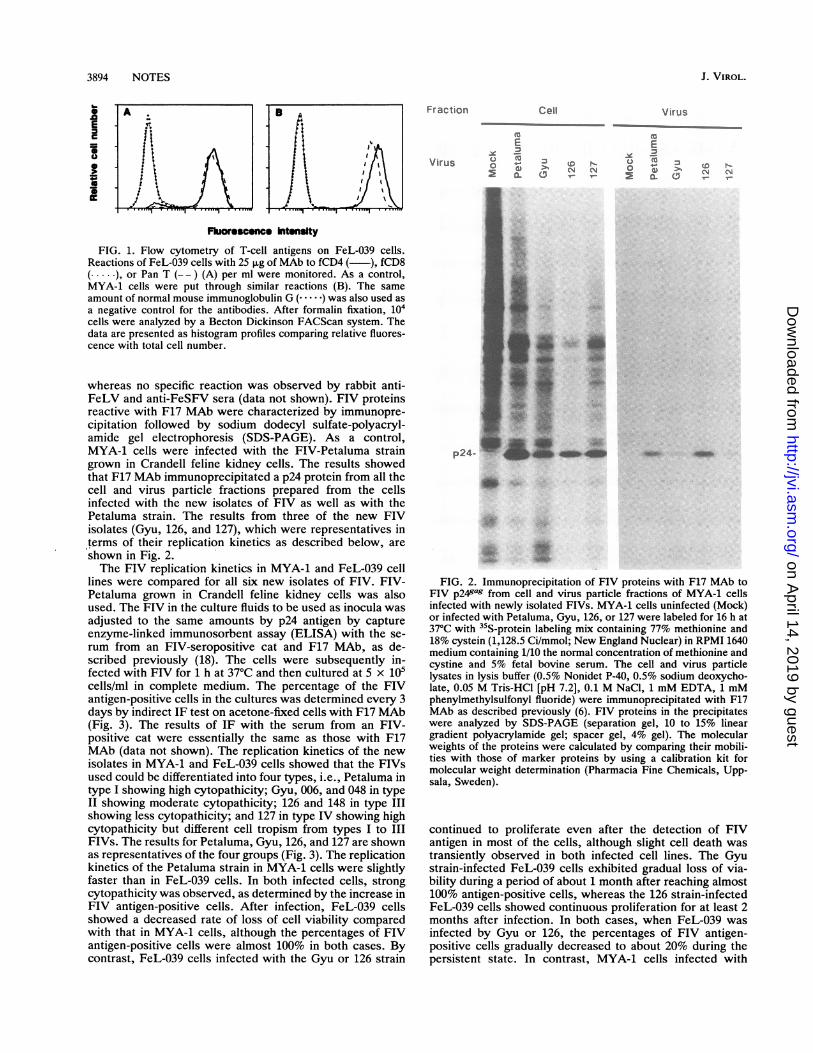
3894 NOTES
E-
B. A
£11X~~~~~~~~~~~~~~~J. VIROL.
Fraction
EY 'EVirus o <X)o
0.
Fluorescence hItenslty
FIG. 1. Flow cytometry of T-cell antigens on FeL-039 cells.Reactions of FeL-039 cells with 25 pLg of MAb to fCD4 ( ), fCD8( ), or Pan T (-- ) (A) per ml were monitored. As a control,MYA-1 cells were put through similar reactions (B). The sameamount of normal mouse immunoglobulin G (. ) was also used asa negative control for the antibodies. After formalin fixation, 104cells were analyzed by a Becton Dickinson FACScan system. Thedata are presented as histogram profiles comparing relative fluores-cence with total cell number.
