ammi visnaga (l.) lamarck (apiaceae): associated ... · toothpick ammi (ammi visnaga (l.) lamarck,...
TRANSCRIPT

Biological Agriculture and Horticulture, 1989, Vol 6, pp. 24 1-268
0 1989 A B Academic Publishers Printed in Great Britain
0 144-8765/89 $10
Ammi visnaga (L.) Lamarck (Apiaceae): Associated Beneficial Insects and Implications for Biological Control, with Emphasis on the Bell-Pepper Agroecosystem
Robert L. Buggl" and L. Theodore Wilson2
'Division of Biological Control, University of California, 1050 San Pablo Avenue, Albany, CA 94706, U.S.A.; and 2Department of Entomology, University of California, Davis, CA 9561 6, U.S.A.
ABSTRACT
Toothpick ammi (Ammi visnaga (L.) Lamarck, Apiaceae), a summer annual of Eurasian origin, commonly occurs along agricultural field margins in the Sacramento Valley of northern California, typically flowering from June to August. Observations during 1979-1984 yielded 65 distinguishable insect taxa attending the flowers, including 60 groups that were at least in part entomophagous. Peak attendances occurred for Braconidae, Ichneumonidae, Sarcophagidae, Sphecidae, Tachinidae, and Vespidae during early morning hours.
Hay alfalfa (Medicago sativa L., Fabaceae) was grown in replicated plots with borders ofeither: ( 1 ) toothpick ammi, ( 2 ) common knotweed (Polygonum aviculare L., Polygonaceae), (3) prostrate pigweed (Amaranthus graecizans L.. Amaranthaceae), or (4) Clean-cultivated edges. Borders of flowering toothpick ammi attracted numerous entomophaga, but had no effect on densities of minute pirate bug (Orius trisricolor [White], Hemiptera: Anthocoridae) occurring in alfalfa.
Pepper (Capsicum annuum L., Solanaceae) was grown with borders of either: ( I ) Vegetative Ammi, from which the flowers were repeatedly clipped, ( 2 ) Normally-flowering Amwi; or (3) Clean-cultivated borders (control) to assess impact on abundance of entomophaga. Predator densities and efficiencies within the plots were assessed weekly by: ( I ) Weekly whole-plant visual inspection; (2) Use of sentinel egg masses of beet armyworm (Spodoptera exigua (Hiibner), Lepidoptera: Noctuidae) or omnivorous leafroller (Platynota srultana Walshingham, Lepidoptera: Tortricidae), glued to the apical foliage and later inspected for occupation, or damage, by predators; and (3) Chunks of tuna placed on cards at ground level and later inspected for occupancy by bigeyed bugs (Geocoris spp., Hemiptera: Lygaeidae). Whole-plant visual inspection revealed a complex of generalist predators; those found on sentinel egg masses were in decreasing order of frequency Orius rristicolor, Geocoris pallens StAl, Lygus spp. (Hemiptera: Miridae), G. africolor Montandon, Collops vitratus (Say) (Coleoptera: Melyridae), and Nabis sp.
*Present address: University of Georgia, Department of Entomology, Coastal Plain Experiment Station, Tifton, GA 31793-0748, U.S.A.
24 1

242 ROBERT L. BUGG AND L. THEODORE WILSON
(Hemiptera: Nabidae). Weekly inspection of whole pepper plants failed to indicate differences in Orius density due to border regime. Likewise, there were no significant differences among weed regimes in numbers of Orius per sentinel egg mass placed on pepper plants amid the plots. However, another trial indicated that Orius recruitment to egg masses was higher on pepper plants with immediately-adjacent Ammi. Per-plant abundance of Geocoris and predation efficiency (Geocoris per lepidopterous egg mass) were separately regressed against Geocoris per tuna bait. Statistically-significant relationships were obtained when data were assessed across all dates, employing the mean counts for each date.
Pending further testing, toothpick ammi should be tolerated in field- and road-side settings, as it is seldom a problem in crops and apparently supplies sustenance to many entomophagous insects.
I NTR ODUCTIO N
Flowering weeds have frequently been cited as important food resources for entomophagous insects (see reviews by: Altieri & Whitcomb, 1979; Altieri & Letourneau, 1982; and Bugg et al., 1987). Many of the relevant studies have emphasized the value of flowering Apiaceae that supply dietary pollen and nectar, leading to increased longevity and fecundity in adult parasitic insects, Generalist predators may also rely to varying extents on nectar and pollen (Bugg et al., 1987).
Toothpick ammi (Ammi visnaga [L.] Lamarck, Apiaceae), a summer annual of Eurasian origin (Munz & Keck, 1973) commonly occurs along agricultural field margins in the Sacramento Valley of northern California. Typically flowering from mid-June through mid-August, the plant features umbels of 5.0 to 11.4cm diameter, comprising numerous small flowers. We were particularly interested in the possibility of enhancing the density and efficiency of minute pirate bug (Orius tristicolor [White], Hemiptera: Anthocoridae), a generalist predator that attacks many pests of vegetable crops, and which can occur on toothpick ammi at densities as high as 22 per umbel (W. Barclay, unpublished data). Several Anthocoridae, including 0. tristicolor, have been shown to respond favorably to vegetational diversifica- tion (Barney et al., 1984; Letourneau & Altieri, 1983; Solomon, 1980), although this has not consistently been the case (Bugg et a/ . , 1987). Salas- Aguilar (1976) suggested the use of flowering plants to enhance biological control by 0. tristicolor, which is known to feed on thrips and pollen (Salas- Aguilar & Ehler, 1977).
The present study was designed to assess patterns of visitation by insects to flowers of Ammi, and implications for biological control of insect pests in agroecosystems.

FLOWERING AMMl VISNAGA AND BIOCONTROL 243
METHODS
The present study comprises four sets of experiments or observations conducted from 1981-1984 in Yolo County, California. The first involved cataloguing insects attending flowers of Ammi. The second set concerned the pattern of daylight attendance by insects. The third involved hay alfalfa (Medicago sativa L., Fabaceae) grown in replicated plots with borders of Ammi or other weeds, and assessed seasonal patterns of floral visitation by entomophagous insects. Also, sweepnet samples were made in the alfalfa, to assess possible effects of weedy borders on densities of Orius tristicolor. The final set involved bell pepper (Capsicum annuum L., Solanaceae) grown in replicated plots with clean-cultivated margins or borders' of flowering or vegetative Ammi. We conducted a census of flower visitors to detect seasonal patterns of attendance, and detailed assessment of generalist predators occurring amid pepper foliage and at ground level. The latter data not only provided information on possible effects of flowering Ammi, but also contributed new information about generalist predators in the bell-pepper agroecosystem.
List of Flower Visitors
A list of taxa whose members visit Ammi flowers was compiled during the years 1979-1984, based on observations and collections made in Yolo County, California.
Diel Patterns of Flower Visitation
To investigate timing of flower visitation, on August 5th, 1981, insects were collected at half-hour intervals from dawn until after dusk (approximately 0600-1830 Pacific Daylight Time) from a specimen of Ammi discovered some 5 km east of Woodland, Yolo Co., growing in a 74-ha field of overwintered sugarbeet (Beta vulgaris L. convar. crassa [Alefeld] J. Helm, Chenopodiaceae). The toothpick ammi plant had 27 umbels, and was located near the northwest corner of the field, with a field of overwintered alfalfa 50 m to the west, and a stockyard 30 m to the north. At each half-hour interval, we collected for 15 minutes, using an aerial net (net aperture'diameter 38-cm, fine mesh) to try to capture all insects visiting the Ammi plant. Insects were preserved in ethanol, then sorted to taxa.

