antitumor and antimetastatic activity of ribozymes...
TRANSCRIPT

Antitumor and Antimetastatic Activity of Ribozymes Targeting theMessenger RNA of Vascular Endothelial Growth Factor Receptors
Pamela A. Pavco,1 Karyn S. Bouhana,Anna M. Gallegos, Arun Agrawal,Karin S. Blanchard, Susan L. Grimm,Kristi L. Jensen, Lori E. Andrews,Francine E. Wincott, Patrice A. Pitot,Robert J. Tressler,2 Cynthia Cushman,3
Mark A. Reynolds,4 and Tom J. Parry5
Ribozyme Pharmaceuticals, Inc., Boulder, Colorado 80301 [P. A. Pa.,K. S. Bo., A. M. G., A. A., K. S. Bl., S. L. G., K. L. J., L. E. A.,F. E. W., M. A. R., T. J. P.], and Chiron Corp., Emeryville, California94608 [P. A. Pi., R. J. T., C. C.]
ABSTRACTChemically stabilized hammerhead ribozymes are nu-
clease-resistant, RNA-based oligonucleotides that selectivelybind and cleave specific target RNAs. Due to their potentialfor specifically inhibiting gene expression, ribozymes arebeing investigated for therapeutic applications as well as forthe elucidation of gene function. In particular, we haveinvestigated ribozymes that target the mRNA of the vascularendothelial growth factor (VEGF) receptors because VEGFsignaling is an important mediator of tumor angiogenesisand metastasis. Here we report pharmacodynamic studiestesting anti-Flt-1 (VEGFR-1) and anti-KDR (VEGFR-2) ri-bozymes in animal models of solid tumor growth and me-tastasis. Ribozymes targeting eitherFlt-1 or KDR signifi-cantly inhibited primary tumor growth in a highlymetastatic variant of Lewis lung carcinoma. However, onlytreatment with the anti-Flt-1 ribozyme resulted in a statis-tically significant and dose-dependent inhibition of lung me-tastasis in this model. The anti-Flt-1 ribozyme was thentested in a xenograft model of human metastatic colorectalcancer in which significant inhibition of liver metastasis wasobserved. Taken together, these data represent the firstdemonstration that synthetic ribozymes targeting VEGF re-
ceptor mRNA reduced the growth and metastasis of solidtumors in vivo.
INTRODUCTIONIn the early 1970s (1), Folkman hypothesized that solid
tumor growth and metastasis are critically dependent on angio-genesis, the formation of new blood vessels from preexistingvasculature. Over the past few decades, many mediators ofangiogenesis have been characterized, providing new and im-portant targets for drug discovery research. Considerable efforthas been directed toward the development of pharmacologicalagents that modulate specific pathways associated with angio-genesis.
Among the many known triggers of tumor angiogenesis,VEGF6 has emerged as a relatively specific effector (2, 3). Infact, VEGF expression has been observed in many human tumortypes (4–10), is up-regulated in response to hypoxia (11, 12),and has been specifically linked with tumor neovascularization(13–15). Tumor cells engineered to express VEGF constitu-tively exhibit enhanced tumor growth and angiogenic pheno-types (16). Conversely, treatments with anti-VEGF monoclonalantibodies have been shown to inhibit the growth of a variety ofsolid tumors in murine models (8, 17, 18).
Inhibition of VEGF signaling at the receptor level has beenproposed as a means of inhibiting VEGF-dependent tumorgrowth and metastasis (19–21). The mitogenic selectivity ofVEGF for endothelial cells is due to the relatively exclusivedistribution of VEGF receptors in this cell type (2, 22). Twomajor human VEGF receptor subtypes have been identified andare known as Flt-1 (VEGFR-1) and KDR (VEGFR-2 or Flk-1 inthe mouse) (23). Although Flt-1 and KDR/Flk-1 receptors bothappear to be involved in angiogenesis, their respective roles inthis process are only partially elucidated.
Both Flt-1 and KDR/Flk-1 receptors may play importantroles in VEGF-stimulated cell proliferation. For example, theexpression of the murine Flk-1 receptor is associated withproliferating endothelial cells (24), and human endothelial cellsexpressing the KDR receptor exhibit shape changes and en-hanced DNA synthesis on stimulation with VEGF (25). Humanumbilical vein endothelial cells expressing Flt-1 receptor mu-tants are resistant to VEGF-stimulated proliferation (26). Inaddition, Barleonet al. (21) demonstrated that purified VEGFand conditioned media from hypoxic tumor cell lines can up-regulate the expression of Flt-1 but not KDR/Flk-1 receptors incultured endothelial cells.
Developmental models provide useful insight into thephysiological roles of these receptorsin vivo. Although muta-tions in either Flk-1 or Flt-1 are fatal to developing mouse
Received 12/8/99; revised 2/23/00; accepted 2/23/00.The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisementin accordance with 18 U.S.C. Section 1734 solely toindicate this fact.1 To whom requests for reprints should be addressed, at RibozymePharmaceuticals, Inc., 2950 Wilderness Place, Boulder, CO 80301.Phone: (303) 449-6500; Fax: (303) 449-6995; E-mail: [email protected] Present address: Matrix Pharmaceuticals, 34700 Campus Drive, Fre-mont, CA 94555.3 Present address: Elan Pharmaceuticals, Avenue One Letchworth, Hert-fordshire, United Kingdom SG62HU.4 Present address: Incyte Microarrays, 6519 Dumberston Circle, Fre-mont, CA 94555.5 Present address: Human Genome Sciences, Inc., 9410 Key WestAvenue, Rockville, MD 20850.
6 The abbreviations used are: VEGF, vascular endothelial growth factor;LLC-HM, Lewis lung carcinoma-highly metastatic variant.
2094Vol. 6, 2094–2103, May 2000 Clinical Cancer Research
Research. on November 2, 2018. © 2000 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

embryos, their respective embryological abnormalities in vas-culogenesis and angiogenesis differ (27–29). Abnormal Flk-1expression leads to defective endothelial cell development (28,29), whereas mutant Flt-1 embryos show normally differentiatedendothelial cells that form highly abnormal and disorganizedvascular structures (27). This body of evidence suggests thatinhibition of VEGF receptor signaling could be an importanttarget for therapeutic intervention in pathological angiogenesis.
Hammerhead ribozymes aretrans-acting, RNA-based en-zymes that specifically bind and cleave target RNA. In recentyears, considerable effort has been directed toward the applica-tion of ribozyme technologies to inhibit specific gene expression(30). Stabilization chemistries have been developed to renderthese molecules highly resistant to serum nucleases (31) andhence suitable forin vivoapplications. As a result, local deliveryof chemically stabilized ribozymes has been reported to inhibitspecifically cytokine-stimulated expression of stromelysinmRNA in a rabbit knee osteoarthritis model (32) and VEGF-stimulated angiogenesis in a rat corneal pocket model (33). Inaddition, a stabilized ribozyme targeting protein kinase CamRNA was shown to reduce sc implanted glioma tumor growthin rats (34).
We reported previously on ribozymes that target specificsites in the mRNA of each of the two major VEGF receptorsubtypes, Flt-1 and KDR (33). Testing in cultured humanmicrovascular endothelial cells was used as a way to choosethe most effective anti-Flt-1 andKDR ribozymes to advanceinto animal studies. These cell culture studies resulted in thechoice of one lead ribozyme site in each mRNA target. Inparticular, ribozymes targetingFlt-1 site 4229 orKDR site726 were chosen from a panel of ribozymes targeting differ-ent sites spanning the length of the target mRNAs. Thesechoices were based on the ability of the ribozyme to inhibitVEGF-stimulated cell proliferation and to decrease specifi-cally their targeted mRNA in cell culture relative to attenu-ated controls. In subsequentin vivo studies, anti-Flt-1andKDR ribozymes targeting these specific sites significantlydecreased angiogenesis when tested in a corneal pocketmodel of VEGF-induced angiogenesis. In these two assays,controls that had the same binding arms as the ribozymes butreduced cleavage activity due to nucleotide changes in thecatalytic core showed little, if any, effect. Here we reportpharmacodynamic studies using systemic delivery of stabi-lized hammerhead ribozymes designed to inhibit the expres-
sion of the human VEGF receptors, Flt-1 or KDR. Eachribozyme is targeted to a conserved sequence in eitherFlt-1or KDR mRNA that is also present in the mouse and rat,enabling rodent studies to be conducted with the same agentin these species. The Lewis lung carcinoma model was cho-sen for initial study because neovascularization in this tumortype is VEGF dependent (35, 36), and the highly metastaticline can be used to study the process of metastasis withoutresection of the primary tumor. In this model, both theanti-Flt-1 and the anti-KDRribozyme inhibited primary tu-mor growth when administered iv by continuous infusion.However, only treatment with the anti-Flt-1 ribozyme re-sulted in a statistically significant and dose-dependent inhi-bition of lung metastasis in this model. In further testing ofthe anti-Flt-1 ribozyme in a model of human colorectalmetastasis, the number of liver metastases was also signifi-cantly reduced.