whereas no specific reaction was observed by rabbit anti-FeLV and anti-FeSFV sera (data not shown). FIV proteinsreactive with F17 MAb were characterized by immunopre-cipitation followed by sodium dodecyl sulfate-polyacryl-amide gel electrophoresis (SDS-PAGE). As a control,MYA-1 cells were infected with the FIV-Petaluma straingrown in Crandell feline kidney cells. The results showedthat F17 MAb immunoprecipitated a p24 protein from all thecell and virus particle fractions prepared from the cellsinfected with the new isolates of FIV as well as with thePetaluma strain. The results from three of the new FIVisolates (Gyu, 126, and 127), which were representatives interms of their replication kinetics as described below, areshown in Fig. 2.The FIV replication kinetics in MYA-1 and FeL-039 cell
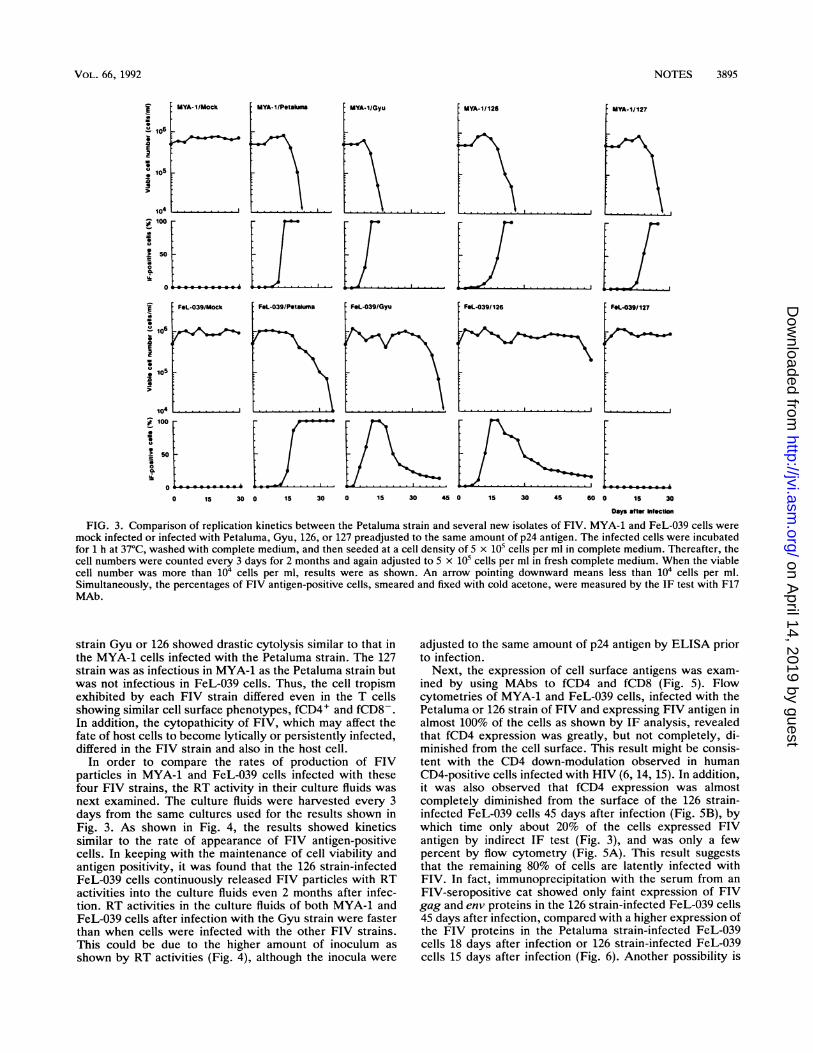
lines were compared for all six new isolates of FIV. FIV-Petaluma grown in Crandell feline kidney cells was alsoused. The FIV in the culture fluids to be used as inocula wasadjusted to the same amounts by p24 antigen by captureenzyme-linked immunosorbent assay (ELISA) with the se-rum from an FIV-seropositive cat and F17 MAb, as de-scribed previously (18). The cells were subsequently in-fected with FIV for 1 h at 37°C and then cultured at 5 x 105cells/ml in complete medium. The percentage of the FIVantigen-positive cells in the cultures was determined every 3days by indirect IF test on acetone-fixed cells with F17 MAb(Fig. 3). The results of IF with the serum from an FIV-positive cat were essentially the same as those with F17MAb (data not shown). The replication kinetics of the newisolates in MYA-1 and FeL-039 cells showed that the FIVsused could be differentiated into four types, i.e., Petaluma intype I showing high cytopathicity; Gyu, 006, and 048 in typeII showing moderate cytopathicity; 126 and 148 in type IIIshowing less cytopathicity; and 127 in type IV showing highcytopathicity but different cell tropism from types I to IIIFIVs. The results for Petaluma, Gyu, 126, and 127 are shownas representatives of the four groups (Fig. 3). The replicationkinetics of the Petaluma strain in MYA-1 cells were slightlyfaster than in FeL-039 cells. In both infected cells, strongcytopathicity was observed, as determined by the increase inFIV antigen-positive cells. After infection, FeL-039 cellsshowed a decreased rate of loss of cell viability comparedwith that in MYA-1 cells, although the percentages of FIVantigen-positive cells were almost 100% in both cases. Bycontrast, FeL-039 cells infected with the Gyu or 126 strain
p24-
Cell Virus
>%N N
Ev ico
-0
0.(C)N N
*iff
*M