244 ROBERT L. BUGG AND L. THEODORE WILSON
Alfalfa Experiments
Several of the entomophaga observed visiting flowers of Ammi are also common in the hay-alfalfa agroecosystem. We therefore tested the possible role of toothpick ammi in attracting and sustaining elevated densities of these entomophaga. During 1984, plots of overwintered hay alfalfa (cv ‘Amador’ were used for further experiments (see: Bugg et al., 1987, for details on establishment practices). The experiment employed a 4 X 4 Latin square design, with 6.1 X 6.1 m plots of alfalfa separated by 6.1 m clean-cultivated alleys. East and west borders of the plots featured strips of 0.61 m width devoted to alternative regimes: (1) toothpick ammi, (2) common knotweed (Polygonum aviculare L., Polygonaceae), (3) prostrate pigweed (Amaranthus graecizans L., Amaranthaceae), and (4) pure alfalfa (control). The alfalfa was mowed only once, in mid April, during the course of the 1984 studies. Flood irrigation was performed as needed.
Common knotweed and prostrate pigweed were established by allowing pre-existing weed borders to reseed naturally, and later uprooting unwanted plant species. Ammi was established by two techniques. Ammi was seeded during mid-June, 1983, into seedling trays with 2.54 cm diameter cells, using a soil mixture of 3: 2: 1 , sandy loam to delta peat to sand. Plants were raised in a lath house, watered as necessary, and transplanted to the appropriate plot borders during early July. Plants were installed, seven to a plot, at ca. 1 m intervals. At the same time, the same borders were sown with Ammi seed, approximately 50 g per border, which was incorporated by raking. The transplants provided flowers during spring of 1984, whereas plants that grew from seed provided flowers during the summer.
Once flowering began, surveys of flower visitors were made during late- morning hours on sixteen occasions between days 108 and 228, by visually inspecting the borders of toothpick ammi. The procedure involved one observer walking slowly from north to south along each border of Ammi, and enumerating observed insects. Data were collected for aphidophagous Syrphidae, Tachinidae, pooled parasitic Hymenoptera (Chalcididae, Ich- neumonidae, and Braconidae), and pooled predatory Hymenoptera (Eumenidae, Vespidae and Sphecidae).
Based on prior observations, minute pirate bug was known to occur at relatively high densities on both toothpick ammi (R.L. Bugg, pers. obs.) and common knotweed (Bugg et a/., 1987). Therefore sweepnet samples (net aperture diameter 38 cm, heavy cotton bag of depth 63.50 cm, handle length 66cm) were taken from the alfalfa on days 152, 159, 177, 183, and 188. On each date, during the early- to mid-afternoon hours, two sweeps were taken at each of four stations within each of the sixteen plots. Stations were located amid the alfalfa, ca. 0.61 m from borders. Both nymphal and adult Orius were

FLOWERING AMMI VISNAGA AND BIOCONTROL 24 5
recorded. Untransformed data for Orius (nymphs and adults pooled) were analyzed by separate ANOVA for each date, and an ANOVA for overall totals. Initially, the mean square for “among plots, corrected for border regime, row, and column” was used as the error term in a test for effects due to border regime, row and column. However, since no effects due to row or column were detected, these two terms were discarded from the model and one-way ANOVA was employed to test for effect due to border regime.
Pepper Studies
During 1984, replicated plots of bell pepper (cultivars ‘Emerald Giant’ [Northup King] and ‘Shamrock’ [Asgrow], in mixed culture) were used to test for influences of adjacent flowering Ammion biological control of insect pests by Orius tristicolor, and to assess seasonal patterns of predator activity in the bell-pepper agroecosystem. Among generalist predators attending flowers of Ammi visnaga, Orius was by far the most consistently encountered. Moreover, Bugg et al. (1987) found that Orius occurred amid bell-pepper foliage through much of the summer. Thus, this predator was the focal point of our attempted enhancement scheme. At the same time, because biological pest control is still a poorly-understood aspect of vegetable culture, we decided to document seasonal occurrence of other entomophaga in the bell-pepper agroecosystem.
The experimental site was a 70X80m parcel located at the Student Experimental Farm, University of California, Davis Campus. The experiment involved a randomized complete block design featuring four blocks, each with three border regimes: (1) Vegetative Ammi, from which developing inflorescence were repeatedly clipped prior to flowering, at approximately weekly intervals; (2) Normally-flowering Ammi; and (3) Control-clean- cultivated borders. Pepper plots were 5.49 X 5.32 m, with 0.76 m borders on east and west sides. Plots were separated by 15.25 m clean-cultivated alleys.
On May 1, the parcel was disced, and 0.76 m beds were created, running in the east-west direction. Pepper plants had been seeded in early April into 2.54 cm-width Speedling@ trays, and grown in the greenhouse: toothpick ammi was obtained by excavating small plants found near Woodland, Yo10 County, California. Pepper and toothpick ammi were transplanted to the plots on May 18th to 20th (days 139 to 141). Pepper plants were installed in single rows at 0.43 m intra-row spacing; toothpick ammi was planted along the east and west borders of the appropriate eight plots, with one plant installed atop each of the seven rows. Care involved weekly furrow irrigation and application of fish emulsion (N:P:K 5:l:l) as needed.
From days 191 to 231, flowering borders were inspected at approximately weekly intervals during late-morning hours, using the same procedure as

246 ROBERT I,. BUGG AND L. THEODORE WILSON
mentioned above for the alfalfa study, with data being collected for aphidophagous Syrphidae, Tixhinidae, parasitic Hymenopotera, and Ves- poidea.
In weekly assessments of predator densities and efficiencies amid the bell pepper plants, we employed diagonal transects running from northwest to southeast through each plot. Each of these transects featured four points located in alternate rows. Pepper plants sampled were randomly chosen from those occurring near the transect points.
Thirteen, approximately-weekly inspections of whole pepper plants were made from days 177 to 254, and per plant counts of Orius(adu1ts and nymphs pooled) were assessed. Two-way ANOVA was employed, after transforming the overall means for each plot (y’= SQRTCy]), with the F-test for effect due to border regime using as denominator the mean square for “among plots, corrected for border regime and block”. In no event was a significant effect due to block found; therefore, this term was discarded, and a one-way ANOVA conducted. Where not otherwise noted, this was the case in all subsequent F-tests. A one-tailed, unpaired t-test was employed to test for effect of pepper cultivar on Oriudplant, using point on the transect as replicate, with untransformed counts pooled across dates.
In testing for predation efficiency by Orius, we employed sentinel egg L masses of beet armyworm (BAW-Spodoptera exigua (Hubner); Lepidoptera: Noctuidae) or omnivorous leafroller (OLR-Plarynota stultana Walshingham; Lepidoptera: Tortricidae), obtained from laboratory culture. Egg masses of both species are known to be attacked by several generalist predators, including Orius (Atkins et a/., 1957; Eveleens et a/., 1973). Paper towels (BAW) or cardboard cartons (OLR) on which egg masses had been deposited were cut into approximately 5 X 5 mm pieces, each including 15 to 30 eggs, which were then refrigerated until needed. Sentinel egg masses were used on 17 occasions from days 162 through 258. OLR egg masses were used on days 195, 197 and 208; on all other dates, BAW eggs were used. During the early afternoon hours on each date, an egg mass was glued to the apical foliage of each of four plants occurring along diagonal transects within.each plot; on days 223 and 227, an additional two baits were used per plot. Thus, 840 egg masses were used in this experiment. Following one-hour exposure periods, egg masses were inspected for number of Orius occurring on or within 5cm of the baits. Because Orius was not observed on egg masses until day 188, only data obtained on or after that date were analyzed. Two-way ANOVA was used, as mentioned -for the ‘data from whole-plant inspection. A two-tailed, unpaired t-test was employed to test for effect of pepper cultivar on Oriudegg mass, using points on the transect as replicates, with untransformed data pooled across dates.
On six dates during September, sentinel BAW egg masses were used along

