antibodies raised against receptor-binding domain of plasmodium knowlesi duffy binding protein...
TRANSCRIPT

Antibodies raised against receptor-binding domain of Plasmodiumknowlesi Duffy binding protein inhibit erythrocyte invasion
Agam P. Singh a, Sunil K. Puri b, Chetan E. Chitnis a,*a Malaria Research Group, International Centre for Genetic Engineering and Biotechnology (ICGEB), P.O. Box 10504, Aruna Asaf Ali Marg, New
Delhi 110067, Indiab Central Drug Research Institute (CDRI), Lucknow, India
Received 12 November 2001; received in revised form 8 January 2002; accepted 15 January 2002
Abstract
Erythrocyte invasion by malaria parasites requires specific receptor-ligand interactions. Plasmodium vivax and Plasmodium
knowlesi are completely dependent on binding the Duffy blood group antigen to invade human erythrocytes. P. knowlesi invades
rhesus erythrocytes by multiple pathways using the Duffy antigen as well as alternative receptors. Plasmodium falciparum binds
sialic acid residues on glycophorin A as well as other sialic acid-independent receptors to invade human erythrocytes. Parasite
proteins that mediate these interactions belong to a family of erythrocyte binding proteins, which includes the P. vivax Duffy
binding protein, 175 kDa P. falciparum erythrocyte binding antigen (EBA-175), P. knowlesi a protein, which binds human and
rhesus Duffy antigens, and P. knowlesi b and g proteins, which bind Duffy-independent receptors on rhesus erythrocytes. The
receptor-binding domains of these proteins lie in conserved, N-terminal, cysteine-rich regions that are referred to as region II. Here,
we have examined the feasibility of inhibiting erythrocyte invasion with antibodies directed against receptor-binding domains of
erythrocyte binding proteins. Region II of P. knowelsi a protein (PkaRII), which binds the Duffy antigen, was expressed as a
secreted protein in insect cells and purified from culture supernatants. Rabbit antibodies raised against recombinant PkaRII were
tested for inhibition of erythrocyte binding and invasion. Antibodies raised against PkaRII inhibit P. knowlesi invasion of both
human and rhesus erythrocytes. These data provide support for the development of recombinant vaccines based on the homologous
binding domains of P. vivax Duffy binding protein and P. falciparum EBA-175. # 2002 Elsevier Science B.V. All rights reserved.
Keywords: Erythrocyte invasion; Erythrocyte binding proteins; Host-parasite interactions; Malaria vaccines
1. Introduction
Malaria parasites have an obligate intra-erythrocytic
stage in vertebrate hosts. Invasion of erythrocytes by
malaria parasites requires specific receptor-ligand inter-
actions [1]. For example, Plasmodium vivax is absolutely
dependent on interaction with the Duffy blood group
antigen to invade human erythrocytes [2]. As a result,
Duffy negative individuals are completely resistant to P.
vivax malaria. Plasmodium falciparum binds sialic acid
residues on glycophorin A as well as alternative
receptors to invade human erythrocytes by multiple
pathways [3�/10]. Like P. vivax , the simian malaria
parasite Plasmodium knowlesi is also absolutely depen-
dent on interaction with the Duffy antigen to invade
human erythrocytes [11]. However, P. knowlesi , like P.
falciparum , uses multiple pathways to invade rhesus
erythrocytes [12]. P. knowlesi can bind the rhesus Duffy
antigen as well as alternative receptors on rhesus
erythrocytes for invasion [12].
Parasite ligands that mediate interaction of P. vivax ,
P. falciparum and P. knowlesi with erythrocyte recep-
Abbreviations: BSA, bovine serum albumin; CD, circular
dichroism; DBL, Duffy-binding-like; EBA-175, 175 kDa P.
falciparum erythrocyte binding antigen; FCS, fetal calf serum; GP67,
67 kDa envelope surface glycoprotein of Autographa californica
nuclear polyhedrosis virus; HSV gD, Herpes simplex virus
glycoprotein D; NTA, nitrilotriacetic acid; OD, optical density; PBS,
phosphate buffered saline; PCR, polymerase chain reaction; PkaRII,
region II of P. knowlesi a protein; PkbRII, region II of P. knowlesi bprotein; PkgRII, region II of P. knowlesi g protein; SDS-PAGE,
sodium dodecyl sulfate-polyacrylamide gel electrohporesis; Sf21,
Spodoptera frugiperda cells.
* Corresponding author. Tel.: �91-11-618-7695; fax: �91-11-616-
2316.
E-mail address: [email protected] (C.E. Chitnis).
Molecular & Biochemical Parasitology 121 (2002) 21�/31
www.parasitology-online.com
0166-6851/02/$ - see front matter # 2002 Elsevier Science B.V. All rights reserved.
PII: S 0 1 6 6 - 6 8 5 1 ( 0 2 ) 0 0 0 1 7 - 8

tors described above share similar features and belong
to a family of erythrocyte binding proteins [13]. The
erythrocyte binding protein family includes the 175 kDa
P. falciparum erythrocyte binding antigen (EBA-175),which binds sialic acid residues on glycophorin A, P.
vivax Duffy binding protein, and three related P.
knowlesi proteins referred to as a, b and g [12�/18].
The extracellular regions of each of these proteins
contain two conserved cysteine-rich domains that are
referred to as regions II and VI [13]. The functional
receptor-binding domain of each member of the ery-
throcyte binding protein family lies in the N-terminal,conserved cysteine-rich region, region II [19�/21]. These
conserved, cysteine-rich regions are referred to as Duffy-
binding-like (DBL) domains after the first binding
domain identified from P. vivax Duffy binding protein
[19]. DBL domains of the erythrocyte binding proteins
have diverse binding specificity. Whereas region II of P.
vivax Duffy binding protein binds the human Duffy
antigen, region II of P. knowlesi a protein (PkaRII)binds both human and rhesus Duffy antigens [19,21].
Region II of P. knowlesi b protein (PkbRII) binds sialic
acid residues on rhesus erythrocytes and region II of P.
knowlesi g protein (PkgRII) binds as yet unidentified
Duffy-independent receptors on rhesus erythrocytes
[19,21]. P. knowlesi b and g proteins may be responsible
for the Duffy-independent pathways used by P. knowlesi
to invade rhesus erythrocytes. Region II of P. falci-
parum EBA-175 contains tandem duplication of DBL
domains referred to as F1 and F2 [17]. The receptor
binding domain of EBA-175 lies in region F2 [20]. Genes
encoding paralogs of EBA-175 are present in the P.
falciparum genome. Such EBA-175 paralogs may bind
sialic acid�/glycophorin A independent receptors to
mediate invasion by alternative pathways [22�/25].
The binding domains of malaria parasite proteins thatbind erythrocyte receptors to mediate invasion are
attractive candidates for development of blood-stage
malaria vaccines. Antibodies directed against such
receptor-binding domains may block erythrocyte bind-
ing and invasion by merozoites. Indeed, antibodies
raised against the recombinant sialic acid binding
domain, region II, of EBA-175 expressed as a secreted
protein in insect cells have been shown to blockerythrocyte binding and invasion by P. falciparum
[26�/28]. The binding domain of P. vivax Duffy binding
protein has also been produced in its functional form
[29,30] and antibodies raised against this functional
domain have been shown to block erythrocyte binding
[30].
Here, we have expressed the binding domain of P.
knowlesi a protein, PkaRII, which binds the Duffyblood group antigen, as a secreted protein in insect cells.
Rabbit antibodies raised against recombinant PkaRII
purified from insect cell culture supernatants were tested
for inhibition of erythrocyte binding and invasion by P.
knowlesi . Antibodies raised against PkaRII inhibit
invasion of both human and rhesus erythrocytes by P.
knowlesi . These data suggest that immunization with the
homologous binding domains of P. vivax Duffy bindingprotein and P. falciparum EBA-175 may elicit antibo-
dies that inhibit multiplication of blood-stage P. vivax
and P. falciparum .