MATERIALS AND METHODSRibozyme Synthesis
Hammerhead ribozymes and their corresponding attenu-ated controls were synthesized and purified as described previ-ously (31, 32, 37). Sequences and the locations of modifiednucleotides are given in Table 1. The five remaining unmodifiedribonucleotides in the catalytic core are essential for cleavageactivity (31). The four phosphorothioate linkages at the 59-endand the 39-39inverted deoxyabasic residue at the 39-end provideadditional resistance to exonucleases (31). The attenuated ri-bozyme controls maintain the binding arm sequence of theparent ribozyme and thus are still capable of binding the mRNAtarget. However, they have two nucleotide changes in the coresequence (Table 1) that substantially reduce their ability to carryout the cleavage reaction (Fig. 1). Ribozymes were designed totarget Flt-1 or KDR/Flk-1 mRNA sites conserved in human,mouse, and rat.
In Vitro Ribozyme Cleavage AssaysRNA cleavage rates were measured for the ribozymes and
their paired attenuated controls using a 15-nucleotide syntheticcomplementary RNA substrate as described previously (38).Briefly, the ribozymes (500 nM) were incubated with stoichio-metric amounts of substrate (and trace amounts of32P-59-end-labeled substrate) in 50 mM Tris-HCl buffer (pH 8.0) and 40 mM
Table 1 Ribozyme and attenuated control sequences with locations of modified nucleotides
Target sitea Ribozyme
Ribozyme sequenceb (59–39)
ActivityBinding arm Core & stem II Binding arm
Flt-4229 RPI.4610 gsasgsusug cUGAuGaggccgaaaggccGaa AgucugB ActiveFlt-4229 RPI.4611 gsasgsusug cUuAuGaggccgaaaggccGau AgucugB AttenuatedKDR-726 RPI.4733 usascsasauu cUGAuGaggccgaaaggccGaa AagacaaB ActiveKDR-726 RPI.4734 usascsasauu cUuAuGaggccgaaaggccGau AagacaaB Attenuated
a Ribozyme target sites are named according to the cleavage position in humanFlt-1 (European Molecular Biology Laboratory accession numberX51602) or humanKDR mRNA (European Molecular Biology Laboratory accession number L04947).
b Modifications are indicated as follows: 29-O-methyl nucleotides, lowercase letters; ribonucleotides, uppercase G and A; 29-C-allyl uridine,U;inverted 39-39deoxyabasic,B. The positions of four phosphorothioate linkages at the 59-end are indicated by an inferior s. Nucleotides comprisingthe base paired region of stem II are initalic. Nucleotide changes in the core of the attenuated controls areunderlined.
2095Clinical Cancer Research
Research. on November 2, 2018. © 2000 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

MgCl2 at 37°C. The magnesium-dependent cleavage reactionswere quenched with EDTA at specified time points and assayedby PAGE with band quantitation using a Molecular DynamicsPhosphorImager (model 425E).
Animal ExperimentationAll animal experimentation was performed in accordance
with the Guide for the Care and Use of Laboratory Animals (39)and the policies and procedures of the Ribozyme Pharmaceuti-cals, Inc. and Chiron Corp. animal care and use programs.
LLC-HMTumor Inoculations. Female 6–8-week-old C57BL/6
mice (Harlan Bioproducts, Indianapolis, IN) were housed ingroups of four and inoculated sc in the left flank with 53 105
LLC-HM cells from brei preparations of tumors exhibiting ahighly metastatic phenotype in mice (six passages; the originalLLC-HM cell line was kindly provided by Dr. MichaelO’Reilly; Children’s Hospital and Harvard Medical School,Boston, MA).
Ribozyme Administrations. Three days after tumor in-oculation, animals were anesthetized with 125 mg/kg ketamineand 8 mg/kg xylazine in sterile veterinary saline and placed in asupine position on a 37°C warming pad. The left jugular veinwas exposed, and sc fat and fascia were cleared from the vessel.After tying off the jugular vein distally, a small incision wasmade in the jugular vein, and a catheter constructed from PE 50polyethylene medical tubing (prefilled with sterile saline) wasintroduced into the vessel to approximately 0.5 cm. The vessel
was secured to the catheter proximal to the incision with threeligatures and anchored to the surrounding muscle tissue using4-0 silk. The catheter was fed through a trochar implanted sc,passing from the neck wound dorsally toward the nuchal region.When the catheter was in place and its volume had been re-placed with test solution, it was attached to an Alzet osmoticminipump (flow rate, 12ml/day). Pumps were filled with eitherribozyme, attenuated control, or saline solutions and implantedsc through a dorsal incision to a final position caudal to thescapulae. Animals were allowed to recover on 37°C warmingpads.
Test substances were dissolved in saline at concentrationsranging from 1.7–167 mg/ml. Beginning 3 days after tumorinoculation, animals were dosed at 0, 1, 3, 10, 30, and 100mg/kg/day for 14 days. These doses correspond to 0, 3, 9, 30,90, and 300 mg/m2/day. Ten animals were used for each treat-ment group. Less than 10% of the 210 animals in the LLC-HMstudy were removed early, primarily because of catheter failures(i.e., detaching from the vessel or pump), or because the animaldid not survive pump implantation. Thus, 7–10 animals re-mained in each group at the end of the study.
Measurement of Tumor Growth and Metastasis. Inpilot experiments, LLC-HM-injected animals began to becomemoribund after about 24 days due to substantial lung tumorburden. To assess both primary tumor growth and metastaticdisease in the same animal, the experiment was terminated onday 25, before control animals became moribund. Primary tu-mor length and width were measured with microcalipers everyother day from postinoculum days 4–24. Tumor volume was
Fig. 1 In vitro ribozyme cleav-age analysis. A time course ofinvitro cleavage analysis on a com-plementary synthetic RNA sub-strate was carried out for the ri-bozymes and paired attenuatedcontrols targeting (A) Flt-1 site4229 and (B) KDR site 726. Theupper and lower bandson thepolyacrylamide gel correspond tofull-length and cleaved RNA sub-strate, respectively. Along withstoichiometric amounts of unla-beled RNA substrate, a traceamount of 32P-59-end-labeledRNA substrate was included inthe reactions for detection pur-poses. Based on PAGE analysisof the time course of cleavage,ribozyme cleavage rates werecalculated using a double expo-nential curve fit as described pre-viously (38). According to thedata, the first-order rate constants(k1) for the anti-Flt-1 ribozymeand attenuated control were0.464 6 0.005 min21 and0.001 6 4 3 1025 min21, re-spectively. The correspondingk1
values for the anti-KDRribozymeand control were 0.7806 0.071min21 and 0.00026 2 3 1024
min21, respectively.
2096Anticancer Effects of Ribozymes Targeting Angiogenesis
Research. on November 2, 2018. © 2000 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
calculated using the following formula: 0.5(length)(width2).Twenty-five days after inoculation, animals were killed, andtumors and lungs were removed. The numbers of surface mac-rometastases on the lung were counted under a dissecting mi-croscope at a magnification of325.
Data Analysis. Group tumor volume means from eachtreatment group obtained from days 12–24 postinoculation weresubjected separately to normality testing and one-way ANOVA.Treatment group means for ribozymes and their paired attenu-ated controls were compared using a Tukey-Kramerpost hoctest for significance (a 5 0.05). The mean numbers of pulmo-nary metastases from each treatment group (including the salinecontrol) were subjected to normality testing and one-wayANOVA, followed by a Dunnett’s test for significance (a 50.05) using the saline group as the control.
