amino acid sequence of the ,8 subunit of follicle ... · phan occurs at position 33 in human fsh-/3...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 251, No. 4, Issue of February 25. pp. 993-1005. 1976
Printed in US’.A.
Amino Acid Sequence of the ,8 Subunit of Follicle-stimulating Hormone from Human Pituitary Glands*
(Received for publication, June 26, 1975)
BRIJ B. SAXENA$ AND PREMILA RATHNAM
From the Division of Endocrinology, Department of Medicine, Cornell University Medical College, New York, New York 10021
The p subunit of follicle-stimulating hormone (FSH-@) from human pituitary glands was reduced and S-aminoethylated prior to thermolytic, tryptic, and chymotryptic digestions. Each digest was gel-filtered on Sephadex G-50 to separate the glycopeptides from the peptides. The glycopeptides and the peptides were isolated by high voltage paper electrophoresis at pH 6, 3.5, and 2.0. The purity of the isolated peptides was confirmed by amino acid analyses. The amino acid sequences of peptides were determined by Edman degradation followed by subtractive amino acid analysis and, in certain cases, confirmed by dansylation. COOH-terminal sequences of the peptides were determined by digestion with carboxypepti- dases A and B and by hydrazinolysis.
The tryptophan content of human follicle-stimulating hormone, of the /3 subunit of human follicle-stimulating hormone, and of the glycopeptides obtained from the enzymic digests was determined by fluorescence spectra, titration against N-bromosuccinimide, calorimetric estimation with p-dimethyl aminobenzaldehyde, hydrolysis with methane sulfonic acid containing 0.2% tryptamine followed by amino acid analysis, microbiological assay, and sequence analysis. The presence of 1 tryptophan residue in the p subunit was indicated.
Human FSH-P consists of 118 amino acid residues, with a predominant proportion of molecules having 108 residues due to microheterogeneity at the NH, and COOH termini. The sequence indicates an abundance of threonine, glutamic acid, and cysteine residues. The NH,-terminal portion (up to 32 residues) shows homology with the /3 subunits of other glycoprotein hormones such as luteinizing hormone, human chorionic gonadotropin, and thyroid-stimulating hormone. However, the sequence differs a great deal from the others in the rest of the molecule, confirming the specificity of the fi subunit of these hormones. There are asparagine residues at positions 1, 7, 24, and 41; glutamine residues at positions 48, 81, and 111; and two carbohydrate moieties linked to asparagine at positions 7 and 24.
The primary structure of human FSH-P reported here has sequences between residues 1 and 20, 28 and 43, and 47 and 108 similar to those reported by Shome and Parlow (Shome, B., and Parlow, A. F. (1974) J. Clin. Endocrinol. Metab. 39, 187-190), but different at positions 44 to 46, 21 to 28, and at the COOH terminus. The alignment of the cysteines in the first 32 residues of the @ subunit of human follicle-stimu- lating hormone (hFSH-P), luteinizing hormone (hLH-P), thyroid-stimulating hormone (hTSH-P), and chorionic gonadotropin (hCG-p) showed that the sequence of the p subunit of human follicle-stimulating hormone between alignment positions 30 and 38 is identical with the other hormones except that trypto- phan occurs at position 33 in human FSH-/3 instead of isoleucine. When compared to bovine thyroid- stimulating hormone-p subunit (bTSH-P) and ovine luteinizing hormone-P subunit (oLH-P), the sequence between positions 28 and 38 is identical in both hFSH-fi and bTSH-@, except that valine occurs at position 33 in bTSH-0 in place of tryptophan in hFSH-0. Human FSH-0, hCG-fi, and oLH-fl contain carbohydrate moieties at locations corresponding to alignment position 13, whereas bovine and human TSH-P and hLH-fi do not. It is noteworthy that the sequence between alignment positions 26 and 29 is Cys-Leu-Thr-Ileu in hFSH-P, hTSH-& and bTSH-P, whereas oLH-fi contains the sequence Cys-Ileu- Thr-Phe, and hCG-@ and hLH-P contain the sequence Cys-Ileu-Thr-Val.
*This work was supported by Grants ROl HD 06543 and R 1 CA Gonadotropins, Bangalore, India, 1973. 13908 from the National Institutes of Health and the Ford Foundation Grant 670.455A. Part of this work was presented at the International
$ Career Scientist Awardee, Health Research Council of the City of New York, Contract I-621.
Sym.posium on Advances in Chemistry, Biology, and Immunology of
993
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from

994 Amino Acid Sequence of FSH-/3
Several investigators have prepared FSH’ in highly purified form (l-6). These preparations have differed in their specific biopotencies, but have shown only minor differences in amino acid composition (7, 8). Similarly to LH (g-13), TSH (14-19) and hCG (20, al), FSH has been shown to consist of a hormone-nonspecific o( subunit and a hormone-specific fl subunit (22-24). The primary structure of the hor’hrone-non-
specific cr subunit has been elucidated (25). That of the hormone-specific /I subunit is described in this paper.
MATERIALS AND METHODS
Amino Acid Sequence of FSH-/3
Human pituitary FSH and its subunits were prepared from three batches of 15,000 glands, as described earlier (25). From a total yield of 500 mg of highly purified and physicochemically homogenous FSH, approximately 120 mg of highly purified fl subunit were obtained. After repurification by gel filtration on a column of Sephadex G-100, the FSH-P was used for amino acid sequence determination. The methods used for the amino acid and NH,- and COOH-terminal residue analyses, for the reduction and S-aminoethylation of FSH-P prior to enzymic digestions with trypsin and thermolysin and chymotrypsin, for the derivatization of certain peptides with SO,PhNCS and sequen- tial degradation of peptides, for the digestion of peptides with proteolytic enzymes, for the determination of amides, and for assign- ment of carbohydrate side chains were the same as described earlier for the FSH-a subunit (25).
NH,-Terminal Analyses
The following experiments were performed to quantitatively deter- mine the NH, terminal amino acid(s) ofhFSH-P.
Digestion with Leucine Aminopeptidase-Aliquots of 200 pg of hFSH-p were digested with 12.5 pg of leucine aminopeptidase in 0.1 M
Tris-HCI buffer at pH 8.6, containing 0.0025 M MgCl,, and incubated f&l, 4. and 19 hours at :X7”. The solution was acidified and the free amino acids were separated from residual protein (26) and identified by amino acid analysis on a Durrum model IX500 ammo acid analyzer.
Dinitrofluorobenzene Method (27)-One hundred nanomoles of hFSH-fl were dissolved in 4 ml of 0.05 N KC1 and adjusted to pH 8 in a pH-stat with 0.05 N KOH. One hundred microliters of a solution of 1.fluoro2,4-dinitrobenzene were added and the mixture was main- tained at pH 8 at 40’ in the dark until the alkali uptake plateaued. The solution was extracted with 3 ml of ether three times to remove excess l-fluoro-2,4-dinitrobenzene. Since the dinitrophenyl-protein was solu- ble, the solution was made to 6 N HCI and hydrolyzed in an evacuated sealed tube for 16 hours at 100”. The hydrolysate was diluted to approximately 1 N HCI with water and extracted five times with 1 ml of ether each time. The ether extracts were combined, evaporated under nitrogen, and analyzed by two-dimensional paper chromatography, according to Levy (28, 29), using the upper phase of toluene/pyri- dine/Zchloroethanol/OJ? N NH,OH in the first dimension and 1.5 M
phosphate buffer, pH 6, in the second dimension.
