an exploration of the binding site of aldolase using alkanediol monoglycolate bisphosphoric esters
TRANSCRIPT

384 Biochimica et Biophysica A cta, 742 (1983) 384-390 Elsevier Biomedical Press
BBA31470
AN EXPLORATION OF THE BINDING SITE OF ALDOLASE USING ALKANEDIOL MONOGLYCOLATE BISPHOSPHORIC ESTERS
H. OGATA a, K. TAKEO b, A. KUWAHARA b, R. SUZUNO b, M. FUJIMOTO t, and J. SHIMIZU c
a Division of Chemistry and Biochemistry, The School of Allied Health Sciences, Yamaguchi University, Ube, Yamaguchi-ken, 755, b Department of Biochemistry, Yamaguchi University School of Medicine, Ube, Yamaguchi-ken, 755, and "Department of Food and Nutrition, Yamaguchi Women's University, Yamaguchi, Yamaguchi-ken 753 (Japan)
(Received August 10th, 1982)
Key words: Aldolase," Bisphosphoric ester," Active site; Binding constant; Substrate analog
Alkanediol monoglycolate bisphosphoric esters (P- - O - - CH 2-- CO- - O - - (CH 2) , - - O - - P), which are ana- logues of the aldolase (D-fructose-l,6-bisphosphate D-glyceraldehyde-3-phosphate-lyase, EC 4.1.2.13) sub- strate fructose 1,6-bisphosphate, were synthesized and used for probing its active site. The K i value was lowest when the maximum distance between the phosphorus atoms of the bisphosphate was brought close to that of fructose 1,6-bisphosphate. The binding constants estimated from difference spectra correlate well with K i values for the substrate analogues. Propanediol monoglycolate bisphosphoric ester protected aldolase from inactivation by 1,2-cyclohexanedione, which preferentially atacks arginine-55. However, propanoi phosphate had little protective effect. The synthesized phosphate compounds protected the enzyme against inactivation by trypsin, and also against spontaneous denaturation. These results suggest that the synthesized phosphate compounds bind to aldolase at the active site, which tends to keep the distance constant between the two phosphate-binding sites for the open-chain form of fructose 1,6-bisphosphate, and stabilize the natural conformation of the enzyme. Both arginine-55 and lysine-146 are shown to participate in the phosphate-bind- ing site for the C-l-phosphate of fructose 1,6-bisphosphate.
Introduction
Aldolase (EC 4.1.2.13) cleaves fructose 1,6-bis- phosphate (Fru-l ,6-P2) to dihydroxyacetone phosphate and glyceraldehyde 3-phosphate. Fru- 1,6-P 2 can exist in carbonyl or hemiacetal forms in aqueous solution. At the active site of aldolase, Fru-1,6-P 2 may be fixed exclusively in one of these forms. In the case of muscle aldolase, it has been established that the open-chain carbonyl form can be active, since xylulose 1,5-bisphosphate is a good substrate [1]. Rose's group [2] recently revealed that the concentration of the open-chain carbonyl
Abbreviation: Fru-l ,6-P 2, fructose 1,6-bisphosphate.
0167-4838/83/0000-0000/$03.00 © 1983 ~ . , y i e r Biomedical Press
form of Fru-l ,6-P 2 in aqueous solution is sufficient to account for the steady-state kinetics of the aldolase reaction. Formation of the enzyme-sub- strate complex is followed by elimination of one mole of water between the carbonyl group of Fru-l ,6-P 2 and the 0~-amino group of Lys-227 to form the catalytic Schiff base intermediate [3,4]. This also suggests that Fru-l ,6-P 2 binds to the active site in its open-chain form.
