alveolar macrophages in neonatal mice are inherently
TRANSCRIPT

Alveolar Macrophages in Neonatal Mice Are Inherently Unresponsiveto Pneumocystis murina Infection
Cathryn Kurkjian,a Melissa Hollifield,a,d J. Louise Lines,a Amy Rogosky,a Kerry M. Empey,b* Mahboob Qureshi,a,c* Stephen A. Brown,d
and Beth A. Garvya,c,d
Departments of Microbiology, Immunology, and Molecular Genetics,a Clinical Pharmaceutical Sciences,b and Internal Medicine,c University of Kentucky, Lexington,Kentucky, USA, and Veteran’s Administration Medical Center, Lexington, Kentucky, USAd
Pneumocystis pneumonia was first diagnosed in malnourished children and has more recently been found in children with upperrespiratory symptoms. We previously reported that there is a significant delay in the immune response in newborn mice infectedwith Pneumocystis compared to adults (Garvy BA, Harmsen AG, Infect. Immun. 64:3987–3992, 1996, and Garvy BA, Qureshi M,J. Immunol. 165:6480 – 6486, 2000). This delay is characterized by the failure of neonatal lungs to upregulate proinflammatorycytokines and attract T cells into the alveoli. Here, we report that regardless of the age at which we infected the mice, they failedto mount an inflammatory response in the alveolar spaces until they were 21 days of age or older. Anti-inflammatory cytokineshad some role in dampening inflammation, since interleukin-10 (IL-10)-deficient pups cleared Pneumocystis faster than wild-type pups and the neutralization of transforming growth factor beta (TGF-�) with specific antibody enhanced T cell migrationinto the lungs at later time points. However, the clearance kinetics were similar to those of control pups, suggesting that there isan intrinsic deficiency in the ability of innate immunity to control Pneumocystis. We found, using an adoptive transfer strategy,that the lung environment contributes to association of Pneumocystis organisms with alveolar macrophages, implying no intrin-sic deficiency in the binding of Pneumocystis by neonatal macrophages. Using both in vivo and in vitro assays, we found thatPneumocystis organisms were less able to stimulate translocation of NF-�B to the nucleus of alveolar macrophages from neona-tal mice. These data indicate that there is an early unresponsiveness of neonatal alveolar macrophages to Pneumocystis infectionthat is both intrinsic and related to the immunosuppressive environment found in neonatal lungs.
Pneumocystis species are opportunistic fungal pathogens thatcause pneumonia in immunosuppressed mammalian hosts.
Outbreaks of Pneumocystis pneumonia (PCP) were first describedin orphan homes among malnourished children (18). Most chil-dren encounter Pneumocystis infection by the age of 2 years, asevidenced by the presence of specific antibodies in the peripheralblood (33), and recent studies suggest that up to 30% of youngchildren carry Pneumocystis in the lungs (50, 51). Clearance ofPneumocystis is dependent on a functional CD4� T cell compart-ment as well as on B cells and alveolar macrophages (23, 28–30,42). Infants with AIDS tend to have a more fulminate course ofPCP than do adults with AIDS, possibly due to the immaturity ofthe immune system (8, 39).
Susceptibility of newborn infants to infection is thought to bedue to the inexperience of the adaptive immune system along withfunctional deficits of both innate and adaptive immune responses(1, 26). We have previously published that it takes 3 weeks for miceinfected at 2 days of age to mount a CD4� T cell response into thealveolar spaces in response to mouse-specific Pneumocystismurina infection (20, 21, 37). In contrast, CD4� T cells infiltratedthe alveoli within 4 to 5 days postinfection in adult mice chal-lenged with a comparable dose. This delay in lung T cell infiltra-tion is associated with delayed infiltration and activation of mac-rophages, delayed chemokine and cytokine production, anddelayed adhesion molecule upregulation (12, 13, 35–37). We alsofound that lung mRNA expression of transforming growth factor�2 (TGF-�2) and TGF-�3 isoforms was upregulated in the unin-fected lungs of infant mice, and TGF-�1 mRNA expression wassimilar in adult and infant mice (37).
Since TGF-� is known to have anti-inflammatory activity, wehypothesized that TGF-� is constitutively expressed in postnatally
developing lungs as a protective mechanism against overly exu-berant inflammatory responses to infectious agents or foreignparticles. Rapid alveolarization takes place during the first 2 weeksafter birth in mice and the first 6 months in human infants (7, 25,48, 54). Inflammation in the lungs during this time has the poten-tial to cause significant damage that could be irreversible. To de-termine whether the immune response to P. murina is develop-mentally regulated, we infected mice at various times after birth todetermine when the immune response was initiated. We also ex-amined TGF-� protein levels in the lungs of infant and adult mice.Our data confirm that TGF-� protein levels are developmentallycontrolled in the lungs and that TGF-� contributes to a delayedimmune response to P. murina in mice infected prior to 3 weeks ofage. However, other intrinsic host factors contribute to the de-layed response to P. murina in neonatal mice, since neutralizationof TGF-� in interleukin-10 (IL-10)-deficient mice did not signif-
Received 28 July 2011 Returned for modification 20 August 2011Accepted 23 May 2012
Published ahead of print 4 June 2012
Editor: G. S. Deepe, Jr.
Address correspondence to Beth A. Garvy, [email protected].
* Present address: Kerry M. Empey, Department of Pharmacy and Therapeutics,University of Pittsburgh, School of Pharmacy, Pittsburgh, Pennsylvania, USA;Mahboob Qureshi, Department of Basic Sciences, Touro University-Nevada,Henderson, Nevada, USA.
Supplemental material for this article may be found at http://iai.asm.org/.
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/IAI.05707-11
August 2012 Volume 80 Number 8 Infection and Immunity p. 2835–2846 iai.asm.org 2835
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 15
Nov
embe
r 20
21 b
y 12
1.12
9.11
0.11
8.

icantly affect clearance of the organisms, even though the inflam-matory response was more intense. Adoptive transfer experimentssuggested that the neonatal lung environment affects the interac-tion of alveolar macrophages with P. murina. However, there isalso an intrinsic inability of neonatal alveolar macrophages to re-spond to P. murina, as confirmed by the failure to activate NF-�Bboth in vitro and in vivo.
MATERIALS AND METHODSMice. Eight-week-old BALB/c, B6D2F1/J, C57BL/6, B6.129P2-IL-10tm1Cgn/J (IL-10�/�), or C57BL/6-Tg(UBC-GFP)30Scha/J (GFP) micewere purchased from Charles River (Wilmington, MA), Taconic (Hud-son, NY), or Jackson Laboratories (Bar Harbor, ME) and bred in ouranimal facilities. Mice were maintained at the Veterinary Medical Unit ofthe Veterans Administration Medical Center (VAMC) or University ofKentucky Department of Laboratory Animal Resources (DLAR) underspecific-pathogen-free conditions. C.B-Igh-1b/ICrTac-Prkdcscid (desig-nated SCID) or C1.29S6(B6)-Rag2tm1FwaN12 (designated RagKO) mice,originally from Taconic (Germantown, NY), were used to maintain asource of P. murina and were bred at the VAMC or DLAR in microisolatorcages with sterilized food and water. The VAMC Institutional AnimalCare and Use Committee (IACUC) and University of Kentucky IACUCapproved all protocols regarding animal use.
P. murina isolation, infection, adoptive transfers, and TGF-� neu-tralization. Lungs were excised from P. murina-infected SCID or RagKOmice and pushed through stainless steel mesh in Hank’s balanced saltsolution (HBSS). Cell debris was removed by centrifugation at 100 � g for3 min. The organisms were pelleted after lysis of red blood cells with water,resuspended in HBSS, and enumerated for inoculation for clearance stud-ies. For purification of trophozoites for in vitro studies, red blood cellswere lysed with water and organisms suspended in HBSS containing 0.5%glutathione at pH 7.3. Organisms were incubated with 200 U DNase at37°C, and clumps were broken up by aspirating through a 26-gauge nee-dle. Cell debris was removed by low-speed centrifugation. The superna-tant was spun at 400 � g to pellet cysts. The supernatant from the 400 � gspin was centrifuged at 1,300 � g to pellet trophozoites. Trophozoiteswere resuspended in HBSS and filtered through 20-�m Magna nylonfilters (GE Osmonics). This preparation results in more than 99% puretrophozoites. The pellet from the 400 � g spin was resuspended in HBSSand contained a mixed population of cysts and trophozoites, and thispopulation was used as an enriched cyst population for in vitro studies.Aliquots of lung homogenates or purified organisms were spun onto glassslides, fixed with methanol, and stained with DiffQuik (Siemens Health-care Diagnostics, Inc., Deerfield, IL). P. murina organisms were enumer-ated by microscopy as described previously (20, 22). Freshly isolated or-ganisms were used for clearance experiments and routinely consisted of aratio of 1:10 cysts to trophic forms.