KM12L4a Human Colorectal Carcinoma/LiverMetastasis Model
Tumor Inoculations. Male 6–8-week-old nude mice(Charles River Laboratories) were housed in groups of five. Onday 0, each spleen was inoculated with 53 105 KM12L4a cellswithin the splenic capsule as described below. KM12L4a cellswere obtained from Dr. Jerald Killion (University of TexasM. D. Anderson Cancer Center, Houston, TX). Cells weregrown in DMEM with 2 mM glutamine, 10% FBS with nones-sential amino acids, sodium pyruvate, and vitamins. On day 5after tumor cell inoculation, the spleens were removed from theanimals.
The tumor cell inoculation was performed as follows: (a)animals were anesthetized with 125 mg/kg ketamine and 8 mg/kgxylazine; (b) an abdominal incision of;3 mm was made to the leftof the midline; and (c) the spleen was gently exteriorized. TheKM12L4a cell suspension was inoculated into the spleen afterslowly inserting a 28-gauge needle into the large pole of the spleenuntil the bevel of the syringe became visible in the central portionof the spleen just beneath the splenic capsule.
Ribozyme Administrations. Three days after inocula-tion, animals were surgically implanted with sc 14-day Alzetosmotic minipumps (flow rate, 12ml/day) filled with eithersaline or anti-Flt-1ribozyme solutions. Animals were dosed at0, 12, 36, and 100 mg/kg/day for 14 days. These administereddoses correspond to 0, 36, 108, and 300 mg/m2/day. On day 18,the initial pumps were removed and replaced with freshminipumps containing either saline or the same doses of anti-Flt-1 ribozyme. Animals were observed daily after each surgery,and healing of the surgical incision sites was unaffected by anytreatment as compared with the saline control group. The studywas ended on day 41.
Fifteen animals were included in each treatment group. Iftumor cells successfully seed the liver, metastatic tumor forma-tion in this xenograft model is usually rapid and aggressive;however, a poor tumor take is possible. Animals were randomlydistributed before assignment to treatment groups to address thevariable outgrowth properties of this model. Several animalswere removed from the study as follows: (a) saline group, oneanimal was killed on day 33 because of the tumor burden; (b) 12mg/kg/day group, one animal was killed early (day 17) becauseof continued loss of body weight, and one animal was founddead on day 35; (c) 100 mg/kg/day, one animal did not survive
cell implantation, and one animal did not survive pump replace-ment. Thus, the final sample size for the saline-treated groupand the 12, 36, and 100 mg/kg/day dose groups was 14, 13, 15,and 13 mice, respectively.
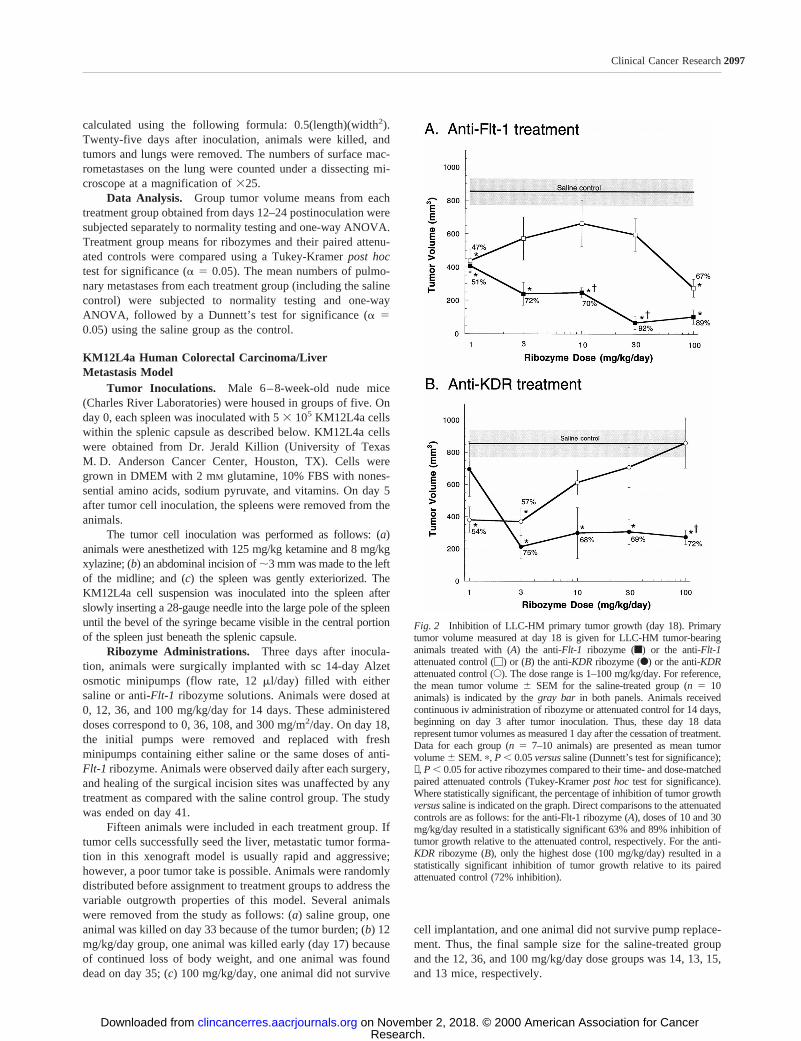
Fig. 2 Inhibition of LLC-HM primary tumor growth (day 18). Primarytumor volume measured at day 18 is given for LLC-HM tumor-bearinganimals treated with (A) the anti-Flt-1 ribozyme (f) or the anti-Flt-1attenuated control (M) or (B) the anti-KDRribozyme (F) or the anti-KDRattenuated control (E). The dose range is 1–100 mg/kg/day. For reference,the mean tumor volume6 SEM for the saline-treated group (n 5 10animals) is indicated by thegray bar in both panels. Animals receivedcontinuous iv administration of ribozyme or attenuated control for 14 days,beginning on day 3 after tumor inoculation. Thus, these day 18 datarepresent tumor volumes as measured 1 day after the cessation of treatment.Data for each group (n 5 7–10 animals) are presented as mean tumorvolume6 SEM.p, P , 0.05versussaline (Dunnett’s test for significance);✝, P , 0.05 for active ribozymes compared to their time- and dose-matchedpaired attenuated controls (Tukey-Kramerpost hoctest for significance).Where statistically significant, the percentage of inhibition of tumor growthversussaline is indicated on the graph. Direct comparisons to the attenuatedcontrols are as follows: for the anti-Flt-1 ribozyme (A), doses of 10 and 30mg/kg/day resulted in a statistically significant 63% and 89% inhibition oftumor growth relative to the attenuated control, respectively. For the anti-KDR ribozyme (B), only the highest dose (100 mg/kg/day) resulted in astatistically significant inhibition of tumor growth relative to its pairedattenuated control (72% inhibition).
2097Clinical Cancer Research
Research. on November 2, 2018. © 2000 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Measurement of Liver Weight and Liver Metastasis.On day 41, all surviving animals were killed by CO2 inhalation.Livers were removed, weighed, and scored for liver metastases.In cases where the number of tumor foci was too numerous tocount (i.e.,.100), a count of 101 was applied.
Data Analysis. All statistical testing was performed atthe a 5 0.05 level. Because the individual numbers of livermetastases in each treatment group were not normally distrib-uted, group means were compared using the Kruskal-Wallisnonparametric rank test at 95% confidence limits. Individualliver weight means were normally distributed. The liver weightdata were subjected to ANOVA followed by Dunnett’s test forsignificance. Differences between treatment groups with respectto the presence or absence of metastases were compared usingthe x2 test.