Aldolase has two binding sites for the phos- phate groups of substrates (or inhibitors). One of these sites can bind the phosphate group at C-1 of Fru-l,6-P2, while another binds the phosphate group at C-6. Ginsburg and Mehler [5] indicated that aldolase has a highly specific bindng site for phosphate and an additional site with lower affin-

ity. Further, a relatively weaker binding site was also proposed by Shapiro et al. [6], as the site for binding of the C-6-phosphate group. The amino acid residues, which bind to the C-l-phosphate group of Fru-1,6-P 2 were proposed as Lys-146 [7,8] and Arg-55 [9]. The distance between the two binding sites for phosphate groups was estimated to be 10-12 A [10]. Aldolase has three tryptophan residues in one subunit and it is reported that one of these may be in or near the active site. By ultraviolet difference spectroscopy, Crowder et al. [11] estimated that binding of D-arabinitol 1,5-bis- phosphate (a potent inhibitor) causes a change in the environment of a tryptophan residue on this enzyme.
Propanediol monoglycolate bisphosphoric ester, an analogue of the substrate Fru-l,6-P 2, and re- lated phosphate compounds were synthesized and used as probes for exploring the binding site of aldolase. Inhibitor constants, difference spectra with aldolase and protection against modification or denaturation of the enzyme were determined.
Material and Methods
Chemicals were obtained from the following sources; Fru- 1,6-P 2 tetraeyclohexylammonium salt, glycerophosphate dehydrogenase/triose phosphate isomerase mixture (2 mg/ml, protein ratio 10: 1) and trypsin from Boehringer Mannheim; NADH from Sigma; 1,3-propanediol, 1,4-butanediol, 1,5- pentanediol and bisphenyl chlorophosphate from aldrich Chemical Co.; glycolic acid and ethylen- eglycol from Katayama Chemical Co.; 1,2- cyclohexanedione from Nakarai Chemical Co.
Rabbit muscle aldolase was prepared by the method of Taylor [ 12], and a three-times recrystal- lized enzyme was used. Enzyme solutions were freed of amonium sulfate by dialysis against 10 mM triethanolamine-HC1 buffer (pH 7.5). The concentration of the aldolase was determined from its absorbance a0.1mg _ 0.938 [13], and M r 158000
" " 2 8 0 n m - -
[14]. The aldolase assay used was essentially that described by Blostein and Rutter [ 15]. Each cuvette contained 3.0 ml 0.1 M triethanolamine-HC1 buffer (pH 7.5), 0.1 ml Fru-l,6-P 2 solution (30 mg tetra- cyclohexylammonium salt/ml), 0.05 ml 8 mg/ml NADH, 0.02 ml glycerophosphate dehydro- genase/triose phosphate isomerase mixture, and
385
0.02 ml 0.2 mg /ml aldolase. Values of g m and g i were estimated according to the method of Lin- eweaver and Burk [16].
Difference spectra on binding of ligands to aldolase were measured at 25°C in two pairs of cuvettes with 1.00 cm path lengths. Tracings were taken from 360 nm to 240 nm.
The preparation of 1,3-propanediol monoglyco- late bisphosphoric ester is described as an exam- ple. 1,3-Propanediol monoglycolate: 0.5 mol glyco- lic acid and 1.5 mol 1,3-propanediol were mixed with 100 ml dry benzene. The solution was heated in a paraffin bath at 90-92°C and water was removed for 3 days. The reaction mixture was distilled under reduced pressure and the ester frac- tion was collected at 143°C under 4 mmHg. The bisphosphoric ester was prepared as follows: 90 mmol diphenyl chlorophosphate in 50 ml dry ben- zene were added dropwise to a stirred solution of 30 mmol 1,3-propanediol monoglycolate and 180 mmol pyridine in 100 ml dry benzene maintained at 0°C, then the mixture was stored at 4°C over-
A B
' ! l
Fig. 1. Thin-layer chromatogram of synthetic phosphate com- pounds and their hydrolyzates. Plate: cellulose. Solvents: iso- propanol/acetic acid/water (6:3 : 1). Samples: (A) orthophos- phate (1), 1,3-propanediol monoglycolate bisphosphoric ester (2), phosphoglycolate (3), hydrolyzate of 1,3-propanediol de- rivative (4) and 1,3-propanediol monophosphate (5). (B) hydro- lyzates of; 1,2-ethanediol derivative (1), 1,3-propanediol deriva- tive (2), 1,4-butanediol derivative (3) and 1,5-pentanediol de- rivative (4).