Mice were anesthetized lightly with halothane or isoflurane anesthesia,and the P. murina inoculum (5 � 105 organisms/g of body weight in 10 �lfor neonates and 50 �l for adults) was placed over both nares (intranasally[i.n.]). For some experiments, organisms purified as described above tothe point of filtration (mixed cysts and trophozoites) were stained withCellTrace Far Red DDAO-SE (DDAO), which has an active succinimidylester that binds amine groups, by incubating organisms with the dye ac-cording to the manufacturer’s instructions (Molecular Probes, Invitro-gen, Carlsbad, CA) prior to infecting mice. The labeling of organisms wasconfirmed by flow cytometry and fluorescence microscopy. Control ani-mals were given inoculations of DDAO-labeled preparations of unin-fected RagKO lungs to confirm that the dye did not bind to any leftoverlung debris (see Fig. S1 in the supplemental material). For the adoptivetransfer of alveolar macrophages, anesthetized adult or 8-day-old micereceived i.n. inoculations of 1.5 � 105 or 5 � 104 green fluorescent pro-tein-positive (GFP�) cells, respectively. For neutralization of TGF-�, neo-natal mice were given intraperitoneal injections of 10 or 50 to 200 �g/ganti-TGF-�1, TGF-�2, and TGF-�3 monoclonal antibody (TGF-�1,2,3
MAb) once or twice per week (11) (clone 1D11; R&D Systems, Minneap-olis, MN; or BioXcell, West Lebanon, NH). A second set of mice wasinjected with an irrelevant mouse IgG1 as an isotype-matched control.
Isolation of cells from alveolar spaces, lungs, and lymph nodes. Micewere exsanguinated under deep halothane or isoflurane anesthesia, andlungs were lavaged with 5 washes of HBSS containing 3 mM EDTA. Bron-chial alveolar lavage fluid (BALF) from the first wash was saved for quan-tification of cytokines. For isolation of alveolar macrophages, cells werepelleted from BALF and resuspended in appropriate media for flow cy-tometry, culture, or adoptive transfer. Right lung lobes were excised,minced, and digested in RPMI containing 3% heat-inactivated fetal calfserum, 1 mg/ml collagenase A, and 50 U/ml DNase for 1 h at 37°C. Di-gested lungs were pushed through 70-�m nylon mesh screens to obtainsingle-cell suspensions, and aliquots were taken for enumeration of P.murina. Tracheobronchial lymph nodes (TBLN) were also excised andpushed through 70-�m nylon mesh screens in HBSS. Erythrocytes wereremoved using a hypotonic lysing buffer. Cells were washed and counted.
Enumeration of Pneumocystis in the lungs of mice. Aliquots of lunghomogenates were diluted, and 100-�l aliquots were spun onto glassslides. Slides were fixed in methanol and stained with DiffQuik (Siemens).P. murina nuclei were enumerated microscopically as previously de-scribed (20). Lung burden is expressed as log10 P. murina nuclei per rightlung lobe, and the limit of detection was 3.23 log10 nuclei per lung.
Flow-cytometric analysis. Lung lavage, lung digest, and TBLN cellswere washed with phosphate-buffered saline (PBS) containing 0.1% bo-vine serum albumin and 0.02% NaN3 and stained with appropriate con-centrations of fluorochrome-conjugated antibodies specific for murine Tcells (CD4, CD8, CD44, and CD62L) or macrophages (CD11c, CD11b,F4/80, and Ia). Antibodies were purchased from BD Biosciences-Phar-mingen (San Diego, CA) or eBioscience (San Diego, CA). Expression ofthese molecules on the surface of the cells was determined by multiparam-eter flow cytometry using a FACSCalibur or LSRII cytofluorimeter (Bec-ton, Dickinson, Mountain View, CA). Ten thousand to 50,000 events wereroutinely acquired.
Analysis of cytokine levels and P. murina-specific IgG in BALF. Cellsand debris were removed from the first wash of BALF by centrifugation,and the supernatant was frozen at �80°C for later use. The quantitation ofmultiple cytokines in the same sample of BALF was performed using beadarray kits purchased from Upstate Cell Signaling Solutions (Lake Placid,NY) or BD Biosciences-Pharmingen. Assays were performed according tothe manufacturer’s directions and analyzed using a Luminex 100 system(Luminex Corp., Austin, TX) or flow cytometry on a FACSCalibur. P.murina-specific IgG was measured by enzyme-linked immunosorbent as-say (ELISA). A sonicate of P. murina organisms (10 �g protein/ml) wascoated onto 96-well plates, and wells were blocked with 5% dry milk inHBSS containing 0.05% Tween 20. Sera were diluted and incubated onplates overnight. Plates were extensively washed, and bound IgG was de-tected using alkaline phosphatase-conjugated anti-mouse IgG (Sigma).Plates were washed and secondary antibodies detected using p-nitrophe-nylphosphate at 1 mg/ml in diethanolamine buffer. Optical density wasread at 405 nm using a plate reader equipped with KC Junior software(Bio-Tek Instruments, Inc., Winnoski, VT).
Analysis of TGF-�1 levels in lung homogenates with ELISA. Mouseleft lung lobes were snap-frozen in liquid nitrogen at the time of euthana-sia and stored at �80°C for later analysis. Five hundred �l of dilutedprotease inhibitor cocktail (1:100 in PBS; Sigma-Aldrich, St. Louis, MO)was added, and the lungs were homogenized using a Dounce homoge-nizer. The homogenates then were centrifuged at 400 � g for 10 min at4°C. Supernatants were collected and used in a TGF-� ELISA kit (R&DSystems, Minneapolis, MN, or eBioscience). The ELISA was performedaccording to the manufacturer’s instructions. Data were normalized tolung protein concentrations, which were determined using a Bio-Rad DCprotein assay kit (Hercules, CA) according to the manufacturer’s instruc-tions.
Kurkjian et al.
2836 iai.asm.org Infection and Immunity
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 15
Nov
embe
r 20
21 b
y 12
1.12
9.11
0.11
8.

Analysis of NF-�B translocation ex vivo and in vitro. Mice wereinfected with P. murina at 2 days of age or as adults as previously de-scribed. At multiple time points, lungs were removed and snap-frozen forsubsequent analysis. Whole lung tissue was homogenized and nuclei ex-tracted as previously described (53, 57, 58), and protein concentrations ofnuclear extracts were determined using the DC protein assay kit fromBio-Rad Laboratories. Nuclear extracts (5 �g of protein) were incubatedwith electrophoretic mobility shift assay (EMSA) binding buffer and 10fmol of 32P-labeled NF-�B probe (double-stranded oligonucleotides con-taining consensus �B sequence 5=-TCAGAGGGGACTTTCCGAGAGG-3=, underlining denotes the NF-�B finding site) for 20 min at room tem-perature. Reaction mixtures were separated in a 6% nondenaturingpolyacrylamide gel by electrophoresis. The gel was transferred to blotterpaper, dried, and exposed to X-ray film using intensifying screens at�70°C. Relative nuclear binding activities for NF-�B were quantified byscanning densitometry.
For examining NF-�B activation in alveolar macrophages, mice wereinfected with P. murina at 2 days of age or as adults as previously de-scribed. At multiple time points, lungs were lavaged and alveolar macro-phages pooled from up to 15 mice. Nuclear extracts were isolated using akit from Panomics (Fremont, CA) per the manufacturer’s instructions.Nuclear extract protein levels were quantified using the microassay procedureof the DC protein assay kit from Bio-Rad Laboratories. Equal protein con-centrations were used in a Chemicon NF-�Bp65 nuclear translocation color-imetric plate-based assay (Millipore, Bellerica, MA) according to the manu-facturer’s instructions. Data are expressed as the optical density read at 450nm in up to triplicate wells per experimental group.
For in vitro assays, alveolar macrophages pooled from adult and 10- to14-day-old mice were placed into RPMI medium containing 5% fetalbovine serum, antibiotics, and 2-mercaptoethanol.
Cells were rested overnight at 2 � 106 cells/ml in 6-well plates or at 2 �105 on chamber slides and then stimulated with 100 ng/ml lipopolysac-charide (LPS) (from Escherichia coli; Sigma), a 1:50 dilution of sonicatedP. murina organisms (at 108 nuclei/ml), 25 mg/ml zymosan, trophozoitesat 10 per 1 macrophage, or a mix of cysts and trophozoites (1:10) at 10cysts per 1 macrophage for up to 4 h. Medium was removed and frozen.Adherent cells in 6-well plates were lysed and nuclei extracted using aChemicon nuclear extraction kit per the manufacturer’s instructions(Millipore). NF-�B translocation was determined using the plate-basedChemicon kit as described above. Adherent cells cultured on chamberslides were fixed in 10% formalin for 15 min, washed in PBS, and perme-abilized in methanol for 10 min. After blocking with 5% normal goatserum in PBS plus 0.3% Triton X-100, cells were incubated overnight withanti-NF-�B p65 (Cell Signaling Technology, Beverly, MA) followed byanti-rabbit IgG-Alexa 594 for 2 h. Nuclei were stained with 4=,6-di-amidino-2-phenylindole (DAPI) prior to examination using a Zeiss im-ager Z1 fluorescence microscope equipped with an AxioCam HRc andAxioVision Rel.4.8 software or a Leica TSP SPS inverted confocal micro-scope. The proportion of cells with translocated NF-�B was determinedmicroscopically by counting a minimum of 100 cells and determining thenumber with colocalization of NF-�B and nuclear staining.
