animal behavior - wfisdanimal behavior concept outline 26.1 ethology focuses on the natural history...
TRANSCRIPT

533
26Animal Behavior
Concept Outline
26.1 Ethology focuses on the natural history ofbehavior.
Approaches to the Study of Behavior. Field biologistsfocus on evolutionary aspects of behavior.Behavioral Genetics. At least some behaviors aregenetically determined.
26.2 Comparative psychology focuses on how learninginfluences behavior.
Learning. Association plays a major role in learning.The Development of Behavior. Parent-offspringinteractions play a key role in the development of behavior.The Physiology of Behavior. Hormones influence manybehaviors, particularly reproductive ones.Behavioral Rhythms. Many behaviors are governed byinnate biological clocks.
26.3 Communication is a key element of many animalbehaviors.
Courtship. Animals use many kinds of signals to courtone another.Communication in Social Groups. Bees and other socialanimals communicate in complex ways.
26.4 Migratory behavior presents many puzzles.
Orientation and Migration. Animals use many cuesfrom the environment to navigate during migrations.
26.5 To what degree animals “think” is a subject oflively dispute.
Animal Cognition. It is not clear to what degree animals“think.”
Organisms interact with their environment in manyways. To understand these interactions, we need to
appreciate both the internal factors that shape the way ananimal behaves, as well as aspects of the external environ-ment that affect individuals and organisms. In this chapter,we explore the mechanisms that determine an animal’s be-havior (figure 26.1), as well as the ways in which behaviordevelops in an individual. In the next chapter, we will con-sider the field of behavioral ecology, which investigateshow natural selection has molded behavior through evolu-tionary time.
FIGURE 26.1Rearing offspring involves complex behaviors. Living in groupscalled prides makes lions better mothers. Females share theresponsibilities of nursing and protecting the pride’s young,increasing the probability that the youngsters will survive intoadulthood.

other males and to attract a female to reproduce; this isthe ultimate, or evolutionary, explanation for the male’svocalization.
The study of behavior has had a long history of contro-versy. One source of controversy has been the question ofwhether behavior is determined more by an individual’sgenes or its learning and experience. In other words, is be-havior the result of nature (instinct) or nurture (experi-ence)? In the past, this question has been considered an “ei-ther/or” proposition, but we now know that instinct andexperience both play significant roles, often interacting incomplex ways to produce the final behavior. The scientificstudy of instinct and learning, as well as their interrelation-ship, has led to the growth of several scientific disciplines,including ethology, behavioral genetics, behavioral neuro-science, and comparative psychology.
Ethology
Ethology is the study of the natural history of behavior.Early ethologists (figure 26.2) were trained in zoology andevolutionary biology, fields that emphasize the study of an-imal behavior under natural conditions. As a result of thistraining, they believed that behavior is largely instinctive,or innate—the product of natural selection. Because behav-ior is often stereotyped (appearing in the same way in dif-ferent individuals of a species), they argued that it must bebased on preset paths in the nervous system. In their view,these paths are structured from genetic blueprints andcause animals to show a relatively complete behavior thefirst time it is produced.
The early ethologists based their opinions on behav-iors such as egg retrieval by geese. Geese incubate theireggs in a nest. If a goose notices that an egg has beenknocked out of the nest, it will extend its neck toward theegg, get up, and roll the egg back into the nest with aside-to-side motion of its neck while the egg is tuckedbeneath its bill. Even if the egg is removed during re-trieval, the goose completes the behavior, as if driven bya program released by the initial sight of the egg outsidethe nest. According to ethologists, egg retrieval behavioris triggered by a sign stimulus (also called a key stimu-lus), the appearance of an egg out of the nest; a compo-nent of the goose’s nervous system, the innate releasingmechanism, provides the neural instructions for themotor program, or fixed action pattern (figure 26.3).More generally, the sign stimulus is a “signal” in the en-vironment that triggers a behavior. The innate releasingmechanism is the sensory mechanism that detects the sig-nal, and the fixed action pattern is the stereotyped act.
534 Part VII Ecology and Behavior
Approaches to the Study of BehaviorDuring the past two decades, the study of animal behaviorhas emerged as an important and diverse science thatbridges several disciplines within biology. Evolution, ecol-ogy, physiology, genetics, and psychology all have naturaland logical linkages with the study of behavior, each disci-pline adding a different perspective and addressing differ-ent questions.
Research in animal behavior has made major contribu-tions to our understanding of nervous system organization,child development, and human communication, as well asthe process of speciation, community organization, and themechanism of natural selection itself. The study of the be-havior of nonhuman animals has led to the identification ofgeneral principles of behavior, which have been applied,often controversially, to humans. This has changed the waywe think about the origins of human behavior and the waywe perceive ourselves.
Behavior can be defined as the way an organism re-sponds to stimuli in its environment. These stimuli mightbe as simple as the odor of food. In this sense, a bacterialcell “behaves” by moving toward higher concentrations ofsugar. This behavior is very simple and well-suited to thelife of bacteria, allowing these organisms to live and repro-duce. As animals evolved, they occupied different environ-ments and faced diverse problems that affected their sur-vival and reproduction. Their nervous systems andbehavior concomitantly became more complex. Nervoussystems perceive and process information concerning envi-ronmental stimuli and trigger adaptive motor responses,which we see as patterns of behavior.
When we observe animal behavior, we can explain it intwo different ways. First, we might ask how it all works,that is, how the animal’s senses, nerve networks, or inter-nal state provide a physiological basis for the behavior. Inthis way, we would be asking a question of proximatecausation. To analyze the proximate cause of behavior,we might measure hormone levels or record the impulseactivity of neurons in the animal. We could also ask whythe behavior evolved, that is, what is its adaptive value?This is a question concerning ultimate causation. Tostudy the ultimate cause of a behavior, we would attemptto determine how it influenced the animal’s survival or re-productive success. Thus, a male songbird may sing dur-ing the breeding season because it has a level of thesteroid sex hormone, testosterone, which binds to hor-mone receptors in the brain and triggers the productionof song; this would be the proximate cause of the malebird’s song. But the male sings to defend a territory from
26.1 Ethology focuses on the natural history of behavior.