386
J L
HO-CHfC-O-CH2-CH2-C H2-OH 0
i
(p)-O-CH2-Q-O-C H2-CHf CHfO-(P) 0 i ~--~ NH:z s a l t
(ppm)
% x
3
i ' 3 ' [FOp]' 10'
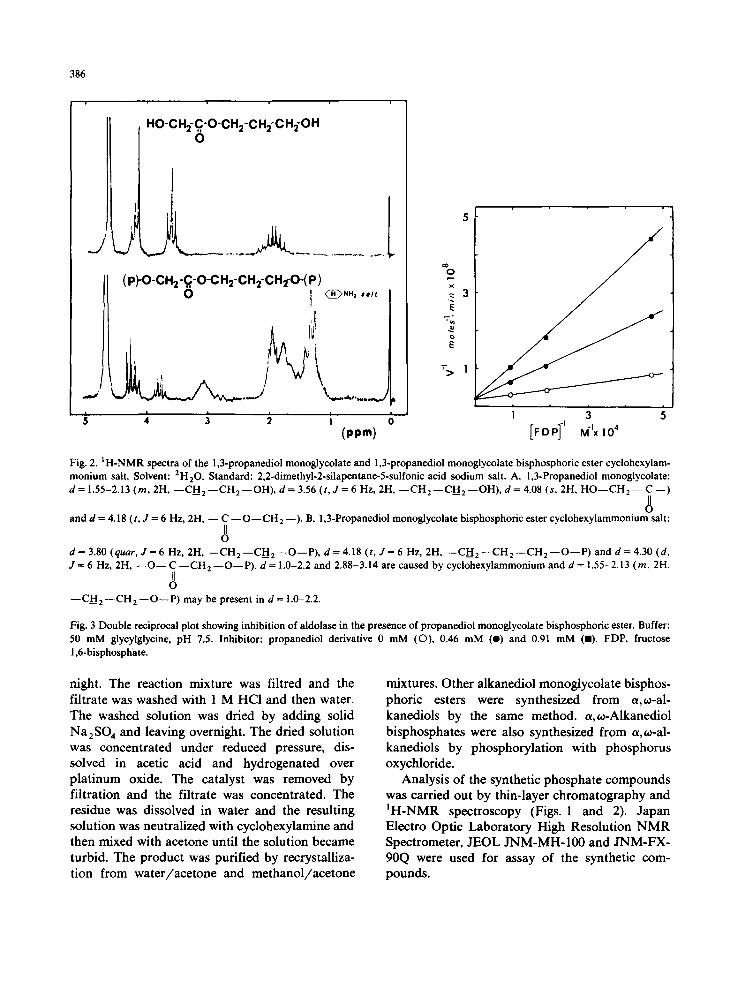
Fig. 2. IH-NMR spectra of the 1,3-propanediol monoglycolate and 1,3-propanediol monoglycolate bisphosphoric ester cyclohexylam- monium salt. Solvent: 2H20. Standard: 2,2-dimethyl-2-silapentane-5-sulfonic acid sodium salt. A. 1,3-Propanediol monoglycolate: d = 1.55-2.13 (m, 2H, - -CFI 2 - - C H 2 - -OH) , d = 3.56 (t, J = 6 Hz, 2H, - - C H 2 - - C H 2 - -OH) , d = 4.08 (s, 2H, H O - - C H 2 - - C - - )
II o
and d = 4.18 (t, J = 6 Hz, 2H, - - C --O--CH2 --). B. 1,3-Propanediol monoglycolate bisphosphoric ester cyclohexylammonium salt: II O
d = 3.80 (quar, J = 6 Hz, 2H, --CH 2 --CH 2 --O--P), d = 4.18 (t, J = 6 Hz, 2H, --CH 2 --CH 2 --CH 2 - -O--P) and d = 4.30 (d, J = 6 Hz, 2H, - - O - - C --CH 2 --O--P). d = 1.0-2.2 and 2,88-3.14 are caused by cyclohexylammonium and d = 1.55-2.13 (m, 2H,
II O
--CH 2 --CH 2 - -O--P) may be present in d = 1.0-2.2.
