an overview of the higher level classification of
TRANSCRIPT

An overview of the higher level classification of Pucciniomycotina based oncombined analyses of nuclear large and small subunit rDNA sequences
M. Catherine Aime1
USDA-ARS, Systematic Botany and Mycology Lab,Beltsville, Maryland 20705
P. Brandon MathenyBiology Department, Clark University, Worcester,Massachusetts 01610
Daniel A. HenkUSDA-ARS, Systematic Botany and Mycology Lab,Beltsville, Maryland 20705
Elizabeth M. FriedersDepartment of Biology, University of Wisconsin,Platteville, Wisconsin 53818
R. Henrik NilssonGoteborg University, Department of Plant andEnvironmental Sciences, Goteborg, Sweden
Meike PiepenbringJ.W. Goethe-Universitat Frankfurt, Department ofMycology, Frankfurt, Germany
David J. McLaughlinDepartment of Plant Biology, University of Minnesota,St Paul, Minnesota 55108
Les J. SzaboUSDA-ARS, Cereal Disease Lab, University ofMinnesota, St Paul, Minnesota 55108
Dominik BegerowUniversitat Tubingen, Spezielle Botanik und Mykologie,Tubingen, Germany
Jose Paulo SampaioCREM, SABT, Faculdade de Ciencias e Tecnologia,Universidade Nova de Lisboa, 2829-516 Caparica,Portugal
Robert BauerMichael WeißFranz Oberwinkler
Universitat Tubingen, Spezielle Botanik und Mykologie,Tubingen, Germany
David HibbettBiology Department, Clark University, Worcester,Massachusetts 01610
Abstract: In this study we provide a phylogeneticallybased introduction to the classes and orders ofPucciniomycotina (5Urediniomycetes), one of three
subphyla of Basidiomycota. More than 8000 species ofPucciniomycotina have been described includingputative saprotrophs and parasites of plants, animalsand fungi. The overwhelming majority of these(,90%) belong to a single order of obligate plantpathogens, the Pucciniales (5Uredinales), or rustfungi. We have assembled a dataset of previouslypublished and newly generated sequence data fromtwo nuclear rDNA genes (large subunit and smallsubunit) including exemplars from all known majorgroups in order to test hypotheses about evolutionaryrelationships among the Pucciniomycotina. Theutility of combining nuc-lsu sequences spanning theentire D1-D3 region with complete nuc-ssu sequencesfor resolution and support of nodes is discussed. Ourstudy confirms Pucciniomycotina as a monophyleticgroup of Basidiomycota. In total our results supporteight major clades ranked as classes (Agaricostilbo-mycetes, Atractiellomycetes, Classiculomycetes, Cryp-tomycocolacomycetes, Cystobasidiomycetes, Microbo-tryomycetes, Mixiomycetes and Pucciniomycetes) and18 orders.
Key words: basidiomycetes, molecular phylo-genetics, parasitic fungi, rDNA systematics, Uredi-niomycetes
INTRODUCTION
One-third of described Basidiomycota belong tosubphylum Pucciniomycotina (5Urediniomycetessensu Swann and Taylor 1995). Roughly 7000 ofthese, or ,90%, belong to a single order, Pucciniales(5Uredinales G. Winter) or the rust fungi (Kirk et al2001). The remaining fungi within the Pucciniomy-cotina are remarkably diverse ecologically, biological-ly and physiologically; only with the relatively recentadvent of ultrastructure, biochemical and molecularsystematic studies have the relatedness of the variedfungi now placed in the Pucciniomycotina becomeapparent. These include Microbotryales, phytopatho-gens once classified in the Ustilaginomycotina (truesmuts and relatives, Ustilaginomycetes sensu Bauer etal 2001); numerous basidiomycetous yeasts formerlyallied with Auriculariales and other groups ofAgaricomycotina (mushroom-forming fungi and re-lated taxa; Hymenomycetes sensu Swann and Taylor1993); and at least one species, Mixia osmundae,formerly classified within the Ascomycota (Nishidaet al 1995).
Acepted for publication 7 September 2006.1 Corresponding author. E-mail: [email protected]
Mycologia, 98(6), 2006, pp. 896–905.# 2006 by The Mycological Society of America, Lawrence, KS 66044-8897
896

Most species of Pucciniomycotina are parasitic andthe group includes phytopathogens (e.g. Pucciniales,Microbotryales), mycoparasites (e.g. Tuberculina,Spiculogloea) and entomopathogens (e.g. Septobasi-diales). A few are putatively saprotrophic or are ofunknown trophic habit. Pucciniomycotina species arepresent in most habitats including freshwater (Baueret al 2003) and marine environments (Swann et al2001). Dimorphism, cryptic fructifications, heteroe-cism and multiple spore types all have evolved in atleast some lineages of Pucciniomycotina, hamperingthe collection of complete biological and life cycledata for many taxa. Yet species of Pucciniomycotinaare causal agents of some of the most devastatingdiseases of crops, may cause opportunistic disease inhumans and have shown potential as biologicalcontrol organisms of invasive plants and of otherpathogenic fungi (e.g. Evans 1993, Swann et al 2001).
A few Pucciniomycotina species produce basidio-carps, these typically being simple stipitate-capitate(stilboid) or resupinate (FIG. 1b) structures. SomePucciniomycotina species form hyphae (filamentous)or yeasts or both (dimorphic) (FIG. 1d). Members ofthe Pucciniales produce spore-filled fruiting struc-tures termed sori (FIG. 1f ). Basidia of teleomorphicspecies may take the form of teliospores, holobasidiaor phragmobasidia, and basidiospores may be sessile,gasteroid or forcibly discharged. The Pucciniomyco-tina is distinguished from the other two major cladesof Basidiomycota by the possession of simple septalpores that lack membrane-bound caps (Swann et al2001, Weiß et al 2004 and references therein). Inaddition the cell wall sugar composition of thePucciniomycotina differs from that in the Ustilagino-mycotina and Agaricomycotina (Prillinger et al2002).
Diverse approaches have been used to delimit taxawithin the Pucciniomycotina, including studies ofultrastructural, physiological, biochemical and eco-logical characters. Molecular characters have hada major impact on our understanding of thephylogenetic relationships of Pucciniomycotina. Asearly as 1985 Gottschalk and Blanz showed that thesecondary structure of 5S ribosomal RNA in somespecies of Pucciniomycotina differs from that of someother lineages of Basidiomycota. The pioneeringstudies of Swann and colleagues (Swann and Taylor1993, 1995; Swann et al 1999) used nuclear smallsubunit (nuc-ssu) rDNA sequences to support themonophyly of the Pucciniomycotina and a number ofmajor clades. Other studies have employed a regionof about 500–650 bp at the 59 end of the nuclear largesubunit (nuc-lsu) rDNA, containing the highly vari-able divergent domains D1 and D2 (Hassouna et al1984) providing resolution for many clades within the
Pucciniomycotina (e.g. Fell et al 2001, Weiß et al2004, Bauer et al 2006). However D1–D2 sequenceshave provided only weak support for monophyly ofPucciniomycotina (Begerow et al 1997, Berres et al1995), or have resolved the group as a paraphyleticassemblage from which the Ustilaginomycotina andAgaricomycotina have been derived (Weiß et al 2004).Nor were analyses of a third gene, that coding for b-tubulin, able to recover a monophyletic Pucciniomy-cotina (Begerow et al 2004). Because of the publisheddiscrepancies in tree topologies and support values,monophyly for the group has yet to be establishedconclusively by molecular analyses.
Current major classifications of Pucciniomycotinainclude those of Kirk et al (2001), Swann et al (2001),Weiß et al (2004) and Bauer et al (2006). The latterthree classifications have many similarities (SUPPLE-
MENTARY TABLE I). However there are also differencesbetween these systems in both resolution andcomposition of taxa. Kirk et al (2001) divide thePucciniomycotina (as Urediniomycetes) into fiveorders and do not include subclasses; thus theirclassification is quite different from the others.
The goal of the present study is to reassess thehigher level systematics of the Pucciniomycotina withcombined analyses of nuc-lsu rDNA and nuc-ssurDNA sequences. Two other studies have includeda simultaneous analysis of nuc-lsu and nuc-ssu rDNAsequences of Pucciniomycotina (Lutzoni et al 2004,Bauer et al 2006). However the former was focused onrelationships across the Fungi and it included only 14species of Pucciniomycotina, of which 10 weremembers of the Pucciniales and the combinedanalysis in the latter employed only 25 species ofPucciniomycotina and did not cover all majorlineages. In the present study we compile and analyzedatasets containing new and previously publishedsequences that represent 174 species from all majorgroups of Pucciniomycotina. We assess the ability ofdifferent rDNA genes and gene regions to recoverand support each clade, and we provide an overviewof the classes and orders of Pucciniomycotina.
MATERIALS AND METHODS
This study uses both newly generated sequences andsequences that were retrieved from GenBank from morethan 80 studies (SUPPLEMENTARY TABLE II). Some Pucci-niales sequences were generated with the methods de-scribed in Aime (2006); sequences of Jola spp. weregenerated as in Frieders (1997). Descriptions of laboratorymethods pertaining to the majority of new sequencesproduced at Clark University can be found online (SUPPLE-
MENTARY APPENDIX 1).The entire data matrix contains 208 operational taxo-
nomic units (OTUs) including six Ascomycota that were
AIME ET AL: PUCCINIOMYCOTINA OVERVIEW 897

FIG. 1. Representatives of the Pucciniomycotina. a. Jola cf. javensis (Platygloeales) fruiting on Sematophyllum swartzii. b.Septobasidium burtii (Septobasidiales) fungal mat completely covering scale insects. c. Eocronartium muscicola (Platygloeales)fruiting on undetermined moss. d. Yeast and filamentous cells of Sporidiobolus pararoseus (Sporidiobolales). e. Cultures of twoSporidiobolus species in the S. pararoseus clade (Sporidiobolales). f. Phragmidium sp. (Pucciniales) on Rosa rubiginosa.
898 MYCOLOGIA

used for rooting purposes, nine Ustilaginomycotina, fourAgaricomycotina and 189 Pucciniomycotina that represent174 species (26 species of Pucciniomycotina are representedby two OTUs and one species is represented by threeOTUs). One OTU, Septobasidium sp., is represented bya nuc-lsu rDNA sequence deposited as Septobasidium sp. anda nuc-ssu rDNA sequence deposited as Septobasidiumcanescens. All other OTUs include sequences from the sameputative species and where possible from the samecollection or isolate (SUPPLEMENTARY TABLE II). Alignmentsare available from the first and/or last author on request.We performed analyses using two sets of OTUs. Dataset 1contains the 128 OTUs that have both nuc-lsu and nuc-ssurDNA, including all the Ascomycota, Ustilaginomycotinaand Agaricomycotina, and 109 Pucciniomycotina thatinclude representatives of all the major groups in theclassifications of Swann et al (2001), Weiß et al (2004) andBauer et al (2006) except the Cryptomycocolacomycetes.Dataset 2 contains all 208 OTUs, 74 of which have only nuc-lsu rDNA, including Cryptomycocolax abnormis (Cryptomy-cocolacomycetes), and six OTUs that have only the nuc-ssurDNA. To compare the phylogenetic utility of the short(D1–D2 region only) versus long (D1–D3 region, seeSUPPLEMENTARY APPENDIX 1) nuc-lsu rDNA sequences, aswell as various combinations of data, we assembled a thirddataset (Dataset 3) that contains 20 Pucciniomycotina andtwo other Basidiomycota that have nuc-lsu rDNA sequencesof at least 1117 bp and nuc-ssu rDNA sequences of at least1667 bp (indicated by an asterisk in SUPPLEMENTARY TABLE
II). Datasets were analyzed with equally weighted maximumparsimony (MP); Dataset 1 also was analyzed with neighborjoining (NJ) and Bayesian methods. (Details of analyticalmethods are provided online in SUPPLEMENTARY APPENDIX
1.)
RESULTS AND DISCUSSION
MP analyses of Dataset 1 recovered 36 trees of 8215steps (CI 5 0.345, RI 5 0.731), one of which isdepicted (FIG. 2, an expanded version is available inonline supplementary materials). MP analyses ofDataset 2 recovered 1000 trees of 9748 steps (CI 5
0.310, RI 5 0.729); the strict consensus tree forDataset 2 is presented (SUPPLEMENTARY FIG. 3).Detailed results and discussion of analyses for allthree datasets are provided online (SUPPLEMENTARY
APPENDIX 2).This work brings together data from more than 80
studies along with newly generated data to providethe first multigene molecular analyses for all majorclades of Pucciniomycotina. The present study strong-ly supports monophyly of the Pucciniomycotina(FIG. 2). We recover eight major clades containing18 subclades (SUPPLEMENTARY FIG. 3), which mostclosely correspond to the classification of Bauer et al(2006). All higher taxa of Pucciniomycotina recog-nized in the present study are supported by bootstrapvalues of at least 90% in one or more of our analyses
and have Bayesian posterior probabilities of at least0.98 with the exception of the Sporidiobolales,Agaricostilbomycetes including Spiculogloeales, andthe Cryptomycocolacales which is represented by onlya single nuc-lsu sequence (SUPPLEMENTARY FIG. 3).
The names of higher taxa used here follow theclassification formalized in Bauer et al (2006). Theclassification of Bauer et al (2006) is different fromearlier classifications (SUPPLEMENTARY TABLE I) in thatmore groups are named and recognized as formaltaxa (e.g. Cystobasidiomycetes); groups formerlyranked as subclasses were elevated to the rank ofclass (e.g. Microbotryomycetes); and higher levelnames prefixed ‘‘Uredinio-’’ or ‘‘Uredin-’’ have beenreplaced by names based on Puccinia, the largestgenus within the Pucciniomycotina (Kirk et al 2001).
Many of the genera of Pucciniomycotina are notmonophyletic, notably anamorphic yeasts such asRhodotorula and Sporobolomyces (Weiß et al 2004,FIG. 2). One of the obvious obstacles to diagnosePucciniomycotina taxa is the homoplasious nature ofthe micromorphological characters available (Bauerand Oberwinkler 1991a, Bauer et al 2006). Ultrastruc-tural characters, for example, structure of spindlepole body or the presence of colacosomes in manyMicrobotryomycetes (Swann et al 1999) appearpromising for delimiting natural groups. However atpresent the major obstacle to applying meaningfuldiagnoses to these groups is the dearth of ultrastruc-tural, ecological and life-cycle data for most Pucci-niomycotina members.
Descriptions of higher level taxa.—Pucciniomycetes. Thisis the most speciose class of Pucciniomycotina. Wehave resolved four orders (Helicobasidiales, Platy-gloeales s. str., Pucciniales and Septobasidiales). Afifth order, Pachnocybales (Bauer et al 2006), isrepresented by a single species, Pachnocybe ferruginea(SUPPLEMENTARY FIG. 3) although molecular evidencefrom other studies places this taxon within theSeptobasidiales (Berres et al 1995, Frieders 1997,Henk and Vilgalys 2006). With the exception of P.ferruginea, which as a holobasidia-producing sapro-troph (Kropp and Corden 1986, Bauer and Ober-winkler 1990) holds an isolated position within theclass, and Platygloea disciformis, which may besaprotrophic, all known members are parasitic andproduce phragmobasidia or teliospores.
The members of the Helicobasidiales producea dikaryophase (Helicobasidium) that is parasitic onplant roots and a haplophase (Tuberculina) that isparasitic on rust fungi (Lutz et al 2004b). Species ofTuberculina are capable only of infecting rusts in thehaploid stage of development (Thirumalachar 1941)and the host-specificity of some species (Lutz et al
AIME ET AL: PUCCINIOMYCOTINA OVERVIEW 899

FIG. 2. Phylogenetic relationships in the Pucciniomycotina based on analyses of Dataset 1 (128 OTUs, each with nuc-lsurDNA and nuc-ssu rDNA sequences). Phylogram representing one of 36 equally most parsimonious trees (8215 steps, CI 5
0.345, RI 5 0.731). Nodes with asterisks collapse in the strict consensus of equally most parsimonious trees. Support valuesinclude MP bootstrap frequencies above 70% (first value, before slash), NJ bootstrap frequencies above 70% (second value)
900 MYCOLOGIA

