anolis heterodermus
TRANSCRIPT

Seediscussions,stats,andauthorprofilesforthispublicationat:http://www.researchgate.net/publication/265249772
UnknownEvolutionaryLineagesandPopulationDifferentiationinAnolisheterodermus(Squamata:Dactyloidae)fromtheEasternandCentralCordillerasofColombiaRevealedbyDNASeq...
ARTICLEinSOUTHAMERICANJOURNALOFHERPETOLOGY·SEPTEMBER2014
DOI:10.2994/SAJH-D-13-00013.1
CITATION
1
READS
99
2AUTHORS:
MarioVargas-Ramírez
MuseumofZoologySenckenbergDresden
30PUBLICATIONS184CITATIONS
SEEPROFILE
RafaelA.Moreno-Arias
NationalUniversityofColombia
18PUBLICATIONS12CITATIONS
SEEPROFILE
Availablefrom:MarioVargas-Ramírez
Retrievedon:17October2015

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions,research libraries, and research funders in the common goal of maximizing access to critical research.
Unknown Evolutionary Lineages and Population Differentiation in Anolisheterodermus (Squamata: Dactyloidae) from the Eastern and CentralCordilleras of Colombia Revealed by DNA Sequence DataAuthor(s): Mario Vargas-Ramírez and Rafael Moreno-AriasSource: South American Journal of Herpetology, 9(2):131-141. 2014.Published By: Brazilian Society of HerpetologyDOI: http://dx.doi.org/10.2994/SAJH-D-13-00013.1URL: http://www.bioone.org/doi/full/10.2994/SAJH-D-13-00013.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological,and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and bookspublished by nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercialinquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.

Unknown Evolutionary Lineages and Population
Differentiation in Anolis heterodermus (Squamata:
Dactyloidae) from the Eastern and Central Cordilleras of
Colombia Revealed by DNA Sequence Data
Mario Vargas-Ramírez1,2,3, Rafael Moreno-Arias2
1 Museum of Zoology, Senckenberg Dresden, A.B. Meyer Building, D-01109 Dresden, Germany.2 Grupo de Biodiversidad y Sistemática Molecular, Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá, Colombia.3 Corresponding author. E-mail: [email protected]
Abstract. Anolis heterodermus is a poorly known, high elevation anole lizard from northern Andes, currently under threat due to habitat
destruction. Although it has been suggested that this taxon corresponds to a species complex, different evolutionary lineages have not been
identified. We examined phylogenetic relationships between individuals of A. heterodermus from the Eastern and Central Cordilleras of
Colombia and other species of the A. heterodermus series using mitochondrial DNA (partial ND2 gene with adjacent tRNA genes and partial
COI gene) and nuclear DNA (partial RAG1 gene) sequences and assessed divergence times between these lineages to elucidate their historical
biogeography. We performed genetic analyses for two populations from the middle portion of the Eastern Cordillera of Colombia based on
the COI gene. We identified three independently evolving evolutionary lineages within A. heterodermus based on two lines of evidence: (i) all
phylogenetic analyses showed A. heterodermus to comprise three strongly supported subgroups, being polyphyletic with respect to other
recognized species of the A. heterodermus series and (ii) the uncorrected p-distances of the ND2 gene revealed that the divergence between
A. heterodermus lineages exceed the divergence between recognized species of the A. heterodermus series. These lineages should be considered
as different evolutionary significant units (ESUs) and candidate species. We found that the diversification of the studied lineages dates to
the Middle Miocene to Pleistocene, falling within a period of major orogenic and climatic events in northern South America. The popula-
tion genetic analyses revealed two management units (MUs) in one of the newly reported lineages from the middle portion of the Eastern
Cordillera. Due to the accelerated destruction of their natural habitat, steps should be taken to ensure the conservation of the identified
ESUs and MUs.
Keywords. Anoles; Dactyloa; mitochondrial DNA; Northern Andes; nuclear DNA; Phenacosaurus.
Resumen. Anolis heterodermus es un lagarto poco estudiado de los Andes del norte, actualmente amenazado por destrucción de su hábitat.
Aunque se ha sugerido que este taxón corresponde a un complejo de especies, no se han identificado linajes evolutivos diferentes. En este
estudio examinamos las relaciones filogenéticas entre individuos de A. heterodermus de las Cordilleras Oriental y Central de Colombia y otras
especies de la serie A. heterodermus, usando ADN mitocondrial (ND2 parcial, genes tARN adyacentes y COI parcial) y ADN nuclear (RAG1
parcial). Adicionalmente, evaluamos los tiempos de divergencia entre estos taxones para dilucidar aspectos biogeográficos históricos. Por
otra parte, llevamos a cabo análisis genéticos de dos poblaciones de la parte media de la Cordillera Oriental de Colombia usando COI. Repor-
tamos tres linajes evolutivos independientes dentro de A. heterodermus basados en dos líneas de evidencia: (i) todos los análisis filogenéticos
revelaron tres subgrupos fuertemente soportados dentro de A. heterodermus, siendo polifilético con respecto a otras especies reconocidas
de la serie A. heterodermus y (ii) las distancias p no corregidas del gen ND2 revelaron que la divergencia entre linajes de A. heterodermus,
exceden la diferenciación entre varias especies reconocidas de la serie A. heterodermus. Estos linajes deben ser considerados como unidades
evolutivas significativas (UES) y especies candidatas. Encontramos que los tiempos de diversificación de las especies estudiadas datan del
Mioceno medio al Pleistoceno, un periodo de eventos orogénicos y climáticos importantes en el norte de Sudamérica. El análisis de genética
de poblaciones reveló dos unidades de manejo (UM) en uno de los linajes identificados de la Cordillera Oriental de Colombia. Debido a la ace-
lerada destrucción de su hábitat natural, medidas de conservación independientes para las UES y UM identificadas deben ser garantizadas.
2001) and considered one of the most threatened regions
with the highest conservation needs in the world (Stadt-
müller, 1987; Olson and Dinerstein, 1997). Therefore,
studies aimed at increasing knowledge about the tropical
Andean biota, to support their conservation and manage-
ment, are crucial.
The high elevation anole lizards of the Andes pre-
viously placed in the genus Phenacosaurus (Lazell, 1969;
Myers and Donnelly, 1996; Williams et al., 1996) are
currently placed in either the heterodermus series of the
unranked “Dactyloa clade” (Castañeda and de Queiroz,
INTRODUCTION
The tropical Andes are regarded as one of the world’s
greatest biodiversity hotspots, harboring high levels of
vertebrate and plant endemism (Olson and Dinerstein,
1997; Myers et al., 2000; Primack et al., 2001; Orme et al.,
2005). Their complex geological and environmental his-
tory is thought to have played a key role in promoting
speciation (Lynch, 1999; Willmott et al., 2001; Guarnizo
et al., 2009). Nevertheless, the tropical Andes are seri-
ously endangered by habitat destruction (Primack et al.,
South American Journal of Herpetology, 9(2), 2014, 131–141
© 2014 Brazilian Society of Herpetology
Submitted: 01 May 2013
Accepted: 25 July 2014
Handling Editor: Guarino Rinaldi Colli
doi: 10.2994/SAJH-D-13-00013.1

2011, 2013) within the genus Anolis (Poe, 1998, 2004;
Jackman et al., 1999; Nicholson et al., 2005; Castañeda
and de Queiroz, 2011, 2013) or the heterodermus species
group of the genus Dactyloa (Nicholson et al., 2012). They
occur in Colombia, Ecuador, Peru and western Venezuela
at elevations of 1,300–3,750 m, reaching the highest el-
evations of any anole (Castañeda and de Queiroz, 2011).
The A. heterodermus series includes six species from Co-
lombia, Ecuador and Venezuela: A. euskalerriari, A. hetero-
dermus, A. inderenae, A. nicefori, A. vanzolinii and A. tetarii
(Castañeda and de Queiroz, 2011, 2013). These lizards
are characterized by lamellar subdigital scales, well-devel-
oped casquing, and the presence of heterogeneous dorsal
scales in all species but A. euskalerriari (Castañeda and de
Queiroz, 2013).