Fig. 3 Double reciprocal plot showing inhibition of aldolase in the presence of propanediol monoglycolate bisphosphoric ester. Buffer: 50 mM glycylglycine, pH 7.5. Inhibitor: propanediol derivative 0 mM (©), 0.46 mM (e) and 0.91 mM (11). FDP, fructose 1,6-bisphosphate.
night. The reac t ion mixture was f i l t red and the f i l t rate was washed with 1 M HCI and then water. The washed solut ion was dr ied by add ing solid N a 2SO4 and leaving overnight . The dr ied solut ion was concen t ra ted under reduced pressure, dis- solved in acetic acid and hydrogena ted over p l a t i num oxide. The cata lys t was removed by f i l t ra t ion and the f i l t rate was concent ra ted . The res idue was dissolved in water and the resul t ing solut ion was neut ra l ized with cyc lohexylamine and then mixed with acetone until the solut ion became turbid. The p roduc t was pur i f ied by recrystaUiza- t ion f rom w a t e r / a c e t o n e and m e t h a n o l / a c e t o n e
mixtures. Other a lkanedio l monoglyco la te bisphos- phor ic esters were synthesized f rom a,~o-al- kanedio ls by the same method, a , ~ - A l k a n e d i o l b i sphospha tes were also synthesized from a ,~ -a l - kanedio ls by phosphory la t ion with phosphorus oxychlor ide.
Analys is of the synthet ic phospha te compounds was c a r d e d out by thin- layer c h r o m a t o g r a p h y and I H - N M R spec t roscopy (Figs. l and 2). J apan Elect ro Opt ic L a b o r a t o r y High Resolu t ion N M R Spectrometer , J E O L J N M - M H - 1 0 0 and J N M - F X - 90Q were used for assay of the synthet ic com- pounds .
Results
The synthetic esters competitively inhibited muscle aldolase activity (Fig. 3). The inhibitor constants of the synthetic compounds were esti- mated (Table I).
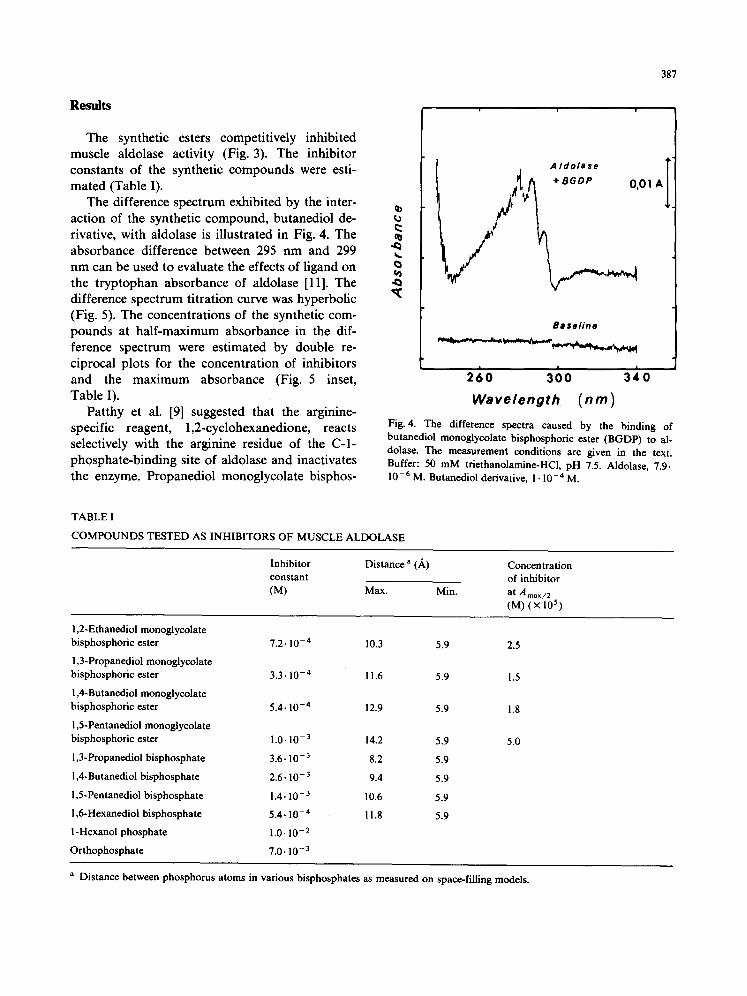
The difference spectrum exhibited by the inter- action of the synthetic compound, butanediol de- rivative, with aldolase is illustrated in Fig. 4. The absorbance difference between 295 nm and 299 nm can be used to evaluate the effects of ligand on the tryptophan absorbance of aldolase [11]. The difference spectrum titration curve was hyperbolic (Fig. 5). The concentrations of the synthetic com- pounds at half-maximum absorbance in the dif- ference spectrum were estimated by double re- ciprocal plots for the concentration of inhibitors and the maximum absorbance (Fig. 5 inset, Table I).