FIG. 2. Immunoprecipitation of FIV proteins with F17 MAb toFIV p24gag from cell and virus particle fractions of MYA-1 cellsinfected with newly isolated FIVs. MYA-1 cells uninfected (Mock)or infected with Petaluma, Gyu, 126, or 127 were labeled for 16 h at37°C with 35S-protein labeling mix containing 77% methionine and18% cystein (1,128.5 Ci/mmol; New England Nuclear) in RPMI 1640medium containing 1/10 the normal concentration of methionine andcystine and 5% fetal bovine serum. The cell and virus particlelysates in lysis buffer (0.5% Nonidet P-40, 0.5% sodium deoxycho-late, 0.05 M Tris-HCl [pH 7.2], 0.1 M NaCl, 1 mM EDTA, 1 mMphenylmethylsulfonyl fluoride) were immunoprecipitated with F17MAb as described previously (6). FIV proteins in the precipitateswere analyzed by SDS-PAGE (separation gel, 10 to 15% lineargradient polyacrylamide gel; spacer gel, 4% gel). The molecularweights of the proteins were calculated by comparing their mobili-ties with those of marker proteins by using a calibration kit formolecular weight determination (Pharmacia Fine Chemicals, Upp-sala, Sweden).
continued to proliferate even after the detection of FIVantigen in most of the cells, although slight cell death wastransiently observed in both infected cell lines. The Gyustrain-infected FeL-039 cells exhibited gradual loss of via-bility during a period of about 1 month after reaching almost100% antigen-positive cells, whereas the 126 strain-infectedFeL-039 cells showed continuous proliferation for at least 2months after infection. In both cases, when FeL-039 wasinfected by Gyu or 126, the percentages of FIV antigen-positive cells gradually decreased to about 20% during thepersistent state. In contrast, MYA-1 cells infected with
A.
if
*t
on April 14, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
NOTES 3895
A106
41
106
I
i0100
104
1 5030
MYA-1/Mock
...
MYA.1/Petakom MYA-1/Gyu
.. . . -.
I YA-1/126
_ F&L-039/Mock
0
S 100_
* 50.
0 15 30
FaL-039/Petaluma
0. 3
, ...I
0 15 30
FdL-039/Gyu
0 iS 30 45 0 15 30 45 s0
FIG. 3. Comparison of replication kinetics between the Petaluma strain and several new isolates of FIV. MYA-1 and FeL-039 cells weremock infected or infected with Petaluma, Gyu, 126, or 127 preadjusted to the same amount of p24 antigen. The infected cells were incubatedfor 1 h at 37°C, washed with complete medium, and then seeded at a cell density of 5 x 105 cells per ml in complete medium. Thereafter, thecell numbers were counted every 3 days for 2 months and again adjusted to 5 x 105 cells per ml in fresh complete medium. When the viablecell number was more than 104 cells per ml, results were as shown. An arrow pointing downward means less than 104 cells per ml.Simultaneously, the percentages of FIV antigen-positive cells, smeared and fixed with cold acetone, were measured by the IF test with F17MAb.
strain Gyu or 126 showed drastic cytolysis similar to that inthe MYA-1 cells infected with the Petaluma strain. The 127strain was as infectious in MYA-1 as the Petaluma strain butwas not infectious in FeL-039 cells. Thus, the cell tropismexhibited by each FIV strain differed even in the T cellsshowing similar cell surface phenotypes, fCD4+ and fCD8-.In addition, the cytopathicity of FIV, which may affect thefate of host cells to become lytically or persistently infected,differed in the FIV strain and also in the host cell.