FLOWERING AMMI VISNAGA AND BIOCONTROL 247
the east and west margins of control plots and those with flowering Ammi. Six egg masses were used per plot on days 250,252, and 258, and 14 on days 258, 264, and 270; thus, this experiment involved a total of 480 egg masses. In all cases, one egg mass was used per plant on any given date, and baits were allotted equally between the east and west edges of all plots. Inspections for Orius within 5cm of egg masses were made after 24 hour exposures. The resulting data were analyzed for effect due to border regime by one-tailed paired and unpaired t-tests, after transformation of overall means for each plot as mentioned above.
Overall seasonal patterns of predator activity were assessed using data from the above-mentioned weekly placement of sentinel ‘egg masses and the inspection of whole plants. Lists were made of predators observed by both methods. Border regime, block, pepper cultivar, and plot were ignored, and by-date means and standard errors of means were generated for minute pirate bug and western bigeyed bug (Geocoris pallens StAl, Hemiptera: Lygaeidae), using transect point as the sample units.
Chunks of tuna were also used to assess abundance of Geocoris spp. At 14, approximately-weekly intervals, beginning on June 7th (day 159), 2.5 X 2.5 cm chunks of canned tuna (chunk light, packed in vegetable oil) were placed on 5 X 7.5 cm cards. Four baits were used per plot, and were placed on the ground at the bases of four pepper plants situated along the diagonal transects. After approximately 1-hour exposure periods, baits were inspected for western bigeyed bug occurring on or within 5cm of baits.
Regression analysis was used to explore the relationship between counts from the easier-to-use tuna baiting and other sampling techniques (whole- plant inspection and sentinel egg masses). Data for bigeyed bugs (G . pallens and G. atricolor Montandon, all postembryonic stages) were employed, grouped according to the week of sampling. In each case, the proposed predictive variable was used as the independent variable. In instances where one sampling procedure was employed on more occasions than was another, the “extra” observations were paired with data obtained from the nearest date.
RESULTS
List of Flower Visitors
Table 1 contains the list of insects observed visiting the flowers of Ammi visnaga. Sixty-five distinguishable insect taxa were observed, including 60 groups that were at least in-part entomophagous. Predatory taxa observed included 6 Hemiptera, 3 Coleoptera, 7 aphidophagous Syrphidae (Diptera),

~ , 248 ROBERT L. BUGG AND L. THEODORE WILSON
TABLE 1
Insects feeding at flowers of toothpick ammi, Ammi visnaga (L.) Lamarck (Apiaceae), in Yo10 County, California, 1979-1984.
Order Family Subordinate taxon and authority
Hemiptera Lygaeidae
Miridae Nabidae Anthocoridae Reduviidae
Coleoptera Melyridae Coccinellidae
Neuroptera Chrysopidae Diptera Syrphidae
Tachinidae
Sarcophagidae
Hymenoptera Braconidae
Ichneumonidae
Geocoris pallens Stl l tieocoris punctipes (Say) Lygus sp. Nabis sp. Orius tristicolor (White) Zelus renardii Kolenati Sinea sp. Collops vittatur (Say) Coccinella novemnotata Herbst Hippodamia convcrgens Guerin-Meneville Chrysoperla carnea (Stephens) Allograpta obliqua (Say) Eristalis tenax (L.) Erisralis sp. Eumerus sp. Eupeodes volucris Osten Sacken Helophilus sp. prob. Melanostoma mellinum (L.) Toxomerus marginata (Say) Metasyrphus sp. Paragus tibialis (Fallen) Platycheirus sp. Scaeva pyrastri (L.) Sphaerophoria sp. Syritta pipiens (L.) Archytas apicifer (Walker) prob. Cylindromya sp. prob. Gymnosoma fuligonsum Robinson-Desvoidy Peleteria texensis Curran Lepesia archippivora (Riley) Leucostoma aterrimum (Villers) prob. Trichopoda plumipes (Fabricius) Senotainia sp. Wohlfahrtia vigil (Walker) Microgaster sp. Chelonus insularis Cresson Chelonus (Microchelonus) sp. Biolysia sp. prob tristis (Gravenhorst) Charitopes albilabris Townes Compsocryptus calypterus (Say) Cryptus rufovinctus (Pratt) Ethylurgus nigriventris Townes prob. Hyposoter sp. Patrocloides montanus (Cresson) Pristomerus spinator (F.) Pterocormus cupitus (Cresson) Pterocormus sp. undet. spp.

FLOWERING AMMI VISNAGA AND BIOCONTROL 249
Order Family Subordinate taxon and authority
Chalcidoidea undet. spp. Chalcididae Spilochalcis delumbis (Cresson) Dryinidae undet. spp. Formicidae Conomyrma bicolor (Wheeler)
Vespidae
Eumenidae Pompilidae Sphecidae
Conomyrma insana (Buckley) Formica sp. Iridomyrmex humilis @la yr) Solenopsis sp. Polistes apachus Saussure Polistes fuscatus (Fabricius) undet. spp. undet. spp. Ammophila sp. undet. Cercerini Chlorion sp. Philanthus sp. Prionyx sp. Tachytes sp. prob. distinctus Smith undet. spp.
TABLE 2
Numbers of insects collected by aerial net from a flowering specimen of Ammi visnaga (L.) Woodland, Yo10 County, California, August 5, 1981.
Taxa Time (Pacific Daylight Saving Time)/No. Collected Per 2, IcMinute Periods
600 700 800 900 1000 1100 1200 1300 1400 1500
Diptera Tachinidae 2 6 3 3 2 1 3 1 0 0 Sarcophagidae 2 3 3 0 1 0 0 2 2 1
Braconidae 0 4 7 4 3 0 1 3 5 3 Ichneumonidae 5 5 3 1 0 0 0 0 1 1 Vespidae 2 1 1 1 0 0 0 0 0 0 Sphecidae 3 1 1 0 2 1 1 0 0 0
Hymenoptera
and among the Hymenoptera, 2 Vespidae and at least seven Sphecidae. Dipterous parasites observed included at least 7 Tachinidae and two Sarcophagidae, whereas hymenopterous parasites comprised at least 3 Braconidae and 10 Ichneumonidae. Several pests were also observed, including Lygus spp. and Calocoris norwegicus Gmelin (Hemiptera: Miridae) and housefly (Musca domestica L., Diptera: Muscidae),
Diel Patterns of Flower Visitation
Table 2 summarizes results from day-long observations and collections at the single specimen of toothpick ammi for Braconidae, Ichneumonidae,

250 ROBERT L. BUGG AND L. THEODORE WILSON
30
a0
10
0 6 7 8 9 10 11 I2 13 14 15 16 17
Time (PDT)
FIGURE la. from Ammi visnaga, Woodland, California, August 5th, 1981.
Numbers of pooled entomophagous insects collected per two, IS-minute periods
6 1 Vespidae Ichneumonidae
6 7 8 9 10 11 I2 13 14 15 16 17
Time (PDT)
FIGURE lb. periods from Ammi visnaga, Woodland, California, August 5th, 198 1.
Ichneumonidae and Vespidae (Hymenoptera) collected per two, 15-minute

FLOWERING AMMI VISNAGA AND BIOCONTROL 25 I
108 113 114 127 139 144
Julian Date
FIGURE 2a. Aphidophagous Syrphidae (Diptera) per 8 borders of flowering Ammi visnuga, alfalfa study, Davis, California, 1984.
Biolesia m
" 10s 113 114 l!N 139 144
Julian Date
FIGURE 2b. Biolysiu sp. prob. tristis (Gravenhorst) (Hymenoptera: Ischneumonidae) per 8 borders of flowering Ammi visnuga, alfalfa study, Davis, California, 1984.

252 ROBERT L. BUGG AND L. THEODORE WILSON
Chalcididae Ichneumonidae
a
0 3 1 - 152 157 180 187 I93 a00 203 218 222 !ZB
JulianDate
120
E 90
a0 80
f & PI
i? 3 3 0 ts
0 152 157 180 187 193 a08 218 !U2 228
JulianDate b

00
C
FLOWERING AMMI VISNACA AND BIOCONTROL 253
152 157 180 187 193 200 206 218 !Z22 228
JulianDate
FIGURE 3a. Parasitic Hymenoptera per 8 borders of flowering Ammi visnugu, alfalfa study, Davis, California, 1984.
FIGURE 3b. Predatory wasps (Hymenoptera: pooled Sphecidae, Polistes apuchus and P. fuscutus [Vespidae], and pooled Eumenidae) per 8 borders of flowering Ammi visnuga, alfalfa study, Davis, California, 1984.
FIGURE 3c. study, Davis, California, 1984.
Tachinidae (Diptera) collected per 8 borders of flowering Ammi visnugu, alfalfa