RESULTSCatalytic Cleavage Rates of Ribozymes and Paired At-
tenuated Controls. In previousin vivo studies, anti-Flt-1andKDR ribozymes significantly decreased angiogenesis whentested in a corneal pocket model of VEGF-induced angiogenesis(33). Attenuated controls were included that retained the bindingarm sequence of the parent ribozymes and hence were stillcapable of binding to their intended mRNA target. However,these controls had two nucleotide changes in the core region(sequences shown in Table 1) that substantially reduced theirability to carry out the cleavage reaction. These controls hadnegligible effect on the end points measured previously in cellculture and only reduced angiogenesis at the highest dosestested in the corneal model. Thus, for direct comparison, similarcontrols were included in the initial Lewis lung carcinomastudies reported here.
Although these initialin vivo data were established using3-bp stem II ribozymes, the 4-bp stem II versions (Table 1) wereused in the current studies. This decision was based, in part, onthe higher in vitro cleavage activity of the 4-bp stem II ri-bozymes compared with that of the 3-bp stem II ribozymes (seebelow), although their activities in cell culture (33) and in thecorneal pocket model are comparable (data not shown).
Fig. 1 shows the results ofin vitro catalytic cleavage assaysconducted for the 4-bp stem II anti-Flt-1 and anti-KDRri-bozymes, along with their paired attenuated controls. Thechanges in the core resulted in attenuated analogues that haveinvitro cleavage rates that are at least 2 orders of magnitude lowerthan those of the active ribozymes. For the anti-Flt-1 ribozymeand its paired attenuated control, the first-order rate constants(k1) were 0.4646 0.005 min21 and 0.0016 4 3 1025 min21,respectively. For the anti-KDR ribozyme and its paired control,k1 values were 0.7806 0.071 min21 and 0.00026 2 3 1024
min21, respectively. For comparison,k1 values for the 3-bpstem II versions of these ribozymes reported previously in thecorneal model study (33) were lower (0.0816 0.007 and0.4346 0.024 min21 for the anti-Flt-1and anti-KDRribozyme,respectively). These cleavage data, together with the originalcell culture and corneal model results (33), demonstrate that theeffect observed for active ribozymes occurs through a ribozyme-specific (i.e., cleavage) mechanism.
Effects of Anti-Flt-1 or KDR Ribozymes on LLC-HMPrimary Tumor Growth. Neovascularization of Lewis lungcarcinoma tumors depends on VEGF (35, 36). A highly meta-static variant of Lewis lung carcinoma (LLC-HM) was selectedin the present study because it allows for the assessment ofprimary tumor growth and metastasis without resection of theprimary tumor. Thus, both the antitumor and antimetastaticeffects of the anti-Flt-1and anti-KDRribozymes (together withtheir respective attenuated controls) were evaluated in theLLC-HM model after continuous iv administration. Pilot studiesconducted before the current study clearly demonstrated that theanti-Flt-1or the anti-KDRribozyme did not affect the healing of
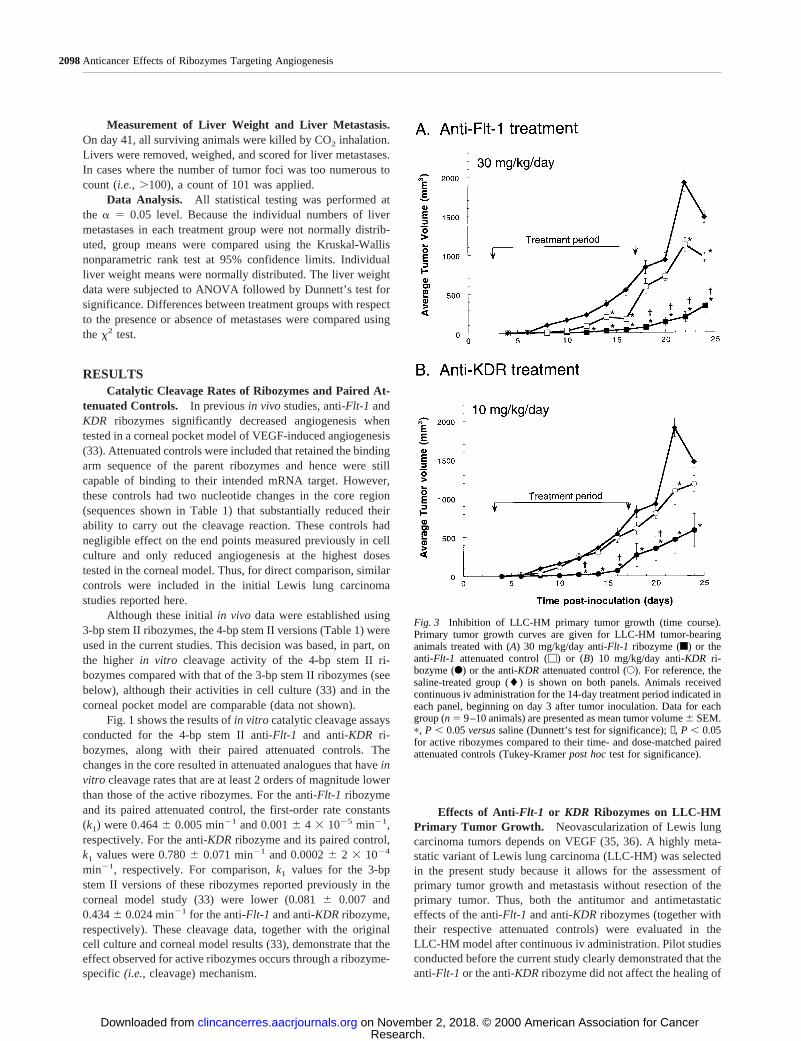
Fig. 3 Inhibition of LLC-HM primary tumor growth (time course).Primary tumor growth curves are given for LLC-HM tumor-bearinganimals treated with (A) 30 mg/kg/day anti-Flt-1ribozyme (f) or theanti-Flt-1 attenuated control (M) or (B) 10 mg/kg/day anti-KDRri-bozyme (F) or the anti-KDRattenuated control (E). For reference, thesaline-treated group (l) is shown on both panels. Animals receivedcontinuous iv administration for the 14-day treatment period indicated ineach panel, beginning on day 3 after tumor inoculation. Data for eachgroup (n5 9–10 animals) are presented as mean tumor volume6 SEM.p, P , 0.05versussaline (Dunnett’s test for significance);✝, P , 0.05for active ribozymes compared to their time- and dose-matched pairedattenuated controls (Tukey-Kramerpost hoctest for significance).
2098Anticancer Effects of Ribozymes Targeting Angiogenesis
Research. on November 2, 2018. © 2000 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
any surgical wounds associated with jugular catheterizationand/or implantation of the Alzet minipump in mice (data notshown).
In general, both anti-Flt-1and anti-KDRribozymes inhib-ited primary tumor growth within the 1–100 mg/kg/day iv doserange. The effect on primary tumor growth observed immedi-ately after the dosing period (day 18) is given in Fig. 2. Theanti-Flt-1 ribozyme (Fig. 2A) reduced LLC-HM primary tumorgrowth in a dose-dependent manner as compared with eithersaline or its corresponding attenuated control. At the lowest dose(1 mg/kg/day), both the anti-Flt-1 ribozyme and its attenuatedcontrol reduced primary tumor growth similarlyversussalinecontrols (;50%;P , 0.001 for both observations). However,with increasing dose, active anti-Flt-1ribozyme reduced pri-mary tumor growth to a greater extent than the attenuatedcontrol (P, 0.001 for 10 and 30 mg/kg/day dose groups). Theattenuated control showed no further tumor growth inhibitionuntil the highest dose (100 mg/kg/day). The greatest inhibitionof tumor growth by the active anti-Flt-1ribozyme (92% ascompared with saline;P 5 0.0001) and the largest significantdifference between activeFlt-1 ribozyme and its attenuatedcontrol (89%;P 5 0.0001) occurred at a dose of 30 mg/kg/day.Mean tumor volumes for the active anti-Flt-1 ribozyme andattenuated control groups at this dose were 65.346 37.18 and585.456 98.67 mm3, respectively, compared with 830.86682.53 mm3 for the saline control group.