2004a) might make them candidates for biocontrol.Lutz et al (2004c) discuss the confused taxonomichistory of Tuberculina. The position of H. corticioidescould not be confirmed with nuc-ssu data alone(SUPPLEMENTARY FIG. 3).
Until relatively recently all basidiomycetes withtransversely septate basidia (termed auricularioid)were classified in the rusts, smuts (Ustilaginales) orAuriculariales (Agaricomycotina). Moore (1990) cre-ated the Platygloeales by removing those species thatproduced simple (rather than dolipore) septal poresfrom the Auriculariales. The order now is consideredto contain mostly phytoparasitic species in the generaPlatygloea, Insolibasidium, Herpobasidium, Eocronar-tium and Jola. These last two genera include speciesthat are parasitic on mosses, produce conidia anddevelop basidiomata which, in the case of Eocronar-tium, are clavarioid fruiting bodies that may reach upto 15 mm tall (Atkinson 1902, Frieders and McLaugh-lin 2001) (FIG. 1a, c). Fitzpatrick (1918) providesdescriptions and illustrations of the life cycle,cytology, host-range and taxonomy of E. muscicola,and the host-parasite interaction is described inBoehm and McLaughlin (1988). The columnar typeof fructification (FIG. 1c) similar to that of the rustgenus Cronartium led Atkinson (1902) to hypothesizea close relationship between Eocronartium and thePucciniales, which is supported by molecular analyses(FIG. 2). The unsupported association of Platygloeapustulata with the Septobasidiales (SUPPLEMENTARY
FIG. 3) might be an artifact of our supermatrixapproach (see SUPPLEMENTARY APPENDIX 2) for whichadditional gene sampling might be necessary toresolve.
Species of Septobasidiales are unique in that theyare parasites of scale insects. This is the second largestorder of Pucciniomycotina, with more than 170described species (Kirk et al 2001), nearly all ofwhich belong to the genus Septobasidium. Theprincipal reference is Couch’s 1938 publication‘‘The Genus Septobasidium’’. These fungi are foundas mats of hyphae covering infected scale insects onplants (FIG. 1b). The life cycle (SUPPLEMENTARY FIG. 4)of Septobasidium has been relatively well characterizedand includes the only known occurrence of a yeaststage in the Pucciniomycetes. Members of the orderlive primarily as dikaryons and undergo meiosis
within probasidia (Couch 1938, Olive 1943). Auricu-loscypha, Coccidiodictyon, Uredinella and Ordonia arethe other genera included in the Septobasidiales(Oberwinkler 1989, Henk and Vilgalys 2006).
Pucciniales species are obligate parasites of vascularplants (FIG. 1f) and have developed the mostcomplex life cycles of any Fungi (Cummins andHiratsuka 2003) (SUPPLEMENTARY FIG. 5). Rusts arephenotypically and genetically plastic organisms withmany characters that set them apart from otherBasidiomycota including the development of spermo-gonia, heteroecism and the occurrence of up to sixdifferent spore types (Laundon 1973, Hennen andBuritica 1980, Cummins and Hiratsuka 2003). Ap-proximately 115 (Cummins and Hiratsuka 2003) to163 (Kirk et al 2001) genera of Pucciniales currentlyare recognized and family classification remainscontentious (Hennen and Buritica 1980, Ono andHennen 1983, Kirk et al 2001). Three recent studiesof molecular data from nuc-lsu (Maier et al 2003),nuc-ssu (Wingfield et al 2004) and combined nucrDNA genes (Aime 2006) have provided confirmationfor some families (e.g. Melampsoraceae) and high-lighted others requiring revision (e.g. Pucciniaceae).The origins of the Pucciniales and their relationshipto other Basidiomycota have been a ripe field forconjecture. Many early hypotheses of basidiomyceteevolution consider Pucciniales as the most ‘‘primi-tive’’ Basidiomycota from which all other lineages arederived (e.g. Linder 1940, Savile 1955). Our resultsconfirm other molecular studies (Prillinger et al 2002,Lutzoni et al 2004) that show the Pucciniales asderived from lineages that include insect and non-vascular plant parasites (FIG. 2).
Cystobasidiomycetes.—Our data resolve two orders ofCystobasidiomycetes. A third order, Naohideales, hasbeen proposed (Bauer et al 2006) but it is hererepresented solely by nuc-lsu sequences of Naohideasebacea, a mycoparasite (SUPPLEMENTARY FIG. 3).Three mostly monotypic taxa, Sakaguchia (isolatedfrom marine habitats), Cyrenella and Bannoa, cannotbe placed into orders with our data. The Cystobasi-diales include both anamorphic yeasts and dimorphicyeasts that produce clamp connections and auricular-ioid basidia without basidiocarps. Genera included inthis order are Occultifur (O. internus is a mycoparasite
rand Bayesian posterior probabilities above 0.70 (below branch). Outgroup taxa are condensed into a single branch forAscomycota (six taxa represented); Agaricomycotina (four taxa represented); Ustilaginomycotina (nine taxa represented).Duplicate sequences have been collapsed into a single branch, which are indicated in parentheses after the taxon name, e.g.Occultifer externus (2). The small cladogram depicts the topology of Microbotryomycetidae obtained in Bayesian analyses, withposterior probabilities indicated only for strongly supported nodes that conflict with the MP topology. (An expanded versionof FIG. 2 is presented in the online supplementary materials).
AIME ET AL: PUCCINIOMYCOTINA OVERVIEW 901

of Dacrymycetales, Bauer et al 2006), Cystobasidium(which like Occultifur produces tremelloid haustorialcells that are indicative of mycoparasitism) andsome Rhodotorula spp. (anamorphic yeasts). TheErythrobasidiales include many yeasts currentlyplaced in Rhodotorula pro parte (p.p.) and Sporobo-lomyces p.p., as well as the monotypic mitosporic yeastErythrobasidium. Other studies (Sampaio 2004, Baueret al 2006) support the inclusion of Bannoa within theErythrobasidiales. No teleomorph has been found inthis order.
Atractiellomycetes.—The Atractiellomycetes containa single order, Atractiellales, originally erected toaccommodate gasteroid auricularioid species withsimple septa formerly placed in the Auriculariales(Oberwinkler and Bandoni 1982). Walker (1984)and Gottschalk and Blanz (1985) recognized theartificiality of this group, which, as supported by ourdata, is limited to include the genera Atractiella,Saccoblastia, Hobsonia, Pleurocolla, Helicogloea, Platy-gloea p.p. and Phleogena. No yeast state is known forthese fungi (Swann et al 2001) but some may produceconidia. Atractiella and Phleogena produce stilboidfruitbodies. All members of this class studied thus farpossess unique ultrastructural organelles of unknownfunction termed symplechosomes (Oberwinkler andBauer 1989, McLaughlin 1990, Bauer and Oberwink-ler 1991b).
Agaricostilbomycetes.—Our data strongly supporta monophyletic Agaricostilbales that is supported assister of the Spiculogloeales in Bayesian analyses(FIG. 2). Blanz and Gottschalk (1986) found thatstructure of the 5S rRNA in Agaricostilbum pulcherri-mum, a small gasteroid fungus found worldwide onpalms, was unique from all examined basidiomycetesand posited that this taxon held an isolated positionwithin the heterobasidiomycetes. Agaricostilbum isalso unusual in that the nuclei divide in the parentcell rather than in the bud cell (Frieders andMcLaughlin 1996), and a similar mitotic pattern wasfound in the closely related Stilbum vulgare but not inthe more distantly related (SUPPLEMENTARY FIG. 3)yeast Bensingtonia yuccicola (McLaughlin et al 2004).The yeast phase of the life cycle of A. pulcherrimum isillustrated in Frieders and McLaughlin (1996). Othergenera placed in the Agaricostilbales include Ben-singtonia, Kondoa, Sterigmatomyces, Sporobolomycesp.p., Chionosphaera, Stilbum, Mycogloea p.p. andKurtzmanomyces. These include gasteroid yeasts withauricularioid basidia (e.g. Agaricostilbum hyphaenes),gasteroid yeasts with holobasidia (e.g. Chionosphaeraapobasidialis), and teliospore-forming yeasts (e.g.Kondoa malvinella). Chionosphaera species produceclamps and microscopic stipitate-capitate fruiting
structures and are mycophilic, associated with lichensand beetle gallery fungi (Kirschner et al 2001a).
The order Spiculogloeales has been placed withinthe Agaricostilbomycetes by Bauer et al (2006) andcontains Mycogloea p.p., Spiculogloea and someanamorphic Sporobolomyces spp. Characters sharedby the teleomorphic species of this order includedimorphism, mycoparasitism and tremelloid hausto-rial cells subtended by clamp connections (Bandoni1998). Mycogloea does not appear monophyletic withthese data (SUPPLEMENTARY FIG. 3) and additionalmolecular (combined nuc-lsu and nuc-ssu sequencescould not be obtained for any Spiculogloea orMycogloea species for this study) and taxonomicsampling, including the type species of Mycogloea,are needed to resolve their placement.
Microbotryomycetes.—This group includes mycopara-sites, phytopathogens and putative saprotrophs witha diversity of micromorphological characters. Manyspecies contain organelles, termed colacosomes, thatare associated with mycoparasitism (Bauer andOberwinkler 1991a). Four orders are recognized:Heterogastridiales (Oberwinkler et al 1990b), whichare mycoparasites and include the aquatic genusHeterogastridium; Leucosporidiales (Sampaio et al2003), which includes the teliospore-forming yeastLeucosporidium and two other genera, Mastigobasi-dium and Leucosporidiella; the Sporidiobolales (Sam-paio et al 2003), including the genera Rhodotorulap.p., Sporidiobolus (FIG. 1d, e), Sporobolomyces p.p. andRhodosporidium (a teliospore-forming yeast illustratedin SUPPLEMENTARY FIG. 6); and the Microbotryales(Bauer et al 1997). Members of Microbotryales areteliospore-forming phytopathogens that once wereclassified within the Ustilaginomycotina and includethe genera Bauerago, Fulvisporium, Aurantiosporium,Rhodotorula p.p., Ustilentyloma, Liroa, Microbotryumand Sphacelotheca. Microbotryum violaceum has be-come an important model organism for co-evolution-ary and population genetics studies between patho-gens and their hosts (e.g. Antonovics et al 2002).
Additional taxa that belong to this class but couldnot be assigned to an order are Colacogloea (amonotypic, mycoparasitic genus that possesses colaco-somes, Oberwinkler et al 1990a), Kriegeria (a mono-typic sedge parasite with an anamorphic yeast stageand unique multiperforate septa, Doubles andMcLaughlin 1991, Swann et al 1999), Camptobasidium(a monotypic aquatic hyphomycete associated withdecaying leaf litter, Marvanova and Suberkropp1990), Rhodotorula p.p., Sporobolomyces p.p., Bensing-tonia yamatoana and Leucosporidium antarcticum.
Classiculomycetes.—This class contains a single order,the Classiculales, containing aquatic fungi of fresh-
902 MYCOLOGIA

water habitats (Bauer et al 2003). Classicula fluitans isthe teleomorph of the hyphomycete Naiadellafluitans and produces auricularioid basidia and clampconnections. Jaculispora submersa is an aquatic hy-phomycete for which no teleomorph has beendiscovered. Classicula is unique in forming subapi-cally swollen sterigmata (Bauer et al 2003). Classiculafluitans forms tremelloid haustorial cells that wereobserved to occasionally self-parasitize in culture,which might indicate a mycoparasitic habit for thisfungus (Bauer et al 2003).
Cryptomycocolacomycetes.—This class contains a singleorder, Cryptomycocolacales, represented by the ge-nus Cryptomycocolax, a mycoparasitic fungus thatinfects ascomycete sclerotia (Oberwinkler and Bauer1990) and has been placed in the Ustilaginomycotina(Kirk et al 2001). Our study confirms the placementof this clade within the Pucciniomycotina (SUPPLE-
MENTARY FIG. 3). Cryptomycocolacales species are theonly fungi outside the Microbotryomycetes thatpossess colacosomes, which might indicate a sistergroup relationship between the two. Cryptomycocolaxhas a unique mode of basidium development that onthe other hand might be indicative of a basal positionwithin the Pucciniomycotina (Oberwinkler and Bauer1990). Transmission electron microscopy studies ofanother genus of colacosome-producing mycopara-sites, Colacosiphon, indicate that it belongs in theCryptomycocolacales (Kirschner et al 2001b). Bothgenera are monotypic.
Mixiomycetes.—This class, containing the Mixiales, ismonotypic containing a single fern parasite, Mixiaosmundae. Until recently M. osmundae was classifiedwithin the Ascomycota (Nishida et al 1995) andremains isolated within the Pucciniomycotina with noknown close relatives (Swann et al 1999, 2001; FIG. 2).Mixia osmundae is a blastosporic yeast, although it hasnot been established whether it produces meiosporesor mitospores. The hyphae are multinucleate contain-ing few septa (Kramer 1958, Nishida et al 1995, Swannet al 2001).
Incertae sedis.—Reniforma strues was isolated frombiofilms in a wastewater treatment plant in WestVirginia (Pore and Sorenson 1990). It is a yeast that,unlike any other known basidiomycete, produceskidney-shaped (reniform) somatic cells. Its classifica-tion within the Basidiomycota is confirmed with thesedata, yet its position within the Pucciniomycotinacould not be fully resolved with nuc-lsu data alone(SUPPLEMENTARY FIG. 3).
Concluding remarks.—While our results provide thefirst support for many of the higher level clades ofPucciniomycotina formalized in the classification of
Bauer et al (2006), many nodes, especially along thebackbone, still are not resolved. Future molecularsystematic studies aimed at filling in missing datain Dataset 2 and targeting additional gene regionsfor exemplar taxa from all the groups recovered bythis study should be the next steps toward recoveringa robust phylogenetic hypothesis for these fungi. Ad-ditional taxonomic sampling might benecessary before a fully resolved phylogeny for thePucciniomycotina is attainable. However, excludingthe Pucciniales, we have sampled 54 genera, or nearlythe entire known generic diversity in the subphylum(Kirk et al 2001). The Pucciniales is often consideredone of the best documented groups of fungi. Yetrecent estimates indicate that only 10–30% of theirdiversity in the neotropics alone has been discovered(Hennen and McCain 1993). Many genera ofPucciniomycotina have been discovered and de-scribed only since the 1990s, often from specialtyniches (e.g. beetle galleries, biofilms) or neotropicalhabitats (e.g. Oberwinkler and Bauer 1990; Ober-winkler et al 1990a, 1990b; Pore and Sorenson 1990;Kirschner et al 2001a, 2001b; Sampaio et al 2003).Furthermore many of these are representative oflineages that are monotypic or problematic withregard to phylogenetic resolution (e.g. Reniformastrues). This would indicate that a tremendousamount of diversity still awaits discovery within thisubiquitous group of fungi.
ACKNOWLEDGMENTS
The authors thank Conrad Schoch and CBS for providingseveral cultures and Pat Crane for Pucciniales material usedfor the generation of sequence data. We thank Rytas Vilgalysand Lisa Bukovnik for access to the DNA sequencing facilityat Duke University, Cindy Park for technical support atSBML and Manfred Binder for assistance with Bayesiananalyses. This work was supported in part by grants from theNational Science Foundation, DEB 0228657, DBI 0320875and DEB-0090301, Research Coordination Network: APhylogeny for Kingdom Fungi to M. Blackwell, J.W.Spatafora and J.W. Taylor. Mention of trade names orcommercial products in this publication is solely for thepurpose of providing specific information and does notimply recommendation or endorsement by the U.S. De-partment of Agriculture.
LITERATURE CITED
Aime MC. 2006. Toward resolving family-level relationshipsin rust fungi (Uredinales). Mycoscience 47:112–122.
Antonovics J, Hood ME, Partain J. 2002. The ecology andgenetics of a host-shift: Microbotryum as a model system.Am Nat 160:S40–S53.
Atkinson GF. 1902. Preliminary note on two new genera ofbasidiomycetes. J Mycol 8:106–107.
AIME ET AL: PUCCINIOMYCOTINA OVERVIEW 903

Bandoni RJ. 1998. On some species of Mycogloea. My-coscience 39:31–36.
Bauer R, Begerow D, Oberwinkler F, Marvanova L. 2003.Classicula: the teleomorph of Naiadella fluitans.Mycologia 95:756–764.
———, ———, ———, Piepenbring M, Berbee ML. 2001.Ustilaginomycetes. In: McLaughlin DJ, McLaughlinEG, Lemke PA, eds. Systematics and evolution. Berlin:Springer, The Mycota VII, Part B: 57–83.
———, ———, Sampaio JP, Weiß M, Oberwinkler F. 2006.The simple-septate basidiomycetes: a synopsis. MycolProg 5:41–66.
———, Oberwinkler F. 1990. Meiosis, spindle pole bodycycle and taxonomy of the heterobasidiomycetePachnocybe ferruginea. Plant Syst Evol 172:241–261.
———, ———. 1991a. The colacosomes: new structures atthe host-parasite interface of a mycoparasitic basidio-mycete. Bot Acta 104:53–57.
———, ———. 1991b. The symplechosome: a unique cellorganelle of some basidiomycetes. Bot Acta 104:93–97.
———, ———, Vanky K. 1997. Ultrastructural markers andsystematics in smut fungi and allied taxa. Can J Bot 75:1273–1314.
Begerow D, Bauer R, Oberwinkler F. 1997. Phylogeneticstudies on nuclear LSU rDNA sequences of smut fungiand related taxa. Can J Bot 75:2045–2056.
———, John B, Oberwinkler F. 2004. Evolutionary relation-ships among b-tubulin gene sequences of basidiomyce-tous fungi. Mycol Res 108:1257–1263.
Berres ME, Szabo LJ, McLaughlin DJ. 1995. Phylogeneticrelationships in auriculariaceous basidiomycetes basedon 25S ribosomal DNA sequences. Mycologia 87:821–840.
Blanz PA, Gottschalk M. 1986. Systematic position ofSeptobasidium, Graphiola and other basidiomycetes asdeduced on the basis of their 5S ribosomal RNAnucleotide sequences. Sys Appl Microbiol 8:121–127.
Boehm EWA, McLaughlin DJ. 1988. Eocronartium muscicola:a basidiomycetous moss parasite exploiting gameto-phytic transfer cells. Can J Bot 66:762–770.
Couch JN. 1938. The genus Septobasidium. Chapel Hill,North Carolina: UNC Press.
Cummins GB, Hiratsuka Y. 2003. Illustrated genera of rustfungi. 3rd ed. St Paul, Minesota: American Phytopath-ological Society. 225 p.
Doubles JC, McLaughlin DJ. 1991. A new basidiomycetousseptal type: the multiperforate septum in Kriegeriaeriophori. Am J Bot 78:1542–1548.
Evans H. 1993. Studies on the rust Maravalia cryptostegiae,a potential biological control agent of rubber-vine weed(Cryptostegia grandiflora, Asclepiadaceae: Periplocoi-deae) in Australia, I: Life-cycle. Mycopathologia 124:163–174.
Fell JW, Boekhout T, Fonseca A, Sampaio JP. 2001.Basidiomycetous yeasts. In: McLaughlin DJ, McLaugh-lin EG, Lemke PA, eds. Systematics and evolution.Berlin: Springer-Verlag, The Mycota VII Part B: 3–35.
Fitzpatrick HM. 1918. The life history and parasitism ofEocronartium muscicola. Phytopathology 8:197–218.
Frieders EM. 1997. An integrated approach to understand-
ing the moss parasites and their role in basidiomyceteevolution [Doctoral thesis]. St Paul: University ofMinnesota. 185 p.
———, McLaughlin DJ. 2001. The heterobasidiomycetemoss parasites Jola and Eocronartium in culture:cytology, ultrastructure and anamorph. Mycol Res105:734–744.
———, ———. 1996. Mitosis in the yeast phase ofAgaricostilbum pulcherrimum and its evolutionary sig-nificance. Can J Bot 74:1392–1406.
Gottschalk M, Blanz PA. 1985. Untersuchungen an 5Sribosomalen Ribonukleinsauren als Beitrag zurKlarung von Systematik und Phylogenie der Basidio-myceten. Z Mykol 51:205–243.
Hassouna N, Michot B, Bachellerie J. 1984. The completenucleotide sequence of mouse 28S rRNA. Implicationsfor the process of size increase of the large subunitrRNA in higher eukaryotes. Nucl Acid Res 12:3563–3583.
Henk DA, Vilgalys R. 2006. Molecular phylogeny suggestsa single origin of insect symbiosis in the Urediniomy-cetidae with support for some relationships within thegenus Septobasidium. Am J Bot. (Accepted for publica-tion).
Hennen JF, Buritica P. 1980. A brief summary of modernrust taxonomic and evolutionary theory. Rept TottoriMycol Inst 18:243–256.
———, McCain JW. 1993. New species and records ofUredinales from the Neotropics. Mycologia 85:970–986.
Kirk PM, Cannon PF, David JC, Stalpers JA. 2001. Dictionaryof the fungi. 9th ed. Wallingford, UK: CABI. 655 p.
Kirschner R, Begerow D, Oberwinkler F. 2001a. A newChionosphaera species associated with conifer inhabit-ing bark beetles. Mycol Res 105:1403–1408.
———, Bauer R, Oberwinkler F. 2001b. Colacosiphon: a newgenus described for a mycoparasitic fungus. Mycologia93:634–644.
Kramer CL. 1958. A new genus in the Protomycetaceae.Mycologia 50:916–926.
Kropp BR, Corden ME. 1986. Morphology and taxonomy ofPachnocybe ferruginea. Mycologia 78:334–342.
Laundon GF. 1973. Uredinales. In: Ainsworth CG, SparrowFK, Sussman AS, eds. The fungi. New York: AcademicPress, IVB:247–279.
Linder DH. 1940. Evolution of the basidiomycetes and itsrelation to the terminology of the basidium. Mycologia32:419–447.
Lutz M, Bauer R, Begerow D, Oberwinkler F. 2004a.Tuberculina-Helicobasidium: host specificity of theTuberculina-stage reveals unexpected diversity withinthe group. Mycologia 96:1316–1329.
———, ———, ———, ———. 2004b. Tuberculina-Thana-tophytum/Rhizoctonia crocorum-Helicobasidium: a un-ique mycoparasitic-phytoparasitic life strategy. MycolRes 108:227–238.
———, ———, ———, ———, Triebel D. 2004c. Tubercu-lina: rust relatives attack rusts. Mycologia 96:614–626.
Lutzoni F, Kauff F, Cox CJ, 41 others. 2004. Assembling the
904 MYCOLOGIA