Similarly, a frog unfolds its long, sticky tongue at thesight of a moving insect, and a male stickleback fish willattack another male showing a bright red underside. Suchresponses certainly appear to be programmed and in-stinctive, but what evidence supports the ethological viewthat behavior has an underlying neural basis?
Behavior as a Response to Stimuli in theEnvironment
In the example of egg retrieval behavior in geese, a goosemust first perceive that an egg is outside of the nest. To re-spond to this stimulus, it must convert one form of energywhich is an input to its visual system—the energy of pho-tons of light—into a form of energy its nervous system canunderstand and use to respond—the electrical energy of anerve impulse. Animals need to respond to other stimuli inthe environment as well. For an animal to orient from afood source back to its nest, it might rely on the position ofthe sun. To find a mate, an animal might use a particularchemical scent. The electromagnetic energy of light andthe chemical energy of an odor must be converted to theelectrical energy of a nerve impulse. This is done throughtransduction, the conversion of energy in the environmentto an action potential, and the first step in the processing ofstimuli perceived by the senses. For example, rhodopsin isresponsible for the transduction of visual stimuli.Rhodopsin is made of cis-retinal and the protein opsin.Light is absorbed by the visual pigment cis-retinal causing it
to change its shape to trans-retinal (see chapter 55). This inturn changes the shape of the companion protein opsin,and induces the first step in a cascade of molecular eventsthat finally triggers a nerve impulse. Sound, odor, andtastes are transduced to nerve impulses by similarprocesses.
Ethologists study behavior from an evolutionaryperspective, focusing on the neural basis of behaviors.
Chapter 26 Animal Behavior 535
FIGURE 26.2The founding fathers of ethology: Karl von Frisch, Konrad Lorenz, and Niko Tinbergen pioneered the study of behavioralscience. In 1973, they received the Nobel Prize in Physiology or Medicine for their path-making contributions. Von Frisch led the studyof honeybee communication and sensory biology. Lorenz focused on social development (imprinting) and the natural history ofaggression. Tinbergen examined the functional significance of behavior and was the first behavioral ecologist.
FIGURE 26.3Lizard prey capture. The complex series of movements of thetongue this chameleon uses to capture an insect represents a fixedaction pattern.
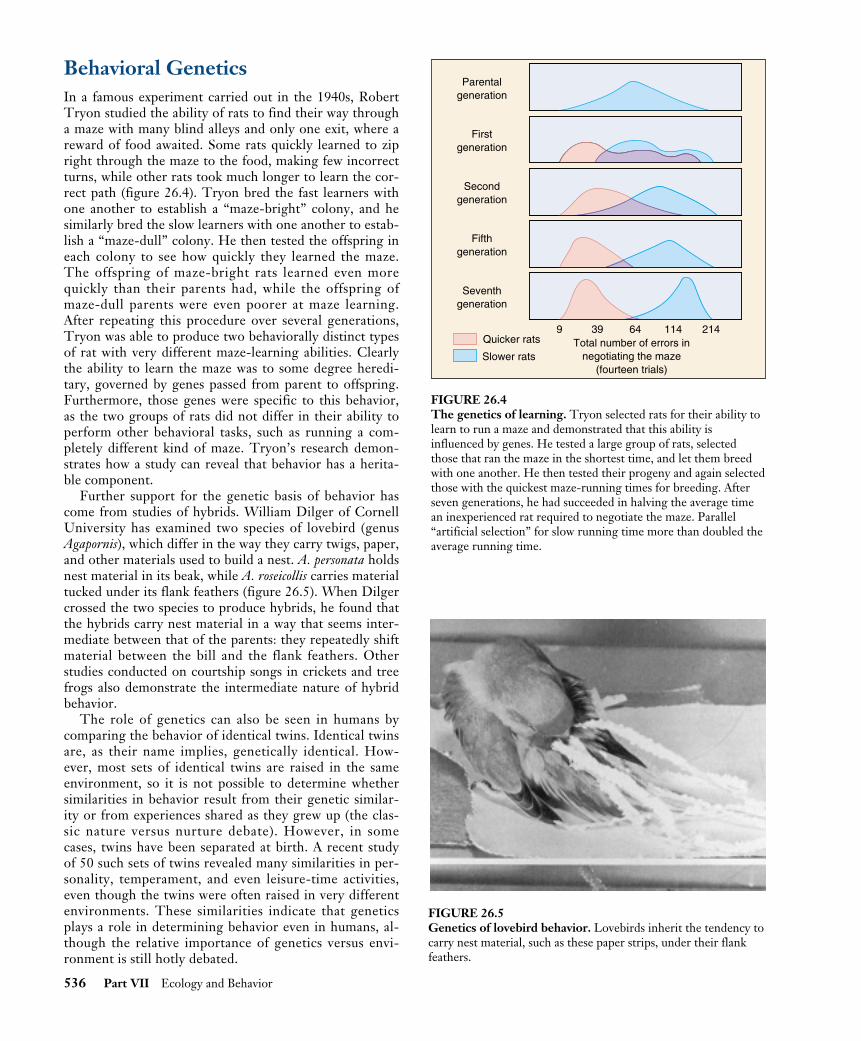
Behavioral GeneticsIn a famous experiment carried out in the 1940s, RobertTryon studied the ability of rats to find their way througha maze with many blind alleys and only one exit, where areward of food awaited. Some rats quickly learned to zipright through the maze to the food, making few incorrectturns, while other rats took much longer to learn the cor-rect path (figure 26.4). Tryon bred the fast learners withone another to establish a “maze-bright” colony, and hesimilarly bred the slow learners with one another to estab-lish a “maze-dull” colony. He then tested the offspring ineach colony to see how quickly they learned the maze.The offspring of maze-bright rats learned even morequickly than their parents had, while the offspring ofmaze-dull parents were even poorer at maze learning.After repeating this procedure over several generations,Tryon was able to produce two behaviorally distinct typesof rat with very different maze-learning abilities. Clearlythe ability to learn the maze was to some degree heredi-tary, governed by genes passed from parent to offspring.Furthermore, those genes were specific to this behavior,as the two groups of rats did not differ in their ability toperform other behavioral tasks, such as running a com-pletely different kind of maze. Tryon’s research demon-strates how a study can reveal that behavior has a herita-ble component.
Further support for the genetic basis of behavior hascome from studies of hybrids. William Dilger of CornellUniversity has examined two species of lovebird (genusAgapornis), which differ in the way they carry twigs, paper,and other materials used to build a nest. A. personata holdsnest material in its beak, while A. roseicollis carries materialtucked under its flank feathers (figure 26.5). When Dilgercrossed the two species to produce hybrids, he found thatthe hybrids carry nest material in a way that seems inter-mediate between that of the parents: they repeatedly shiftmaterial between the bill and the flank feathers. Otherstudies conducted on courtship songs in crickets and treefrogs also demonstrate the intermediate nature of hybridbehavior.
The role of genetics can also be seen in humans bycomparing the behavior of identical twins. Identical twinsare, as their name implies, genetically identical. How-ever, most sets of identical twins are raised in the sameenvironment, so it is not possible to determine whethersimilarities in behavior result from their genetic similar-ity or from experiences shared as they grew up (the clas-sic nature versus nurture debate). However, in somecases, twins have been separated at birth. A recent studyof 50 such sets of twins revealed many similarities in per-sonality, temperament, and even leisure-time activities,even though the twins were often raised in very differentenvironments. These similarities indicate that geneticsplays a role in determining behavior even in humans, al-though the relative importance of genetics versus envi-ronment is still hotly debated.
536 Part VII Ecology and Behavior
Parentalgeneration
Firstgeneration
Secondgeneration
Fifthgeneration
Seventhgeneration
Total number of errors innegotiating the maze
(fourteen trials)
9 39 64 114 214Quicker rats
Slower rats
FIGURE 26.4The genetics of learning. Tryon selected rats for their ability tolearn to run a maze and demonstrated that this ability isinfluenced by genes. He tested a large group of rats, selectedthose that ran the maze in the shortest time, and let them breedwith one another. He then tested their progeny and again selectedthose with the quickest maze-running times for breeding. Afterseven generations, he had succeeded in halving the average timean inexperienced rat required to negotiate the maze. Parallel“artificial selection” for slow running time more than doubled theaverage running time.
FIGURE 26.5Genetics of lovebird behavior. Lovebirds inherit the tendency tocarry nest material, such as these paper strips, under their flankfeathers.
Single Gene Effects on Behavior
The maze-learning, hybrid, and identical twins studies justdescribed suggest genes play a role in behavior, but recentresearch has provided much greater detail on the geneticbasis of behavior. In the fruit fly Drosophila, and in mice,many mutations have been associated with particular be-havioral abnormalities.
In fruit flies, for example, individuals that possess alter-native alleles for a single gene differ greatly in their feedingbehavior as larvae; larvae with one allele move around agreat deal as they eat, whereas individuals with the alterna-tive allele move hardly at all. A wide variety of mutations atother genes are now known in Drosophila which affect al-most every aspect of courtship behavior.
The ways in which genetic differences affect behaviorhave been worked out for several mouse genes. For example,some mice with one mutation have trouble remembering in-formation that they learned two days earlier about where ob-jects are located. This difference appears to result becausethe mutant mice do not produce the enzyme α-calcium-calmodulin-dependent kinase II, which plays an importantrole in the functioning of a part of the brain, the hippocam-pus, that is important for spatial learning.
Modern molecular biology techniques allow the role ofgenetics in behavior to be investigated with ever greaterprecision. For example, male mice genetically engineered(as “knock-outs”) to lack the ability to synthesize nitricoxide, a brain neurotransmitter, show increased aggressivebehavior.
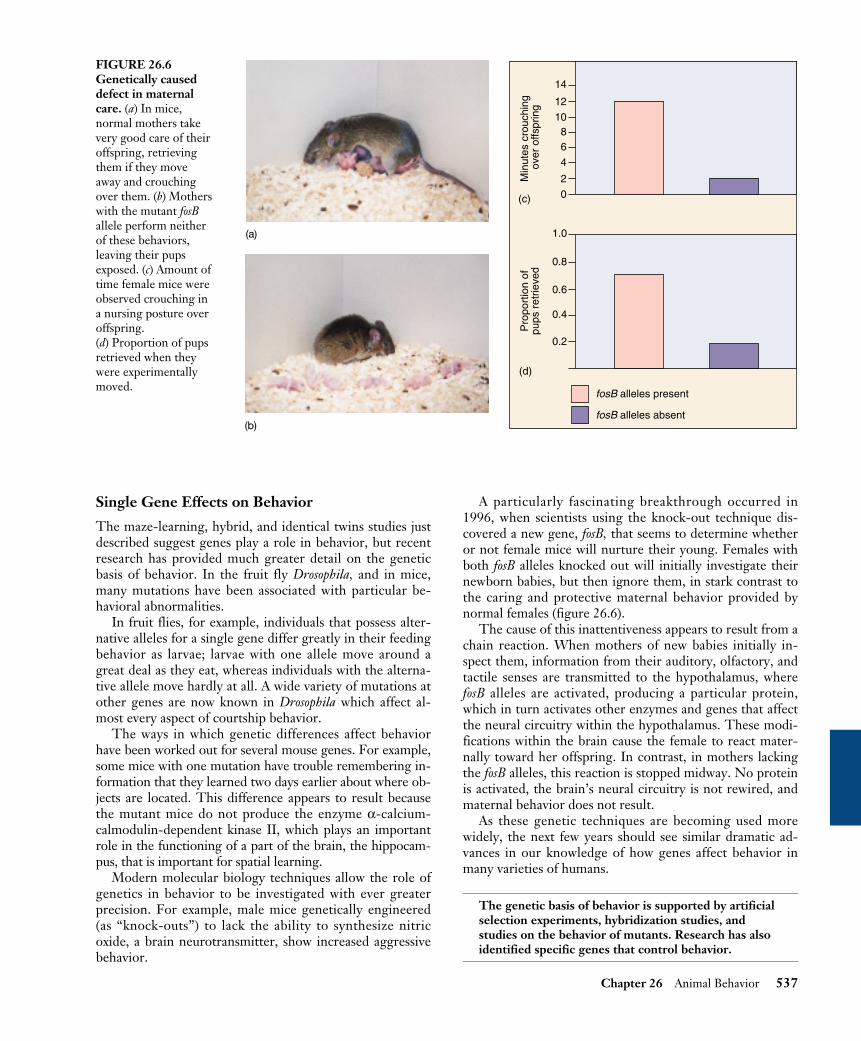
A particularly fascinating breakthrough occurred in1996, when scientists using the knock-out technique dis-covered a new gene, fosB, that seems to determine whetheror not female mice will nurture their young. Females withboth fosB alleles knocked out will initially investigate theirnewborn babies, but then ignore them, in stark contrast tothe caring and protective maternal behavior provided bynormal females (figure 26.6).
The cause of this inattentiveness appears to result from achain reaction. When mothers of new babies initially in-spect them, information from their auditory, olfactory, andtactile senses are transmitted to the hypothalamus, wherefosB alleles are activated, producing a particular protein,which in turn activates other enzymes and genes that affectthe neural circuitry within the hypothalamus. These modi-fications within the brain cause the female to react mater-nally toward her offspring. In contrast, in mothers lackingthe fosB alleles, this reaction is stopped midway. No proteinis activated, the brain’s neural circuitry is not rewired, andmaternal behavior does not result.
As these genetic techniques are becoming used morewidely, the next few years should see similar dramatic ad-vances in our knowledge of how genes affect behavior inmany varieties of humans.
The genetic basis of behavior is supported by artificialselection experiments, hybridization studies, andstudies on the behavior of mutants. Research has alsoidentified specific genes that control behavior.
Chapter 26 Animal Behavior 537
14
12
1086
4
2
0
1.0
0.8
0.6
0.4
0.2
(c)
(d)
fosB alleles present
fosB alleles absent
Min
utes
crou
chin
gov
erof
fspr
ing
Pro
port
ion
ofpu
psre
trie
ved
(a)
(b)
FIGURE 26.6Genetically causeddefect in maternalcare. (a) In mice,normal mothers takevery good care of theiroffspring, retrievingthem if they moveaway and crouchingover them. (b) Motherswith the mutant fosBallele perform neitherof these behaviors,leaving their pupsexposed. (c) Amount oftime female mice wereobserved crouching ina nursing posture overoffspring.(d) Proportion of pupsretrieved when theywere experimentallymoved.
LearningWhile ethologists were attempting to explain behavior asan instinctive process, comparative psychologists focusedheavily on learning as the major element that shapes behav-ior. These behavioral scientists, working primarily on ratsin laboratory settings, identified the ways in which animalslearn. Learning is any modification of behavior that resultsfrom experience rather than maturation.
The simplest type of learning, nonassociative learn-ing, does not require an animal to form an associationbetween two stimuli or between a stimulus and a re-sponse. One form of nonassociative learning is habitua-tion, which can be defined as a decrease in response to arepeated stimulus that has no positive or negative conse-quences (that is, no reinforcement). In many cases, thestimulus evokes a strong response when it is first encoun-tered, but the magnitude of the response gradually de-clines with repeated exposure. For example, young birdssee many types of objects moving overhead. At first, theymay respond by crouching down and remaining still.Some of the objects, like falling leaves or members oftheir own species flying by, are seen very frequently andhave no positive or negative consequence to thenestlings. Over time, the young birds may habituate tosuch stimuli and stop responding. Thus, habituation canbe thought of as learning not to respond to a stimulus.Being able to ignore unimportant stimuli is critical to ananimal confronting a barrage of stimuli in a complex en-vironment. Another form of nonassociative learning issensitization, characterized by an increased responsive-ness to a stimulus. Sensitization is essentially the oppositeof habituation.
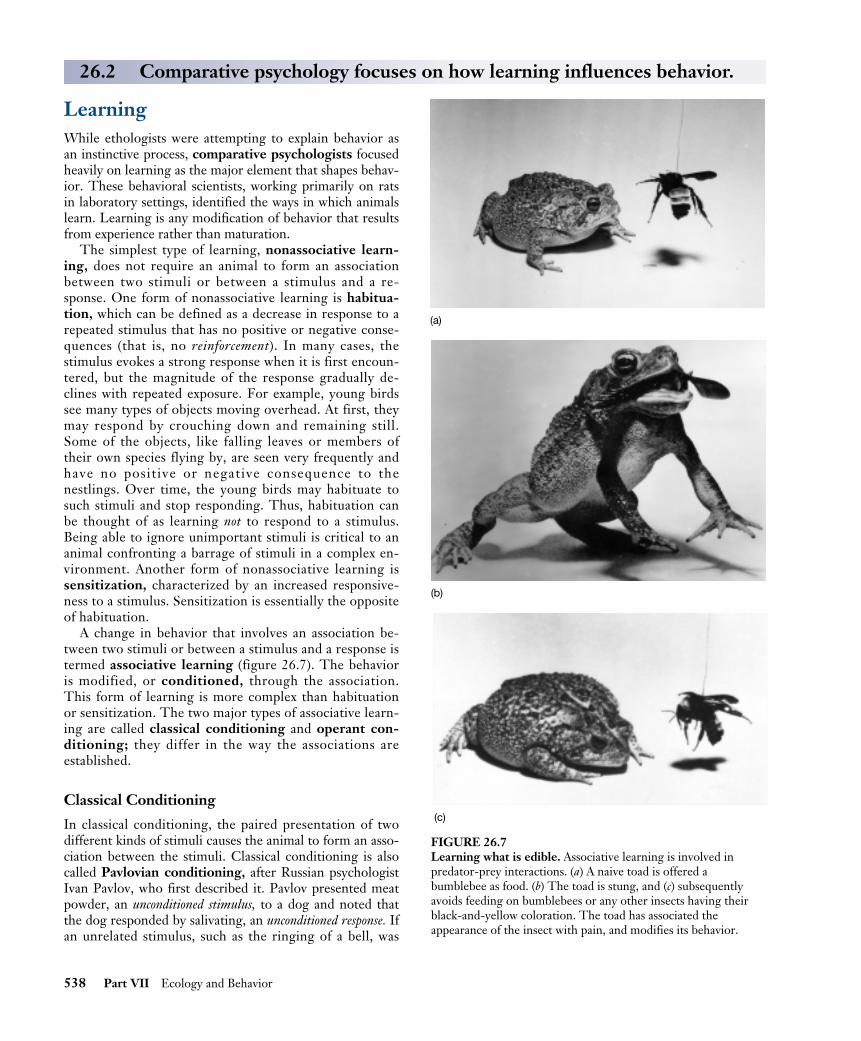
A change in behavior that involves an association be-tween two stimuli or between a stimulus and a response istermed associative learning (figure 26.7). The behavioris modified, or conditioned, through the association.This form of learning is more complex than habituationor sensitization. The two major types of associative learn-ing are called classical conditioning and operant con-ditioning; they differ in the way the associations areestablished.
Classical Conditioning
In classical conditioning, the paired presentation of twodifferent kinds of stimuli causes the animal to form an asso-ciation between the stimuli. Classical conditioning is alsocalled Pavlovian conditioning, after Russian psychologistIvan Pavlov, who first described it. Pavlov presented meatpowder, an unconditioned stimulus, to a dog and noted thatthe dog responded by salivating, an unconditioned response. Ifan unrelated stimulus, such as the ringing of a bell, was
538 Part VII Ecology and Behavior
26.2 Comparative psychology focuses on how learning influences behavior.
FIGURE 26.7Learning what is edible. Associative learning is involved inpredator-prey interactions. (a) A naive toad is offered abumblebee as food. (b) The toad is stung, and (c) subsequentlyavoids feeding on bumblebees or any other insects having theirblack-and-yellow coloration. The toad has associated theappearance of the insect with pain, and modifies its behavior.
(a)
(b)
(c)