254 ROBERT L. BUGG AND L. THEODORE WILSON
Sarcophagidae, Sphecidae, Tachinidae, and Vespidae. Two major periods of activity were apparent, one peaking at 0700 hours and the other at 1400 (Fig. la). The diel patterns of flower attendance varied considerably among the groups of entomophaga. Vespids, at one extreme, were captured between 0600 and 0900 hours; ichneumonids occurred during two distinct periods: from 0600 to 0900 and from 1400 to 1700 hours (Fig. lb). The general decrease in entomophage activity during late morning and the general increase beginning around 1200 hou,rs cannot be explained by temperature effects alone, because maximum temperature is reached around 1400 to 1500 hours.
Alfalfa Study
Data for visual inspection of Ammi flowers bordering alfalfa plots are summarized in Figures 2a-b for spring flowering and Figures 3a-c for summer flowering. Peak attendance for pooled aphidophagous Syrphidae occurred on day 127; the onset of hot weather thereafter apparently suppressed Scaevapyrastri (L.) and Eupeodes volucris Osten-Sacken (Fig. 2a). Peak incidence for Biolysia sp. (Hymenoptera: Ichneumonidae) occurred on day 108, the first day of sampling (Fig. 2b). For pooled parasitic Hymenoptera (Ichneumonidae, Braconidae, and Chalcididae), maximum abundance occurred on day 222 (Fig. 3a). For pooled predatory Hymenoptera (Vespidae, Eumenidae, and Sphecidae), the peak occurred on day 228 (Fig. 3b), and for Tachinidae on day 222 (Fig. 3c). The data did not indicate any clear seasonal trends, but the latter three groups all peaked toward the end of the sampling, in mid August.
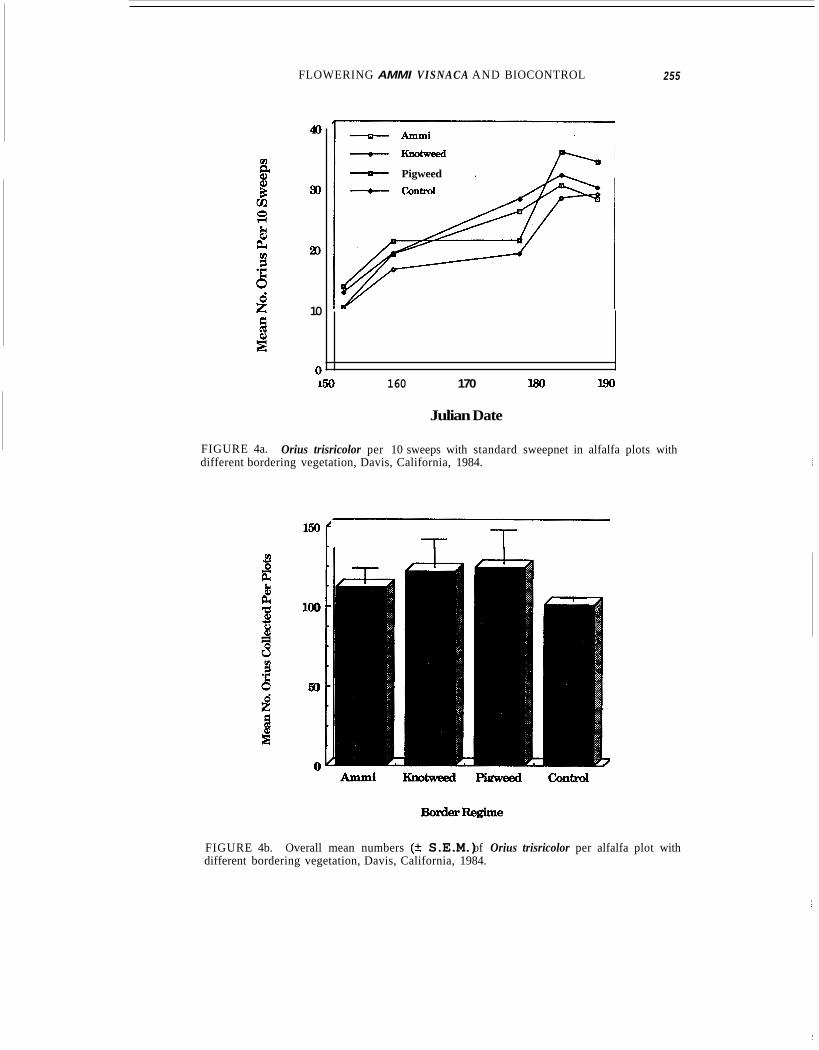
In no case did ANOVA of sweepnet-sampling data indicate any significant influence by border regime on Orius densities in adjoining alfalfa (Fig. 4a-b). Values of P obtained by one-way ANOVA for May 31st; June 7th, 25th, and 29th; July 5th; and overall totals (counts for each plot pooled across all dates) were respectively 0.6967,0.8498, 0.3 187, 0.7503, 0.8857, and 0.6678. Despite the lack of significant differences, on all but the last sampling date mean densities were consistently higher in plots with weedy borders (Fig. 4a),
Pepper Studies
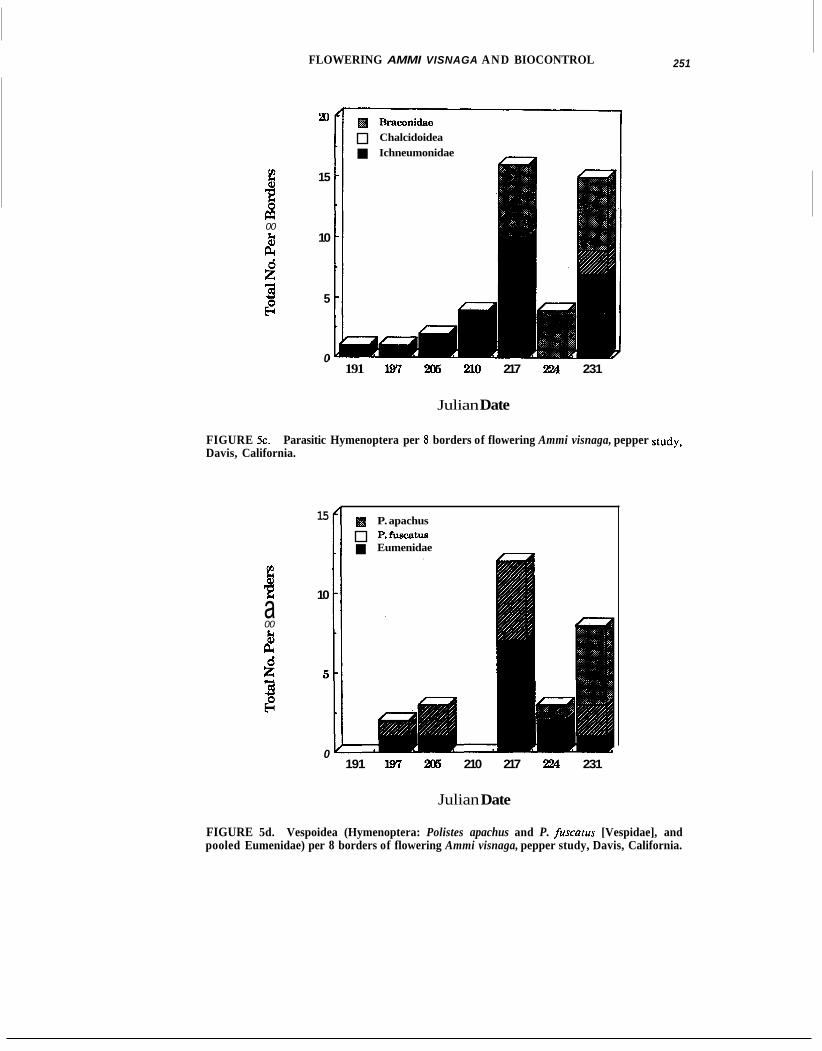
In the pepper studies, weekly visual inspection of flowering borders indicated at least two peaks of entomophage activity, on day 205 and later on day 217 or 224 (Fig. 5a-d).
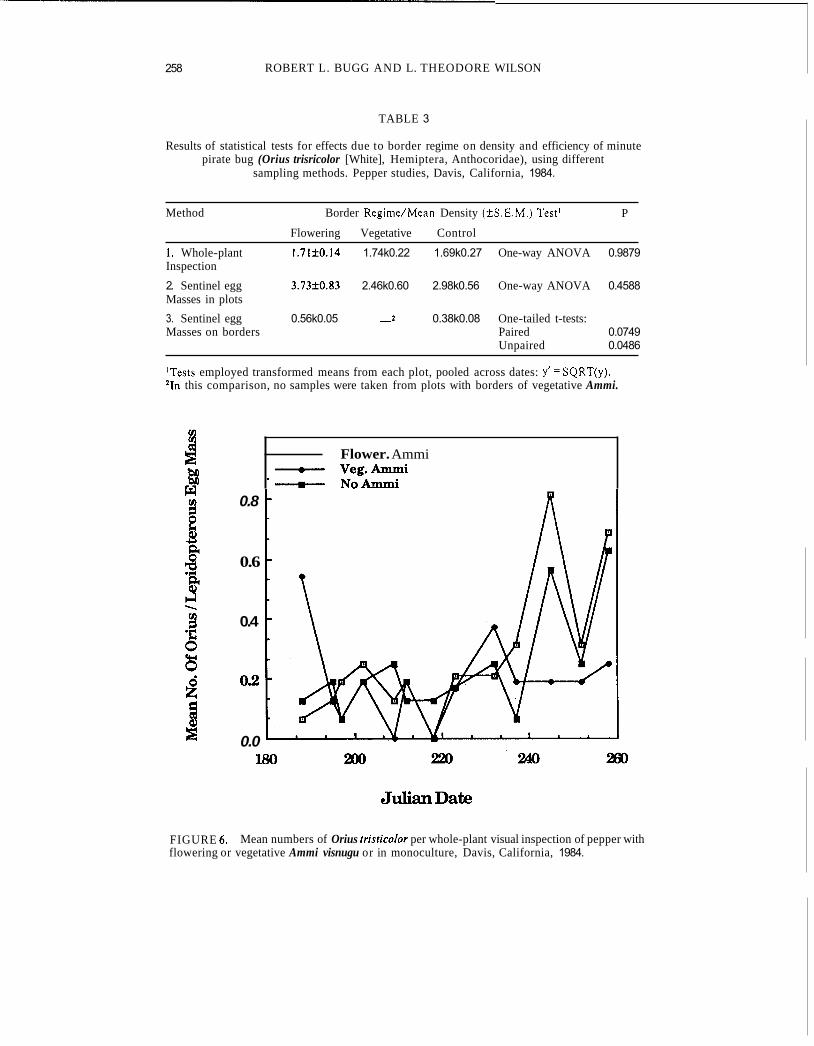
Table 3 summarizes results of various tests for effects due to border regime on density and efficiency of Orius trisitcolor. Inspection of whole pepper
FLOWERING AMMI VISNACA AND BIOCONTROL 255
40
30
a0
10
n
- Pigweed - control
- 150 160 170 Lso 190
Julian Date
FIGURE 4a. different bordering vegetation, Davis, California, 1984.
Orius trisricolor per 10 sweeps with standard sweepnet in alfalfa plots with
150 T - r
100
50
n
BorderF&gime
FIGURE 4b. Overall mean numbers (+ S.E.M.) of Orius trisricolor per alfalfa plot with different bordering vegetation, Davis, California, 1984.