Anolis heterodermus is a large species (maxi-
mum snout–vent length [SVL]: females = 86 mm,
males = 85 mm; Lazell, 1969; M. Vargas-Ramírez, un-
published data) inhabiting Andean scrubland and for-
est (Dunn, 1944) and conforming to a twig ecomorph
(Miyata, 1983; Torres-Carvajal et al., 2010, Nicholson
et al., 2012). It occurs in the northern Ecuadorian Andes
and all three cordilleras of the Colombian Andes at eleva-
tions above 1,600 m (Fig. 1; Torres-Carvajal et al., 2010;
Castañeda and de Queiroz, 2011). Some information on
its ecology and natural history has been gathered (Dunn,
1944; Osorno-Mesa and Osorno-Mesa, 1946; Lazell,
1969; Miyata, 1983; Torres-Carvajal et al., 2010, More-
no-Arias and Urbina-Cardona, 2013), and it was included
in three recent molecular phylogenetic studies of anoles
(Castañeda and de Queiroz, 2011, 2013; Nicholson et al.,
2012).
Previous efforts to recognize different species with-
in Anolis heterodermus based on morphology were un-
successful. Lazell (1969) showed that the morphological
characters used to describe A. richteri (Dunn, 1944) and
A. paramoensis (Hellmich, 1949) fall within the variation
in A. heterodermus, which he considered a senior synonym
of the former two. Later, due to the large morphologi-
cal variability observed within this taxon, Williams et al.
(1996) suggested that A. heterodermus is an unresolved
complex of sibling species. Likewise, Castañeda and de
Queiroz (2011) suggested that the species might repre-
sent a species complex, based on the observation that for
the RAG1 dataset A. heterodermus was not monophyletic
(A. inderenae was nested within it) and the fact that geo-
graphic distance between multiple samples of A. hetero-
dermus were in conflict with their evolutionary relation-
ships revealed by the ND2 and COI genes. Despite these
suggestions, the identification of independent evolution-
ary lineages within the species is still lacking.
Herein, we used mitochondrial and nuclear DNA se-
quence data to (i) assess the phylogenetic relationships
between populations of Anolis heterodermus from the
Eastern and Central cordilleras of Colombia and of allied
taxa of the A. heterodermus series (excluding A. tetarii)
and (ii) study the population genetics of A. heterodermus
from the middle portion of the Eastern Cordillera of Co-
lombia. Furthermore, past expansion of such populations
and divergence times between taxa of the A. heterodermus
series were assessed to elucidate historical biogeographic
aspects of these Andean anoles.
MATERIALS AND METHODS
Sampling and laboratory procedures
A total of 63 samples of Anolis heterodermus from
eight localities in the middle portion of the Eastern Cor-
dillera of Colombia (Fig. 1, inset) were collected in several
field trips from 2010–2012. Samples were obtained by
toe-clipping and stored in 96% ethanol Individuals were
released immediately after sampling. To assess phylo-
genetic relationships, fragments of mitochondrial DNA
Figure 1. Geographic distribution of the six species of the A. hetero-
dermus series. Red stars within the inset indicate sampled localities for
Anolis heterodermus in the middle portion of the Eastern Cordillera. Red
stars outside inset indicate localities for Anolis heterodermus individuals
from Central Cordillera included in this study. Letters A, B, and C show
the location of subgroups. Other species records from Dunn (1944), La-
zell (1969), Rueda and Hernández-Camacho (1988), Mueses-Cisneros
(2006), Torres-Carvajal et al. (2010), Castañeda and De Queiroz (2011,
2013), HerpNet database (accessed 21 April 2013), Instituto Alexander
von Humboldt data base (information provided by the institution), Uni-
versidad de Antioquia herpetological database (accessed 21 April 2013),
and Universidad Industrial de Santander herpetological database (ac-
cessed 21 April 2013).
Unknown Evolutionary Lineages and Population Differentiation in Anolis heterodermus (Squamata: Dactyloidae)
from the Eastern and Central Cordilleras of Colombia Revealed by DNA Sequence Data
Mario Vargas-Ramírez, Rafael Moreno-Arias
132
South American Journal of Herpetology, 9(2), 2014, 131–141

(mtDNA) partial NADH dehydrogenase subunit II (ND2),
the adjacent transfer-RNA, and cytochrome oxidase sub-
unit I (COI), and the nuclear (nDNA) recombination ac-
tivating gene 1 (RAG1) were sequenced for representa-
tive samples of each locality. For the population genetics
analyses, COI sequences were obtained for all 63 sampled
individuals (Appendix S1).
Total genomic DNA was extracted using a Qiagen
DNA tissue extraction kit (Qiagen, Hilden, Germany)
following the manufacturer’s instructions. All gene frag-
ments were amplified using newly designed primer pairs.
Primer sequences and PCR conditions are shown in Ta-
ble 1. PCRs were carried out in a total volume of 50 μl
containing 1 unit Taq polymerase (Bioron, Ludwigshafen,
Germany), 1 × buffer (as recommended by the supplier),
0.5 μM of each primer, and 0.2 mM of each dNTP (Fer-
mentas, St. Leon-Rot, Germany). PCR products were puri-
fied using the ExoSAP-IT enzymatic cleanup (USB Europe
GmbH, Staufen, Germany; modified protocol: 30 min at
37°C, 15 min at 80°C) and sequenced on an ABI 3130xl
Genetic Analyzer (Applied Biosystems, Foster City, CA,
USA) using the BigDye Terminator v3.1 Cycle Sequencing
Kit (Applied Biosystems). Remaining DNA extractions
are stored at -80°C in the tissue sample collection of the
Museum of Zoology, Senckenberg Dresden, Germany (for
voucher numbers see Appendix S1).
Phylogenetic analyses and divergence dating
Phylogenetic relationships between populations
of Anolis heterodermus from the middle portion of the
Eastern and Central Cordilleras of Colombia, and the
species belonging of the A. heterodermus series (A. euskal-
erriari, A. inderenae, A. nicefori and A. vanzolinii) were as-
sessed using a fragment of 1,036 bp of ND2 and 397 bp
corresponding to the adjacent five transfer-RNA genes
(tRNA-Trp, tRNA-Ala, tRNA-Asn, tRNA-Cys, tRNA-Tyr).
Additional analyses were conducted using the same taxa
minus A. nicefori (for which only a single sequence of
ND2 + tRNA was available) and the following datasets:
(i) a 534 bp fragment of COI, (ii) a 1,967 bp mtDNA data-
set comprosed of partial ND2, adjacent tRNA genes, and
partial COI, (iii) a 1,761 bp nDNA fragment of RAG1 and,
(iv) a combined matrix of 3,728 bp, containing the mtD-
NA and nuDNA. For tree rooting, A. neblininus, A. calimae,
and Polychrus marmoratus were used as outgroups. For
phylogenetic inference, the newly obtained sequences of
A. heterodermus from the middle portion of the Eastern
Cordillera were combined with GenBank data from indi-
viduals of the same species from the Central Cordillera.
GenBank sequences included one individual from an un-
known locality, four species of the A. heterodermus series,
and the outgroups (see Appendix S1).
The ND2 + tRNAs, COI, mitochondrial, RAG1, and
combined datasets were analyzed using the following par-
tition schemes: (i) unpartitioned, (ii) partitioned by gene
(i.e., each gene treated as a distinct partition, all tRNA
genes grouped into a single partition), and (iii) maximum
partitioning (i.e., each codon of each protein-coding gene
and all tRNA genes treated as distinct partitions). The
software PARTITIONFINDER (Lanfear et al., 2012) was
used to find the best partitioning scheme and substitu-
tion models for phylogenetic analyses, resulting in the
selection of the maximum partitioning scheme for all da-
tasets. Based on this scheme and individual best-fit mod-
els for nucleotide substitution (Table S1), Bayesian analy-
ses (BA) were performed in MrBayes v.3.1 (Ronquist and
Huelsenbeck, 2003). The substitution models were incor-
porated into a single tree search (mixed model partition
approach; Nylander et al., 2004) and two parallel runs
were carried out using four Markov chains, each starting
from a random tree. The Markov chains were run for 10
million generations, sampling every 100 generations. The
burn-in was set to sample only the plateau of the most
likely trees, which were used for generating a 50% major-
ity rule consensus. The posterior probabilities in this tree
are a measure of clade frequency and, thus, credibility.