2. Materials and methods
2.1. Plasmid construct for expression of recombinant
PkaRII in insect cells
DNA fragments encoding PkaRII, the binding do-
main of P. knowlesi a protein (amino acids 202�/536),
fused to hexa-histidine at the carboxyl end were
amplified by polymerase chain reaction (PCR) and
cloned downstream of the signal sequence of the 67
kDa envelope surface glycoprotein (GP67) and polyhe-
drin promoter of Autographa californica nuclear poly-hedrosis virus in transfer vector pAcGP67B
(Pharmingen) as follows. Primers APM1 (5? CGC
GGA TCC AAT CA AAC TTT TCT TCA A 3?) and
APM4 (5?GAC GAA TTC TTT GTT ATA TTG GTA
GT-3?) were used for PCR with plasmid p1cg [16], which
contains the 5? end of gene encoding P. knowlesi aprotein, as template to amplify a 288 bp DNA fragment
(PCR product 1) encoding the N-terminal region ofPkaRII by PCR. Primers APM3 (5? TGC GAA TTC
TGT AAG GAT AT AAG ATG 3?) and APM2 (5?ATA GTT TAG CGG CCG CTC AGA GAT GAT
GAT GAT GAT GT TCA GTT ATC GGA TTA GA
3?) were used for PCR with plasmid p6D [16], which
contains the 3? end of gene encoding P. knowlesi aprotein, as template to amplify a 764 bp DNA fragment
(PCR product 2) encoding the C-terminal region ofPkaRII fused to a 6-histidine tag. PCR products 1 and 2
were digested with BamHI, EcoRI and EcoRI, NotI
respectively. The transfer vector pAcGP67B (Pharmin-
gen) was digested with BamHI, NotI and used in a
three-way ligation reaction with the two digested PCR
products to yield expression plasmid
pGP67BKADR2.1. Sequencing using an ABI 310 auto-
mated sequencer (Applied Biosystems) was used toconfirm that the DNA fragment encoding PkaRII in
pGP67BKADR2.1 has the correct orientation and no
mutations have been introduced by PCR. Recombinant
transfer plasmid, pGP67BKADR2.1, which contains a
DNA fragment encoding PkaRII as a BamHI�/NotI
fragment fused to the signal sequence of GP67 protein
downstream of the polyhedrin promoter, was used to
obtain recombinant baculovirus for expression of re-combinant PkaRII as a secreted protein in Spodoptera
frugiperda cells (Sf21 insect cells, Pharmingen) as
described below.
A.P. Singh et al. / Molecular & Biochemical Parasitology 121 (2002) 21�/3122

2.2. Cotransfection for construction of recombinant virus
for expression of secreted, recombinant PkaRII in insect
cells
Sf21 insect cells were co-transfected with
pGP67BKADR2.1 and Baculogold baculoviral genomic
DNA (Pharmingen) as recommended by the manufac-
turer. Isolation of recombinant plaques, plaque assays
and amplification to obtain high titre recombinant virus
(BVGP67PKADR2.1) were performed using standard
procedures.
2.3. Expression and purification of recombinant PkaRII
Sf21 insect cells (4�/106 cells ml�1) adapted forgrowth in suspension cultures in Sf900-II serum-free
medium (GIBCO) were infected with recombinant virus
BVGP67KADR2.1 at multiplicity of infection of 5 and
cultured for 96 h at 27 8C in suspension culture. Culture
supernatant was collected after 96 h, centrifuged to
remove cell debris, dialyzed extensively against dialysis
buffer (50 mM sodium phosphate, 300 mM NaCl, pH
8.0) at 4 8C, filtered through 0.45 mm filter (Millipore)and loaded onto a column containing Ni-nitrilotriacetic
acid (Ni-NTA) matrix (Qiagen) pre-equilibrated with
dialysis buffer. The Ni-NTA column was washed with
20 column volumes of dialysis buffer containing 10 mM
imidazole and 3 column volumes of 50 mM sodium
acetate pH 4.6. Recombinant PkaRII was eluted with
elution buffer containing 50 mM sodium phosphate pH
8.0, 300 mM NaCl, 250 mM imidazole, 10% glycerol.
2.4. Analysis of purified PkaRII by reverse phase
chromatography
PkaRII purified by Ni-NTA chromatography as
described above was loaded on a reverse phase C8
column. The gradient used for elution was developed
using Buffer A (0.05% trifluoroacetic acid in water) and
Buffer B (0.05% trifluoroacetic acid in 90% acetonitrile,
10% water). The column was initially equilibrated with90% Buffer A and 10% Buffer B and reached a
composition of 10% Buffer A and 90% Buffer B in 40
min.
2.5. Circular dichroism (CD) spectroscopy of
recombinant PkaRII
CD spectra were recorded on a Jasco-J720 spectro-
polarimeter. Spectra of purified PkaRII in 10 mM
phosphate buffer, pH 7.0 were recorded in the far-UV
region from 184 to 260 nm using a cuvette with a pathlength of 0.1 cm and the following instrument para-
meters-instrument sensitivity, 1 mdeg; response time, 2
s; scan speed, 50 nm min�1. Deconvolution of the CD
spectra was performed using the method of Bohm et al.
[31].
2.6. Erythrocyte binding assay with recombinant PkaRII
Blood collected in 10% citrate phosphate dextrose was
stored at 4 8C for up to 4 weeks and washed three times
in RPMI 1640 culture medium (GIBCO) before use.
Duffy phenotypes of erythrocytes were determined by
standard blood typing methods using two antisera (anti-
Fya and anti-Fyb) (Ortho-Clinical Diagnostics). Duffy
positive human erythrocytes and rhesus erythrocytes
were treated with chymotrypsin as described earlier [14].Recombinant PkaRII (10 mg) was incubated for 1 h at
room temperature with normal and chymotrypsin-
treated erythrocytes (100 ml packed erythrocytes) in a
total volume of 600 ml containing 10% fetal calf serum
(FCS) (GIBCO). The reaction mixture was layered over
dibutylpthalate (Sigma) and centrifuged to collect ery-
throcytes. Bound protein was eluted from the erythro-
cytes with 300 mM NaCl. One-third of the elute wasseparated by SDS-PAGE and detected by Western
blotting using a mouse monoclonal antibody raised
against penta-histidine (Qiagen). Purified PkaRII (1 mg)
was used as a positive control for Western blotting.
2.7. Animals and immunization
Rabbits used in this study were procured from theAnimal Facility of the International Center for Genetic
Engineering and Biotechnology, New Delhi, India.
Animals were housed, fed, and used in experiments
according to the guidelines set forth in the National
Institutes of Health manual titled, Guide for the Care
And Use of Laboratory Animals (National Institutes of
Health Publication No. 86-23, US Department of
Health and Human Services, Washington DC). Two16-week-old New Zealand White rabbits were immu-
nized with 400 mg of recombinant PkaRII emulsified in
complete Freund’s adjuvant delivered by the subcuta-
neous route. The rabbits were boosted thrice with 200 mg
of PkaRII formulated in incomplete Freund’s adjuvant
delivered by the subcutaneous route 4, 8 and 12 weeks
after primary immunization. One rabbit was immunized
with adjuvant alone according to the schedule describedabove to provide control serum. Rabbits were bled
before immunization (pre-immune sera) and 2 weeks
after each boost.
2.8. ELISA
Rabbit sera were tested for recognition of recombi-
nant PkaRII by ELISA. Serial dilutions of sera startingwith 1:1000 fold dilution were tested. Briefly, wells of
flat-bottom Immulon-2 plates (Dynatech Laboratories)
were coated with 0.1 mg of PkaRII and blocked with 3%
A.P. Singh et al. / Molecular & Biochemical Parasitology 121 (2002) 21�/31 23

BSA in PBS (blocking buffer). Antigen-coated wells
were incubated for 90 min at 37 8C with 100 ml of rabbit
serum diluted in 0.1% blocking buffer. Wells were
washed with phosphate-buffered saline (PBS) and0.05% Tween 20 (wash buffer). A total of 100 ml of
horseradish peroxidase-labeled anti-rabbit IgG antibody
(Sigma) diluted 1:2000 fold was then added to each well,
incubated for 60 min at 37 8C and washed three times
with wash buffer prior to development of enzyme
reaction with o-phenylenediamine dihydrochloride as
the chromogen and hydrogen peroxide as the substrate.