Fungal Tree of Life: progress, classification andevolution of subcellular traits. Am J Bot 91:1446–1480.
Maier W, Begerow D, Weiß M, Oberwinkler F. 2003.Phylogeny of the rust fungi: an approach using nuclearlarge subunit ribosomal DNA sequences. Can J Bot 81:12–23.
Marvanova L, Suberkropp K. 1990. Camptobasidium hydro-philum and its anamorph, Crucella subtilis: a newheterobasidiomycete from streams. Mycologia 82:208–217.
McLaughlin DJ. 1990. A new cytoplasmic structure in thebasidiomycete Helicogloea: the microscala. Expl Mycol14:331–338.
———, Hanson Jr RW, Frieders EM, Swann EC, Szabo LJ.2004. Mitosis in the yeast phase of the basidiomycetesBensingtonia yuccicola and Stilbum vulgare and itsphylogenetic implications. Am J Bot 91:808–815.
Moore RT. 1990. Order Platygloeales ord. nov. Mycotaxon39:245–248.
Nishida H, Ando K, Ando Y, Hirata A, Sugiyama J. 1995.Mixia osmundae: transfer from the Ascomycota to theBasidiomycota based on evidence from molecules andmorphology. Can J Bot 73(1):S660–S666.
Oberwinkler F. 1989. Coccidiodictyon, new genus andOrdonia, two genera in the Septobasidiales. Op Bot100:185–192.
———, Bandoni RJ. 1982. A taxonomic survey of thegasteroid, auricularioid Heterobasidiomycetes. CanJ Bot 60:1726–1750.
———, Bauer R. 1989. The systematics of gasteroid,auricularioid Heterobasidiomycetes. Sydowia 41:224–256.
———, ———. 1990. Cryptomycocolax: a new mycoparasiticheterobasidiomycete. Mycologia 82:671–692.
———, ———, Bandoni RJ. 1990a. Colacogloea: a newgenus in the auricularioid Heterobasidiomycetes.Can J Bot 68:2531–2536.
———, ———, ———. 1990b. Heterogastridiales: a neworder of Basidiomycetes. Mycologia 82:48–58.
Olive LS. 1943. Cytology of various basidial types in thegenus Septobasidium. Mycologia 35:557–572.
Ono Y, Hennen JF. 1983. Taxonomy of the chaconiaceousgenera (Uredinales). Trans Mycol Soc Jap 24:369–402.
Pore RS, Sorenson WG. 1990. Reniforma strues, a new yeastfrom wastewater. Mycologia 82:549–553.
Prillinger H, Lopandic K, Schweigkofler W, Deak R, AartsHJM, Bauer R, Sterflinger K, Kraus GF, Maraz A. 2002.Phylogeny and systematics of the fungi with specialreference to the Ascomycota and Basidiomycota. In:Breitenbach M, Crameri R, Lehrer SB, eds., Fungalallergy and pathogenicity. Chem Immunol vol 81. 207–295.
Sampaio JP. 2004. Diversity, phylogeny and classification ofbasidiomycetous yeasts. In: Agerer R, Piepenbring M,Blanz P, eds. Frontiers in Basidiomycote mycology.Eching, Germany: IHW-Verlag. p 49–80.
———, Gadanho M, Bauer R, Weiß M. 2003. Taxonomicstudies in the Microbotryomycetidae: Leucosporidiumgolubevii sp. nov., Leucosporidiella gen. nov. and thenew orders Leucosporidiales and Sporidiobolales.Mycol Prog 2:53–68.
Savile DBO. 1955. A phylogeny of the Basidiomycetes.Can J Bot 33:60–104.
Swann EC, Frieders EM, McLaughlin DJ. 1999. Microbo-tryum, Kriegeria and the changing paradigm in basid-iomycete classification. Mycologia 91:51–66.
———, ———, ———. 2001. Urediniomycetes. In:McLaughlin DJ, McLaughlin EG, Lemke PA, eds.Systematics and evolution. Berlin: Springer-Verlag,The Mycota VII Part B: 37–56.
———, Taylor JW. 1993. Higher taxa of basidiomycetes: an18S rRNA gene perspective. Mycologia 85:923–936.
———, ———. 1995. Phylogenetic perspectives on basidio-mycete systematics: evidence from the 18S rRNA gene.Can J Bot 73:S862–S868.
Thirumalachar MJ. 1941. Tuberculina on Uromyces hobsoniiVize. J Indian Bot Soc 20:107–110.
Walker WF. 1984. 5S rRNA sequences from Atractiellales,and basidiomycetous yeasts and fungi imperfecti. SysAppl Microbiol 5:352–359.
Weiß M, Bauer R, Begerow D. 2004. Spotlights onheterobasidiomycetes. In: Agerer R, Piepenbring M,Blanz P, eds. Frontiers in Basidiomycote mycology.Eching, Germany: IHW-Verlag. p 7–48.
Wingfield BD, Ericson L, Szaro T, Burdon JJ. 2004.Phylogenetic patterns in the Uredinales. AustralasPlant Pathol 33:327–335.
AIME ET AL: PUCCINIOMYCOTINA OVERVIEW 905

APPENDIX 1
MATERIAL AND METHODS
DNA extractions, PCR, and sequencing.—New sequenceswere obtained from live or lyophilized cultures, herbariummaterial and fresh collections. DNA was isolated with theEZNA Fungal DNA Kit or EZNA Forensic DNA Kit (OmegaBio-tek, Doraville, Georgia) following the manufacturer’sprotocols. Primer sequences for nuc-ssu are from White et al(1990) and O’Donnell et al (1998) and can be found onlineat http://aftol.org/. Primer pairs PNS1/NS41 and NS19b/NS8 were used to generate two nuc-ssu rDNA products,sequences of which overlap and yield a total product size ofabout 1780 bp. On some occasions NS51 was paired withITS2 to amplify the 39 end of the gene when the primercombination NS19b/NS8 was not optimal. Additionalprimers for sequencing included NS19bc, NS51 and NS6.Due to the presence of multiple introns only partial nuc-ssurDNA sequences were obtained for Colacogloea peniophorae(sample No. 3) and Entyloma holwayi. Only the 18S regionbetween PNS1/NS41 (,1200 bp) at the 59 end of the genewas obtained for Gymnosporangium juniperi-virginianae,Kondoa malvinella, Kuehneola uredinis, Malassezia pachy-dermatis, Sporisorium formosanum and Tilletia controversa.Nuc-lsu rDNA sequences of approximately 1400 bp wereamplified with primers LR0R and LR7 and sequenced withthese primers and LR3R, LR16 and LR5 (Vilgalys andHester 1990; http://www.biology.duke.edu/fungi/mycolab/primers.htm). This region includes the first three divergentdomains (D1–D3) of the large subunit rDNA (Hassouna et al1984, Hopple and Vilgalys 1999) as opposed to mostpublished Pucciniomycotina nuc-lsu rDNA sequences, whichconsist of the first ,600 bp of the region containing thefirst two divergent domains (D1–D2). In several instances,especially for taxa of the Pucciniales, LR16 was not a suitablesequencing primer. For Pileolaria toxicodendri individualnuc-lsu rDNA amplicons were cloned with the TOPO TAcloning kit (Invitrogen, Carlsbad, California) but in all othercases the PCR products were sequenced directly. DNAsequencing reactions were performed with ABI Prism dyeterminator chemistry and were run on an ABI 377XL or 3700automated DNA sequencer (the latter at the DNA sequenc-ing facility of the Duke University Biology Department, R.Vilgalys laboratory).
Assessing conflict.—The nuc-lsu and nuc-ssu rDNA arelinked in the nuclear rDNA repeat, which reduces thechances of phylogenetic conflicts among the loci due tohybridization, lineage sorting or horizontal gene trans-mission. Nevertheless the majority of operational taxonomicunits (OTUs) combine sequences obtained from differentstudies (SUPPLEMENTARY TABLE II) so misidentifications,laboratory errors or differing species concepts could besources of incongruence. To assess conflict we performed1000 replicate bootstrapped neighbor joining (NJ) analysesof the nuc-lsu and nuc-ssu rDNA partitions independently,using PAUP* 4.0b12 (Swofford 2002) with Kimura two-
parameter distances. OTUs with differing placements thatreceived at least 70% NJ bootstrap support in both datapartitions were considered to demonstrate conflict. Poten-tial conflicts were detected between the nuc-lsu and nuc-ssurDNA partitions in Filobasidiella neoformans, Melampsoraeuphorbiae and Mycogloea sp. (trees not shown). One of theconflicting nodes involving F. neoformans was stronglysupported only marginally (bootstrap 5 73%) so the datafor that taxon were combined. The data for M. euphorbiaeand Mycogloea sp. were not combined and only nuc-lsu datawere used for these OTUs in Dataset 2.
Data analyses.—Datasets 1 and 2 were analyzed with equallyweighted maximum parsimony (MP) and bootstrapped MP.Dataset 1 also was analyzed with neighbor joining (NJ) andBayesian methods. MP analyses were implemented inPAUP* with a two-step search strategy consisting of aninitial search with 1000 heuristic searches (each witha starting tree generated by taxon addition with a randomsequence, TBR branch swapping, and keeping up to twotrees per replicate) followed by a second heuristic searchusing the shortest trees found in the initial search as startingtrees, with TBR branch swapping and MAXTREES set to 1000.Bootstrapped MP analyses used 1000 replicates, each withone heuristic search with a starting tree generated byrandom taxon addition, TBR branch swapping, and keepingup to 10 trees per replicate.
Modeltest 3.7 (Posada and Crandall 1998) was used toevaluate models of molecular evolution for use in NJ andBayesian analyses. Optimal models were identified for thenuc-ssu rDNA alone, nuc-lsu rDNA alone and the combineddataset. For each data partition the optimal model accord-ing to the Akaike information criterion was the general timereversible model, with a proportion of invariable sites andamong-site rate heterogeneity modeled with a discretegamma distribution (i.e. the GTR+G+I model). Howeverhierarchical likelihood ratio tests selected the TIM+I+Gmodel for the nuc-ssu rDNA alone and the combineddataset and the TrN+I+G model for the nuc-lsu rDNA datapartition.
NJ and bootstrapped NJ analyses (with 1000 replicates)were implemented in PAUP* with the BIONJ algorithm(Gascuel 1997) with maximum likelihood distances thatwere obtained using the GTR+I+G model. Model parame-ters were fixed based on values obtained from Modeltest, asfollows: base frequencies A 5 0.2498, C 5 0.2045, G 5
0.2772, T 5 0.2685; substitution rates A«C 5 1.1857, A«G5 2.4999, A«T 5 1.5885, C«G 5 1.0903, C«T 5 4.7743,G«T 5 1.0000; proportion of invariable sites 5 0.2680;gamma distribution shape parameter 5 0.5577 (with fourrate classes).
Bayesian analyses were performed using MrBayes 3.0(Ronquist and Huelsenbeck 2003) with the GTR+I+Gmodel with all parameter values allowed to vary. Modelparameters for the nuc-lsu rDNA and nuc-ssu rDNApartitions obtained with Modeltest were similar (values not

shown), so the data were not partitioned. Three indepen-dent Metropolis-coupled Markov chain Monte Carlo anal-yses were performed, each with four chains, three of whichwere ‘‘heated’’ (with default temperature settings). Eachanalysis was run 1 3 106 generations, with trees sampledevery 100 generations. A total of 8500 trees were uploadedfrom each run after a burn-in of the first 1501 trees.Posterior probabilities were calculated from the 25 500 treesproduced by each of the three independent runs.
To assess the phylogenetic resolution and supportafforded by different rDNA regions we performed two setsof partitioned analyses. The first set of partitioned analysesused MP, bootstrapped MP and bootstrapped NJ analyses ofthe nuc-lsu rDNA and nuc-ssu rDNA data partitions ofDataset 1, with the same analytical settings as the combinednuc-lsu/nuc-ssu rDNA analysis. The nuc-lsu rDNA datapartition in Dataset 1 contains many sequences that includeonly the D1–D2 region, as well as sequences up to 1778 bp(SUPPLEMENTARY TABLE II). To compare the phylogeneticutility of the short (D1–D2) versus long (D1–D3) nuc-lsurDNA sequences, as well as various combinations of data, weassembled a third dataset (Dataset 3) that contains 21species of Pucciniomycotina and two other Basidiomycotathat have nuc-lsu rDNA sequences of at least 1117 bp andnuc-ssu rDNA sequences of at least 1667 bp (OTUs selectedfor Dataset 3 are indicated by an asterisk in SUPPLEMENTARY
TABLE II). We performed analyses of Dataset 3 with sixdifferent data partitions, including: (i) The first approxi-mately 650 bp of nuc-lsu rDNA containing the D1–D2region, with the same set of included positions used inanalyses of Datasets 1 and 2; (ii) The D1–D2 region of nuc-lsu rDNA but including two hypervariable regions that wereexcluded in analyses of Datasets 1 and 2 (the hypervariableregions were aligned with Clustal X 1.83 [Thompson et al1997] with default settings); (iii) Longer D1–D3 nuc-lsurDNA sequences (excluding the hypervariable regions); (iv)Complete nuc-ssu rDNA sequences; (v) Combined D1–D2and nuc-ssu rDNA sequences (excluding the hypervariableregions of nuc-lsu rDNA); and (vi) combined D1-D3 nuc-lsurDNA and nuc-ssu rDNA sequences (excluding the hyper-variable regions of nuc-lsu rDNA).
Each data partition was analyzed with MP, bootstrappedMP and bootstrapped NJ. Analytical settings were the sameas described above, except that the MP analysis used a single-step heuristic search with 1000 replicates, each witha starting tree generated with a random taxon addition
sequence, TBR branch swapping, and MAXTREES set toautoincrease.
LITERATURE CITED
Gascuel O. 1997. BIONJ: an improved version of the NJalgorithm based on a simple model of sequence data.Mol Biol Evol 14:685–695.
Hassouna N, Michot B, Bachellerie J. 1984. The completenucleotide sequence of mouse 28S rRNA. Implicationsfor the process of size increase of the large subunitrRNA in higher eukaryotes. Nucleic Acid Res 12:3563–3583.
Hopple JS, Vilgalys R. 1999. Phylogenetic relationships inthe mushroom genus Coprinus and dark-spored alliesbased on sequence data from the nuclear gene codingfor the large ribosomal subunit RNA: divergentdomains, outgroups and monophyly. Mol PhylogenetEvol 13:1–19.
O’Donnell K, Cigelnik E, Benny GL. 1998. Phylogeneticrelationships among the Harpellales and Kickxellales.Mycologia 90:624–639.
Posada D, Crandall KA. 1998. Modeltest: testing the modelof DNA substitution. Bioinformatics 14:817–818.
Ronquist F, Huelsenbeck JP. 2003. MrBayes 3: Bayesianphylogenetic inference under mixed models. Bioinfor-matics 19:1572–1574.
Swofford DL. 2002. PAUP*: phylogenetic analysis usingparsimony (*and other methods). Version 4. Sunder-land, Massachusetts: Sinauer Associates.
Thompson JD, Gibson TBJ, Plewniak F, Jeanmougin F,Higgins DG. 1997. The Clustal X windows interface:flexible strategies for multiple sequence alignmentaided by quality analysis tools. Nucleic Acid Res 25:4876–4882.
Vilgalys R, Hester M. 1990. Rapid genetic identification andmapping of enzymatically amplified ribosomal DNAfrom several Cryptococcus species. J Bacteriol 172:4238–4246.
White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification anddirect sequencing of fungal ribosomal RNA genes forphylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ,White TJ, eds. PCR protocols: a guide to methods andapplications. San Diego: Academic Press. p 315–322.