COOH-Terminal Analysis by Hydrazinoly,sis
A procedure of Reichert et al. (30) modified by Landefeld et al. (31) was used. A sample of 100 nmol of hormone was dried in a vacuum oven at 80” over silica gel for 20 hours, anhydrous hydrazine (0.5 ml) was added. and the tube evacuated and sealed. Hydrazinolysis was allowed to continue for 20 hours at 80”. The hydrazinolysate was dried m a desiccator over concentrated H,SO, and KOH. The sample was dissolved in 3 ml of water and extracted with 1 ml of benzaldehyde for time periods of 0.5. 1.0, and 3.0 hours. The mixtures were shaken constantly on a Vortex mixer and centrifuged. This step was followed by extraction with 2 ml of ethyl acetate followed by 3 ml of ether. The -
‘The abbreviations used are: FSH-@, p subunit of folhcle-stimulat- ing hormone; hCG. human chorionic gonadotropin: Ae-FSH-0, S- aminoethylated FSH-@; hLH-/3, p subunit of human luteinizing hormone; oLH-/J, @ subunit of ovine-luteinizing hormone; bTSH-/3, 0 subunit of bovine thyroid-stimulating hormone; hTSH-p,fi subunit of human thyroid-stimulating hormone; SO,PhNCS 4sulfophenylisothi- ocyanate sodium monohydrate; hGH, human growth hormone; HMG, human menopausal gonadotropin.
remaining ether was removed by heating the water phase at 60” for 15 min. The sample was then lyophilized, redissolved in pH 2.2 citrate buffer, and analyzed on a Durrum amino acid analyzer. An amino acid calibration mixture was subjected to the conditions of hydrazinolysis in order to obtain correction factors.
Separation of Tryptic Peptides’of S-Aminoethylated Ae-FSH-fl
The tryptic digest was applied to a column (1.5 x 250 cm) of Sephadex G-50 (superfine) and equilibrated in 0.1 M ammonium bicarbonate containing 5% propanol (32). The column was developed with 0.1 M ammonium bicarbonate and a flow rate of 18 ml/hour was maintained. The eluate was monitored at 280 nM and 206 nM (Uvicord III, LKB Instruments, Stockholm, Sweden) to locate peptides. Ali- quots of 5 ml were collected in a refrigerated fraction collector and pooled as shown in Fig. 1. Fraction I consisted of a small amount of a large molecular weight material. Fraction II consisted of undigested material. Fractions III and IV contained glycopeptides, and Fractions V to VIII contained peptides. Fractions III to VIII were further purified by two-dimensional high voltage electrophoresis at pH 6.0, and/or pH 3.5, and/or pH 2.0 as described earlier (25).
Separation of Thermolytic Peptides of FSH-@
The thermolytic digest was applied to a column (1.5 x 250 cm) of Sephadex G-50 (superfine) and equilibrated in 0.1 M ammonium bicarbonate containing 5% propanol. The column was eluted with the same solvent as described earlier for the tryptic peptides. Fractions were pooled as shown in Fig. 2. Fraction I consisted of undigested material. Fraction II containing a mixture of glycopeptides was further purified sequentially by gel filtration on columns (1 x 100 cm) of Sephadex G-25 and G-15 and on a column (1 x 30 cm) of Bio-Gel P2 (Bio-Rad Laboratories, Richmond, Calif.). All gel filtration columns were eluted with 0.1 M ammonium bicarbonate buffer containing 5% propanol. Two glycopeptides were isolated by two-dimensional high voltage electrophoresis at pH 3.5 and pH 6.0.
Fractions III and IV and V were individually gel-filtered on columns (1 x 100 cm) of Sephadex G-25 and G-15 followed by Bio-Gel P2. The fractions containing peptides recovered from gel filtration columns were further purified by two-dimensional high voltage electrophoresis on Whatman No. 1 paper at pH 2.0, and/or 3.5, and/or pH 6.0 as described earlier.
Separation of Chymotryptic Peptides of FSH-8
The column (1.5 x 250 cm) of Sephadex G-50 (superfine) was developed as described for the tryptic and thermolytic peptides. Fractions were pooled as shown in Fig. 3. Fractions II to V were purified by high voltage electrophoresis at pH 6.0. and/or 3.5. and/or 2.0 on Whatman No. 1 paper as described earlier. The purity of tryptic, thermolytic, and chymotryptic peptides was established by electropho- retie homogeneity at pH 6, 3.5, and 2 and confirmed by the integral ratio of residues obtained by amino acid analysis.
Examination of FSH-/3 for Tryptophan
Evidence for the apparent presence of tryptophan in FSH has been obtained previously (1, 33) by spectrophotometric methods. To estab- lish unambiguously the presence of tryptophan in FSH-@ the following direct tests were performed.
FIG. 1. Gel filtration of a tryptic digest of human pituitary Ae-FSH- @ on a column (1.5 x 250 cm) of Sephadex G-50 (superfine).
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Amino Acid Sequence of FSH-fl 995
Staining of Glycopeptides with p-Dimethyl Aminobenzaldehyde
3 0.8-
E
: Oh- A P
2 ; 0.4-
:: 4
(39)-Five to ten nanomoles of glycopeptide T-6 were spotted on a Whatman No. 1 filter paper and sprayed with 1% (w/v) methionine and dried. The paper was then sprayed with a freshly made solution of p-dimethyl aminobenzaldehyde in HCl-acetone (1 g of p-dimethyl aminobenzaldehyde, 90 ml of acetone, and 10 ml of concentrated HCl). The color developed was compared with tryptophan spotted as control.
6il 7b 8b 9b lb0 ii0 li0
Fraction number
FIG. 2. Gel filtration of a thermolysin digest of human pituitary Ae-FSH-P on a column (1.5 x 250 cm) of Sephadex G-50 (superfine).
Amino Acid Analysis of FSH-@ and Glycopeptides with Methane Sulfonic Acid (IO)-Samples containing 2 to 5 nmol of tryptophan were hydrolyzed overnight with 25 al of 4 N methane sulfonic acid (Cata- logue No. 25600, Pierce Chemical Co., Rockford, Ill.) under vacuum. The samples were neutralized with 25 ~1 of 3.5 N NaOH and analyzed on a Durrum amino acid analyzer. The effect of hFSH-p on the recovery of tryptophan from methane sulfonic acid hydrolysates was examined by hydrolysis of 10 nmol of tryptophan and 3 nmol of bovine serum albumin individually with and without the addition of 8 nmol of hFSH-p followed by amino acid analysis.
Microbiological Assay for Tlyptophan-The procedure recom- mended by Difco Laboratories (Detroit, Mich.) was used for the assay, using Lactobacillusplantarum ATCC 8014, with slight modifications.
I GEL FILTRATION : SEPHADEX G-50 SUPERFINE
1.0 4’J ll
Inoculum for assay was prepared by subculturing L. plantarum ATCC 8014, from a stock culture, into a tube containing 10 ml of Bacto-Lactobacilli broth AOAC. After 24 hours of incubation at 37”, the cells were centrifuged under aseptic conditions, and the superna- tant liquid decanted. The cells were resuspended in 10 ml of sterile 0.85% sodium chloride. The cell suspension was then diluted 1:lOO with sterile 0.85% sodium chloride. One drop of this latter suspension was then used to inoculate each of the assay tubes.