Statistical analysis. Data were analyzed utilizing the SigmaStat statis-tical software package (SPSS Inc., Chicago, IL). Analysis of variance(ANOVA) was used to determine differences between and within groups.Student Newman Keul’s post hoc tests were applied to discriminate differ-ences between groups at individual time points. Data were determined tobe significantly different when the P value was less than 0.05 using atwo-tailed test. For data that did not meet the assumptions of equal vari-ance or normal distribution, nonparametric tests were used.
RESULTSResolution of Pneumocystis infection is age dependent. We havepreviously shown that clearance of P. murina is delayed in miceinfected as neonates compared to clearance in adult mice (20). Todetermine whether P. murina organism clearance and inflamma-
tion is developmentally controlled, mice were infected intrana-sally at 2, 7, or 14 days of age, and their ability to resolve P. murinainfection was compared to that of adult mice. As we have previ-ously reported, organism burden had peaked by day 7 postinfec-tion in adult mice, and they cleared P. murina by day 21 postin-fection (Fig. 1A). In contrast, lung burdens of mice infected as 2-,7-, or 14-day-old pups peaked 1 to 2 weeks later than in adultmice. Furthermore, none of the mice infected as pups were able tocontrol their lung burden until after they reached 3 weeks of age.Mice infected at 2 weeks of age were able to control P. murinaburden better than mice infected at 2 and 7 days of age, althoughthey were still unable to clear the organisms until 4 weeks postin-fection, or about 6 weeks of age (Fig. 1A). Notably, mice wereinfected with equivalent numbers of organisms based on bodyweight, which means that adults received approximately 10-foldmore organisms than 2-day-old mice. Variability in the size of themice between groups due to the age at infection and the inoculumdose, which was normalized to weight, likely contribute to thedifferences in lung burden between the groups at day 7 postinfec-tion. Alternatively, there may be a developmental difference in thelungs during the first 3 postnatal weeks that is more permissive forP. murina growth at 7 days than at 14 days of age. Regardless of theinitial inoculum, organism burden increased in the lungs of allpup groups through 2 weeks postinfection, indicating permissive-ness of Pneumocystis growth in the lungs of young mice. Thesedata confirm that between 14 and 21 days of age is a critical time inpup immune development in the context of P. murina clearance.
The immune response to Pneumocystis is age dependent. Al-veolar macrophages have been shown to be the effector cells thatkill P. murina, and specific antibody can have a significant role inopsonizing and targeting organisms for clearance (13, 28, 43, 47).P. murina-specific IgG is elevated in the blood of adult mice at day14 postinfection; however, mice infected at 2, 7, or 14 days of agedid not have measurable specific IgG in the blood until at least 3weeks postinfection (Fig. 1B). Mice infected at 2 days of age didnot have measurable P. murina-specific IgG until after day 21postinfection, the time at which we terminated the experiment forthese mice. We have previously published that specific IgG is notdetected until at least 4 weeks postinfection when mice are in-fected at 2 days of age (20). In addition to the delay in antibodyresponses, activation of alveolar macrophages was delayed in themice infected with P. murina at 2, 7, and 14 days of age comparedto adults (Fig. 1C). Activated macrophages as defined by the up-regulation of major histocompatibility complex class II (MHC-II)(Ia) and CD11b had already peaked at day 7 postinfection in adultmice, whereas the peak for the mice infected as pups lagged behindby as much as 3 weeks. This corresponded to significantly lowerlevels of tumor necrosis factor (TNF) in the BALF of mice infectedas pups compared to mice infected as adults (Fig. 1E). TNF hasbeen shown to induce the expression of adhesion molecules and tobe required early in the host response against P. murina for reso-lution to occur (9, 36). Macrophages have been shown to produceTNF in response to interactions with Pneumocystis �-glucans (24),so the delay in production of TNF in the lungs of mice infected at2 weeks of age and younger is consistent with delayed activation ofalveolar macrophages.
It is well known that CD4� T cells are required for mountingan effective host response to P. murina infection (5, 22, 43), andwe have reported that there is a delay in the infiltration of lympho-cytes into the alveolar spaces of the lungs in mice infected as neo-
Macrophage Function in Neonatal Lungs
August 2012 Volume 80 Number 8 iai.asm.org 2837
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 15
Nov
embe
r 20
21 b
y 12
1.12
9.11
0.11
8.

nates compared to adults (20). Therefore, we determined howincreasing age affects infiltration of T cells into the alveolar spacesof the lung during P. murina infection. As shown in Fig. 1D, mi-gration of activated CD4� T cells into the alveolar spaces wasdelayed in pups infected at 2, 7, or 14 days of age compared toadults. The number of CD4� T cells peaked in adult lungs by day7 postinfection. In contrast, alveolar CD4� T cells peaked at day14 postinfection in pups infected at 2 weeks of age but not untilafter day 14 in pups infected at 2 days and 1 week of age. Notably,infiltration of CD4� T cells into the alveoli was not detected in anygroup of mice until they were 3 weeks of age or older (Fig. 1D).
IL-10 is partially responsible for delaying clearance of Pneu-mocystis from neonatal lungs. We have previously shown thatthere is constitutive mRNA expression of IL-10 and TGF-� iso-
forms in the lungs of neonatal mice, and that these levels werecomparable to or higher than those in adults (37). Furthermore,we reported that adult IL-10�/� mice clear P. murina faster anddevelop a more intense inflammatory response than do wild-typemice (38). Moreover, IL-10 produced by epithelial cells has beenassociated with controlling inflammation in adult lungs (16). Todetermine whether the presence of IL-10 is responsible for thedelayed inflammatory response to P. murina in neonatal mice,IL-10�/� pups were infected with organisms at 2 days of age. LungP. murina burden was cleared with slightly faster kinetics in IL-10�/� pups than in wild-type C57BL/6 pups (Fig. 2A). There weresignificant differences in lung organism burdens between IL-10�/� and wild-type pups by days 24 and 31 postinfection (Fig.2A). The faster clearance kinetics in IL-10�/� mice corresponded
FIG 1 Immune response to and clearance of P. murina is delayed in young mice infected at up to 2 weeks of age compared to adults. Mice were infected at 2, 7,or 14 days (young) or at 8 weeks (adult) of age with 5 � 105 organisms/g of body weight. At the indicated time points, lung organism (PC) burden (A), P.murina-specific serum IgG (B), percent activated alveolar macrophages in the BALF (C), percent CD4� T cells in the BALF (D), and TNF concentrations in theBALF (E) were determined as described in Materials and Methods. Activation status of CD11c� F4/80� macrophages was determined by expression of CD11band MHC-II (Ia). Data represent the means � standard deviations (SD) from 5 mice per group and are representative of two separate experiments. For some datapoints, error bars are smaller than the symbols. *, P � 0.05 compared to adults at the same time point. O.D. 405 nm, optical density at 405 nm.
Kurkjian et al.
2838 iai.asm.org Infection and Immunity
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 15
Nov
embe
r 20
21 b
y 12
1.12
9.11
0.11
8.

to elevated numbers of activated CD4� T cells in lung digests ofIL-10�/� mice around 3 weeks postinfection (Fig. 2C). BALF alsohad elevated, though not statistically significant, numbers of acti-vated CD4� T cells in IL-10�/� pups (Fig. 2B). The elevation ofactivated T cells in IL-10�/� mice was due to increased propor-tions of cells in the lung digest or BALF as opposed to differencesin absolute numbers of cells (data not shown). There were reducednumbers of activated CD4� T cells in the TBLN of IL-10�/� mice,likely due to increased migration to the lungs (Fig. 2D). The ele-vated T cells at later time points in the IL-10�/� pups did not affecthelp to B cells, since there were no differences in P. murina-spe-cific IgG in the sera (Fig. 2E). Examination of lung proinflamma-tory cytokines by RNase protection assay or ELISA on BALF indi-cated that at later time points (at 3 weeks of age or greater),
infected IL-10�/� mice had elevated levels of TNF-, IL-1�, IL-6,and gamma interferon (IFN-) compared to wild-type mice insome experiments, but this was not consistent among all experi-ments performed (5 separate experiments) (data not shown). To-gether, these data suggest that IL-10 has a role in controlling in-flammation in neonatal lungs. However, because differences werenot seen in the IL-10�/� pups until after the mice were 3 weeks ofage, there are clearly other mechanisms responsible for the lack ofresponsiveness to P. murina in neonates compared to adults.