Patthy et al. [9] suggested that the arginine- specific reagent, 1,2-cyclohexanedione, reacts selectively with the arginine residue of the C-l- phosphate-binding site of aldolase and inactivates the enzyme. Propanediol monoglycolate bisphos-
387
A +BGDP 0,0| A
Basel ine
t !
260 300 3~0 Wavelength (nm)
Fig. 4. The difference spectra caused by the binding of butanediol monoglycolate bisphosphoric ester (BGDP) to al- dolase. The measurement conditions are given in the text. Buffer: 50 m M triethanolamine-HCl, pH 7.5. Aldolase, 7.9. 10 -6 M. Butanediol derivative, 1 • 10 -4 M.
TABLE I
C O M P O U N D S TESTED AS INHIBITORS OF MUSCLE ALDOLASE
Inhibitor constant (M)
Distance a (A)
Max. Min.
Concentration of inhibitor
at A max/2 (M) ( × 10 s)
1,2-Ethanedioi monogiycolate bisphosphoric ester
1,3-Propanediol monoglycolate bisphosphoric ester
1,4-Butanediol monogiycolate bisphosphoric ester
1,5-Pentanediol monoglycolate bisphosphoric ester
1,3-Propanediol bisphosphate
1,4-Butanediol bisphosphate
1,5-Pentanediol bisphosphate
1,6-Hexanedioi bisphospbate
l-Hexanol phosphate
Orthophosphate
7.2.10 -4 10.3 5.9 2.5
3.3.10 - 4 11.6 5.9 1.5
5.4- 10 -4 12.9 5.9 1.8
1.0- 10 -3 14.2 5.9 5.0
3.6- 10- 3 8.2 5.9
2 .6 .10- 3 9.4 5.9
1.4.10- 3 10.6 5.9
5.4.10 -4 11.8 5.9
1.0- 10- 2
7.0.10 -3
a Distance between phosphorus atoms in various bisphosphates as measured on space-filling models.
388
0,010 E o, o,
I
0,005
i i
S ' / o:t/ / 0 ~ ,'o
[ P G D P ] i M I x l O 4
I O0 5 110 [PGD P] MxIO -5
Fig. 5. The difference in absorbance, between A 2 9 5 . m and .4299nm, produced by binding of propanediol monoglycolate bisphosphoric ester (PGDP) to aldolase plotted against the concentration of the synthesized phosphate compound. Buffer: 50 mM triethanolamine-HCi pH 7.5. The inset shows a double-reciprocai plot for the concentrations of the inhibitor and the differences in absorbance.