The software TRACER 1.5.4 (Rambaut and Drummond,
2007) was used to assess an acceptable level of the MCMC
Table 1. Primers developed and used in this study and thermocycling conditions. ID = initial denaturing, C = No. cycles, D = denaturing, A = annealing,
PE = primer extension, FE = final elongation.
Frag-
mentPrimers Forward/Reverse primers (5’–3’)
Thermocycling conditions
ID C D A PE FE
ND2 PheIfor/PheIIrev CTAGCCTTGCAACCGGAA/ 5 min, 94°C 35–38 45 s, 94°C 45 s, 53–55°C 60 s, 72°C 10 min, 72°C
AGGTAAAATGGCCGAACGT
ND2 HetN1F/HetN1R TTGCAACCGGAACCATTATT/ 5 min, 94°C 35–38 45 s, 94°C 45 s, 53–55°C 60 s, 72°C 10 min, 72°C
GCTATTAAAATTGGTAGGGG
COI Hetco1f/Hetco1r CATTGGCACCCTTTACTTAA/ 5 min, 94°C 35 45 s, 94°C 45 s, 52°C 60 s, 72°C 10 min, 72°C
AGGTGTTTAAGTTACGGTCT
Rag1 HetRaf/HetRar TTGAGAAGCCACTTCCTGAT/ 5 min, 95°C 35 45 s, 95°C 60 s, 63–66°C 60 s, 72°C 10 min, 72°C
TAGGAAAGCAAGGATAGCGA
Rag1 HetRbf/HetRbr AGAACCTGCATCCTSAAATG/ 5 min, 95°C 35 45 s, 95°C 60 s, 63–66°C 60 s, 72°C 10 min, 72°C
TTGCATTGCCATGAGTGACA
Unknown Evolutionary Lineages and Population Differentiation in Anolis heterodermus (Squamata: Dactyloidae)
from the Eastern and Central Cordilleras of Colombia Revealed by DNA Sequence Data
Mario Vargas-Ramírez, Rafael Moreno-Arias
133
South American Journal of Herpetology, 9(2), 2014, 131–141

chains mixing and to estimate effective sample sizes for
all parameters. Additionally, also using the maximum par-
titioning scheme for each dataset, maximum likelihood
(ML) analyses were run in RAxML 7.2.6 (Stamatakis,
2006), implemented in the graphical user interface rax-
mlgui v.0.93 (Silvestro and Michalak, 2012). The default
GTR + G model was used across all partitions. To explore
the robustness of the branching patterns, five indepen-
dent ML searches were run using the fast bootstrap algo-
rithm. Subsequently, 1,000 thorough bootstrap replicates
were calculated and plotted on the tree with the highest
likelihood value. For assessing intra- and interspecific
variability within Anolis heterodermus and between spe-
cies of the A. heterodermus series, respectively, uncorrect-
ed p-distances based on the partial ND2 gene calculated
with MEGA v.4.0.2 (Tamura et al., 2011) were compared.
Divergence time estimation of the Anolis hetero-
dermus series lineages was performed using the relaxed
Bayesian molecular clock approach implemented in the
program BEAST 1.7.5 (Drummond et al., 2012), using
the subprogram BEAUti v1.7.5 to set the analysis param-
eters. Due to the lack of appropriate Anolis fossil records
to calibrate the phylogeny, the average rate of evolution
of the fragment of the ND2 gene of 0.65% mutations per
lineage per million years (Macey et al., 1998) was used.
This evolutionary rate has already been implemented to
estimate divergence times in several Anolis species (e.g.,
Creer et al., 2001; Glor et al., 2003; Jackman et al., 2002;
Jezkova et al., 2009; Rodríguez-Robles et al., 2008; Gart-
ner et al., 2013). The following parameters were used to
run the analysis: lognormal clock model, speciation Yule
process tree prior, random starting tree and the model of
sequence evolution HKY; selected as substitution model
using the Bayesian information criterion in PARTITION-
FINDER. Two BEAST analyses were run for 100 million
generations each, sampling every 10,000 generations.
Posterior distributions were examined in TRACER, con-
sidering a burn-in of the initial 10% of samples. The sam-
ples of probable trees were summarized using TreeAnno-
tator v.1.7.5 (Drummond et al., 2012), and the resulting
trees visualized using FigTree v.1.3.1 (Rambaut, 2009).
Population genetics analyses
The 63 COI sequences (533 bp) were collapsed into
haplotypes using TCS v.1.21 (Clement et al., 2000). Based
on the phylogenetic results, population structure was
evaluated for each evolutionary lineage of Anolis hetero-
dermus from the middle portion of the Eastern Cordil-
lera using BAPS v.5.3 (Corander and Marttinen, 2006;
Corander et al., 2008). For the genetic mixture analyses
performed at the level of individuals, the maximum num-
ber of populations (Kmax), was set from 1–4, with 10
replicate runs each. The analyses were performed using
the non-spatial model. The admixture analyses based on
mixture clustering were performed with 500 iterations,
30 reference individuals and 20 iterations per reference
individual. To detect genetic differentiation within the
revealed populations, the nearest-neighbour statistic
Snn (Hudson, 2000) was calculated with DNASP v.5 (Li-
brado and Rozas, 2009). The same program was used to
calculate haplotype diversity (Hd), nucleotide diversity
(π), and segregation sites (S) for each population. TCS
was also used to assess genealogical relationships within
populations, by calculating parsimony networks for the
observed COI haplotypes. To test the hypothesis of a re-
cent demographic expansion for the same populations, a
mismatch distribution test was run in ARLEQUIN v.3.11
(Excoffier et al., 2005) and the expansion parameters , 1
and 0 were calculated. ARLEQUIN estimates parameters
of a sudden demographic expansion using a generalized
least-square approach (Schneider and Excoffier, 1999).
Additionally, two tests of selective neutrality (Tajima’s D
and Fu’s FS) were also calculated in ARLEQUIN to test for
signatures of recent demographic expansion.
RESULTS
Phylogenetic analyses and divergence dating
Both tree-building methods revealed for the individ-
ual mitochondrial and nuclear datasets, as well as for the
concatenated mitochondrial and combined matrix datas-
ets, a congruent phylogenetic pattern consisting of three
subgroups (Fig. 2, Figs. S1–S2): subgroup C was shown as
a sister to (A + B). Similarly, in all but the COI tree, Ano-
lis euskalerriari was placed as sister of all three subgroups
with maximum support. However, trees differed in the
placement of some taxa within subgroups and branch sup-
port values. Since only one sequence of the ND2 + tRNA
fragment was available for A. nicefori, detailed descrip-
tion of this dataset’s resulting phylogeny, which is the
most taxonomically complete one, is presented. Trees and
corresponding descriptions for the COI, RAG1, and com-
bined mtDNA datasets are presented in Appendix S2 and
Figures S1–S2. The resulting phylogenetic trees for the
ND2 + tRNA mtDNA and combined datasets are depicted
in Figure 2.
For the ND2 + tRNA dataset, both phylogenetic
analyses revealed the three, well-supported subgroups.
Within the maximally supported subgroup A, Anolis
heterodermus from Majuy (A. heterodermus Maj) was re-
vealed with strong support (Bayesian posterior probabil-
ity of 0.98, ML bootstrap support of 95%) to be sister of
a moderately supported clade formed by individuals of
A. heterodermus from Conejera sites 1 and 2 (Con1 and
Con2; A. heterodermus Con). Both phylogenetic analyses
recovered the sequence of A. heterodermus AY296144 of
Unknown Evolutionary Lineages and Population Differentiation in Anolis heterodermus (Squamata: Dactyloidae)
from the Eastern and Central Cordilleras of Colombia Revealed by DNA Sequence Data
Mario Vargas-Ramírez, Rafael Moreno-Arias
134
South American Journal of Herpetology, 9(2), 2014, 131–141

unknown locality as sister taxon of A. heterodermus from
Majuy with moderate support and placed A. heterodermus
from Madrid sites 1 and 2 (Mad1 and Mad2; A. hetero-
dermus Mad) as a moderately supported sister clade (BA:
0.95; ML: 72%).