Increased levels of TGF-� may play a role in the delayed mi-gration of T cells in pups. Since we found that there are factorsother than IL-10 that must contribute to the unresponsiveness ofneonatal mice to P. murina, we examined whether TGF-� is acontributing factor. TGF-�1 has long been associated with immu-
FIG 2 IL-10�/� pups clear P. murina faster than do wild-type mice. Two-day-old IL-10�/� and C57BL/6 (wild-type) pups were infected with P. murina or wereinoculated with buffer. At the indicated time points, lung organism (PC) burden (A) and activated CD4� T cells in the BALF (B), lung parenchyma (C), anddraining lymph nodes (D) were determined as described in Materials and Methods. For flow cytometry, cells were gated on CD4� lymphocytes and analyzed forCD44 and CD62L expression. (E) P. murina-specific IgG levels in sera were examined by ELISA. Data represent means � SD from 4 to 8 mice per group and arerepresentative of five separate experiments. *, P � 0.05 compared to infected wild-type mice at the same time point.
Macrophage Function in Neonatal Lungs
August 2012 Volume 80 Number 8 iai.asm.org 2839
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 15
Nov
embe
r 20
21 b
y 12
1.12
9.11
0.11
8.

nosuppression, and mice deficient in this cytokine die of a lym-phoproliferative disease by about 4 weeks of age (45). Moreover,TGF-� is one of several growth factors elevated in the lungs ofpostnatally developing mice (2, 32). A TGF-�1-specific ELISA wasperformed on supernatants from lung homogenates of uninfectedand P. murina-infected pups and adults. Lung homogenates wereused because we were unable to detect TGF-� in BALF. Unin-fected pups had significantly higher levels of total TGF-�1 per mgof lung protein at 9 and 16 days of age than uninfected adults (Fig.3A). TGF-�1 concentrations dropped somewhat in the lungs ofpups infected at 2 days of age with P. murina (Fig. 3B). At day 7postinfection, TGF-�1 levels in infected pups had dropped about30% compared to those of uninfected pups. However, the concen-tration of TGF-�1 in the lungs of infected pups at day 7 was stillabout 2-fold greater than in P. murina-infected adult lungs (Fig.3B). The higher TGF-�1 levels in pups decreased to adult levels byday 21 in both uninfected and infected pups, suggesting thatTGF-� is developmentally controlled but can be affected by aninfectious agent.
Neutralization of TGF-� in IL-10�/� mice results in in-creased inflammation and modest control of Pneumocystisgrowth. To determine whether TGF-� is responsible for the de-
layed response to P. murina in neonatal compared to adult mice,we neutralized TGF-� in vivo by injection of 10 �g/g of specificmonoclonal antibody at days �1, 4, 11, and 17 postinfection.Neutralization of TGF-� had no effect on clearance of P. murina,although we did see trends toward elevated proinflammatory cy-tokines in the lungs (data not shown). We next neutralized TGF-�1,2 in IL-10�/� mice to determine whether reduction of twoanti-inflammatory mediators would induce the neonatal immuneresponse to control P. murina growth comparable to adult mice.Since we saw only modest reduction in lung TGF-�1 levels withour previous antibody treatments, for this experiment we injectedanti-TGF-�1,2,3 MAb twice per week with doses of 50 to 200 �g/gbeginning on the day of infection with P. murina. The level ofgrowth of P. murina was not reduced in the IL-10�/� pups treatedwith anti-TGF-� (Fig. 4A). However, increased infiltration ofCD4� T cells into the alveoli of anti-TGF-�-treated IL-10�/�
mice was observed (Fig. 4B and C). Moreover, though not statis-tically significant, neutralization of TGF-� consistently resulted ina modest increase in TNF- in IL-10�/� pups at about 2 weekspostinfection (Fig. 4D). Together, these data suggest that IL-10and TGF-� contribute to the control of inflammation during theclearance phase of P. murina infection.
The neonatal lung environment contributes to reduced asso-ciation of alveolar macrophages with P. murina. Figure 1 indi-cates that P. murina grows virtually unchecked in the lungs ofinfant mice until they are about 3 weeks of age. Neutralization ofanti-inflammatory mediators did not significantly affect thegrowth of the organisms during these first 2 to 3 weeks (Fig. 2Aand 4A). We used an adoptive transfer approach to determinewhether the neonatal lung environment had a significant effect onalveolar macrophage association with P. murina. Alveolar macro-phages were isolated from adult or 7-day-old mice expressinggreen fluorescent protein (GFP) under the control of the actinpromoter and inoculated into the lungs of 8-day-old or adultmice, respectively. Adult and pup GFP� alveolar macrophagesexpressed CD11c but were CD11b negative at the time of transfer(data not shown) and were considered unactivated. Mice wereinfected 24 h later with P. murina labeled with DDAO, and 18 hafter infection lungs were lavaged and analyzed by flow cytometry.As shown in Fig. 5A, donor neonatal and host adult alveolar mac-rophages could be distinguished by expression of GFP. The pro-portion of CD11c� CD11b� cells that had bound P. murina andwere positive for DDAO was not different between the adult GFP�
host adult cells and the GFP� transferred pup cells (Fig. 5B).Moreover, the proportion of CD11c� cells that upregulatedCD11b and bound P. murina was similar in donor pups and hostadults (Fig. 5B). Together, these data confirm that alveolar mac-rophages from neonatal mice associate with P. murina and be-come activated, as determined by CD11b expression in an envi-ronment that is less immunosuppressive than the neonatal lung.
To confirm whether the neonatal lung environment is suppres-sive for alveolar macrophage interactions with P. murina, weadoptively transferred GFP� alveolar macrophages from adultmice into the lungs of 7-day-old mice, followed by infection withDDAO-labeled P. murina the next day. As shown in Fig. 5C, lessthan 3% of either host pup or transferred adult alveolar macro-phages were positive for DDAO 24 h after infection, indicative ofa lack of association with P. murina. This corresponded to lessthan 20% of the alveolar macrophages from either host or donorexpressing CD11b upon infection. Together, these data demon-
FIG 3 TGF-�1 is constitutively expressed at high levels in neonatal lungs.Lungs were obtained from uninfected pups or from mice infected with P.murina at 2 days of age or as adults and were snap-frozen. Lungs were homog-enized in protease inhibitor cocktail, and TGF-�1 concentrations in the super-natants were determined by ELISA and normalized for total protein levels.Two separate experiments show TGF-�1 concentrations in uninfected mice(A) and TGF-�1 concentrations in mice infected as neonates or adults (B).Data represent means � SD from 3 to 5 mice per group and are representativeof two separate experiments. *, P � 0.05 compared to pups at the same timepoint.
Kurkjian et al.
2840 iai.asm.org Infection and Immunity
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 15
Nov
embe
r 20
21 b
y 12
1.12
9.11
0.11
8.
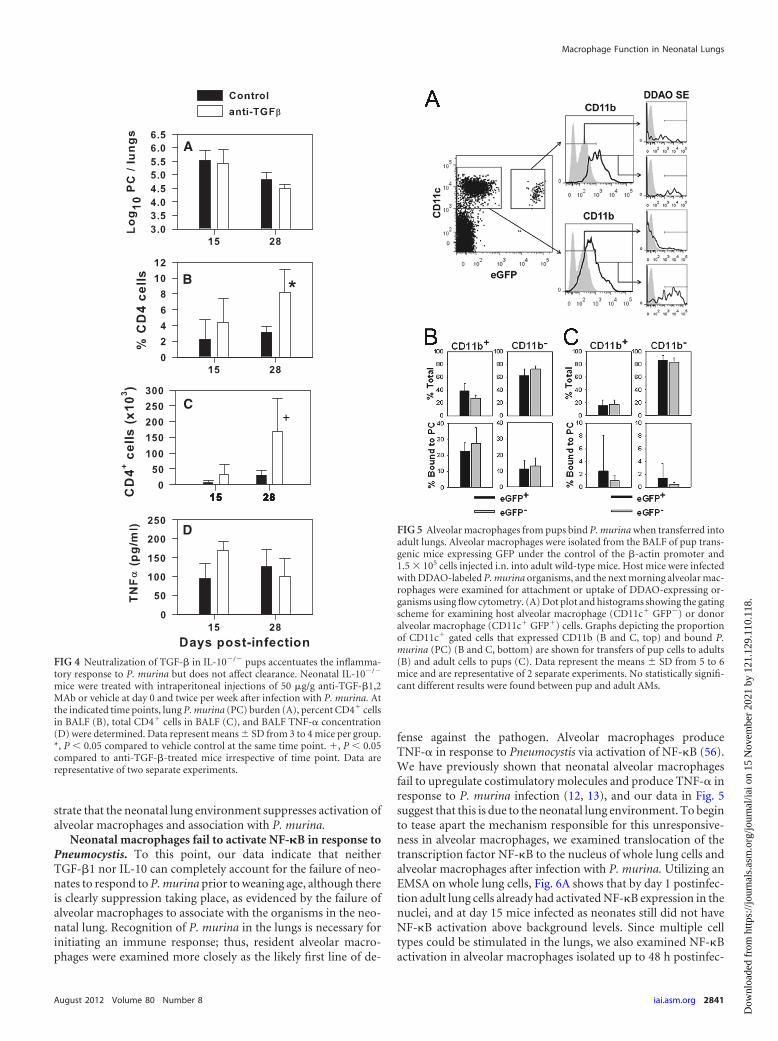
strate that the neonatal lung environment suppresses activation ofalveolar macrophages and association with P. murina.