The differences between the antitumor effects of the anti-KDR ribozyme and its corresponding attenuated control weremore complex (Fig. 2B). At a dose of 1 mg/kg/day, the anti-KDR ribozyme had no effect on primary tumor growthversusthe saline control. However, at all higher doses (3–100 mg/kg/day), the anti-KDRribozyme significantly inhibited primarytumor growth as compared with the saline control by about thesame degree (68–75%;P , 0.0001 for all observations). Theantitumor dose relationship profile of the attenuated control wasmore complicated. Doses of 1 and 3 mg/kg/day showed signif-icant tumor growth inhibition compared with saline (54–57%;P 5 0.0019 for both observations). In contrast, higher doses ofthe attenuated control had no effect on tumor growth. For theanti-KDR ribozyme, 10 mg/kg/day was the lowest dose thatresulted in significant inhibition of primary tumor volume (68%;P 5 0.0001) where the corresponding attenuated control had nosignificant effect (versussaline). A statistically significant dif-ference between the effects of the active anti-KDR ribozyme andits attenuated control was detected at the 100 mg/kg/day dose(72%; P , 0.001).
A more complete view of the integrated difference betweenthe active ribozymesversustheir attenuated controls is providedby the primary tumor growth characteristics from the entire24-day growth period. Overall, the greatest difference betweenthe ribozymes and their controls, even after dosing was com-pleted, was observed with a dose of 30 mg/kg/day for theanti-Flt-1 ribozyme and a dose of 10 mg/kg/day for the anti-KDR ribozyme. Primary tumor growth curves for these twotreatments are shown in Fig. 3. For the anti-Flt-1ribozyme (Fig.3A), the maximum tumor volume reduction compared with thesaline control was 92% immediately after the last day of dosing(day 18) and 77% at day 24. Moreover, the maximum tumorvolume reduction for the anti-Flt-1ribozyme compared with its
attenuated control was 89% on day 18 and 65% at day 24. Itshould be noted that the reduction observed with the anti-Flt-1ribozyme from day 18 through day 24 was significant comparedwith both the saline and attenuated controls, even though ri-bozyme treatment was discontinued on day 17 (P , 0.05).
With 10 mg/kg/day anti-KDRribozyme (Fig. 3B), themaximal tumor volume reduction compared with saline was68% on day 18 (P5 0.0001) and 60% on day 24 (P , 0.05). Infact, the reduction in tumor growth at this dose was statisticallysignificant when compared with saline from days 12–24 (P ,0.05 for all observations). In contrast, LLC-HM tumor growthwas not significantly reduced by treatment with the anti-KDRattenuated control at this dose, except as noted on day 22. Even
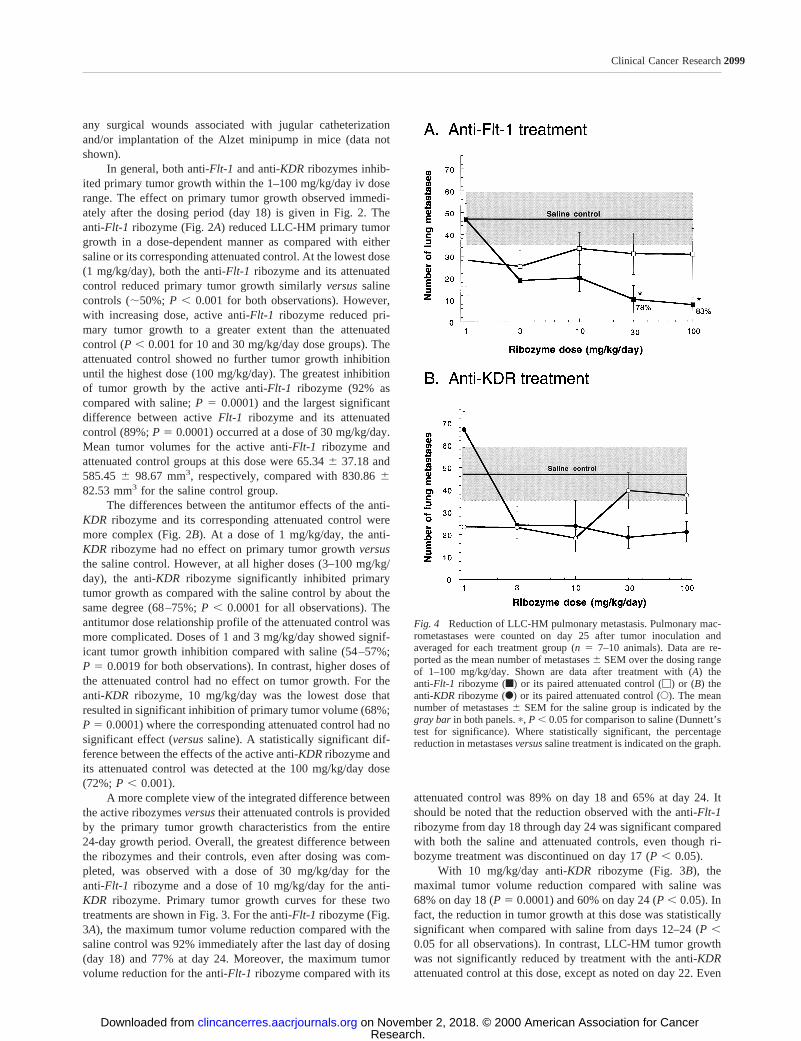
Fig. 4 Reduction of LLC-HM pulmonary metastasis. Pulmonary mac-rometastases were counted on day 25 after tumor inoculation andaveraged for each treatment group (n5 7–10 animals). Data are re-ported as the mean number of metastases6 SEM over the dosing rangeof 1–100 mg/kg/day. Shown are data after treatment with (A) theanti-Flt-1 ribozyme (f) or its paired attenuated control (M) or (B) theanti-KDR ribozyme (F) or its paired attenuated control (E). The meannumber of metastases6 SEM for the saline group is indicated by thegray bar in both panels.p, P , 0.05 for comparison to saline (Dunnett’stest for significance). Where statistically significant, the percentagereduction in metastasesversussaline treatment is indicated on the graph.
2099Clinical Cancer Research
Research. on November 2, 2018. © 2000 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

so, the differences in tumor volume reduction between the activeanti-KDRribozyme and its attenuated control were statisticallydifferent only on a limited number of days,e.g.,day 20 wherea 56% difference was noted (P, 0.05).
Antimetastatic Activity of Anti- Flt-1 and KDR Ri-bozymes in LLC-HM Tumor-bearing Mice. The antimeta-static activities of the anti-Flt-1and anti-KDRribozymes andtheir attenuated controls were also determined (Fig. 4). For thisassessment, lung macrometastases were counted at the end ofthe study (postinoculation day 25 and 8 days after the cessationof treatment). As shown in Fig. 4A, the anti-Flt-1ribozymereduced the number of pulmonary metastases with respect tosaline controls in a dose-dependent manner. The greatest anti-metastatic activity with respect to saline controls (78% and 83%inhibition) was observed at doses of 30 and 100 mg/kg/dayanti-Flt-1 ribozyme, respectively (P5 0.005 for both observa-tions). In contrast, no antimetastatic activity was observed with
the anti-Flt-1attenuated control; the data were statistically in-distinguishable from saline controls at all doses and showed nodose-dependent trend.
Treatment with the anti-KDRribozyme also reduced thenumber of pulmonary metastases (Fig. 4B), but not to the samedegree as the anti-Flt-1ribozyme. For the dose range of 3–100mg/kg/day, the anti-KDRribozyme reduced the number of me-tastases;50–60%, although this was not significantly differentcompared with the saline control group. Interestingly, any re-duction in the number of pulmonary metastases present at thelowest doses of the attenuated control was not observed at thehigher doses. No dose of the anti-KDR attenuated control re-sulted in a statistical reduction of lung metastases (versussalinecontrol).
Inhibition of Liver Metastasis in a Model of HumanColorectal Cancer. Because the anti-Flt-1ribozyme exhib-ited a slightly more robust inhibition of primary tumor growthand also significantly inhibited lung metastasis in the LLC-HMmodel, this ribozyme was tested further in a metastatic model ofhuman colorectal cancer. In this model, human colorectal cancercells (KM12L4a) were implanted into the spleens of mice, andafter 5 days, the spleens were removed. Continuous sc admin-istration of the anti-Flt-1ribozyme was initiated 3 days after thesplenic inoculation and continued for 28 days. At the end of thestudy (day 41), livers were weighed, and spontaneous metasta-ses to the liver were counted. Although iv dosing was used in theLLC-HM study reported above, subsequent pharmacokineticstudies with the anti-Flt-1ribozyme in mice also supported scdosing. The anti-Flt-1ribozyme is;80% bioavailable after scadministration, and itst1/2 in plasma after a single administra-tion is similar to that seen with iv dosing (40). Thus, a sc routeof administration was used in this xenograft model in an effortto avoid the surgical complications associated with chronicvascular catheterization in immunocompromised mice.
