aquaaqua-aquapress.com/pdf/aqua11(4).pdf · today known as the rivulidae by garman (1895), until...
TRANSCRIPT

aquaJournal of Ichthyology and Aquatic
BiologyVol. 11 (4), October 2006
AquapressISSN
0945-9871

Managing Editor:
Heiko BleherVia G. Falcone 11,27010 Miradolo Terme (PV), ItalyTel.: +39-0382-754707/08Fax: +39-0382-754129E-mail: [email protected]
Scientific Editor:
Friedhelm KruppCurator of FishesSenckenberg Research Instituteand Natural History MuseumSenckenberganlage 2560325 Frankfurt am Main, GermanyTel: +49-69-7542.255Fax: +49-69-7542.253E-mail: [email protected]
Editorial Board:
Gerald R. AllenDepartment of Aquatic Zoology,Western Australian Museum, Perth, Australia
Nina G. BogutskayaZoological Institute of the Russian Academy of Sci-ences, St. Petersburg, Russia
Wilson J. E. M. CostaLaboratório de Ictiologia Geral e Aplicada,Departamento de Zoologia,Universidade Federal do Rio de Janeiro, Brasil
Axel MeyerLehrstuhl für Zoologie und Evolutionsbiologie,Universität Konstanz, Germany
Paolo ParentiDepartment of Enviromental Sciences,University of Milano-Bicocca,Milan, Italy
Mário de PinnaMuseu de Zoologia da USP, São Paulo, Brazil
John E. RandallBishop Museum, Honolulu, Hawaii, U.S.A.
Richard WinterbottomCentre of Biodiversity & Conservation Biology,Royal Ontario Museum,Toronto, Canada
Scopeaqua is an international journal which publishes originalscientific articles in the fields of systematics, taxonomy,biogeography, ethology, ecology, and general biology offishes. Papers on freshwater, brackish, and marine fisheswill be considered. aqua is fully refereed and aims at pub-lishing manuscripts within 2-4 months of acceptance. Inview of the importance of color patterns in species iden-tification and animal ethology, authors are encouraged tosubmit color illustrations in addition to descriptions ofcoloration. It is our aim to provide the international sci-entific community with an efficiently published journalmeeting high scientific and technical standards.
Call for papersThe editors welcome the submission of original manu-scripts which should be sent in digital format to the scien-tific editor. Full length research papers and short notes willbe considered for publication. There are no page chargesand color illustrations will be published free of charge.Authors will receive one free copy of the issue in whichtheir paper is published and an e-print in PDF format.
Subscription NoticeAt least one volume (4 issues) of aqua is being publishedper year, each issue comprising 48 pages (including cover).The subscription rate (for one volume = 4 issues) isfrom volume 12 on: Personal subscription: Euro 75,00(incl. priority mail); Institutional subscription: Euro 140,00(incl. priority mail). Subscription enquires should be sentto the publisher at the address given below or by e-mailto: [email protected]
aqua binderBinders for Volumes of aqua are available at cost priceEuro 12,50 (US$ 15.00) plus postage Euro 8,00 (US$10.00). Notice: aqua Volumes 1(1)-5(4) = 1st. binder;Volumes 6(1)-9(4) = 2nd. binder; Volumes 10(1)-13(4)= 3rd. binder.
Special PublicationSince 2003 Aquapress publishes a series of Special Publi-cations, which are produced at irregular intervals. All Spe-cial Publications have about 100 or more pages and areavailable separately from regular issues of aqua. Enquiriesabout subscriptions and prices should be sent to the pub-lisher at the address given here above or by e-mail to:[email protected]
ISSN 0945-9871Publisher: Aquapress, Redazione aqua,I-27010 Miradolo Terme (Pavia), ItalyPrinter: SATE s.r.l. (Bergamo), ItalyCopyediting and layout: Rossella Bulla© 2006 aqua, Journal of Ichthyology and Aquatic Biology
aqua - Journal of Ichthyology and Aquatic Biology

aqua vol. 11 no. 4 - 2006133
AbstractRelationships among species of Rivulus endemic to the
Brazilian Amazonas river basin are evaluated through ananalysis of morphological characters, including terminaltaxa representative of all Rivulus lineages. Based on the pre-sent morphological phylogeny and previous molecularanalyses, seven subgenera are recognized: Rivulus, Cyn-odonichthys, Anablepsoides, Benirivulus n. subgen.,Laimosemion, Owiyeye n. subgen., and Melanorivulus n. sub-gen. Benirivulus is diagnosed by a unique colour pattern andincludes a single species, R. beniensis, from the rio Madeiradrainage; Owiyeye is diagnosed by the absence of dermo-sphenotic, a derived pattern of frontal squamation, and acolour pattern of the mandibular region, and includesspecies from the Amazonas and Orinoco river basins;Melanorivulus is diagnosed by an apomorphic morphologyof preopercle, and derived colour patterns of male postor-bital region and female unpaired fins, and includes speciesfrom southern Amazonian tributaries, Paraná-Paraguay riversystem, and Parnaíba and São Francisco river basins. Eigh-teen species from the Amazonas river basin of Brazil are rec-ognized as valid: R. urophthalmus, R. taeniatus, R. micropus,R. ornatus, R. beniensis, R. strigatus, and R. dibaphus, whichare herein redescribed, and R. atratus, R. romeri, R. kirovskyi,R. uatuman, R. uakti, R. amanapira, R. tecminae, R. modes-tus, R. zygonectes, R. violaceus, and R. litteratus, described orredescribed recently. Field data indicate that species of dif-ferent lineages included in the clade comprising Benirivulus,Laimosemion, Owiyeye, and Melanorivulus are found in dif-ferent kinds of aquatic environments, suggesting a divergentevolution for habitat preference.
ResumoAs relações de parentesco entre espécies de Rivulus endêmi-
cas da bacia amazônica brasileira são avaliadas através deuma análise de caracteres morfológicos, incluindo táxonsterminais representativos de todas as linhagens de Rivulus.Com base na presente filogenia morfológica e em análisesmoleculares anteriores, sete subgêneros são reconhecidos:Rivulus, Cynodonichthys, Anablepsoides, Benirivulus n. sub-gen., Laimosemion, Owiyeye n. subgen., e Melanorivulus n.subgen. Benirivulus é diagnosticado por um padrão de col-
orido exclusivo e inclui uma única espécie, R. beniensis, dadrenagem do rio Madeira; Owiyeye é diagnosticado pelaausência de dermosfenótico, um padrão de escamaçãofrontal derivado e um padrão de colorido da regiãomandibular, e inclui espécies das bacias dos rios Amazonas eOrinoco; Melanorivulus é diagnosticado pela morfologiaapomórfica do pré-opérculo e pelos padrões de coloridoderivados de região pós-orbital de macho e de nadadeirasímpares de fêmeas, e inclui espécies dos tributários amazôni-cos meridionais, do sistema de rios Paraná-Paraguai, e baciasdos rios Paranaíba e São Francisco. Dezoito espécies da baciado rio Amazonas do Brasil são reconhecidas como válidas: R.urophthalmus, R. taeniatus, R. micropus, R. ornatus, R.beniensis, R. strigatus e R. dibaphus, que são aqui redescritas,e R. atratus, R. romeri, R. kirovskyi, R. uatuman, R. uakti, R.amanapira, R. tecminae, R. modestus, R. zygonectes, R. vio-laceus e R. litteratus, recentemente descritas ou redescritas.Dados de campo indicam que espécies de distintas lin-hagens, incluídas no clado que compreende Benirivulus,Laimosemion, Owiyeye e Melanorivulus, são encontradas emtipos deferentes de ambientes aquáticos, sugerindo umaevolução divergente para preferência de habitats.
ZusammenfassungDie verwandtschaftlichen Beziehungen der Arten der Gat-
tung Rivulus, die endemisch im Amazonasbecken vorkom-men, werden anhand von morphologischen Merkmalenuntersucht und die Taxa der verschiedenen Linien innerhalbder Gattung abschließend bewertet. Auf der Grundlage dergegenwärtig bekanntenmorphologisch begründeten Abstam-mungsverhältnisse und der vorhandenen genetischen Analy-sen lassen sich sieben Untergattungen unterscheiden: Rivulus,Cynodonichthys, Anablepsoides, Benirivulus n. subgen., Laimo-semion, Owiyeye n. subgen. und Melanorivulus n. subgen. Be-nirivulus lässt sich durch die einmalige Färbung eindeutigerkennen und umfasst nur eine Art: R. beniensis vom RioMadeira und seinem Einzugsgebiet. Owiyeye kann amFehlen des “dermosphenotischen” Knochens, am sekundärenMuster der Frontalbeschuppung und am Farbmuster desUnterkieferbereichs bestimmt werden; zu dieser Untergat-tung zählen Arten vom Amazonas- und vomOrinocobecken.Melanorivulus wird gekennzeichnet durch: apomorphe (evo-
aqua, Journal of Ichthyology and Aquatic Biology
Relationships and taxonomy of the killifish genus Rivulus(Cyprinodontiformes: Aplocheiloidei: Rivulidae) from the Brazilian Amazonas
river basin, with notes on historical ecology
Wilson J. E. M. Costa
Laboratório de Ictiologia Geral e Aplicada, Departamento de Zoologia, Universidade Federal do Rio de Janeiro,Caixa Postal 68049, CEP 21944-970, Rio de Janeiro, Brasil. E-mail: [email protected]
Received: 08 April 06 – Accepted: 15 August 06

aqua vol. 11 no. 4 - 2006 134
lutiv neue) Gestaltung des Präoperculums, abgeleitete Farb-muster der Postorbital-Region bei Männchen und unpaareFlossen bei Weibchen; Arten der südlichen Nebenflüsse desAmazonas, des Paraná-Paraguay-Systems und der Parnaiba-und São-Francisco-Flussbecken gehören dazu. AchtzehnArten des brasilianischen Amazonasbeckens werden als gültiganerkannt: R. urophthalmus, R. taeniatus, R. micropus, R.ornatus, R. beniensis, R. strigatus und R. dibaphus, die hiermitwiederbeschrieben wird, sowie R. atratus, R. romeri, R.kirovskyi, R. uatuman, R. uakti, R. amanapira, R. tecminae, R.modestus, R. zygonectes, R. violaceus und R. litteratus, die in let-zter Zeit neu oder wiederbeschrieben wurden. Nach Frei-landbeobachtungen kommen die Arten verschiedenerAbstammungslinien der Klade mit Benirivulus, Laimosemion,Owiyeye und Melanorivulus in unterschiedlichen aquatischenLebensräumen vor, sodass von einer divergenten Evolutiongemäß dem bevorzugten Habitat auszugehen ist.
RésuméDes affinités entre des espèces de Rivulus endémiques du
bassin brésilien de l’Amazone sont évaluées au moyen d’uneanalyse de caractéristiques morphologiques, incluant des tax-ons terminaux représentatifs de tous les lignages de Rivulus.Sur base de la phylogénie morphologique actuelle et d’analy-ses moléculaires antérieures, sept sous-genres sont identifiés:Rivulus, Cynodonichthys, Anablepsoides, Benirivulus n. sub-gen., Laimosemion, Owiyeye n. subgen. et Melanorivulus n.subgen. Benirivulus se distingue par un patron de colorationunique et comprend une seule espèce, R. beniensis, du systèmedu Rio Madeira; Owiyeye est distinct par l’absence de dermo-sphénotique, un patron dérivé d’écailles frontales, et unpatron de coloration de la région mandibulaire, et inclut desespèces des bassins de l’Amazone et de l’Orénoque; Melano-rivulus se distingue par une morphologie apomorphique dupré opercule, des patrons de colorations dérivés de la régionpostorbitale du mâle et des nageoires impaires de la femelle etcompte des espèces des tributaires sud de l’Amazone, du sys-tème du Paraná-Paraguay, du bassin du Paranaíba et du SãoFrancisco. Dix-huit espèces du bassin brésilien de l’Amazonesont reconnues: R. urophthalmus, R. taeniatus, R. micropus, R.ornatus, R. beniensis, R. strigatus et R. dibaphus, qui sontredécrits ici, et R. atratus, R. romeri, R. kirovskyi, R. uatuman,R. uakti, R. amanapira, R. tecminae, R. modestus, R. zygonectes,R. violaceus et R. litteratus, décrits ou redécrits récemment.Des donnés de terrain révèlent que des espèces de différentslignages faisant partie du clade comprenant Benirivulus,Laimosemion, Owiyeye et Melanorivulus se trouvent dansdiverses sortes d’environnements aquatiques, suggérant uneévolution divergente en matière de choix d’habitat.