Five- to twenty-five nanomole aliquots of hFSH, hFSH-8, hLH, bovine serum albumin hGH, and ribonuclease were digested in lo-ml culture tubes with caps, with 5 yg each of thermolysin and protease in 200 al of 0.1 M ammonium bicarbonate buffer at pH 8.0 for 72 hours at 37” prior to assay. On the day of the assay, 1 ml of distilled water and 1 ml of freshly prepared Bacto-Tryptophan assay medium (Code 0327, Difco Laboratories, Detroit, Mich.) were added. The tubes were sterilized by autoclaving for 10 min at 15 pounds pressure. After the tubes had cooled, they wereinoculated with the L. plantarum and then incubated at 37” for 72 hours. At the end of the incubation period, the turbidity was determined by reading in a Beckman spectrophotometer at 540 nm and the acid liberated was measured by titration against 0.04 N NaOH. Suitable controls and solutions of tryptophan containing 0.5 to 2 agltube were also assayed simultaneously.
FIG. 3. Gel filtration of a chymotryptic digest of human pituitary Ae-FSH-Don a column (1.5 x 250 cm) of Sephadex G-50 (superfine). RESULTS AND DISCUSSION
Fluorescence Spectra-The fluorescence spectrum of FSH-P was determined as described by Henning et al. (34) in a Hitachi model MPF-3 spectrophotofluorimeter. Aliquots of 0.1 amol of ribonuclease and FSH-fl were subjected to spectral analysis, whereas the reference enzymes were analyzed at concentrations equivalent to 0.1 amol of tryptophanyl residue. Five to twenty nanomoles of glycopeptides T-6 and C-4 were also subjected to spectral analysis. The samples were dissolved in 0.01 M potassium phosphate buffer, pH 7.3, containing 0.1 M sodium chloride (35). Excitation maxima of 290, 290, 280, 282, 282, and 282 were used for chymotrypsin, bovine serum albumin, ribonu- clease, FSH-P, glycopeptide T-6, and glycopeptide C-4, respectively.
Titration with N-Bromosuccinimide-The reaction of N-bromosuc- cinimide with FSH-@ was carried out by adding a 0.002 M aqueous solution of N-bromosuccinimide to the FSH-fl in 0.1 M sodium acetate buffer, pH 4.0 (34). The correction factor and extinction coefficient of Patchornik et al. (36) were used in the calculation of the values given in Table II. An aliquot of N-bromosuccinimide-treated FSH was also assayed for biological activity, with suitable controls.
Calorimetric Method-The tryptophan content of FSH-/3, of FSH, and of reference proteins in Table III was estimated calorimetrically by the method of Spies and Chambers (37) as modified by Opienska- Blauth et al. (38). Two miniliters of a solution of acetic acid containing 56 mg of Fes+/liter (0.27 g of FeCl,.GH,O dissolved in 0.5 ml of water and made to 1 liter with glacial acetic acid) were mixed with samples in 1 ml of water. Two milliliters of concentrated sulfuric acid were added and the solution was shaken vigorously and allowed to stand for 15 min. The extinction at 545 nM was measured and the tryptophan content was calculated from a standard curve of tryptophan similarly treated.
Amino Acid Analysis-Highly purified human FSH contain- ing 4000 to 14000 i.u. of 2nd IRP-HMG/mg has been prepared (l-6); however, FSH used by Shome and Parlow (41, 42) for sequence studies contained 1800 i.u./mg. The FSH used in the present studies contained 9765 h 291 i.u./mg (25). The amino acid composition of FSH-8 (Table I) obtained from the analysis of 24-, 48-, and 72-hour acid hydrolysates is compati- ble with the amino acid sequence presented in Fig. 5. The amino acid composition showed a few differences in proline, threonine, cysteine, and tyrosine from that of a 24-hour hydrolysate of an earlier preparation of FSH-P (43) which was not purified by gel filtration on Sephadex G-100. These differences were resolved by the performance of amino acid analysis with 24-, 48-, and 72-hour hydrolysates of FSH-P purified by gel filtration on Sephadex G-100. The amino acid analysis ‘of FSH-@ is comparable with that reported by Reichert and Ward (23), except for minor differences in threonine, proline, glycine, and isoleucine. The amino acid analysis of FSH-8, as reported by Shome and Parlow (44), differs in having slightly higher arginine, glycine, and tyrosine. The amino acid residues calculated from the first proposal of the sequence of human FSH-/3 by Shome and Parlow (41), however, has 1 additional residue of aspartic acid, threonine, glutamic acid, and isoleucine and 1 residue less of proline, alanine, tyrosine, and phenylalanine and 2 residues less of
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from

996 Amino Acid Sequence of FSH-/3
TABLE I Amino acid analysis of FSH-8 from human pituitary glands
Amino acid 24 hr
Acid hydrolysis for
48 hr 72 hr
Average or extrapolated to zero time (69)
Residues/13,297 g” Residues from of protein sequence data
mollmol ofprotein” Tryptophan 1’ Aspartic acid 9.5 9.6 9.3 9.4 9 Threonine 11.8 10.0 9.6 12.7 13 Serine 5.7 5.2 4.2 6.5 6-7
Glutamine 10.9 10.4 10.7 10.6 10 Proline 5.1 6.1 6.0 5.7 6
Glycine 5.7 5.5 6.0 5.7 6
Alanine 6.2 6.6 6.8 6.5 6-7
Half-cystined 11.0 11.0 11.5 11.1 11
Valine 6.8 7.2 7.7 7.1 7-8
Methionine 1.1 1.7 1.0 1.7 l-2
Isoleucine 5.1 5.1 4.6 4.9 5
Leucine 6.1 5.9 5.2 6.6 6-7
Tyrosine 5.7 3.3 4.0 6.0 6
Phenylalanine 3.5 3.3 3.7 3.5 3-4
Histidine 2.4 3.4 3.7 3.5 3
Lysine 7.0 7.1 7.4 6.8 7
Arginine 4.5 4.6 4.4 4.5 5
a Calculated molecular weight for protein moiety from sequence data, b Calculated on the basis of total micrograms of residues recovered using a molecular weight of 13,297. c See text for methods of determination. d Also determined as Cys(Ae) in an aliquot of Ae-FSH-8.
1 9
13 I
10 6
6
7
12 6 1 5 7
8
3 3
7
5 13,297 g
leucine as compared to the amino acid residues presented in Fig. 5 and Table I.
The presence of tryptophan in human FSH has been indicated (1, 42, 45) by ultraviolet absorption methods of Beaven and Holiday (46) and Goodwin and Morton (47). In present studies also, FSH-@ and glycopeptides T-6 and C-4 (Fig. 4) exhibited a spectrum similar to tryptophan-containing proteins like bovine serum albumin and chymotrypsin and unlike that of ribonuclease, a tryptophanless protein. Shome and Parlow (41), in their first proposal of the amino acid sequence of FSH-8, have shown the presence of tryptophan (44). Also, during the sequence analysis of human FSH-/I in the present studies, tryptophan was recovered, but in submolar quantities. Difficulties have been encountered in the quantita- tion of tryptophan by the spectrophotometric method, espe- cially when tryptophan was present in the protein at a low level and the tyrosine content was high (48). Similarly, side reac- tions of sialic acid (49) or the hexosamines (50) with Ehrlich’s reagent (39) have also been shown to occur. For example, the A protein component of tryptophan synthetase of Escherichia coli was considered to contain tryptophan until the absence of this amino acid was proven by titration with N-bromosuccini- mide, amino acid analysis of alkaline hydrolysate, and mi- crobiological assay (34). Hence, further experiments were performed in order to unambiguously establish the presence of tryptophan in human FSH. Glycopeptide T-6 (Fig. 5) gave a positive staining on paper with Ehrlich’s reagent, suggesting the presence of tryptophan. In Table II are presented the results of titration against N-bromosuccinimide. The values obtained for bovine serum albumin, ribonuclease, and chymo- trypsin compare well with values reported in the literature. A value of 0.45 mol of tryptophan was obtained per mol of hFSH-8. The results of the calorimetric determination also indicated the presence of tryptophan in FSH and FSH-8, and glycopeptide C-4, in submolar quantities (Table III). The
~- BSA -------- RNfiE
o--o--o CHYMOTRYPSIN
-. \
ii \
C--.-S-. hFSH+
I \
60
t
___---_ Bs‘,
- - - GLYCOPEPTIDE T- 6
C-O--. GLYCOPEPTIM C-4
I I I I I I I 300 320 340 360 360 400
WAVE LENGTH (nm)
FIG. 4. Fluorescence spectra. The proteins were dissolved in 2.5 ml of 0.01 M potassium phosphate buffer, pH 7.3, and 0.1 M sodium chloride. Bovine serum albumin (MA), 0.05 rmol (2 mol of trypto- phan/mol of protein (61)); chymotrypsin, 0.01 pmol (10 mol of tryptophan/mol of protein (62)); ribonuclease (RNuse), 0.1 flmol (no tryptophan (63)); FSH-j3, 0.1 pmol; glycopeptides T-6and C-4, 10 no.