Neonatal macrophages fail to activate NF-�B in response toPneumocystis. To this point, our data indicate that neitherTGF-�1 nor IL-10 can completely account for the failure of neo-nates to respond to P. murina prior to weaning age, although thereis clearly suppression taking place, as evidenced by the failure ofalveolar macrophages to associate with the organisms in the neo-natal lung. Recognition of P. murina in the lungs is necessary forinitiating an immune response; thus, resident alveolar macro-phages were examined more closely as the likely first line of de-
fense against the pathogen. Alveolar macrophages produceTNF- in response to Pneumocystis via activation of NF-�B (56).We have previously shown that neonatal alveolar macrophagesfail to upregulate costimulatory molecules and produce TNF- inresponse to P. murina infection (12, 13), and our data in Fig. 5suggest that this is due to the neonatal lung environment. To beginto tease apart the mechanism responsible for this unresponsive-ness in alveolar macrophages, we examined translocation of thetranscription factor NF-�B to the nucleus of whole lung cells andalveolar macrophages after infection with P. murina. Utilizing anEMSA on whole lung cells, Fig. 6A shows that by day 1 postinfec-tion adult lung cells already had activated NF-�B expression in thenuclei, and at day 15 mice infected as neonates still did not haveNF-�B activation above background levels. Since multiple celltypes could be stimulated in the lungs, we also examined NF-�Bactivation in alveolar macrophages isolated up to 48 h postinfec-
FIG 4 Neutralization of TGF-� in IL-10�/� pups accentuates the inflamma-tory response to P. murina but does not affect clearance. Neonatal IL-10�/�
mice were treated with intraperitoneal injections of 50 �g/g anti-TGF-�1,2MAb or vehicle at day 0 and twice per week after infection with P. murina. Atthe indicated time points, lung P. murina (PC) burden (A), percent CD4� cellsin BALF (B), total CD4� cells in BALF (C), and BALF TNF- concentration(D) were determined. Data represent means � SD from 3 to 4 mice per group.*, P � 0.05 compared to vehicle control at the same time point. �, P � 0.05compared to anti-TGF-�-treated mice irrespective of time point. Data arerepresentative of two separate experiments.
FIG 5 Alveolar macrophages from pups bind P. murina when transferred intoadult lungs. Alveolar macrophages were isolated from the BALF of pup trans-genic mice expressing GFP under the control of the �-actin promoter and1.5 � 105 cells injected i.n. into adult wild-type mice. Host mice were infectedwith DDAO-labeled P. murina organisms, and the next morning alveolar mac-rophages were examined for attachment or uptake of DDAO-expressing or-ganisms using flow cytometry. (A) Dot plot and histograms showing the gatingscheme for examining host alveolar macrophage (CD11c� GFP�) or donoralveolar macrophage (CD11c� GFP�) cells. Graphs depicting the proportionof CD11c� gated cells that expressed CD11b (B and C, top) and bound P.murina (PC) (B and C, bottom) are shown for transfers of pup cells to adults(B) and adult cells to pups (C). Data represent the means � SD from 5 to 6mice and are representative of 2 separate experiments. No statistically signifi-cant different results were found between pup and adult AMs.
Macrophage Function in Neonatal Lungs
August 2012 Volume 80 Number 8 iai.asm.org 2841
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 15
Nov
embe
r 20
21 b
y 12
1.12
9.11
0.11
8.

tion. Cells were lavaged from the lungs of mice infected at 2 days ofage and as adults and pooled by group. The purity of alveolarmacrophages was determined by differential counts and was 92%in pups at both time points and 86 and 91% in adults at 24 and 48h postinfection, respectively. The rest of the cells were neutrophilswith no lymphocytes detected in any of the pools of cells. Utilizinga plate-based assay for nuclear extraction and NF-�B detection,we found that by 48 h, macrophages from adult lungs had signif-icantly elevated levels of translocated NF-�B, whereas macro-phages from neonatal mice did not (Fig. 6B). To determinewhether this was an intrinsic defect or due to the lung environ-ment, we next isolated alveolar macrophages from adult and 10-to 14-day-old mice and stimulated them in vitro with either LPS orsonicated P. murina antigen. The purity of the alveolar macro-phages was more than 95%. As shown in Fig. 6C, LPS and soni-cated P. murina induced significant activation of NF-�B in cellsfrom adult mice within 1 h; however, NF-�B activation in cellsfrom pups was at or below background levels. This correspondedto failure of pup alveolar macrophages to produce TNF- abovebackground levels within 24 h after in vitro stimulation (data notshown). We next utilized fluorescence microscopy to determine
the age at which alveolar macrophages respond similarly to adults.Alveolar macrophages were isolated from mice of ages rangingfrom 14 days to adult, stimulated with P. murina in vitro, andstained for p65 of NF-�B using fluorescently labeled antibody (seeFig. S2 in supplemental material). The proportion of alveolarmacrophages with p65 colocalized in the nucleus (stained withDAPI) was quantitated microscopically. Figure 6D shows that theproportion of macrophages with activated NF-�B increased overtime, and by about 5 weeks of age the proportion of activatedmacrophages was approaching that of adults. Together, these datademonstrate not only that the neonatal lung environment con-tributes to unresponsiveness to P. murina but also that neonatalalveolar macrophages are intrinsically unresponsive to stimula-tion with P. murina as well as LPS.
Recognition of unopsonized P. murina by alveolar macro-phages occurs through pattern recognition receptors, includingDectin-1, which recognize �-glucan in the cyst cell wall (47), ormannose receptors, which recognize mannosylated glycoproteinsexpressed on the cell surface of both cyst and trophozoite forms ofthe organisms (15). Dectin-1 transduces a signal through anITAM-like motif that results in activation of NF-�B, while man-
FIG 6 Activation of NF-�B is delayed in the lungs and alveolar macrophages of neonates in response to P. murina. (A and B) Neonatal and adult mice wereinfected with 5 � 105 P. murina organisms/g. At the indicated time points, lungs were snap-frozen and then homogenized prior to nuclear extraction (A), or theywere lavaged and nuclei extracted from alveolar macrophages (B). (A) Nuclear translocation of NF-�B was determined for lungs from individual mice by EMSA,and data are expressed as means � SD of relative densitometry units (arbitrary units) from 3 mice per group. �, P � 0.05 compared to pup lungs irrespective oftime point. (B) Nuclear translocation of pooled samples was determined by a colorimetric plate-based commercial assay, and data are expressed as means � SDof replicate wells (optical density read at 450 nm). *, P � 0.05 compared to pups at the same time point. Alveolar macrophages were isolated from uninfected10-day-old (C) and adult mice or mice of various ages (D) and rested in culture medium overnight. (C) Alveolar macrophages were stimulated with LPS orsonicated P. murina, and 1 h later cells were lysed and nuclei extracted. NF-�B translocation was determined from in vitro-stimulated cells using the plate-basedassay. Data are expressed as a percentage of the uninfected control, because the background NF-�B activation was consistently higher in neonatal macrophagesthan in adults. The means � SD from replicate wells normalized to controls are shown. No statistical differences were found. (D) Alveolar macrophages frommice at the indicated ages were stimulated with P. murina and stained with DAPI and fluorescent antibody specific for p65. The proportion of alveolarmacrophages with colocalized p65 and DAPI was determined by counting microscopically. Data represent the proportion of alveolar macrophages (AMs) withp65 in the nuclei per minimum of 100 cells counted. Fluorescent micrographs of alveolar macrophages can be found in Fig. S2 in the supplemental material.
Kurkjian et al.
2842 iai.asm.org Infection and Immunity
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 15
Nov
embe
r 20
21 b
y 12
1.12
9.11
0.11
8.