SommarioI rapporti di parentela tra le specie del genere Rivulus
endemiche della regione amazzonica del Brasile sono stativalutati con un’analisi dei caratteri morfologici, includendotaxa rappresentativi di tutte linee evolutive di Rivulus. Sullabase di questa analisi filogenetica e di precedenti filogenesimolecolari sono riconosciuti sette sottogeneri: Rivulus, Cyn-odonichthys, Anablepsoides, Benirivulus n. subgen., Laimo-
semion, Owiyeye n. subgen. e Melanorivulus n. subgen. Beni-rivulus è diagnosticato dalla singolare colorazione e com-prende una sola specie, R. beniensis, presente nel bacino idro-grafico del rio Madeira; Owiyeye è diagnosticato per l’assenzadel dermosfenotico, per un modello derivato di disposizionedelle squame frontali e per la colorazione della regionemandi-bolare, e include specie dei bacini del Rio delle Amazzoni edell’Orinoco; Melanorivulus è diagnosticato da una mor-fologia apomorfica del preopercolo e una colorazione derivatadella regione postorbitale del maschio e delle pinne imparidelle femmine, e include specie diffuse nei tributari merid-ionali del Rio delle Amazzoni, e nei bacici del Paraná-Paraguay, del Parnaíba e del São Francisco. Diciotto specie so-no riconosciute come valide nel bacino amazzonico brasil-iano: R. urophthalmus, R. taeniatus, R. micropus, R. ornatus, R.beniensis, R. strigatus e R. dibaphus, che sono qui ridescritte, eR. atratus, R. romeri, R. kirovskyi, R. uatuman, R. uakti, R.amanapira, R. tecminae, R. modestus, R. zygonectes, R. violaceuse R. litteratus, che sono state descritte o ridescritte recente-mente. Dati di campionamento indicano che le specie dellediverse linee evolutive incluse nell’unità cladistica compren-dente Benirivulus, Laimosemion, Owiyeye e Melanorivulus so-no diffuse in varie tipologie di ambiente acquatico, sugge-rendo un’evoluzione divergente nella preferenza dell’habitat.
IntroductionRivulus Poey, 1860 is the most geographically wide-
spread and diversified genus of Neotropicalaplocheiloid killifishes (e.g. Costa 2003a). It is alsothe most taxonomically problematic genus ofNeotropical aplocheiloids, in spite of recent efforts toclarify the taxonomic status of included species,among which many are still poorly defined. Taxo-nomic problems are due to: great number ofincluded species (over 100 nominal species) in a widegeographic range (from Mexico, about 20° N, toArgentina, about 30° S, including Caribbeanislands), making it difficult to develop taxonomicstudies encompassing comparative material from allregions; brief descriptions, often including only a fewcharacters insufficient to diagnose taxa; bad preserva-tion of type specimens; equivocal synonymies (e.g.Garman 1895); and, unknown data on colour pat-terns of live specimens in older descriptions.Rivulus was erected to include a single species from
Cuba, R. cylindraceus Poey, 1860 (Poey 1860). Sincethen, over 120 nominal species were incorporatedinto Rivulus, some subgenera were described, andsome species groups were diagnosed (e.g. Hoede-man 1959; Huber 1992; Costa 1995a, 2003a).Some studies have been directed to the taxonomy ofRivulus in some regularly and recently explored zoo-geographical areas, including: Central America (e.g.Bussing 1980; Etzel & Berkenkamp 1992;
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin

aqua vol. 11 no. 4 - 2006135
Berkenkamp & Etzel 1992 1993a, 1993b, 1995,1997, 1999), Guianas (Hoedeman 1959; Huber1979, 1991; Vermeulen & Isbrücker 2000), ríoOrinoco basin (Thomerson et al. 1991a, 1991b,1992; Lasso et al. 1992; Thomerson & Taphorn1992, 1993), Peruvian Amazon (e.g. Fels & Huber1985; Fels & de Rham 1982), and central andsouth-eastern Brazil (e.g. Seegers 1984; Costa 1989,1991, 1995a, 2003c, 2004g, 2005a; Costa & Brasil1991). However, species of Rivulus endemic to theBrazilian Amazon have received little attention,except for recent descriptions of new species fromthe rio Negro drainage and near localities (e.g. Costa2003b, 2004c, 2004d, 2004e). Among species ofRivulus from the Brazilian Amazon are some of theoldest species names presently assigned to the genus,therefore making them important keys in solvingtaxonomic problems.The only genus today considered to be a synonym
of Rivulus is Cynodonichthys Meek, 1904, which wasdescribed to include C. tenuis Meek from Mexico(Meek 1904). Cynodonichthys has been placed in thesynonymy of Rivulus since Regan (1907). Vomerivu-lus Fowler, 1944 was proposed as a new monotypicsubgenus of Rivulus based on the possession ofvomerine teeth in its type species, R. leucurus Fowler,1944, from the río Juradó, Colombia (Fowler1944). However, possession of vomerine teeth is aplesiomorphic condition in rivulids, occurring inmost aplocheiloids, including R. cylindraceus, thetype species of Rivulus (e.g. Costa 1998a, 1998b).Hoedeman (1959, 1961), based on combinations
of morphological features such as presence of caudalspot, details of colour patterns, maximum adult size,frontal squamation pattern, and number of longitu-dinal scales, divided Rivulus into species groups.Huber (1992) adopted this classification, recogniz-ing a total of 18 assemblages (i.e., superspecies),besides describing a new monotypic subgenus,Anablepsoides Huber, 1992, for R. atratus Garman,1895. Subsequently, two Huber’s superspecies wereformally recognized as new subgenera, OditichthysHuber, 1999 and Laimosemion Huber, 1999 (Huber1999). Some species groups have been consistentlysupported both by osteological (Costa 1998a) andmolecular data (Murphy & Collier 1996, Murphy etal. 1999, Hrbek & Larson 1999, Hrbek et al. 2004).Since the first taxonomic revision of the species
grouped into the cyprinodontiform assemblagetoday known as the Rivulidae by Garman (1895),until recent years (Parenti 1981), Rivulus wasdefined by a combination of plesiomorphic mor-
phological features. In other words, Rivulus wasdiagnosed by the absence of the distinctive features(i.e., apomorphies) that diagnose the other rivulidgenera (e.g. long fins in Pterolebias Garman, 1895,vs. short fins in Rivulus; numerous dorsal- and anal-fin rays in Cynolebias Steindachner, 1876, vs. fewrays in Rivulus) (Garman 1895, Regan 1912, Myers1927). In addition, Rivulus contained species livingin perennial aquatic biotopes, whereas the remain-ing rivulids comprised species inhabiting seasonalpools.Weitzman & Wourms (1967) suggested that
rivulids were derived from a Rivulus-like ancestor.Parenti (1981), based on morphological characters,provided the first phylogenetic analysis of rivulids,in which Rivulus was considered a paraphyleticgroup with their nominal species included at least intwo basal rivulid lineages. Among the five nominalspecies of Rivulus examined in Parenti’s analysis, R.stellifer Thomerson & Turner, 1973 (transferred toRachovia Myers by Costa 1998a) and R. hartii(Boulenger, 1890) were placed in an assemblagetermed “Rivulus”, that would be more closely relatedto all other rivulid genera (e.g. Cynolebias;Pterolebias; Trigonectes Myers, 1925) than to Rivulus,which would include the type species R. cylindraceus,and R. marmoratus Poey, 1880 (transferred to Kryp-tolebias Costa, 2004 by Costa 2004a) and R. tenuis(Meek, 1904), among the species analysed in thisstudy. Consequently, Rivulus was considered to bethe sister group to a clade including “Rivulus” and allother rivulids, which was diagnosed by an apomor-phic cartilaginous interhyal (vs. ossified in Rivulus)and a derived number of pelvic-fin rays (seven in“Rivulus” and the remaining rivulids, vs. six in Rivu-lus). However, Parenti (1981) did not find synapo-morphies corroborating the monophyly of Rivulusor of “Rivulus”.Costa (1990), in a phylogenetic analysis based on
morphological and behavioural traits, confirmed thepresence of an ossified interhyal in some species thenplaced in Rivulus, but demonstrated that the num-ber of seven pelvic-fin rays is not informative todefine a rivulid clade. Rivulus was considered to be amonophyletic group based on the morphology ofthe shoulder girdle and the neural prezygapophysesof caudal vertebrae. A similar general phylogenetichypothesis among rivulid lineages was obtained byMurphy & Collier (1997) using mitochondrialgenes of seven rivulid species, but monophyly ofRivulus was not properly tested since only twospecies of Rivulus were included in the analysis.
Wilson J. E. M. Costa

aqua vol. 11 no. 4 - 2006 136
Costa (1998a), in a subsequent phylogenetic studyof rivulids, including new taxa and re-evaluatingpreviously described morphological and behaviouralcharacters, noted that, in fact, R. cylindraceus, thetype species of Rivulus, not available in the previousstudy, has a cartilaginous interhyal. Among the newcharacters analysed, Costa (1998a) found thatspecies of Rivulus with an ossified interhyal, i.e. R.brasiliensis (Valenciennes, 1821), R. caudomarginatusSeegers, 1984, and R. ocellatus Hensel, 1868, all latertransferred to Kryptolebias by Costa (2004a) alsoexhibit other putative plesiomorphic conditions,and thus monophyly of Rivulus was weakly sup-ported. Subsequent molecular analyses (Murphy etal. 1999, Hrbek & Larson 1999, Hrbek et al. 2004)indicated Rivulus as a paraphyletic group, with dif-ferent assemblages as successive sister groups to aclade including annual fish genera.Costa (2004a) described Kryptolebias to include
species previously placed in Rivulus with an osseousinterhyal, among other morphological features.Kryptolebias was considered to be the sister group toa clade including Rivulus and all other rivulids (Costa2004a, 2004b, 2004c), a hypothesis supported bymolecular data (Vermeulen & Hrbek 2005).Since the description of Kryptolebias and the con-
sequent transference of some species of Rivulus toKryptolebias, a re-evaluation of the generic place-ment of species maintained in Rivulus became nec-essary, as well as a reassessment of morphologicalcharacters employed to define subgroups of Rivulus.The objectives of the present study are: to testhypotheses of relationships among species of Rivulusfrom the Brazilian Amazon to other rivulids, basedon morphological characters discussed and illus-trated in previous studies (e.g. Costa 1990, 1998,2004, 2005b); to diagnose and to update data onthe taxonomy and distribution of species of Rivulusfrom the Brazilian Amazonas basin, providingredescriptions of poorly known species; to re-evalu-ate morphological diagnoses for nominal subgeneraand to purpose a new classification in order to for-mally accommodate hypotheses derived from recentphylogenetic studies.
Material and methodsMaterial pertinent to the taxonomic study on
Rivulus from the Brazilian Amazonas river basin islisted in the taxonomical accounts below. Data onmaterial is given in the following order: cataloguenumber, number of specimens; country: majorpolitical division: locality; collectors, and date of col-
lection. Acronyms for institutions are: ANSP, Acad-emy of Natural Sciences, Philadelphia; BMNH,TheNatural History Museum, London; CAS, CaliforniaAcademy of Sciences, San Francisco; CBF, Collec-ción Boliviana de Fauna, Museo Nacional de Histo-ria Natural, La Paz; FMNH, Field Museum of Nat-ural History, Chicago; INPA, Instituto Nacional dePesquisas da Amazônia, Manaus; MCNG, Museode Ciencias Naturales de la Universidad NacionalExperimental de los Llanos Occidentales, Guanare;MCP, Museu de Ciências e Tecnologia da PontifíciaUniversidade Católica do Rio Grande do Sul, PortoAlegre; MCZ, Museum of Comparative Zoology,Cambridge; MHNC, Museo Nacional de HistoriaNatural, La Havana; MNRJ, Museu Nacional, Uni-versidade Federal do Rio de Janeiro, Rio de Janeiro;MZUSP, Museu de Zoologia, Universidade de SãoPaulo, São Paulo; NMW, NaturhistorischesMuseum Wien, Vienna; UFPB, Universidade Fed-eral da Paraíba, João Pessoa; UFRJ, Instituto deBiologia, Universidade Federal do Rio de Janeiro,Rio de Janeiro; UMMZ, University of Michigan,Museum of Zoology, Ann Arbor; UNT, Universi-dade Federal do Tocantins, Porto Nacional; USNM,National Museum of Natural History (formerUnited States National Museum), SmithsonianInstitution, Washington; ZFMK, ZoologischesFarschungsinstitut und Museum Alexander Koenig,Bonn; and, ZUEC, Departamento de Zoologia daUniversidade Estadual de Campinas, Campinas.Geographic entities are presented by their regionalnames (e.g. rio or río, igarapé, lago), which are notcapitalized following Portuguese and Spanish gram-mars; this practice is important to avoid Englishstyle and generalizations when translating SouthAmerican localities, which become data inaccurateand may incur errors (see also Menezes &Weitzman1990, and Reis 1997 for further justification for thiswidely employed practice). EPA, that appears as col-lectors of some material, means Expedição Perma-nente da Amazônia, a cooperative project betweenMZUSP, INPA and Museu Paraense Emilío Goeldi,Belém; NEAMB means Núcleo de Estudos Ambi-entais da Universidade Federal do Tocantins. Theremaining material of Aplocheiloidei taxa examinedin the present study is listed in the Appendix 1.Measurements and counts follow Costa (1995b).