presented at the same time as the meat powder, over re-peated trials the dog would salivate in response to thesound of the bell alone. The dog had learned to associatethe unrelated sound stimulus with the meat powder stimu-lus. Its response to the sound stimulus was, therefore, con-ditioned, and the sound of the bell is referred to as a condi-tioned stimulus.
Operant Conditioning
In operant conditioning, an animal learns to associate itsbehavioral response with a reward or punishment. Ameri-can psychologist B. F. Skinner studied operant condition-ing in rats by placing them in an apparatus that came to becalled a “Skinner box.” As the rat explored the box, itwould occasionally press a lever by accident, causing a pel-let of food to appear. At first, the rat would ignore thelever, eat the food pellet, and continue to move about.Soon, however, it learned to associate pressing the lever(the behavioral response) with obtaining food (the reward).When it was hungry, it would spend all its time pressingthe lever. This sort of trial-and-error learning is of majorimportance to most vertebrates.
Comparative psychologists used to believe that any twostimuli could be linked in classical conditioning and thatanimals could be conditioned to perform any learnablebehavior in response to any stimulus by operant condi-tioning. As you will see below, this view has changed.Today, it is thought that instinct guides learning by deter-mining what type of information can be learned throughconditioning.
Instinct
It is now clear that some animals have innate predisposi-tions toward forming certain associations. For example, if arat is offered a food pellet at the same time it is exposed toX rays (which later produces nausea), the rat will rememberthe taste of the food pellet but not its size. Conversely, if arat is given a food pellet at the same time an electric shockis delivered (which immediately causes pain), the rat will re-member the size of the pellet but not its taste. Similarly, pi-geons can learn to associate food with colors but not withsounds; on the other hand, they can associate danger withsounds but not with colors.
These examples of learning preparedness demon-strate that what an animal can learn is biologically influ-enced—that is, learning is possible only within the bound-aries set by instinct. Innate programs have evolvedbecause they underscore adaptive responses. Rats, whichforage at night and have a highly developed sense ofsmell, are better able to identify dangerous food by itsodor than by its size or color. The seed a pigeon eats mayhave a distinctive color that the pigeon can see, but itmakes no sound the pigeon can hear. The study of learn-
ing has expanded to include its ecological significance, sothat we are now able to consider the “evolution of learn-ing.” An animal’s ecology, of course, is key to understand-ing what an animal is capable of learning. Some species ofbirds, like Clark’s nutcracker, feed on seeds. Birds storeseeds in caches they bury when seeds are abundant so theywill have food during the winter. Thousands of seedcaches may be buried and then later recovered. Onewould expect the birds to have an extraordinary spatialmemory, and this is indeed what has been found (figure26.8). Clark’s nutcracker, and other seed-hoarding birds,have an unusually large hippocampus, the center formemory storage in the brain (see chapter 54).
Habituation and sensitization are simple forms oflearning in which there is no association betweenstimuli and responses. In contrast, associative learning(classical and operant conditioning) involves theformation of an association between two stimuli orbetween a stimulus and a response.
Chapter 26 Animal Behavior 539
FIGURE 26.8The Clark’s nutcracker has an extraordinary memory. AClark’s nutcracker can remember the locations of up to 2000 seedcaches months after hiding them. After conducting experiments,scientists have concluded that the birds use features of thelandscape and other surrounding objects as spatial references tomemorize the locations of the caches.