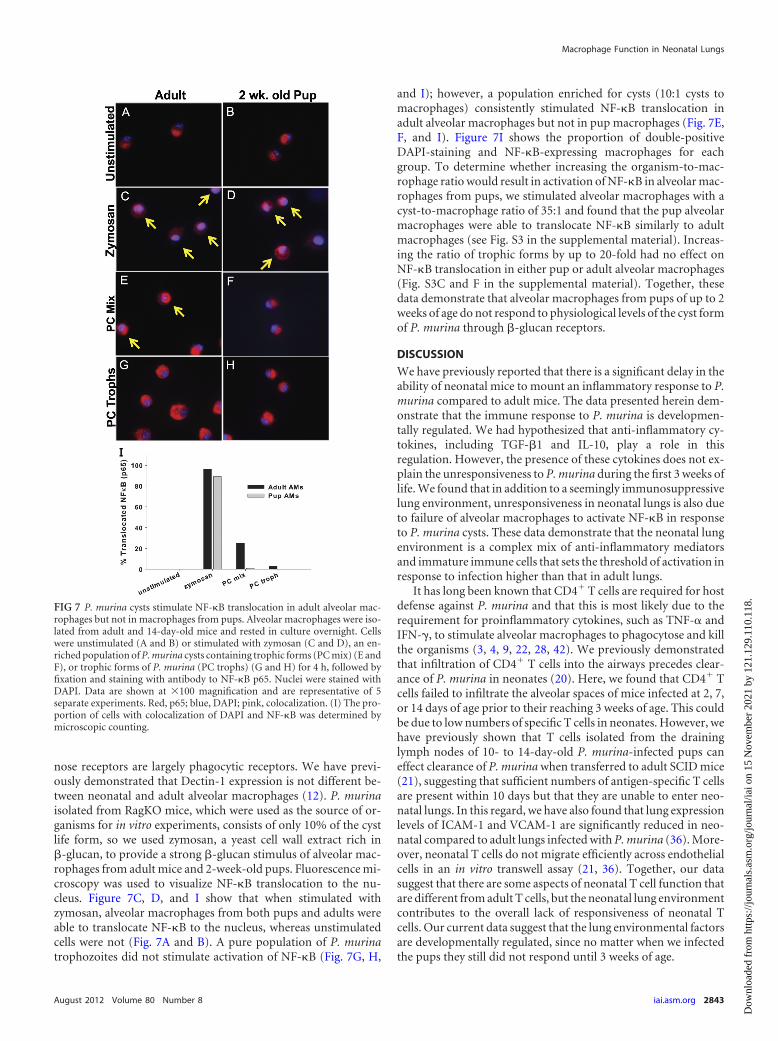
nose receptors are largely phagocytic receptors. We have previ-ously demonstrated that Dectin-1 expression is not different be-tween neonatal and adult alveolar macrophages (12). P. murinaisolated from RagKO mice, which were used as the source of or-ganisms for in vitro experiments, consists of only 10% of the cystlife form, so we used zymosan, a yeast cell wall extract rich in�-glucan, to provide a strong �-glucan stimulus of alveolar mac-rophages from adult mice and 2-week-old pups. Fluorescence mi-croscopy was used to visualize NF-�B translocation to the nu-cleus. Figure 7C, D, and I show that when stimulated withzymosan, alveolar macrophages from both pups and adults wereable to translocate NF-�B to the nucleus, whereas unstimulatedcells were not (Fig. 7A and B). A pure population of P. murinatrophozoites did not stimulate activation of NF-�B (Fig. 7G, H,
and I); however, a population enriched for cysts (10:1 cysts tomacrophages) consistently stimulated NF-�B translocation inadult alveolar macrophages but not in pup macrophages (Fig. 7E,F, and I). Figure 7I shows the proportion of double-positiveDAPI-staining and NF-�B-expressing macrophages for eachgroup. To determine whether increasing the organism-to-mac-rophage ratio would result in activation of NF-�B in alveolar mac-rophages from pups, we stimulated alveolar macrophages with acyst-to-macrophage ratio of 35:1 and found that the pup alveolarmacrophages were able to translocate NF-�B similarly to adultmacrophages (see Fig. S3 in the supplemental material). Increas-ing the ratio of trophic forms by up to 20-fold had no effect onNF-�B translocation in either pup or adult alveolar macrophages(Fig. S3C and F in the supplemental material). Together, thesedata demonstrate that alveolar macrophages from pups of up to 2weeks of age do not respond to physiological levels of the cyst formof P. murina through �-glucan receptors.
DISCUSSION
We have previously reported that there is a significant delay in theability of neonatal mice to mount an inflammatory response to P.murina compared to adult mice. The data presented herein dem-onstrate that the immune response to P. murina is developmen-tally regulated. We had hypothesized that anti-inflammatory cy-tokines, including TGF-�1 and IL-10, play a role in thisregulation. However, the presence of these cytokines does not ex-plain the unresponsiveness to P. murina during the first 3 weeks oflife. We found that in addition to a seemingly immunosuppressivelung environment, unresponsiveness in neonatal lungs is also dueto failure of alveolar macrophages to activate NF-�B in responseto P. murina cysts. These data demonstrate that the neonatal lungenvironment is a complex mix of anti-inflammatory mediatorsand immature immune cells that sets the threshold of activation inresponse to infection higher than that in adult lungs.
It has long been known that CD4� T cells are required for hostdefense against P. murina and that this is most likely due to therequirement for proinflammatory cytokines, such as TNF- andIFN-, to stimulate alveolar macrophages to phagocytose and killthe organisms (3, 4, 9, 22, 28, 42). We previously demonstratedthat infiltration of CD4� T cells into the airways precedes clear-ance of P. murina in neonates (20). Here, we found that CD4� Tcells failed to infiltrate the alveolar spaces of mice infected at 2, 7,or 14 days of age prior to their reaching 3 weeks of age. This couldbe due to low numbers of specific T cells in neonates. However, wehave previously shown that T cells isolated from the draininglymph nodes of 10- to 14-day-old P. murina-infected pups caneffect clearance of P. murina when transferred to adult SCID mice(21), suggesting that sufficient numbers of antigen-specific T cellsare present within 10 days but that they are unable to enter neo-natal lungs. In this regard, we have also found that lung expressionlevels of ICAM-1 and VCAM-1 are significantly reduced in neo-natal compared to adult lungs infected with P. murina (36). More-over, neonatal T cells do not migrate efficiently across endothelialcells in an in vitro transwell assay (21, 36). Together, our datasuggest that there are some aspects of neonatal T cell function thatare different from adult T cells, but the neonatal lung environmentcontributes to the overall lack of responsiveness of neonatal Tcells. Our current data suggest that the lung environmental factorsare developmentally regulated, since no matter when we infectedthe pups they still did not respond until 3 weeks of age.
FIG 7 P. murina cysts stimulate NF-�B translocation in adult alveolar mac-rophages but not in macrophages from pups. Alveolar macrophages were iso-lated from adult and 14-day-old mice and rested in culture overnight. Cellswere unstimulated (A and B) or stimulated with zymosan (C and D), an en-riched population of P. murina cysts containing trophic forms (PC mix) (E andF), or trophic forms of P. murina (PC trophs) (G and H) for 4 h, followed byfixation and staining with antibody to NF-�B p65. Nuclei were stained withDAPI. Data are shown at �100 magnification and are representative of 5separate experiments. Red, p65; blue, DAPI; pink, colocalization. (I) The pro-portion of cells with colocalization of DAPI and NF-�B was determined bymicroscopic counting.
Macrophage Function in Neonatal Lungs
August 2012 Volume 80 Number 8 iai.asm.org 2843
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 15
Nov
embe
r 20
21 b
y 12
1.12
9.11
0.11
8.

TGF-� and IL-10 are known to play prominent roles in down-regulating immune responses, and both can be secreted by a num-ber of cell types (27, 34). In addition, TGF-� is involved in theintricate orchestration of lung branching morphogenesis duringdevelopment (25, 31). Both TGF-� and IL-10 are known to beconstitutively expressed in adult lungs, presumably to regulate cellturnover, host defense, and matrix protein metabolism (10, 16).We previously found that adult IL-10�/� mice clear P. murinaslightly faster than wild-type mice, and this correlated with a moreintense inflammatory response (38). This is consistent with otherstudies that demonstrate that IL-10 negatively regulates macro-phage function (6, 16, 17, 38, 40, 46, 55). As with our studies ofadult IL-10�/� mice, we observed improvement in clearance of P.murina in IL-10�/� pups, arguing that IL-10 contributes to theimmunosuppressive environment of neonatal lungs but is not theonly factor involved. Although we can conclude that IL-10 con-tributes to the overall suppressive environment of neonatal lungs,deficiency of IL-10 only had an effect on mice after they reached 3weeks of age, suggesting alternate mechanisms of immune sup-pression in the lungs of very young mice.