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Amino Acid Sequence of FSH-$ 997
SEQUENCE OF FSH-$
10 Asn - Ser - Cys - Glu - Leu - Thr - Asn(CH0) - Ileu - Thr - Ileu - Ala - Ileu - Glu - Lys - Glu - Glu - Cys- -Th-l- - Th-2 - -Th-3 - --h-4- i Th-5
- Th-la-+ - Th-Za l - Th-4a - T-l- < T-2 * -T-3- - C-l : + c-2 ------------------------------
+ CZTh-l- +CZTh-2- + CZTh-3 + 4 CZTh-4
20 30 Arg - Phe - Cys - Leu - Thr - Ileu - Asn(CH0) - Thr - Thr - Trp - Cys - Ala - Gly - Tyr - Cys - Tyr - Thr - - i---h-6--+ < Th-7 b c-- Th-8- + - Th-9- - Th-10 - - c'l'h-Ya-
tT-4-t -T-5--+ < T-6 + - T-T..=- - -c-3---+ + c-4 C-5- -C-6- -
----+cCZThZ+
40 50 Arg - Asp - Leu - Val - Tyr - Lys - Asn - Pro - Ala - Arg - Pro - Lys - Ileu - Gln - Lys - Thr - Cys - Thr -
Th-11 Th-12 + Th-9b-
- +---------T-9 + -T-10 - +-T-11---+ - T-12 - -T-13--~ - c-7- 4 C-8 --------______-____-_----------~
x- CBTh-l- C8Th-2 +
60 70 Phe - Lys - Glu - Leu - Val - Tyr - Glu - Thr - Val - Arg - Val - Pro - Gly - Cys - Ala - His - His- Ala - --h-13-----+ - Th-14 b --h-15 l - Th-16
- -
T-14- < T-15 k -T-16-u T-17 +-T17Cl*T17C2::
c-9 c-10 * cc-11 ;; +CSTh3+
80 Asp - Ser - Leu - Tyr - Thr - Tyr - Pro - Val - Ala - Thr - Gln - Cys - His - Cys - Gly - Lys - Cys - Asp -
Th-17 Th-18 Th-16a- c-- Th-17ap c-Th-18a---t +Th-18W - Th-18T-l- Th-16b- +Th-17d- +Th-17b-
+Thl7c+ tThl7c-t ---------------------------------------- CT-18~ +--T-19---+ +TZW c--
T-17C3 T-17C4 : : T17C5+ c-12 c-13 : c-14- -c-15
+C15Tl++C15T2---HC15T3++-----
90 100 Ser - Asp - Ser - Thr - Asp - Cys - Thr - Val - Arg - Gly - Leu - Gly - Pro - Ser - Tyr - Cys - Ser - Phe -
+ -Th-19- i Th-20 3 <
T-21 F -T-22- < T-23 f T-23a + 4
---__--_-_______________________________----- C-16-------t -c-17- C15T4 b - C15T5-C15T6+
110 118 Gly - Glu - Met - Lys - Gln - Tyr - Pro - Thr - Ala - Leu - Ser - Tyr Th-21 + Th-2la- Th2lb-t 1 T-24 T-25 l
T-24a- +- C-18-
FIG. 5. Amino acid sequence of human pituitary FSH-6 subunit
quantities of tryptophan obtained for reference proteins were treatment of FSH with N-bromosuccinimide resulted in a comparable to those reported in the literature. The glycopep- complete loss of biological activity.
tides C-2 and Th-7 did not yield a positive reaction in the Since hydrolysis in 5.7 N HCl causes complete destruction of above test for tryptophan. It is interesting to note that the tryptophan, hFSH, hFSH-/?, glycopeptides C-2, C-4, Th-7, and
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Amino Acid Sequence of FSH-@ 998
TABLE II TABLE IV
Titration of tryptophan with N-bromosuccinimide Determination of tryptophan after hydrolysis of sample with 4 N
methane sulfonic acid” Maximal Trvotoohanlmol
Protein” decrease in Tryptophan _ ;f protein absorbance 1” SamDIe
(280 nm) Found Reported0
pm01 mol
Bovine serum albumin 0.15 0.108 2.1 2 (66) (0.05 /.Lmol)
RNAse (0.1 rmol) 0 0 0 0 (‘38) Chymotrypsin (0.05 0.67 0.486 9.5 10 (67)
rmo1) hFSH-@ (0.04 rmol) 0.03 0.018 0.45
u Molar concentration of protein was based on the following molecu-
lar weights: bovine serum albumin, 65,000 (66); ribonuclease, 14,800 (68); FSH-/3, 13,000; and chymotrypsin, 25,000 (67).
b The figures in parentheses are references
TABLE III
Calorimetric determination of tryptophan
Sample” Tryptophan content
Found Reported
Bovine serum albumin
Chymotrypsin RNASE
FSH FSH-/3
Glycopeptide C-2
Glycopeptide C-4 Glycopeptide Th-7
HCG- ab
HCG”
nmol/nmolprotein
1.9 2 (66)
10.8 10 (67)
0 0 (68)
0.5 0.4
0
0.2 0
0
0
“Molecular weight of 65,000 (66), 25,000 (67), 24,000, and 13,000
were used for bovine serum albumin, chymotrypsin, FSH, and FSH-/3, respectively.
‘Gift from Dr. 0. P. Bahl (State University of New York at Buffalo).
T-6, and peptides shown in Table IV, as well as equine FSH and FSH-P,’ were also subjected to hydrolysis with 4 N
methane sulfonic acid containing 0.2% tryptamine, to protect the tryptophan from destruction. Both human and equine FSH and FSH-/3 showed the presence of tryptophan; however, the yield for the glycopeptides C-4 and T-6 was in submolar quantities (Table IV). The glycopeptides C-2 and Th-7 were devoid of tryptophan. The values for the reference proteins, however. were in accordance with those reported in the litera- ture. It appears that the increase in the carbohydrate content of the glycopeptides C-4 and T-6, as compared to the subunit, causes greater destruction of tryptophan (40). This was further
indicated by a 43% and 32% loss of tryptophan when 10 nmol of tryptophan and 3 nmol of bovine serum albumin were hydro- lyzed separately with 8 nmol of hFSH-P. As shown by Inglis and Liu (51), the interaction of the adjacent cysteine residue may also contribute to the destruction of tryptophan.