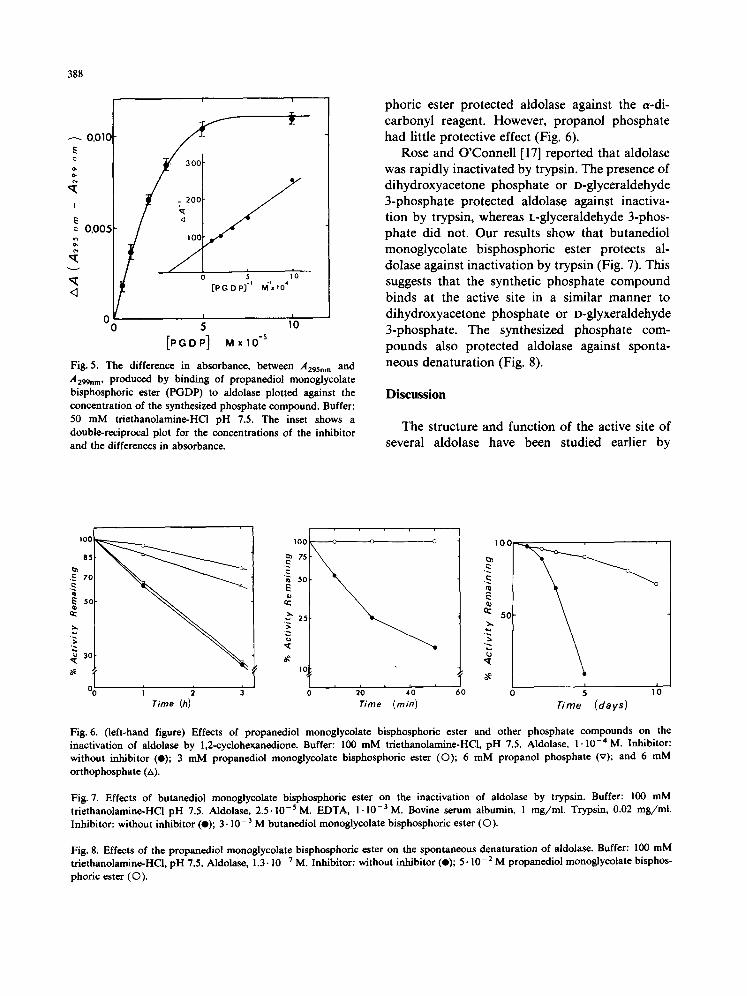
phoric ester protected aldolase against the ePdi- carbonyl reagent. However, propanol phosphate had little protective effect (Fig. 6).
Rose and O'Connell [17] reported that aldolase was rapidly inactivated by trypsin. The presence of dihydroxyacetone phosphate or D-glyceraldehyde 3-phosphate protected aldolase against inactiva- tion by trypsin, whereas L-glyceraldehyde 3-phos- phate did not. Our results show that butanediol monoglycolate bisphosphoric ester protects al- dolase against inactivation by trypsin (Fig. 7). This suggests that the synthetic phosphate compound binds at the active site in a similar manner to dihydroxyacetone phosphate or D-glyxeraldehyde 3-phosphate. The synthesized phosphate com- pounds also protected aldolase against sponta- neous denaturation (Fig. 8).
D i s c u s s i o n
The structure and function of the active site of several aldolase have been studied earlier by
100
85
.~ 7G
~ 5c Iu
' ~ 30
i 2 3 Time (h)
"! "-._ !
IC
' 210 ' 410 '
Time (rain) 60
,ooi
5o~ \
i 0 5 10
Time (days)
Fig. 6. (left-hand figure) Effects of propanediol monoglycolate bisphosphoric ester and other phosphate compounds on the inactivation of aldolase by 1,2-cyclohexancdione. Buffer: 100 mM triethanolamine-HCl, pH 7.5. Aldolase, 1.10 -4 M. Inhibitor: without inhibitor (e); 3 mM propanediol monoglycolate bisphosphoric ester (O); 6 mM propanol phosphate (v); and 6 mM orthophosphate (/,).
Fig. 7. Effects of butanedioi monoglycolate bisphosphoric ester on the inactivation of aldolase by trypsin. Buffer: 100 mM triethanolamine-HC1 pH 7.5. Aldolase, 2.5.10 -5 M. EDTA, I. 10 -3 M. Bovine serum albumin, 1 mg/ml. Trypsin, 0.02 mg/ml. Inhibitor: without inhibitor (e); 3.10 -3 M butanediol monoglycolate bisphosphoric ester (©).
Fig. 8. Effects of the propanediol monoglycolate bisphosphoric ester on the spontaneous denaturation of aldolase. Buffer: 100 mM triethanolamine-HC1, pH 7.5. Aldolase, 1.3-10 -7 M. Inhibitor: without inhibitor (e); 5.10 -2 M propanediol monoglycolate bisphos- phoric ester (©).
steady-state kinetics and the effect of inhibitors [2,5,10,11,18], and by chemical modification of the active site [1,6,7-9,17], and the amino acid se- quence has been determined [4].