The reaction was terminated by addition of sulfuric acidand the absorbance at 490 nm (OD490 nm) was recorded
in each well using an ELISA microplate reader (Mole-
cular Devices). Pre-immune sera as well as control sera
raised against adjuvant alone were used at similar
dilutions.
2.9. Immunoflourescence assay
Rabbit serum raised against recombinant PkaRIIwere tested for recognition of PkaRII, PkbRII and
PkgRII expressed on surface of mammalian COS7 cells
by immunofluorescence assay. Plasmid DNA constructs
pHKADR22, pHKBDR22 and pHKGDR22, which are
designed to express PkaRII, PkbRII and PkgRII on the
surface of COS7 cells, have been described earlier [19].
Briefly, plasmids pHKADR22, pHKBDR22 and
pHKGDR22 contain DNA encoding PkaRII, PkbRIIand PkgRII respectively fused to DNA encoding the
signal sequence of Herpes simplex virus glycoprotein D
(HSV gD) at the 5? end and DNA encoding the
transmembrane segment and cytoplasmic domain
HSVgD at the 3? end cloned in a mammalian cell
expression vector [19,32]. COS7 cells were transfected
with plasmids pHKADR22, pHKBDR22 and
pHKGDR22 using Lipofectin (GIBCO) as describedby the manufacturer. Expression of PkaRII, PkbRII
and PkgRII on the surface of COS7 cells was confirmed
by immunofluorescence assay using murine monoclonal
antibody DL6, which is directed against HSV gD
sequences in the recombinant fusion proteins, as de-
scribed earlier [19,32]. Serial dilutions of rabbit serum
raised against recombinant PkaRII were used in im-
munofluorescence assays with transfected COS7 cellsexpressing PkaRII, PkbRII and PkgRII on the surface
to determine end point titres.
2.10. Inhibition of erythrocyte binding with rabbit serum
raised against PkaRII
Transfected COS7 cells expressing PkaRII, PkbRII
and PkgRII on the surface were used in erythrocytebinding assays with human and rhesus erythrocytes as
described earlier [19,30]. Erythrocyte binding assays
were performed in the presence of different dilutions
of rabbit serum raised against recombinant PkaRII as
described earlier [30]. Pre-immune rabbit serum and
rabbit serum raised against Freund’s adjuvant alone
were used as control. The number of COS7 cells havingrosettes of bound erythrocytes was scored in 30 fields at
200 fold magnification. Each dilution of serum was
tested in triplicate. The average percent inhibition of
binding (and S.D.) was determined at each dilution.
2.11. Purification of antibodies from rabbit serum
Rabbit antibodies (IgG) were purified using Protein
A-Sepharose (Pharmacia) as recommended by the
manufacturer. Briefly, 5.0 ml Protein A slurry was
loaded in a column, washed with 10 ml PBS and
equilibrated with 10 ml 100 mM Tris pH 8.0. Rabbitserum (4.5 ml) was mixed with 0.5 ml 1 M Tris pH 8.0
and loaded on the Protein A column using gravity flow.
The column was washed with 35 ml 100 mM Tris pH 8.0
and 20 ml 10 mM Tris pH 8.0. Bound IgG was eluted
with 100 mM Glycine pH 3.0. Elutes (0.5 ml each) were
collected in 1.5 ml tubes containing 50 ml 1 M Tris pH
8.0. Purified IgG was separated by SDS-PAGE and
detected by Coomasie-staining. Proteins of expected size(25 and 55 kDa) were detected and the purity of IgG was
�/99%. Purified IgG was dialyzed against RPMI 1640
(GIBCO) prior to use in invasion assays. Concentration
of antibodies was determined by measuring OD280 nm.
2.12. Inhibition of erythrocyte invasion by P. knowlesi
with purified rabbit antibodies raised against PkaRII
Parasitized blood was collected from rhesus monkeys
infected with P. knowlesi H strain by venipuncture when
parasitemia was �/5% and majority of parasites were in
the schizont stage. P. knowlesi schizonts were isolated to�/95% purity by centrifugation of infected blood on
45% Percoll. Purified P. knowlesi schizonts were used
for in vitro erythrocyte invasion assays as described
earlier [33]. Briefly, human or rhesus erythrocytes (1�/
107) were incubated with P. knowlesi schizonts (1�/106)
in the presence of 10% FCS and different dilutions of
purified rabbit antibodies raised against PkaRII or
Freund’s adjuvant alone for 8�/10 h at 37 8C under90% N2, 5% O2, 5% CO2. Different concentrations of
antibodies were tested (2.7, 0.9, 0.3 and 0.1 mg ml�1).
Two thousand erythrocytes were scored for presence of
P. knowlesi rings on Giemsa-stained smears. The
percentage of erythrocytes infected with rings was
calculated to determine invasion rates. Invasion inhibi-
tion efficiencies were determined using invasion rates in
presence of different concentrations (conc a) of anti-PkaRII antibodies (Inv(PkaRII)conc a) and antibodies
raised against adjuvant alone (Inv(Adj.)conc a) as fol-
lows:
A.P. Singh et al. / Molecular & Biochemical Parasitology 121 (2002) 21�/3124
Inhibition efficiency (%)
�(1�Inv(PkaRII)conc a=Inv(Adj:)conc a)� 100
Invasion assays were performed in triplicate at eachantibody concentration. Average invasion rates and
S.D. are reported. P values were calculated by one-
way ANOVA and non-parametric test for invasion
rates. P values greater than 0.05 were considered not
significant.
3. Results
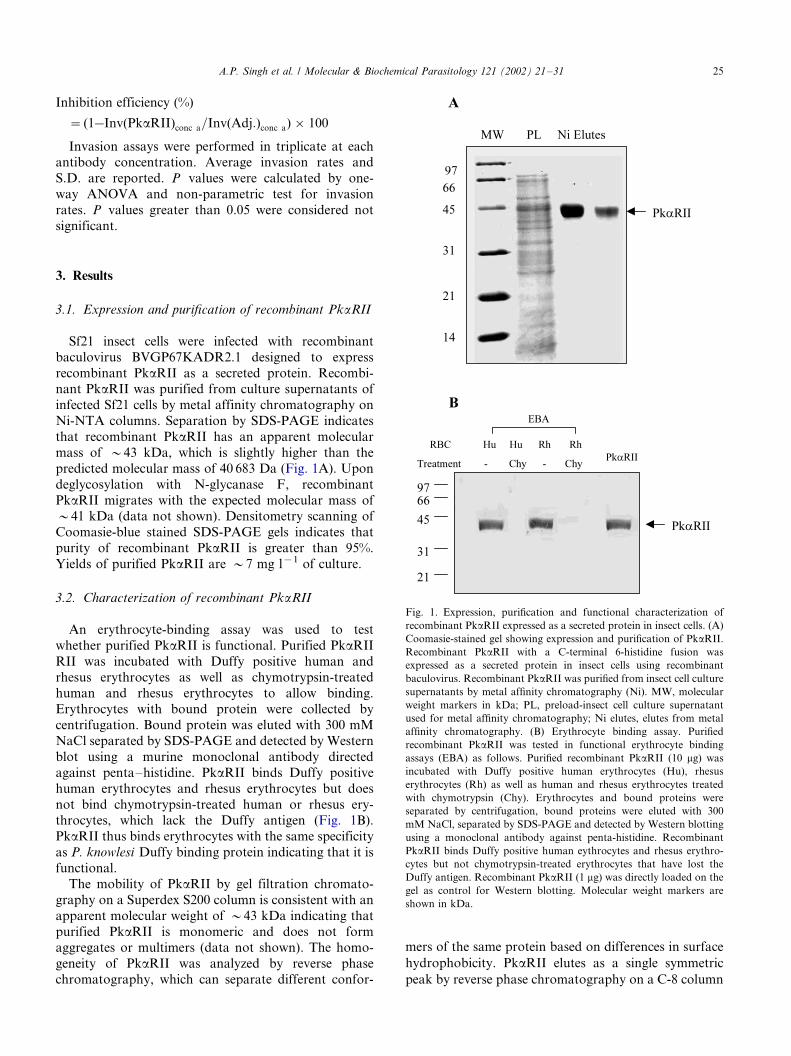
3.1. Expression and purification of recombinant PkaRII
Sf21 insect cells were infected with recombinant
baculovirus BVGP67KADR2.1 designed to express
recombinant PkaRII as a secreted protein. Recombi-nant PkaRII was purified from culture supernatants of
infected Sf21 cells by metal affinity chromatography on
Ni-NTA columns. Separation by SDS-PAGE indicates
that recombinant PkaRII has an apparent molecular
mass of �/43 kDa, which is slightly higher than the
predicted molecular mass of 40 683 Da (Fig. 1A). Upon
deglycosylation with N-glycanase F, recombinant
PkaRII migrates with the expected molecular mass of�/41 kDa (data not shown). Densitometry scanning of
Coomasie-blue stained SDS-PAGE gels indicates that
purity of recombinant PkaRII is greater than 95%.