Within subgroup B, both tree building methods re-
vealed Anolis heterodermus from Antioquia JN112690
(A. heterodermus Ant) and A. heterodermus from Huila
JN112688 (A. heterodermus Hul) as sister taxa with low
support (BA: 0.86, ML: 53%), and A. heterodermus from
Caldas JN112689 (A. heterodermus Cal) as sister of this
clade, with maximum support. Anolis vanzolinii and A. in-
derenae appeared as moderately supported successive sis-
ter taxa (BA: 1, ML: 72% and 79%, respectively).
Within group C, an individual of Anolis heterodermus
from Gachancipa (A. heterodermus Gac) and individuals
from Tabio sites 1 and 2 (Tab1 and Tab2; A. heterodermus
Tab) were placed in both analyses in a maximally support-
ed polytomy (A. heterodermus Gac/Tab lineage). Finally, in
both analyses A. nicefori was inferred to be sister taxon of
Figure 2. Bayesian inference trees showing the evolutionary relationships between Anolis heterodermus from middle portion of the Eastern and Central
cordilleras of Colombia and other species of the A. heterodermus series. Left: Tree based on 1,433 bp mtDNA (partial ND2 gene and tRNA genes). Right:
Tree based on 3,728 bp of combined evidence (1,967 bp mtDNA of concatenated mitochondrial sequences and 1,761 bp RAG1 nDNA). Trees rooted with
A. neblininus, A. calimae, and Polychrus marmoratus (not shown). Posterior probability values for the Bayesian analysis (BA) above and bootstrap support
percentage values for the maximum likelihood (ML) analysis below. Support values along branches are thorough bootstrap values > 50. Bold branches are
supported by maximum posterior probability in BA and maximum bootstrap support in ML analyses.
Table 2. Mean uncorrected p-distances between individuals per locality of Anolis heterodermus, species of the Anolis heterodermus series, A. neblininus, and
A. calimae, based on a 1,036 bp fragment of ND2. Values below the diagonal are average percentages, above the diagonal are standard error (500 bootstrap
replicates), and along the diagonal and in bold are within-locality divergences (X ± SE). Con = A. heterodermus Conejera, Mad = A. heterodermus Madrid,
Gb = A. heterodermus Genbank AY296144, Cal = A. heterodermus Caldas, Ant = A. heterodermus Antioquia, Hul = A. heterodermus Huila, vanz = A. vanzolinii,
inde = A. inderenae, Gac/Tab = A. heterodermus Gachancipa/Tabio, nicef = A. nicefori, eusk = A. euskalerriari, nebli = A. neblininus, calim = A. calimae, n =
number of sequences.
n Con Maj Mad Gb Cal Ant Hul vanz inde Gac/Tab nicef eusk nebli calim
Con 2 0.41 ± 0.40 0.42 0.45 0.35 0.83 0.90 0.84 0.80 0.95 1.12 1.19 1.60 2.46 2.27
Maj 1 0.43 0.37 0.74 0.76 0.71 0.19 0.18 0.93 1.15 1.17 1.59 2.44 2.25
Mad 2 0.86 0.32 0.54 ± 0.40 0.54 0.11 0.13 0.82 0.83 0.96 1.13 1.20 1.61 2.42 2.23
Gb 1 1.51 1.29 2.53 0.83 0.90 0.82 0.80 0.93 1.12 1.17 1.58 2.44 2.21
Cal 1 8.49 8.24 6.81 6.37 0.46 0.44 0.65 0.86 1.11 1.22 1.53 2.53 2.21
Ant 1 8.46 8.71 7.71 7.26 1.95 0.46 0.66 0.91 1.14 1.25 1.54 2.55 2.25
Hul 1 8.43 8.18 7.03 6.63 1.94 1.83 0.64 0.83 1.13 1.21 1.53 2.46 2.21
vanz 1 7.50 7.26 6.54 5.89 3.83 4.31 3.94 0.81 1.15 1.17 1.60 2.37 2.16
inde 1 11.35 11.09 8.54 8.25 6.86 7.13 6.49 5.40 1.14 1.12 1.61 2.45 2.24
Gac/Tab 3 11.26 12.07 11.61 11.26 11.52 11.13 10.99 11.42 11.13 0.55 ± 0.40 0.93 1.35 2.41 2.26
nicef 1 12.38 12.56 12.68 11.82 12.61 12.89 12.21 12.12 11.67 7.40 1.42 2.36 2.18
eusk 1 19.06 20.20 18.28 18.43 17.36 17.36 17.07 18.07 18.78 16.82 16.49 2.46 2.22
nebli 1 35.84 39.06 36.24 35.63 37.15 37.41 36.07 34.71 36.55 35.28 34.54 35.43 2.20
calim 1 31.34 31.24 30.66 30.28 29.71 30.95 29.57 29.56 30.75 30.64 30.12 30.87 29.29
Unknown Evolutionary Lineages and Population Differentiation in Anolis heterodermus (Squamata: Dactyloidae)
from the Eastern and Central Cordilleras of Colombia Revealed by DNA Sequence Data
Mario Vargas-Ramírez, Rafael Moreno-Arias
135
South American Journal of Herpetology, 9(2), 2014, 131–141
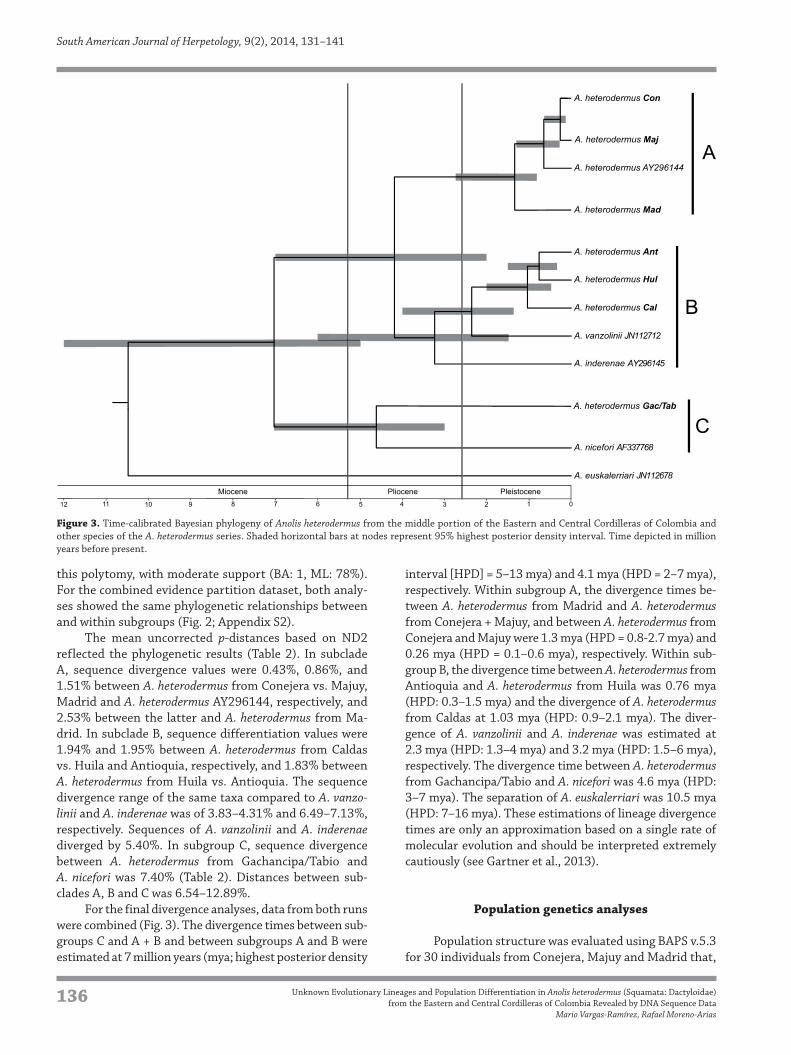
this polytomy, with moderate support (BA: 1, ML: 78%).