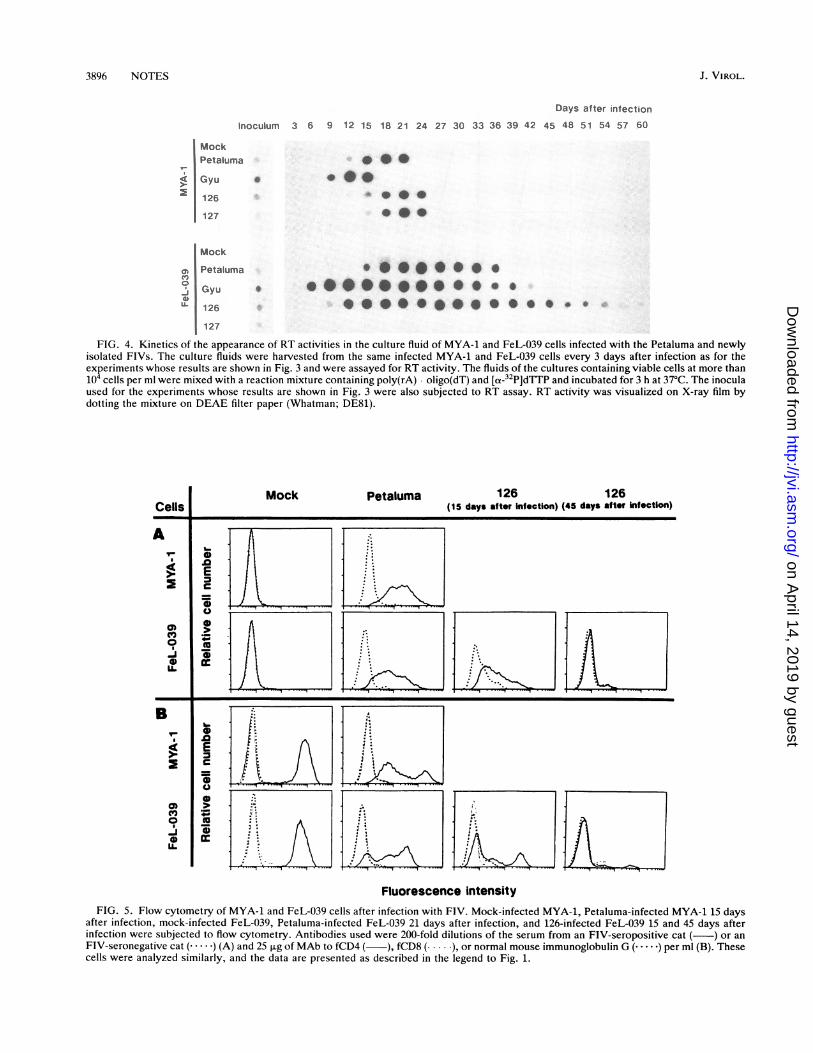
In order to compare the rates of production of FIVparticles in MYA-1 and FeL-039 cells infected with thesefour FIV strains, the RT activity in their culture fluids wasnext examined. The culture fluids were harvested every 3days from the same cultures used for the results shown inFig. 3. As shown in Fig. 4, the results showed kineticssimilar to the rate of appearance of FIV antigen-positivecells. In keeping with the maintenance of cell viability andantigen positivity, it was found that the 126 strain-infectedFeL-039 cells continuously released FIV particles with RTactivities into the culture fluids even 2 months after infec-tion. RT activities in the culture fluids of both MYA-1 andFeL-039 cells after infection with the Gyu strain were fasterthan when cells were infected with the other FIV strains.This could be due to the higher amount of inoculum as
shown by RT activities (Fig. 4), although the inocula were
adjusted to the same amount of p24 antigen by ELISA priorto infection.