APPENDIX 2
RESULTS AND DISCUSSION
The newly generated nuc-lsu rDNA sequences rangefrom 917 to 1778 bp (SUPPLEMENTARY TABLE II).Seventy percent (140) of the nuc-lsu rDNA sequencesin the complete dataset (Dataset 2) are between 501and 648 bp, reflecting the prevalence of studies thatsolely use the region containing the highly variabledivergent domains D1 and D2 (Hassouna et al 1984).The newly generated nuc-ssu rDNA sequences rangefrom 829 to 2146 bp. Eighty percent (107) of the nuc-ssu rDNA sequences in the Dataset 2 are more than1600 bp (SUPPLEMENTARY TABLE II) and containdivergent domains D1–D3. Introns ranging from255 to 416 bp are present in the nuc-lsu rDNAsequence of Entyloma holwayi and in the nuc-ssurDNA sequences of Bensingtonia ciliata, Colacogloeapeniophorae, Entyloma holwayi and Ustilago maydis.The aligned length of the complete nuc-lsu rDNAsequences is 2069 bp. The analyzed region of nuc-lsurDNA (excluding introns and regions that weconsidered to be of ambiguous alignment) contains1357 positions, of which 699 are variable and 550(40.5%) are parsimony informative (in the context ofDataset 1). The nuc-ssu rDNA has an aligned lengthof 3736 positions. Excluding introns and regions ofambiguous alignment the nuc-ssu rDNA contains1843 positions, including 1007 variable positionsand 710 (38.5%) parsimony informative positions(in the context of Dataset 1; SUPPLEMENTARY TABLE
III).
Analyses of Dataset 1.—MP analyses of Dataset 1recovered 36 trees of 8215 steps (CI 5 0.345, RI 5
0.731; SUPPLEMENTARY TABLE III). Thirty-one of themost parsimonious trees were found in 125 of the1000 heuristic searches in the first part of the analysis.The strict consensus of all equally most parsimonioustrees is highly resolved but there is a large polytomy atthe base of the Microbotryomycetes (FIG. 2). ThePucciniomycotina was supported as monophyleticwith 100% MP and NJ bootstrap support andposterior probability of 1.0 (FIG. 2). The Atractiello-mycetes received MP bootstrap support of 80%, butthe other five classes for which there were at least twoOTUs each received MP bootstrap support of 100%
(FIG. 2 and SUPPLEMENTARY TABLE IV). All six classeswith more than two OTUs received NJ bootstrapsupport of at least 96% and posterior probabilities of1.0. In total 52 nodes in the Pucciniomycotinareceived MP bootstrap support greater than 90%, 62nodes received NJ bootstrap support greater than
90% and 100 nodes received Bayesian posteriorprobabilities greater than 0.95 (FIG. 2). However thebasal nodes in the ‘‘backbone’’ of the Pucciniomyco-tina are weakly supported (FIG. 2).
MP analyses of Dataset 1 using only the nuc-lsurDNA recovered 1000 trees of 3799 steps (CI 5 0.324,RI 5 0.695); 33 nodes received MP bootstrap supportexceeding 90% and 27 nodes received NJ bootstrapsupport exceeding 90% (results summarized inSUPPLEMENTARY TABLE III). MP analyses of Dataset 1using only the nuc-ssu rDNA recovered 1000 trees of4136 steps (CI 5 0.373, RI 5 0.766); 44 nodesreceived MP bootstrap support exceding 90% and 50nodes received NJ bootstrap support exceeding 90%
(SUPPLEMENTARY TABLE III).
Analyses of Dataset 2.—MP analyses of Dataset 2recovered 1000 trees of 9748 steps (CI 5 0.310, RI 5
0.729). Four trees were recovered in two of the 1000heuristic searches in the first part of the analysis.Despite the large number of equally parsimonioustrees the strict consensus is highly resolved (SUPPLE-
MENTARY FIG. 3). For the most part the same majorgroups that are strongly supported in analyses ofDataset 1 were resolved in analyses of Dataset 2, butMP bootstrap support values in analyses of Dataset 2are generally lower than those in analyses of Dataset 1(SUPPLEMENTARY TABLE IV). For example, MP boot-strap support for the Pucciniomycotina dropped from100% (Dataset 1) to 83% (Dataset 2) and support forthe Microbotryomycetes dropped from 100% to 74%
(SUPPLEMENTARY TABLE IV).
Analyses of Dataset 3.—Dataset 3 was analyzed with sixdifferent data partitions (SUPPLEMENTARY TABLE III).The D1–D2 region alone had the worst performance,with only four (MP) or five (NJ) nodes receivingbootstrap support of 90% or greater. The Clustal X-aligned hypervariable subregions of the D1–D2 re-gion had an aligned length of 153 bp, with 97 variablepositions and 81 (53%) parsimony informative posi-tions. Addition of these data to the more conservedpart of the D1–D2 dataset resulted in seven nodesreceiving bootstrap support of at least 90%. Indepen-dent analyses of the longer (D1–D3) nuc-lsu rDNAand nuc-ssu rDNA regions alone yielded similarresults, with 8–10 nodes receiving at least 90%
bootstrap support. Combined analyses of D1–D2 orlonger nuc-lsu sequences and nuc-ssu rDNA se-quences yielded 10 or 11 nodes supported by at least90% of the bootstrap replicates.

Combined vs. independent analyses of nuc-lsu rDNAand nuc-ssu rDNA.—Combined analyses of nuc-lsurDNA and nuc-ssu rDNA sequences provide strongersupport for the phylogenetic relationships of Pucci-niomycotina than is attained through analyses ofeither region alone (SUPPLEMENTARY TABLES III andIV). In analyses of Dataset 1 (with 128 OTUs)combined analyses recovered about twice as manystrongly supported nodes as analyses of nuc-lsu rDNAalone, and about 1.2–1.4 times as many stronglysupported nodes as analyses of nuc-ssu rDNA alone.Combined analyses of Dataset 1 also provide greaterresolution than the single-gene analyses; both single-partition analyses of Dataset 1 recovered 1000 equallymost parsimonious trees (this was the MAXTREES
setting, so there are probably many more equallyparsimonious trees), whereas the combined analysisrecovered only 36 trees.
The nuc-lsu rDNA data partition in Dataset 1includes taxa with a range of nuc-lsu sequence lengths(SUPPLEMENTARY TABLE II). Analyses of Dataset 3 allowassessment of the impact of the longer nuc-lsu rDNAsequences which incorporate divergent domains D1–D3 compared to shorter nuc-lsu sequences coveringonly D1–D2, as well as the resolving power of differentcombinations of data (SUPPLEMENTARY TABLE III). TheD1-D2 region is the smallest data partition analyzed,and not surprisingly it provides the weakest resolu-tion. Inclusion of the hypervariable parts of the D1–D2 region increases the number of strongly sup-ported nodes (SUPPLEMENTARY TABLE III) but theextreme variability of these regions also increases thedanger of making incorrect hypotheses of homologywhen aligning the sequences. The strongest resultswere obtained in the combined analyses of the nuc-lsuand nuc-ssu rDNA sequences. There was essentially nodifference in the strength of support in analyses thatcombined nuc-ssu rDNA sequences with the D1–D2regions vs. those that combined nuc-ssu rDNAsequences with extended-length nuc-lsu rDNA se-quences (SUPPLEMENTARY TABLE III). Nevertheless theextended-length nuc-lsu rDNA sequences have about1.4 times as many variable and parsimony-informative
sites as the D1–D2 region alone, and the extended-length nuc-lsu rDNA provides stronger resolutionthan the D1–D2 region when those regions areanalyzed independently (SUPPLEMENTARY TABLE III).
Analyses of Datasets 1 and 3 indicate that combiningnuc-lsu rDNA and nuc-ssu rDNA sequences improvesphylogenetic resolution and support and also that theutility of D1–D2 sequences can be enhanced signifi-cantly by extending them to include the completeLR0R-LR7 fragment covering D1–D3. Another strategyfor building on the existing data would be to combinethem with a supermatrix approach. Such approaches,where taxon sampling is extensive but not all taxa havebeen sampled at all loci resulting in a large amount ofmissing data, nonetheless have shown potential forphylogenetic reconstruction in large datasets (Driskellet al 2004). Analyses of Dataset 2 (in which 74 OTUshave only nuc-lsu rDNA sequences and six OTUs haveonly nuc-ssu rDNA sequences) resolve the same majorclades as analyses of Dataset 1 (FIG. 2 and SUPPLEMEN-
TARY FIG. 3), but many more equally most parsimoni-ous trees were derived from Dataset 2 than wereobtained in analyses of Dataset 1 and the bootstrapvalues are lower in Dataset 2 (FIG. 2, SUPPLEMENTARY
FIG. 3, and SUPPLEMENTARY TABLE IV). Thus super-matrix analyses might provide a useful means toconstruct comprehensive phylogenetic trees that in-corporate the large database of D1–D2 sequences, butthey probably will not provide the resolution andsupport that could be attained through combinedanalyses that have no missing data.
LITERATURE CITED
Driskell AC, Ane C, Burleigh JG, McMahon MM, O’MearaBC, Sanderson MJ. 2004. Prospects for building thetree of life from large sequence databases. Science 306:1172–1174.
Hassouna N, Michot B, Bachellerie J. 1984. The completenucleotide sequence of mouse 28S rRNA. Implicationsfor the process of size increase of the large subunitrRNA in higher eukaryotes. Nucleic Acid Res 12:3563–3583.

LITERATURE CITED
Bauer R, Begerow D, Sampaio JP, Weiß M, Oberwinkler F.2006. The simple-septate basidiomycetes: a synopsis.Mycol Progress 5:41–66.
Swann EC, Frieders EM, McLaughlin DJ. 2001. Uredinio-mycetes. In: McLaughlin DJ, McLaughlin EG, Lemke
PA, eds. Systematics and Evolution. Berlin: Springer-Verlag, The Mycota VIIB: 37–56.
Weiß M, Bauer R, Begerow D. 2004. Spotlights on
heterobasidiomycetes. In: Agerer R, Piepenbring M,
Blanz P, eds. Frontiers in Basidiomycote Mycology.
Eching, Germany: IHW-Verlag. p 7–48.
SUPPLEMENTARY TABLE I. Comparison of classifications of higher taxa of Pucciniomycotina. Exact compositions of taxa varyacross classification. Major lineages in each classification are indicated by boldface.
Swann et al (2001) Weiß et al (2004) Bauer et al (2006)
UREDINIOMYCETES UREDINIOMYCETES PUCCINIOMYCOTINA
Urediniomycetidae Urediniomycetidae PucciniomycetesUredinales Uredinales PuccinialesHelicobasidium Helicobasidium subgroup HelicobasidialesPlatygloeales s. str. Platygloeales PlatygloealesSeptobasidiales Septobasidiales Septobasidiales
Pachnocybaceaea Pachnocybaceae PachnocybalesErythrobasidiumb clade Cystobasidiaceae lineage CystobasidiomycetesCystobasidiumc Cystobasidiaceae group Cystobasidiales
Erythrobasidium Erythrobasidium subgroup ErythrobasidialesNaohidea Naohidea subgroup Naohideales
Atractiellales Atractiellales lineage AtractiellomycetesAttractiellales
Agaricostilbomycetidae Agaricostilbomycetidae AgaricostilbomycetesAgaricostilbales Agaricostilbales AgaricostilbalesMycogloea Mycogloea group Spiculogloeales
Microbotryomycetidae Microbotryomycetidae MicrobotryomycetesHeterogastridiales Heterogastridiales HeterogastridialesSporidiobolaceae p.p. Leucosporidiales LeucosporidialesMicrobotryales Microbotryales MicrobotryalesSporidiobolaceae p.p. Sporidiobolales SporidiobolalesCamptobasidiaceae Camptobasidiaceae group incertae sedisd
Cryptomycocolacales Cryptomycocolacales CryptomycocolacomycetesCryptomycocolacales
Naiadellac Classiculales lineage ClassiculomycetesClassiculales
Mixiaceae Mixiaceae lineage MixiomycetesMixiales
a Placed within Septobasidiales.b Full name: Erythrobasidium, Naohidea, Sakaguchia clade.c Placed incertae sedis within Urediniomycetes.d Placed within Microbotryomycetes.

SUP
PL
EM
EN
TA
RY
TA
BL
EII
I.R
eso
luti
on
and
sup
po
rtaf
ford
edb
yd
iffe
ren
td
ata
par
titi
on
su
sin
gD
atas
ets
1an
d3
Dat
aset
1(1
28O
TU
s)D
atas
et3
(22
OT
Us)
nu
c-ls
ulo
ng
(D1-
D3)
nu
c-ss
uco
mp
lete
nu
c-ss
uco
mp
lete
+n
uc-
lsu
lon
gn
uc-
lsu
sho
rt(D
1-D
2)
nu
c-ls
ush
ort
+h
yper
-var
iab
lere
gio
ns
nu
c-ls
ulo
ng
nu
c-ss
uco
mp
lete
nu
c-ss
uco
mp
lete
+n
uc-
lsu
sho
rt
nu
c-ss
uco
mp
lete
+n
uc-
lsu
lon
g
Ali
gned
len
gth
2069
3736
5805
840
993
2069
3736
4576
5805
Incl
ud
edp
osi
tio
ns
1357
1843
3200
557
700
1357
1843
2400
3200
Var
iab
lep
osi
tio
ns
699
1007
1706
271
368
521
567
838
1088
Info
rmat
ive
po
siti
on
s55
0(4
0%)
710
(38%
)12
60(4
0%)
197
(35%
)27
8(4
0%)
369
(27%
)35
2(1
9%)
549
(23%
)72
1(2
2%)
No
.M
Ptr
ees
1000
1000
361
13
61
1M
Ptr
eele
ngt
h37
9941
3682
1577
511
8613
5112
4720
3926
16C
I0.
324
0.37
30.
345
0.52
80.
515
0.55
40.
621
0.58
10.
583
no
des
wit
hM
Pb
s.
90%
3344
624
78
911
11n
od
esw
ith
MP
bs
.70
%58
6575
1212
1310
1415
no
des
wit
hN
Jb
s.
90%
2750
685
710
1010
10n
od
esw
ith
NJ
bs
.70
%44
6984
1213
1614
1717

SUP
PL
EM
EN
TA
RY
TA
BL
EIV
.Su
pp
ort
for
maj
or
gro
up
so
fP
ucc
inio
myc
oti
na
inth
isan
do
ther
stu
die
s.K
eyto
sup
po
rtva
lues
:b
s5
bo
ots
trap
%;
MP
5m
axim
um
par
sim
on
y;N
J5
nei
ghb
or
join
ing;
ML
5m
axim
um
like
lih
oo
d;
ME
5m
inim
um
evo
luti
on
;B
pp
5B
ayes
ian
po
ster
ior
pro
bab
ilit
y;–
5gr
ou
pis
no
tre
solv
edas
mo
no
ph
ylet
icin
op
tim
alo
rco
nse
nsu
str
ee(s
);N
A5
no
tap
pli
cab
leb
ecau
segr
ou
pw
asn
ot
sam
ple
do
ris
mo
no
typ
ic.
Ifth
ed
elim
itat
ion
of
the
gro
up
inth
eci
ted
wo
rkco
nfl
icts
wit
hth
atin
the
pre
sen
tst
ud
y,th
esu
pp
ort
valu
esin
this
tab
leco
rres
po
nd
toth
egr
ou
pas
del
imit
edh
ere,
exce
pt
asn
ote
d.
Th
isst
ud
yD
atas
et1
Th
isst
ud
yD
atas
et2
Oth
erst
ud
ies
No
.O
TU
sM
Pb
sN
Jbs
Bp
pN
o.
OT
Us
MP
bs
Stu
dy
Dat
aN
o.
OT
Us
Sup
po
rt1
PU
CC
INIO
MY
CO
TIN
A10
910
010
01.
0018
983
Bau
eret
al(2
006)
lsu
87–
NJb
sB
auer
etal
(200
6)ls
u+
ssu
2510
0N
Jbs
Wei
ßet
al(2
004)
lsu
42–
Bp
p–
NJb
sSw
ann
etal
(199
9)ss
u25
100
MP
bs
100
NJb
sL
utz
on
iet
al(2
004)
lsu
+ss
u15
1.00
Bp
p10
0N
Jbs
Ber
res
etal
(199
5)ls
u9
37M
Pb
s65
ML
bs
–M
Eb
sM
cLau
ghli
net
al(1
995,
fig
2)m
orp
h7
–M
PP
ucc
inio
myc
etes
1910
010
01.
0041
86B
auer
etal
(200
6)ls
u16
99N
Jbs
Bau
eret
al(2
006)
lsu
+ss
u3
100
NJ
Wei
ßet
al(2
004)
lsu
121.
00B
pp
92N
Jbs
Sam
pai
o(2
004,
fig.
1)ls
u12
1.00
Bp
pSw
ann
etal
(199
9)ss
u4
100
MP
bs
100
NJb
sL
utz
on
iet
al(2
004)
lsu
+ss
u11
1.00
Bp
p10
0N
Jbs
Sep
tob
asid
iale
s1
NA
NA
NA
449
Hel
ico
bas
idia
les
296
981.
0010
87P
laty
glo
eale
s4
100
100
1.00
899
Wei
ßet
al(2
004)
lsu
31.
00B
pp
99N
Jbs
Pu
ccin
iale
s12
100
100
1.00
1688
Wei
ßet
al(2
004)
lsu
61.
00B
pp
99N
Jbs
Win
gfie
ldet
al(2
004)
ssu
72,
50M
Pb
sP
ach
no
cyb
ales
0N
AN
AN
A1
NA