The results of the microbiological assay for tryptophan (52) are given in Table V. FSH and FSH-fi and the glycopeptides C-4 and T-6 indicated the presence of tryptophan and were unlike the tryptophanless protein ribonuclease. These results
indicated the presence of tryptophan in glycopeptides C-4 and T-6. The yield of tryptophan from glycopeptides in submolar quantities and lower recoveries during sequence studies sug- gested significant destruction of tryptophan during experimen-
Tryptophan content Sampleb
Found Reportedc
nmollnmol protein
Bovine serum albumin 2.0 (3)d 2 (66) Chymotrypsin 9.3 (3) 10 (67) RNASE 0 0 (68) FSH 0.3 (3)
FSH-/3 0.2 (3)
Glycopeptide C-2 0 Glycopeptide C-4 0.1
Glycopeptide Th-7 0
Glycopeptide T-6 0.1
Peptides’ 0
Equine FSH 0.6 Equine FSH-@ 0.5
‘Containing 0.2% 3.(2.aminoethyl)indole.
bMolecular weights of 65,000 (66), 25,000 (67), 24,000, and 13,000
were used for bovine serum albumin, chymotrypsin, FSH, and FSH-(3, respectively.
‘The figures in parentheses are references. d Mean (number of determinations).
‘Peptides T-l, T-5, T-9, T-10, T-11, T-19, T-20, T-21, T-22, T-25,
Th-2, Th-3, Th-4, Th-5, Th-7, Th-9, Th-Sa, Th-10, Th-12, Th-13, Th-14, Th-15, Th-16, Th-16a, Th-17, Th-18b, Th-19, Th-20, Th-2la,
Th-21, and C-3.
TABLE V
Microbiological assay for tryptophan (48)
Fraction (nmol of protein used
for assay)
Turbidometric determination”
Resldueslmol reported in literature
Residues”lmol
Bovine serum albumin (6.7 nmol) 2 2
Ribonuclease (13 nmol) 0 0 hFSH (13 nmol) 0.6 hFSH-/3 (15 nmol) 0.3
CM-hFSH-P (15 nmol) 0.4
Glycopeptide C-4 (20 nmol) 0.2 Glycopeptide T-6 (20 nmol) 0.2
“Data were corrected for controls using thermolysin and protease alone without protein or peptide samples.
‘Values corrected for recovery, assuming 2 residues of tryptopham
mol of bovine serum albumin.
tal procedures. The presence of tryptophan in the amino acid analysis of equine FSH and FSH-P, hydrolyzed in 4 N methane sulfonic acid containing 0.2% tryptamine, is noteworthy.
The total amino acid residues from the amino acid analysis and sequence studies of FSH-a (25) and of FSH-fi are compared with the total amino acid residues obtained from 24-, 48-, and 72.hour hydrolyses of human pituitary FSH. As shown in Table VI, the total residues from (Y and 0 are
comparable to those obtained from intact human pituitary FSH.
Amino Acid Sequences-The amino acid composition and
3The quantitative data on the amino acid compositions, yields, and sequences of the peptides and the supporting analytical data in Table VIII and Appendices A, B, and C are published as a miniprint supplement immediately following this paper. For the convenience of those who prefer to obtain supplementary material in the form of 23 pages of full size photocopies, these same data are available as Journal of Biological Chemistry 75M-851. Orders for supplementary material should specify the title. authors, and reference to this paper and the
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from
Amino Acid Sequence of FSH-/3 999
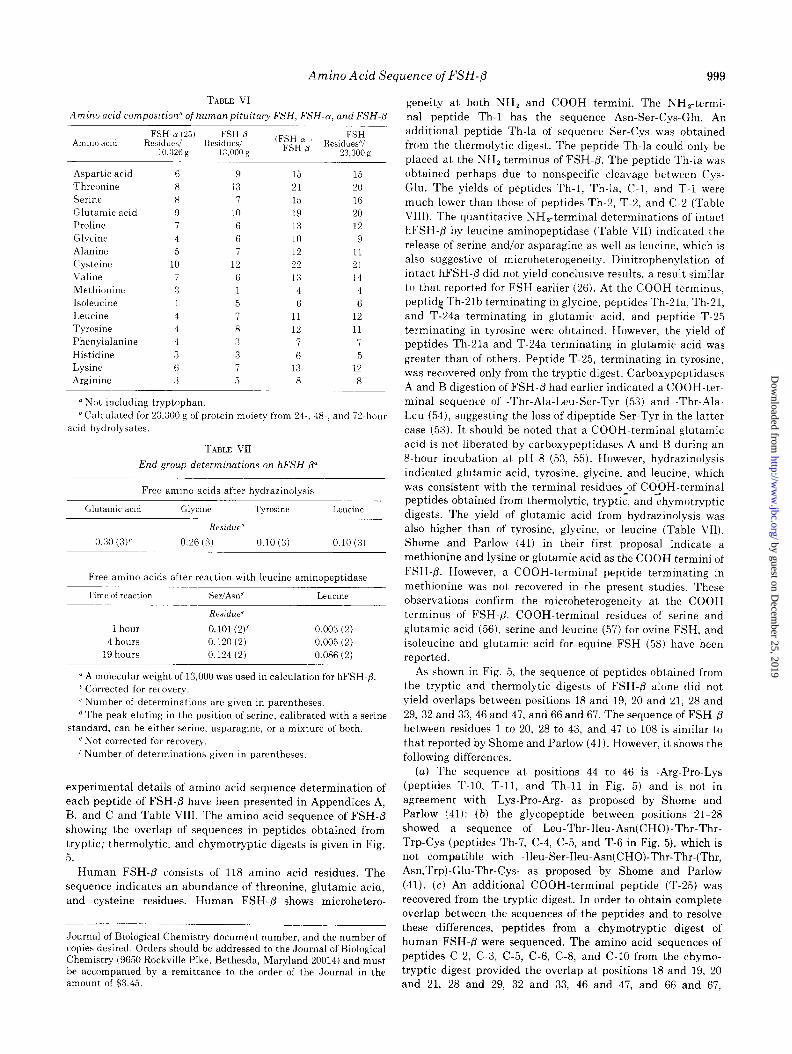
TABLE VI
Amino acid composition” of human pituitary FSH, FSH-a, and FSH-8
Amino acid FSH-a (25) FSH-8 (FSH~a + FSH
Residues/ Residues/ FSH-p Residues”/ 10.326 g 13.000 g 23.300 g
Aspartic acid
Threonine
Swine
Glutamic acid
Proline Glycine
Alanine
Cysteine
Valine
Methionine Isoleucine
Leucine
Tyrosine
Phenylalanine
Histidine Lysine
Arginine
4
5
10
3 1
4
4
4
0 6
3
9 13
10
6
6
1%
6
5
15 15
21 20
15 16
19 20
13 12
10 9
12 11 22 21
13 14
4 4
6 6
11 12 12 11
7 7
6 5
13 12
8 8
“Not including tryptophan.
‘Calculated for 23,300 g of protein moiety from 24-, 48., and 72 acid hydrolysates.
TABLE VII
End group determinations on hFSH-@
-hour
Free amino acids after hydrazinolysis
Glutamic acid Glycine Tyrosine Leucine
Residue”
0.26 (3, 0.10 (3) 0.10 (3)
Free amino acids after reaction with leucine aminopeptidase
Time ofreactirm SeIfASlI” Leucine
1 hour
4 hours 19 hours
Residue’
O.lOl(2)’ 0.120 (2)
0.124 (2)
0.003 (2)
0.005 (2)
0.086 (2)
0 A molecular weight of 13,000 was used in calculation for hFSH-8.
* Corrected for recovery.
‘Number of determinations are given in parentheses.
d The peak eluting in the position of serine, calibrated with a serine
standard, can be either serine, asparagine, or a mixture of both. e Not corrected for recovery.