256
m L
15
10
5 -
0 4
ROBERT L. BUGG A N D L. THEODORE WILSON
-
-
191 197 #36 210 217 224 231
Julian Date FIGURE 5a. Aphidophagous Syrphidae (Diptera) per 8 borders of flowering Ammi visnaga, pepper study, Davis, California.
191 197 206 210 217 224 231
Julian Date
FIGURE 5b. , Tachinidae (Diptera) per 8 borders of flowering Ammi visnuga, pepper study, Davis, California.
00
FLOWERING AMMI VISNAGA AND BIOCONTROL
a0 Chalcidoidea Ichneumonidae
15
10
5
0 191 197 206 a0 217 224 231
251
Julian Date
FIGURE 5c. Davis, California.
Parasitic Hymenoptera per 8 borders of flowering Ammi visnaga, pepper study,
P. apachus
Eumenidae P.fuscatus
0 191 197 206 210 217 224 231
15
d 10 a 00
k
Julian Date
FIGURE 5d. Vespoidea (Hymenoptera: Polistes apachus and P. fuscatus [Vespidae], and pooled Eumenidae) per 8 borders of flowering Ammi visnaga, pepper study, Davis, California.
258 ROBERT L. BUGG AND L. THEODORE WILSON
Flower. Ammi - Veg.Ammi -- NoAmmi
TABLE 3
Results of statistical tests for effects due to border regime on density and efficiency of minute pirate bug (Orius trisricolor [White], Hemiptera, Anthocoridae), using different
sampling methods. Pepper studies, Davis, California, 1984.
Method Border Regime/Mean Density (kS.E.M.) Test1 P
Flowering Vegetative Control
1. Whole-plant 1.71k0.14 1.74k0.22 1.69k0.27 One-way ANOVA 0.9879 Inspection
2. Sentinel egg 3.73k0.83 2.46k0.60 2.98k0.56 One-way ANOVA 0.4588 Masses in plots 3. Sentinel egg 0.56k0.05 -2 0.38k0.08 One-tailed t-tests: Masses on borders Paired 0.0749
Unpaired 0.0486
ITests employed transformed means from each plot, pooled across dates: y' = SQRT(y). *In this comparison, no samples were taken from plots with borders of vegetative Ammi.
0.8
0.6
0.4
02
0.0 180
JulianDate
FIGURE 6. flowering or vegetative Ammi visnugu or in monoculture, Davis, California, 1984.
Mean numbers of Orius rrisricdor per whole-plant visual inspection of pepper with