Th
isst
ud
yD
atas
et1
Th
isst
ud
yD
atas
et2
Oth
erst
ud
ies
No
.O
TU
sM
Pb
sN
Jbs
Bp
pN
o.
OT
Us
MP
bs
Stu
dy
Dat
aN
o.
OT
Us
Sup
po
rt1
Cys
tob
asid
iom
ycet
es27
100
961.
0033
90B
auer
etal
(200
6)ls
u10
90N
Jbs
Bau
eret
al(2
006)
lsu
+ss
u2
100
NJ
Wei
ßet
al(2
004)
lsu
50.
74B
pp
–N
Jbs
Sam
pai
o(2
004,
fig.
1)ls
u11
0.92
Bp
pSa
mp
aio
(200
4,fi
g.2)
lsu
260.
98B
pp
Fel
let
al(2
001)
lsu
1998
MP
bs
Cys
tob
asid
iale
s12
100
100
1.00
1298
Wei
ßet
al(2
004)
lsu
20.
96B
pp
95N
Jbs
Sam
pai
o(2
004,
fig.
2)ls
u8
1.00
Bp
pF
ell
etal
(200
1)ls
u5
100
MP
bs
Ery
thro
bas
idia
les
1483
911.
0015
43Sa
mp
aio
(200
4,fi
g.2)
lsu
181.
00B
pp
Fel
let
al(2
001)
lsu
1369
MP
bs
Nao
hid
eale
s1
NA
NA
NA
298
Aga
rico
stil
bo
myc
etes
25–
–1.
0041
–B
auer
etal
(200
6)ls
u15
–N
Jbs
Bau
eret
al(2
006)
lsu
+ss
u4
89N
JA
gari
cost
ilb
ales
2210
010
01.
0034
98W
eiß
etal
(200
4)ls
u3
0.94
Bp
p,
50N
Jbs
Sam
pai
o(2
004,
fig.
1)ls
u7
1.00
Bp
pSa
mp
aio
(200
4,fi
g.2)
lsu
231.
00B
pp
Swan
net
al(1
999)
ssu
310
0M
Pb
s10
0N
Jbs
Fel
let
al(2
001)
lsu
2464
MP
bs
Spic
ulo
glo
eale
s3
100
100
1.00
774
Bau
eret
al(2
006)
lsu
3–
NJb
sB
auer
etal
(200
6)ls
u+
ssu
290
NJb
ps
Wei
ßet
al(2
004)
lsu
21.
00B
pp
55N
Jbs
Sam
pai
o(2
004,
fig.
2)ls
u3
1.00
Bp
pF
ell
etal
(200
1)ls
u3
100
MP
bs
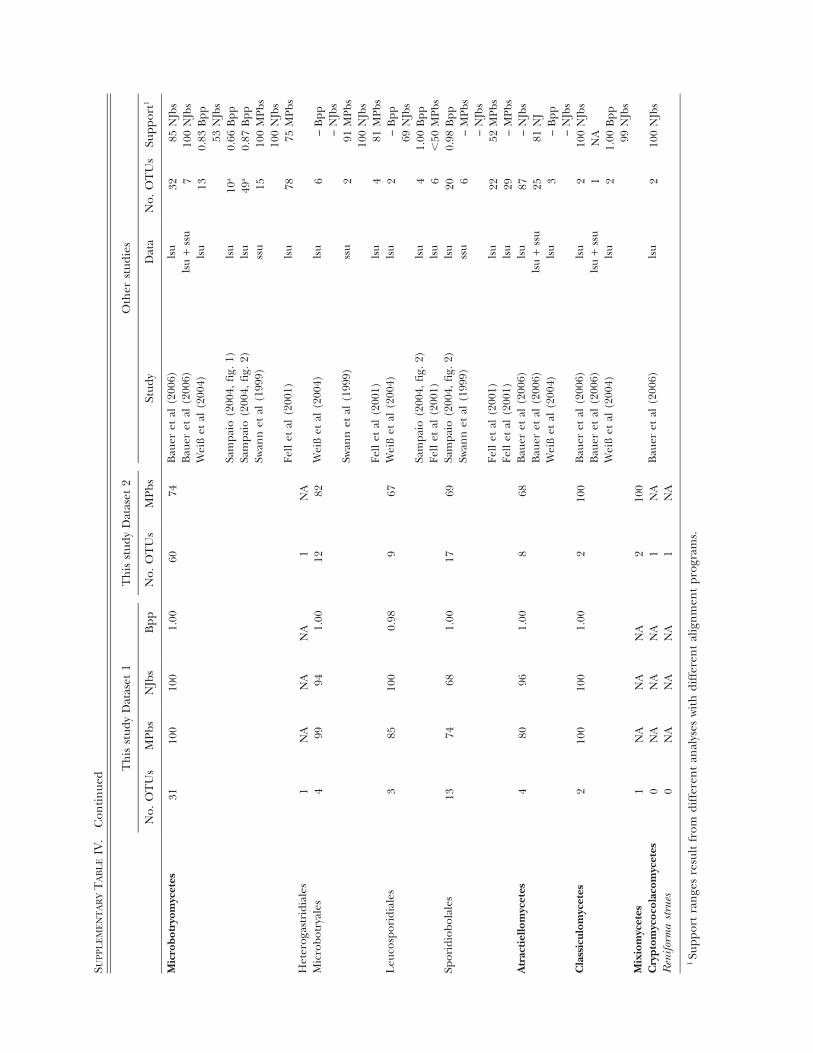
SUP
PL
EM
EN
TA
RY
TA
BL
EIV
.C
on
tin
ued
Th
isst
ud
yD
atas
et1
Th
isst
ud
yD
atas
et2
Oth
erst
ud
ies
No
.O
TU
sM
Pb
sN
Jbs
Bp
pN
o.
OT
Us
MP
bs
Stu
dy
Dat
aN
o.
OT
Us
Sup
po
rt1
Mic
rob
otr
yom
ycet
es31
100
100
1.00
6074
Bau
eret
al(2
006)
lsu
3285
NJb
sB
auer
etal
(200
6)ls
u+
ssu
710
0N
Jbs
Wei
ßet
al(2
004)
lsu
130.
83B
pp
53N
Jbs
Sam
pai
o(2
004,
fig.
1)ls
u10
a0.
66B
pp
Sam
pai
o(2
004,
fig.
2)ls
u49
a0.
87B
pp
Swan
net
al(1
999)
ssu
1510
0M
Pb
s10
0N
Jbs
Fel
let
al(2
001)
lsu
7875
MP
bs
Het
ero
gast
rid
iale
s1
NA
NA
NA
1N
AM
icro
bo
trya
les
499
941.
0012
82W
eiß
etal
(200
4)ls
u6
–B
pp
–N
Jbs
Swan
net
al(1
999)
ssu
291
MP
bs
100
NJb
sF
ell
etal
(200
1)ls
u4
81M
Pb
sL
euco
spo
rid
iale
s3
8510
00.
989
67W
eiß
etal
(200
4)ls
u2
–B
pp
69N
Jbs
Sam
pai
o(2
004,
fig.
2)ls
u4
1.00
Bp
pF
ell
etal
(200
1)ls
u6
,50
MP
bs
Spo
rid
iob
ola
les
1374
681.
0017
69Sa
mp
aio
(200
4,fi
g.2)
lsu
200.
98B
pp
Swan
net
al(1
999)
ssu
6–
MP
bs
–N
Jbs
Fel
let
al(2
001)
lsu
2252
MP
bs
Fel
let
al(2
001)
lsu
29–
MP
bs
Atr
acti
ello
myc
etes
480
961.
008
68B
auer
etal
(200
6)ls
u87
–N
Jbs
Bau
eret
al(2
006)
lsu
+ss
u25
81N
JW
eiß
etal
(200
4)ls
u3
–B
pp
–N
Jbs
Cla
ssic
ulo
myc
etes
210
010
01.
002
100
Bau
eret
al(2
006)
lsu
210
0N
Jbs
Bau
eret
al(2
006)
lsu
+ss
u1
NA
Wei
ßet
al(2
004)
lsu
21.
00B
pp
99N
Jbs
Mix
iom
ycet
es1
NA
NA
NA
210
0C
ryp
tom
yco
cola
com
ycet
es0
NA
NA
NA
1N
AB
auer
etal
(200
6)ls
u2
100
NJb
sR
enif
orm
ast
rues
0N
AN
AN
A1
NA
1Su
pp
ort
ran
ges
resu
ltfr
om
dif
fere
nt
anal
yses
wit
hd
iffe
ren
tal
ign
men
tp
rogr
ams.
SUP
PL
EM
EN
TA
RY
TA
BL
EIV
.C
on
tin
ued

LITERATURE CITED
Bauer R, Begerow D, Sampaio JP, Weiß M, Oberwinkler F.2006. The simple-septate basidiomycetes: a synopsis.Mycol Progress 5:41–66.
Berres ME, Szabo LJ, McLaughlin DJ. 1995. Phylogeneticrelationships in auriculariaceous basidiomycetes basedon 25S ribosomal DNA sequences. Mycologia 87:821–840.
Fell JW, Boekhout T, Fonseca A, Sampaio JP. 2001.Basidiomycetous yeasts. In: McLaughlin DJ, McLaugh-lin EG, Lemke PA, eds. Systematics and Evolution.Berlin: Springer-Verlag, The Mycota VIIB: 3–35.
Lutzoni F, Kauff F, Cox CJ, 41 others. 2004. Assembling theFungal Tree of Life: progress, classification, andevolution of subcellular traits. Am J Bot 91:1446–1480.
McLaughlin DJ, Berres ME, Szabo LJ. 1995. Molecules and
morphology in basidiomycete phylogeny. Can J Bot73(1):S684–692.
Sampaio JP. 2004. Diversity, phylogeny and classification of
basidiomycetous yeasts. In: Agerer R, Piepenbring M,
Blanz P, eds. Frontiers in Basidiomycote Mycology.
Eching, Germany: IHW-Verlag. p 49–80.Swann EC, Frieders EM, McLaughlin DJ. 1999. Microbo-
tryum, Kriegeria and the changing paradigm in basid-iomycete classification. Mycologia 91:51–66.
Weiß M, Bauer R, Begerow D. 2004. Spotlights onheterobasidiomycetes. In: Agerer R, Piepenbring M,Blanz P, eds. Frontiers in Basidiomycote Mycology.Eching, Germany. IHW-Verlag. p 7–48.
Wingfield BD, Ericson L, Szaro T, Burdon JJ. 2004.Phylogenetic patterns in the Uredinales. AustralasPlant Pathol 33:327–335.

SUP
PL
EM
EN
TA
RY
TA
BL
EII
.G
enB
ank
nu
mb
ers
and
sou
rces
for
seq
uen
ces
anal
yzed
inth
isst
ud
y.D
atas
et1
con
tain
sth
ose
taxa
wit
hb
oth
nu
c-ls
uan
dn
uc-
ssu
seq
uen
ces.
Dat
aset
2co
nta
ins
all
seq
uen
ces.
Tax
au
sed
for
Dat
aset
3ar
ein
dic
ated
by
anas
teri
sk.
Do
ub
leas
teri
sks
ind
icat
ea
nu
c-ss
use
qu
ence
der
ived
fro
ma
dif
fere
nt
iso
late
/co
llec
tio
nth
anth
en
uc-
lsu
seq
uen
ce.
Spec
ies
nu
c-ls
urD
NA
nu
c-ss
urD
NA
Gen
Ban
kn
o.
len
gth
(bp
)R
efer
ence
(cit
edin
Gen
ban
k)G
enB
ank
no
.le
ngt
h(b
p)
Ref
eren
ce(c
ited
inG
enb
ank)
AS
CO
MY
CO
TA
Leo
tia
lubr
ica
(Sco
p.)
Per
s.A
Y544
644
1396
Lu
tzo
ni
etal
(200
4)A
Y544
687
1729
Lu
tzo
ni
etal
(200
4)
Mor
chel
lacf
.el
ata
Fr.
AY5
4466
513
86L
utz
on
iet
al(2
004)
AY5
4470
917
29L
utz
on
iet
al(2
004)
Neo
lect
avit
elli
na
(Bre
s.)
Ko
rf&
J.K
.R
oge
rsA
F27
9401
1252
Bh
atta
char
yaet
al(2
000)
Z27
393*
*17
22L
and
vik
etal
(199
3)
Sacc
haro
myc
esce
revis
iae
Mey
enex
E.C
.H
anse
nJ0
1355
3911
Bay
evet
al(1
981)
J013
5317
98R
ub
tso
vet
al(1
980)
Sor
dari
afi
mic
ola
(Ro
ber
geex
Des
m.)
Ces
.&
De
No
t.A
Y545
728
1343
Lu
tzo
ni
etal
(200
4)X
6985
1**
1796
Wil
mo
tte
etal
(199
3)
Taphri
na
wie
sner
i(R
ath
ay)
Mix
AY5
4829
214
11L
utz
on
iet
al(2
004)
AY5
4829
317
25L
utz
on
iet
al(2
004)
BA
SID
IOM
YC
OT
AA
GA
RIC
OM
YC
OT
INA
Au
ricu
lari
asp
.A
Y634
277
1433
Mat
hen
yet
al(2
004)
un
pu
bli
shed
DQ
2345
4217
64M
ath
eny
etal
(200
5)u
np
ub
lish
ed
Bot
ryob
asi
diu
mbo
tryo
sum
(Bre
s.)
J.E
riks
s.D
Q08
9013
1297
Nil
sso
net
al(2
005)
un
pu
bli
shed
AY6
6266
717
66N
ilss
on
etal
(200
5)u
np
ub
lish
ed
Calo
cera
corn
ea(B
atsc
h)
Fr.
*A
Y701
526
1367
Mat
hen
yet
al(2
004)
un
pu
bli
shed
AY7
7161
017
64M
ath
eny
etal
(200
4)u
np
ub
lish
ed
Fil
obasi
die
lla
neo
form
an
sK
wo
n-C
hu
ng
AJ5
5129
072
4K
idd
(200
3)u
np
ub
lish
edX
6018
3**
1769
Van
de
Pee
ret
al(1
992)
US
TIL
AG
INO
MY
CO
TIN
A
En
tylo
ma
hol
wayi
Syd
.A
Y745
721
1778
Mat
hen
yet
al(2
004)
un
pu
bli
shed
DQ
2345
6220
45M
ath
eny
etal
(200
5)u
np
ub
lish
ed
Mala
ssez
iafu
rfu
r(C
.P.
Ro
bin
)B
aill
.*A
Y745
725
1373
Mat
hen
yet
al(2
004)
un
pu
bli
shed
AY0
8322
3**
1628
McI
lhat
ton
and
Cu
rran
(200
2)u
np
ub
lish
edM
ala
ssez
iapach
yder
mati
s(W
eid
man
)C
.W.
Do
dge
AY7
4572
413
89M
ath
eny
etal
(200
4)u
np
ub
lish
edD
Q45
7640
1179
this
stu
dy
Spor
isor
ium
form
osan
um
(Saw
ada)
Van
kyA
Y740
134
594
Sto
llet
al(2
005)
DQ
8322
2610
96th
isst
ud
y
Til
leti
aco
ntr
over
saJ.
G.
Ku
hn
DQ
8322
4413
73th
isst
ud
yD
Q83
2245
829
this
stu
dy
Til
leti
ari
aan
omala
Ban
do
ni
&B
.N.
Joh
riA
Y745
715
1381
Mat
hen
yet
al(2
004)
un
pu
bli
shed
AY8
0375
217
71M
ath
eny
etal
(200
4)u
np
ub
lish
ed
Til
leti
opsi
sm
inor
Nyl
and
AY7
4571
314
04M
ath
eny
etal
(200
4)u
np
ub
lish
edD
8319
017
27T
akas
him
aan
dN
akas
e(1
996)
Til
leti
opsi
sw
ash
ingt
onen
sis
Nyl
and
AY7
4571
413
97M
ath
eny
etal
(200
4)u
np
ub
lish
edD
8319
217
30T
akas
him
aan
dN
akas
e(1
996)
Ust
ilago
mayd
is(D
C.)
Co
rda
AF
4539
3811
15P
iep
enb
rin
get
al(2
002)
X62
396*
*21
83D
eW
ach
ter
etal
(199
2)P
UC
CIN
IOM
YC
OT
INA
Aga
rico
stil
bum
hyp
haen
es(H
ar.
&P
at.)
Ob
erw
.&
Ban
do
ni*
AY6
3427
813
69M
ath
eny
etal
(200
4)u
np
ub
lish
edA
Y665
775
1774
Mat
hen
yet
al(2
004)
un
pu
bli
shed
Aga
rico
stil
bum
pu
lcher
rim
um
(Ber
k.&
Bro
om
e)B
.L.
Bra
dy,
B.
Sutt
on
&Sa
mso
nL
2027
753
9B
erre
set
al(1
995)
AY3
7339
1**
1760
McL
augh
lin
etal
(200
4)A
tract
iell
aso
lan
i(C
oh
n&
J.Sc
hro
t.)
Ob
erw
.&
Ban
do
ni
AY5
1283
155
4B
eger
ow
etal
(200
3)u
np
ub
lish
edA
ura
nti
ospor
ium
subn
iten
s(J
.Sc
hro
t.&
Hen
n.)
M.
Pie
pen
br.
,V
anky
&O
ber
w.
AF
0098
4656
0B
eger
ow
etal
(199
7)B
an
noa
hahaji
men
sis
Ham
am.,
Th
anh
&N
akas
eA
B08
2571
648
Nag
aham
aet
al(2
002)
un
pu
bli
shed