’ Number of determinations given in parentheses.
experimental details of amino acid sequence determination of
each peptide of FSH-@ have been presented in Appendices A, B. and C and Table VIII. The amino acid sequence of FSH-/3 showing the overlap of sequences in peptides obtained from tryptic: thermolytic, and chymotryptic digests is given in Fig.
5. Human FSH-/3 consists of 118 amino acid residues. The
sequence indicates an abundance of threonine, glutamic acid, and cysteine residues. Human FSH-P shows microhetero-
Journal of Biological Chemistry document number, and the number of copies desired. Orders should be addressed to the Journal of Biological Chemistry (9650 Rockville Pike, Bethesda, Maryland 20014) and must be accompanied by a remittance to the order of the Journal in the amount of $3.45.
geneity at both NH, and COOH termini. The NH,-termi-
nal peptide Th-1 has the sequence Asn-Ser-Cys-Glu. An additional peptide Th-la of sequence Ser-Cys was obtained from the thermolytic digest. The peptide Th-la could only be placed at the NH, terminus of FSH-@. The peptide Th-la was obtained perhaps due to nonspecific cleavage between Cys- Glu. The yields of peptides Th-1, Th-la, C-l, and T-l were much lower than those of peptides Th-2, T-2, and C-2 (Table VIII). The quantitative NH,-terminal determinations of intact hFSH-P by leucine aminopeptidase (Table VII) indicated the
release of serine and/or asparagine as well as leucine, which is also suggestive of microheterogeneity. Dinitrophenylation of intact hFSH-/3 did not yield conclusive results, a result similar to that reported for FSH earlier (26). At the COOH terminus, peptid2 Th-Plb terminating in glycine, peptides Th-2la, Th-21, and T-24a terminating in glutamic acid, and peptide T-25 terminating in tyrosine were obtained. However, the yield of peptides Th-2la and T-24a terminating in glutamic acid was greater than of others. Peptide T-25, terminating in tyrosine, was recovered only from the tryptic digest. Carboxypeptidases A and B digestion of FSH-P had earlier indicated a COOH-ter- minal sequence of -Thr-Ala-Leu-Ser-Tyr (53) and -Thr-Ala- Leu (54), suggesting the loss of dipeptide Ser-Tyr in the latter
case (53). It should be noted that a COOH-terminal glutamic acid is not liberated by carboxypeptidases A and B during an 8-hour incubation at pH 8 (53, 55). However, hydrazinolysis indicated glutamic acid, tyrosine, glycine, and leucine, which was consistent with the terminal residues of COOH-terminal peptides obtained from thermolytic, tryptic, andchymotryptic digests. The yield of glutamic acid from hydrazinolysis was also higher than of tyrosine, glycine, or leucine (Table VII). Shome and Parlow (41) in their first proposal indicate a methionine and lysine or glutamic acid as the COOH termini of
FSH-P. However, a COOH-terminal peptide terminating in methionine was not recovered in the present studies. These observations confirm the microheterogeneity at the COOH terminus of FSH-/3. COOH-terminal residues of serine and glutamic acid (56), serine and leucine (57) for ovine FSH, and isoleucine and glutamic acid for equine FSH (58) have been reported.
As shown in Fig. 5, the sequence of peptides obtained from the tryptic and thermolytic digests of FSH-@ alone did not yield overlaps between positions 18 and 19, 20 and 21, 28 and
29, 32 and 33,46 and 47, and 66 and 67. The sequence of FSH-fi between residues 1 to 20, 28 to 43, and 47 to 108 is similar to that reported by Shome and Parlow (41). However, it shows the following differences.
(a) The sequence at positions 44 to 46 is -Arg-Pro-Lys (peptides T-10, T-11, and Th-11 in Fig. 5) and is not in agreement with -Lys-Pro-Arg- as proposed by Shome and Parlow (41): (b) the glycopeptide between positions 21-28 showed a sequence of Leu-Thr-Ileu-Asn(CHO)-Thr-Thr-
Trp-Cys (peptides Th-7, C-4, C-5, and T-6 in Fig. 5), which is not compatible with -Ileu-Ser-Ileu-Asn(CHO)-Thr-Thr-(Thr, Asn,Trp)-Glu-Thr-Cys- as proposed by Shome and Parlow
(41). (c) An additional COOH-terminal peptide (T-25) was recovered from the tryptic digest. In order to obtain complete overlap between the sequences of the peptides and to resolve these differences, peptides from a chymotryptic digest of human FSH-@ were sequenced. The amino acid sequences of peptides C-2, C-3, C-5, C-6, C-8, and C-10 from the chymo- tryptic digest provided the overlap at positions 18 and 19, 20 and 21, 28 and 29, 32 and 33, 46 and 47, and 66 and 67,
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from

1000
hFSH-P
hTSH-P
hLH -p
KG -p
Alijrnment Position
hFSH-P
h TSH-P
hLH -p
hCG -p
Alignment Posit.ion
hFSH-P
h TSH-P
hLH -p
hCG -p
Alignment Position
Amino Acid Sequence of FSH-P
Asn - Ser - Cys - Glu - Leu-Thr -Asn(CHO) - Ileu
Fhe -Cys -Ileu- Pro - Thr -Glx - Tyr
Ser - Arg - Glu - F’ro - Leu -Arg - Pro - Trp - Cys -His - Pro - Ileu - Asn - Ala
Ser - Lys - Gln - Pro - Leu -Arg - Pro - Arg - Cys- Arg - Pro - Ileu -Asn(CHO)- Ala - .- . __-- -.--_~- ~--- __-__I_
5 10
10 20
_ Thr - Ileu - Ala - Ileu - Glu - Lys - Glu - Glu - Cys - Arg - Phe - Cys - Leu - Thr - Ileu
-(Met , Thr , His , Val , Glx)-Arg -Arg - Glx - Cys -Ala - Tyr - Cys - Leu - Thr -Ileu
- Ileu- Leu - Ala - Val - Gln -Lys -Glu - Gly- Cys -Fro - Val -Cys - Ileu - Thr-Val
- Thr - Leu - Ala - Val - Glu - Lys - Glu- Gly - Cys - Pro - Val -Cys - Ileu - Thr- Val
r5 2’0 -___---.-
2’5
30
-Asn(CHO) - Thr - Thr - Trp - Cys -Ala - Gly - Tyr - Cys -
Asn (CHO) - Thr - Thr - Ileu - Cys -Ala - Gly - Tyr - Cys -
-Asn (CHO) - Thr - Thr - Ileu - Cys - Ala - Gly - Tyr - Cys -
-Asn(CHO) - Thr - Thr -1leu - Cys -Ala - Gly - Tyr - Cys -
-5
-- ---
35
FIG. 6. Comparison between the NH,-terminal sequence.i of’hFSH-P. hLH-0, hTSH-p, and hCG-p.
respectively, and confirmed the sequence at (a) positions 44 to 46 to be -Arg-Pro-Lys- (peptides C-8 and C-8Th-1); and (b) positions 21 to 28 (peptides C-4 and C-5). The sequences of peptides between positions 1 to 20 and 28 to 118 show complete overlap (Fig. 5) and do not contain tryptophan (Table IV). The yield of tryptophan during the sequence determination of peptides C-4 and T-6 was low. However, the presence of
cysteine and tryptophan as the COOH-terminal residues of peptides T-6 and C-4, respectively, and the establishment of the amino acid sequences from the NH, terminus of T-6 and C-4 permitted the location of tryptophan at position 27.
It should be pointed out that the cleavage of Arg-Pro at positions 44 and 45 by trypsin (peptides T-10 and T-11) was atypical. Similarly the peptide Tyr-Cys, recovered in a low yield from the tryptic digest (Table VIII), 3 may have arisen by atypical cleavage at either positions 30 and 31 (peptide T-7a) or positions 103 and 104 (peptide T-23a).