Next, the expression of cell surface antigens was exam-
ined by using MAbs to fCD4 and fCD8 (Fig. 5). Flowcytometries of MYA-1 and FeL-039 cells, infected with thePetaluma or 126 strain of FIV and expressing FIV antigen inalmost 100% of the cells as shown by IF analysis, revealedthat fCD4 expression was greatly, but not completely, di-minished from the cell surface. This result might be consis-tent with the CD4 down-modulation observed in humanCD4-positive cells infected with HIV (6, 14, 15). In addition,it was also observed that fCD4 expression was almostcompletely diminished from the surface of the 126 strain-infected FeL-039 cells 45 days after infection (Fig. SB), bywhich time only about 20% of the cells expressed FIVantigen by indirect IF test (Fig. 3), and was only a fewpercent by flow cytometry (Fig. 5A). This result suggeststhat the remaining 80% of cells are latently infected withFIV. In fact, immunoprecipitation with the serum from an
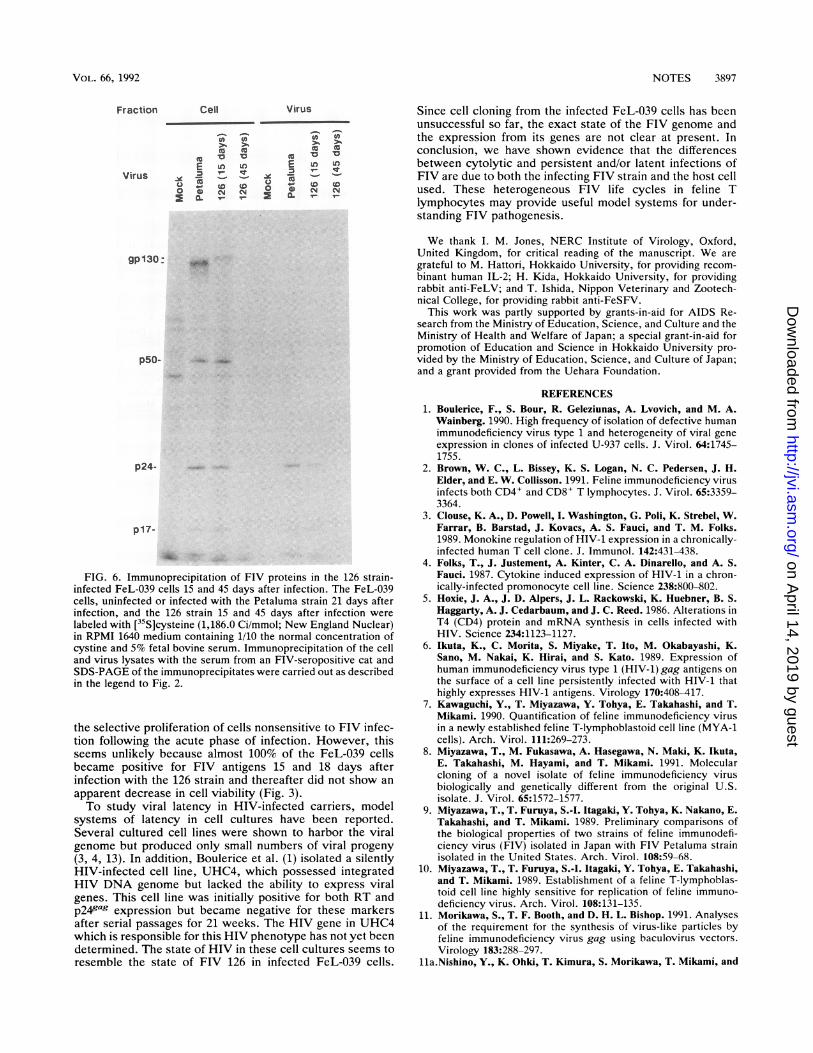
FIV-seropositive cat showed only faint expression of FIVgag and env proteins in the 126 strain-infected FeL-039 cells45 days after infection, compared with a higher expression ofthe FIV proteins in the Petaluma strain-infected FeL-039cells 18 days after infection or 126 strain-infected FeL-039cells 15 days after infection (Fig. 6). Another possibility is
PFaL-039/126
,
FOL-039/127
L. .-
0 15 30
Day attr hInection
I
VOL. 66, 1992
on April 14, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
3896 NOTES
Days after infection
Inoculum 3 6 9 12 15 18 21 24 27 30 33 36 39 42 45 48 51 54 57 60
MockPetaluma
4 Gyu2 126
127
0 *S
0@:*
Mock
O Petalumao
Gyu *
O 126 *
127
FIG. 4. Kinetics of the appearance of RT activities in the culture fluid of MYA-1 and FeL-039 cells infected with the Petaluma and newlyisolated FIVs. The culture fluids were harvested from the same infected MYA-1 and FeL-039 cells every 3 days after infection as for theexperiments whose results are shown in Fig. 3 and were assayed for RT activity. The fluids of the cultures containing viable cells at more than104 cells per ml were mixed with a reaction mixture containing poly(rA) oligo(dT) and [a-32P]dTTP and incubated for 3 h at 37°C. The inoculaused for the experiments whose results are shown in Fig. 3 were also subjected to RT assay. RT activity was visualized on X-ray film bydotting the mixture on DEAE filter paper (Whatman; DE81).