Our studies did not address which cell types in neonatal mouselungs produce TGF-�1 and/or IL-10, although we are quite inter-ested in this. Others have shown that lung epithelial cells can pro-duce these cytokines as well as monocytes, alveolar macrophages,and other cell types (10, 16, 49). We favor the hypothesis that lungstructural cells or macrophages are the source of these cytokinesrather than regulatory T cells, because we see very few CD4� Tcells in the airways of neonatal mice prior to 3 weeks of age. In P.murina-infected pups, CD4� T cells accumulate in the draininglymph nodes and lung parenchyma in very low numbers arounddays 10 to 14 postinfection (data not shown). We have examinedthese cells for expression of CD25 as a cursory look for regulatoryT cells but have not found them in great enough numbers to beconfident of the results. Experiments under way in our laboratorywill determine the source of both TGF-�1 and IL-10 in the lungsof infant mice. We were surprised that neutralization of TGF-�and IL-10 in concert did not result in appreciable changes in alve-olar inflammation or control of P. murina prior to mice reaching3 weeks of age. In spite of this, we found that adult alveolar mac-rophages adoptively transferred into neonatal lungs did not asso-ciate with fluorescently labeled P. murina, although neonatal al-veolar macrophages transferred to adult lungs were able to bindorganisms. This is consistent with an immunosuppressive lungenvironment in neonatal mice that promotes growth of P. murinaand inhibits nonopsonic uptake of organisms. Together, thesedata could be interpreted to mean that anti-inflammatory cyto-kines are important in inhibiting binding and uptake of P. murinaearly after infection in neonates but have a more dramatic effecton induction of adaptive immunity.
Alveolar macrophages have been demonstrated to be the effec-tor cells most responsible for killing of Pneumocystis (28, 47), andexpression of the �-glucan receptor Dectin-1 has been shown tobe an important pathway for macrophage recognition and killingof the organisms (44, 47). Here, we show that the cyst form of theorganisms stimulates NF-�B translocation in adult alveolar mac-rophages but not in macrophages from neonates. We have previ-ously demonstrated that the expression levels of Dectin-1 arecomparable on neonatal and adult alveolar macrophages, whichmay mean that signaling through the Dectin-1 receptor is im-paired in neonatal macrophages (12). Consistent with this, the
data presented herein indicate that alveolar macrophages fromneonates do not activate NF-�B in the same time frame as adultmacrophages. This is an age-dependent response, as alveolar mac-rophages increasingly responded to P. murina in vitro from 2weeks of age through 5 weeks of age. In contrast, zymosan, a yeastcell wall extract, provided a potent signal for NF-�B translocationin neonatal alveolar macrophages, and PC was able to stimulateactivation in adults, albeit to a lesser extent than zymosan. Thisdifference between zymosan and PC in the ability to stimulateneonatal alveolar macrophages could have to do with strength ofsignal, since increasing the P. murina-to-macrophage ratio re-sulted in translocation of NF-�B p65. Alternatively, the signalingpathways differ in response to zymosan and PC. We have somegene expression data suggesting that some signaling intermediatesin the NF-�B pathways are differentially expressed depending onthe stimulus. We are quite interested in these genes and are in theprocess of performing experiments to understand these mecha-nisms.
Pneumocystis �-glucans have been shown to signal NF-�B ac-tivation in alveolar epithelial cells (14, 52). However, a recentstudy with Dectin-1�/� mice indicated that Dectin-1 was impor-tant for recognizing and killing of P. murina by macrophages butnot for production of cytokines, such as TNF- (44). Signalingthrough MyD88 stimulated cytokine production in Dectin-1�/�
macrophages (44). There is some evidence that Dectin-1 can in-teract with TLR-2 (19), and it was recently demonstrated thatPneumocystis induced NF-�B activation and cytokine productionthrough TLR-2 ligation on alveolar macrophages (56). We havealso shown that TLR-2 expression levels are comparable on adultand neonatal alveolar macrophages (12). We hypothesize thatthere is a dysregulation of the signaling pathways of Dectin-1 andTLR-2 in neonatal alveolar macrophages that is responsible for theinherent unresponsiveness to P. murina. We can envision severalmechanisms for this, including overexpression of control proteinssuch as A20 or epigenetic regulation, as has been shown for neo-natal T cells (41). Preliminary data from our laboratory indicatethat several signaling intermediates in the NF-�B pathways aresignificantly downregulated in neonatal alveolar macrophagescompared to adults. We are in the process of determining themechanism of this downregulation.
In summary, we have shown that the response to P. murina isdevelopmentally controlled. This likely is not due solely to thepresence of anti-inflammatory cytokines present in the postna-tally developing lungs but also to factors intrinsic to immune cells,such as alveolar macrophages. Since approximately 80% of alveo-lar space is developed postnatally, one can envision that it is crit-ical to strike a balance between unresponsiveness to dangerouspathogens and an overexuberant immune response that coulddamage sensitive lungs.
ACKNOWLEDGMENTS
This work was funded by Public Health Service grants HL062053 andHL088989 from the National Heart, Lung, and Blood Institute to B.A.G.,fellowship grant RT-051-N from the American Lung Association to M.Q.,VA Merit Review to S.A.B., and resources provided by the Veteran’s Af-fairs Medical Center, Lexington, KY.
REFERENCES1. Adkins B, Leclerc C, Marshall-Clarke S. 2004. Neonatal adaptive immu-
nity comes of age. Nat. Rev. Immunol. 4:553–564.2. Alejandre-Alcazar MA, et al. 2008. TGF-� signaling is dynamically reg-
Kurkjian et al.
2844 iai.asm.org Infection and Immunity
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 15
Nov
embe
r 20
21 b
y 12
1.12
9.11
0.11
8.

ulated during the alveolarization of rodent and human lungs. Dev. Dy-namics 237:259 –269.
3. Beck JM, et al. 1991. Reduction in intensity of Pneumocystis cariniipneumonia in mice by aerosol administration of gamma interferon. In-fect. Immun. 59:3859 –3862.
4. Beck JM, et al. 1991. Inflammatory responses to Pneumocystis carinii inmice selectively depleted of helper T lymphocytes. Am. J. Respir. Cell Mol.Biol. 5:186 –197.
5. Beck JM, Warnock ML, Kaltreider HB, Shellito JE. 1993. Host defensesagainst Pneumocystis carinii in mice selectively depleted of CD4� lympho-cytes. Chest 103:116s–118s.
6. Belkaid Y, et al. 2001. The role of interleukin (IL)-10 in the persistence ofLeishmania major in the skin after healing and the therapeutic potential ofanti-IL-10 receptor antibody for sterile cure. J. Exp. Med. 194:1497–1506.
7. Bourbon JR. 1999. Gene expression in alveolar development. In GaultierC, Bourbon JR, Post M (ed), Lung development. American PhysiologicalSociety, New York, NY.
8. CDC. 1995. 1995 Revised guidelines for prophylaxis against Pneumocystiscarinii pneumonia for children infected with or perinatally exposed tohuman immunodeficiency virus. MMWR Morb. Mortal. Wkly. Rep. 44:1–11.
9. Chen W, Havell EA, Harmsen AG. 1992. Importance of endogenoustumor necrosis factor alpha and gamma interferon in host resistanceagainst Pneumocystis carinii infection. Infect. Immun. 60:1279 –1284.
10. Coker RK, et al. 1996. Diverse cellular TGF-�1 and TGF-�3 gene expres-sion in normal human and murine lung. Eur. Respir. J. 9:2501–2507.
11. Dasch JR, Pace DR, Waegell W, Inenaga D, Ellingsworth L. 1989.Monoclonal antibodies recognizing transforming growth factor-beta.Bioactivity neutralization and transforming growth factor beta 2 affinitypurification. J. Immunol. 142:1536 –1541.
12. Empey KM, Hollifield M, Garvy BA. 2007. Exogenous heat-killed Esch-erichia coli improves alveolar macrophage activity and reduces Pneumo-cystis carinii lung burden in infant mice. Infect. Immun. 75:3382–3393.
13. Empey KM, Hollifield M, Schuer K, Gigliotti F, Garvy BA. 2004. Passiveimmunization of neonatal mice against Pneumocystis carinii f. sp. murisenhances control of infection without stimulating inflammation. Infect.Immun. 72:6211– 6220.
14. Evans SE, et al. 2005. Pneumocystis cell wall beta-glucans stimulate alve-olar epithelial cell chemokine generation through nuclear factor-kappaB-dependent mechanisms. Am. J. Respir. Cell Mol. Biol. 32:490 – 497.
15. Ezekowitz RA, et al. 1991. Uptake of Pneumocystis carinii mediated by themacrophage mannose receptor. Nature 351:155–158.
16. Fernandez S, Jose P, Avidiushko MG, Kaplan AM, Cohen DA. 2004.Inhibition of IL-10 receptor function in alveolar macrophages by toll-likereceptor agonists. J. Immunol. 172:2613–2620.
17. Fiorentino DF, Zlotnik A, Mosmann TR, Howard M, O’Garra A. 1991.IL-10 inhibits cytokine production by activated macrophages. J. Immu-nol. 147:3815–3822.