Measurements are presented as percentages of stan-dard length (SL), except for those related to headmorphology, expressed as percentages of headlength. Fin-ray counts include all elements. Numberof vertebrae, gill rakers, and caudal-fin rays were
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin

aqua vol. 11 no. 4 - 2006137
recorded only from cleared and stained specimens.The compound caudal centrum was counted as asingle element. Osteological preparations were madeaccording to Taylor & Van Dyke (1985). Terminol-ogy for frontal squamation (i.e. A-H) followsHoedeman (1958); determination of frontal squa-mation patterns (i.e. D, E, F) is according to Hoede-man (1958) (i.e. frontal scale with all margins free),except S-pattern (i.e. a scale just posterior to snoutwith all margins free) which is according to Huber(1992). Terminology for cephalic neuromast seriesfollows Costa (2001).The phylogenetic analysis is according to the cladis-
tic methodology applied to morphological characters(e.g. Kitching et al. 1998, Wiens 2000). Terminaltaxa were species of Rivulus occurring in the Brazil-ian Amazonas basin, species of Rivulus representingall main lineages from all zoogeographic regionsinserted in the geographic area of the genus, andspecies of rivulid genera representing lineages closelyrelated to Rivulus according to recent phylogeneticstudies (Costa 1998a, Murphy et al. 1999, Hrbek &Larson 1999): Aphyolebias boticarioi Costa, 2004,Gnatholebias hoignei (Thomerson, 1974), Micro-moema xiphophora (Thomerson & Taphorn, 1992),Moema apurinan Costa, 2004, Neofundulusparaguayensis (Eigenmann & Kennedy, 1903),Pituna compacta (Myers, 1927), Rachovia maculipin-nis (Radda, 1964), Rachovia stellifer, Renova oscariThomerson & Taphorn, 1995, and Trigonectesrubromarginatus Costa, 1990. Four levels of out-groups were included: Prorivulus auriferus Costa,Lima & Suzart, 2004, and Kryptolebias brasiliensis,representing the successive most basal lineages of theRivulidae (Costa 2004a, 2004c); Scriptaphyosemionguignardi (Romand, 1981) of the Nothobranchidae,the sister group of Rivulidae (Murphy & Collier1997, Costa 2004a); and, Aplocheilus panchax(Hamilton-Buchanan, 1822) of the Aplocheilidae,the sister group of Nothobranchidae plus Rivulidae(Murphy & Collier 1997, Costa 2004a).Characters were obtained from previous phyloge-
netic studies (e.g. Parenti 1981; Costa 1990, 1998a,2005b).Character states of multi-state characterswere treated as ordered whenever possible. Bound-aries between states of quantitative characters weretaken from Costa (2005b), which were determinedto accommodate the best estimate of range observedfor each terminal species included in that category(i.e. character state). All characters and characterstates used in the phylogenetic analysis are listed andcoded in the Appendix 2 and are organized in the
data matrix of Appendix 3. Most parsimoniouscladograms, consistency indices (CI) and retentionindices (RI) were obtained using the algorithmmhennig*;bb* of the program Hennig86 (Farris1988). TreeGardener 2.2.1 (Ramos 1996) was usedfor optimisation of character state changes in thestrict consensus tree of most parsimonious clado-grams, using ACCTRAN. Bootstrap analysis(Felsenstein 1985) was used to establish nodal sup-port through the simple heuristic algorithm ofPAUP 4.0 (Swoford 1998) with 1000 replicates.
Results
Phylogenetic analysisThe phylogenetic analysis is based on a re-exami-
nation of previously reported, discussed and illus-trated morphological characters (e.g. Parenti 1981;Costa 1990, 1998a, 2005b), which are listed in theAppendix 2. Their distribution among taxa exam-ined appears in the data matrix (Appendix 3).Six most parsimonious cladograms were found
(tree length = 364, CI = 0.42, RI = 0.72). They dif-fer only in the position of R. igneus, (i.e., as sister toR. hartii or to R. amphoreus Huber, 1979), and R.uatuman Costa, 2004 (i.e., as sister to R. romeriCosta, 2003 or to R. kirovskyi Costa, 2004). Thestrict consensus tree (three length = 367, CI = 0.41,RI = 0.71) is illustrated in Fig. 1.
Taxonomic accountsRivulus is here formally divided into seven subgen-
era, which are supported both by morphological(the present analysis) and molecular data (e.g. Mur-phy et al. 1999, Hrbek & Larson 1999; see discus-sion below). Subgenera of Rivulus and species occur-ring in the Brazilian Amazonas basin are listed anddiagnosed below. Diagnoses of subgenera aresynapomorphies obtained from the above phyloge-netic analysis; numbers in parentheses refer to char-acters states listed in Appendix 2.
Rivulus Poey, 1860
Rivulus Poey, 1860: 299 (type species: Rivulus cylin-draceus Poey; type by monotypy).
Subgenus Rivulus Poey, 1860
Rivulus Poey, 1860: 299 (type species: Rivulus cylin-draceus Poey; type by monotypy).
Wilson J. E. M. Costa

aqua vol. 11 no. 4 - 2006 138
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
Fig. 1. Strict consensus of six equally parsimonious shortest phylogenies among 47 species of the Rivulidae (L = 367, CI =0.41, RI = 0.71). Numbers above branches are bootstrap values.

aqua vol. 11 no. 4 - 2006139
DiagnosisDistinguished from all other congeners by the pos-
session of the following apomorphic features: hypu-rals completely fused to form a single plate (45.3),about 60% of the anterior portion of the caudal fincovered by scales (82.2), four neuromasts on theanterior supraorbital series (85.1), and a dark metal-lic blue humeral blotch in male.
Included speciesR. cylindraceus Poey, 1860, R. insulaepinorum De La
Cruz & Dubitsky, 1976 and R. roloffi Roloff, 1938.
DistributionRivers basins of Cuba and Republica Dominicana.
Subgenus Cynodonichthys Meek, 1904
Cynodonichthys Meek, 1904: 98 (type species: Cyn-odonichthys tenuis Meek; type by original designa-tion).
Vomerivulus Fowler, 1944: 244 (type species: Rivulusleucurus Fowler; type by original designation).
Oditichthys Huber, 1999: 49 (type species: Rivulusigneus Huber; type by original designation).
DiagnosisTentatively diagnosed by the presence of yellow
stripes on dorsal and ventral margins of caudal fin inmale (101.1, 102.1), a condition inconspicuous orabsent in some members of Cynodonichthys (i.e.reversals), but also exhibited by some species ofOwiyeye (i.e. homoplasies).
Included speciesRivulus amphoreus Huber, 1979, R. bahianus
Huber, 1990, R. birkhahni Berkenkamp & Etzel,1992, R. boehlkei Huber & Fels, 1985, R. brunneusMeek & Hildebrand, 1913, R. christinae Huber,1992, R. chucunaque Breder, 1925, R. cryptocallusSeegers & Huber, 1981, R. deltaphilus Seegers,1983, R. depressus Costa, 1991, R. derhami Fels &Huber, 1985, R. elegans Steindachner, 1880, R. elon-gatus Fels & de Rham, 1981, R. erberi Berkenkamp,1989, R. frommi Berkenkamp & Etzel, 1993, R. fus-colineatus Bussing, 1980, R. glaucus Bussing, 1980,R. haraldsiolii Berkenkamp, 1984, R. hartii(Boulenger, 1890), R. hildebrandi Myers, 1927, R.holmiae Eigenmann, 1909, R. igneus Huber, 1991,R. immaculatus Thomerson, Nico & Taphorn,1991, R. intermittens Fels & de Rham, 1981, R. iri-
descens Fels & de Rham, 1981, R. isthmensis Gar-man, 1895, R. janeiroensis Costa, 1991, R. jucundusHuber, 1992, R. kuelpmanni Berkenkamp & Etzel,1993, R. lanceolatus Eigenmann, 1909, R. leucurusFowler, 1944, R. limoncochae Hoedeman, 1962, R.luelingi Seegers, 1984, R. lungi Berkenkamp, 1984,R. magdalenae Eigenmann & Henn, 1916, R.mazaruni Myers, 1924, R. micropus (Steindachner),1863, R. monikae Berkenkamp & Etzel, 1995, R.monticola Staeck & Schindler, 1997, R. montiumHildebrand, 1938, R. nudiventris Costa & Brasil,1991, R. ophiomimus Huber, 1992, R. pacificusHuber, 1992, R. peruanus (Regan, 1903), R.rubripunctatus Bussing, 1980, R. rubrolineatus Fels& de Rham, 1981, R. santensis Köhler, 1906, R.siegfriedi Bussing, 1980, R. simplicis Costa, 2004, R.speciosus Fels & de Rham, 1981, R. stagnatus Eigen-mann, 1909, R. sucubti Breder, 1925, R. taeniatusFowler, 1945, R. tenuis (Meek, 1904), R. tessellatusHuber, 1992, R. uroflammeus Bussing, 1980, R.urophthalmus Günther, 1866, R. villwockiBerkenkamp & Etzel, 2002, R. waimacui Eigen-mann, 1909, R. wassmanni Berkenkamp & Etzel,1999, R. weberi Huber, 1992, and R. xanthonotusAhl, 1926.
DistributionRiver basins of Mesoamerica (between Mexico
and Panama, and southern Caribbean islands), andnorthern (Colombia, Venezuela, Guianas, Peru,Ecuador, Bolivia and northern Brazil) and easternSouth America (coastal plains of south, south-east-ern and north-eastern Brazil).
Rivulus urophthalmus Günther, 1866(Figs. 2-9; Table I)
Rivulus urophthalmus Günther, 1866: 327 (typelocality: Para [Belém, formerly often known asPará, northern Brazil]; lectotype: BMNH1851.12.26, designated by Huber 1991:67; photo-graph of lectotype in Huber 1991: 72, fig. 7).
Rivulus poey Steindachner, 1876: 165 (type locality:canals in the city of Para [now Belém]; typesunknown; neotype: UFRJ 6282, male, 34.9 mmSL; creek at 15 de agosto street, Icoaraci, Belém;collected by A. Sarraf & M. Mourão Jr., 24 Jul.1996 [herein designated]).
Rivulus urophthalmus var. aurata Schreitmüller,1928: 88 (Maharankao [correctly Maranhão],Brazil; type: not designated).