For the combined evidence partition dataset, both analy-
ses showed the same phylogenetic relationships between
and within subgroups (Fig. 2; Appendix S2).
The mean uncorrected p-distances based on ND2
reflected the phylogenetic results (Table 2). In subclade
A, sequence divergence values were 0.43%, 0.86%, and
1.51% between A. heterodermus from Conejera vs. Majuy,
Madrid and A. heterodermus AY296144, respectively, and
2.53% between the latter and A. heterodermus from Ma-
drid. In subclade B, sequence differentiation values were
1.94% and 1.95% between A. heterodermus from Caldas
vs. Huila and Antioquia, respectively, and 1.83% between
A. heterodermus from Huila vs. Antioquia. The sequence
divergence range of the same taxa compared to A. vanzo-
linii and A. inderenae was of 3.83–4.31% and 6.49–7.13%,
respectively. Sequences of A. vanzolinii and A. inderenae
diverged by 5.40%. In subgroup C, sequence divergence
between A. heterodermus from Gachancipa/Tabio and
A. nicefori was 7.40% (Table 2). Distances between sub-
clades A, B and C was 6.54–12.89%.
For the final divergence analyses, data from both runs
were combined (Fig. 3). The divergence times between sub-
groups C and A + B and between subgroups A and B were
estimated at 7 million years (mya; highest posterior density
interval [HPD] = 5–13 mya) and 4.1 mya (HPD = 2–7 mya),
respectively. Within subgroup A, the divergence times be-
tween A. heterodermus from Madrid and A. heterodermus
from Conejera + Majuy, and between A. heterodermus from
Conejera and Majuy were 1.3 mya (HPD = 0.8-2.7 mya) and
0.26 mya (HPD = 0.1–0.6 mya), respectively. Within sub-
group B, the divergence time between A. heterodermus from
Antioquia and A. heterodermus from Huila was 0.76 mya
(HPD: 0.3–1.5 mya) and the divergence of A. heterodermus
from Caldas at 1.03 mya (HPD: 0.9–2.1 mya). The diver-
gence of A. vanzolinii and A. inderenae was estimated at
2.3 mya (HPD: 1.3–4 mya) and 3.2 mya (HPD: 1.5–6 mya),
respectively. The divergence time between A. heterodermus
from Gachancipa/Tabio and A. nicefori was 4.6 mya (HPD:
3–7 mya). The separation of A. euskalerriari was 10.5 mya
(HPD: 7–16 mya). These estimations of lineage divergence
times are only an approximation based on a single rate of
molecular evolution and should be interpreted extremely
cautiously (see Gartner et al., 2013).
Population genetics analyses
Population structure was evaluated using BAPS v.5.3
for 30 individuals from Conejera, Majuy and Madrid that,
Figure 3. Time-calibrated Bayesian phylogeny of Anolis heterodermus from the middle portion of the Eastern and Central Cordilleras of Colombia and
other species of the A. heterodermus series. Shaded horizontal bars at nodes represent 95% highest posterior density interval. Time depicted in million
years before present.
Unknown Evolutionary Lineages and Population Differentiation in Anolis heterodermus (Squamata: Dactyloidae)
from the Eastern and Central Cordilleras of Colombia Revealed by DNA Sequence Data
Mario Vargas-Ramírez, Rafael Moreno-Arias
136
South American Journal of Herpetology, 9(2), 2014, 131–141

according to the phylogenetic analyses, corresponded to
one evolutionary lineage (subgroup A), and 33 individu-
als from Gachancipa and Tabio that corresponded to a dif-
ferent evolutionary lineage (subgroup C). The subgroup
A alignment included 12 variable and 12 parsimony in-
formative sites without missing data and the subgroup C
alignment included five variable and two parsimony in-
formative sites without missing data.
The genetic mixture analyses of BAPS revealed one
population (K = 1) as the most likely division for each
subgroup. The admixture analysis revealed all individu-
als assigned to the corresponding population with 100%
probability. In the Snn comparison test between localities
in subgroup A, Conejera vs. Madrid and Majuy was sta-
tistically significant (P < 0.001), suggesting demographic
independence. Therefore subpopulations from Conejera
and Madrid/Majuy should be considered as different
management units (sensu Moritz, 1994). Under the 95%
criterion, the parsimony haplotype network assigned in-
dividuals from Conejera and Madrid/Majuy to two clearly
distinct haplotype clusters, differing by a minimum of six
mutational steps (Fig. 4.I). Sequences of individuals from
Conejera corresponded to two haplotypes (a–b) and indi-
viduals from Madrid/Majuy to six haplotypes (c–h), the
latter with a maximum of six mutational steps between
them (Fig. 4.I). For the Madrid/Majuy subpopulation,
TCS revealed e as the ancestral haplotype. This subpopu-
lation has higher haplotype and nucleotide diversity than
the Conejera subpopulation (Table 3).
The same analyses revealed a star-like network
for the Gachancipa/Tabio population in subgroup C
Figure 4. Parsimony network based on 533 bp of COI from 30 individu-
als of Anolis heterodermus in subgroup B (I) and 33 individuals in sub-
group C (II). Each circle corresponds to a distinct haplotype and lines
connecting haplotypes to one mutational step. Small black circles are
missing node haplotypes. Inset symbol size indicates haplotype frequen-
cy. Ancestral haplotypes in each network are indicated by a thicker line.
Inset photographs: Above: female of A. heterodermus from the Conejera
population, Cerro de la Conejera, Bogotá D.C., Colombia. Below: female
of A. heterodermus from the Gachancipa/Tabio population from Ga-
chancipa, Cundinamarca, Colombia.
Table 3. Genetic parameters and results of the model of population expansion for each population/subpopulation of Anolis heterodermus. n = sample size;
S = number of segregating sites; h = number of haplotypes; Hd = haplotype diversity and π = nucleotide diversity. Parameters of the population expansion
model: = age of expansion, 0 = population size before expansion,
1 = population size after expansion. When SSD values in the goodness-of-fit test
for the mismatch distribution are non-significant (P > 0.05), the data do not deviate from the expectation of population expansion. Negative significant
D and FS values are expected when population expansion occurs. Significant SSD, D and F
S values asterisked. The number of bootstrap replicates in the
mismatch test was 1,000.
Population Con Mad/Maj Con/Mad/Maj Gac/Tab
Subgroup A A A C
n 7 23 30 33
S 1 5 12 5
h 2 6 8 5
Hd 0.476 0.842 0.883 0.460
π 0.00089 0.00430 0.00830 0.00116
Model parameters
0.717 (0–22.799) 4.258 (0–7.707) 9.219 (0.08–14.434) 0.701 (0.000–1.848)
00 (0–0.021) 0 (0–4.127) 0.002 (0–9.290) 0 (0–0.313)
199999 (1.102–99999) 5.527 (3.030–99999) 6.743 (3.770–99999) 3.523 (0.840–99999)
Goodness-of-fit test
SSD 0.017170 0.013330 0.014260 0.000074
P 0.271 0.541 0.670 0.964
Tajima’s D 0.55902 2.23079 1.55363 -1.32863
P 0.833 0.991 0.962 0.083
Fu’s FS
0.58867 0.28889 1.51809 -1.81211
P 0.459 0.585 0.776 0.050*
Unknown Evolutionary Lineages and Population Differentiation in Anolis heterodermus (Squamata: Dactyloidae)
from the Eastern and Central Cordilleras of Colombia Revealed by DNA Sequence Data
Mario Vargas-Ramírez, Rafael Moreno-Arias
137
South American Journal of Herpetology, 9(2), 2014, 131–141

consisting of five haplotypes (A–E), suggesting popula-
tion expansion (Fig. 4.II). This population showed lower
haplotype and nucleotide diversity than both subpopula-
tions of Anolis heterodermus in subgroup A (Table 3). The
pairwise sequence mismatch distributions were not sig-
nificantly different from the sudden expansion model of
Rogers and Harpending (1992; Table 3). Finally, the popu-
lation from Gachancipa/Tabio in subgroup C also showed
a significantly negative Fu’s Fs value and negative Tajima’s
D (Table 3), suggesting that this population could have ex-
perienced a more pronounced and sudden demographic
expansion event.