The effects on indices of liver metastasis of tumor-bearinganimals after treatment with 12, 36, or 100 mg/kg/day anti-Flt-1ribozyme are listed in Table 2 and are shown graphically in Fig.5. As seen in Table 2, there was no statistical difference in liverweight between ribozyme-treated and control animals. How-ever, there was a clear decrease in metastatic incidence at 100mg/kg/day (P5 0.03) as well as a dose-dependent reduction inthe mean number of spontaneous metastases to the liver (P 5
Fig. 5 Inhibition of colorectal cancer metastasis to the liver by theanti-Flt-1 ribozyme. Metastases to the liver 41 days after splenic inoc-ulation of KM12L4a cells are reported. Data are reported as the numberof metastases in individual mice in each treatment group (F) and as themean number of metastases6 SEM (f). The sample size for thesaline-treated group and the 12, 36, and 100 mg/kg/day dose groups was14, 13, 15, and 13 mice, respectively. Where multilobular tumor clusterswith numerous foci (.100) were present, the value of.101 was used.Continuous sc infusion of 12, 36, or 100 mg/kg/day anti-Flt-1ribozymefrom day 3 to day 28 is compared with the saline control group.p, P ,0.05 for comparison with saline (Kruskal-Wallis nonparametric ranktest).
Table 2 Results of anti-Flt-1ribozyme treatment in the KM12L4a colorectal cancer model of liver metastasisa
TreatmentLiver metastatic
incidencebLiver weightc
Mean6 SEMLiver metastasesd
Mean6 SEMLiver metastasesd
Median
Saline 12/14 2.76 0.5 596 13 101a
12 mg/kg/day 11/13 3.16 0.6 636 14 10136 mg/kg/day 13/15 2.26 0.3 306 12 (P5 0.03) 5 (P5 0.03)100 mg/kg/day 6/13 (P5 0.03) 1.86 0.3 176 10 (P5 0.03) 0 (P5 0.03)Observation group NAe 2.06 0.1 NA NA
a Some livers had large multilobular tumor clusters consisting of numerous (.100) tumor foci that occupied from 30–80% of the liver lobesof some animals. These animals were scored as having.100 tumor foci/liver, and a value of 101 liver metastases was entered for statistical analysis.
b Liver metastatic incidence5 the number of livers with one or more metastases/total number of livers in the group. Differences were assessedby x2 methodology.
c Liver weights were normally distributed (ANOVA) and are reported as mean6 SEM. There were no statistically significant differences in liverweights between saline- and ribozyme-treated groups based on Dunnett’s test for significance.
d Liver metastases were counted, andPs were calculated based on comparison to saline using a nonparametric rank test (Kruskal-Wallis).e NA, not applicable because animals were not inoculated with tumor cells.
2100Anticancer Effects of Ribozymes Targeting Angiogenesis
Research. on November 2, 2018. © 2000 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

0.03 for the 36 and 100 mg/kg/day doses). Moreover, treatmentwith the anti-Flt-1ribozyme at doses of 36 and 100 mg/kg/dayreduced the median number of liver metastases to 5 and 0,respectively (P5 0.03 for both observations).
The significant decrease in metastasis resulting from treat-ment with the anti-Flt-1ribozyme can also be seen in Fig. 6.Hepatic metastases are visible asdarkened areasin photographsof livers from control animals treated only with saline (Fig. 6B).In contrast, the gross morphology of the livers treated with 100mg/kg/day anti-Flt-1ribozyme is similar to that of normal livers(Fig. 6A).
DISCUSSIONVEGF has long been shown to play a major role in the
recruitment and maintenance of blood supply for growing andmetastasizing tumors (3). Recently, the inhibition of VEGFsignaling at the receptor level has been proposed as a therapeuticstrategy for the treatment of solid cancers (19, 20). The presentstudy illustrates the antitumor and antimetastatic effects ofchemically stabilized hammerhead ribozymes targeting eitherthe Flt-1 or the KDR VEGF receptor mRNA in a metastatictumor model in which neovascularization is thought to be de-pendent on VEGF expression (35, 36). In addition, the inhibi-tory effects of the anti-Flt-1ribozyme on liver metastasis in amodel of human colorectal cancer are described. Based in parton these data, the anti-Flt-1ribozyme (ANGIOZYME) is cur-rently being developed for clinical oncology indications.
In supporting cell culture studies (33), a significant de-crease in cell proliferation was demonstrated with the activeanti-Flt-1 and anti-KDRribozymes, but not with their corre-sponding attenuated controls. In addition, these controls be-haved as expected in the corneal model of VEGF-inducedangiogenesis in that their effect on decreasing angiogenesis wassignificant only at the higher doses, if at all. These resultssuggest a ribozyme-specific (cleavage) mechanism of action.
In the pharmacodynamic studies reported here, the active
anti-Flt-1 and KDR ribozymes exhibited antitumor activity.Treatment with the anti-Flt-1ribozyme reduced LLC-HM pri-mary tumor growth (Fig. 2A) and the number of lung metastases(Fig. 4A) in a dose-dependent manner. Moreover, the number ofspontaneous liver metastases in the colorectal cancer model wassignificantly reduced after treatment with the two highest dosesof the anti-Flt-1 ribozyme (Table 2; Figs. 5 and 6). For theanti-KDR ribozyme, all doses except the lowest dose signifi-cantly inhibited LLC-HM primary tumor growth (Fig. 2B). Instark contrast to the anti-Flt-1 ribozyme, there was no signifi-cant effect on lung metastasis at any dose of the anti-KDRribozyme (Fig. 4B).
Whereas the anti-Flt-1attenuated control had minimal ef-fects on primary tumor growth and no effect on metastasis, someunexpected effects of the anti-KDRattenuated controls werenoted on primary tumor growth, particularly at the lowest dosestested (Figs. 2Band 4B). For the anti-KDRattenuated control,the lower doses inhibited primary tumor growth, but the higherdoses showed no inhibition. It is important to note that theanti-KDRcontrol failed to inhibit cell proliferation in culturedhuman endothelial cells (33) and did not statistically inhibitangiogenesis in the VEGF-induced rat corneal model (data notshown). Additional studies would be necessary to characterizeany specific role of this modified oligonucleotide in the complexprocesses involved in tumor growth.
The specific roles of each VEGF receptor in VEGF-induced endothelial cell proliferation and angiogenesis arepoorly understood. It is clear that the KDR/Flk-1 receptor me-diates VEGF-induced endothelial cell proliferation, shape, andmotility changes (25, 41). KDR/Flk-1 is also involved in em-bryonic vasculogenesis and angiogenesis (24, 28, 29). However,evidence is now emerging for the participation of the Flt-1receptor in endothelial cell proliferation and angiogenesis. Bar-leon et al. (21) demonstrated that VEGF stimulation of Flt-1receptors in cultured endothelial cells enhances DNA synthesis.In addition, Fonget al. (27) demonstrated that Flt-1 receptor
Fig. 6 Representative livers from anti-Flt-1ribozyme- and saline-treated mice. Livers treated with (A) 100 mg/kg/day anti-Flt-1ribozyme or (B)saline were removed and photographed 41 days after splenic inoculation of KM12L4a cells. The anti-Flt-1 ribozyme treatment period lasted from day3 to day 28.
2101Clinical Cancer Research
Research. on November 2, 2018. © 2000 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

mutants result in abnormalities characterized by normal endo-thelial cell differentiation but abnormal three-dimensional ves-sel structure in embryonic blood vessels. Previous results fromour own work with anti-Flt-1and anti-KDRribozymes demon-strated that both Flt-1 and KDR/Flk-1 receptors participate inendothelial cell proliferation and angiogenesis (33).