Spec
ies
nu
c-ls
urD
NA
nu
c-ss
urD
NA
Gen
Ban
kn
o.
len
gth
(bp
)R
efer
ence
(cit
edin
Gen
ban
k)G
enB
ank
no
.le
ngt
h(b
p)
Ref
eren
ce(c
ited
inG
enb
ank)
Ben
sin
gton
iach
an
gbaie
nsi
sF
.Y.
Bai
&Q
.M.
Wan
gA
Y233
342
639
Wan
get
al(2
003)
Ben
sin
gton
iaci
liata
Ingo
ld*
AY7
4573
014
02M
ath
eny
etal
(200
4)u
np
ub
lish
edD
3823
320
93T
akas
him
aet
al(1
995a
)
Ben
sin
gton
iain
gold
iiN
akas
e&
Ito
hA
F18
9888
631
Fel
let
al(2
000)
D38
234
1746
Tak
ash
ima
etal
(199
5a)
Ben
sin
gton
iam
isca
nth
i(N
akas
e&
M.
Suzu
ki)
Nak
ase
&B
oek
ho
ut
AF
1898
9164
0F
ell
etal
(200
0)D
3823
617
49T
akas
him
aet
al(1
995a
)B
ensi
ngt
onia
mu
sae
M.
Tak
ash
.,S.
O.
Suh
&N
akas
eA
Y512
833
628
Beg
ero
wet
al(2
003)
un
pu
bli
shed
D43
946*
*17
39T
akas
him
aet
al(1
995b
)B
ensi
ngt
onia
naga
noe
nsi
s(N
akas
e&
M.
Suzu
ki)
Nak
ase
&B
oek
ho
ut
AF
1898
9363
4F
ell
etal
(200
0)D
3836
617
53T
akas
him
aet
al(1
995a
)B
ensi
ngt
onia
phyl
lada
(Van
der
Wal
t&
Y.Ya
mad
a)V
and
erW
alt,
Nak
agaw
a&
Y.Ya
mad
aex
Bo
ekh
ou
tA
Y512
836
642
Beg
ero
wet
al(2
003)
un
pu
bli
shed
D38
237a
**17
48T
akas
him
aet
al(1
995a
)B
ensi
ngt
onia
saka
guch
iiSu
gita
,M
.T
akas
h.,
Ham
am.
&N
akas
eA
F36
3646
599
Sco
rzet
tiet
al(2
002)
AB
0017
4626
53Su
gita
etal
(199
7)
Ben
sin
gton
iaso
rbi
F.Y
.B
ai&
Q.M
.W
ang
AY2
3334
563
1W
ang
etal
(200
3)B
ensi
ngt
onia
subr
osea
(Nak
ase
&M
.Su
zuki
)N
akas
e&
Bo
ekh
ou
tA
F18
9895
640
Fel
let
al(2
000)
D38
238
1744
Tak
ash
ima
etal
(199
5a)
Ben
sin
gton
iaya
mato
an
a(N
akas
e,M
.Su
zuki
&It
oh
)N
akas
e&
Bo
ekh
ou
tA
F18
9896
619
Fel
let
al(2
000)
D38
239
2658
Tak
ash
ima
etal
(199
5a)
Ben
sin
gton
iayu
ccic
ola
(Nak
ase
&M
.Su
zuki
)N
akas
e&
Bo
ekh
ou
t*A
Y745
727
1388
Mat
hen
yet
al(2
004)
un
pu
bli
shed
DQ
2345
4317
75M
ath
eny
etal
(200
5)u
np
ub
lish
edC
am
pto
basi
diu
mhyd
rophil
um
Mar
van
ova
&Su
ber
kr.
[1]
AY2
1299
157
0Sa
mp
aio
etal
(200
3)
Cam
pto
basi
diu
mhyd
rophil
um
[2]
AY5
1283
754
1B
eger
ow
etal
(200
3)u
np
ub
lish
ed
Chio
nos
phaer
aapob
asi
dia
lis
D.E
.C
ox
[1]
AF
3934
7048
7K
irsc
hn
eret
al(2
001)
Chio
nos
phaer
aapob
asi
dia
lis
[2]
AF
1774
0763
2Sa
mp
aio
etal
(199
9b)
U77
662*
*16
58Sw
ann
etal
(199
9)C
hio
nos
phaer
acu
nic
uli
cola
R.
Kir
sch
ner
,B
eger
ow
&O
ber
w.
AF
3934
7348
3K
irsc
hn
eret
al(2
001)
Chry
som
yxa
arc
tost
aphyl
iD
iete
l*A
Y700
192
1180
Mat
hen
yet
al(2
004)
un
pu
bli
shed
AY6
5700
916
67M
ath
eny
etal
(200
4)u
np
ub
lish
edC
lass
icu
lafl
uit
an
sR
.B
auer
,B
eger
ow
,O
ber
w.
&M
arva
no
vaA
Y512
838
559
Beg
ero
wet
al(2
003)
un
pu
bli
shed
AY1
2447
815
16B
auer
etal
(200
3)C
olaco
gloe
apen
iophor
ae
(Bo
urd
ot
&G
alzi
n)
Ob
erw
.,R
.B
auer
&B
and
on
i[1
]A
F18
9898
621
Fel
let
al(2
000)
Col
aco
gloe
apen
iophor
ae
[2]
AY5
1283
961
6B
eger
ow
etal
(200
3)u
np
ub
lish
ed
Col
aco
gloe
apen
iophor
ae
[3]*
AY6
2931
313
15M
ath
eny
etal
(200
4)u
np
ub
lish
edD
Q23
4565
2146
Mat
hen
yet
al(2
005)
un
pu
bli
shed
Col
eosp
oriu
mast
eru
m(D
iete
l)Sy
d.
&P
.Sy
d.
AF
5221
6495
1Sz
aro
and
Bru
ns
(200
2)u
np
ub
lish
edA
Y123
286
1760
Win
gfie
ldet
al(2
004)
Cro
nart
ium
ribi
cola
J.C
.F
isch
.A
F52
2166
963
Szar
oan
dB
run
s(2
002)
un
pu
bli
shed
M94
338
1753
Bru
ns
etal
(199
2)
Cry
pto
myc
ocol
ax
abn
orm
isO
ber
w.
&R
.B
auer
AY5
1284
160
9B
eger
ow
etal
(200
3)u
np
ub
lish
ed
Cyr
enel
lael
egan
sG
och
.A
Y512
842
573
Beg
ero
wet
al(2
003)
un
pu
bli
shed
SUP
PL
EM
EN
TA
RY
TA
BL
EII
.C
on
tin
ued
Spec
ies
nu
c-ls
urD
NA
nu
c-ss
urD
NA
Gen
Ban
kn
o.
len
gth
(bp
)R
efer
ence
(cit
edin
Gen
ban
k)G
enB
ank
no
.le
ngt
h(b
p)
Ref
eren
ce(c
ited
inG
enb
ank)
Cys
toba
sidiu
mfi
met
ari
um
(Sch
um
ach
.)P
.R
ob
erts
AY5
1284
363
3B
eger
ow
etal
(200
3)u
np
ub
lish
edA
Y124
479
1624
Bau
eret
al(2
003)
En
doc
ron
art
ium
hark
nes
sii
Hir
ats.
*A
Y700
193
1200
Mat
hen
yet
al(2
004)
un
pu
bli
shed
AY6
6578
517
64M
ath
eny
etal
(200
4)u
np
ub
lish
ed
Eoc
ron
art
ium
mu
scic
ola
(Per
s.)
Fit
zp.
[1]
AF
0148
2591
4B
run
san
dSz
aro
(199
7)u
np
ub
lish
edA
Y123
323
1763
Win
gfie
ldet
al(2
004)
Eoc
ron
art
ium
mu
scic
ola
[2]
AY5
1284
462
2B
eger
ow
etal
(200
3)u
np
ub
lish
edE
ryth
roba
sidiu
mhase
gaw
ian
um
Ham
am.,
Sugi
y.&
Ko
mag
.[1
]A
F13
1058
603
Sam
pai
oet
al(1
999a
)
Ery
thro
basi
diu
mhase
gaw
ian
um
[2]
AF
1898
9959
8F
ell
etal
(200
0)D
1280
3**
1755
Suh
and
Sugi
yam
a(1
993)
Fu
lvis
por
ium
rest
ifaci
ens
(D.E
.Sh
aw)
Van
kyA
F00
9860
561
Beg
ero
wet
al(1
997)
Gym
nos
por
an
giu
mju
nip
eri-
vir
gin
ian
ae
Sch
wei
n.
AY6
2931
613
48M
ath
eny
etal
(200
4)u
np
ub
lish
edD
Q66
7158
1145
this
stu
dy
Hel
icob
asi
diu
mco
rtic
ioid
esB
and
on
iU
7530
317
47Sw
ann
and
McL
augh
lin(1
996)
un
pu
blis
hed
Hel
icob
asi
diu
mlo
ngi
spor
um
Wak
ef.
AY2
2204
611
32L
utz
etal
(200
4b)
Hel
icob
asi
diu
mm
ompa
Tan
aka
[1]
AY2
5417
811
28L
utz
etal
(200
4a)
Hel
icob
asi
diu
mm
ompa
[2]*
AY2
5417
911
17L
utz
etal
(200
4a)
U77
064
1657
Swan
net
al(1
999)
Hel
icob
asi
diu
mpu
rpu
reu
m(T
ul.
)P
at.
[1]
AY2
2204
911
06L
utz
etal
(200
4b)
Hel
icob
asi
diu
mpu
rpu
reu
m[2
]A
Y254
180
1121
Lu
tzet
al(2
004a
)
Hel
icog
loea
lage
rhei
mii
Pat
.A
Y512
849
510
Beg
ero
wet
al(2
003)
un
pu
bli
shed
AY1
2447
617
21B
auer
etal
(200
3)
Hel
icog
loea
vari
abi
lis
K.
Wel
lsL
2028
255
7B
erre
set
al(1
995)
U78
043
1769
Swan
net
al(1
999)
Her
pob
asi
diu
mfi
lici
nu
m(R
ost
r.)
Lin
d[1
]A
F42
6193
506
Mai
eret
al(2
003)
Her
pob
asi
diu
mfi
lici
nu
m[2
]A
Y512
850
619
Beg
ero
wet
al(2
003)
un
pu
bli
shed
Het
erog
ast
ridiu
mpyc
nid
ioid
eum
Ob
erw
.&
R.
Bau
erA
Y512
851
599
Beg
ero
wet
al(2
003)
un
pu
bli
shed
U41
567*
*16
51Sw
ann
and
Tay
lor
(199
5)
Hob
son
iam
irabi
lis
(Pec
k)L
ind
erA
F28
9663
1720
Sika
roo
di
etal
(200
1)
Hya
lopso
rapol
ypod
ii(P
ers.
)M
agn
.A
Y512
852
516
Beg
ero
wet
al(2
003)
un
pu
bli
shed
AB
0110
1517
47Sj
amsu
rid
zal
etal
(199
9)In
soli
basi
diu
mdef
orm
an
s(C
.J.
Go
uld
)O
ber
w.
&B
and
on
i*A
Y646
099
1413
Mat
hen
yet
al(2
004)
un
pu
bli
shed
AY1
2329
2**
1756
Win
gfie
ldet
al(2
004)
Jacu
lisp
ora
subm
ersa
H.J
.H
ud
s.&
Ingo
ldA
Y512
853
595
Beg
ero
wet
al(2
003)
un
pu
bli
shed
AY1
2447
716
69B
auer
etal
(200
3)
Jola
cf.
javen
sis
Pat
.D
Q41
6207
570
this
stu
dy
DQ
4162
0617
17th
isst
ud
y
Jola
hoo
keri
aru
mA
.M
øll
erD
Q41
6208
568
this
stu
dy
Kon
doa
aer
iaA
.F
on
seca
,J.
P.
Sam
p.
&F
ell
AF
1899
0263
8F
ell
etal
(200
0)K
ondoa
malv
inel
la(F
ell
&I.
L.
Hu
nte
r)Y.
Yam
ada,
Nak
agaw
a&
I.B
ann
oA
Y745
720
1440
Mat
hen
yet
al(2
004)
un
pu
bli
shed
DQ
6671
6411
90th
isst
ud
y
Kon
doa
sp.
AF
1899
0463
7F
ell
etal
(200
0)
Kri
eger
iaer
iophor
iB
res.
[1]*
AY7
4572
813
97M
ath
eny
etal
(200
4)u
np
ub
lish
edD
Q41
9918
1766
this
stu
dy
Kri
eger
iaer
iophor
i[2
]A
F18
9905
b61
9F
ell
etal
(200
0)U
7706
3b**
1684
Swan
net
al(1
999)
Ku
ehn
eola
ure
din
is(L
ink)
Art
hu
rA
Y745
696
1385
Mat
hen
yet
al(2
004)
un
pu
bli
shed
DQ
0929
1913
10M
ath
eny
etal
(200
5)u
np
ub
lish
ed
Ku
rtzm
an
omyc
esin
soli
tus
J.P
.Sa
mp
.&
Fel
lA
F17
7408
630
Sam
pai
oet
al(1
999b
)
SUP
PL
EM
EN
TA
RY
TA
BL
EII
.C
on
tin
ued

Spec
ies
nu
c-ls
urD
NA
nu
c-ss
urD
NA
Gen
Ban
kn
o.
len
gth
(bp
)R
efer
ence
(cit
edin
Gen
ban
k)G
enB
ank
no
.le
ngt
h(b
p)
Ref
eren
ce(c
ited
inG
enb
ank)
Ku
rtzm
an
omyc
esn
ecta
irei
(Ro
dr.
Mir
.)Y.
Yam
ada,
Ito
h,
H.
Kaw
as.,
I.B
ann
o&
Nak
ase
AF
1774
0962
9Sa
mp
aio
etal
(199
9b)
D64
122*
*17
39Su
het
al(1
996b
)
Ku
rtzm
an
omyc
esta
rdu
sG
im.-J
ura
do
&U
den
AF
1774
1062
9Sa
mp
aio
etal
(199
9b)
Leu
cosp
orid
iell
acr
eati
niv
ora
(Go
lub
ev)
J.P
.Sa
mp
.A
F18
9925
c61
9F
ell
etal
(200
0)L
euco
spor
idie
lla
fraga
ria
(J.A
.B
arn
ett
&B
uh
agia
r)J.
P.
Sam
p.
AY5
1287
959
4B
eger
ow
etal
(200
3)u
np
ub
lish
edL
euco
spor
idie
lla
mu
scor
um
(Di
Men
na)
J.P
.Sa
mp
.A
Y213
008
589
Sam
pai
oet
al(2
003)
Leu
cosp
orid
iell
aya
kuti
ca(G
olu
bev
)J.
P.
Sam
p.
AY2
1300
158
3Sa
mp
aio
etal
(200
3)L
euco
spor
idiu
man
tarc
ticu
mF
ell,
Stat
zell
,I.
L.
Hu
nte
r&
Ph
aff
AY5
1285
562
2B
eger
ow
etal
(200
3)u
np
ub
lish
ed
Leu
cosp
orid
ium
fell
iiG
im.-J
ura
do
&U
den
AY5
1285
661
7B
eger
ow
etal
(200
3)u
np
ub
lish
edL
euco
spor
idiu
msc
otti
iF
ell,
Stat
zell
,I.
L.
Hu
nte
r&
Ph
aff
[1]
AF
0704
1961
9F
ell
etal
(199
8)X
5349
9**
1765
Hen
dri
kset
al(1
991)
Leu
cosp
orid
ium
scot
tii
[2]*
AY6
4609
813
89M
ath
eny
etal
(200
4)u
np
ub
lish
edA
Y707
092
1803
Mat
hen
yet
al(2
004)
un
pu
bli
shed
Lir
oaem
oden
sis
(Ber
k.)
Cif
.[1
]A
Y512
858
565
Beg
ero
wet
al(2
003)
un
pu
bli
shed
Lir
oaem
oden
sis
[2]
AY2
1299
258
7Sa
mp
aio
etal
(200
3)
Mast
igob
asi
diu
min
term
ediu
mG
olu
bev
[1]
AF
1898
8961
9F
ell
etal
(200
0)
Mast
igob
asi
diu
min
term
ediu
m[2
]A
Y512
859
603
Beg
ero
wet
al(2
003)
un
pu
bli
shed
D38
235*
*17
44T
akas
him
aet
al(1
995a
)
Mel
am
pso
raeu
phor
biae
(C.
Sch
ub
.)C
asta
gne
DQ
4375
0411
00th
isst
ud
yM
icro
botr
yum
cord
ae
(Lir
o)
G.
Dem
l&
Pri
llin
ger
AY2
1300
258
9Sa
mp
aio
etal
(200
3)M
icro
botr
yum
reti
cula
tum
(Lir
o)
R.
Bau
er&
Ob
erw
.A
Y213
003
586
Sam
pai
oet
al(2
003)
U79
566*
*92
5Sw
ann
etal
(199
9)M
icro
botr
yum
vio
lace
um
(Per
s.)
G.
Dem
l&
Ob
erw
.A
Y512
864
620
Beg
ero
wet
al(2
003)
un
pu
bli
shed
U77
062*
*17
27Sw
ann
and
Tay
lor
(199
5)
Mix
iaos
mu
ndae
(Nis
hid
a)C
.L.
Kra
mer
[1]
AB
0528
4060
0Sj
amsu
rid
zal
etal
(200
2)D
1416
317
80N
ish
ida
etal
(199
5)
Mix
iaos
mu
ndae
[2]
AY5
1286
761
3B
eger
ow
etal
(200
3)u
np
ub
lish
edM
ycog
loea
macr
ospor
a(B
erk.
&B
roo
me)
McN
abb
U41
848
1601
Swan
nan
dT
aylo
r(1
995)
Myc
oglo
ean
ippon
ica
Ban
do
ni
AY1
5965
060
2K
irsc
hn
eret
al(2
002)
un
pu
bis
hed
Myc
oglo
easp
.A
Y512
868
629
Beg
ero
wet
al(2
003)
un
pu
bli
shed
Naoh
idea
seba
cea
(Ber
k.&
Bro
om
e)O
ber
w.
[1]
AF
1310
6154
1Sa
mp
aio
etal
(199
9a)
Naoh
idea
seba
cea
[2]*
AF
5221
7612
03Sz
aro
and
Bru
ns
(200
2)u
np
ub
lish
edA
Y123
302
2094
Win
gfie
ldet
al(2
004)
Occ
ult
ifu
rex
tern
us
J.P
.Sa
mp
.,R
.B
auer
&O
ber
w.
[1]*
AY7
4572
314
23M
ath
eny
etal
(200
4)u
np
ub
lish
edD
Q45
7639
1774
this
stu
dy
Occ
ult
ifu
rex
tern
us
[2]
AF
1899
1162
9F
ell
etal
(200
0)A
Y124
475*
*16
29B
auer
etal
(200
3)
Pach
noc
ybe
ferr
ugi
nea
Ber
k.L
2028
455
4B
erre
set
al(1
995)
SUP
PL
EM
EN
TA
RY
TA
BL
EII
.C
on
tin
ued