Alignment of the cysteine residues in the NH,-terminal portion of hFSH-fi with those of hCG-/3 (59), hTSH-/3 (17), and hLH-/3 (60), as shown in Fig. 6, yields homology up to residue
32 of hFSH-P; however, the sequence -Ileu-Ser-Ileu-Asn(CHO)- Thr-Thr-(Thr,Asn,Trp)-Glu-Thr-Cys-Ala-Gly-Tyr-Cys-, as proposed by Shome and Parlow (41), can only be aligned if “gaps” are introduced in the sequences of the other hormones. The alignment positions are numbered as suggested by Bewley and Li (61). It is interesting to note that the sequence of hFSH-fi between alignment positions 30 and 38 is identical with the other hormones with the sole exception of tryptophan at position 33 of hFSH-P instead of isoleucine in the other hormones. It should be pointed out, however, that tryptophan
may be an acceptable substitute for isoleucine (54, 61), and, therefore, hFSH-0, hCG-8, hLH-P, and hTSH-@ may be con- sidered homologous in the NH,-terminal region (up to residue 32 of hFSH-0) (Fig. 6). This homology is not surprising in view of the recombination capacities of the different /3 subunits with structurally similar 01 subunits.
When compared to bovine TSH-fl and ovine LH-@ (62-64)
(Fig. 7), the sequence between alignment positions 28 and 38 is identical in both hFSH$ and bTSH-p except at alignment position 33, where bovine TSH-/3 contains valine, whereas hFSH-fl has tryptophan. These results emphasize the essential
nature of tryptophan at position 27 of hFSH-P. As also shown in Figs. 6 and 7, hFSH-P, hCG-/3, and oLH-@ contain carbohy- drate moieties at positions corresponding to alignment position 13, whereas bovine and human TSH-fi and hLH-p do not.
Similarly, a carbohydrate moiety is present at alignment position 30 of hFSH-fl and at a similar sequence in human and bovine TSH-jY, in hLH-fi, and in hCG-P but not in oLH-P. The sequence of amino acids around both asparagine-linked carbohydrate chains in hFSH-fl is Asn-X-Thr, which is consist- ent with the amino acid sequence associated with carbohydrate units of this type in other glycoproteins (65). It is interesting to note that the sequence between alignment positions 26 and 29 (Figs. 6 and 7) is Cys-Leu-Thr-Ileu in hFSH-8, hTSH-@, and bTSH-8, whereas oLH-P contains the sequence Cys-Ileu- Thr-Phe, and hCG-@ and hLH-P contain the sequence Cys- Ileu-Thr-Val.
Acknowledgments-We are grateful to the Endocrine Study Section of the National Institutes of Health, Bethesda, Md.
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Amino Acid Sequence of FSH-/3 1001
hFSH-B Asn - Ser - Cys - Glu -
oLH -B Ser - Arg - Glu - Pro - Leu - Arg - Pro - Leu - Cys - Glu -
bTSH-B Phe - Cys - Ileu-
Alignment Position 5 10
hFSH-f3 - Leu - Thr - Asn(CH0) - Ileu - Thr - I& - Ala - Ileu - Glu - Lys -
oLH -6 - Pro - Ileu- Asn(CH0) - Ala - Thr - Leu - Ala - Ala - Glu - Lys -
bTSH-B - Pro - Thr - Glu - Tyr - Met - Met - His - Val - Glu - Arg -
Alignment -- Position 15 20
hFSH-t? - Glu - Glu - Cys - Arg - Phe - Cys - Leu - Thr - Ileu - Asn(CH0) -
oLH -B - Glu - Ala - Cys - Pro - Val - Cys - Ileu - Thr - Phe - Thr
bTSH-B - Lys - Glu - Cys - Ala - Tyr - Cys - Leu - Thr - Ileu - Asn(CH0) - Alignment Position 25 30
hFSH-fi - Thr - Thr - Trp - Cys - Ala - Gly - Tyr - Cys -
oLH -B - Thr - Ser - Ileu - Cys - Ala - Gly - Tyr - Cys -
bTSH-B - Thr - Thr - Val - Cys - Ala - Gly - Tyr - Cys - Alignment Position 35
FIG. 7. Comparison between the amino acid sequences of hFSH-fl, oLH-& and bTSH-p.
for supplying hormone standards and to the National Pituitary Agency, Baltimore, Md. for supplying human pituitary glands. Thanks are due to Dr. S. Moore (The Rockefeller University, New York, New York) and to Dr. 0. P. Bahl (The State University of New York, Buffalo, New York) for encourage- ment and advice. Thanks are also due to Ms. Inessa Hakkier for devoted and excellent technical assistance and to Dr. A. B. Rifkind (Department of Pediatrics) for assistance with studies using a spectrophotofluorimeter.
REFERENCES
1. Roos, P. (1968) Acta Endocrinol. Suppl. (Copenhagen) 131, 1-93 2. Papkoff, H., Mahlmann, L. J., and Li, C. H. (1967) Biochemistry
6, 3976-3982 3. Peckham, W. D., and Parlow, A. F. (1969) Endocrinology 84,
953-957 4. Saxena, B. B., and Rathnam, P. (1967) J. Biol. Chem. 242,
3769-3775 5. Reichert, L. E., Jr., Kathan, R. H., and Ryan, R. J. (1968)
Endocrinology 82, 109-114 6. Ryan, R. J. (1968) J. Clin. Endocrinol. Metab. 28, 886-896 7. Barker, S. A., Gray, C. J., Kennedy, J. F., and Butt, W. R. (1969)
J. Endocrinol. 45, 275-285 8. Geschwind, J. I. (1969) Gen. Comp. Endocrinol. S2, 179-184 9. Shome, B., and Parlow, A. F. (1973) J. Clin. Endocrinol. Metab.
36, 618-621
10. Maghuin-Rogister, G., and Herman, G. (1972) FEBS Lett. 23, 225-229
11. Closset, J., Hennen, G., and Lequin, R. M. (1973) FEBS Lett. 29,
97-100 12. Sairam, M. R., Papkoff, H., and Li, C. H. (1972) Biochem.
Biophys. Res. Commun. 48, 530-537 13. Rathnam, P., and Saxena, B. B. (1971) J. Biol. Chem. 246,
708777094 14. Ward, D. N., Reichert, L. E., Liu, W. K., Nahm, H. S., and
Lamkin, W. M. (1972) in Gonadotropins (Saxena, B. B., Beling, C. G., and Gandy, H. M., eds) pp. 132-143, John Wiley and Sons, New York
15. Maghuin-Rogister, G., and Hennen, G. (1972) FEBS Lett. 25, 57-60
16. Liao, T.-H., and Pierce, J. G. (1971) J. Biol. Chem. 246, 850-865 17. Shame, B., and Parlow, A. F. Abstract of 55th Annual Endocrine
Society Meeting, 1973, p. 23, The Endocrine Society, Maryland 18. Sairam, M. R., and Li, C. H. (1973) Biochem. Biophys. Res.
Commun. 51, 336-342 19 Sairam, M. R., and Li, C. H. (1973) Biochem. Biophys. Res.
Commun. 54, 426-431 20. Swaminathan, N., and Bahl, 0. P. (1970) Biochem. Biophys. Res.