Wilson J. E. M. Costa

aqua vol. 11 no. 4 - 2006 140
Material examinedBrazil: Estado do Pará: UFRJ 6247, 18; stream in
praia de Ariramba, ilha do Mosqueiro, Belém,1°6’34.7”S 48°25’32.7”W; W. J. E. M. Costa, R.Paiva & D. Ramos, 21 Jun. 2004. UFRJ 6248, 11;stream in ilha do Mosqueiro, Belém, 1°9’27.4”S48°20’55.4”W; W. J. E. M. Costa, R. Paiva & D.Ramos, 21 Jun. 2004. UFRJ 6254, 8; UFRJ 6261, 3;stream crossing the road PA-391, ilha do Mosqueiro,Belém, 1°8’59.5”S 48°21’27.9”W;W. J. E.M. Costa,R. Paiva&D. Ramos, 21 Jun. 2004. UFRJ 6255, 11;creek near the road PA-124, near Santa Luzia,0°46’39.1”S 47°16’43.5”W; W. J. E. M. Costa, R.Paiva & D. Ramos, 19 Jun. 2004. UFRJ 6256, 5;pools near stream near the road PA-446, Primavera,0°54’8.9”S 47°2’31.0”W; W.J.E.M. Costa, R. Paiva& D. Ramos, 18 Jun. 2004. UFRJ 6257, 9; swampat Quatipuru, 0°52’12.4”S 47°0’23.6”W;W. J. E.M.Costa, R. Paiva & D. Ramos, 18 Jun. 2004. UFRJ
6258, 15; stream near the road PA-124, near Pindo-rama, Salinópolis, 0°42’28.1”S 47°18’5.6”W; W. J.E. M. Costa, R. Paiva & D. Ramos, 18 Jun. 2004.UFRJ 6259, 4; UFRJ 6290, 34; stream at forest edge,road PA-446, Primavera, 0°56’23.9”S 47°5’21.6”W;W. J. E. M. Costa, R. Paiva & D. Ramos, 19 Jun.2004. UFRJ 4087, 9 (c&s); igarapé Maguary, Belém;A. Sarraf & E. Nogueira, 1 Aug. 1996. UFRJ 3055,6 (c&s); Primavera; A. Werner, May 1988. UFRJ4082, 6; campus of Universidade Federal do Pará,Belém; A. Sarraf & M. Mourão Jr., 30 Jul. 1996.UFRJ 4083, 4; creek at 15 de agosto street, Icoaraci,Belém; A. Sarraf & E. Nogueira, 1 Aug. 1996. UFRJ4079, 5; igarapé Maguary, Belém; A. Sarraf & M.Mourão Jr., 24 Jul. 1996. Estado doTocantins: UFRJ5149, 9; floodplains of left bank of rio Tocantins,about 1200 m from the river margin, SSE Sampaio,5º23’00”S 47°51’41”W; G. C. Brasil, 8 Jun. 2000.UNT 553, 3; rio Brejão, road BR-153, Araguaina; E.
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
Fig. 3. Rivulus urophthalmus, UFRJ 6261, male, 26.7 mmSL (two days after collection); Brazil: Pará: Belém (samepopulation as Fig. 3, with slightly different colour patternson the caudal fin). Photo by W. J. E. M. Costa.
Fig. 4. Rivulus urophthalmus, UFRJ 6261, female, 30.0mm SL (two days after collection); Brazil: Pará: Belém.Photo by W. J. E. M. Costa.
Fig. 2. Rivulus urophthalmus, UFRJ 6261, male, 27.1 mmSL (two days after collection); Brazil: Pará: Belém. Photoby W. J. E. M. Costa.
Fig. 6. Rivulus urophthalmus, UFRJ 6256, male, 26.6 mmSL (one day after collection); Brazil: Pará: Primavera (com-pare with Fig. 5 to observe different colour patterns on thecaudal peduncle). Photo by W. J. E. M. Costa.
Fig. 7. Rivulus urophthalmus, UFRJ 6259, female, 33.2mm SL (one day after collection); Brazil: Pará: Primavera.Photo by W. J. E. M. Costa.
Fig. 5. Rivulus urophthalmus, UFRJ 6259, male, 30.1 mmSL (one day after collection); Brazil: Pará: Primavera.Photo by W. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006141
L. Beerli & L. M. Lima, 22 Nov. 2003. UNT 554, 5;river at the road BR-153, between Filadelphia andAraguaina; E. L. Beerli, 2 Nov. 2003. Estado doAmapá: INPA 2244, 26 (3 c&s); stream belowcachoeira Santo Antônio, rio Jari drainage; M. Jegu& J. Zuanon, 21 Jun. 1987. Estado do Maranhão:UFRJ 5152, 16; swamp about 500 m from the rightmargin of rio Tocantins, 5°12’33”S 48°27’59”W; G.C. Brasil, 8 Jun. 2000. UFRJ 6293, 4; Olho d’Águadas Cunhãs; A. C. De Luca, C. G. C. Wenceslau &M. A. Santos, 7 Jul. 2003.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anteriorportion of trunk slightly deeper than wide, jawsshort, snout blunt, tip of anal fin rounded in male,caudal fin rounded in male, pelvic-fin tip notreaching anus in male, dorsal-fin origin on verticalbetween base of last 3rd and 4th anal-fin rays, dor-sal-fin rays 7-8, anal-fin rays 12-14, frontal squa-mation E-patterned, frontal scales arranged circu-larly, canal preopercular short and opened, contactorgans absent, longitudinal series of scales 37-40,gill rakers of first branchial arch 1-2 + 8, red stripeson flank, jaws not distinctively pigmented, andround black spot on dorsal portion of caudal fin infemale.
Wilson J. E. M. Costa
Fig. 9. Rivulus urophthalmus, UFRJ 6264, female, 28.9mm SL (one day after collection); Brazil: Pará: Altamira.Photo by W. J. E. M. Costa.
Fig. 10. Geographic distribution of: Rivulus modestus (dot), R. romeri (inverted triangle), R. taeniatus (stars), R. urophthalmus(triangles), and R. violaceus (lozenges). One symbol may represent more than one collecting site.
Fig. 8. Rivulus urophthalmus, UFRJ 6264, male, 29.5 mmSL (one day after collection); Brazil: Pará: Altamira. Photoby W. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006 142
DescriptionMorphometric data appear in Table I. Largest spec-
imen, a female 43.9 mm SL. Dorsal profile gentlyconvex from snout to end of dorsal-fin base, approx-imately straight on caudal peduncle. Ventral profileslightly convex from lower jaw to end of anal-finbase, about straight on caudal peduncle. Body slen-der, subcylindrical anteriorly, slightly deeper thanwide, to compressed posteriorly. Greatest body depthat vertical just in front to pelvic-fin base. Jaws short,snout blunt.Tip of dorsal and anal fins rounded. Caudal fin
rounded. Pectoral fin short and rounded, posteriormargin reaching vertical at about 60% of lengthbetween pectoral-fin and pelvic-fin bases. Pelvic finshort and elliptical, tip not reaching anus. Pelvic-finbases medially separated by short interspace. Dorsal-fin origin on vertical between base of 3rd and 4th lastanal-fin rays, and between neural spines of 21st and23rd vertebrae. Anal-fin origin between pleural ribsof 15th and 17th vertebrae. Dorsal-fin rays 7-8; anal-fin rays 12-14; caudal-fin rays 25-28; pectoral-finrays 13; pelvic-fin rays 6.Scales small, cycloid. Body and head entirely
scaled, except anterior ventral surface of head. Bodysquamation extending over anterior 30 % of caudal-fin base; no scales on dorsal- and anal-fin bases.Frontal squamation E-patterned; E-scales not over-lapping medially; scales arranged in regular circularpattern around A-scale without exposed margins.Longitudinal series of scales 37-40; transverse seriesof scales 8; scale rows around caudal peduncle 16.Contact organs absent.Cephalic neuromasts: supraorbital 3 + 3, parietal 1,
anterior rostral 1, posterior rostral 1, infraorbital 1 +1 + 11-15 + 1, preorbital 2-3, otic 1, post-otic 2,supratemporal 1, median opercular 1, ventral oper-cular 2, preopercular 2 + 4, mandibular 4 + 1, lateralmandibular 2-3.Interhyal minute, cartilaginous. Rostral cartilage
longer than wide, width about 80% length. Basihyalsubtriangular, width about 65% length; basihyalcartilage about 10% basihyal length. Six bran-chiostegal rays. Second pharyngobranchial teeth 3-4. Gill rakers of first branchial arch 1-2 + 8. Vomer-ine teeth 5-6. Ventral process of posttemporal short.Total vertebrae 32-35.Coloration in life: Male: Side of body metallic yel-
lowish green to metallic purplish blue ventrally;eight longitudinal rows of red to reddish browndots, often in close proximity, forming stripes; usu-ally three, sometimes four or five rows of red dots
crossing caudal peduncle and reaching caudal-finbase, remaining rows reaching vertical throughanal-fin base; dark chromatophores concentratedon longitudinal zone between flank and dorsum;sometimes dark grey bars on flank anterior to cau-dal peduncle, when fish exposed to strong sunlight.Dorsum light brown. Venter white. Side of headmetallic yellowish green to metallic purplish blue,sometimes dark grey when fish exposed to strongsunlight. Lower jaw light brown. Iris light yellowishto greenish brown. Dorsal fin pale yellow withbrown dots on basal portion, often with bright yel-low zone. Anal fin yellow, basal portion sometimesblue, often with brown or red dots. Caudal fin paleyellow, often with brown dots and dorsal and ven-tral bright yellow zones, sometimes dorsal and ven-tral margins bright orange. Paired fins yellowishhyaline.Female: Side of body light brown, with eight lon-
gitudinal rows of reddish brown dots, often in closeproximity, forming stripes, alternating with longitu-dinal rows of pale golden dots; dark chromatophoresconcentrated on longitudinal zone between flankand dorsum; sometimes dark grey bars on flankanterior to caudal peduncle, when fish exposed tostrong sunlight. Dorsum light brown. Venter white.Side of head pale metallic yellowish green to metal-lic purplish blue, sometimes dark grey when fishexposed to strong sunlight. Lower jaw light brown.Iris light yellowish to greenish brown. Unpaired finsyellowish hyaline with brown dots; round black spoton dorsal portion of caudal fin usually reaching finmargin, anteriorly bordered by small, triangular paleyellow spot. Paired fins hyaline.
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
Fig. 11. Brazil: Pará: Primavera; pool close to creek nearforest border, typical habitat of Rivulus urophthalmus.Photo by W. J. E. M. Costa.
aqua vol. 11 no. 4 - 2006143
Wilson J. E. M. Costa
DistributionLower Amazon basin, including lower Xingu and
Tocantins river basins, and adjacent coastaldrainages to East, northern Brazil (Fig. 10).
HabitatShallow creeks at sunny places, sometimes in forest
border, at depths of about 5-10 cm (Fig. 11).
Rivulus taeniatus Fowler, 1945(Figs 12-16; Table I)
Rivulus taeniatus Fowler, 1945: 129 (type locality:Morelia, río Caqueta drainage, Colombia; holotype:ANSP 71720, illustrated by Fowler 1945: 129).
Material examinedBrazil: Estado do Amazonas: UFRJ 6245, 23; UFRJ
6246, 4 (c&s); creek adjacent to rio Urubuí, rioUrubu basin, Presidente Figueiredo, 2º2’43.9”S60°1’43.2”W; W. J. E. M. Costa, R. Paiva & D.Ramos, 11 Jun. 2004. INPA 1847, 1; INPA 1848, 2;pool near igarapé Acará, Reserva Ducke, Manaus;Rosa, 23 Mar. 1988. INPA 2367, 4; Belém dos Pal-mares, rio Solimões, between Benjamin Constantand São Paulo de Olivença; R. Silva, 1 May 1987.INPA 3905, 3; INPA 3906, 1; igarapé Candiru, km50 of the Manaus-Itacoatiara road; C. Pereira, 11 Jul.1990. UFRJ 3931, 73; pool near igarapé Acará, Re-serva Ducke, Manaus; C. Figueiredo & C. Codeço,
21 Sep. 1996. Estado do Acre: ZUEC non catalo-gued, 4; Campus Cacimba; C. F. D. Haddad & J. R.Santos, 2 Jan. 1982. UFRJ 6291, 7; UFRJ 6292, 82;creek between Rio Branco and Senador Guiomard,rio Acre drainage; W. J. E. M. Costa et al., 12 Jun.2003.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anterior por-tion of trunk slightly deeper than wide, jaws short,snout blunt, tip of anal fin rounded in male, caudalfin rounded in male, pelvic-fin tip usually not reach-ing anus and never reaching the anterior portion ofthe anal-fin base in male, dorsal-fin origin on verti-cal between base of last 2nd and 3rd anal-fin rays, dor-sal-fin rays 7-9, anal-fin rays 12-15, frontal squama-tion E-patterned, frontal scales arranged circularly,canal preopercular short and opened, contact organson flank scales in male, longitudinal series of scales38-40, gill rakers of first branchial arch 2 + 9, red
Table I. Morphometric data of the R. urophthalmus, R. taeniatus, R. micropus, and R. ornatus.