Cells
I-
,v
o IL-
Br
q
0)
0-JIL
N.
.0EC
coZiS.XUS
Mock Petaluma 126 126(15 days after Infection) (45 days afte Infection)
Fluorescence intensityFIG. 5. Flow cytometry of MYA-1 and FeL-039 cells after infection with FIV. Mock-infected MYA-1, Petaluma-infected MYA-1 15 days
after infection, mock-infected FeL-039, Petaluma-infected FeL-039 21 days after infection, and 126-infected FeL-039 15 and 45 days afterinfection were subjected to flow cytometry. Antibodies used were 200-fold dilutions of the serum from an FIV-seropositive cat ( ) or anFIV-seronegative cat (. ) (A) and 25 ,ug of MAb to fCD4 ( ), fCD8 (. ), or normal mouse immunoglobulin G (. ) per ml (B). Thesecells were analyzed similarly, and the data are presented as described in the legend to Fig. 1.
J . VlIROL.
on April 14, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
NOTES 3897
Virus
0t
>1 >-
to
E Luo L
0
0.o 0 CMN2 a. r- I_
gp130: -W
p50-
p24-
p17-
FIG. 6. Immunoprecipitation of FIV proteins in the 126 strain-infected FeL-039 cells 15 and 45 days after infection. The FeL-039cells, uninfected or infected with the Petaluma strain 21 days afterinfection, and the 126 strain 15 and 45 days after infection were
labeled with [35S]cysteine (1,186.0 Ci/mmol; New England Nuclear)in RPMI 1640 medium containing 1/10 the normal concentration ofcystine and 5% fetal bovine serum. Immunoprecipitation of the celland virus lysates with the serum from an FIV-seropositive cat andSDS-PAGE of the immunoprecipitates were carried out as describedin the legend to Fig. 2.
the selective proliferation of cells nonsensitive to FIV infec-tion following the acute phase of infection. However, thisseems unlikely because almost 100% of the FeL-039 cellsbecame positive for FIV antigens 15 and 18 days afterinfection with the 126 strain and thereafter did not show an
apparent decrease in cell viability (Fig. 3).To study viral latency in HIV-infected carriers, model
systems of latency in cell cultures have been reported.Several cultured cell lines were shown to harbor the viralgenome but produced only small numbers of viral progeny
(3, 4, 13). In addition, Boulerice et al. (1) isolated a silentlyHIV-infected cell line, UHC4, which possessed integratedHIV DNA genome but lacked the ability to express viralgenes. This cell line was initially positive for both RT andp249ag expression but became negative for these markersafter serial passages for 21 weeks. The HIV gene in UHC4which is responsible for this HIV phenotype has not yet beendetermined. The state of HIV in these cell cultures seems toresemble the state of FIV 126 in infected FeL-039 cells.
Since cell cloning from the infected FeL-039 cells has beenunsuccessful so far, the exact state of the FIV genome andthe expression from its genes are not clear at present. Inconclusion, we have shown evidence that the differencesbetween cytolytic and persistent and/or latent infections ofFIV are due to both the infecting FIV strain and the host cellused. These heterogeneous FIV life cycles in feline Tlymphocytes may provide useful model systems for under-standing FIV pathogenesis.