Spec
ies
nu
c-ls
urD
NA
nu
c-ss
urD
NA
Gen
Ban
kn
o.
len
gth
(bp
)R
efer
ence
(cit
edin
Gen
ban
k)G
enB
ank
no
.le
ngt
h(b
p)
Ref
eren
ce(c
ited
inG
enb
ank)
Phle
ogen
afa
gin
ea(F
r.)
Lin
kA
Y512
869
537
Beg
ero
wet
al(2
003)
un
pu
bli
shed
AY3
7339
2**
1833
McL
augh
lin
etal
(200
4)P
ileo
lari
ato
xico
den
dri
(Ber
k.&
Rav
enel
)A
rth
ur
[1]
AY7
4569
911
73M
ath
eny
etal
(200
4)u
np
ub
lish
ed
Pil
eola
ria
toxi
coden
dri
[2]
AY7
4569
814
14M
ath
eny
etal
(200
4)u
np
ub
lish
edD
Q09
2921
1774
Mat
hen
yet
al(2
005)
un
pu
bli
shed
Pla
tygl
oea
dis
cifo
rmis
(Fr.
)N
euh
off
*A
Y629
314
1355
Mat
hen
yet
al(2
004)
un
pu
bli
shed
DQ
2345
6317
63M
ath
eny
etal
(200
5)u
np
ub
lish
ed
Pla
tygl
oea
pu
stu
lata
G.W
.M
arti
n&
Cai
nA
Y512
871
585
Beg
ero
wet
al(2
003)
un
pu
bli
shed
Pla
tygl
oea
ves
tita
Bo
urd
ot
&G
alzi
nA
Y512
872
593
Beg
ero
wet
al(2
003)
un
pu
bli
shed
AY1
2448
016
02B
auer
etal
(200
3)
Ple
uro
coll
aco
mpre
ssa
(Ell
is&
Eve
rh.)
Die
hl
AY3
8258
150
1K
irsc
hn
eret
al(2
004)
un
pu
bli
shed
Pu
ccin
iaaru
ndin
ari
ae
Sch
wei
n.
DQ
4152
7710
42th
isst
ud
y
Pu
ccin
iahor
dei
G.H
.O
tth
DQ
3545
2710
74A
ime
(200
6)D
Q41
5278
this
stu
dy
Pu
ccin
iast
rum
goep
per
tian
um
(Ku
ehn
)K
leb
.A
F52
2180
940
Szar
oan
dB
run
s(2
002)
un
pu
bli
shed
AY1
2330
517
54W
ingf
ield
etal
(200
4)
Ren
ifor
ma
stru
esP
ore
&So
ren
son
AF
1899
1255
7F
ell
etal
(200
0)R
hod
ospor
idiu
mdio
bovatu
mS.
Y.N
ewel
l&
I.L
.H
un
ter
AF
1899
1459
7F
ell
etal
(200
0)R
hod
ospor
idiu
mfl
uvia
leF
ell,
Ku
rtzm
an,
Tal
lman
&J.
D.
Bu
ck[1
]A
F07
0422
604
Fel
let
al(1
998)
Rhod
ospor
idiu
mfl
uvia
le[2
]*A
Y745
719
1362
Mat
hen
yet
al(2
004)
un
pu
bli
shed
U77
395*
*17
37Sw
ann
etal
(199
9)R
hod
ospor
idiu
msp
haer
ocarp
um
S.Y.
New
ell
&F
ell
AY5
1287
556
9B
eger
ow
etal
(200
3)u
np
ub
lish
ed
Rhod
ospor
idiu
mto
rulo
ides
Ban
no
AF
0704
2660
3F
ell
etal
(199
8)D
1280
6**
1759
Suh
and
Sugi
yam
a(1
993)
Rhod
otor
ula
arm
enia
caR
.G.
Shiv
as&
Ro
dr.
Mir
.A
F18
9920
629
Fel
let
al(2
000)
AB
1266
4417
83N
agah
ama
etal
(200
3)u
np
ub
lish
ed
Rhod
otor
ula
au
ran
tiaca
(Sai
to)
Lo
dd
erA
Y512
877
626
Beg
ero
wet
al(2
003)
un
pu
bli
shed
AB
0303
54**
1757
Tak
ash
ima
etal
(200
0)R
hod
otor
ula
bogo
rien
sis
(Dei
nem
a)A
rx&
Wei
jman
AF
1899
2359
2F
ell
etal
(200
0)
Rhod
otor
ula
cres
olic
aM
idd
elh
ove
n&
Spaa
ijA
F18
9926
619
Fel
let
al(2
000)
Rhod
otor
ula
dif
flu
ens
(Ru
inen
)T
.H
aseg
.,B
ann
o&
Yam
auch
iA
Y512
878
621
Beg
ero
wet
al(2
003)
un
pu
bli
shed
Rhod
otor
ula
foli
oru
m(R
uin
en)
Ro
dr.
Mir
.&
Wei
jma
AF
3178
0461
9F
ell
(200
0)u
np
ub
lish
ed
Rhod
otor
ula
glu
tin
is(F
rese
n.)
F.C
.H
arri
son
[1]
AF
0704
3060
4F
ell
etal
(199
8)
Rhod
otor
ula
glu
tin
is[2
]*A
Y646
097
1349
Mat
hen
yet
al(2
004)
un
pu
bli
shed
DQ
8321
9417
62th
isst
ud
y
Rhod
otor
ula
gram
inis
Di
Men
na
AF
0704
3160
4F
ell
etal
(199
8)X
8382
7**
1803
Cai
etal
(199
6)
Rhod
otor
ula
hor
dea
Ro
dr.
Mir
.&
Wei
jman
[1]
AY5
1288
162
3B
eger
ow
etal
(200
3)u
np
ub
lish
ed
Rhod
otor
ula
hor
dea
[2]*
AY6
3190
114
12M
ath
eny
etal
(200
4)u
np
ub
lish
edA
Y657
013
1777
Mat
hen
yet
al(2
004)
un
pu
bli
shed
Rhod
otor
ula
lact
osa
T.
Has
eg.
AF
1899
3660
1F
ell
etal
(200
0)D
4536
617
48Su
het
al(1
996a
)
Rhod
otor
ula
lary
ngi
sR
eier
sol
AF
1899
4262
9F
ell
etal
(200
0)A
B12
6649
1782
Nag
aham
aet
al(2
003)
un
pu
bli
shed
Rhod
otor
ula
mari
na
Ph
aff,
Mra
k&
O.B
.W
illi
ams
AF
1899
4462
9F
ell
etal
(200
0)A
B12
6645
1784
Nag
aham
aet
al(2
003)
un
pu
bli
shed
Rhod
otor
ula
min
uta
(Sai
to)
F.C
.H
arri
son
AF
1899
4562
9F
ell
etal
(200
0)D
4536
717
52Su
het
al(1
996a
)
SUP
PL
EM
EN
TA
RY
TA
BL
EII
.C
on
tin
ued

Spec
ies
nu
c-ls
urD
NA
nu
c-ss
urD
NA
Gen
Ban
kn
o.
len
gth
(bp
)R
efer
ence
(cit
edin
Gen
ban
k)G
enB
ank
no
.le
ngt
h(b
p)
Ref
eren
ce(c
ited
inG
enb
ank)
Rhod
otor
ula
mu
cila
gin
osa
(A.
Jorg
.)F
.C.
Har
riso
nA
F07
0432
600
Fel
let
al(1
998)
AB
0216
68**
1760
Ham
amo
toan
dN
akas
e(2
000)
Rhod
otor
ula
pall
ida
Lo
dd
erA
F18
9962
629
Fel
let
al(2
000)
AB
1266
5117
84N
agah
ama
etal
(200
3)u
np
ub
lish
edR
hod
otor
ula
pin
icol
aF
.Y.
Bai
,L
.D.
Gu
o&
J.H
.Z
hao
AF
4442
9362
7Z
hao
etal
(200
2)A
B12
6652
1783
Nag
aham
aet
al(2
003)
un
pu
bli
shed
Rhod
otor
ula
sloo
ffia
eE
.K.
No
vak
&V
oro
s-F
elka
iA
F18
9968
629
Fel
let
al(2
000)
AB
1266
53**
1781
Nag
aham
aet
al(2
003)
un
pu
bli
shed
Rhod
otor
ula
son
ckii
(Ho
psu
-Hav
u,
Tu
nn
ela
&Ya
rro
w)
Ro
dr.
Mir
.&
Wei
jman
[1]
AF
1899
6962
7F
ell
etal
(200
0)
Rhod
otor
ula
son
ckii
[2]
AY2
1300
959
2Sa
mp
aio
etal
(200
3)
Rhod
otor
ula
sp.
1A
B05
5196
d63
6Sj
amsu
rid
zal
etal
(200
1)u
np
ub
lish
edA
B05
5191
d17
42Sj
amsu
rid
zal
etal
(200
1)u
np
ub
lish
ed
Rhod
otor
ula
sp.
2A
B05
5197
d63
6Sj
amsu
rid
zal
etal
(200
1)u
np
ub
lish
edA
B05
5192
d17
43Sj
amsu
rid
zal
etal
(200
1)u
np
ub
lish
ed
Rhod
otor
ula
sp.
3A
B17
6591
e59
1N
akas
eet
al(2
004)
un
pu
bli
shed
AB
1765
30e
1783
Nak
ase
etal
(200
4)u
np
ub
lish
ed
Rhod
otor
ula
sp.
4A
B05
5194
f63
7Sj
amsu
rid
zal
etal
(200
1)u
np
ub
lish
edA
B05
5189
f17
36Sj
amsu
rid
zal
etal
(200
1)u
np
ub
lish
ed
Rhod
otor
ula
sp.
5A
B05
5195
g63
4Sj
amsu
rid
zal
etal
(200
1)u
np
ub
lish
edA
B05
5190
g17
19Sj
amsu
rid
zal
etal
(200
1)u
np
ub
lish
ed
Sacc
obla
stia
fari
nace
a(H
oh
n.)
Do
nk
AJ4
0640
190
3L
ange
r(2
000)
un
pu
bli
shed
Saka
guch
iadacr
yoid
ea(F
ell,
I.L
.H
un
ter
&T
allm
an)
Y.Ya
mad
a,K
.M
aed
a&
Mik
ata
[1]
AF
1899
7257
1F
ell
etal
(200
0)
Saka
guch
iadacr
yoid
ea[2
]A
F18
9973
571
Fel
let
al(2
000)
Sep
toba
sidiu
mbu
rtii
Llo
ydA
Y373
388
1430
McL
augh
lin
etal
(200
4)
Sep
toba
sidiu
mca
nes
cen
sB
urt
AY3
7338
917
87M
cLau
ghli
net
al(2
004)
Sep
toba
sidiu
mca
rest
ian
um
Bre
s.L
2028
954
8B
erre
set
al(1
995)
Sep
toba
sidiu
msp
.hA
Y254
182
570
Lu
tzet
al(2
004a
)A
Y123
320*
*17
29W
ingf
ield
etal
(200
4)Sphace
loth
eca
pol
ygon
i-per
sica
riae
G.
Dem
l&
Ob
erw
.A
F18
9974
619
Fel
let
al(2
000)
Sphace
loth
eca
pol
ygon
i-se
rru
lati
Mai
reA
Y512
884
620
Beg
ero
wet
al(2
003)
un
pu
bli
shed
Sphen
ospor
ake
vor
kian
iiL
ind
erD
Q35
4521
1205
Aim
e(2
006)
Spic
ulo
gloe
asp
.A
Y512
885
601
Beg
ero
wet
al(2
003)
un
pu
bli
shed
Spor
idio
bolu
sjo
hn
son
iiN
ylan
d[1
]A
Y167
608
604
Alv
arez
-Ro
dri
guez
etal
(200
3)L
2226
1**
1760
Swan
nan
dT
aylo
r(1
993)
Spor
idio
bolu
sjo
hn
son
ii[2
]*A
Y745
718
1371
Mat
hen
yet
al(2
004)
un
pu
bli
shed
AB
0216
72**
1807
Ham
amo
toan
dN
akas
e(2
000)
Spor
idio
bolu
spara
rose
us
Fel
l&
Tal
lman
[1]
AF
0704
3760
1F
ell
etal
(199
8)A
B02
1689
**17
59H
amam
oto
and
Nak
ase
(200
0)
Spor
idio
bolu
spara
rose
us
[2]
AF
1899
7860
1F
ell
etal
(200
0)A
B02
1694
**17
58H
amam
oto
and
Nak
ase
(200
0)
Spor
idio
bolu
ssa
lmon
icol
orF
ell
&T
allm
anA
F07
0439
604
Fel
let
al(1
998)
AB
0216
9717
81H
amam
oto
and
Nak
ase
(200
0)
Spor
obol
omyc
esbl
um
eae
M.
Tak
ash
.&
Nak
ase
AY2
1301
057
3Sa
mp
aio
etal
(200
3)A
B03
0321
1756
Tak
ash
ima
and
Nak
ase
(200
0)
Spor
obol
omyc
esco
pro
smae
Ham
am.
&N
akas
eA
F18
9980
629
Fel
let
al(2
000)
D66
880
1757
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
esco
pro
smic
ola
Ham
am.
&N
akas
e[1
]A
F18
9981
630
Fel
let
al(2
000)
D66
879
1749
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
esco
pro
smic
ola
[2]
AY5
1288
863
2B
eger
ow
etal
(200
3)u
np
ub
lish
ed
Spor
obol
omyc
esdra
cophyl
liH
amam
.&
Nak
ase
AF
1899
8263
0F
ell
etal
(200
0)D
6688
217
57H
amam
oto
and
Nak
ase
(200
0)
SUP
PL
EM
EN
TA
RY
TA
BL
EII
.C
on
tin
ued

Spec
ies
nu
c-ls
urD
NA
nu
c-ss
urD
NA
Gen
Ban
kn
o.
len
gth
(bp
)R
efer
ence
(cit
edin
Gen
ban
k)G
enB
ank
no
.le
ngt
h(b
p)
Ref
eren
ce(c
ited
inG
enb
ank)
Spor
obol
omyc
esel
onga
tus
R.G
.Sh
ivas
&R
od
r.M
ir.
AF
1899
8360
1F
ell
etal
(200
0)A
B02
1669
1761
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
esfa
lcatu
sN
akas
e,It
oh
&M
.Su
zuki
AF
0754
9062
1F
ell
etal
(199
9)A
B02
1670
1758
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
esfo
liic
ola
R.G
.Sh
ivas
&R
od
r.M
ir.
AF
1899
8462
9F
ell
etal
(200
0)A
B02
1671
1781
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
esgr
aci
lis
Der
xA
F18
9985
629
Fel
let
al(2
000)
AB
1784
8017
87N
agah
ama
etal
(200
4)u
np
ub
lish
ed
Spor
obol
omyc
esgr
iseo
flavu
sN
akas
e&
M.
Suzu
kiA
F20
7888
610
Ho
ng
etal
(200
0)D
6688
417
58H
amam
oto
and
Nak
ase
(200
0)Spor
obol
omyc
esin
osit
ophil
us
Nak
ase
&M
.Su
zuki
AF
1899
8761
9F
ell
etal
(200
0)A
B02
1673
1781
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
eskl
uyv
eri-
nie
lii
Van
der
Wal
tA
F18
9988
629
Fel
let
al(2
000)
AB
0216
7417
53H
amam
oto
and
Nak
ase
(200
0)Spor
obol
omyc
esla
ctop
hil
us
Nak
ase,
Ito
h,
M.
Suzu
ki&
Ban
do
ni
AF
1774
1162
1Sa
mp
aio
etal
(199
9b)
AB
0216
7516
68H
amam
oto
and
Nak
ase
(200
0)Spor
obol
omyc
esli
nder
ae
Nak
ase,
M.
Tak
ash
.&
Ham
am.
[1]
AF
1899
8963
1F
ell
etal
(200
0)
Spor
obol
omyc
esli
nder
ae
[2]
AF
2078
9062
1H
on
get
al(2
000)
D66
885
1753
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
esn
ylan
dii
M.
Tak
ash
.&
Nak
ase
AF
3871
2357
3V
aler
ioet
al(2
002)
AB
0303
1917
56T
akas
him
aan
dN
akas
e(2
000)
Spor
obol
omyc
esor
yzic
ola
Nak
ase
&M
.Su
zuki
AF
1899
9062
9F
ell
etal
(200
0)A
B02
1677
1756
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
esphyl
lom
ati
sV
and
erW
alt
&Y.
Yam
ada
AF
1899
9162
9F
ell
etal
(200
0)A
B02
1685
1761
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
espoo
nso
okia
eM
.T
akas
h.
&N
akas
eA
F38
7124
570
Val
erio
etal
(200
2)A
B03
0320
1757
Tak
ash
ima
and
Nak
ase
(200
0)Spor
obol
omyc
esru
ber
(Nak
ase,
G.
Oka
da
&Su
giy.
)B
oek
ho
ut
AF
1899
9262
6F
ell
etal
(200
0)A
B02
1686
2072
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
essa
lici
nu
s(B
.N.
Joh
ri&
Ban
do
ni)
Nak
ase
&S.
Ito
AF
1899
9562
5F
ell
etal
(200
0)A
B02
1687
1760
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
essa
sico
laN
akas
e&
M.
Suzu
kiA
F20
7889
613
Ho
ng
etal
(200
0)A
B02
1688
2088
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
essi
ngu
lari
sP
haf
f&
Car
mo
Sou
zaA
F20
7885
609
Ho
ng
etal
(200
0)A
B02
1690
**17
63H
amam
oto
and
Nak
ase
(200
0)Spor
obol
omyc
essu
bbru
nn
eus
Nak
ase
&M
.Su
zuki
*A
Y745
717
1431
Mat
hen
yet
al(2
004)
un
pu
bli
shed
AB
0216
9117
44H
amam
oto
and
Nak
ase
(200
0)
Spor
obol
omyc
esta
upoe
nsi
sH
amam
.&
Nak
ase
AF
1774
1362
3Sa
mp
aio
etal
(199
9b)
D66
886
2044
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
ests
uga
e(P
haf
f&
Car
mo
Sou
za)
Nak
ase
&It
oh
AF
1899
9862
1F
ell
etal
(200
0)A
B02
1692
1760
Ham
amo
toan
dN
akas
e(2
000)
Spor
obol
omyc
esxa
nth
us
(Nak
ase,
G.
Oka
da
&Su
giy.
)B
oek
ho
ut
AF
1774
1461
6Sa
mp
aio
etal
(199
9b)
D64
118
1730
Suh
etal
(199
6b)
Spor
obol
omyc
esyu
nn
an
ensi
sF
.Y.
Bai
,M
.T
akas
h.,
Ham
am.
&N
akas
eA
Y335
162
601
Bai
etal
(200
4)A
F22
9176
1777
Bai
etal
(200
1)
Ste
rigm
ato
myc
esel
via
eSo
nck
&Ya
rr[1
]A
F18
9999
632
Fel
let
al(2
000)
Ste
rigm
ato
myc
esel
via
e[2
]A
F19
0000
632
Fel
let
al(2
000)
AY3
7339
0**
1047
McL
augh
lin
etal
(200
4)
SUP
PL
EM
EN
TA
RY
TA
BL
EII
.C
on
tin
ued

Spec
ies
nu
c-ls
urD
NA
nu
c-ss
urD
NA
Gen
Ban
kn
o.
len
gth
(bp
)R
efer
ence
(cit
edin
Gen
ban
k)G
enB
ank
no
.le
ngt
h(b
p)
Ref
eren
ce(c
ited
inG
enb
ank)
Ste
rigm
ato
myc
eshalo
phil
us
Fel
l*A
Y745
716
1400
Mat
hen
yet
al(2
004)
un
pu
bli
shed
D64
119
1749
Suh
etal
(199
6b)
Sti
lbu
mvu
lgare
To
de
AY3
7338
717
99M
cLau
ghli
net
al(2
004)
Than
ato
phyt
um
croc
oru
m(P
ers.
)N
ees
AY8
8516
8i91
7M
ath
eny
etal
(200
5)u
np
ub
lish
edD
8564
8i **17
42K
un
inag
a(1
996)
un
pu
bli
shed
Tri
phra
gmiu
mu
lmari
ae
(DC
.)L
ink
AF
4262
1951
2M
aier
etal
(200
3)A
Y125
401
1698
Win
gfie
ldet
al(2
004)
Tu
berc
uli
na
maxi
ma
Ro
str.
[1]
AY2
2204
411
19L
utz
etal
(200
4b)
Tu
berc
uli
na
maxi
ma
[2]
AY2
9240
811
08L
utz
etal
(200
4a)
Tu
berc
uli
na
sbro
zzii
Cav
ara
&Sa
cc.
[1]
AY2
2204
559
3L
utz
etal
(200
4b)
Tu
berc
uli
na
sbro
zzii
[2]
AY2
9242
559
3L
utz
etal
(200
4a)
Uro
myc
esappen
dic
ula
tus
(Per
s.ex
Per
s.)
Un
ger*
AY7
4570
413
68M
ath
eny
etal
(200
5)u
np
ub
lish
edD
Q35
4510
1775
Aim
e(2
006)
Ust
ilen
tylo
ma
flu
itan
s(L
iro
)V
anky
AF
0098
8257
9B
eger
ow
etal
(199
7)A
Y124
481
2059
Bau
eret
al(2
003)
aas
Ben
sin
gton
iaphyl
ladu
sY.
Yam
ada,
Nak
agaw
a&
Van
der
Wal
t.bas
Zym
oxen
oglo
eaer
iophor
iD
.J.
McL
augh
lin
&D
ou
ble
s.cas
Rhod
otor
ula
crea
tin
ivor
aG
olu
bev
.d
asR
hod
otor
ula
cass
iico
la(S
jam
suri
dza
let
alu
np
ub
lish
ed).
eas
Rhod
otor
ula
fush
an
ensi
s(N
akas
eet
alu
np
ub
lish
ed).
fas
Rhod
otor
ula
nym
phaea
e(S
jam
suri
dza
let
alu
np
ub
lish
ed).
gas
Rhod
otor
ula
sam
an
eae
(Sja
msu
rid
zal
etal
un
pu
bli
shed
).h
25S
isfr
om
Sep
toba
sidiu
msp
.;18
Sis
fro
mS.
can
esce
ns.
ias
Hel
icob
asi
diu
mpu
rpu
reu
m.
SUP
PL
EM
EN
TA
RY
TA
BL
EII
.C
on
tin
ued