R. urophthalmus R. taeniatus R. micropus R. ornatus
males (10) females (10) males (10) females (10) males (6) females (4) males (5) females (4)
Standard length (mm) 26.7-34.9 24.9-35.4 29.7-61.1 28.8-50.1 31.9-42.8 30.8-41.8 19.4-21.1 19.3-19.7
Percents of standard lengthBody depth 19.6-21.9 19.5-21.9 18.7-20.9 18.6-21.7 18.0-19.6 17.0-19.0 17.4-19.0 16.6-18.4Caudal peduncle depth 12.5-13.9 12.7-13.4 12.6-16.0 12.6-14.3 12.9-14.1 12.4-13.4 12.7-14.4 12.5-13.4Pre-dorsal length 77.0-79.1 76.2-78.8 78.0-79.9 78.6-80.8 77.7-80.6 77.9-81.1 78.5-81.7 81.0-84.6Pre-pelvic length 53.1-55.0 53.3-55.8 53.6-57.7 54.5-57.5 50.5-54.5 52.5-53.9 53.3-57.0 53.6-57.3Length of dorsal-fin base 7.0-9.1 7.1-8.5 6.1-8.5 6.5-8.5 6.1-8.1 6.1-8.3 6.2-7.8 5.0-6.1Length of anal-fin base 16.7-19.3 16.0-18.7 16.6-18.7 16.3-19.1 16.3-18.7 15.5-17.6 14.0-16.4 13.3-15.2Caudal-fin length 30.4-36.2 33.1-37.4 29.0-33.3 29.0-32.4 30.4-36.2 31.4-33.3 46.0-51.6 44.7-45.8Pectoral-fin length 17.4-19.7 17.1-20.9 17.0-20.0 16.7-19.6 18.9-20.7 18.0-19.3 22.1-26.3 22.5-23.6Pelvic-fin length 8.0-10.0 7.7-8.9 7.9-10.6 7.1-9.1 9.9-13.4 9.2-10.0 11.5-14.1 10.4-12.1Head length 23.0-25.4 23.1-26.2 22.8-24.8 23.0-24.9 24.4-25.3 24.3-25.6 27.0-28.6 27.8-28.1Percents of head lengthHead depth 63.7-71.2 64.5-73.7 64.1-73.8 64.7-74.5 57.5-60.3 54.7-58.6 51.4-55.7 48.8-53.6Head width 75.6-82.9 77.9-85.7 78.2-91.8 78.7-88.3 72.8-74.4 71.4-77.4 73.6-79.6 73.3-80.0Snout length 13.1-16.8 13.9-17.2 13.0-17.0 13.9-15.8 14.8-15.1 13.9-14.2 11.5-14.3 12.0-13.5Lower jaw length 19.5-23.2 20.8-25.3 23.3-25.5 23.1-26.1 22.8-24.2 24.6-27.0 20.2-22.8 19.1-20.3Eye diameter 30.5-34.5 29.3-34.5 28.4-36.7 28.0-34.9 30.4-32.1 28.4-32.0 28.6-34.1 28.5-33.5
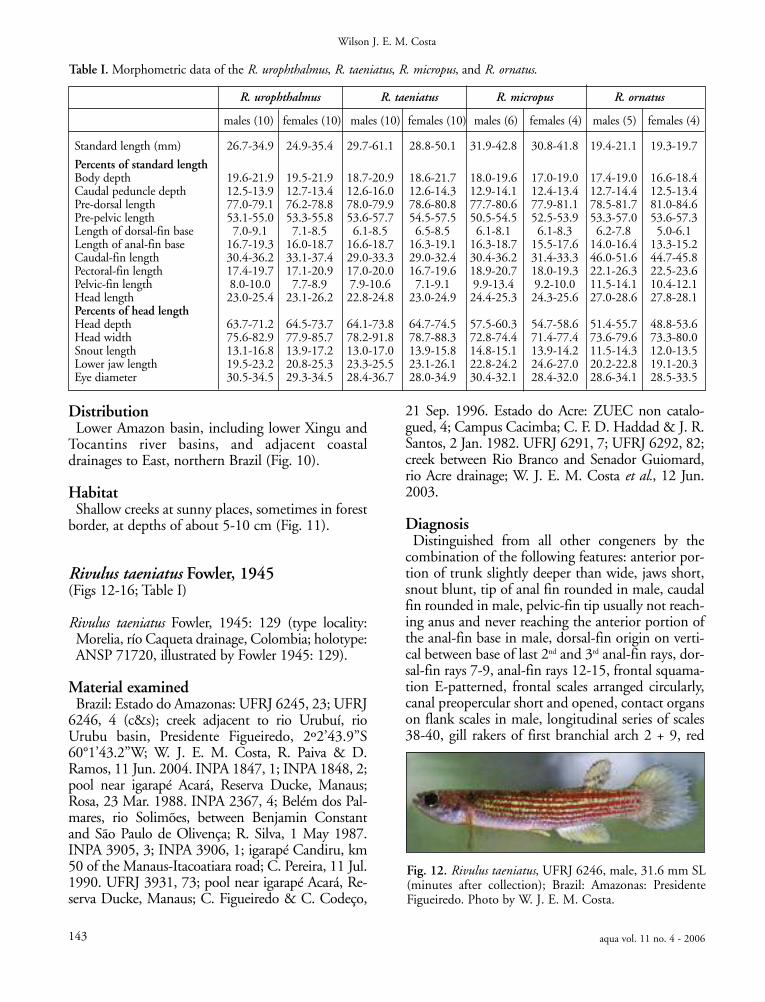
Fig. 12. Rivulus taeniatus, UFRJ 6246, male, 31.6 mm SL(minutes after collection); Brazil: Amazonas: PresidenteFigueiredo. Photo by W. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006 144
stripes on flank, jaws not distinctively pigmentedexcept by red tip of lower jaw, and round black spoton dorsal portion of caudal-fin in female.
DescriptionMorphometric data appear inTable I. Largest spec-
imen, a male 61.1 mm SL. Dorsal profile gentlyconvex from snout to end of dorsal-fin base, approx-imately straight on caudal peduncle. Ventral profileslightly convex from lower jaw to end of anal-finbase, about straight on caudal peduncle. Body slen-der, subcylindrical anteriorly, slightly deeper thanwide, to compressed posteriorly. Greatest bodydepth at vertical just in front to pelvic-fin base. Jawsshort, snout blunt.Tip of dorsal and anal fins rounded. Caudal fin
rounded. Pectoral fin short and rounded, posteriormargin reaching vertical at about 60 % of lengthbetween pectoral- and pelvic-fin bases. Pelvic finshort and elliptical, often tip not reaching anus,sometimes on urogenital papilla in male. Pelvic-finbases medially separated by short interspace. Dorsal-fin origin on vertical between base of last 2nd and 3rd
anal-fin rays, and between neural spines of 21st and23rd vertebrae. Anal-fin origin between pleural ribsof 15th and 16th vertebrae. Dorsal-fin rays 7-9; anal-fin rays 12-15; caudal-fin rays 27-28; pectoral-finrays 13-14; pelvic-fin rays 7.
Scales small, cycloid. Body and head entirelyscaled, except anterior ventral surface of head. Bodysquamation extending over anterior 30 % of caudal-fin base; no scales on dorsal and anal-fin bases.Frontal squamation E-patterned; E-scales not over-lapping medially; scales arranged in regular circularpattern around A-scale without exposed margins.Longitudinal series of scales 38-40; transverse seriesof scales 8; scale rows around caudal peduncle 16-18. Contact organ on each scale of midventral ven-tral portion of flank in male.Cephalic neuromasts: supraorbital 3 + 3, parietal 1,
anterior rostral 1, posterior rostral 1, infraorbital 1 +1 + 13-15 + 1, preorbital 2-3, otic 1, post-otic 2,supratemporal 1, median opercular 1, ventral oper-cular 2, preopercular 2 + 4, mandibular 3-4 + 1, lat-eral mandibular 2.Interhyal minute, cartilaginous. Basihyal subtrian-
gular, width about 55 % length; basihyal cartilageabout 10 % basihyal length. Six branchiostegal rays.Second pharyngobranchial teeth 1. Gill rakers of firstbranchial arch 2 + 9. Vomerine teeth 1. Ventralprocess of posttemporal short. Total vertebrae 33-36.Coloration in life: Male: Side of body metallic yel-
lowish green to metallic purplish blue ventrally; 7-8longitudinal rows of red to reddish brown dots, usu-ally in close proximity, forming stripes; usually three,sometimes four or five rows of red dots crossing cau-
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
Fig. 13. Rivulus taeniatus, UFRJ 6246, female, 33.3 mmSL (minutes after collection); Brazil: Amazonas: PresidenteFigueiredo. Photo by W. J. E. M. Costa.
Fig. 14. Rivulus taeniatus, UFRJ 6291, male, 28.1 mm SL(one day after collection); Brazil: Acre: Rio Branco. Photoby W. J. E. M. Costa.
Fig. 16. Rivulus taeniatus, UFRJ 6291, female, 30.9 mmSL (one day after collection); Brazil: Acre: Rio Branco.Photo by W. J. E. M. Costa.
Fig. 15. Rivulus taeniatus, UFRJ 6291, male, 28.5 mm SL(one day after collection); Brazil: Acre: Rio Branco (com-pare to Fig. 14 to observe slightly different pigmentationpatterns in individuals of the same population). Photo byW. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006145
dal peduncle and reaching caudal-fin base, remainingrows reaching vertical between dorsal and anal-finbases; dark chromatophores concentrated on longi-tudinal zone between flank and dorsum. Dorsumlight brown. Venter white. Side of head metallic yel-lowish green to metallic purplish blue, sometimesdark bluish grey when fish exposed to strong sun-light. Lower jaw light brown, often with red tip. Irislight yellowish to greenish brown. Dorsal fin palelight grey with subbasal oblique row brown spots,often forming stripe; distal portion pale yellow. Analfin pale yellow, basal portion sometimes blue, oftenwith dark grey margin. Caudal fin light grey, dorsalportion pale yellow. Paired fins yellowish hyaline.Female: Side of body light brown, with eight lon-
gitudinal rows of dark red to reddish brown dots,sometimes in close proximity, forming stripes; darkchromatophores concentrated on longitudinalzone between flank and dorsum. Dorsum lightbrown. Venter white. Side of head pale metallicyellowish green to metallic purplish blue, some-times dark bluish grey when fish exposed to strong
sunlight. Lower jaw light brown. Iris light yellow-ish brown. Unpaired fins yellowish hyaline withfaint brown dots; round black spot on dorsal por-tion of caudal fin reaching fin margin, anteriorlybordered by small, triangular pale yellow spot.Paired fins yellowish hyaline.
DistributionWestern and middle rio Amazonas basin (Fig. 10).
HabitatShallow creeks at sunny places, sometimes in forest
border, at depths of about 5-10 cm (Fig. 17).
Rivulus micropus (Steindachner, 1863)(Fig. 18; Table I)
Fundulus micropus Steindachner, 1863: 184 (typelocality: Rio Negro, Brazil; holotype: NMW77762; photographs and X-ray of lectotype inHuber 1991: 67, fig. 2).
Rivulus compressus Henn, 1916: 111 (type locality:Manáos [now Manaus]; emended as igarapé daCachoeira Grande, 2 miles out of Manaos, nearmouth of rio Negro by Eigenmann 1911; holotype:FMNH 57003, illustrated by Henn 1916: fig. 1).
Material examinedBrazil: Estado do Amazonas: UFRJ 6288, 2; creek
at the road Manaus-Manacapuru, 3°13’51.2”S60°32’37.5”W;W. J. E. M. Costa, R. Paiva &D. Ra-mos, 9 Jun. 2004. UFRJ 6289, 3; stream at the roadManacapuru-Novo Airão, lower rio Negro basin,3°10’30.7”S 60°42’34.9”W; W. J. E. M. Costa, R.Paiva&D. Ramos, 9 Jun. 2004.MZUSP 38307, 13;MZUSP 38449, 2 (c&s); lago Buiuçu, Ati-Paraná,NW of Fonte Boa; EPA, 11-12 Oct. 1968. INPA1837, 3 (1c&s); FUA, Manaus; J. Zuanon, J. Porto& E. N. S. Silva, 10 May 1988. INPA 2371, 2; Dis-trito Industrial, Manaus; J. Zuanon, 21 Nov. 1988.
Wilson J. E. M. Costa
Fig. 17. Brazil: Amazonas: Presidente Figueiredo; creekclose to rio Urubuí, habitat of Rivulus taeniatus. Photo byW. J. E. M. Costa.