The Development ofBehaviorBehavioral biologists now recognizethat behavior has both genetic andlearned components, and the schoolsof ethology and psychology are lesspolarized than they once were. Thusfar in this chapter we have discussedthe influence of genes and learningseparately. As we will see, these factorsinteract during development to shapebehavior.
Parent-Offspring Interactions
As an animal matures, it may form so-cial attachments to other individuals orform preferences that will influencebehavior later in life. This process,called imprinting, is sometimes con-sidered a type of learning. In filial im-printing, social attachments form be-tween parents and offspring. Forexample, young birds of some speciesbegin to follow their mother within afew hours after hatching, and their fol-lowing response results in a bond be-tween mother and young. However,the young birds’ initial experience de-termines how this imprint is estab-lished. The German ethologist Kon-rad Lorenz showed that birds will follow the first objectthey see after hatching and direct their social behavior to-ward that object. Lorenz raised geese from eggs, and whenhe offered himself as a model for imprinting, the goslingstreated him as if he were their parent, following him duti-fully (figure 26.9). Black boxes, flashing lights, and water-ing cans can also be effective imprinting objects (figure26.10). Imprinting occurs during a sensitive phase, or acritical period (roughly 13 to 16 hours after hatching ingeese), when the success of imprinting is highest.
Several studies demonstrate that the social interactionsthat occur between parents and offspring during the criticalperiod are key to the normal development of behavior. Thepsychologist Harry Harlow gave orphaned rhesus monkeyinfants the opportunity to form social attachments with twosurrogate “mothers,” one made of soft cloth covering awire frame and the other made only of wire. The infantschose to spend time with the cloth mother, even if only thewire mother provided food, indicating that texture and tac-tile contact, rather than providing food, may be among thekey qualities in a mother that promote infant social attach-ment. If infants are deprived of normal social contact, theirdevelopment is abnormal. Greater degrees of deprivationlead to greater abnormalities in social behavior during
childhood and adulthood. Studies on orphaned human in-fants suggest that a constant “mother figure” is required fornormal growth and psychological development.
Recent research has revealed a biological need for thestimulation that occurs during parent-offspring interactionsearly in life. Female rats lick their pups after birth, and thisstimulation inhibits the release of an endorphin (see chap-ter 56) that can block normal growth. Pups that receivenormal tactile stimulation also have more brain receptorsfor glucocorticoid hormones, longer-lived brain neurons,and a greater tolerance for stress. Premature human infantswho are massaged gain weight rapidly. These studies indi-cate that the need for normal social interaction is based inthe brain and that touch and other aspects of contact be-tween parents and offspring are important for physical aswell as behavioral development.
Sexual imprinting is a process in which an individuallearns to direct its sexual behavior at members of its ownspecies. Cross-fostering studies, in which individuals ofone species are raised by parents of another species, revealthat this form of imprinting also occurs early in life. Inmost species of birds, these studies have shown that the fos-tered bird will attempt to mate with members of its fosterspecies when it is sexually mature.
540 Part VII Ecology and Behavior
(a)
(b)
FIGURE 26.9An unlikely parent. The eager goslingsfollowing Konrad Lorenz think he is theirmother. He is the first object they sawwhen they hatched, and they have usedhim as a model for imprinting.
FIGURE 26.10How imprinting is studied. Ducklingswill imprint on the first object they see,even (a) a black box or (b) a white sphere.

Interaction between Instinct and Learning
The work of Peter Marler and his colleagues on the ac-quisition of courtship song by white-crowned sparrowsprovides an excellent example of the interaction betweeninstinct and learning in the development of behavior.Courtship songs are sung by mature males and arespecies-specific. By rearing male birds in soundproof in-cubators provided with speakers and microphones, Marlercould control what a bird heard as it matured and recordthe song it produced as an adult. He found that white-crowned sparrows that heard no song at all during devel-opment, or that heard only the song of a different species,the song sparrow, sang a poorly developed song as adults(figure 26.11). But birds that heard the song of their ownspecies, or that heard the songs of both the white-crownedsparrow and the song sparrow, sang a fully developed,white-crowned sparrow song as adults. These results sug-gest that these birds have a genetic template, or instinc-tive program, that guides them to learn the appropriatesong. During a critical period in development, the tem-plate will accept the correct song as a model. Thus, songacquisition depends on learning, but only the song of thecorrect species can be learned. The genetic template forlearning is selective. However, learning plays a prominentrole as well. If a young white-crowned sparrow is surgi-cally deafened after it hears its species’ song during thecritical period, it will also sing a poorly developed song asan adult. Therefore, the bird must “practice” listening tohimself sing, matching what he hears to the model histemplate has accepted.
Although this explanation of song development stoodunchallenged for many years, recent research has shownthat white-crowned sparrow males can learn anotherspecies’ song under certain conditions. If a live malestrawberry finch is placed in a cage next to a young malesparrow, the young sparrow will learn to sing the straw-berry finch’s song! This finding indicates that socialstimuli may be more effective than a tape-recorded songin overriding the innate program that guides song devel-opment. Furthermore, the males of some bird specieshave no opportunity to hear the song of their ownspecies. In such cases, it appears that the males instinc-tively “know” their own species’ song. For example, cuck-oos are brood parasites; females lay their eggs in the nestof another species of bird, and the young that hatch arereared by the foster parents (figure 26.12). When thecuckoos become adults, they sing the song of their ownspecies rather than that of their foster parents. Becausemale brood parasites would most likely hear the song oftheir host species during development, it is adaptive forthem to ignore such “incorrect” stimuli. They hear noadult males of their own species singing, so no correctsong models are available. In these species, natural selec-tion has programmed the male with a genetically guidedsong.
Interactions that occur during sensitive phases ofimprinting are critical to normal behavioraldevelopment. Physical contact plays an important rolein the development of psychological well-being andgrowth.
Chapter 26 Animal Behavior 541
54321
6
4
2
Fre
quen
cy(k
Hz)
(a)
(b)0.5 1.0 1.5 2.0
Time (s)
FIGURE 26.11Song development in birds. (a) The sonograms of songsproduced by male white-crowned sparrows that had been exposedto their own species’ song during development are different from(b) those of male sparrows that heard no song during rearing. Thisdifference indicates that the genetic program itself is insufficientto produce a normal song.
FIGURE 26.12Brood parasite. Cuckoos lay their eggs in the nests of otherspecies of birds. Because the young cuckoos (large bird to theright) are raised by a different species (like this meadow pipit,smaller bird to the left), they have no opportunity to learn thecuckoo song; the cuckoo song they later sing is innate.