We thank I. M. Jones, NERC Institute of Virology, Oxford,United Kingdom, for critical reading of the manuscript. We are
grateful to M. Hattori, Hokkaido University, for providing recom-
binant human IL-2; H. Kida, Hokkaido University, for providingrabbit anti-FeLV; and T. Ishida, Nippon Veterinary and Zootech-nical College, for providing rabbit anti-FeSFV.
This work was partly supported by grants-in-aid for AIDS Re-search from the Ministry of Education, Science, and Culture and theMinistry of Health and Welfare of Japan; a special grant-in-aid forpromotion of Education and Science in Hokkaido University pro-
vided by the Ministry of Education, Science, and Culture of Japan;and a grant provided from the Uehara Foundation.
REFERENCES
1. Boulerice, F., S. Bour, R. Geleziunas, A. Lvovich, and M. A.Wainberg. 1990. High frequency of isolation of defective humanimmunodeficiency virus type 1 and heterogeneity of viral gene
expression in clones of infected U-937 cells. J. Virol. 64:1745-1755.
2. Brown, W. C., L. Bissey, K. S. Logan, N. C. Pedersen, J. H.Elder, and E. W. Collisson. 1991. Feline immunodeficiency virusinfects both CD4' and CD8' T lymphocytes. J. Virol. 65:3359-3364.
3. Clouse, K. A., D. Powell, I. Washington, G. Poli, K. Strebel, W.Farrar, B. Barstad, J. Kovacs, A. S. Fauci, and T. M. Folks.1989. Monokine regulation of HIV-1 expression in a chronically-infected human T cell clone. J. Immunol. 142:431-438.
4. Folks, T., J. Justement, A. Kinter, C. A. Dinarello, and A. S.Fauci. 1987. Cytokine induced expression of HIV-1 in a chron-ically-infected promonocyte cell line. Science 238:800-802.
5. Hoxie, J. A., J. D. Alpers, J. L. Rackowski, K. Huebner, B. S.Haggarty, A. J. Cedarbaum, and J. C. Reed. 1986. Alterations inT4 (CD4) protein and mRNA synthesis in cells infected withHIV. Science 234:1123-1127.
6. Ikuta, K., C. Morita, S. Miyake, T. Ito, M. Okabayashi, K.Sano, M. Nakai, K. Hirai, and S. Kato. 1989. Expression ofhuman immunodeficiency virus type 1 (HIV-1) gag antigens on
the surface of a cell line persistently infected with HIV-1 thathighly expresses HIV-1 antigens. Virology 170:408-417.
7. Kawaguchi, Y., T. Miyazawa, Y. Tohya, E. Takahashi, and T.Mikami. 1990. Quantification of feline immunodeficiency virusin a newly established feline T-lymphoblastoid cell line (MYA-1cells). Arch. Virol. 111:269-273.
8. Miyazawa, T., M. Fukasawa, A. Hasegawa, N. Maki, K. Ikuta,E. Takahashi, M. Hayami, and T. Mikami. 1991. Molecularcloning of a novel isolate of feline immunodeficiency virusbiologically and genetically different from the original U.S.isolate. J. Virol. 65:1572-1577.
9. Miyazawa, T., T. Furuya, S.-I. Itagaki, Y. Tohya, K. Nakano, E.Takahashi, and T. Mikami. 1989. Preliminary comparisons ofthe biological properties of two strains of feline immunodefi-ciency virus (FIV) isolated in Japan with FIV Petaluma strainisolated in the United States. Arch. Virol. 108:59-68.
10. Miyazawa, T., T. Furuya, S.-I. Itagaki, Y. Tohya, E. Takahashi,and T. Mikami. 1989. Establishment of a feline T-lymphoblas-toid cell line highly sensitive for replication of feline immuno-deficiency virus. Arch. Virol. 108:131-135.
11. Morikawa, S., T. F. Booth, and D. H. L. Bishop. 1991. Analysesof the requirement for the synthesis of virus-like particles byfeline immunodeficiency virus gag using baculovirus vectors.Virology 183:288-297.