Yields of purified PkaRII are �/7 mg l�1 of culture.
3.2. Characterization of recombinant PkaRII
An erythrocyte-binding assay was used to test
whether purified PkaRII is functional. Purified PkaRII
RII was incubated with Duffy positive human and
rhesus erythrocytes as well as chymotrypsin-treatedhuman and rhesus erythrocytes to allow binding.
Erythrocytes with bound protein were collected by
centrifugation. Bound protein was eluted with 300 mM
NaCl separated by SDS-PAGE and detected by Western
blot using a murine monoclonal antibody directed
against penta�/histidine. PkaRII binds Duffy positive
human erythrocytes and rhesus erythrocytes but does
not bind chymotrypsin-treated human or rhesus ery-throcytes, which lack the Duffy antigen (Fig. 1B).
PkaRII thus binds erythrocytes with the same specificity
as P. knowlesi Duffy binding protein indicating that it is
functional.
The mobility of PkaRII by gel filtration chromato-
graphy on a Superdex S200 column is consistent with an
apparent molecular weight of �/43 kDa indicating that
purified PkaRII is monomeric and does not formaggregates or multimers (data not shown). The homo-
geneity of PkaRII was analyzed by reverse phase
chromatography, which can separate different confor-
mers of the same protein based on differences in surface
hydrophobicity. PkaRII elutes as a single symmetric
peak by reverse phase chromatography on a C-8 column
Fig. 1. Expression, purification and functional characterization of
recombinant PkaRII expressed as a secreted protein in insect cells. (A)
Coomasie-stained gel showing expression and purification of PkaRII.
Recombinant PkaRII with a C-terminal 6-histidine fusion was
expressed as a secreted protein in insect cells using recombinant
baculovirus. Recombinant PkaRII was purified from insect cell culture
supernatants by metal affinity chromatography (Ni). MW, molecular
weight markers in kDa; PL, preload-insect cell culture supernatant
used for metal affinity chromatography; Ni elutes, elutes from metal
affinity chromatography. (B) Erythrocyte binding assay. Purified
recombinant PkaRII was tested in functional erythrocyte binding
assays (EBA) as follows. Purified recombinant PkaRII (10 mg) was
incubated with Duffy positive human erythrocytes (Hu), rhesus
erythrocytes (Rh) as well as human and rhesus erythrocytes treated
with chymotrypsin (Chy). Erythrocytes and bound proteins were
separated by centrifugation, bound proteins were eluted with 300
mM NaCl, separated by SDS-PAGE and detected by Western blotting
using a monoclonal antibody against penta-histidine. Recombinant
PkaRII binds Duffy positive human eythrocytes and rhesus erythro-
cytes but not chymotrypsin-treated erythrocytes that have lost the
Duffy antigen. Recombinant PkaRII (1 mg) was directly loaded on the
gel as control for Western blotting. Molecular weight markers are
shown in kDa.
A.P. Singh et al. / Molecular & Biochemical Parasitology 121 (2002) 21�/31 25
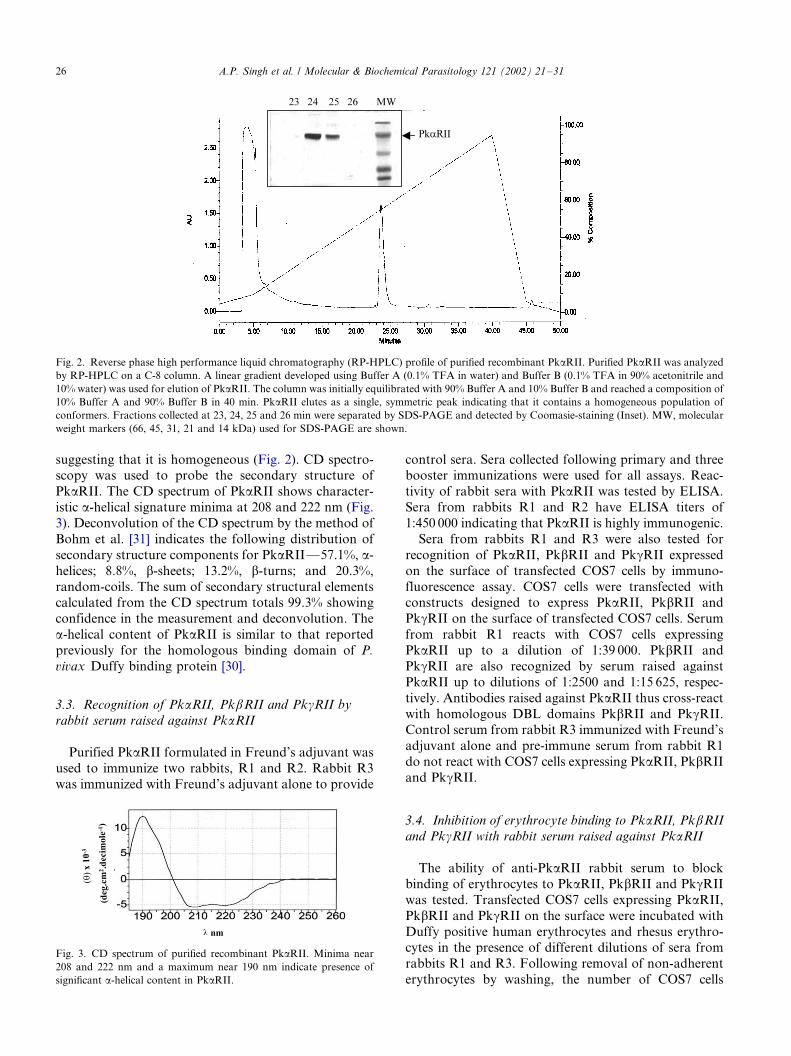
suggesting that it is homogeneous (Fig. 2). CD spectro-scopy was used to probe the secondary structure of
PkaRII. The CD spectrum of PkaRII shows character-
istic a-helical signature minima at 208 and 222 nm (Fig.
3). Deconvolution of the CD spectrum by the method of
Bohm et al. [31] indicates the following distribution of
secondary structure components for PkaRII*/57.1%, a-
helices; 8.8%, b-sheets; 13.2%, b-turns; and 20.3%,
random-coils. The sum of secondary structural elementscalculated from the CD spectrum totals 99.3% showing
confidence in the measurement and deconvolution. The
a-helical content of PkaRII is similar to that reported
previously for the homologous binding domain of P.
vivax Duffy binding protein [30].
3.3. Recognition of PkaRII, PkbRII and PkgRII by
rabbit serum raised against PkaRII
Purified PkaRII formulated in Freund’s adjuvant was
used to immunize two rabbits, R1 and R2. Rabbit R3
was immunized with Freund’s adjuvant alone to provide
control sera. Sera collected following primary and threebooster immunizations were used for all assays. Reac-
tivity of rabbit sera with PkaRII was tested by ELISA.