The active site of aldolase has two binding site for the phosphate groups of the substrate, Fru-1,6- /)2. Hartman and Barker [10] estimated the dis- tance between the two binding sites as 10-12 ,~ using a,o~-alkanediol bisphosphate analogues of Fru-l,6-P 2. The results obtained here for the dif- ference spectra support their proposal. This sug- gests that the active sites may be relatively flexible but tend to keep the distance between two phos- phate binding sites to that of the open-chain form of Fru-l,6-P 2.
Formation of the complex between aldolase and its substrate might be stabilized by electro- static binding of the substrate phosphate groups. The presence of positively charged groups in the active site of aldolase was suggested by Castellino and Barker [19], who distinguished between highly specific binding sites for phosphate and additional sites with lower affinity for phosphate. A weaker electrostatic interaction links the C-6 phosphate group of the substrate and Lys-107 at the active site [4]. Recently, two amino acid residues were proposed for the C-1 phosphate binding site of aldolase: Arg-55 by 1,2-cyclohexanedione-labeling [9], and Lys-146 by interchain cross-link of a triose phosphate derivative with Lys-227, which forms the catalytic Schiff base intermediate [7].
Propanediol monoglycolate bisphosphoric ester is very effective in preventing inactivation by 1,2- cyclohexanedione, which selectively modifies the Arg-55 of aldolase, whereas propanol monophos- phate gives only slight protection.
One of the three tryptophan residues of the subunit of aldolase, Trp-147, is a neighbour of Lys-146, which has the role of binding site for the C-1 phosphate group of the substrate. Results of difference spectra titrations suggest that the tryp- tophan is quite sensitive to binding of the synthe- sized phosphate compounds. This implies that both amino acid residues participate in the binding of the synthesized phosphate compounds.
Aldolase has 15 arginine and 26 lysine residues in a subunit, and Arg-55 [9], Lys-107 [6], Lys-146 [7,8] and Lys-227 [3,4] are said to be components of the active site of aldolase [4]. These amino acids
389
(P)-O-CH2-C-O-CH2-CH2-CH2- O-(P ) 0
Fig. 9. 31P-NMR spectra of the propanediol monoglycolate diphosphoric ester cyclohexylammonium salt. Solvent: 2H20. Standard: orthophosphoric acid. Temperature, 25°C.