It is clear that VEGF is involved in tumor growth andmetastasis. Studies have shown that enhanced VEGF expressionin tumors leads to increased tumor growth (16), whereas ther-apies that target VEGF itself reduce tumor growth and neovas-cularization as well as metastasis (8, 17, 18, 42). Thus, it isreasonable to assume that interfering with VEGF receptor func-tion should also impact primary tumor growth and metastasis.Recently, the inhibition of VEGF receptor function has beendemonstrated as an alternative means of cancer therapy. Theexpression of a tyrosine kinase-deficient Flk-1/KDR mutantresulted in reduced blastoma tumor growth and neovasculariza-tion in vivo (43). Strawnet al. (20) have shown that smallmolecule Flk-1/KDR antagonists reduce angiogenesis in a cho-rioallantoic membrane model. Several of these tyrosine kinaseinhibitors are currently in clinical trials for a variety of clinicalindications. Additionally, Shibuyaet al. (19, 44) presentedevidence that the Flt-1 receptor is involved in tumor angiogen-esis. The results from the current study reinforce the hypothesisthat targeting the VEGF/VEGF receptor system is a promisingstrategy for targeting solid tumor growth and metastasis byinterfering with the process of neovascularization.
In addition to potential roles as therapeutics, ribozymestargeting VEGF receptor mRNA can also be used to determinethe relative role(s) of each subtype in signal transduction and ondownstream biological effects. The anti-Flt-1 and anti-KDRribozymes provide unique tools for elucidating the functionalroles of VEGF receptors in the angiogenesis, growth, and me-tastasis of VEGF-expressing tumors.
ACKNOWLEDGMENTSWe thank Dr. Michael O’Reilly for providing the Lewis lung
carcinoma cell line and protocols for the preparation of LLC-HMinoculates and Dr. Jerald Killion for the KM12L4a human colorectalcancer cell line. We also thank Drs. Lawrence Blatt, Thale Jarvis,Jennifer Sandberg, Nassim Usman, and Shawn Zinnen (Ribozyme Phar-maceuticals, Inc.) for technical advice and support and Dr. ErnestBorden (Cleveland Clinic, Cleveland, OH) and Dr. Lea Aukerman(Chiron Corp., Emeryville, CA) for reviewing the manuscript.
REFERENCES1. Folkman, J. Tumor angiogenesis: therapeutic implications. N. Engl.J. Med.,285: 1182–1186, 1971.2. Thomas, K. A. Vascular endothelial growth factor, a potent andselective angiogenic agent. J. Biol. Chem.,271: 603–606, 1996.3. Martiny-Baron, G., and Marme, D. VEGF-mediated tumour angio-genesis: a new target for cancer therapy. Curr. Opin. Biotechnol.,6:675–680, 1995.4. Myoken, Y., Kayada, Y., Okamoto, T., Kan, M., Sato, G. H., andSato, J. D. Vascular endothelial cell growth factor (VEGF) produced byA-431 human epidermoid carcinoma cells and identification of VEGFmembrane binding sites. Proc. Natl. Acad. Sci. USA,88: 5819–5823,1991.5. Plate, K., Breier, G., Weich, H., and Risau, W. Vascular endothelialgrowth factor is a potential tumour angiogenesis factor in human glio-masin vivo. Nature (Lond.),359: 845–848, 1992.
6. Plate, K. H., Breier, G., Millauer, B., Ullrich, A., and Risau, W.Up-regulation of vascular endothelial growth factor and its cognatereceptors in a rat glioma model of tumor angiogenesis. Cancer Res.,53:5822–5827, 1993.
7. Brown, L. F., Berse, B., Jackman, R. W., Tognazzi, K., Guidi, A. J.,Dvorak, H. F., Senger, D. R., Connolly, J. L., and Schnitt, S. J.Expression of vascular permeability factor (vascular endothelial growthfactor) and its receptors in breast cancer. Hum. Pathol.,26: 86–91,1995.
8. Warren, R., Yuan, H., Matli, M., Gillett, N., and Ferrara, N. Regu-lation by vascular endothelial growth factor of human colon cancertumorigenesis in a mouse model of experimental liver metastasis.J. Clin. Investig.,95: 1789–1797, 1995.9. Mattern, J., Koomagi, R., and Volm, M. Association of vascularendothelial growth factor expression with intratumoral microvessel den-sity and tumour cell proliferation in human epidermoid lung carcinoma.Br. J. Cancer,73: 931–934, 1996.10. Nicol, D., Hii, S-I., Walsh, M., Teh, B., Thompson, L., Kennett, C.,and Gotley, D. Vascular endothelial growth factor expression is in-creased in renal cell carcinoma. J. Urol.,157: 1482–1486, 1997.11. Hlatky, L., Tsionou, C., Hahnfeldt, P., and Coleman, C. N. Mam-mary fibroblasts may influence breast tumor angiogenesis via hypoxia-induced vascular endothelial growth factor up-regulation and proteinexpression. Cancer Res.,54: 6083–6086, 1994.12. Shweiki, D., Itin, A., Soffer, D., and Keshet, E. Vascular endothelialgrowth factor induced by hypoxia may mediate hypoxia-initiated angio-genesis. Nature (Lond.),359: 843–845, 1992.13. Takahashi, A., Sasaki, H., Kim, S., Tobisu, K-I., Kakizoe, T.,Tsukamoto, T., Kumamoto, Y., Sugimura, T., and Terada, M. Markedlyincreased amounts of messenger RNAs for vascular endothelial growthfactor and placenta growth factor in renal cell carcinoma associated withangiogenesis. Cancer Res.,54: 4233–4237, 1994.14. Samoto, K., Ikezaki, K., Ono, M., Shono, T., Kohno, K., Kuwano,M., and Fukui, M. Expression of vascular endothelial growth factor andits possible relation with neovascularization in human brain tumors.Cancer Res.,55: 1189–1193, 1995.15. Potgens, A. J. G., Lubsen, N. H., van Altena, M. C., Schoenmakers,J. G. G., Ruiter, D. J., and de Waal, R. M. Vascular permeability factorexpression influences tumor angiogenesis in human melanoma linesxenografted to nude mice. Am. J. Pathol.,146: 197–209, 1995.16. Zhang, H. T., Craft, P., Scott, P. A., Ziche, M., Weich, H. A.,Harris, A. L., and Bicknell, R. Enhancement of tumor growth andvascular density by transfection of vascular endothelial cell growthfactor into MCF-7 human breast carcinoma cells. J. Natl. Cancer Inst.,87: 213–219, 1995.17. Kim, K. J., Li, B., Winer, J., Armanini, M., Gillett, N., Phillips,H. S., and Ferrara, N. Inhibition of vascular endothelial growth factor-induced angiogenesis suppresses tumour growthin vivo. Nature (Lond.),362: 841–844, 1993.18. Melnyk, O., Shuman, M., and Kim, K. Vascular endothelial growthfactor promotes tumor dissemination by a mechanism distinct from itseffect on primary tumor growth. Cancer Res.,56: 921–924, 1996.19. Shibuya, M. Role of VEGF-flt receptor system in normal and tumorangiogenesis. Adv. Cancer Res.,67: 281–316, 1995.20. Strawn, L., McMahon, G., App, H., Schreck, R., Kuchler, W.,Longhi, M., Hui, T., Tang, C., Levitzki, A., Gazit, A., Chen, I., Keri, G.,Orfi, L., Risau, W., Flamme, I., Ullrich, A., Hirth, P., and Shawyer, L.Flk-1 as a target for tumor growth inhibition. Cancer Res.,56: 3540–3545, 1996.21. Barleon, B., Siemeister, G., Martiny-Baron, G., Weindel, K., Her-zog, C., and Marme, D. Vascular endothelial growth factor up-regulatesits receptor fms-like tyrosine kinase 1 (FLT-1) and a soluble variant ofFLT-1 in human vascular endothelial cells. Cancer Res.,57: 5421–5425,1997.22. Jakeman, L. B., Winer, J., Bennett, G. L., Altar, C. A., and Ferrara,N. Binding sites for vascular endothelial growth factor are localized onendothelial cells in adult rat tissues. J. Clin. Investig.,89: 244–253,1992.