The Physiology ofBehaviorThe early ethologists’ emphasis on in-stinct sometimes overlooked the internalfactors that control behavior. If askedwhy a male bird defends a territory andsings only during the breeding season,they would answer that a bird singswhen it is in the right motivational stateor mood and has the appropriate drive.But what do these phrases mean in termsof physiological control mechanisms?
Part of our understanding of thephysiological control of behavior hascome from the study of reproductivebehavior. Animals show reproductivebehaviors such as courtship only duringthe breeding season. Research onlizards, birds, rats, and other animalshas revealed that hormones play an im-portant role in these behaviors. Changesin day length trigger the secretion ofgonadotropin-releasing hormone by thehypothalamus, which stimulates the re-lease of the gonadotropins, follicle-stimulating hormone (FSH) andluteinizing hormone, by the anterior pi-tuitary gland. These hormones causethe development of reproductive tissuesto ready the animal for breeding. Thegonadotropins, in turn, stimulate the se-cretion of the steroid sex hormones, es-trogens and progesterone in females andtestosterone in males. The sex hor-mones act on the brain to trigger behav-iors associated with reproduction. Forexample, birdsong and territorial behav-ior depend upon the level of testos-terone in the male, and the receptivityof females to male courtship dependsupon estrogen levels.
Hormones have both organizationaland activational effects. In the exampleof birdsong given above, estrogen in themale causes the development of thesong system, which is composed ofneural tissue in the forebrain and itsconnections to the spinal cord and thesyrinx (a structure like our larynx that allows the bird tosing). Early in a male’s development, the gonads produceestrogen, which stimulates neuron growth in the brain. Inthe mature male, the testes produce testosterone, whichactivates song. Thus, the development of the neural sys-tems that are responsible for behavior is first organized,then activated by hormones.
Research on the physiology of repro-ductive behavior shows that there areimportant interactions among hor-mones, behavior, and stimuli in both thephysical and social environments of anindividual. Daniel Lehrman’s work onreproduction in ring doves provides anexcellent example of how these factorsinteract (figure 26.13). Male courtshipbehavior is controlled by testosteroneand related steroid hormones. Themale’s behavior causes the release ofFSH in the female, and FSH promotesthe growth of the ovarian follicles (seechapter 59). The developing follicles re-lease estrogens, which affect other re-productive tissues. Nest constructionfollows after one or two days. The pres-ence of the nest then triggers the secre-tion of progesterone in the female, initi-ating incubation behavior after the eggis laid. Feeding occurs once the eggshatch, and this behavior is also hormon-ally controlled.
The research of Lehrman and hiscolleagues paved the way for many addi-tional investigations in behavioral en-docrinology, the study of the hormonalregulation of behavior. For example,male Anolis lizards begin courtship aftera seasonal rise in temperature, and themale’s courtship is needed to stimulatethe growth of ovarian follicles in the fe-male. These and other studies demon-strate the interactive effects of the phys-ical environment (for example,temperature and day length) and the so-cial environment (such as the presenceof a nest and the courtship display of amate) on the hormonal condition of ananimal. Hormones are, therefore, aproximate cause of behavior. To controlreproductive behavior, they must be re-leased when the conditions are most fa-vorable for the growth of young. Otherbehaviors, such as territoriality anddominance behavior, also have hormonalcorrelates.
Hormones may interact with neuro-transmitters to alter behavior. Estrogen affects the neuro-transmitter serotonin in female mice, and may be in partresponsible for the “mood swings” experienced by somehuman females during the menstrual cycle.
Hormones have important influences on reproductiveand social behavior.
542 Part VII Ecology and Behavior
(1)
(2)
(3)
(4)
(5)
FIGURE 26.13Hormonal control of reproductivebehavior. Reproduction in the ring doveinvolves a sequence of behaviorsregulated by hormones: (1) courtshipand copulation; (2) nest building; (3) egglaying; (4) incubation; and (5) feedingcrop milk to the young after they hatch.

Behavioral RhythmsMany animals exhibit behaviors that vary at regular inter-vals of time. Geese migrate south in the fall, birds sing inthe early morning, bats fly at night rather than during theday, and most humans sleep at night and are active in thedaytime. Some behaviors are timed to occur in concertwith lunar or tidal cycles (figure 26.14). Why do regular re-peating patterns of behavior occur, and what determineswhen they occur? The study of questions like these has re-vealed that rhythmic animal behaviors are based on bothendogenous (internal) rhythms and exogenous (external)timers.
Most studies of behavioral rhythms have focused on be-haviors that appear to be keyed to a daily cycle, such assleeping. Rhythms with a period of about 24 hours arecalled circadian (“about a day”) rhythms. Many of thesebehaviors have a strong endogenous component, as if theywere driven by a biological clock. Such behaviors are saidto be free-running, continuing on a regular cycle even in theabsence of any cues from the environment. Almost all fruitfly pupae hatch in the early morning, for example, even ifthey are kept in total darkness throughout their week-longdevelopment. They keep track of time with an internalclock whose pattern is determined by a single gene, calledthe per (for period ) gene. Different mutations of the pergene shorten or lengthen the daily rhythm. The per geneproduces a protein in a regular 24-hour cycle in the brain,serving as the fly’s pacemaker of activity. The protein prob-ably affects the expression of other genes that ultimatelyregulate activity. As the per protein accumulates, it seemsto turn off the gene. In mice, the clock gene is responsiblefor regulating the animal’s daily rhythm.
Most biological clocks do not exactly match the rhythmsof the environment. Therefore, the behavioral rhythm ofan individual deprived of external cues gradually drifts outof phase with the environment. Exposure to an environ-mental cue resets the biological clock and keeps the behav-ior properly synchronized with the environment. Light isthe most common cue for resetting circadian rhythms.
The most obvious circadian rhythm in humans is thesleep-activity cycle. In controlled experiments, humanshave lived for months in underground apartments, whereall light is artificial and there are no external cues whatso-ever indicating day length. Left to set their own schedules,most of these people adopt daily activity patterns (onephase of activity plus one phase of sleep) of about 25 hours,although there is considerable variation. Some individualsexhibit 50-hour clocks, active for as long as 36 hours duringeach period! Under normal circumstances, the day-nightcycle resets an individual’s free-running clock every day toa cycle period of 24 hours.
What constitutes an animal’s biological clock? In someinsects, the clock is thought to be located in the optic lobesof the brain, and timekeeping appears to be based on hor-mones. In mammals, including humans, the biological
clock lies in a specific region of the hypothalamus calledthe suprachiasmatic nucleus (SCN). The SCN is a self-sustaining oscillator, which means it undergoes sponta-neous, cyclical changes in activity. This oscillatory activityhelps the SCN to act as a pacemaker for circadian rhythms,but in order for the rhythms to be entrained to externallight-dark cycles, the SCN must be influenced by light. Infact, there are both direct and indirect neural projectionsfrom the retina to the SCN.
The SCN controls circadian rhythms by regulating thesecretion of the hormone melatonin by the pineal gland.During the daytime, the SCN suppresses melatonin secre-tion. Consequently, more melatonin is secreted over a 24-hour period during short days than during long days. Vari-ations in melatonin secretion thus serve as an indicator ofseasonal changes in day length, and these variations partici-pate in timing the seasonal reproductive behavior of manymammals. Disturbances in melatonin secretion may be par-tially responsible for the “jet-lag” people experience whenair travel suddenly throws their internal clocks out of regis-ter with the day-night cycle.
Many important behavioral rhythms have cycle periodslonger than 24 hours. For example, circannual behaviorssuch as breeding, hibernation, and migration occur on ayearly cycle. These behaviors seem to be largely timed byhormonal and other physiological changes keyed to ex-ogenous factors such as day length. The degree to whichendogenous biological clocks underlie circannual rhythmsis not known, as it is very difficult to perform constant-environment experiments of several years’ duration. Themechanism of the biological clock remains one of themost tantalizing puzzles in biology today.
Endogenous circadian rhythms have free-running cycleperiods of approximately 24 hours; they are entrainedto a more exact 24-hour cycle period by environmentalcues.
Chapter 26 Animal Behavior 543
FIGURE 26.14Tidal rhythm. Oysters open their shells for feeding when thetide is in and close them when the tide is out.