Sera from rabbits R1 and R2 have ELISA titers of
1:450 000 indicating that PkaRII is highly immunogenic.
Sera from rabbits R1 and R3 were also tested for
recognition of PkaRII, PkbRII and PkgRII expressed
on the surface of transfected COS7 cells by immuno-
fluorescence assay. COS7 cells were transfected withconstructs designed to express PkaRII, PkbRII and
PkgRII on the surface of transfected COS7 cells. Serum
from rabbit R1 reacts with COS7 cells expressing
PkaRII up to a dilution of 1:39 000. PkbRII and
PkgRII are also recognized by serum raised against
PkaRII up to dilutions of 1:2500 and 1:15 625, respec-
tively. Antibodies raised against PkaRII thus cross-react
with homologous DBL domains PkbRII and PkgRII.Control serum from rabbit R3 immunized with Freund’s
adjuvant alone and pre-immune serum from rabbit R1
do not react with COS7 cells expressing PkaRII, PkbRII
and PkgRII.
3.4. Inhibition of erythrocyte binding to PkaRII, PkbRII
and PkgRII with rabbit serum raised against PkaRII
The ability of anti-PkaRII rabbit serum to block
binding of erythrocytes to PkaRII, PkbRII and PkgRII
was tested. Transfected COS7 cells expressing PkaRII,
PkbRII and PkgRII on the surface were incubated with
Duffy positive human erythrocytes and rhesus erythro-cytes in the presence of different dilutions of sera from
rabbits R1 and R3. Following removal of non-adherent
erythrocytes by washing, the number of COS7 cells
Fig. 2. Reverse phase high performance liquid chromatography (RP-HPLC) profile of purified recombinant PkaRII. Purified PkaRII was analyzed
by RP-HPLC on a C-8 column. A linear gradient developed using Buffer A (0.1% TFA in water) and Buffer B (0.1% TFA in 90% acetonitrile and
10% water) was used for elution of PkaRII. The column was initially equilibrated with 90% Buffer A and 10% Buffer B and reached a composition of
10% Buffer A and 90% Buffer B in 40 min. PkaRII elutes as a single, symmetric peak indicating that it contains a homogeneous population of
conformers. Fractions collected at 23, 24, 25 and 26 min were separated by SDS-PAGE and detected by Coomasie-staining (Inset). MW, molecular
weight markers (66, 45, 31, 21 and 14 kDa) used for SDS-PAGE are shown.
Fig. 3. CD spectrum of purified recombinant PkaRII. Minima near
208 and 222 nm and a maximum near 190 nm indicate presence of
significant a-helical content in PkaRII.
A.P. Singh et al. / Molecular & Biochemical Parasitology 121 (2002) 21�/3126
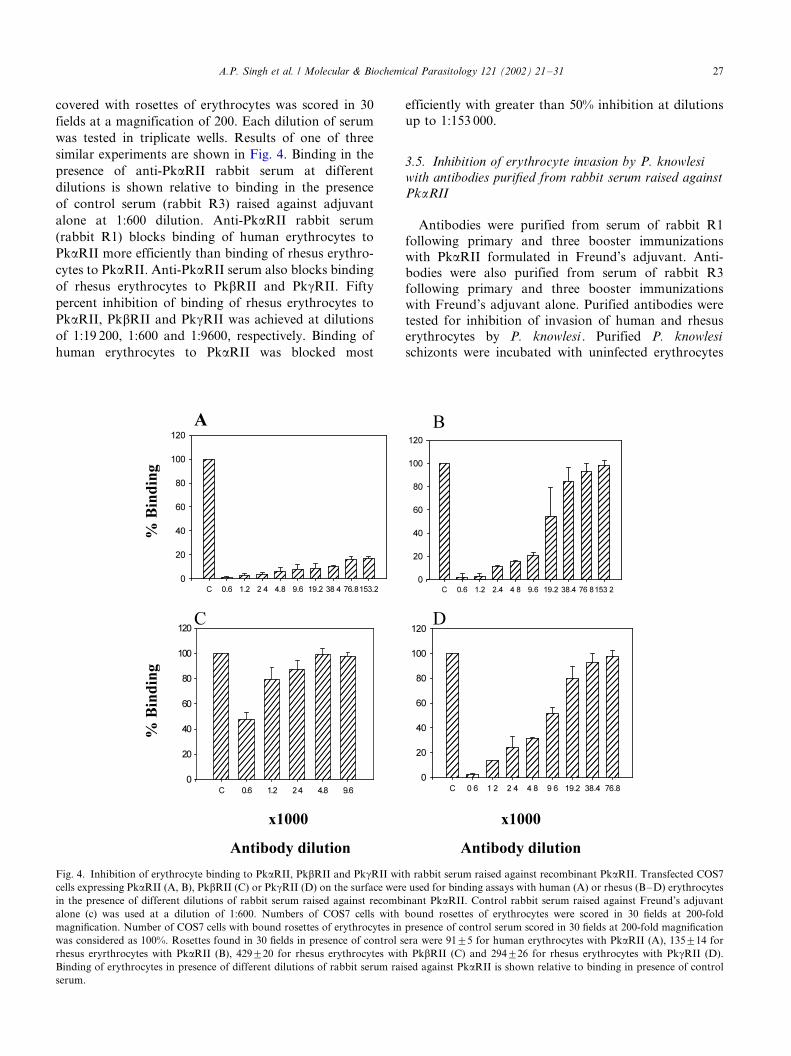
covered with rosettes of erythrocytes was scored in 30
fields at a magnification of 200. Each dilution of serum
was tested in triplicate wells. Results of one of three
similar experiments are shown in Fig. 4. Binding in the
presence of anti-PkaRII rabbit serum at different
dilutions is shown relative to binding in the presence
of control serum (rabbit R3) raised against adjuvant
alone at 1:600 dilution. Anti-PkaRII rabbit serum
(rabbit R1) blocks binding of human erythrocytes to
PkaRII more efficiently than binding of rhesus erythro-
cytes to PkaRII. Anti-PkaRII serum also blocks binding
of rhesus erythrocytes to PkbRII and PkgRII. Fifty
percent inhibition of binding of rhesus erythrocytes to
PkaRII, PkbRII and PkgRII was achieved at dilutions
of 1:19 200, 1:600 and 1:9600, respectively. Binding of
human erythrocytes to PkaRII was blocked most
efficiently with greater than 50% inhibition at dilutions
up to 1:153 000.
3.5. Inhibition of erythrocyte invasion by P. knowlesi
with antibodies purified from rabbit serum raised against
PkaRII
Antibodies were purified from serum of rabbit R1
following primary and three booster immunizations
with PkaRII formulated in Freund’s adjuvant. Anti-
bodies were also purified from serum of rabbit R3
following primary and three booster immunizations
with Freund’s adjuvant alone. Purified antibodies were
tested for inhibition of invasion of human and rhesuserythrocytes by P. knowlesi . Purified P. knowlesi
schizonts were incubated with uninfected erythrocytes
Fig. 4. Inhibition of erythrocyte binding to PkaRII, PkbRII and PkgRII with rabbit serum raised against recombinant PkaRII. Transfected COS7
cells expressing PkaRII (A, B), PkbRII (C) or PkgRII (D) on the surface were used for binding assays with human (A) or rhesus (B�/D) erythrocytes
in the presence of different dilutions of rabbit serum raised against recombinant PkaRII. Control rabbit serum raised against Freund’s adjuvant
alone (c) was used at a dilution of 1:600. Numbers of COS7 cells with bound rosettes of erythrocytes were scored in 30 fields at 200-fold
magnification. Number of COS7 cells with bound rosettes of erythrocytes in presence of control serum scored in 30 fields at 200-fold magnification
was considered as 100%. Rosettes found in 30 fields in presence of control sera were 9195 for human erythrocytes with PkaRII (A), 135914 for
rhesus eryrthrocytes with PkaRII (B), 429920 for rhesus erythrocytes with PkbRII (C) and 294926 for rhesus erythrocytes with PkgRII (D).