11a.Nishino, Y., K. Ohki, T. Kimura, S. Morikawa, T. Mikami, and
Fraction Cell
0 0
V00E Ln Ln
Virus V : !.( D tD0 0 N N
a.X-_ _
VOL. 66, 1992
on April 14, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

3898 NOTES
K. Ikuta. Major core proteins, p24s, of human, simian, andfeline immunodeficiency viruses are partly expressed on thesurface of the virus-infected cells. Vaccine, in press.
12. Pedersen, N. C., E. W. Ho, M. L. Brown, and J. K. Yamamoto.1987. Isolation of a T-lymphotropic virus from domestic catswith an immunodeficiency-like syndrome. Science 235:790-793.
13. Pomerantz, R. J., D. Trono, M. B. Feinberg, and D. Baltimore.1990. Cells nonproductively infected with HIV-1 exhibit anaberrant pattern of viral RNA expression: a molecular model forlatency. Cell 61:1271-1276.
14. Salmon, P., R. Olivier, Y. Riviere, E. Brisson, J.-C. Gluckman,M.-P. Kieny, L. Montagnier, and D. Klatzman. 1988. Loss ofCD4 membrane expression and CD4 mRNA during acute hu-man immunodeficiency virus replication. J. Exp. Med. 168:1953-1969.
15. Stevenson, M., X. Zhang, and D. J. Volsky. 1987. Downregula-tion of cell surface molecule during noncytopathic infection of Tcells with human immunodeficiency virus. J. Virol. 61:3741-3748.
16. Talbott, R. L., E. E. Sparger, K. M. Lovelace, W. M. Fitch,N. C. Pedersen, P. A. Luciw, and J. H. Elder. 1989. Nucleotidesequence and genomic organization of the feline immunodefi-ciency virus. Proc. Natl. Acad. Sci. USA 86:5743-5747.
17. Theilen, G. H., T. G. Kawakami, J. D. Rush, and R. J. Munn.1969. Replication of cat leukaemia virus in cell suspensioncultures. Nature (London) 222:589-590.
18. Tilton, G. K., T. P. O'Connor, Jr., C. L. Seymour, K. L.Lawrence, N. D. Cohen, P. R. Andersen, and Q. J. Tonelli. 1990.Immunoassay for detection of feline immunodeficiency virus
core antigen. J. Clin. Microbiol. 28:898-904.19. Tochikura, T. S., K. A. Hayes, C. M. Cheney, A. Tanabe-
Tochikura, J. L. Tojko, L. E. Mathes, and R. G. Olsen. 1990. Invitro replication and cytopathogenicity of the feline immunode-ficiency virus for feline T4 thymic lymphoma 3201 cells. Virol-ogy 179:492-497.
20. Willey, R. L., D. H. Smith, L. A. Laskey, T. S. Theodore, P. L.Earl, B. Moss, D. J. Capon, and M. A. Martin. 1988. In vitromutagenesis identifies a region within the envelope gene of thehuman immunodeficiency virus that is critical for infectivity. J.Virol. 62:139-147.
21. Yamamoto, J. K., C. D. Ackley, H. Zochlinski, H. Louie, E.Pembroke, M. Torten, H. Hansen, R. Munn, and T. Okuda.1991. Development of IL-2-independent feline lymphoid celllines chronically infected with feline immunodeficiency virus:importance for diagnostic reagents and vaccines. Intervirology32:361-375.
22. Yamamoto, J. K., H. Hansen, E. W. Ho, T. Y. Morishita, T.Okuda, T. R. Sawa, R. M. Nakamura, and N. C. Pedersen. 1989.Epidemiologic and clinical aspects of feline immunodeficiencyvirus infection in cats from the continental United States andCanada and possible mode of transmission. J. Am. Vet. Med.Assoc. 194:213-220.
23. Yamamoto, J. K., F. Sparger, E. W. Ho, P. R. Anderson, T. P.O'Connor, C. P. Mandell, L. Lowenstine, R. Mune, and N. C.Pedersen. 1988. Pathogenesis of experimentally induced felineimmunodeficiency virus infection in cats. Am. J. Vet. Res.48:1246-1258.
J. VIROL.
on April 14, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from