Fig. 18. Rivulus micropus, UFRJ 6288, male, 31.9 mm SL(one day after collection); Brazil: Amazonas: Manacapuru.Photo by W. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006 146
DiagnosisDistinguished from all other congeners by the
combination of the following features: anterior por-tion of trunk slightly deeper than wide, jaws slightlyelongated, snout pointed, tip of anal fin slightlypointed in male, caudal fin rounded in male, pelvic-fin tip reaching anterior portion of anal-fin base inmale, dorsal-fin origin on vertical between base ofpenultimate and last anal-fin ray, dorsal-fin rays 7-9,anal-fin rays 13-15, frontal squamation E or D-pat-terned, frontal scales arranged circularly, canal pre-opercular short and opened, contact organs on flankscales in male, longitudinal series of scales 40-45, gillrakers of first branchial arch 2 + 10, pink stripes onflank, jaws not distinctively pigmented, and roundblack spot on dorsal portion of caudal fin in female.
DescriptionMorphometric data appear inTable I. Largest spec-
imen, a male 42.8 mm SL. Dorsal profile gentlyconvex from snout to end of dorsal-fin base, approx-imately straight on caudal peduncle. Ventral profileslightly convex from lower jaw to end of anal-finbase, about straight on caudal peduncle. Body slen-der, subcylindrical anteriorly, slightly deeper than
wide, to compressed posteriorly. Greatest bodydepth at vertical just in front to pelvic-fin base. Jawsslightly elongated, snout pointed.Tip of dorsal rounded; tip of anal fin slightly
pointed in male, rounded in female. Caudal finrounded. Pectoral fin short and rounded, posteriormargin reaching vertical at about 80 % of distancebetween pectoral and pelvic-fin bases. Pelvic fin ellip-tical, tip reaching between urogenital papilla and baseof 2nd anal-fin ray in male, and urogenital papilla infemale. Pelvic-fin bases medially separated by shortinterspace. Dorsal-fin origin on vertical between baseof penultimate and last anal-fin ray, and betweenneural spines of 24th and 25th vertebrae. Anal-fin ori-gin between pleural ribs of 16th and 17th vertebrae.Dorsal-fin rays 7-9; anal-fin rays 13-15; caudal-finrays 29-30; pectoral-fin rays 15; pelvic-fin rays 7-8.Scales small, cycloid. Body and head entirely
scaled, except anterior ventral surface of head. Bodysquamation extending over anterior 30-40% of cau-dal-fin base; no scales on dorsal and anal-fin bases.Frontal squamation E or D-patterned; E-scales notoverlapping medially; scales arranged in regular cir-cular pattern around A-scale without exposed mar-gins. Longitudinal series of scales 40-45; transverse
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
Fig. 19. Geographic distribution of: Rivulus beniensis (lozenges), R. dibaphus (dots), R. micropus (stars), R. strigatus (triangles),and R. tecminae (inverted triangles). One symbol may represent more than one collecting site.

aqua vol. 11 no. 4 - 2006147
series of scales 8; scale rows around caudal peduncle16. Contact organ on each scale of midventral ven-tral portion of flank in male.Cephalic neuromasts: supraorbital 3 + 3, parietal 1,
anterior rostral 1, posterior rostral 1, infraorbital 1 +16 + 1, preorbital 3, otic 1, post-otic 2, supratempo-ral 1, median opercular 1, ventral opercular 2, preop-ercular 2 + 4, mandibular 4 + 2, lateral mandibular 3.Interhyal minute, cartilaginous. Basihyal subtrian-
gular, width about 55% length; basihyal cartilageabout 25% basihyal length. Six branchiostegal rays.Second pharyngobranchial teeth 3-4. Gill rakers offirst branchial arch 2 + 10. Vomerine teeth 1. Ven-tral process of posttemporal long. Total vertebrae 35.Coloration in life: Male: Side of body dark blue,
with 8 narrow pink stripes, of which 3-5 reachingcaudal-fin base; dark chromatophores concentratedon longitudinal zone between flank and dorsum.Dorsum light brown. Venter white. Side of head darkpurplish blue. Jaws dark brown. Iris brown.Unpaired fins pale yellow with brown spots. Pairedfins hyaline.Female: Similar to male, but flank stripes darker,
and round black spot on dorsal portion of caudal finreaching fin margin, anteriorly bordered by small,triangular pale yellow spot.
DistributionMiddle rio Amazonas basin (Fig. 19).
HabitatMargins of streams and lakes at sunny places,
sometimes in forest border, at depths of about 10-50cm (Fig. 20).
Subgenus Anablepsoides Huber, 1992
Anablepsoides Huber, 1992: 43 (type species: Rivulusatratus Garman; type by original designation).
DiagnosisDistinguished from the remaining subgenera of
Rivulus by the following apomorphic features:rounded rostral cartilage (15.0), external medial teethof premaxilla and dentary laterally displaced (20.1),long, acuminate caudal fin (76.1, 77.3), frontal squa-mation S-patterned (80.4; also occurring in Owiy-eye), oblique dark grey on flank in male whenexposed to sunlight (93.3), and transverse stripethrough chin (98.2; also occurring in Owiyeye).
Included speciesRivulus atratus Garman, 1895 and R. ornatus Gar-
man, 1895.
DistributionAmazonas river basin, in Brazil and Peru.
Rivulus ornatus Garman, 1895(Fig. 21; Table I)
Rivulus ornatus Garman, 1895: 139 (type locality:Silva, Cudajas, Lago Aleixo, Lake Hyanuary [fourdistinct localities in the Brazilian Amazon, Estadodo Amazonas: Silves, Codajás, lago Aleixo andparaná do Janauari]; lectotype: MCZ 6266, desig-nated by Huber 1992: 347).
Rivulus obscurus Garman, 1895: 140 (type locality:Lake Hyanuary [paraná do Janauari, Estado doAmazonas, Brazil]; lectotype: MCZ 6853, desig-nated by Huber 1992: 336).
Wilson J. E. M. Costa
Fig. 20. Brazil: Amazonas: Manacapuru; stream near roadManaus-Manacapuru, habitat of Rivulus micropus. Photoby W. J. E. M. Costa.
Fig. 21. Rivulus ornatus, not preserved, male, about 20.0mm SL (about one week after collection); Brazil: Ama-zonas: Manaus. Photo by W. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006 148
Material examinedBrazil: Estado do Amazonas: MCZ 6854, 4 para-
lectotypes; paraná do Janauari, 3°12’S 60º5’W; L.Agassiz, 27 Oct. 1865. MCZ 98664, 1 paralecto-type of R. obscurus; same data as MCZ 6854. UFRJ7499, 28; stream tributary to rio Sanasani, Silves;EPA, 7 Oct. 1967. INPA 3908, 3; igarapé Candiru,km 50 of the road Manaus-Itacoatiara; C. Pereira,11 Jul. 1990. Estado do Pará: MZUSP 37208, 3;lago Jacaré, rio Trombetas; EPA, 7 Nov. 1969.MZUSP 38440, 2 (c&s); rio Curuinapui; EPA, 26Jan. 1968.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anteriorportion of trunk slightly wider than deep, jawsshort, snout blunt, tip of anal fin rounded in male,caudal fin elliptical to acuminate in male, pelvic-fin tip reaching the anterior portion of the anal-finbase in male, dorsal-fin origin posterior to anal-finbase, dorsal-fin rays 5-7, anal-fin rays 9-11, frontalsquamation S-patterned, contact organs absent,frontal scales arranged circularly, canal preopercu-lar short and opened, longitudinal series of scales
29-32, gill rakers of first branchial arch 1 + 7,oblique rows of red dots on flank in male, trans-verse black bar through the chin, and no black spoton dorsal portion of caudal-fin.
DescriptionMorphometric data appear in Table I. Largest
specimen, a male 21.1 mm SL. Dorsal profile gen-tly convex from snout to end of dorsal-fin base,approximately straight on caudal peduncle. Ventralprofile weakly convex from lower jaw to end ofanal-fin base, about straight on caudal peduncle.Body slender, subcylindrical anteriorly, wider thandeep, to compressed posteriorly. Greatest bodydepth at vertical just in front of pelvic-fin base.Jaws short, snout blunt.Tip of dorsal and anal fins rounded. Caudal fin
elliptical, often pointed in male. Pectoral finrounded, posterior margin reaching verticalbetween pelvic-fin base and anus in male, and ver-tical just anterior to pelvic-fin base in female.Pelvic fin elliptical, tip reaching between urogeni-tal papilla and base of 4th anal-fin ray in male,between urogenital papilla and anal-fin origin infemale. Pelvic-fin bases medially separated by short
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
Fig. 22. Geographic distribution of: Rivulus ornatus (stars), R. uakti (inverted triangle), R. uatuman (dot), and R. zygonectes(lozenges). One symbol may represent more than one collecting site.

aqua vol. 11 no. 4 - 2006149
interspace. Dorsal-fin origin on vertical just poste-rior to anal-fin base, between neural spines of 20th
and 21st vertebrae. Anal-fin origin between pleuralribs of 13th and 14th vertebrae. Dorsal-fin rays 5-7;anal-fin rays 9-11; caudal-fin rays 24; pectoral-finrays 12-13; pelvic-fin rays 7.Scales small, cycloid. Body and head entirely
scaled, except anterior ventral surface of head.Body squamation extending over anterior 20 % ofcaudal-fin base; no scales on dorsal and anal-finbases. Frontal squamation S-patterned; E-scalesnot overlapping medially; scales arranged in regu-lar circular pattern around A-scale without exposedmargins. Longitudinal series of scales 29-32; trans-verse series of scales 7; scale rows around caudalpeduncle 16. Contact organs absent.Cephalic neuromasts: supraorbital 3 + 3, parietal 1,
anterior rostral 1, posterior rostral 1, infraorbital 1 +12 + 1, preorbital 2, otic 1, post-otic 1, supra-temporal 1, median opercular 1, ventral opercular 2,preopercular 2 + 4, mandibular 3-4 + 1, lateral man-dibular 3.Interhyal minute, cartilaginous. Basihyal subtri-
angular, width about 50% length; basihyal carti-lage about 10% basihyal length. Six branchiostegal
rays. Second pharyngobranchial teeth 1-2. Gillrakers of first branchial arch 1 + 7. Vomerine teeth1. Ventral process of posttemporal absent. Totalvertebrae 29-30.Coloration in life: Male: Side of body pale blue,
usually with 8 narrow light brown vertical lines, over-lapped by oblique rows of red dots; dark chro-matophores concentrated on longitudinal zonebetween flank and dorsum. Dorsum light brownwith transverse dark brown bars. Venter brown. Sideof head and infraorbital region dark metallic bluewith red dots. Jaws light brown; dark brown to blacktransverse bar just below lower jaw. Iris brown. Dor-sal and anal fins pale yellow to pale blue, with red toreddish brown bars. Dorsal portion of caudal fin paleyellow to pale blue with dark red narrow bars, medianportion dark brown, and ventral portion reddishorange. Pectoral fin hyaline. Pelvic fin pale yellow.Female: Side of body light brown with 8 narrow
brown vertical lines, and 5-7 oblique dark brownbars; dark chromatophores concentrated on longi-tudinal zone between flank and dorsum. Dorsumlight brown with transverse brown bars. Venterlight brown. Side and ventral portion of headbrown. Jaws light brown; dark brown to black
Wilson J. E. M. Costa
Fig. 23. Geographic distribution of: Rivulus amanapira (inverted triangle), R. atratus (stars), R. kirovskyi (dot), and R. litter-atus (lozenges). One symbol may represent more than one collecting site.

aqua vol. 11 no. 4 - 2006 150
transverse bar just below lower jaw. Iris brown.Unpaired fins hyaline with transverse narrow darkbrown bars. Paired fins hyaline.
DistributionMiddle rio Amazonas basin, Brazil (Fig. 22).
HabitatStagnant waters in sunny places near forest bor-
der, with dense aquatic vegetation, about 50-150cm deep.
Rivulus atratus Garman, 1895
Rivulus atratus Garman, 1895: 140 (type locality:Jutahy [now Jutaí, Estado do Amazonas, Brazil];lectotype: MCZ 6318, designated by Huber 1992:102).
Material examinedBrazil: Estado do Amazonas: MCZ 98675, 1 para-
lectotype; Jutaí; Thayer Expedition, Sep. 1865.MZUSP 38310, 1; MZUSP 38449, 1 (c&s); rioIçapó, mouth of rio Jutaí; EPA, 22 Oct. 1968.MZUSP 38494, 12; lago Miuá, above Codajás;EPA, 25 Sep. 1968.