Much of the research in animal behavior is devoted to ana-lyzing the nature of communication signals, determininghow they are perceived, and identifying the ecological rolesthey play and their evolutionary origins.
CourtshipDuring courtship, animals produce signals to communicatewith potential mates and with other members of their ownsex. A stimulus-response chain sometimes occurs, inwhich the behavior of one individual in turn releases a be-havior by another individual (figure 26.15).
Courtship Signaling
A male stickleback fish will defend the nest it builds on thebottom of a pond or stream by attacking conspecific males(that is, males of the same species) that approach the nest.Niko Tinbergen studied the social releasers responsible forthis behavior by making simple clay models. He found thata model’s shape and degree of resemblance to a fish were
544 Part VII Ecology and Behavior
26.3 Communication is a key element of many animal behaviors.
Female giveshead-up displayto male
1
Male swims zigzagto female and then leads her to nest
Male showsfemale entrance to nest
2
3 4
5
Female enters nestand spawns whilemale stimulates tail Male enters nest
and fertilizes eggs
unimportant; any model with a red underside (like the un-derside of a male stickleback) could release the attack be-havior. Tinbergen also used a series of clay models todemonstrate that a male stickleback recognizes a female byher abdomen, swollen with eggs.
Courtship signals are often species-specific, limiting com-munication to members of the same species and thus play-ing a key role in reproductive isolation. The flashes of fire-flies (which are actually beetles) are such species-specificsignals. Females recognize conspecific males by their flashpattern (figure 26.16), and males recognize conspecific fe-males by their flash response. This series of reciprocal re-sponses provides a continuous “check” on the species iden-tity of potential mates.
Visual courtship displays sometimes have more than onecomponent. The male Anolis lizard extends and retracts hisfleshy and often colorful dewlap while perched on a branchin his territory (figure 26.17). The display thus involvesboth color and movement (the extension of the dewlap aswell as a series of lizard “push-ups”). To which componentof the display does the female respond? Experiments inwhich the dewlap color is altered with ink show that color isunimportant for some species; that is, a female can becourted successfully by a male with an atypically coloreddewlap.
FIGURE 26.15A stimulus-response chain. Stickleback courtship involves a sequence of behaviors leading to the fertilization of eggs.

Pheromones
Chemical signals also mediate interactions between malesand females. Pheromones, chemical messengers used forcommunication between individuals of the same species,serve as sex attractants among other functions in many ani-mals. Even the human egg produces a chemical attractantto communicate with sperm! Female silk moths (Bombyxmori) produce a sex pheromone called bombykol in a glandassociated with the reproductive system. Neurophysiologi-cal studies show that the male’s antennae contain numeroussensory receptors specific for bombykol. These receptorsare extraordinarily sensitive, enabling the male to respondbehaviorally to concentrations of bombykol as low as onemolecule in 1017 molecules of oxygen in the air!
Many insects, amphibians, and birds produce species-specific acoustic signals to attract mates. Bullfrog males callto females by inflating and discharging air from their vocalsacs, located beneath the lower jaw. The female can distin-guish a conspecific male’s call from the call of other frogsthat may be in the same habitat and mating at the sametime. Male birds produce songs, complex sounds composedof notes and phrases, to advertise their presence and to at-tract females. In many bird species, variations in the males’songs identify particular males in a population. In thesespecies, the song is individually specific as well as species-specific.
Level of Specificity
Why should different signals have different levels of speci-ficity? The level of specificity relates to the function ofthe signal. Many courtship signals are species-specific tohelp animals avoid making errors in mating that wouldproduce inviable hybrids or otherwise waste reproductiveeffort. A male bird’s song is individually specific because itallows his presence (as opposed to simply the presence ofan unidentifiable member of the species) to be recognizedby neighboring birds. When territories are being estab-lished, males may sing and aggressively confront neighbor-ing conspecifics to defend their space. Aggression carriesthe risk of injury, and it is energetically costly to sing.After territorial borders have been established, intrusionsby neighbors are few because the outcome of the contestshave already been determined. Each male then “knows”his neighbor by the song he sings, and also “knows” thatmale does not constitute a threat because they have alreadysettled their territorial contests. So, all birds in the popula-tion can lower their energy costs by identifying theirneighbors through their individualistic songs. In a similarway, mammals mark their territories with pheromones thatsignal individual identity, which may be encoded as ablend of a number of chemicals. Other signals, such as themobbing and alarm calls of birds, are anonymous, convey-ing no information about the identity of the sender. Thesesignals may permit communication about the presence of apredator common to several bird species.
Courtship behaviors are keyed to species-specific visual,chemical, and acoustic signals.
Chapter 26 Animal Behavior 545
1
2
3
4
5
6
7
8
9
FIGURE 26.16Firefly fireworks. The bioluminescent displays of these lampyridbeetles are species-specific and serve as behavioral mechanisms ofreproductive isolation. Each number represents the flash patternof a male of a different species.
FIGURE 26.17Dewlap display of a male Anolis lizard. Under hormonalstimulation, males extend their fleshy, colored dewlaps to courtfemales. This behavior also stimulates hormone release and egg-laying in the female.
Communication inSocial GroupsMany insects, fish, birds, and mam-mals live in social groups in which in-formation is communicated betweengroup members. For example, someindividuals in mammalian societiesserve as “guards.” When a predatorappears, the guards give an alarm call,and group members respond by seek-ing shelter. Social insects, such as antsand honeybees, produce alarmpheromones that trigger attack be-havior. Ants also deposit trailpheromones between the nest and afood source to induce cooperationduring foraging (figure 26.18). Honey-bees have an extremely complex dancelanguage that directs nestmates torich nectar sources.
The Dance Language of theHoneybee
The European honeybee, Apis mellifera,lives in hives consisting of 30,000 to40,000 individuals whose behaviors areintegrated into a complex colony.Worker bees may forage for miles fromthe hive, collecting nectar and pollenfrom a variety of plants and switchingbetween plant species and popula-tions on the basis of how energeti-cally rewarding their food is. Thefood sources used by bees tend tooccur in patches, and each patch of-fers much more food than a singlebee can transport to the hive. Acolony is able to exploit the resourcesof a patch because of the behavior ofscout bees, which locate patches andcommunicate their location to hivemates through a dancelanguage. Over many years, Nobel laureate Karl vonFrisch was able to unravel the details of this communica-tion system.
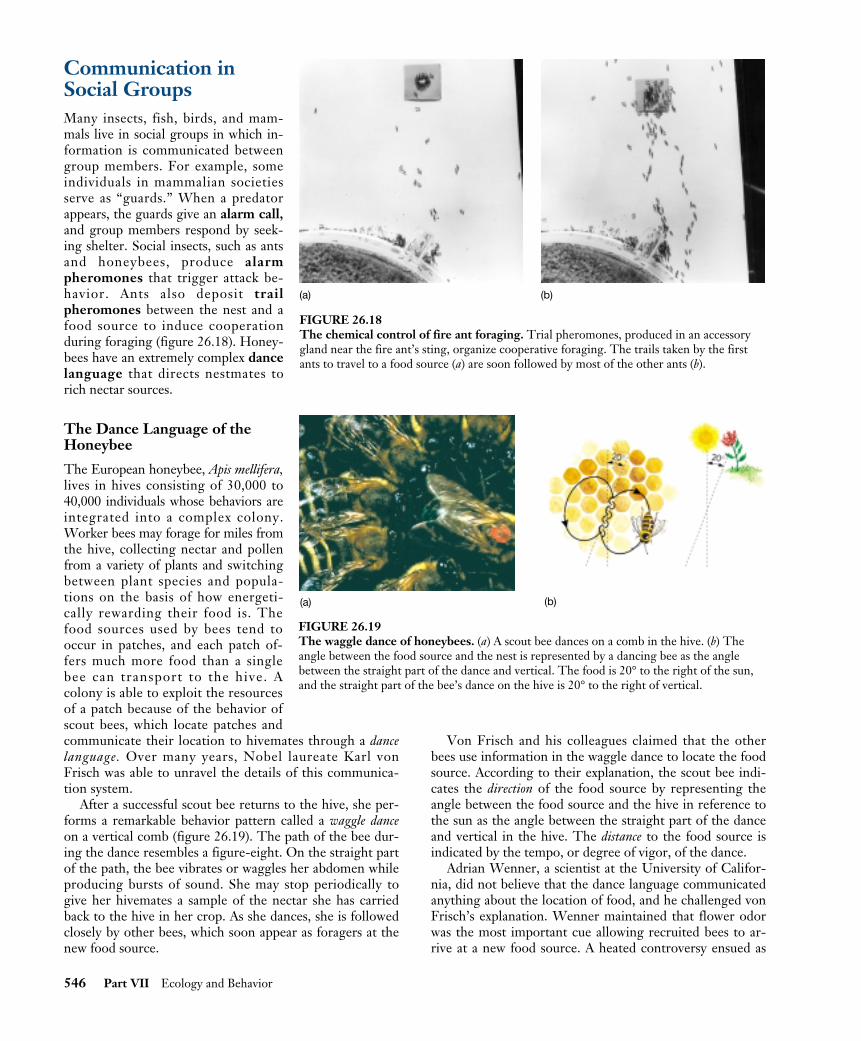
After a successful scout bee returns to the hive, she per-forms a remarkable behavior pattern called a waggle danceon a vertical comb (figure 26.19). The path of the bee dur-ing the dance resembles a figure-eight. On the straight partof the path, the bee vibrates or waggles her abdomen whileproducing bursts of sound. She may stop periodically togive her hivemates a sample of the nectar she has carriedback to the hive in her crop. As she dances, she is followedclosely by other bees, which soon appear as foragers at thenew food source.
Von Frisch and his colleagues claimed that the otherbees use information in the waggle dance to locate the foodsource. According to their explanation, the scout bee indi-cates the direction of the food source by representing theangle between the food source and the hive in reference tothe sun as the angle between the straight part of the danceand vertical in the hive. The distance to the food source isindicated by the tempo, or degree of vigor, of the dance.
Adrian Wenner, a scientist at the University of Califor-nia, did not believe that the dance language communicatedanything about the location of food, and he challenged vonFrisch’s explanation. Wenner maintained that flower odorwas the most important cue allowing recruited bees to ar-rive at a new food source. A heated controversy ensued as
546 Part VII Ecology and Behavior
(a) (b)
FIGURE 26.18The chemical control of fire ant foraging. Trial pheromones, produced in an accessorygland near the fire ant’s sting, organize cooperative foraging. The trails taken by the firstants to travel to a food source (a) are soon followed by most of the other ants (b).
(a) (b)
FIGURE 26.19The waggle dance of honeybees. (a) A scout bee dances on a comb in the hive. (b) Theangle between the food source and the nest is represented by a dancing bee as the anglebetween the straight part of the dance and vertical. The food is 20° to the right of the sun,and the straight part of the bee’s dance on the hive is 20° to the right of vertical.