Binding of erythrocytes in presence of different dilutions of rabbit serum raised against PkaRII is shown relative to binding in presence of control
serum.
A.P. Singh et al. / Molecular & Biochemical Parasitology 121 (2002) 21�/31 27
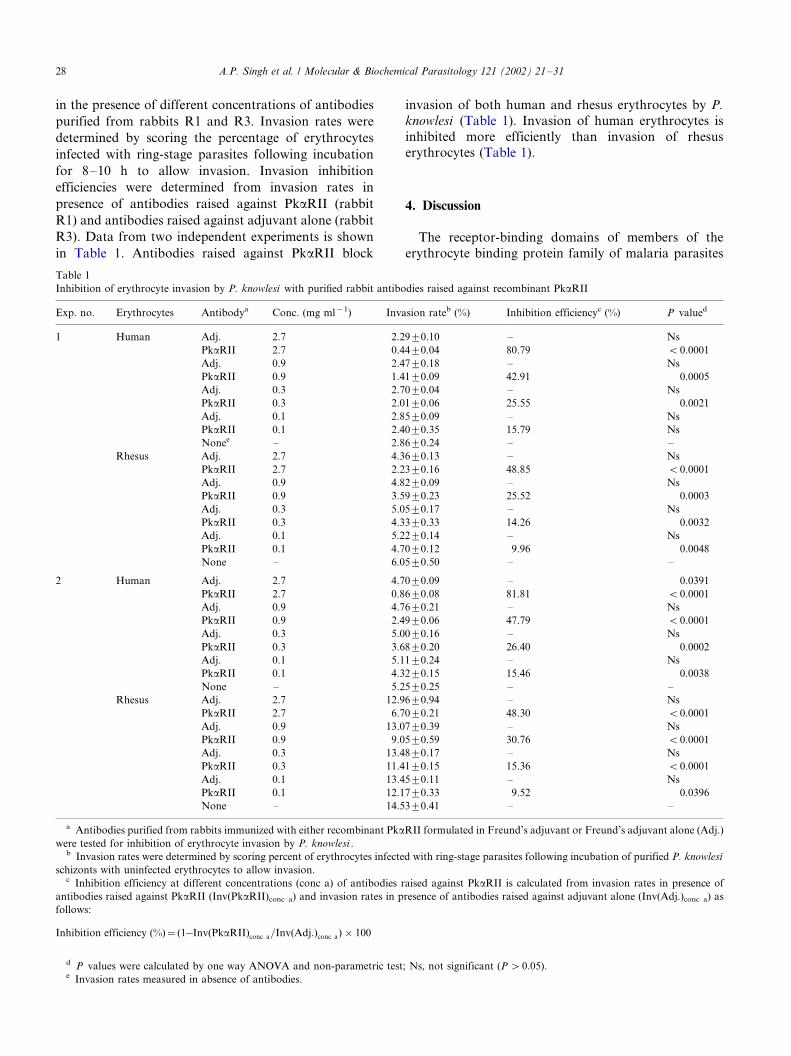
in the presence of different concentrations of antibodies
purified from rabbits R1 and R3. Invasion rates were
determined by scoring the percentage of erythrocytes
infected with ring-stage parasites following incubation
for 8�/10 h to allow invasion. Invasion inhibition
efficiencies were determined from invasion rates in
presence of antibodies raised against PkaRII (rabbit
R1) and antibodies raised against adjuvant alone (rabbit
R3). Data from two independent experiments is shown
in Table 1. Antibodies raised against PkaRII block
invasion of both human and rhesus erythrocytes by P.
knowlesi (Table 1). Invasion of human erythrocytes is
inhibited more efficiently than invasion of rhesus
erythrocytes (Table 1).
4. Discussion
The receptor-binding domains of members of the
erythrocyte binding protein family of malaria parasites
Table 1
Inhibition of erythrocyte invasion by P. knowlesi with purified rabbit antibodies raised against recombinant PkaRII
Exp. no. Erythrocytes Antibodya Conc. (mg ml�1) Invasion rateb (%) Inhibition efficiencyc (%) P valued
1 Human Adj. 2.7 2.2990.10 �/ Ns
PkaRII 2.7 0.4490.04 80.79 B0.0001
Adj. 0.9 2.4790.18 �/ Ns
PkaRII 0.9 1.4190.09 42.91 0.0005
Adj. 0.3 2.7090.04 �/ Ns
PkaRII 0.3 2.0190.06 25.55 0.0021
Adj. 0.1 2.8590.09 �/ Ns
PkaRII 0.1 2.4090.35 15.79 Ns
Nonee �/ 2.8690.24 �/ �/
Rhesus Adj. 2.7 4.3690.13 �/ Ns
PkaRII 2.7 2.2390.16 48.85 B0.0001
Adj. 0.9 4.8290.09 �/ Ns
PkaRII 0.9 3.5990.23 25.52 0.0003
Adj. 0.3 5.0590.17 �/ Ns
PkaRII 0.3 4.3390.33 14.26 0.0032
Adj. 0.1 5.2290.14 �/ Ns
PkaRII 0.1 4.7090.12 9.96 0.0048
None �/ 6.0590.50 �/ �/
2 Human Adj. 2.7 4.7090.09 �/ 0.0391
PkaRII 2.7 0.8690.08 81.81 B0.0001
Adj. 0.9 4.7690.21 �/ Ns
PkaRII 0.9 2.4990.06 47.79 B0.0001
Adj. 0.3 5.0090.16 �/ Ns
PkaRII 0.3 3.6890.20 26.40 0.0002
Adj. 0.1 5.1190.24 �/ Ns
PkaRII 0.1 4.3290.15 15.46 0.0038
None �/ 5.2590.25 �/ �/
Rhesus Adj. 2.7 12.9690.94 �/ Ns
PkaRII 2.7 6.7090.21 48.30 B0.0001
Adj. 0.9 13.0790.39 �/ Ns
PkaRII 0.9 9.0590.59 30.76 B0.0001
Adj. 0.3 13.4890.17 �/ Ns
PkaRII 0.3 11.4190.15 15.36 B0.0001
Adj. 0.1 13.4590.11 �/ Ns
PkaRII 0.1 12.1790.33 9.52 0.0396
None �/ 14.5390.41 �/ �/
a Antibodies purified from rabbits immunized with either recombinant PkaRII formulated in Freund’s adjuvant or Freund’s adjuvant alone (Adj.)
were tested for inhibition of erythrocyte invasion by P. knowlesi .b Invasion rates were determined by scoring percent of erythrocytes infected with ring-stage parasites following incubation of purified P. knowlesi
schizonts with uninfected erythrocytes to allow invasion.c Inhibition efficiency at different concentrations (conc a) of antibodies raised against PkaRII is calculated from invasion rates in presence of
antibodies raised against PkaRII (Inv(PkaRII)conc a) and invasion rates in presence of antibodies raised against adjuvant alone (Inv(Adj.)conc a) as
follows:
Inhibition efficiency (%)�(1�Inv(PkaRII)conc a=Inv(Adj:)conc a)�100
d P values were calculated by one way ANOVA and non-parametric test; Ns, not significant (P �0.05).e Invasion rates measured in absence of antibodies.
A.P. Singh et al. / Molecular & Biochemical Parasitology 121 (2002) 21�/3128

lie in conserved N-terminal cysteine-rich regions referred
to as region II [19�/21]. Here, we have produced
recombinant PkaRII, region II of P. knowlesi a protein,
which binds the Duffy antigen on human and rhesuserythrocytes, as a secreted protein in insect cells.