DiagnosisDistinguished from all other congeners by the com-
bination of the following features: anterior portion oftrunk wider than deep, jaws short, snout blunt, tip ofanal fin rounded in male, caudal fin elliptical toacuminate in male, pelvic-fin tip not reaching to uro-genital papilla in male, dorsal-fin origin posterior toanal-fin base, dorsal-fin rays 5-6, anal-fin rays 8-10,frontal squamation S-patterned, frontal scalesarranged circularly, canal preopercular short andopened, contact organs absent, longitudinal series ofscales 32-34, gill rakers of first branchial arch 1 + 7,dark brown oblique bars on flank, transverse blackbar through the chin, and no black spot on dorsalportion of caudal fin. Also distinguished from allother congeners in having venter dark brown to blackand dark brown spots on pectoral fin.
DistributionMiddle rio Amazonas basin, Brazil and Peru (Fig.
23).
HabitatStill waters close to river and lake margins.
Benirivulus, new subgenus
Type species: Rivulus beniensis Myers, 1927.
DiagnosisTwo autapomorphic conditions were found in
Benirivulus, which are not present in other con-geners: a colour pattern of anal fin, consisting of oneor two subbasal rows of reddish brown close dots,sometimes forming stripes, and a broad dark greydistal zone on the dorsal fin (Fig. 24).
EtymologyThe name Benirivulus is derived from Beni, an
important Amazonian river, and rivulus, from theLatin meaning stream, in reference to the occurrenceof the new taxon in streams of that river basin. Gen-der masculine.
Included speciesOnly R. beniensis, the type species of the subgenus.
DistributionMadeira river drainage, Amazonas river basin, in
Bolivia and Brazil.
Rivulus beniensis Myers, 1927(Fig. 24; Table II)
Rivulus strigatus (non R. strigatus Regan); Pearson,1925: 51 (misidentification).
Rivulus beniensis Myers, 1927b: 133 (type locality:Ivon, Rio Beni [lower río Beni, just above conflu-ence with río Madre de Díos, Departamento deBeni], Bolivia; lectotype: USNM 86776, desig-nated by Fels & Huber 1985: 35).
Rivulus beniensis lacustris Myers, 1927b: 133 (typelocality: lagoons along shores of Lake Rogoagua[lago Rogoaguado, río Yata basin, which drainsinto río Mamoré, immediately above confluence
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
Fig. 24. Rivulus beniensis, UFRJ 5885, male, 19.6 mm SL(one day after collection); Brazil: Rondônia: betweenMutum Paraná and Abunã. Photo by W. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006151
with río Beni], Bolivia; lectotype: UMMZ 66396,designated by Fels & Huber 1985: 35).
Rivulus bolivianus Seegers, 1988: 172 (type locality:laguna Suarez, ca. 4 km SWTrinidad, río Mamoré,Bolivia; holotype: ZFMK 14602).
Material examinedBolivia: Departamento de Beni: CBF 1886, 54;
CBF 1873, 39; Espírito, Estación Biológica Beni;J. Sarmiento, 23-24 Apr. 1987. Brazil: Estado deRodônia: UFRJ 5884, 67; UFRJ 5885, 4 (c&s);swamp near road BR-364, between Mutum-Paranáand Abunã, rio Madeira basin, 9º40’34.2”S65º4’9.9”W; W. J. E. M. Costa, S. Lima & A.Pinto, 6 Jun. 2003. UFRJ 5881, 6; swamp close torio Mamoré, near Guajará-Mirim, 10º45’53.5”S65º20’21.7”W; W. J. E. M. Costa, S. Lima & A.Pinto, 7 Jun. 2003. Estado de Mato Grosso:swamp at km 2 of the road between Vila Bela andCasal Vasco, near rio Guaporé; W. J. E. M. Costa,M. I. Landim, R. D’Arrigo & C. Moreira, 29 Apr.1996.
DiagnosisDistinguished from all other congeners by the com-
bination of the following features: anterior portion oftrunk slightly deeper than wide, jaws short, snoutblunt, tip of anal fin rounded in male, caudal finrounded in male, pelvic-fin tip reaching the anteriorportion of the anal-fin base in male, dorsal-fin originon vertical between base of 9th and 10th anal-fin rays,
dorsal-fin rays 7-8, anal-fin rays 11-13, frontal squa-mation E-patterned, frontal scales arranged circu-larly, canal preopercular short and opened, contactorgans on flank scales in male, longitudinal series ofscales 29-32, gill rakers of first branchial arch 1 + 7,longitudinal rows of reddish brown dots stripes onflank in male, jaws black, and round black spot ondorsal portion of caudal-fin in female.
DescriptionMorphometric data appear in Table II. Largest
specimen, a female 28.8 mm SL. Dorsal profile gen-tly convex from snout to end of dorsal-fin base,approximately straight on caudal peduncle. Ventralprofile slightly convex from lower jaw to end of anal-fin base, about straight on caudal peduncle. Bodyslender, subcylindrical anteriorly, slightly deeperthan wide, to compressed posteriorly. Greatest bodydepth at vertical just in front to pelvic-fin base. Jawsshort, snout blunt.Tip of dorsal and anal fins rounded. Caudal fin
rounded. Pectoral fin rounded, posterior marginreaching vertical at about 80% length between pec-toral-fin and pelvic-fin bases. Pelvic fin elliptical, tipreaching between urogenital papilla and anal-fin ori-gin in male, between anus and urogenital papilla infemale. Pelvic-fin bases medially separated by shortinterspace. Dorsal-fin origin on vertical betweenbase of 9th and 10th anal-fin rays, and between neuralspines of 19th and 21st vertebrae. Anal-fin originbetween pleural ribs of 15th and 16th vertebrae. Dor-
Wilson J. E. M. Costa
Table II. Morphometric data of Rivulus beniensis, R. strigatus, and R. dibaphus.
R. beniensis R. strigatus R. dibaphus
males (10) females (10) males (10) females (7) males (9) females (6)
Standard length (mm) 19.6-23.0 19.4-25.8 22.2-26.5 18.7-22.2 19.2-26.5 17.8-23.4
Percents of standard lengthBody depth 19.8-22.0 19.5-21.3 18.8-21.9 19.2-20.8 19.7-23.3 19.6-21.5Caudal peduncle depth 13.0-14.3 12.7-14.4 13.4-15.5 12.7-14.6 13.0-14.9 13.1-14.4Pre-dorsal length 76.2-78.3 76.8-79.9 70.4-73.3 71.2-74.1 70.5-73.1 71.4-72.9Pre-pelvic length 53.0-55.2 52.7-55.8 52.3-57.1 53.1-56.0 55.0-57.4 53.6-57.1Length of dorsal-fin base 8.2-9.9 7.0-9.2 10.0-12.1 10.7-12.1 12.0-13.6 10.3-12.7Length of anal-fin base 17.0-20.2 16.5-17.9 17.7-19.5 16.5-18.5 18.9-21.2 16.6-19.1Caudal-fin length 34.5-39.3 34.0-37.4 36.0-41.6 36.7-42.6 36.2-41.0 36.9-41.0Pectoral-fin length 18.2-21.1 18.8-21.2 18.8-21.1 19.4-22.7 20.6-23.2 20.1-21.9Pelvic-fin length 9.9-11.9 8.7-10.6 9.5-11.3 8.5-10.8 9.3-10.7 9.2-10.8Head length 25.1-27.6 24.7-26.8 25.4-27.4 25.9-27.3 26.1-28.5 26.5-28.9Percents of head lengthHead depth 63.0-67.9 60.2-66.3 63.4-68.8 63.8-68.0 61.0-67.7 60.1-64.8Head width 71.7-79.6 72.7-77.8 73.4-79.7 73.8-80.2 71.7-77.0 73.7-79.3Snout length 14.2-16.4 12.7-16.4 11.7-14.4 12.2-14.0 11.9-15.1 12.3-16.3Lower jaw length 15.8-20.8 16.7-20.3 19.1-23.0 19.0-22.0 19.3-23.5 19.8-21.9Eye diameter 33.3-36.2 31.5-37.1 33.1-35.4 32.2-37.0 30.1-33.8 32.7-35.1

aqua vol. 11 no. 4 - 2006 152
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
sal-fin rays 7-8; anal-fin rays 11-13; caudal-fin rays23-26; pectoral-fin rays 13; pelvic-fin rays 6.Scales small, cycloid. Body and head entirely
scaled, except anterior ventral surface of head. Bodysquamation extending over anterior 25 % of caudal-fin base; no scales on dorsal and anal-fin bases.Frontal squamation E-patterned; E-scales not over-lapping medially; scales arranged in regular circularpattern around A-scale without exposed margins.Longitudinal series of scales 29-32; transverse seriesof scales 7; scale rows around caudal peduncle 16.Contact organ on each scale of midventral portionof flank in male.Cephalic neuromasts: supraorbital 3 + 3, parietal 1,
anterior rostral 1, posterior rostral 1, infraorbital 1 +12 + 1, preorbital 2, otic 1, post-otic 1, supratempo-ral 1, median opercular 1, ventral opercular 1, preop-ercular 2 + 4, mandibular 3 + 1, lateral mandibular 2.Interhyal minute, cartilaginous. Basihyal subtri-
angular, width about 35 % length; basihyal carti-lage about 30 % basihyal length. Six branchioste-gal rays. Second pharyngobranchial teeth 2. Gillrakers of first branchial arch 1 + 7. Vomerine teeth2. Ventral process of posttemporal absent. Totalvertebrae 30-32.Coloration in life: Male: Side of body metallic yel-
lowish green; six longitudinal rows of reddish browndots, of which often two or three rows with sparsedots or absent, middle row forming stripe; dark chro-matophores concentrated on longitudinal zonebetween flank and dorsum, often forming shortoblique dark brown bars when fish exposed to strongsunlight. Dorsum light brown. Venter white. Side of
head metallic yellowish green with small dark brownspots. Lower jaw black. Iris light yellowish with blackvertical stripe. Dorsal fin pale yellow with subbasalrow of reddish brown dots and dark grey distal stripe.Anal fin light blue on basal portion and pale yellowon distal portion, with one or sometimes two sub-basal rows of reddish brown dots and dark grey distalstripe. Caudal fin light grey. Paired fins hyaline, ante-rior margin of pelvic fin dark grey.Female: Side of body with irregularly shaped
brown blotches, forming longitudinal row ofrounded light brown interspaces on ventral portion;sometimes almost inconspicuous rows of browndots coalesced to brown blotches; dark chro-matophores concentrated on longitudinal zonebetween flank and dorsum, often forming shortoblique dark brown bars when fish exposed tostrong sunlight. Dorsum light brown. Venter white.Side of head pale metallic yellowish green with smalldark brown spots. Lower jaw black. Iris light yel-lowish with black vertical stripe. Dorsal fin hyalinewith two subbasal rows of brown dots and grey dis-tal stripe. Anal fin hyaline with two or three subbasalrows of brown dots and grey distal stripe. Caudal finlight grey; round black spot on dorsal portion ofcaudal fin reaching caudal-fin margin, anteriorlybordered by small, triangular pale yellow spot.Paired fins hyaline, anterior margin of pelvic fingrey.
DistributionRio Madeira basin, Bolivia and Brazil (Fig. 19).
HabitatStagnant waters in sunny places near forest border,
with dense aquatic vegetation, about 50-150 cmdeep (Fig. 25).
Subgenus Laimosemion Huber, 1999
Laimosemion Huber, 1999: 49 (type species: Rivu-lus geayi Vaillant; type by original designation).
DiagnosisDistinguished from other rivulids by having bifid
epipleural ribs (43.1).
Included speciesRivulus agilae Hoedeman, 1954, R. dibaphus
Myers, 1927, R. breviceps Eigenmann, 1909, R.cladophorus Huber, 1991, R. corpulentus Thomerson
Fig. 25. Brazil: Rondônia: between Mutum Paraná andAbunã; lake at the border of forest, habitat of Rivulusbeniensis. Photo by W. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006153
Wilson J. E. M. Costa
& Taphorn, 1993, R. frenatus Eigenmann, 1912, R.geayi Vaillant, 1899, R. gransabanae Lasso, Taphorn& Thomerson, 1992, R. lyricauda Thomerson,Berkenkamp &Taphorn, 1991, R. nicoi Thomerson& Taphorn, 1992, R. strigatus Regan, 1912, R. tor-renticola Vermeulen & Isbrücker, 2000, and R.xiphidius Huber, 1979.
DistributionOrinoco river basin, Guiana rivers, eastern Ama-
zonas river basin and adjacent coastal areas inVenezuela, Guiana, Suriname, Guyana and Brazil.