the two groups of researchers published articles supportingtheir positions.
The “dance language controversy” was resolved (in theminds of most scientists) in the mid-1970s by the creativeresearch of James L. Gould. Gould devised an experimentin which hive members were tricked into misinterpretingthe directions given by the scout bee’s dance. As a result,Gould was able to manipulate where the hive memberswould go if they were using visual signals. If odor were thecue they were using, hive members would have appeared atthe food source, but instead they appeared exactly whereGould predicted. This confirmed von Frisch’s ideas.
Recently, researchers have extended the study of thehoneybee dance language by building robot bees whosedances can be completely controlled. Their dances are pro-grammed by a computer and perfectly reproduce the nat-ural honeybee dance; the robots even stop to give foodsamples! The use of robot bees has allowed scientists to de-termine precisely which cues direct hivemates to foodsources.
Primate Language
Some primates have a “vocabulary” that allows individualsto communicate the identity of specific predators. The vo-calizations of African vervet monkeys, for example, distin-guish eagles, leopards, and snakes (figure 26.20). Chim-panzees and gorillas can learn to recognize a large numberof symbols and use them to communicate abstract con-cepts. The complexity of human language would at first ap-pear to defy biological explanation, but closer examinationsuggests that the differences are in fact superficial—all lan-guages share many basic structural similarities. All of theroughly 3000 languages draw from the same set of 40 con-
sonant sounds (English uses two dozen of them), and anyhuman can learn them. Researchers believe these similari-ties reflect the way our brains handle abstract information,a genetically determined characteristic of all humans.
Language develops at an early age in humans. Humaninfants are capable of recognizing the 40 consonant soundscharacteristic of speech, including those not present in theparticular language they will learn, while they ignore othersounds. In contrast, individuals who have not heard certainconsonant sounds as infants can only rarely distinguish orproduce them as adults. That is why English speakers havedifficulty mastering the throaty French “r,” French speak-ers typically replace the English “th” with “z,” and nativeJapanese often substitute “l” for the unfamiliar English “r.”Children go through a “babbling” phase, in which theylearn by trial and error how to make the sounds of lan-guage. Even deaf children go through a babbling phaseusing sign language. Next, children quickly and easily learna vocabulary of thousands of words. Like babbling, thisphase of rapid learning seems to be genetically pro-grammed. It is followed by a stage in which children formsimple sentences which, though they may be grammaticallyincorrect, can convey information. Learning the rules ofgrammar constitutes the final step in language acquisition.
While language is the primary channel of human com-munication, odor and other nonverbal signals (such as“body language”) may also convey information. However,it is difficult to determine the relative importance of theseother communication channels in humans.
The study of animal communication involves analysis ofthe specificity of signals, their information content, andthe methods used to produce and receive them.
Chapter 26 Animal Behavior 547
0
1
0.5 secondsEagle
2
3
4
5
6
7
8
0.5 secondsLeopard
0
1Fre
quen
cy (
kilo
cycl
espe
r se
cond
)
Fre
quen
cy (
kilo
cycl
espe
r se
cond
)
2
3
4
5
6
7
8
(a)
(b)
FIGURE 26.20Primate semantics. (a) Predators, like this leopard, attack and feed on vervetmonkeys. (b) The monkeys give different alarm calls when eagles, leopards, andsnakes are sighted by troupe members. Each distinctive call elicits a different andadaptive escape behavior.

Orientation and MigrationAnimals may travel to and from a nest to feed or move reg-ularly from one place to another. To do so, they must ori-ent themselves by tracking stimuli in the environment.
Movement toward or away from some stimulus is calledtaxis. The attraction of flying insects to outdoor lights is anexample of positive phototaxis. Insects that avoid light, suchas the common cockroach, exhibit negative phototaxis. Otherstimuli may be used as orienting cues. For example, troutorient themselves in a stream so as to face against the cur-rent. However, not all responses involve a specific orienta-tion. Some animals just become more active when stimulusintensity increases, a responses called kineses.
Long-range, two-way movements are known as migra-tions. In many animals, migrations occur circannually.Ducks and geese migrate along flyways from Canada acrossthe United States each fall and return each spring.Monarch butterflies migrate each fall from central andeastern North America to several small, geographically iso-lated areas of coniferous forest in the mountains of centralMexico (figure 26.21). Each August, the butterflies begin aflight southward to their overwintering sites. At the end ofwinter, the monarchs begin the return flight to their sum-mer breeding ranges. What is amazing about the migrationof the monarch, however, is that two to five generationsmay be produced as the butterflies fly north. The butter-flies that migrate in the autumn to the precisely locatedoverwintering grounds in Mexico have never been therebefore.
When colonies of bobolinks became established in thewestern United States, far from their normal range in theMidwest and East, they did not migrate directly to theirwinter range in South America. Instead, they migrated eastto their ancestral range and then south along the originalflyway (figure 26.22). Rather than changing the originalmigration pattern, they simply added a new pattern.
How Migrating Animals Navigate
Biologists have studied migration with great interest, andwe now have a good understanding of how these feats ofnavigation are achieved. It is important to understand thedistinction between orientation (the ability to follow abearing) and navigation (the ability to set or adjust a bear-ing, and then follow it). The former is analogous to using acompass, while the latter is like using a compass in con-junction with a map. Experiments on starlings indicate thatinexperienced birds migrate by orientation, while olderbirds that have migrated previously use true navigation(figure 26.23).
Birds and other animals navigate by looking at the sunand the stars. The indigo bunting, which flies during theday and uses the sun as a guide, compensates for themovement of the sun in the sky as the day progresses byreference to the north star, which does not move in thesky. Buntings also use the positions of the constellationsand the position of the pole star in the night sky, cuesthey learn as young birds. Starlings and certain otherbirds compensate for the sun’s apparent movement in the
548 Part VII Ecology and Behavior
26.4 Migratory behavior presents many puzzles.
SanFrancisco
NewYork
LosAngeles
MexicoCity
Summerbreedingranges
Overwinteringaggregationareas
(a) (b) (c)
FIGURE 26.21Migration of monarch butterflies. (a) Monarchs from western North America overwinter in areas of mild climate along the PacificCoast. Those from the eastern United States and southeastern Canada migrate to Mexico, a journey of over 3000 kilometers that takesfrom two to five generations to complete. (b) Monarch butterflies arriving at the remote fir forests of the overwintering grounds and (c)forming aggregations on the tree trunks.