259 FLOWERING AMMI VISNAGA AND BIOCONTROL
TABLE 4
Season-long totals for entomophaga observed on or within 2cm of lepidopterous egg masses glued to apical pepper foliage (approximately 1 hour diurnal exposure), Davis, CA, 1984.
- Predator Species Baits (Number Exposed)
Spodoptera exigua (672) Platynota stultana (144) Orius tristicolor (Whitel
34 5 Nymphs 9 I
Nymphs 9 4
Adults 4 0
Adults 0 1 Nymphs 8 2
Adults 1 1
Adults 3 3
Adults
Geocoris paliens Sthl Adults 9 6
G. atricolor Montandon
Lygus SPP.
Nabis spp.
Collops vittatus (White)
Hippodamia convergens Guerin-Meneville
Diabrotica undecimpunctata undecimpunctata Mannerheim
Chalcidoidea
Adult I 0
Adult 0 1
(undet. egg parasite) 2 0 Tetramorium caespitum (L.) 0 1
(undet. mite, prob. Anystidae) 1 1 Forficula auricularia L. I 0 Araneae (undet. spider) 1 0
Acari
Total 83 26
plants failed to indicate differences in per-plant Orius counts due to border regime. Respective overall means (rt S.E.M.) for “Emerald Giant” and “Shamrock” plants were 2.07 k 0.21 (n = 17) and 1.59 4 0.12 (n=22), and were deemed significantly different (P= 0.0429). However, the absolute difference was not great, and the use of two pepper cultivars probably did not interfere with detection of possible effects due to border regime.
The test of predation efficiency involving pepper plants arrayed throughout the plots yielded no significant differences among weed regimes in numbers of Oriusper egg mass (Fig. 6). Respective means (2 S.E.M.)for “Emerald Giant” and “Shamrock” plants were 0.191 4 0.033 (n = 29) and 0.282 rt 0.036 (n = 30), and were not significantly different (P ~0.0691).
Late-season use of sentinel egg masses glued to plants along borders
ROBERT L. BUGG AND L. THEODORE WILSON I 260
9
a
b
T
140 160 180 a00 220 240 2tio JulianDate
1.2
1.0
0.8
0.6
0.4
02
0.0 140 160 180 a00 220 240 a60
JulianDate
26 I FLOWERING AMMl VISNAGA AND BIOCONTROL
0.6
0.5
0.4
0.3
0.2
0.1
0.0
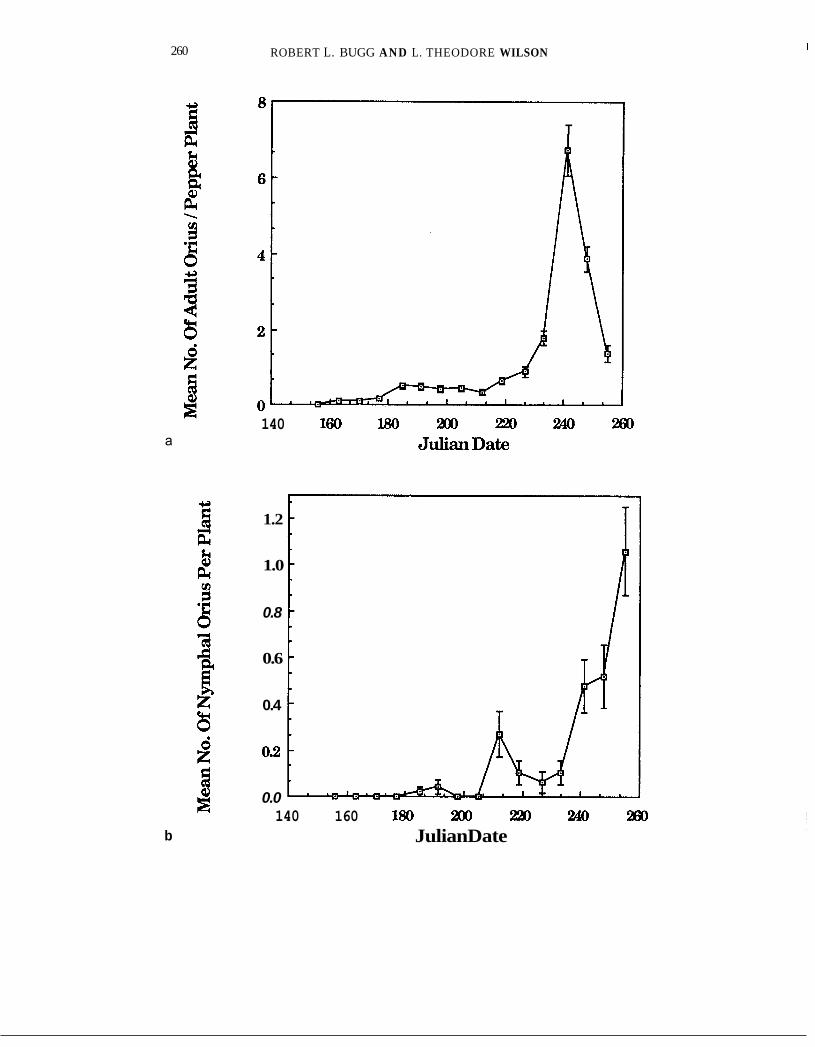
FIGURE 7a. pepper plants (data pooled for across all border regimes), Davis, California, 1984.
Mean numbers of adult Orius frisficolor (k S.E.M.) per visual inspection of whole
FIGURE 7b. Mean numbers of nymphal Orius frisricolor (k S.E.M.) per visual inspection of whole pepper plants (data pooled across all border regimes), Davis, California, 1984.
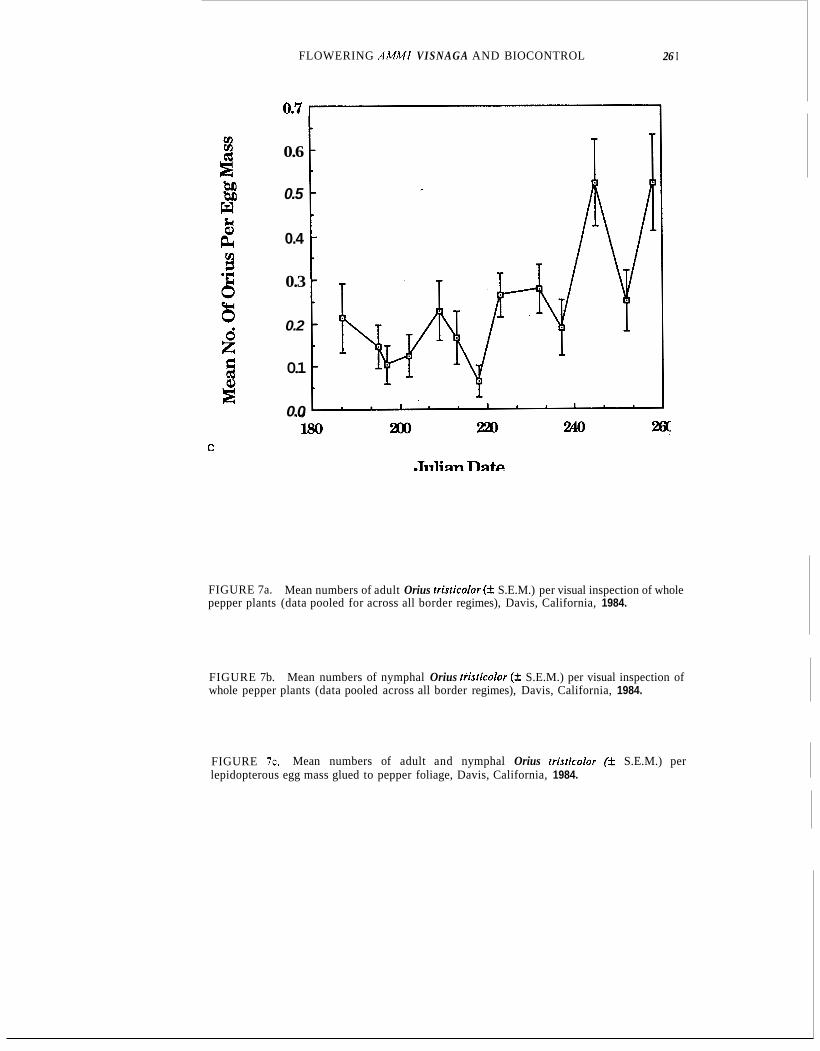
FIGURE 7c. lepidopterous egg mass glued to pepper foliage, Davis, California, 1984.
Mean numbers of adult and nymphal Orius fristicolor (k S.E.M.) per

262
F9
B k
$:
200
1.50
1.00
0.50
0.00
ROBERT L. BUGG AND L. THEODORE WILSON
T Y Adults
140 160 180 200 m, 240 m JdianDate
FIGURE 8. Mean numbers of adult and nymphal Geocoris pallens (+ S.E.M.) per tuna bait placed at bases of pepper plants (data pooled across all border regimes), Davis, California, 1984.
produced what must be termed equivocal results. The paired and unpaired one-tailed t-tests of transformed data yielded non-significant and significant differences, respectively (Table 3). It is noteworthy that mean numbers of Oriuslegg mass were consistently higher on pepper plants with adjoining flowering Ammi, for all dates tested.
Whole-plant visual inspection revealed a complex of generalist predators, including minute pirate bug, bigeyed bugs (G. atricolor and G. pallens), Lygus sp., Collops vittatus (Say) (Coleoptera: Melyridae), an assassin bug (Zelus renardii Kolenati, Hemiptera: Reduviidae), ants (Conomyrma insana [Buckley], Teframorium caespitum [L.], Hymenoptera: Formicidae) and black widow spider (Latrodectus mactans [Fabricius], Arachnida: Araneae: Theridiidae), and several other spiders of the Clubionidae, Thomisidae, and Salticidae. Predators encountered on sentinel egg masses were, in decreasing order of frequency observed, Orius, Geocoris pallens, Lygus spp., G. atricolor, Collops, and Nabis (Table 4).
Whole-plant inspection and use of sentinel egg masses both indicated low early-season activity by Orius, with increases occurring after 220 and peaks on or after day 240 (Fig, 7a-c). Densities of adults consistently exceeded those of nymphs.
Tuna baiting, whole-plant inspection, and data from sentinel egg masses all indicated early-season abundance of Geocoris pallens through mid-July,