DISCUSSION
Previous morphological and molecular data indicat-
ed that more than one species might be grouped within
Anolis heterodermus (Lazell, 1969; Williams et al., 1996;
Castañeda and de Queiroz, 2011). The present study con-
firms A. heterodermus as a species complex and identifies
three independent evolutionary lineages within it based
on the following lines of evidence. First, congruent results
from mtDNA and nDNA analyses revealed that A. hetero-
dermus comprises three strongly supported subgroups
and is polyphyletic with respect to the other species of the
A. heterodermus series (Fig. 2). Second, uncorrected p-dis-
tances within A. heterodermus exceed the values found be-
tween different recognized species of the A. heterodermus
series (Table 2) and fall within the differentiation between
closely related Anolis species of 5.0–22.5% (for review see
Gartner et al., 2013). Consequently, relying on molecu-
lar criteria we conclude that the individuals of the three
clades correspond to independently evolving evolutionary
lineages and therefore qualify as distinct species (Avise
and Ball, 1990). However, to fully resolve the taxonomic
problems within the A. heterodermus series, an integrative
approach is necessary, combining evidence from molecular
markers with an extensive examination of the morphol-
ogy and color patterns of live and museum specimens.
The diversification times of the studied species of
the Anolis heterodermus series date to the Middle Miocene
to Pleistocene, falling within a period of significant oro-
genic and climatic events in northern South America. The
divergence between subgroups C and A + B was estimated
to have occurred during the Middle–Late Miocene, dur-
ing an episode of volcanism and uplift of the Central and
Eastern Cordilleras (Hoorn et al., 1995; Guerrero, 1997).
The estimated dates of divergence of subgroups A and B,
within subgroup C and A. inderenae fall within a period
in the Pliocene when the northern Andes experienced
extensive uplift, especially along the eastern Cordilleras
(Gregory-Wodzicki, 2000; Mora et al., 2008; Hoorn et al.,
2010). The widespread Pleistocene differentiation of
A. vanzolinii, A. heterodermus from Antioquia, Huila, and
Caldas within subgroup B and A. heterodermus from Cone-
jera, Majuy, and Madrid within subgroup A suggest that
the species complex was deeply affected by the dramatic
climatic changes that characterize this period. Similarly,
the evidence for recent, sudden demographic expansion
in A. heterodermus populations from the middle portion
of the Eastern Cordillera suggests that these populations
might have suffered isolation and decline due to extreme
environmental conditions followed by recent popula-
tion expansion. The overall cooling of temperatures in
the Pleistocene may have led to downhill glacier expan-
sion and severe uphill compression of montane vegeta-
tion areas (Hooghiemstra and van der Hammen, 2004;
Hooghiemstra et al., 2006). Resulting dispersal barriers
might have caused the isolation and reduction of A. het-
erodermus populations at higher elevations. After such
dramatic climatic changes, favorable conditions might
have allowed secondary contact, as evidenced by the cur-
rent possible sympatric/parapatric distribution of A. het-
erodermus lineages in the middle portion of the Eastern
Cordillera and other species of the A. heterodermus series
elsewhere (see Fig. 1).
Due to the fact that effective conservation measures
depend largely on a good knowledge of the systematics
of the species (Mace, 2004), the definition of indepen-
dent genealogical lineages within Anolis heterodermus is
critical for their assessment and future management. Our
analyses revealed three cryptic, independent evolutionary
lineages that should be considered as evolutionary signifi-
cant units (sensu Moritz, 1994) and unconfirmed candi-
date species for conservation purposes (Vieites et al.,
2009). Furthermore, each population of A. heterodermus
within subgroup A (Conejera and Madrid/Majuy) should
be considered a different management unit (sensu Moritz,
1994), each meriting conservation measures. In this pre-
liminary study, only a small portion of the distribution
range of the species was sampled. A range wide molecular
and morphological evaluation aimed at describing these
and identifying any further independently evolving evo-
lutionary lineages within A. heterodermus complex, as well
as determining their distribution ranges, is warranted.
Etter (1993) reported that in Colombia only 27% of
the northern Andean montane tropical forest remains, and
Armenteras et al. (2003) reported that on the Eastern Cor-
dillera only 41% and 45% of the original, pre-transformed
Andean and sub-Andean forest remains. Currently, those
fragments, which comprise the restricted habitat of Anolis
heterodermus, are further threatened by high human pop-
ulation density and extreme continuous transformation.
Therefore, scientific research aimed at (i) providing infor-
mation to increase the general knowledge about Andean
anoles and (ii) guiding the design and implementation of
conservation measures, as well as protection of what is
left of Andean montane tropical forests and environmen-
tal education, should be promptly implemented
Unknown Evolutionary Lineages and Population Differentiation in Anolis heterodermus (Squamata: Dactyloidae)
from the Eastern and Central Cordilleras of Colombia Revealed by DNA Sequence Data
Mario Vargas-Ramírez, Rafael Moreno-Arias
138
South American Journal of Herpetology, 9(2), 2014, 131–141

Excoffier L., Laval G., Schneider S. 2005. ARLEQUIN: an integrated
software package for population genetics data analysis, Version
3.11. Evolutionary Bioinformatics Online 1:47–50.
Etter A. 1993. Diversidad ecosistemica en Colombia hoy. Pp. 43–61,
in Cárdenas, S., Correa, H.D. (Eds.), Nuestra Diversidad Biológica.
Fundación Alejandro Escobar, Colección María Restrepo de Angel.
CEREC, Bogotá.
Gartner G.E.A., Gamble T., Jaffe A.L., Harrison A., Losos J.B.
2013. Left–right dewlap asymmetry and phylogeography of Anolis
lineatus on Aruba and Curaçao. Biological Journal of the Linnean
Society 110: 409–426. doi:10.1111/bij.12131
Glor R.E., Kolbe J.J, Powell R, Larson A, Losos J.B. 2003.
Phylogenetic analysis of ecological and morphological
diversification in Hispaniolan trunk–ground anoles (Anolis cybotes
group). Evolution 57: 2383–2397. doi:10.1554/02-369
Gregory-Wodzicki K.M. 2000. Uplift history of the central and
northern Andes: a review. Geological Society of America Bulletin 112:
1091–1105. doi:10.1130/0016-7606(2000)112<1091:UHOTCA>2.
0.CO;2
Guarnizo C.E., Amézquita A., Bermingham E. 2009. The relative
roles of vicariance versus elevational gradients in the genetic
differentiation of the high Andean tree frog, Dendropsophus labialis.
Molecular Phylogenetics and Evolution 50:84–92. doi:10.1016/j.
ympev.2008.10.005
Guerrero J. 1997. Stratigraphy, sedimentary environments, and the
Miocene uplift of the Colombian Andes. Pp. 15–43, in Kay R.F.,
Madden R.H., Cifelli R.L., Flynn J.J. (Eds.), Vertebrate Paleontology
in the Neotropics: The Miocene Fauna of La Venta, Colombia.
Smithsonian Institution Press, Washington D.C.
Hooghiemstra H., Van der Hammen T. 2004. Quaternary ice-age
dynamics in the Colombian Andes: developing an understanding of
our legacy. Philosophical Transactions of the Royal Society of London B
359:173–180. doi:10.1098/rstb.2003.1420
Hooghiemstra H., Wijninga V.M., Cleef A.M. 2006. The
paleobotanical records of Colombia: implications for biogeography
and biodiversity. Annals of the Missouri Botanical Garden 96:297–325.