sky by using an internal clock. If such birds are shown anexperimental sun in a fixed position while in captivity,they will change their orientation to it at a constant rateof about 15° per hour.
Many migrating birds also have the ability to detect theearth’s magnetic field and to orient themselves with respectto it. In a closed indoor cage, they will attempt to move inthe correct geographical direction, even though there areno visible external cues. However, the placement of a pow-erful magnet near the cage can alter the direction in whichthe birds attempt to move. Magnetite, a magnetized ironore, has been found in the heads of some birds, but the sen-sory receptors birds employ to detect magnetic fields havenot been identified.
It appears that the first migration of a bird is innatelyguided by both celestial cues (the birds fly mainly atnight) and the earth’s magnetic field. These cues give thesame information about the general direction of the mi-gration, but the information about direction provided bythe stars seems to dominate over the magnetic informa-tion when the two cues are experimentally manipulated togive conflicting directions. Recent studies, however, indi-cate that celestial cues tell northern hemisphere birds tomove south when they begin their migration, while mag-netic cues give them the direction for the specific migra-tory path (perhaps a southeast turn the bird must makemidroute). In short, these new data suggest that celestialand magnetic cues interact during development to fine-tune the bird’s navigation.
We know relatively little about how other migratinganimals navigate. For instance, green sea turtles migratefrom Brazil halfway across the Atlantic Ocean to Ascen-sion Island, where the females lay their eggs. How dothey find this tiny island in the middle of the ocean,which they haven’t seen for perhaps 30 years? How dothe young that hatch on the island know how to findtheir way to Brazil? Recent studies suggest that wave ac-tion is an important cue.
Many animals migrate in predictable ways, navigatingby looking at the sun and stars, and in some cases bydetecting magnetic fields.
Chapter 26 Animal Behavior 549
FIGURE 26.22Birds on the move. (a) Thesummer range of bobolinksrecently extended to the far Westfrom their more established rangein the Midwest. When theymigrate to South America in thewinter, bobolinks that nested inthe West do not fly directly to thewinter range; instead, they fly tothe Midwest first and then use theancestral flyway. (b) The goldenplover has an even longermigration route that is circular.These birds fly from Arcticbreeding grounds to winteringareas in southeastern SouthAmerica, a distance of some 13,000kilometers.
Winteringrange
Breedingrange
Holland
Switzerland
Spain
Bobolink Golden plover
Summernestingrange
Winterrange
Summernestingrange
Winterrange
(a) (b)
FIGURE 26.23Migratory behavior of starlings. The navigational abilities ofinexperienced birds differ from those of adults who have made themigratory journey before. Starlings were captured in Holland,halfway along their full migratory route from Baltic breedinggrounds to wintering grounds in the British Isles; these birds weretransported to Switzerland and released. Experienced older birdscompensated for the displacement and flew toward the normalwintering grounds (blue arrow). Inexperienced young birds keptflying in the same direction, on a course that took them towardSpain (red arrow). These observations imply that inexperiencedbirds fly by orientation, while experienced birds learn truenavigation.

Animal CognitionIt is likely each of us could tell an anecdotal story about thebehavior of a pet cat or dog that would seem to suggest thatthe animal had a degree of reasoning ability or was capableof thinking. For many decades, however, students of animalbehavior flatly rejected the notion that nonhuman animalscan think. In fact, behaviorist Lloyd Morgan stated thatone should never assume a behavior represents consciousthought if there is any other explanation that precludes theassumption of consciousness. The prevailing approach wasto treat animals as though they responded to the environ-ment through reflexlike behaviors.
In recent years, serious attention has been given to thetopic of animal awareness. The central question iswhether animals show cognitive behavior—that is, dothey process information and respond in a manner thatsuggests thinking (figure 26.24)? What kinds of behaviorwould demonstrate cognition? Some birds in urban areasremove the foil caps from nonhomogenized milk bottlesto get at the cream beneath, and this behavior is known to
have spread within a population to other birds. Japanesemacaques learned to wash potatoes and float grain to sep-arate it from sand. A chimpanzee pulls the leaves off of atree branch and uses the stick to probe the entrance to atermite nest and gather termites. As we saw earlier, vervetmonkeys have a vocabulary that identifies specific preda-tors.
Only a few experiments have tested the thinking abilityof nonhuman animals. Some of these studies suggest thatanimals may give false information (that is, they “lie”).Currently, researchers are trying to determine if some pri-mates deceive others to manipulate the behavior of theother members of their troop. There are many anecdotalaccounts that appear to support the idea that deception oc-curs in some nonhuman primate species such as baboonsand chimpanzees, but it has been difficult to devise field-based experiments to test this idea. Much of this type of re-search on animal cognition is in its infancy, but it is sure togrow and to raise controversy. In any case, there is nothingto be gained by a dogmatic denial of the possibility of animalconsciousness.
550 Part VII Ecology and Behavior
26.5 To what degree animals “think” is a subject of lively dispute.
(a) (b)
FIGURE 26.24Animal thinking? (a) This chimpanzee is stripping the leaves from a twig, which it will then use to probe a termite nest. This behaviorstrongly suggests that the chimpanzee is consciously planning ahead, with full knowledge of what it intends to do. (b) This sea otter isusing a rock as an “anvil,” against which it bashes a clam to break it open. A sea otter will often keep a favorite rock for a long time, asthough it has a clear idea of what it is going to use the rock for. Behaviors such as these suggest that animals have cognitive abilities.

In any case, some examples, particularly those involvingproblem-solving, are hard to explain in any way other thanas a result of some sort of mental process. For example, in aseries of classic experiments conducted in the 1920s, achimpanzee was left in a room with a banana hanging fromthe ceiling out of reach. Also in the room were severalboxes, each lying on the floor. After unsuccessful attemptsto jump up and grab the bananas, the chimpanzee suddenlylooks at the boxes and immediately proceeds to move themunderneath the banana, stack one on top of another, andclimb up to claim its prize (figure 26.25).
Perhaps it is not so surprising to find obvious intelli-gence in animals as closely related to us as chimpanzees.But recent studies have found that other animals alsoshow evidence of cognition. Ravens have always been con-sidered among the most intelligent of birds. Bernd Hein-rich of the University of Vermont recently conducted anexperiment using a group of hand-reared crows that livedin an outdoor aviary. Heinrich placed a piece of meat onthe end of a string and hung it from a branch in theaviary. The birds liked to eat meat, but had never seenstring before and were unable to get at the meat. Afterseveral hours, during which time the birds periodicallylooked at the meat but did nothing else, one bird flew tothe branch, reached down, grabbed the string, pulled itup, and placed it under his foot. He then reached downand grabbed another piece of the string, repeating this ac-tion over and over, each time bringing the meat closer(figure 26.26). Eventually, the meat was within reach andwas grasped. The raven, presented with a completelynovel problem, had devised a solution. Eventually, threeof the other five ravens also figured out how to get to themeat. Heinrich has conducted other similarly creative ex-periments that can leave little doubt that ravens have ad-vanced cognitive abilities.
Research on the cognitive behavior of animals is in itsinfancy, but some examples are compelling.
Chapter 26 Animal Behavior 551
FIGURE 26.26Problem solving by a raven. Confronted with a problem it hadnever previously confronted, the raven figures out how to get themeat at the end of the string by repeatedly pulling up a bit ofstring and stepping on it.
FIGURE 26.25Problem solving by achimpanzee. Unable toget the bananas byjumping, the chimpanzeedevises a solution.

552 Part VII Ecology and Behavior
Chapter 26 Summary Questions Media Resources
26.1 Ethology focuses on the natural history of behavior.
• Behavior is an adaptive response to stimuli in theenvironment. An animal’s sensory systems detect andprocess information about these stimuli.
1. How does a hybrid lovebird’smethod of carrying nestmaterials compare with that ofits parents? What does thiscomparison suggest aboutwhether the behavior isinstinctive or learned?
• Behavior is both instinctive (influenced by genes) andlearned through experience. Genes are thought tolimit the extent to which behavior can be modifiedand the types of associations that can be made.
• The simplest forms of learning involve habituationand sensitization. More complex associative learning,such as classical and operant conditioning, may bedue to the strengthening or weakening of existingsynapses as well as the formation of entirely newsynapses.
• An animal’s internal state influences when and how abehavior will occur. Hormones can change an ani-mal’s behavior and perception of stimuli in a way thatfacilitates reproduction.
2. How does associative learningdiffer from nonassociativelearning? How does classicalconditioning differ from operantconditioning?3. What is filial imprinting?What is sexual imprinting? Whydo some young animals imprinton objects like a moving box?4. How does Marler’s work onsong development in white-crowned sparrows indicate thatbehavior is shaped by learning?How does it indicate thatbehavior is shaped by instinct?
26.2 Comparative psychology focuses on how learning influences behavior.
• Animals communicate by producing visual, acoustic,and chemical signals. These signals are involved inmating, finding food, defense against predators, andother social situations.
5. How do communicationsignals participate inreproductive isolation? Give oneexample of a signal that isspecies-specific. Why are somesignals individually specific?
26.3 Communication is a key element of many animal behaviors.
• Animals use cues such as the position of the sun andstars to orient during daily activities and to navigateduring long-range migrations.
6. What is the definition oftaxis? What are kineses? Whatcues do migrating birds use toorient and navigate during theirmigrations?
26.4 Migratory behavior presents many puzzles.
• Many anecdotal accounts point to animal cognition,but research is in its infancy.
7. What evidence would youaccept that an animal is“thinking”?
26.5 To what degree animals “think” is a subject of lively dispute.
www.mhhe.comraven6e www.biocourse.com
• On Science Article:Polyandry in Hawks