Commun. 40, 422-427 21. Morgan, F. J., and Canfield, R. E. (1971) Endocrinology 88,
1045-1053 22. Saxena, B. B., and Rathnam, P. (1971) J. Biol. Chem. 246,
3549-3554 23. Reichert, L. E., Jr., and Ward, D. N. (1974) Endocrinology 94,
655-664 24. Ryan, R. J., Jiang, N. S., and Hanlon, M. S. (1969) in 51st Meeting
of Endocrine Society, Abstract 40, The Endocrine Society, Maryland
25. Rathnam, P., and Saxena, B. B. (1975) J. Biol. Chem. 250, 6735-6746
26. Saxena, B. B., and Rathnam, P. (1968) in Gonadotropins, 1968
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from

1002 Amino Acid Sequence of FSH-0
(Rosemberg, E., ed) pp. :1-12, Geron-X Inc., Los Altos, Califor- 50. Elson, L. A., and Morgan, W. T. J. (1933) Biochem. J. 27, nia 1824-1828
27. Sanger, F. (1945) Biochem. J. 39,507-515 28. Levy, A. L. (1954) Nature 174, 126-127 29. Narita, K. (1970) in Protein Seqknce Determination (Needleman,
S. B., ed) p. 36, Springer-Verlag, New York 30. Reichert, L. E., Jr., Rasco, M. A., Ward, D. N., Niswender, G. D.,
and Midgley, A. R., Jr. (1969) J. Biol. Chem. 244,5110-5117 31. Landefeld. T. D., Grimek H. J., and McShan, W. H. (1972)
Biochem. Biophys. Res. Comma. 46, 463-469 32. Cunningham, B. A., Wang, J. L., Berggard, I., and Peterson, P. A.
(1973) Biochemistry 12, 4811-4822 33. Beaven, G. H., and Holiday, E. R. (1952) Adu. Protein Chem. 7,
3 19-386
51. Inglis, A. S., and Liu, T.-Y. (1970) J. Biol. Chem. 245, 112-116 52. Snell, E. E. (1957) Methods Enzymol. 111,477 53. Rathnam, P., and Saxena, B, B. (1974) in Gonadotropins and
Gonadal Function (Moudgal, N. R., ed) pp. 23-41, Academic Press, New York
54. Rathnam, P., and Saxena, B. B. (1972) Abstract of Four Interna- tional Congress of Endocrinology, p. 437, The Endocrine So- ciety, Maryland
34. Henninp. U.. Helinski. D. R., Chao, F. C.. and Yanofskv. C. (1962)
55. Narita, K. (1970) in Protein Sequence Determination (Needleman, S. B., ed) pp. 74-77, Springer-Verlag, New York
56. Grimek, H. J., Nuti, L. C., and McShan, W. H. (1971) Biochem. Biophys. Res. Commun. 45,879-886
57. Papkoff, H. (1971) in Structure-Actiuity Relationships of Protein and Polypeptide Hormones (Margoulies, M., and Greenwood, F. C., eds) pp. 73-79, Excerpta Medica, Amsterdam
58. Nuti, L. C., Grimek, H. J., Braselton, W. E., and McShan, W. H. (1972) Endocrinology 91, 1418-1422
59. Carlsen, R. B., Bahl, 0. P., and Swaminathan, N. (1973) J. Biol. Chem. 248, 6810-6827
J. Biil. Chem. 237, 1523-1530 35. Velick, S. F., Parker, C. W., and Eisen, H. N. (1960) Proc. N&l.
Acad. Sci. U. S. A. 46, 1470-1482 36. Patchomik. A., Lawson. W. B.. Gross. E.. and Witkou. N. (1960) J.
Am. Chem. Sot. 82, 5923-5927 37. Snies. J. R., and Chambers, D. C. (1949) Anal. Chem. 21.
‘1249-1266 38. Opienska-Blauth, J., Charezinski, M., and Berbec, H. (1963) Anal.
Biochem. 6, 69-76 39. Scoffone, E., and Fontana, A. (1970) in Protein SequenceDetermi-
nation (Needleman, S. B., ed) pp. 208-210, Springer-Verlag, New York
40. Liu, T.-Y., and Chang, Y. H. (1971) J. Biol. Chem. 246, 2842-2848 41. Amoss, M. S., Jr., Monahan, M. W., and Verlander, M. S. (1974)
J. Clin. Endocrinol. Metab. 39, 187-190 42. Parlow, A. F., and Shame, B. (1974) J. Clin. Endocrinol. Metab.
39, 1955198 43. Saxena, B. B., and Rathnam, P. (1971) in Structure-Actiuity
Relationships of Protein and Polypeptide Hormones (Margou- lies, M., and Greenwood, F. C., eds) pp. 122-131, Excerpta Medica, Amsterdam
44. Shame, B., and Parlow, A. F. (1974) J. Clin. Endocrinol. Metab. 39, 199-202
45. Saxena, B. B., and Rathnam, P. (1968) in Gonadotropins, 1968 (Rosemberg, E., ed) pp. :1-l& Geron-X Inc., Los Altos, California
46. Beaven, G. H., and Holiday, R. E. (1952) Adu. Protein Chem. 7, 319-386
47. Goodwin. T. W.. and Morton, R. A. (1946) Biochem. J. 40, 628-632 48. Scoffone, E., and Fontana, A. (1970) in Protein Sequence De-
termination (Needleman, S. B., ed) p. 185, Springer-Verlag, New York
49. Werner, I.. and Odin, L. (19521 Acta Sot. Med. Upsal. 57,230
60. Sairam, M. R., and Li, C. H. (1973) International Res. Comm. System 1, 14
61. Bewley, T. A., and Li, C. H. (1974) in Lactogenic Hormones, Fetal Nutrition and Lactation (Josimovich, J. B., Reynolds, N., and Cobo, E., eds) pp. 19-32, J. Wiley & Sons, New York
62. Pierce, J. G., Liao, T. H., Cornell, J. S., and Carlsen, R. B. (1971) in Structure-Actiuity Relationships of Protein and Polypeptide Hormones, Part I (Margoulies, M., and Greenwood, F. C., eds) pp. 91-98, Excerpta Medica, Amsterdam
63. Ward, D. N., and Liu, W. K. (1971) in Structure-Actiuity Relationships of Protein and Polypeptide Hormones, Part I (Margoulies, M., and Greenwood, F. C., eds) pp. 80-90, Ex- cerpta Medica, Amsterdam
64. Papkoff, H. (1971) in Structure-Actiuity Relationships of Protein and Polypeptide Hormones (Margoulies, M., and Greenwood, F. C., eds) pp. 7:1-79, Excerpta Medica, Amsterdam
65. Winzler. R. J. (1973) in Hormonal Proteins and Peotides (Li. C. H., ed) Vol. I, pp. l-15, Academic Press, New York 1
66. Hughes, W. L. (1954) in Proteins (Neurath. H.. and Bailev. K.. eds) $01. 2, pp. 663-664, Academic Press, New York
67. Northrop, J. H., Kunitz, M., and Herriot, R. N. (1948) in Crystalline Enzymes, p. 105, Columbia University Press, New York
68. Hirs, C. H. W., Stein, W. H., and Moore, S. (1954) J. Biol. Chem. 211,941-950
69. Smith, E. L., and Stockell, A. (1954) J. Biol. Chem. 207, 501-514
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Amino Acid Sequence of FSH-P 1003
: i
E
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Amino Acid Sequence of FSH-p
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Amino Acid Sequence of FSH-B 1005
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from

B B Saxena and P Rathnamhuman pituitary glands.
Amino acid sequence of the beta subunit of follicle-stimulating hormone from
1976, 251:993-1005.J. Biol. Chem.
http://www.jbc.org/content/251/4/993Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/251/4/993.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on Decem
ber 25, 2019http://w
ww
.jbc.org/D
ownloaded from