Hoorn C., J., Sarmiento G., Lorente M. 1995. Andean
tectonics as a cause for the changing drainage patterns in
Miocene northern South America. Geology 23: 237–240.
doi:10.1130/0091-7613(1995)023<0237:ATAACF>2.3.CO;2
Hoorn C., Wesselingh F.P., ter Steege H., Bermudez M.A., Mora A.,
Sevink J., ..., Antonelli A. 2010. Amazonia through time: Andean
uplift, climate change, landscape evolution, and biodiversity. Science
330: 927–931. doi:10.1126/science.1194585
Hudson R.R. 2000. A new statistic for detecting genetic differentiation.
Genetics 155:2011–2014.
Jackman T.R., Larson A., de Queiroz K., Losos J.B.
1999. Phylogenetic relationships and tempo of early
diversification in Anolis lizards. Systematic Biology 48:254–285.
doi:10.1080/106351599260283
Jackman T.R., Irschick D.J., de Queiroz K., Losos J.B., Larson A.
2002. Molecular phylogenetic perspective on evolution of lizards of
the Anolis grahami series. Journal of Experimental Zoology (Molecular
and Developmental Evolution) 294: 1–16.
Jezkova T., Leal M., Rodríguez-Robles J.A. 2009. Living together
but remaining apart: comparative phylogeography of Anolis
poncensis and A. cooki, two lizards endemic to the aridlands of
Puerto Rico. Biological Journal of the Linnean Society 96: 617–634.
doi:10.1111/j.1095-8312.2008.01147.x
Lanfear R., Calcott B., Ho S.Y.W., Guindon S. 2012. PartitionFinder:
combined selection of partitioning schemes and substitution
models for phylogenetic analyses. Molecular Biology and Evolution
29:1695–1701. doi:10.1093/molbev/mss020
Lazell J.D. Jr. 1969. The genus Phenacosaurus (Sauria: Iguanidae).
Breviora 325:1–24.
Librado P., Rozas J. 2009. DnaSP v5: a software for comprehensive
analysis of DNA polymorphism data. Bioinformatics 25:1451–1452.
doi:10.1093/bioinformatics/btp187
ACKNOWLEDGMENTS
We thank the Grupo de Conservación y Biodivers-
idad del Instituto de Ciencias Naturales de la Universidad
Nacional de Colombia and Fundación Biodiversa Colom-
bia for institutional and logistical support. We are grate-
ful to Olga Victoria Castaño-Mora, Gladys Cárdenas-Are-
valo, Michel Estefan-Agudelo and Ecotreck foundation for
fieldwork assistance. Thanks to Claudia Munera from In-
stituto de Investigaciones biológicas Alexander von Hum-
boldt (IAvH) in Colombia for information on distribution
of high elevation anole species. Thanks for help with the
manuscript style go to Fiona Mulvey. We thank the Asso-
ciate Editor and two anonymous reviewers for their con-
structive comments and suggestions that have improved
the quality of the manuscript. Sampling and access to ge-
netic resources allowed by Resolución 0255 of the ANLA,
Ministerio de Ambiente y Desarrollo Sostenible de Colom-
bia (Permiso Marco otorgado a la Universidad Nacional de
Colombia de recolección de especímenes de especies silves-
tres con fines de investigación científica no comercial para
el proyecto “Estructuración a pequeña escala en reptiles
de alta montaña en la cordillera Oriental de Colombia”).
Mario Vargas-Ramírez’s research in Germany is funded by
the Humboldt Foundation (Georg Forster fellowship).
REFERENCES
Armenteras D., Gast F., Villareal H. 2003. Andean forest
fragmentation and representativeness of protected natural areas in
the eastern Andes, Colombia. Biological Conservation 113:245–256.
doi:10.1016/S0006-3207(02)00359-2
Avise J.C., Ball R.M.1990. Principles of genealogical concordance in
species concepts and biological taxonomy. Pp. 45–67, in Futuyma,
D., Antonovics, J. (Eds.), Surveys in Evolutionary Biology. Oxford
University Press, Oxford.
Castañeda M.R., de Queiroz K. 2011. Phylogenetic relationships
of the Dactyloa clade of Anolis lizards based on nuclear and
mitochondrial DNA sequence data. Molecular Phylogenetics and
Evolution 61:784–800. doi:10.1016/j.ympev.2011.07.004
Castañeda M.R., de Queiroz K. 2013. Phylogeny of the Dactyloa
clade of Anolis lizards: New insights from combining morphological
and molecular data. Bulletin of the Museum of Comparative Zoology
160:345–398. doi:10.3099/0027-4100-160.7.345
Clement M., Posada D., Crandall K.A. 2000. TCS: a computer
program to estimate gene genealogies. Molecular Ecology 9:1657–
1660. doi:10.1046/j.1365-294x.2000.01020.x
Corander J., Marttinen P. 2006. Bayesian identification of admixture
events using multilocus molecular markers. Molecular Ecology
15:2833-2843. doi:10.1111/j.1365-294X.2006.02994.x
Corander J., Siren J., Arjas E. 2008. Bayesian spatial modeling of
genetic population structure. Computational Statistics 23:111–129.
doi:10.1007/s00180-007-0072-x
Creer D.A., de Queiroz K., Jackman T.R., Losos J.B., Larson A.
2001. Systematics of the Anolis roquet series of the southern Lesser
Antilles. Journal of Herpetology 35: 428–441. doi:10.2307/1565961
Dunn E.R. 1944. The lizards of genus Phenacosaurus. Caldasia 3:57–62.
Drummond A.J., Suchard M.A., Xie D., Rambaut A. 2012. Bayesian
phylogenetics with BEAUti and the BEAST 1.7. Molecular Biology
and Evolution 29: 1969–1973. doi:10.1093/molbev/mss075
Unknown Evolutionary Lineages and Population Differentiation in Anolis heterodermus (Squamata: Dactyloidae)
from the Eastern and Central Cordilleras of Colombia Revealed by DNA Sequence Data
Mario Vargas-Ramírez, Rafael Moreno-Arias
139
South American Journal of Herpetology, 9(2), 2014, 131–141

Primack R., Rozzi R., Feisinger P., Dirzo R., Massardo F.
2001. Fundamentos de Conservación Biológica: Perspectivas
Latinoamericanas. Fondo de Cultura Economica, México D.F.
Rambaut A., Drummond A.J. 2007. Tracer v1.4. Available from
http://beast.bio.ed.ac.uk/software/tracer/
Rambaut A. 2009. FigTree v1.3.1. Available from http://tree.bio.ed.ac.
uk/software/figtree/
Rodríguez-Robles J.A., Jezkova T., Leal M. 2008. Genetic
structuring in the threatened ‘Lagartijo del Bosque Seco’ (Anolis
cooki) from Puerto Rico. Molecular Phylogenetics and Evolution 46:
503–514. doi:10.1016/j.ympev.2007.11.006
Rogers A.R., Harpending H. 1992. Population growth makes waves
in the distribution of pairwise genetic differences. Molecular Biology
and Evolution 9:552–569.
Ronquist F., Huelsenbeck J.P. 2003. MrBayes 3: Bayesian phylogenetic
inference under mixed models. Bioinformatics 19:1572–1574.
doi:10.1093/bioinformatics/btg180
Rueda J.V., Hernández-Camacho J.I. 1988. Phenacosaurus inderenae
(Sauria: Iguanidae), nueva especie gigante, proveniente de la
Cordillera Oriental de Colombia. Trianea 2: 339–350.
Schneider S., Excoffier L. 1999. Estimation of past demographic
parameters from the distribution of pairwise differences when
the mutation rates vary among sites: application to human
mitochondrial DNA. Genetics 152:1079–1089.
Silvestro D., Michalak I. 2012. RAxMLGUI: a graphical frontend for
RAxML. Organisms Diversity & Evolution 12:335–337. doi:10.1007/
s13127-011-0056-0
Stadtmüller T. 1987. Cloud Forest in the Humid Tropics: A
Bibliographic Review. United Nations University y Centro
Agronómico de Investigación y Enseñanza. Turrialba. Costa Rica.
Stamatakis A. 2006. RAxML-VI-HPC: maximum likelihood-based
phylogenetic analyses with thousands of taxa and mixed models.