Recombinant PkaRII was purified from culture super-
natants of insect cells by metal affinity chromatography
and characterized to show that it is pure, homogenous
and functional. Mobility of recombinant PkaRII on
SDS-PAGE gels suggests that it has an apparent
molecular mass of �/43 kDa, which is higher than the
predicted molecular mass of 40 683 Da. PkaRII containsthree putative N-glycosylation sites namely, NQT, NNT
and NES. Treatment with N-glycanase F results in
reduction of the apparent molecular mass to �/41 kDa
indicating that recombinant PkaRII is glycosylated. The
CD spectrum of recombinant PkaRII indicates presence
of significant a-helical content (57.1%), which is similar
to the a-helical content reported previously for the
homologous binding domain from P. vivax Duffybinding protein [30] and as predicted by sequence
analysis of DBL domains [34]. Importantly, recombi-
nant PkaRII is functional in that it binds normal Duffy
positive human erythrocytes and rhesus erythrocytes but
not chymotrypsin-treated human or rhesus erythrocytes
that have lost the Duffy antigen.
Recombinant PkaRII was used to immunize rabbits.
Antibodies raised against this functional receptor-bind-ing domain were tested for inhibition of erythrocyte
binding and invasion by P. knowlesi in vitro. Results of
these experiments are of direct relevance for vaccine
development efforts based on the homologous receptor-
binding domains of P. vivax Duffy binding protein and
P. falciparum EBA-175. Recombinant PkaRII is highly
immunogenic and elicits high titre antibodies that
recognize PkaRII expressed on surface of COS7 cells.Rabbit serum raised against PkaRII cross-reacts with
the homologous DBL domains PkbRII and PkgRII to
varying extents. Anti-PkaRII rabbit serum was tested
for inhibition of erythrocyte binding to PkaRII, PkbRII
and PkgRII expressed on the surface of COS7 cells.
Anti-PkaRII serum inhibits binding of both human and
rhesus erythrocytes to PkaRII. Binding of human
erythrocytes to PkaRII is inhibited more efficientlythan binding of rhesus erythrocytes. Anti-PkaRII serum
also inhibits binding of rhesus erythrocytes to PkbRII
and PkgRII although less efficiently than it inhibits
binding to PkaRII. Immunofluorescence titres of anti-
PkaRII serum for recognition of PkaRII, PkbRII and
PkgRII correlate with the efficiency of inhibition of
erythrocyte binding to these domains.
Antibodies raised against recombinant PkaRII havegreater cross-reactivity with PkgRII than PkbRII. Pair
wise comparison of amino acid sequences of PkaRII
with amino acid sequences of PkbRII and PkgRII
indicates that the percent sequence identity between
these DBL domains is similar, around 70%. One would
therefore not predict that antibodies raised against
PkaRII would have greater cross-reactivity with
PkgRII than PkbRII. The cross-reactivity data suggeststhat PkaRII shares some cross-reactive epitopes with
PkgRII that are not present in PkbRII. Sequence
analysis does reveal the presence of a number of
hydrophilic amino acid residues that are unique to
PkaRII and PkgRII. Such residues are likely to be
exposed and may be responsible for presence of cross-
reactive epitopes shared by PkaRII and PkgRII.
Antibodies purified from rabbit serum raised againstPkaRII were tested for inhibition of invasion of human
and rhesus erythrocytes by P. knowlesi in vitro. Anti-
bodies raised against PkaRII inhibit invasion of human
erythrocytes by around 80% at the highest concentration
tested (2.7 mg ml�1) in two independent experiments.
Like P. knowlesi , P. vivax is also completely dependent
on interaction with the Duffy antigen for invasion of
human erythrocytes. The observation that antibodiesdirected against PkaRII block invasion of human
erythrocytes by P. knowlesi efficiently suggests that
antibodies elicited by a recombinant vaccine based on
the binding domain of P. vivax Duffy binding protein
should block multiplication of blood-stage P. vivax
effectively.
Antibodies raised against PkaRII also inhibit inva-
sion of rhesus erythrocytes by P. knowlesi . However,inhibition of invasion of rhesus erythrocytes (around
48% at antibody concentration of 2.7 mg ml�1) is less
efficient than inhibition of invasion of human erythro-
cytes (around 80% at antibody concentration of 2.7 mg
ml�1). Anti-PkaRII serum blocks binding of rhesus
erythrocytes to PkaRII less efficiently than binding of
human erythrocytes. Moreover, while P. knowlesi is
completely dependent on the interaction of a proteinwith the Duffy antigen for invasion of human erythro-
cytes, it can use alternative pathways mediated by b and
g proteins to invade rhesus erythrocytes. Although
antibodies raised against PkaRII cross-react with
PkbRII and PkgRII, they inhibit binding of rhesus
erythrocytes to PkbRII and PkgRII less efficiently
than binding of human erythrocytes to PkaRII. The
availability of multiple pathways for invasion ofrhesus erythrocytes and the lower efficiency for inhibi-
tion of binding of rhesus erythrocytes to PkaRII,
PkbRII and PkgRII compared to human erythro
cytes may be responsible for the lower inhibition of
invasion of rhesus erythrocytes compared to human
erythrocytes.
Like P. knowlesi , P. falciparum also uses multiple
pathways to invade human erythrocytes [3�/10]. Para-logs of P. knowlesi a protein, namely b and g proteins,
mediate Duffy-independent alternative invasion path-
ways. Similarly, EBA-175 paralogs, which contain
receptor-binding DBL domains, may bind sialic acid�/
A.P. Singh et al. / Molecular & Biochemical Parasitology 121 (2002) 21�/31 29

glycophorin A independent receptors to mediate inva-
sion by alternative pathways [22�/25]. The observations
made here with P. knowlesi suggest that antibodies
raised against the binding domain of EBA-175 maycross-react with binding domains of EBA-175 paralogs
to inhibit invasion by alternative pathways, although the
inhibition may not be very efficient. Indeed, it has been
observed that antibodies raised against recombinant
region II of EBA-175 expressed as a secreted protein in
insect cells inhibit invasion by P. falciparum isolates that
use multiple invasion pathways [28]. The efficiencies
reported for inhibition of P. falciparum invasion byantibodies raised against recombinant EBA-175 region
II [28] are similar to the efficiencies described here for
inhibition of P. knowlesi invasion of rhesus erythrocytes
by antibodies raised against PkaRII.
In conclusion, antibodies raised against the receptor-
binding domain of P. knowlesi a protein, which binds
the Duffy antigen, block invasion of human erythro-
cytes by P. knowlesi efficiently. By analogy, antibodiesraised against the receptor-binding domain of P. vivax
Duffy binding protein should effectively block erythro-
cyte invasion by P. vivax . This provides support for the
development of a vaccine based on region II of P. vivax
Duffy binding protein. Antibodies raised against
PkaRII also inhibit P. knowlesi invasion of rhesus
erythrocytes although at reduced efficiency compared
to inhibition of invasion of human erythrocytes. Thisobservation suggests that in case of P. falciparum , which
like P. knowlesi uses multiple invasion pathways,
antibodies raised against the binding domain of EBA-
175 should block invasion of human erythrocytes,
although at reduced efficiency. It has been shown that
immunization of Aotus monkeys by a variety of prime-
boost combinations using EBA-175 region II DNA
immunization constructs as well as recombinant EBA-175 region II produced in insect cells results in reduction
of parasite densities and partial protection against
blood-stage P. falciparum challenge [35]. These data
support the inclusion of the binding domain of EBA-175
in a vaccine for P. falciparum malaria. However, it may
be necessary to combine EBA-175 with other parasite
proteins that play functional roles in invasion to block
erythrocyte invasion efficiently and provide significantprotection against P. falciparum malaria.
Acknowledgements
We thank Drs Gary Cohen and Roselyn Eisenberg for
providing plasmid pRE4 and monoclonal antibody
DL6, Amit Sharma and Virander S. Chauhan for
comments on the manuscript, and Andrew Lynn forCD spectroscopy. This investigation was supported by a
grant from the European Commission (IC18CT980369)
to SKP and CEC and an International Research
Scholar’s grant from the Howard Hughes Medical
Institute to CEC.
References
[1] Chitnis CE. Molecular insights into receptors used for erythrocyte
invasion by malaria parasites. Curr Op Hematol 2001;8:85�/91.
[2] Miller LH, Mason SJ, Clyde DF, McGinnis MH. The resistance
factor to Plasmodium vivax in Blacks: Duffy blood group
genotype, FyFy. N Engl J Med 1976;295:302�/4.