FLOWERING AMMI VISNAGA A N D BIOCONTROL
0.6
0.5 T -
140 160 180 200 m, 240 m Julian Date
FIGURE 9a. Mean numbers of adult Geocorispallens(+ S.E.M.)per visual inspection ofwhole pepper plants (data pooled for across all border regimes), Davis, California, 1984.
0.40 1 I 0.30
020
0.10
0.00 140 160 180 a00 220 240 260
Julian Date
FIGURE 9b. whole pepper plants (data pooled for across all border regimes), Davis, California, 1984.
Mean numbers of nymphal Geocorispallens (+ S.E.M.) per visual inspection of
ROBERT L. BUGG AND L. THEODORE WILSON
O.3 I 0.2
0.1
0.0
T I- I
160 180 200 220 2.10
Julian Date
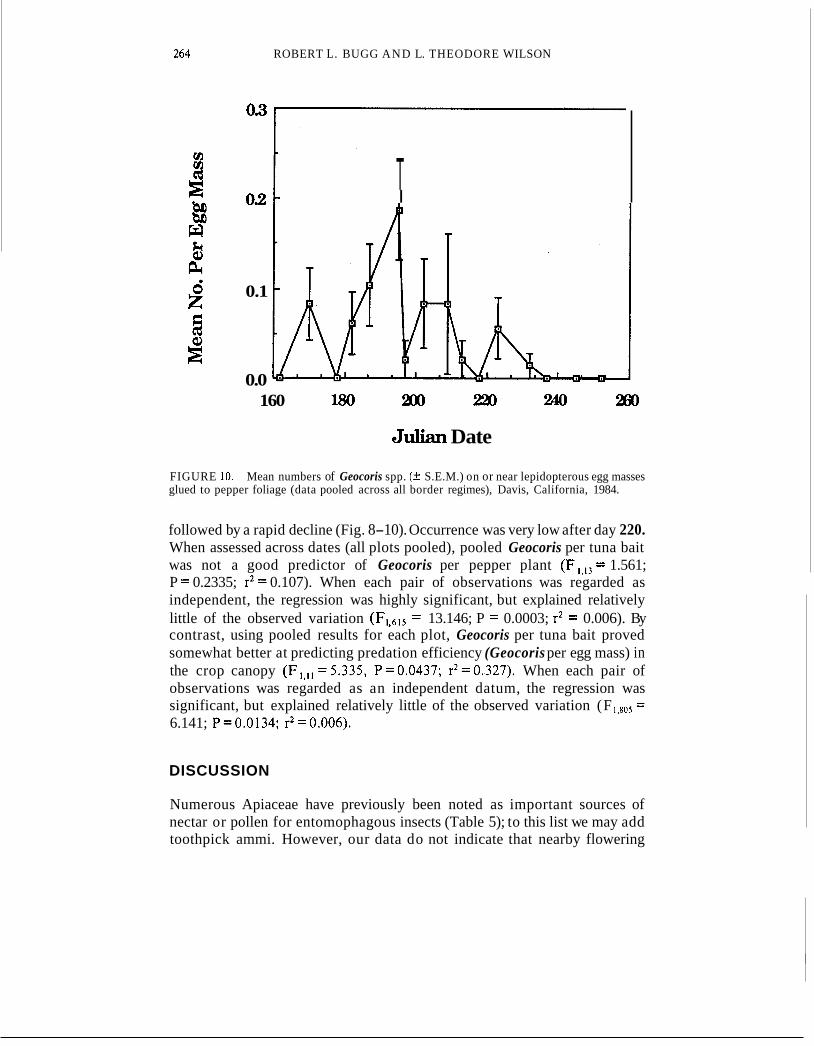
FIGURE 10. glued to pepper foliage (data pooled across all border regimes), Davis, California, 1984.
Mean numbers of Geocoris spp. (+ S.E.M.) on or near lepidopterous egg masses
followed by a rapid decline (Fig. 8-10). Occurrence was very low after day 220. When assessed across dates (all plots pooled), pooled Geocoris per tuna bait was not a good predictor of Geocoris per pepper plant (Fl,13 = 1.561; P = 0.2335; r2 = 0.107). When each pair of observations was regarded as independent, the regression was highly significant, but explained relatively little of the observed variation (F1,615 = 13.146; P = 0.0003; r2 = 0.006). By contrast, using pooled results for each plot, Geocoris per tuna bait proved somewhat better at predicting predation efficiency (Geocoris per egg mass) in the crop canopy (F , , l l ~ 5 . 3 3 5 , P=0.0437; r2=0.327). When each pair of observations was regarded as an independent datum, the regression was significant, but explained relatively little of the observed variation (F 1,805 = 6.141; P=0.0134; r2=0.006).
DISCUSSION
Numerous Apiaceae have previously been noted as important sources of nectar or pollen for entomophagous insects (Table 5); to this list we may add toothpick ammi. However, our data do not indicate that nearby flowering

265 FLOWERING AMMI VISNAGA A N D BlOCONTROL
TABLE 5
Apiaceae known to be important sources of nectar for entomophagous insects.
Plant Species & Authority References Angelica spp. Daucus carota L.
Reviewed by Spradbery, 1973 Bohart and Nye, 1960; Judd, 1970; Hirose, 1966;
Rogochaya, 1971; Syme, 1975 & 1977; Leius, 1963 & 1967; Van Emden, 1963
Coriandrum sativum L. Adesh kevich, 197 1 Heracleum sphondylium L. Spradbery, 1973 Anethum graveolens L. Chumakova, 1971 Eryngium sp. Petroselinum crispum [P. Miller] Peucedanum oreoselinum L. Sawoniewicz, 1973 Conium maculatum L. Foeniculum vulgare P. Miller var. duke Battandier & Trabut Heracleum sibiricum L. Rogochaya., 1971 Pastinaca sativa L.
Reviewed by Spradbery, 1973 Ozols, 1964
Leius, 1967; Review by Spradbery, 1973
Maingay et al.. in prep.
Buntin, 1983; Foster & Ruesink, 1984
toothpick ammi enhanced densities of minute pirate bug in alfalfa. These results contrast with those of Barney et al. (1984) who found some evidence for elevated densities of Orius insidiosus (Say) in grass-infested alfalfa stands. In the latter study, however, the weeds were interspersed among the alfalfa plants, whereas our trial involved discrete borders of weeds. Also, the vacuum-sampling technique employed by Barney et al. (op. cit.) did not distinguish between insects actually occurring on the alfalfa versus those on the weeds, whereas our approach concerned those on the alfalfa.
Our results do not unequivocally indicate increased predator density or efficiency in bell pepper, although the data from use of sentinel egg masses are interesting in light of the relatively low values of P obtained,
As noted by Bugg et al. (1987), there have been few definitive field experiments confirming enhanced biological control through provision of nectariferous plants. As with many prior studies, the alfalfa and pepper studies presented here suffer from several constraints, including the relatively- small distances between consecutive plots (6.10 m and 15.25 m, respectively), the low degree of replication (4), and the short term of the study (1 year). All of these factors might reduce the power to discern effects due to weed regime. In addition, the toothpick ammi did not stop flowering until the studies were nearly complete, Thus, weed phenology would probably not promote dispersal of predators to the crops of interest. In fact, one might well expect Ammi, which afforded nectar, pollen, and thrips, to serve as a sink rather than a source of predators during the relevant time frame. Future experiments should take into careful account the phenological correspondence of crop plants, pests, insectary plants, and biological-control agents.
The data for predator densities and efficiencies in the bell pepper system

266 ROBERT L. BUGG AND L. THEODORE WILSON
indicated a complex of generalist predators attacking lepidopterous egg masses. In a prior study of predators associated with organically-grown bell pepper in Yo10 County (Bugg et af., 1987), one of us also noted that Anthicus biguttufus LeConte, A. punctulatus LeConte, and A . servinus LaFerte (Coleoptera: Anthicidae) occurred on pepper foliage (R.L. Bugg, unpubl. data). In the present study, activity by Geocoris spp. peaked during early summer and declined rapidly after mid-July. This decline was in contrast to a late-season increase in Orius tristicofor abundance. In the field, we have often observed Geocoris preying on Orius, but it is unclear whether the observed pattern of late-season Orius increase is due to release from predation by Geocoris.
Both whole-plant inspection and baiting with lepidopterous egg masses yielded higher estimates of adult than of nymphal Orius. This may indicate use of biased sampling procedures, or that the bell-pepper agroecosystem as exemplified here may not provide good habitat for breeding by Orius.
It has long been clear that Geocoris and Orius assist in suppressing pests of hay alfalfa (Bisabri-Ershadi & Ehler, 1981) and cotton (Ehler & Van Den Bosch, 1974; Ehler, 1977; Eveleens, van den Bosch & Ehler, 1973). Our present studies indicate that they may also benefit bell pepper production, though the extent of this benefit is still unclear.
The future role of nectar sources in Californian agroecosystems also remains unclear, but toothpick ammi should be tolerated in field- and road- side settings, as it is seldom a problem in crops and apparently supplies sustenance to many entomophagous insects.
ACKNOWLEDGEMENTS
We gratefully acknowledge assistance by the following: John Anderson, William Barclay, Robert W. Carlson (determination of Ichneumonidae), Lincoln Constance (determination of Ammi visnaga), William Crepps, Sean S. Duffey, Kevin Etnst, E.E. Grissell (determination of Spi/ochalcis delumbis), Kennth S . Hagen (determination of Anthicidae, and assistance in identifying Geocoris spp., and Syrphidae), Richard Hamm, Jalil Hejazi, Michael Hoffmann, Carolyn Rech, Stephen Risch, Dru Rivers, John Saponara, Hans P. Sauter, Jan Smilanick, Solomon Teklu, Robert Waegell, Philip S. Ward (determination of Formicidae), and N.E. Woodley.
Ref e rences
Adashkevich, B.P. (1971). The role of nectar plants in the attraction of aphidophages to the vegetable pea. In Biological Methods of Protecting Fruit and Vegetable Crops from Pests, Diseases, and Weeds as Bases for Integrated Systems, pp. 2-3. Ministry of Agriculture of the U.S.S.R., All-Union Order of Lenin, V.I. Lenin Academy of Agricultural Sciences, All-Union Scientific Research Institute of Biological Methods of Plant Protection; Kishinev, Union of Soviet Socialist Republics.