2102Anticancer Effects of Ribozymes Targeting Angiogenesis
Research. on November 2, 2018. © 2000 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

23. Ferrara, N., Heinsohn, H., Walder, C. E., Bunting, S., and Thomas,G. R. The regulation of blood vessel growth by vascular endothelialgrowth factor. Ann. N. Y. Acad. Sci.,752: 246–255, 1995.24. Millauer, B., Wizigmann-Voos, S., Schnurch, H., Martinez, R.,Moller, N. P., Risau, W., and Ullrich, A. High affinity VEGF bindingand developmental expression suggest Flk-1 as a major regulator ofvasculogenesis and angiogenesis. Cell,72: 835–846, 1993.25. Waltenberger, J., Claesson-Welsh, L., Siegbahn, A., Shibuya, M.,and Heldin, C. H. Different signal transduction properties of KDR andFlt1, two receptors for vascular endothelial growth factor. J. Biol.Chem.,269: 26988–26995, 1994.26. Barleon, B., Totzke, F., Herzog, C., Blanke, S., Kremmer, E.,Siemeister, G., Marme, D., and Martiny-Baron, G. Mapping of the sitesfor ligand binding and receptor dimerization at the extracellular domainof the vascular endothelial growth factor receptor FLT-1. J. Biol. Chem.,272: 10382–10388, 1997.27. Fong, G. H., Rossant, J., Gertsenstein, M., and Breitman, M. L.Role of the Flt-1 receptor tyrosine kinase in regulating the assembly ofvascular endothelium. Nature (Lond.),376: 66–70, 1995.28. Shalaby, F., Rossant, J., Yamaguchi, T. P., Gertsenstein, M., Wu,X. F., Breitman, M. L., and Schuh, A. C. Failure of blood-islandformation and vasculogenesis in Flk-1-deficient mice. Nature (Lond.),376: 62–66, 1995.29. Shalaby, F., Ho, J., Stanford, W. L., Fischer, K. D., Schuh, A. C.,Schwartz, L., Bernstein, A., and Rossant, J. A requirement for Flk1 inprimitive and definitive hematopoiesis and vasculogenesis. Cell,89:981–990, 1997.30. Usman, N., and Stinchcomb, D. T. Design, synthesis, and functionof therapeutic hammerhead ribozymes.In: R. Eckstein and D. M. J.Lilley (eds.), Nucleic Acids and Molecular Biology, Vol. 10, pp. 243–264. Berlin: Springer-Verlag, 1996.31. Beigelman, L., McSwiggen, J., Draper, K., Gonzalez, C., Jensen,K., Karpeisky, A., Modak, A., Matulic-Adamic, J., DiRenzo, A.,Haeberli, P., Sweedler, D., Tracz, D., Grimm, S., Wincott, F., Thackray,V., and Usman, N. Chemical modification of hammerhead ribozymes.Catalytic activity and nuclease resistance. J. Biol. Chem.,270: 25702–25708, 1995.32. Flory, C. M., Pavco, P. A., Jarvis, T. C., Lesch, M. E., Wincott,F. E., Beigelman, L., Hunt, S. W., III, and Schrier, D. J. Nuclease-resistant ribozymes decrease stromelysin mRNA levels in rabbit syno-vium following exogenous delivery to the knee joint. Proc. Natl. Acad.Sci. USA,93: 754–758, 1996.33. Parry, T. J., Cushman, C., Gallegos, A. M., Agrawal, A. B., Rich-ardson, M., Andrews, L. E., Maloney, L., Mokler, V. R., Wincott, F. E.,and Pavco, P. A. Bioactivity of anti-angiogenic ribozymes targetingFlt-1 andKDR mRNA. Nucleic Acids Res.,27: 2569–2577, 1999.
34. Sioud, M., and Sørensen, D. R. A nuclease-resistant protein kinaseC a ribozyme blocks glioma cell growth. Nat. Biotechnol.,16: 556–561, 1998.35. O’Reilly, M. S., Holmgren, L., Shing, Y., Chen, C., Rosenthal,R. A., Moses, M., Lane, W. S., Cao, Y., Sage, E. H., and Folkman, J.Angiostatin: a novel angiogenesis inhibitor that mediates the suppres-sion of metastases by a Lewis lung carcinoma. Cell,79: 315–328, 1994.36. Folkman, J. Angiogenesis in cancer, vascular, rheumatoid and otherdisease. Nat. Med.,1: 27–31, 1995.37. Wincott, F., DiRenzo, A., Shaffer, C., Grimm, S., Tracz, D., Work-man, C., Sweedler, D., Gonzalez, C., Scaringe, S., and Usman, N.Synthesis, deprotection, analysis and purification of RNA and ri-bozymes. Nucleic Acids Res.,23: 2677–2684, 1995.38. Burgin, A. B., Gonzalez, C., Matulic-Adamic, J., Karpeisky, A. M.,Usman, N., McSwiggen, J. A., and Beigelman, L. Chemically modifiedhammerhead ribozymes with improved catalytic rates. Biochemistry,35: 14090–14097, 1996.39. Clark, J. D., Baldwin, R. L., Bayne, K. A., Brown, M. J., Gebhart,G. F., Gonder, J. C., Gwathmey, J. K., Keeling, M. E., Kohn, D. F.,Robb, J. W., Smith, O. A., Steggerda, J-A. D., Vandenbergh, J. G.,White, W. J., Williams-Blangero, S., and VandeBerg, J. L. Guide for theCare and Use of Laboratory Animals. Washington DC: National Acad-emy Press, 1996.40. Sandberg, J. A., Bouhana, K. S., Gallegos, A. M., Agrawal,A. B., Grimm, S. L., Wincott, F. E., Reynolds, M. A., Pavco, P. A., andParry, T. J. Pharmacokinetics of an antiangiogenic ribozyme(ANGIOZYMEy) in the mouse. Antisense & Nucleic Acid Drug De-velopment,9: 271–277, 1999.41. Keyt, B. A., Nguyen, H. V., Berleau, L. T., Duarte, C. M., Park, J.,Chen, H., and Ferrara, N. Identification of vascular endothelial growthfactor determinants for binding KDR and FLT-1 receptors. Generationof receptor-selective VEGF variants by site-directed mutagenesis.J. Biol. Chem.,271: 5638–5646, 1996.42. Yuan, F., Chen, Y., Dellian, M., Safabakhsh, N., Ferrara, N., andJain, R. K. Time-dependent vascular regression and permeabilitychanges in established human tumor xenografts induced by an anti-vascular endothelial growth factor/vascular permeability factor anti-body. Proc. Natl. Acad. Sci. USA,93: 14765–14770, 1996.43. Millauer, B., Shawver, L. K., Plate, K. H., Risau, W., and Ullrich,A. Glioblastoma growth inhibitedin vivo by a dominant-negative Flk-1mutant. Nature (Lond.),367: 576–579, 1994.44. Shibuya, M., Seetharam, L., Ishii, Y., Sawano, A., Gotoh, N.,Matsushime, H., and Yamaguchi, S. Possible involvement of VEGF-FLT tyrosine kinase receptor system in normal and tumor angiogenesis.Proceedings of the International Symposium Princess Takahatsu CancerResearch Fund,24: 162–170, 1994.
2103Clinical Cancer Research
Research. on November 2, 2018. © 2000 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

2000;6:2094-2103. Clin Cancer Res Pamela A. Pavco, Karyn S. Bouhana, Anna M. Gallegos, et al. Receptorsthe Messenger RNA of Vascular Endothelial Growth Factor Antitumor and Antimetastatic Activity of Ribozymes Targeting
Updated version
http://clincancerres.aacrjournals.org/content/6/5/2094
Access the most recent version of this article at:
Cited articles
http://clincancerres.aacrjournals.org/content/6/5/2094.full#ref-list-1
This article cites 38 articles, 15 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/6/5/2094.full#related-urls
This article has been cited by 20 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/6/5/2094To request permission to re-use all or part of this article, use this link
Research. on November 2, 2018. © 2000 American Association for Cancerclincancerres.aacrjournals.org Downloaded from