[3] Breuer WV, Ginsburg H, Cabantchik ZI. An assay of malaria
parasite invasion into human erythrocytes. The effects of chemical
and enzyme modification of erythrocyte membrane components.
Biochem Biophys Acta 1983;755:263�/71.
[4] Friedman MJ, Blankenburt T, Sensabaugh G, Tenforde TS.
Recognition and invasion of human erythrocytes by malarial
parasites: contribution of sialo-glycoproteins to attachment and
host specificity. J Cell Biol 1982;98:1682�/7.
[5] Miller LH, Haynes JO, McAuliffe FM, Shiroishi T, Durocher J,
McGinnis MH. Evidence for differences in erythrocyte surface
receptors for the malaria parasites, Plasmodium falciparum and P.
knowlesi . J Exp Med 1977;146:277�/81.
[6] Pasvol G, Wainscoat JS, Weatherall DJ. Erythrocytes deficient in
glycophorin resist invasion by the malarial parasite Plasmodium
falciparum . Nature 1982;297:64�/6.
[7] Perkins ME, Holt EH. Erythrocyte receptor recognition varies in
Plasmodium falciparum isolates. Mol Biochem Parasitol
1988;27:23�/34.
[8] Mitchell GH, Hadley TJ, McGinniss MH, Klotz FW, Miller LH.
Invasion of erythrocytes by Plasmodium falciparum malaria
parasites. Evidence for receptor heterogeneity and two receptors.
Blood 1986;67:1519�/21.
[9] Dolan SA, Proctor JL, Alling DW, Okubo Y, Wellems TE, Miller
LH. Glycophorin B as an EBA-175 independent Plasmodium
falciparum receptor of human erythrocytes. Mol Biochem Para-
sitol 1994;64:55�/63.
[10] Okoyeh JN, Pillai CR, Chitnis CE. Plasmodium falciparum field
isolates commonly use erythrocyte invasion pathways that are
independent of sialic acid residues of glycophorin A. Infect
Immun 1999;67:5784�/91.
[11] Miller LH, Mason SJ, Dvorak JA, McGinnis MH, Rothman IK.
Erythrocyte receptors for Plasmodium knowlesi malaria: Duffy
blood group determinants. Science 1975;189:561�/3.
[12] Haynes JD, Dalton JP, Klotz FW, et al. Receptor-like specificity
of a Plasmodium knowlesi malarial protein that binds to Duffy
antigen ligands on erythrocytes. J Exp Med 1988;167:1873�/81.
[13] Adams JH, Sim BKL, Dolan SA, Fang X, Kaslow DC, Miller
LH. A family of erythrocyte binding proteins of malaria parasites.
Proc Natl Acad Sci (USA) 1992;89:7085�/9.
[14] Camus D, Hadley TJ. A Plasmodium falciparum antigen that
binds to host erythrocytes and merozoites. Science 1985;230:553�/
5.
[15] Wertheimer SP, Barnwell JW. Plasmodium vivax interaction with
the human Duffy blood group glycoprotein: identification of a
receptor like protein. Exp Parasitol 1989;69:340�/50.
[16] Adams JH, Hudson DH, Torii M, et al. The Duffy receptor
family of Plasmodium knowlesi is located within micronemes of
invasive malaria merozoites. Cell 1990;63:141�/53.
[17] Sim BKL, Orlandi PA, Haynes JD, et al. Primary structure of the
175K Plasmodium falciparum erythrocyte binding antigen and
identification of a peptide, which elicits antibodies that inhibit
malaria merozoite invasion. J Cell Biol 1990;111:1877�/84.
[18] Fang X, Kaslow DC, Adams JH, Miller LH. Cloning of the of
Plasmodium vivax Duffy receptor. Mol Biochem Parasitol
1991;44:125�/32.
A.P. Singh et al. / Molecular & Biochemical Parasitology 121 (2002) 21�/3130

[19] Chitnis CE, Miller LH. Identification of the erythrocyte binding
domains of Plasmodium vivax and Plasmodium knowlesi proteins
involved in erythrocyte invasion. J Exp Med 1994;180:497�/506.
[20] Sim BKL, Chitnis CE, Wasniowska K, Hadley TJ, Miller LH.
Receptor and ligand domains for invasion of erythrocytes by of
Plasmodium falciparum . Science 1994;264:1941�/4.
[21] Ranjan A, Chitnis CE. Mapping regions containing binding
residues within functional domains of Plasmodium vivax and
Plasmodium knowlesi erythrocyte-binding proteins. Proc Natl
Acad Sci (USA) 1999;96:14067�/72.
[22] Peterson DS, Miller LH, Wellems TE. Isolation of multiple
sequences from the Plasmodium falciparum genome that encode
conserved domains homologous to those in erythrocyte-binding
proteins. Proc Natl Acad Sci (USA) 1995;92:7100�/4.
[23] Peterson DS, Wellems TE. EBL-1, a putative erythrocyte binding
protein of Plasmodium falciparum , maps within a favored linkage
group in two genetic crosses. Mol Biochem Parasitol
2000;105:105�/13.
[24] Thompson JK, Triglia T, Reed MB, Cowman AF. A novel ligand
from Plasmodium falciparum that binds to a sialic acid-containing
receptor on the surface of human erythrocytes. Mol Microbiol
2001;41:47�/58.
[25] Mayer DCG, Kaneko O, Taylor DEH, Reid ME, Miller LH.
Characterization of a Plasmodium falciparum erythrocyte-binding
protein paralogous to EBA-175. Proc Natl Acad Sci (USA)
2001;98:5222�/7.
[26] Liang H, Narum DL, Fuhrmann SR, Luu T, Sim BKL. A
recombinant baculovirus-expressed Plasmodium falciparum re-
ceptor-binding domain of erythrocyte binding protein EBA-175
biologically mimics native protein. Infect Immun 2000;68:3564�/8.
[27] Ockenhouse CF, Barbosa A, Blackall DP, et al. Sialic acid
binding of baculovirus-expressed recombinant antigens from
Plasmodium falciparum EBA-175 to glycophorin A. Mol Biochem
Parasitol 2001;113:9�/21.
[28] Narum DL, Haynes JD, Fuhrmann S, et al. Antibodies against
the Plasmodium falciparum receptor binding domain of EBA-175
block invasion pathways that do not involve sialic acids. Infect
Immun 2000;68:1964�/6.
[29] Dutta S, Daugherty JR, Ware LA, Lanar DE, Ockenhouse CF.
Expression, purification and characterization of a functional
region of the Plasmodium vivax Duffy binding protein. Infect
Immun 2000;109:179�/84.
[30] Singh S, Pandey K, Chattopadhayay R, et al. Biochemical,
biophysical and functional characterization of bacterially ex-
pressed and refolded receptor binding domain of Plasmodium
vivax Duffy binding protein. J Biol Chem 2001;206:17111�/6.
[31] Bohm G, Muhr R, Jaenicke R. Quantitative analysis of protein
far UV circular dichroism spectra by neural networks. Prot Eng
1992;5:191�/5.
[32] Cohen GH, Wilcox WC, Sodora DL, et al. Expression of herpes
simplex virus type 1 glycoprotein D deletion mutants in mamma-
lian cells. J Virol 1988;62:1932�/40.
[33] Horuk R, Chitnis CE, Darbonne WC, et al. A receptor for the
malarial parasite Plasmodium vivax : the erythrocyte chemokine
receptor. Science 1993;261:1182�/4.
[34] Smith JD, Subramanian G, Gamain B, Baruch DI, Miller LH.
Classification of adhesive domains in the Plasmodium falciparum
erythrocyte membrane protein 1 family. Mol Biochem Parasitol
2000;110:293�/310.
[35] Jones TR, Narum DL, Gozalo AS, et al. Protection of Aotus
monkeys by Plasmodium falciparum EBA-175 region II DNA
prime-protein boost immunization regimen. J Infect Dis
2001;183:303�/12.
A.P. Singh et al. / Molecular & Biochemical Parasitology 121 (2002) 21�/31 31