FLOWERING AMMI VISNAC‘A AND BlOCONTROL 26 7
Altieri, M.A. & Letourneau, D.K. (1982). Vegetation management and biological control in agroecosystems. Crop Protection. 1, 405-430,
Altieri, M.A. & Whitcomb, W.H. (1979). The potential use of weeds in the manipulation of beneficial insects. HortScience, 14, 12-18.
Atkins, E.L. Jr., Frost, M.H. Jr., Anderson, L.D., & Deal, AS. (1957). The “omnivorous leaf roller,” Plarynota stultana Wlshm., on cotton in California: nomenclature, life history, and bionomics (Lepidoptera: Tortricidae). Annals of the Enromological Society of America, 50,
Barney, R.J., Lamp, W.O., Armbrust, E.J. & Kapusta, G. (1984). Insect predatorcommunityand its response to weed management in spring-planted alfalfa. Protection Ecology, 6, 23-33.
Blsabri-Ershadi, B. & Ehler, L.E. (198 I). Natural biological control of western yellow-striped armyworm, Spodoprera praeflca (Grote), in hay alfalfa in northern California. Hilgardia, 49,
Bohart, G.E. & Nye, W.P. (1960). Insect pollinators ofcarrots in Utah. United States Department of Agriculture, Agricultural Research Service Bulletin 419.
Bugg, R.L., Ehler, L.E. & Wilson, L.T. (1987). Effect of common knotweed (Polygonum aviculare) on abundance and efficiency of insect predators of crop pests. Hilgardia, 55, (7),
Buntin, L.A. (1983). The relationship of wild parsnip, Pastinaca sariva L., to parasitoid populations associated with soybean pests in central Iowa (Biocontrol, Green Cloverworm, Natural Enemies). Ph.D. dissertation. Iowa State University, Ames.
Chumakova, B.M. (1971). The role of supplementary feeding in increasing the effectiveness of parasitic Hymenoptera in the agrobiocenosis. In Biological Methods of Prorecting Fruit and Vegetable Crops from Pests. Diseases, and Weeds as Bases for Inregrated Systems, pp. 83-84. Ministry of Agriculture 01 the U.S.S.R., All-Union Order of Lenin, V.I. Lenin Academy of Agricultural Sciences, All-Union Scientific Research Institute of Biological Methods of Plant Protection; Kishinev, Union of Soviet Socialist Republics.
Ehler, L.E. (1977). Natural enemies of cabbage looper on cotton in the San Joaquin Valley. Hilgardia, 45, 73-106.
Ehler, L.E. & van den Bosch, R. (1974). An analysis of the natural biological control of Trichoplusia ni (Lepidoptera: Noctuidae) on cotton in California. Canadian Entomologist, 106,
Eveleens, K.G., van den Bosch, R. & Ehler, L.E. (1973). Secondary outbreak induction of beet armyworm by experimental insecticide applications in cotton in California. Environmental Entomology, 2, 497-503.
Hirose, Y. (1966). Parasitic Hymenoptera visiting the flowers ofcarrot planted in the truck crop field. Science Bulletin of the Faculty of Agriculture, Kyushu University, Series 2, 22(3), 2 17-223.
Judd, W.W. (1970). Insects associated with flowering wild carrot, Daucus carota L., in southern Ontario. Proceedings of the Entomological Society of Ontario, 100, 176-18 1.
Leius, K. (1963). Effects of pollens on fecundity and longevity of adult Scambus buoliunae (Htg.) (Hymenoptera: Ichneumonidae). Canadian Entomologisr, 95, 202-7.
Leius, K. (1967). Food sources and preferences of adults of a parasite, Scambusbuolianae (Hym.: Ichn.), and their consequences. Canadian Entomologist, 99, 865-871.
Letourneau, D.K. & Altieri, M.A. (1983). Abundance patterns of a predator, Orius tristicolor (Hemiptera: Anthocoridae), and its prey, Frankliniella occidentalis (Th ysanoptera: Thripidae): habitat attraction in polycultures versus monocultures. Environmental Entomology, 12,
Munz, P.A. & Keck, D.D. (1973). A California Flora (with Supplement by P.A. Munz). University of California Press; Berkeley, California.
OZO~S, E.Y. (1964). Nutritional base of imaginal stages of entomophagous insects in agrocenoses. In Biological Control of Agricultural and Forest Pests. (A.I. Cherapanov, ed.), pp. 134-135. Academy of Sciences of the U.S.S.R. Siberian Branch, Biological Institute. Israeli Program for Scientific Translations; Jerusalem, 1969.
Rogochaya, L.G. (1971). The question of the nutritive relationship of tachinids with wild flowering plants and their attraction into orchards. In Biological Methods of Protecting Fruit and Vegetable Crops from Pests, Diseases, and Weeds as Bases for Integrated Systems, pp. 65-66.
25 1-259.
1-23.
1-53.
1067- 1073.
1464- 1469.

268 ROBERT L. BUGG AND L. THEODORE WILSON
Ministry of Agriculture of the U.S.S.R., All-Union Order of Lenin, V.I. Lenin Academy of Agricultural Sciences, All-Union Scientific Research Institute of Biological Methods of Plant Protection; Kishinev, Union of Soviet Socialist Republics.
Salas-Aguilar, J. (1976). Studies on Orius tristicolor (White) in northern California. M.S. thesis, University of California, Davis; 3 1 pp.
Salas-Aguilar, J. & Ehler, L.E. (1977). Feeding habits of Orius tristicolor. Annals of the Entomological Society of America, 70, 60-62.
Sawoniewicz, J. (1973). Gasienicznikowate (Ichneumonidae, Hymenoptera) odwiedzajace Kwiaty Goryszu-Peucedanum oreoselinum L. (Umbelliferae). Folia Forestalia Polonica, Seria
Solomon, M.G. (1980). Windbreaks as a source of orchard pests and predators. In Pests. Pathogens and Vegetation: The Role of Weeds and Wild Plants in the Ecology of Crop Pests and Diseases. (J.M. Thresh, ed.), pp. 273-283. Pitman Advanced Publishing Program; Boston, Massachusetts.
Spradbery, J.P. (1973). Wasps: an Account of the Biologyand Natural History of Solitary andSocia1 Wasps. Sidgwick & Jackson; London.
Syme, P.D. (1975). The effects of flowers on the longevity and fecundity of two native parasites of the European pine shoot moth in Ontario. Environmental Entomologist, 4, 337-346.
Syme, P.D. (1977). Observations on the longevity and fecundity of Orgilus obscurator (Hymenoptera: Braconidae) and the effects of certain foods on longevity. Canadian Entomologist, 109, 995-1000.
van Emden, H.F. (1963). Observations on the effect of flowers on the activity of parasitic Hymenoptera. Entomologist’s Monthly Magazine, 98, 265-270.
A , 21,43-78.
(Received 28th September, 1988; accepted 9th January, 1989).