may be present on the surface of the enzyme, but could also be protected against trypsin digestion by the synthesized phosphate compounds. The protection against trypsin digestion and against spontaneous denaturation indicates that propane- diol monoglycolate bisphosphoric ester binds at the active site of aldolase and stabilizes the native conformation of the enzyme.
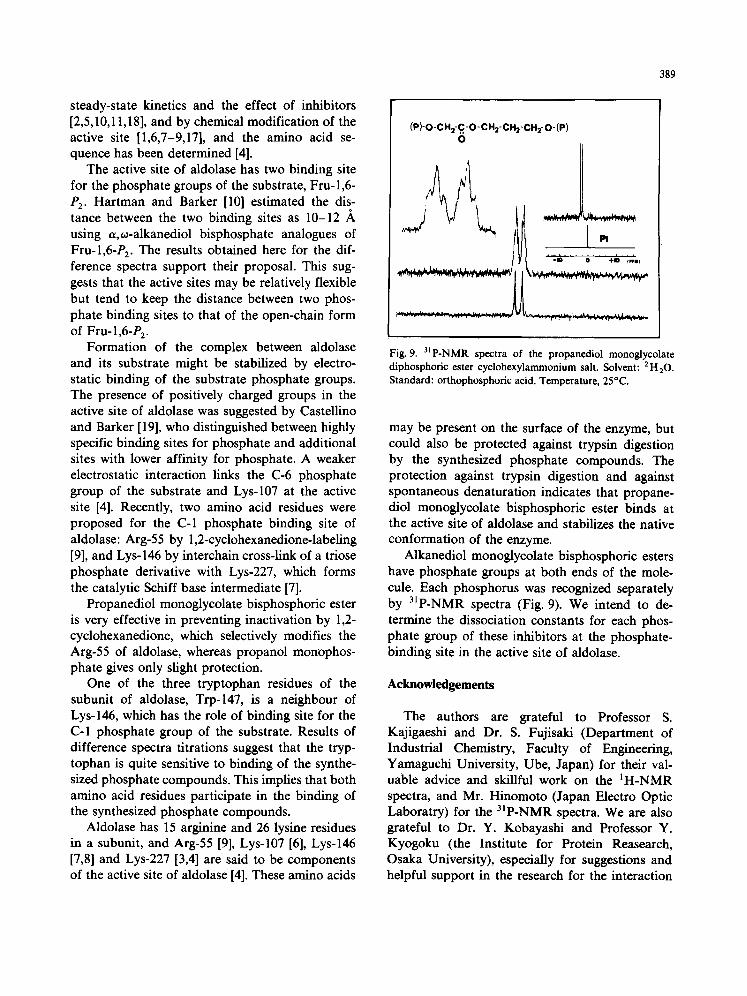
Alkanediol monoglycolate bisphosphoric esters have phosphate groups at both ends of the mole- cule. Each phosphorus was recognized separately by 31p-NMR spectra (Fig. 9). We intend to de- termine the dissociation constants for each phos- phate group of these inhibitors at the phosphate- binding site in the active site of aldolase.
Acknowledgements
The authors are grateful to Professor S. Kajigaeshi and Dr. S. Fujisaki (Department of Industrial Chemistry, Faculty of Engineering, Yamaguchi University, Ube, Japan) for their val- uable advice and skillful work on the IH-NMR spectra, and Mr. Hinomoto (Japan Electro Optic Laboratry) for the 31p-NMR spectra. We are also grateful to Dr. Y. Kobayashi and Professor Y. Kyogoku (the Institute for Protein Reasearch, Osaka University), especially for suggestions and helpful support in the research for the interaction

390
between enzyme and phosphate compound using NMR.
References
1 Mehler, A.H. and Cusic M.E., Jr. (1967) Science 155, 1101-1103
2 Mid¢lfort, C.F., Gupta, R.K. and Rose, I.A. (1976) Bio- chemistry 15, 2178-2185
3 Grazi, E., Cheng, T. and Horecker, B.L. (1962) Biochim. Biophys. Acta 7, 250-253
4 Lai, C.Y., Nakai, N. and Chang, D. (1974) Science 183, 1204-1206
5 Ginsburg, A. and Mehler, A.H. (1966) Biochemistry 5, 2623-2634
6 Shapiro, S., Enser, M., Pugh, E. and Horecker, B.L. (1968) Arch. Biochem. Biophys. 128, 554-562
7 Lubini, D.G.E. and Christen, P. (1979) Proc. Natl. Acad. Sci. U.S.A. 76, 2527-2531
8 Hartman, F.C. and Brown, J.P. (1970) J. Biol. Chem. 251, 3057-3062
9 Patthy, L., V~rani, A., Thrsz, J. and Kovhcs, K. (1979) Eur. J. Biochem. 99, 309-313
10 Hartman, F.C. and Barker, R. (1965) Biochemistry 6, 1068-1075
11 Crowder, A.L., III, Barker, R. and Swenson, C.A. (1973) Biochemistry 12, 2078-2083
12 Taylor, J.F. (1955) Methods Enzymol. !, 310-315 Academic Pres New York
13 Donovan, J.W. (1964) Biochemistry 3, 67-74 14 Kawahara, K. and Tanford, C. (1966) Biochemistry 5,
1578-1584 15 Blostein, R. and Rutter, W.J. (1963) J. Biol. Chem. 238,
3280-3285 16 Lineweaver, H. and Burk, D. (1934) J. Am. Chem. Soc. 56,
658-666 17 Rose, I.A. and O'Connell, E.L. (1969) J. Biol. Chem. 244,
126-134 18 Suh, B. and Barker, R. (1971)J. Biol. Chem. 246, 7041-7050 19 Castellino, F.J. and Barker, R. (1966) Biochem. Biophys.
Res. Commun. 23, 182-187