18. Gajdusek DC. 1957. Pneumocystis carinii– etiological agent of interstitialplasma cell pneumonia of premature and young infants. Pediatrics 19:543–565.
19. Gantner BN, Simmons RM, Canavera SJ, Akira S, Underhill DM. 2003.Collaborative induction of inflammatory responses by Dectin-1 and Toll-like receptor 2. J. Exp. Med. 197:1107–1117.
20. Garvy BA, Harmsen AG. 1996. Susceptibility to Pneumocystis cariniiinfection: host responses of neonatal mice from immune or naive mothersand of immune or naive adults. Infect. Immun. 64:3987–3992.
21. Garvy BA, Qureshi M. 2000. Delayed inflammatory response to Pneu-mocystis carinii infection in neonatal mice is due to an inadequate lungenvironment. J. Immunol. 165:6480 – 6486.
22. Harmsen AG, Stankiewicz M. 1990. Requirement for CD4� cells inresistance to Pneumocystis carinii pneumonia in mice. J. Exp. Med. 172:937–945.
23. Harmsen AG, Stankiewicz M. 1991. T cells are not sufficient for resistanceto Pneumocystis carinii pneumonia in mice. J. Protozool. 38:44S– 45S.
24. Hoffman OA, Standing JE, Limper AH. 1993. Pneumocystis cariniistimulates tumor necrosis factor-alpha release from alveolar macrophagesthrough a beta-glucan-mediated mechanism. J. Immunol. 150:3932–3940.
25. Jaskoll T, Choy HA, Melnick M. 1996. The glucocorticoid-glucocorticoid receptor signal transduction pathway, transforminggrowth factor-�, and embryonic mouse lung development in vivo. Pediatr.Res. 39:749 –759.
26. Levy O. 2007. Innate immunity of the newborn: basic mechanisms andclinical correlates. Nat. Rev. Immunol. 7:379 –390.
27. Li MO, Wan YY, Sanjabi S, Roberston A-KL, Flavell RA. 2006. Trans-forming growth factor-� regulation of immune responses. Annu. Rev.Immunol. 24:99 –146.
28. Limper AH, Hoyte JS, Standing JE. 1997. The role of alveolar macro-phages in Pneumocystis carinii degradation and clearance from the lung. J.Clin. Investig. 99:2110 –2117.
29. Lund FE, Schuer K, Hollifield M, Randall TD, Garvy BA. 2003. Clear-ance of Pneumocystis carinii in mice is dependent on b cells but not on P.carinii-specific antibody. J. Immunol. 171:1423–1430.
30. Marcotte H, et al. 1996. Pneumocystis carinii infection in transgenic Bcell-deficient mice. J. Infect. Dis. 173:1034 –1037.
31. Mendelson CR. 2000. Role of transcription factors in fetal lung develop-ment and surfactant protein gene expression. Annu. Rev. Physiol. 62:875–915.
32. Nakanishi H, Sugiura T, Streisand JB, Lonning SM, Roberts JD, Jr.2007. TGF-�-neutralizing antibodies improve pulmonary alveologenesisand vasculogenesis in the injured newborn lung. Am. J. Physiol. Lung CellPhysiol. 293:L151–L161.
33. Peglow SL, et al. 1990. Serologic responses to Pneumocystis carinii anti-gens in health and disease. J. Infect. Dis. 161:296 –306.
34. Pestka S, et al. 2004. Interleukin-10 and related cytokines and receptors.Annu. Rev. Immunol. 22:929 –979.
35. Qureshi M, Empey KM, Garvy BA. 2005. Modulation of proinflamma-tory responses to Pneumocystis carinii f. sp. muris in neonatal mice bygranulocyte-macrophage colony-stimulating factor and IL-4: role ofAPCs. J. Immunol. 174:441– 448.
36. Qureshi MH, Cook-Mills J, Doherty DE, Garvy BA. 2003. TNF-alpha-dependent ICAM-1- and VCAM-1-mediated inflammatory responses aredelayed in neonatal mice infected with Pneumocystis carinii. J. Immunol.171:4700 – 4707.
37. Qureshi MH, Garvy BA. 2001. Neonatal T cells in an adult lung environ-ment are competent to resolve Pneumocystis carinii pneumonia. J. Immu-nol. 166:5704 –5711.
38. Qureshi MH, Harmsen AG, Garvy BA. 2003. IL-10 modulates hostresponses and lung damage induced by Pneumocystis carinii infection. J.Immunol. 170:1002–1009.
39. Rogers MF, et al. 1987. Acquired immunodeficiency syndrome in chil-dren: report of the Centers for Disease Control national surveillance,1982–1985. Pediatrics 79:1008 –1014.
40. Romani L, et al. 1994. Neutralization of IL-10 up-regulates nitric oxideproduction and protects susceptible mice from challenge with Candidaalbicans. J. Immunol. 152:3514 –3521.
41. Rose S, Lichtenheld M, Foote MR, Adkins B. 2007. Murine neonatalCD4� cells are poised for rapid Th2 effector-like function. J. Immunol.178:2667–2678.
42. Roths JB, Sidman CL. 1992. Both immunity and hyperresponsiveness toPneumocystis carinii result from transfer of CD4� but not CD8� T cellsinto severe combined immunodeficiency mice. J. Clin. Investig. 90:673–678.
43. Roths JB, Sidman CL. 1993. Single and combined humoral and cell-mediated immunotherapy of Pneumocystis carinii pneumonia in immu-nodeficient SCID mice. Infect. Immun. 61:1641–1649.
44. Saijo S, et al. 2007. Dectin-1 is required for host defense against Pneumo-cystis carinii but not against Candida albicans. Nat. Immunol. 8:39 – 46.
45. Shull MM, et al. 1992. Targeted disruption of the mouse transforminggrowth factor-�1 gene results in multifocal inflammatory disease. Nature359:393–399.
46. Soltys J, Bonfield T, Chmiel J, Berger M. 2002. Functional IL-10 defi-ciency in the lung of cystic fibrosis (cftr�/�) and IL-10 knockout micecauses increased expression and function of B7 costimulatory moleculeson alveolar macrophages. J. Immunol. 168:1903–1910.
47. Steele C, et al. 2003. Alveolar macrophage-mediated killing of Pneumo-cystis carinii f. sp. muris involves molecular recognition by the Dectin-1�-glucan receptor. J. Exp. Med. 198:1677–1688.
48. Ten Have-Opbroek AAW. 1981. The development of the lung in mam-mals: an analysis of concepts and findings. Am. J. Anat. 162:201–219.
49. Toossi Z, et al. 1996. Decreased production of TGF-beta 1 by humanalveolar macrophages compared with blood monocytes. J. Immunol. 156:3461–3468.
50. Vargas SL, et al. 2001. Search for primary infection by Pneumocystis
Macrophage Function in Neonatal Lungs
August 2012 Volume 80 Number 8 iai.asm.org 2845
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 15
Nov
embe
r 20
21 b
y 12
1.12
9.11
0.11
8.

carinii in a cohort of normal, healthy infants. Clin. Infect. Dis. 32:855–861.
51. Vargas SL, et al. 1999. Association of primary Pneumocystis carinii infec-tion and sudden infant death syndrome. Clin. Infect. Dis. 29:1489 –1493.
52. Wang J, et al. 2005. Pneumocystis carinii activates the NF-kappa B sig-naling pathway in alveolar epithelial cells. Infect. Immun. 73:2766 –2777.
53. Wang Y, et al. 2004. Activation of nuclear factor kappa B in vivo selec-tively protects the murine small intestine against ionizing radiation-induced damage. Cancer Res. 64:6240 – 6246.
54. Wert SE. 2004. Normal and abnormal structural development of the lung.In Polin RA, Fox WA, Abman SH (ed), Fetal and neonatal physiology, 3rded. Saunders, Philadelphia, PA.
55. Willment JA, et al. 2003. Dectin-1 expression and function are enhancedon alternatively activated and GM-CSF-treated macrophages and are neg-atively regulated by IL-10, dexamethasone, and lipopolysaccharide. J. Im-munol. 171:4569 – 4573.
56. Zhang C, et al. 2006. Toll-like receptor 2 mediates alveolar macrophageresponse to Pneumocystis murina. Infect. Immun. 74:1857–1864.
57. Zhou D, et al. 1999. A high dose of ionizing radiation induces tissue-specific activation of nuclear factor-�B in vivo. Radiat. Res. 151:703–709.
58. Zhou D, et al. 2001. Effects of NF-�B1 (p50) targeted gene disruption onionizing radiation-induced NF-�B activation and TNF, IL-1, IL-1�,and IL-6 mRNA expression in vivo. Int. J. Radiat. Oncol. Biol. Phys. 77:763–772.
Kurkjian et al.
2846 iai.asm.org Infection and Immunity
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 15
Nov
embe
r 20
21 b
y 12
1.12
9.11
0.11
8.