LITERATURE CITED
Aime MC. 2006. Toward resolving family-level relationshipsin rust fungi (Uredinales). Mycoscience 47:112–122.
Alvarez-Rodriguez ML, Belloch C, Villa M, Uruburu F,Larriba G, Coque JJ. 2003. Degradation of vanillic acidand production of guaiacol by microorganisms isolatedfrom cork samples. FEMS Microbiol Lett 220:49–55.
Bai FY, Cai Y, Wang QM, Ohkubo H. 2004. Rhodotorulaoryzae sp. Nov., a novel basidiomycetous yeast speciesisolated from paddy rice. Antonie Van Leeuwenhoek86:295–299.
———, Takashima M, Hamamoto M, Nakase T. 2001.Sporobolomyces yunnanensis sp. nov., a Q-10(H2)-con-taining yeast species with a close phylogenetic relation-ship to Erythrobasidium hasegawianum. Int J Syst EvolMicrobiol 51:231–235.
Bauer R, Begerow D, Oberwinkler F, Marvanova L. 2003.Classicula: the teleomorph of Naiadella fluitans.Mycologia 95:756–764.
Bayev A, Georgiev OI, Hadjiolov AA, Nikolaev N, SkryabinKG, Zakharyev VM. 1981. The structure of the yeastribosomal RNA genes. 3. Precise mapping of the 18 Sand 25 S rRNA genes and structure of the adjacentregions. Nucleic Acid Res 9:789–799.
Begerow D, Bauer R, Oberwinkler F. 1997. Phylogeneticstudies on nuclear LSU rDNA sequences of smut fungiand related taxa. Can J Bot 75:2045–2056.
Berres ME, Szabo LJ, McLaughlin DJ. 1995. Phylogeneticrelationships in auriculariaceous basidiomycetes basedon 25S ribosomal DNA sequences. Mycologia 87:821–840.
Bhattacharya D, Lutzoni F, Reeb V, Simon D, Nason J,Fernandez F. 2000. Widespread occurrence of spliceo-somal introns in the rDNA genes of ascomycetes. MolBiol Evol 17:1971–1984.
Bruns TD, Vilgalys R, Barns SM, Gonzalez D, Hibbett DS,Lane DJ, Simon L, Stickel S, Szaro TM, Weisburg WG,Sogin ML. 1992. Evolutionary relationships within theFungi: analyses of nuclear small subunit rRNA se-quences. Mol Phylog Evol 1:231–241.
Cai J, Roberts IN, Collins MD. 1996. Phylogenetic relation-ships among members of the ascomycetous yeastgenera Brettanomyces, Debaryomyces, Dekkera, and Kluy-veromyces deduced by small-subunit rRNA gene se-quences. Int J Syst Bacteriol 46:542–549.
de Wachter R, Neefs JM, Goris A, van de Peer Y. 1992. Thegene coding for small ribosomal subunit RNA in thebasidiomycete Ustilago maydis contains a group Iintron. Nucleic Acid Res 20:1251–1257.
Fell JW, Blatt GM, Statzell-Tallman A. 1998. Validation ofthe basidiomycetous yeast, Sporidiobolus microsporus sp.nov., based on phenotypic and molecular analyses.Antonie Van Leeuwenhoek 74:265–270.
———, Boekhout T, Fonseca A, Scorzetti G, Statzell-Tallman A. 2000. Biodiversity and systematics ofbasidiomycetous yeasts as determined by large-subunitrDNA D1/D2 domain sequence analysis. Int J Syst EvolMicrobiol 50:1351–1371.
———, Roeijmans H, Boekhout T. 1999. Cystofilobasidiales,
a new order of basidiomycetous yeasts. Int J SystBacteriol 49:907–913.
Hamamoto M, Nakase T. 2000. Phylogenetic analysis of theballistoconidium-forming yeast genus Sporobolomycesbased on 18S rDNA sequences. Int J Syst Evol Microbiol50:1373–1380.
Hendriks L, de Baere R, van de Peer Y, Neefs J, Goris A, deWachter R. 1991. The evolutionary position of therhodophyte Porphyra umbilicalis and the basidiomyceteLeucosporidium scottii among other eukaryotes asdeduced from complete sequences of small ribosomalsubunit RNA. J Mol Evol 32:167–177.
Hong SG, Chun J, Nam JS, Park Y-D, Bae KS. 2000.Phylogenetic analysis of the genus Sporobolomyces basedon partial sequences of 26S rDNA. J Microbiol Bio-technol 10:363–366.
Kirschner R, Begerow D, Oberwinkler F. 2001. A newChionosphaera species associated with conifer inhabit-ing bark beetles. Mycol Res 105:1403–1408.
Landvik S, Eriksson OE, Gargas A, Gustafsson P. 1993.Relationships of the genus Neolecta (Neolectales ordo.nov., Ascomycotina) inferred from 18S rDNA se-quences. Sys Ascomycetum 11:107–118.
Lutz M, Bauer R, Begerow D, Oberwinkler F. 2004a.Tuberculina - Thanatophytum/Rhizoctonia crocorum -Helicobasidium: A unique mycoparasitic-phytoparasiticlife strategy. Mycol Res 108:227–238.
———, ———, ———, ———, Triebel D. 2004b. Tubercu-lina: rust relative attacks rusts. Mycologia 96:614–626.
Lutzoni F, Kauff F, Cox CJ, McLaughlin D, Celio G,Dentinger B, Padamsee M, Hibbett D, James TY,Baloch E, Grube M, Reeb V, Hofstetter V, Schoch C,Arnold AE, Miadlikowska J, Spatafora J, Johnson D,Hambleton S, Crockett M, Shoemaker R, Sung G-H,Lucking R, Lumbsch T, O’Donnell K, Binder M,Diederich P, Ertz D, Gueidan C, Hansen K, HarrisRC, Hosaka K, Lim Y-W, Matheny PB, Nishida H, PfisterD, Rogers J, Rossman A, Schmitt I, Sipman H, Stone J,Sugiyama J, Yahr R, Vilgalys R. 2004. Assembling theFungal Tree of Life: progress, classification, andevolution of subcellular traits. Am J Bot 91:1446–1480.
Maier W, Begerow D, Weiß M, Oberwinkler F. 2003.Phylogeny of the rust fungi: an approach using nuclearlarge subunit ribosomal DNA sequences. Can J Bot 81:12–23.
McLaughlin DJ, Hanson RW Jr, Frieders EM, Swann EC,Szabo LJ. 2004. Mitosis in the yeast phase of thebasidiomycetes Bensingtonia yuccicola and Stilbumvulgare and its phylogenetic implications. Am J Bot91:808–815.
Nishida H, Ando K, Ando Y, Hirata A, Sugiyama J. 1995.Mixia osmundae: transfer from the Ascomycota to theBasidiomycota based on evidence from molecules andmorphology. Can J Bot 73(Suppl 1):S660–S666.
Piepenbring M, Stoll M, Oberwinkler F. 2002. The genericposition of Ustilago maydis, Ustilago scitaminea, andUstilago esculenta (Ustilaginales). Mycol Progress 1:71–80.
Rubtsov PM, Musakhanov MM, Zakharyev VM, Krayev AS,Skryabin KG, Bayev AA. 1980. The structure of the yeast

ribosomal RNA genes. I. The complete nucleotidesequence of the 18S ribosomal RNA gene fromSaccharomyces cerevisiae. Nucleic Acid Res 8:5779–5794.
Sampaio JP, Bauer R, Begerow D, Oberwinkler F. 1999a.Occultifur externus sp. nov., a new species of a simple-pored auricularioid heterobasidiomycete from plantlitter in Portugal. Mycologia 91:1094–1101.
———, Fell JW, Gadanho M, Bauer R. 1999b. Kurtzmano-myces insolitus sp. nov., a new anamorphic heterobasi-diomycetous yeast species. Syst Appl Microbiol 22:619–625.
———, Gadanho M, Bauer R, Weiß M. 2003. Taxonomicstudies in the Microbotryomycetidae: Leucosporidiumgolubevii sp. nov., Leucosporidiella gen. nov. and thenew orders Leucosporidiales and Sporidiobolales.Mycol Prog 2:53–68.
Scorzetti G, Fell JW, Fonseca A, Statzell-Tallman A. 2002.Systematics of basidiomycetous yeasts: a comparison oflarge subunit D1/D2 and internal transcribed spacerrDNA regions. FEMS Yeast Res 2:495–517.
Sikaroodi M, Lawrey JD, Hawksworth DL, DePriest PT. 2001.The phylogenetic position of selected lichenicolousfungi: Hobsonia, Illosporium, and Marchandiomyces.Mycol Res 105:453–460.
Sjamsuridzal W, Nishida H, Ogawa H, Kakishima M,Sugiyama J. 1999. Phylogenetic positions of rust fungiparasitic on ferns: Evidence from 18S rDNA sequenceanalysis. Mycoscience 40:21–27.
———, ———, Yokota A. 2002. Phylogenetic position ofMixia osmundae inferred from 28S rDNA comparison.J Gen Appl Microbiol 48:121–123.
Stoll M, Begerow D, Oberwinkler F. 2005. Molecularphylogeny of Ustilago, Sporisorium, and related taxabased on combined analyses of rDNA sequences. MycolRes 109:342–356.
Sugita T, Takashima M, Hamamoto M, Budhaka P, NakaseT. 1997. Bensingtonia sakaguchii sp. nov. isolated froma leaf of Bischofia javanica in the Ogasawara Island.J Gen Appl Microbiol 43:231–235.
Suh S, Sugiyama J. 1993. Phylogeny among the basidiomy-cetous yeasts inferred from small subunit ribosomalDNA sequence. J Gen Microbiol 139:1595–1598.
———, Takashima M, Nakase T. 1996a. Phylogenetic studyof the anamorphic basidiomycetous yeasts Rhodotorulalactosa and R. minuta, and related taxa based on 18Sribosomal DNA sequences. J Gen Appl Microbiol 42:1–6.
———, Takematsu A, Takashima M, Nakase T. 1996b.Molecular phylogenetic study on stalked conidium-forming yeasts and related basidiomycetous yeast taxabased on 18S rDNA sequences. Microbiol Cult Coll 12:79–86.
Swann EC, Frieders EM, McLaughlin DJ. 1999. Microbo-tryum, Kriegeria and the changing paradigm in basid-iomycete classification. Mycologia 91:51–66.
———, Taylor JW. 1993. Higher taxa of basidiomycetes: an18S rRNA gene perspective. Mycologia 85:923–936.
———, ———. 1995. Phylogenetic perspectives on basidio-mycete systematics: evidence from the 18S rRNA gene.Can J Bot 73:S862–S868.
Takashima M, Hamamoto M, Nakase T. 2000. Taxonomicsignificance of fructose in the class Urediniomycetes:distribution of fructose in cell wall and phylogeny ofurediniomycetous yeasts. Syst Appl Microbiol 23:63–70.
———, Nakase T. 1996. A phylogenetic study of the genusTilletiopsis, Tilletiaria anomala and related taxa basedon the small subunit ribosomal DNA sequences. J GenAppl Microbiol 42:421–429.
———, ———. 2000. Four new species of the genusSporobolomyces isolated from leaves in Thailand. My-coscience 41:357–369.
———, Suh S, Nakase T. 1995a. Phylogenetic relationshipsamong the species of the genus Bensingtonia andrelated taxa based on the small subunit ribosomal DNAsequences. J Gen Appl Microbiol 41:131–141.
———, ———, ———. 1995b. Bensingtonia musae sp. nov.isolated from a dead leaf of Musa paradisiaca and itsphylogenetic relationship among basidiomycetousyeasts. J Gen Appl Microbiol 41:143–151.
Valerio E, Gadanho M, Sampaio JP. 2002. Sporobolomycesodoratus sp. nov., a new species in the Sporidiobolusruineniae clade. FEMS Yeast Res 2:9–16.
van de Peer Y, Hendriks L, Goris A, Neefs A, VancanneyetM, Kersters K, Berny JF, Hennebert GL, de Wachter R.1992. Evolution of basidiomycetous yeasts as deducedfrom small ribosomal subunit RNA sequences. SystAppl Microbiol 15:250–258.
Wang QM, Bai FY, Zhao JH, Jia JH. 2003. Bensingtoniachangbaiensis sp. nov. and Bensingtonia sorbi sp. nov.,novel ballistoconidium-forming yeast species fromplant leaves. Int J Syst Evol Microbiol 53:2085–2089.
Wilmotte AM, Van de Peer Y, Goris A, Chapelle S, De BaereR, Nelissen B, Neefs JM, Hennebert GL, De Wachter R.1993. Evolutionary relationships among higher fungiinferred by small ribosomal subunit RNA sequenceanalysis. Syst Appl Microbiol 16:436–444.
Wingfield BD, Ericson L, Szaro T, Burdon JJ. 2004.Phylogenetic patterns in the Uredinales. AustralasPlant Pathol 33:327–335.
Zhao J-H, Bai F-Y, Guo L-D, Jia J-H. 2002. Rhodotorulapinicola sp. nov., a basidiomycetous yeast speciesisolated from xylem of pine twigs. FEMS Yeast Res 2:159–163.

SUPPLEMENTARY FIG. 2A (Expanded). Phylogenetic relationships of Pucciniomycotina based on analyses of Dataset 1 (128OTUs, each with nuc-lsu rDNA and nuc-ssu rDNA sequences). Phylogram representing one of 36 equally most parsimonioustrees (8215 steps, CI 5 0.345, RI 5 0.731). Nodes with asterisks collapse in the strict consensus of equally most parsimonioustrees. Support values include MP bootstrap frequencies above 70% (first value, before slash), NJ bootstrap frequencies above70% (second value) and Bayesian posterior probabilities above 0.70. A. Ascomycete outgroups, Ustilaginomycotina,Agaricomycotina, and Pucciniomycotina p.p.

SUPPLEMENTARY FIG. 2B (Expanded). Phylogenetic relationships of Pucciniomycotina based on analyses of Dataset 1 (128OTUs, each with nuc-lsu rDNA and nuc-ssu rDNA sequences). Phylogram representing one of 36 equally most parsimonioustrees (8215 steps, CI 5 0.345, RI 5 0.731). Nodes with asterisks collapse in the strict consensus of equally most parsimonioustrees. Support values include MP bootstrap frequencies above 70% (first value, before slash), NJ bootstrap frequencies above70% (second value) and Bayesian posterior probabilities above 0.70. B. Pucciniomycotina p.p. The small cladogram depicts thetopology of Microbotryomycetidae obtained in Bayesian analyses, with posterior probabilities indicated only for stronglysupported nodes that conflict with the MP topology.

SUPPLEMENTARY FIG. 3A. Phylogenetic relationships in the Pucciniomycotina based on analyses of Dataset 2 (208 OTUs, 128with nuc-lsu rDNA and nuc-ssu rDNA sequences, 74 with only nuc-lsu rDNA sequences, indicated by ‘‘(lsu)’’ after taxon name,and 6 with only nuc-ssu rDNA sequences, indicated by ‘‘(ssu)’’ after taxon name). Strict consensus of 1000 equally mostparsimonious trees (9748 steps, CI50.31, RI50.729). MP bootstrap values above 70% are indicated. A. Ascomycete outgroups,Ustilaginomycotina, Agaricomycotina, and Pucciniomycotina p.p.

SUPPLEMENTARY FIG. 3B. Phylogenetic relationships in the Pucciniomycotina based on analyses of Dataset 2 (208 OTUs, 128with nuc-lsu rDNA and nuc-ssu rDNA sequences, 74 with only nuc-lsu rDNA sequences, indicated by ‘‘(lsu)’’ after taxon name,and 6 with only nuc-ssu rDNA sequences, indicated by ‘‘(ssu)’’ after taxon name). Strict consensus of 1000 equally mostparsimonious trees (9748 steps, CI50.31, RI50.729). MP bootstrap values above 70% are indicated. B. Pucciniomycotina p.p.

SUPPLEMENTARY FIG. 4. Life cycle of Septobasidium meredithae (Septobasidiales). A. Transversely septate basidium givesrise to forcibly discharged spores. B. Spores are subdivided by septa, and each section gives rise to bud cells that also bud toform yeast colonies in culture. C. When spores encounter a young scale insect, the clusters of bud cells form hyphae thatpenetrate the cuticle of the scale insect and begin to form coiled structures called haustoria. Infected scale insect settles on thehost plant and becomes completely invaded by the fungus. The fungus grows out of the insect to form a mycelial mat thatcovers the host plant and adjacent uninfected scale insects, producing basidia along the upper surface. SUPPLEMENTARY FIG. 5.Life cycle of the rust Gymnosporangium juniperi-virginianae (Pucciniales). A. Teliospores germinate to produce basidia andhaploid basidiospores that are wind-borne to the leaf of a rosaceous host. B. Spermogonia containing haploid spermatia onthe adaxial leaf surface. C. The products from compatible spermatia fuse to form limited dikaryotic hyphae that emerge fromthe abaxial leaf surface forming spore-filled sori known as aecia. The aecia contain long chains of dikaryotic aeciospores thatare wind-borne to the alternate host. D. Juniperus spp. are infected with aeciospores, producing galls from which gelatinouscolumns of telia emerge. Karyogamy occurs within the teliospores. SUPPLEMENTARY FIG. 6. Life cycle of Rhodosporidiumtoruloides (Sporidiobolales). A. A transversely septate basidium gives rise to spores. B. The spores bud and persist as yeasts. C.Yeast cells of the proper mating types fuse via a thin hyphal connection to form a dikaryon. D. The dikaryon forms hyphae thatwill eventually give rise to teliospores.