Bioinformatics 22:2688–2690. doi:10.1093/bioinformatics/btl446
Tamura K., Peterson D., Peterson N., Stecher G., Nei M., Kumar
S. 2011. MEGA 5: Molecular evolutionary genetics analysis
using maximum likelihood, evolutionary distance, and maximum
parsimony methods. Molecular Biology and Evolution 28:2731–2739.
doi:10.1093/molbev/msr121
Torres-Carvajal O., Ayala F., Carvajal-Campos A. 2010. Reptilia,
Squamata, Iguanidae, Anolis heterodermus Duméril, 1851:
Distribution extension, first record for Ecuador and notes on color
variation. Ckeck List 6:189–190.
Vences M., Guayasamin J.M., Miralles A., de La Riva I. 2013.
To name or not to name: Criteria to promote economy of change
in Linnaean classification schemes. Zootaxa 3636:201–244.
doi:10.11646/zootaxa.3636.2.1
Vieites D.R., Wollenberg K.C., Andreone F., Köhler J., Glaw
F., Vences M., 2009. Vast underestimation of Madagascar’s
biodiversity evidenced by an integrative amphibian inventory.
Proceedings of the National Academy of Sciences USA 106:8267–8272.
doi:10.1073/pnas.0810821106
Williams E.E., Praderio M.J., Gorzula S. 1996. A phenacosaur from
Chimantá Tepui, Venezuela. Breviora 506:1–15.
Willmott K., Hall J.W., Lamas G. 2001. Systematics of Hypanartia
(Lepidoptera: Nymphalidae: Nymphalinae) with a test for
geographical speciation mechanisms in the Andes. Systematic
Entomology 26:369–399. doi:10.1046/j.1365-3113.2001.00157.x
Lynch J.D. 1999. Ranas pequeñas, la geometría de la evolución, y la
especiación en los Andes Colombianos. Revista de la Academia
Colombiana de Ciencias Exactas, Físicas y Naturales 23:143–159.
Mace G.M. 2004. The role of taxonomy in species conservation.
Philosophical Transactions of the Royal Society of London B 359:711–
719. doi:10.1098/rstb.2003.1454
Macey J.R., Schulte II J.A., Ananjeva N.B., Larson A., Rastegar-
Pouyani N., Shammakov S.M., Papenfuss T.J. 1998.
Phylogenetic relationships among agamid lizards of the Laudakia
caucasia species group: testing hypotheses of biogeographic
fragmentation and an area cladogram for the Iranian Plateau.
Molecular Phylogenetics and Evolution 10:118–131.
Miyata K. 1983. Notes on Phenacosaurus heterodermus in the Sabana de
Bogotá, Colombia. Journal of Herpetology 17:102–105.
Mora A., Parra M., Strecker M.R., Sobel E.R., Hooghiemstra
H., Torres V., Vallejo-Jaramillo J. 2008. Climatic forcing
of asymmetric orogenic evolution in the Eastern Cordillera of
Colombia. Geological Society of America Bulletin 120: 930–949.
doi:10.1130/B26186.1
Moreno-Arias R.A., Urbina-Cardona N. 2013. Population dynamics
of the Andean lizard Anolis heterodermus: fast-slow demographic
strategies in fragmented scrubland landscapes. Biotropica
45:253–261.
Moritz C.1994. Defining ‘Evolutionarily Significant Units’
for conservation. Trends in Evolution & Ecology 9:373–375.
doi:10.1016/0169-5347(94)90057-4
Mueses-Cisneros J.J. 2006. Anolis heterodermus. Distribution.
Herpetological review 37: 493.
Myers C.W., Donnelly M.A. 1996. A new herpetofauna from Cerro
Yaví, Venezuela: first results of the Robert G. Goelet American
Museum-Terramar Expedition to the northwestern Tepuis.
American Museum Novitates 3172:1–56.
Myers N., Mittermeier R.A., Mittermeier C.G., Fonseca G.A.B.,
Kent J. 2000. Biodiversity hotspots for conservation priorities.
Nature 24:853–858.
Nicholson K.E., Glor R.E., Kolbe J.J., Larson A., Hedges S.B.,
Losos J.B. 2005. Mainland colonization by island lizards. Journal of
Biogeography 32:929–938. doi:10.1111/j.1365-2699.2004.01222.x
Nicholson K.E., Crother B.I., Guyer C., Savage J.M. 2012. It is time
for a new classification of anoles (Squamata: Dactyloidae). Zootaxa
3477:1–108.
Nylander J.A.A., Ronquist F., Huelsenbeck J.P., Nieves-Aldrey J.L.
2004. Bayesian phylogenetic analysis of combined data. Systematic
Biology 53:47–67. doi:10.1080/10635150490264699
Olson D.M., Dinerstein E. 1997. Global 2000: A representation
approach to conserving the earth�s distinctive ecoregions. World
Wildlife Foundation, Washington, D.C.
Orme C.D.L., Davies R.G., Burgess M., Eigenbrod F., Pickup N.,
Olson V.A., ..., Owens I.P.F. 2005. Global hotspots of species
richness are not congruent with endemism or threat. Nature
18:1016–1019. doi:10.1038/nature03850
Osorno-Mesa H., Osorno-Mesa E. 1946. Anotaciones sobre lagartos
del género Phenacosaurus. Caldasia 17:123–130.
Poe S. 1998. Skull characters and the cladistic relationships of the
Hispaniolan dwarf twig Anolis. Herpetological Monographs 12:192–
236. doi:10.2307/1467021
Poe S. 2004. Phylogeny of anoles. Herpetological Monographs 18:37–89.
doi:10.1655/0733-1347(2004)018[0037:POA]2.0.CO;2
Unknown Evolutionary Lineages and Population Differentiation in Anolis heterodermus (Squamata: Dactyloidae)
from the Eastern and Central Cordilleras of Colombia Revealed by DNA Sequence Data
Mario Vargas-Ramírez, Rafael Moreno-Arias
140
South American Journal of Herpetology, 9(2), 2014, 131–141

ONLINE SUPPORTING INFORMATION
The following Supporting Information is available
for this article online:
Appendix S1. GenBank accession numbers of DNA se-
quences used in this study.
Appendix S2. Description of the results of the phyloge-
netic analyses of COI, combined mitochondrial, and com-
bined evidence partitions.
Figure S1. Bayesian inference tree for the COI fragment
showing the evolutionary relationships between Anolis
heterodermus from middle portion of the Eastern and
Central Cordilleras of Colombia and other species of the
A. heterodermus series. Trees rooted with A. neblininus,
A. calimae, and Polychrus marmoratus (not shown). Pos-
terior probability values for the Bayesian analysis (BA)
above and bootstrap support percentage values for the
maximum likelihood (ML) analysis below. Support values
along branches are thorough bootstrap values > 50%. Bold
branches are supported by maximum posterior probabili-
ty in BA and maximum bootstrap support in ML analyses.
Figure S2. Bayesian inference trees showing the evolu-
tionary relationships between Anolis heterodermus from
middle portion of the Eastern and Central Cordilleras of
Colombia and other species of the A. heterodermus series.
Left: Combined mitochondrial tree based on 1,967 bp
mtDNA (partial ND2, adjacent tRNA genes, and COI).
Right: Tree based on 1,761 bp of RAG1. Trees rooted
with A. neblininus, A. calimae, and Polychrus marmoratus
(not shown). Posterior probability values for the Bayes-
ian analysis (BA) above and bootstrap support percentage
values for the maximum likelihood (ML) analysis below.
Support values along branches are thorough bootstrap
values > 50%. Bold branches are supported by maximum
posterior probability in Bayesian analyses and maximum
bootstrap support in ML analyses.
Table S1. Evolutionary models selected by the Bayesian
Information Criterion in partitionfinder (Lanfear et al.
2012) for the optimal partitioning schemes.
Unknown Evolutionary Lineages and Population Differentiation in Anolis heterodermus (Squamata: Dactyloidae)
from the Eastern and Central Cordilleras of Colombia Revealed by DNA Sequence Data
Mario Vargas-Ramírez, Rafael Moreno-Arias
141
South American Journal of Herpetology, 9(2), 2014, 131–141