Rivulus strigatus Regan, 1912(Figs. 26-28; Table II)
Rivulus strigatus Regan, 1912: 502 (type locality:Amazon [Belém, Pará, Brazil]; holotype: BMNH1912.8.30).
R. geayi (non R. geayi Vaillant); Huber 1991: 69-70(equivocal synonymy).
Material examinedBrazil: Estado do Pará: UFRJ 6249, 25; UFRJ
6250, 6; UFRJ 6251, 4 (c&s); stream at the roadPA-446, Primavera, 0°57’57.3”S 47°8’46.8”W; W.J. E. M. Costa, R. Paiva & D. Ramos, 19 Jun. 2004.UFRJ 6262, 1; stream at ilha do Mosqueiro, roadPA-391, Belém, 1°8’59.5”S 48°21’27.9”W; W. J. E.M. Costa, R. Paiva & D. Ramos, 21 Jun. 2004.UFRJ 4075, 16; UFRJ 4078, 11; igarapé Maguary,Belém; A. Sarraf & E. Nogueira, 1 Aug. 1996. UFRJ1820, 1 (c&s); Belém; M. Lacerda, 1990.
DiagnosisDistinguished from all other congeners by the com-
bination of the following features: anterior portion oftrunk slightly deeper than wide, jaws short, snoutblunt, tip of anal fin slightly pointed in male, caudalfin rounded in male, pelvic-fin tip reaching the ante-rior portion of the anal-fin base in male, dorsal-finorigin on vertical between base of 7th and 8th anal-finrays, dorsal-fin rays 8-9, anal-fin rays 11-13, frontalsquamation F-patterned, frontal scales arranged cir-cularly, canal preopercular short and opened, contactorgans on flank scales in male, longitudinal series ofscales 30-32, gill rakers of first branchial arch 1 + 7,longitudinal rows of red spots on anterior portion offlank and red chevron-like oblique bars on posteriorportion of flank in male, jaws black, and no blackspot on dorsal portion of caudal-fin.
DescriptionMorphometric data appear in Table II. Largest
specimen, a male 26.5 mm SL. Dorsal profileslightly convex from snout to end of dorsal-fin base,approximately straight on caudal peduncle. Ventralprofile weakly convex from lower jaw to end of anal-fin base, about straight on caudal peduncle. Bodyslender, subcylindrical anteriorly, slightly deeperthan wide, to compressed posteriorly. Greatest bodydepth at vertical just in front to pelvic-fin base. Jawsshort, snout blunt.Tip of dorsal and anal fins slightly pointed in male,
rounded in female. Caudal fin rounded. Pectoral finrounded, posterior margin reaching vertical at about75 % length between pectoral-fin and pelvic-finbases. Pelvic fin elliptical, tip reaching between baseof 1st and 2nd anal-fin rays in male, between urogen-ital papilla and base of 1st anal-fin ray in female.Pelvic-fin bases medially in contact. Dorsal-fin ori-
Fig. 26. Rivulus strigatus, UFRJ 6262, male, 23.0 mm SL(two days after collection); Brazil: Pará: Belém. Photo byW. J. E. M. Costa.
Fig. 27. Rivulus strigatus, UFRJ 6250, male, 26.5 mm SL(one day after collection); Brazil: Pará: Primavera. Photoby W. J. E. M. Costa.
Fig. 28. Rivulus strigatus, UFRJ 6250, female, 22.2 mm SL(one day after collection); Brazil: Pará: Primavera. Photoby W. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006 154
gin on vertical between base of 7th and 8th anal-finrays, and between neural spines of 18th and 19th ver-tebrae. Anal-fin origin between pleural ribs of 14th
and 15th vertebrae. Dorsal-fin rays 8-9; anal-fin rays11-13; caudal-fin rays 28; pectoral-fin rays 14-15;pelvic-fin rays 6.Scales small, cycloid. Body and head entirely
scaled, except anterior ventral surface of head. Bodysquamation extending over anterior 25 % of caudal-fin base; no scales on dorsal- and anal-fin bases.Frontal squamation F-patterned; E-scales not over-lapping medially; scales arranged in regular circularpattern around A-scale without exposed margins.Longitudinal series of scales 30-32; transverse seriesof scales 7; scale rows around caudal peduncle 16.Contact organ on each scale of midventral portionof flank in male.Cephalic neuromasts: supraorbital 3 + 3, parietal 1,
anterior rostral 1, posterior rostral 1, infraorbital 1 +12-13 + 1, preorbital 2, otic 1, post-otic 1,supratemporal 1, median opercular 1, ventral oper-
cular 2, preopercular 2 + 4, mandibular 3 + 1, lateralmandibular 2-3.Interhyal minute, cartilaginous. Basihyal subtrian-
gular, width about 50 % length; basihyal cartilageabout 25 % basihyal length. Six branchiostegal rays.Second pharyngobranchial teeth absent. Gill rakersof first branchial arch 1 + 7. Vomerine teeth absent.Ventral process of posttemporal absent. Total verte-brae 30-31.Coloration in life: Male: Side of body metallic blue
or greenish blue; four to five longitudinal rows ofred dots on anterior half of flank, and red chevron-like oblique bars with vertex anteriorly directed onposterior half of flank; dark blue humeral blotch.Dorsum light brown. Venter white. Side of headmetallic greenish blue with small red spots; infraor-bital region yellow. Jaws black. Iris brown, anteriorand posterior portion green. Dorsal fin light bluewith red bars on basal and posterior zones, red onanterior and distal portions; sometimes pale yellowanterior margin. Anal fin light blue with grey dotson basal and posterior portions; narrow dark reddishgrey distal stripe. Upper two-thirds of caudal finlight red with transverse rows of small blue spots,lower third greenish blue; narrow dark reddish greyventral stripe. Pectoral fin hyaline. Pelvic fin lightblue with dark reddish grey anterior margin.Female: Side of body light grey with small dark
purplish brown spots arranged both in overlappedlongitudinal and oblique rows; dark grey humeralblotch; dark chromatophores concentrated on longi-tudinal zone between flank and dorsum, often form-ing short transverse dark brown bars when fishexposed to strong sunlight. Dorsum light brown.Venter white. Side of head pale blue with small darkbrown spots; infraorbital region pale yellow. Jawsblack. Iris brown. Dorsal fin pale yellow on basaland posterior portions, light purplish grey on distaland anterior portions. Anal fin pale yellow to palepink, basal portion light blue; distal margin darkgrey. Caudal fin pale yellow with dark brown bars onupper two- thirds of fin. Pectoral fin hyaline. Pelvicfin pale yellow with dark grey anterior margin.
DistributionRiver basins close to the right bank of rio Ama-
zonas mouth (Fig. 19).
HabitatMargins of clear water streams with moderate
current, sand bottom, at depth of about 20-40 cm(Fig. 29).
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
Fig. 29. Brazil: Pará: Primavera; clear water stream, habitatof Rivulus strigatus. Photo by W. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006155
Rivulus dibaphus Myers, 1927(Figs 30-32; Table II)
Rivulus dibaphus Myers, 1927: 121 (type locality:Igarapé do Ajamuri, Lower Amazon [corrected by S.O. Kullander in Huber 1991: 70 as Ajamuri or Aga-muri, a beach in Lago Grande at mouth of rio Mai-curu, just east of Santarém, Pará, Brazil]; lectotype:CAS 44214, designated by Huber 1992: 184).
R. geayi (non R. geayi Vaillant); Huber, 1991: 69-70(equivocal synonymy).
Material examinedBrazil: Estado do Pará: CAS 44214, lectotype;
CAS 44215, 6; Igarapé do Ajamuri, Santarém; C.Ternetz, Jul. 1924. UFRJ 6283, 10; UFRJ 6284, 6(c&s); UFRJ 6285, 4; stream 5 km S of Alter doChão, Santarém, 2°33’1.3”S 54°57’21.1”W; W. J.E. M. Costa, R. Paiva & D. Almeida, 14 Jun.
2004. UFRJ 6286, 2; stream tributary to rio Tapa-jós, Porto Novo, Belterra, 2°37’45.9”S54°58’41.7”W; W. J. E. M. Costa, R. Paiva & D.Almeida, 14 Jun. 2004. UFRJ 6286, 7; stream trib-utary to rio Tapajós, near Porto Novo, Belterra,2°37’54.0”S 54°58’19.6”W; W. J. E. M. Costa, R.Paiva & D. Almeida, 14 Jun. 2004. INPA 2370, 8(1c&); igarapé do Patenós, rio Trombetas basin; J.Zuanon & E. Ferreira, 2 Dec. 1988.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anterior por-tion of trunk slightly deeper than wide, jaws short,snout blunt, tip of anal fin slightly pointed in male,caudal fin subtruncate in male, pelvic-fin tip reach-ing the anterior portion of the anal-fin base in male,dorsal-fin origin on vertical between base of 7th and8th anal-fin rays, dorsal-fin rays 9-10, anal-fin rays12-13, frontal squamation E-patterned, frontalscales arranged circularly, canal preopercular shortand opened, contact organs on flank scales in male,longitudinal series of scales 30-32, gill rakers of firstbranchial arch 1 + 9, longitudinal rows of red spotson anterior portion of flank and red chevron-likeoblique bars on posterior portion of flank in male,jaws black, and no black spot on dorsal portion ofcaudal-fin.
DescriptionMorphometric data appear in Table II. Largest
specimen, a male 26.5 mm SL. Dorsal profileslightly convex from snout to end of dorsal-fin base,approximately straight on caudal peduncle. Ventralprofile weakly convex from lower jaw to end of anal-fin base, about straight on caudal peduncle. Bodyslender, subcylindrical anteriorly, slightly deeperthan wide, to compressed posteriorly. Greatest bodydepth at vertical just in front to pelvic-fin base. Jawsshort, snout blunt.Tip of dorsal and anal fins slightly pointed in male,
rounded in female. Caudal fin subtruncate in male,rounded in female. Pectoral fin rounded, posteriormargin reaching vertical at about 75-90 % lengthbetween pectoral-fin and pelvic-fin bases. Pelvic finelliptical, tip reaching between base of 2nd and 3rd
anal-fin rays in male, between urogenital papilla andbase of 2nd anal-fin ray in female. Pelvic-fin basesmedially in contact. Dorsal-fin origin on verticalbetween base of 7th and 8th anal-fin rays, andbetween neural spines of 16th and 18th vertebrae.Anal-fin origin between pleural ribs of 13th and 14th
Wilson J. E. M. Costa
Fig. 30. Rivulus dibaphus, UFRJ 6284, male, 24.8 mm SL(one day after collection); Brazil: Pará: Santarém. Photo byW. J. E. M. Costa.
Fig. 31. Rivulus dibaphus, UFRJ 6284, male, 22.3 mm SL(one day after collection); Brazil: Pará: Santarém. Photo byW. J. E. M. Costa.
Fig. 32. Rivulus dibaphus, UFRJ 6284, female, 21.8 mmSL (one day after collection); Brazil: Pará: Santarém. Photoby W. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006 156
vertebrae. Dorsal-fin rays 9-10; anal-fin rays 12-13;caudal-fin rays 28-31; pectoral-fin rays 14-15;pelvic-fin rays 6-7.Scales small, cycloid. Body and head entirely scaled,
except anterior ventral surface of head. Body squa-mation extending over anterior 30 % of caudal-finbase; no scales on dorsal and anal-fin bases. Frontalsquamation E-patterned; E-scales not overlappingmedially; scales arranged in regular circular patternaround A-scale without exposed margins. Longitudi-nal series of scales 30-32; transverse series of scales 7;scale rows around caudal peduncle 16. Contact organon each scale of midventral portion of flank in male.Cephalic neuromasts: supraorbital 3 + 3, parietal 1,
anterior rostral 1, posterior rostral 1, infraorbital 1 +12-16 + 1, preorbital 2-3, otic 1, post-otic 1,supratemporal 1, median opercular 1, ventral oper-cular 2, preopercular 2 + 4, mandibular 3 + 1, lateralmandibular 2-3.Interhyal minute, cartilaginous. Basihyal subtrian-
gular, width about 45% length; basihyal cartilageabout 25% basihyal length. Six branchiostegal rays.Second pharyngobranchial teeth absent. Gill rakersof first branchial arch 1 + 9. Vomerine teeth absent.Ventral process of posttemporal absent. Total verte-brae 30-31.
Coloration in life: Male: Side of body metallicblue, bluish black on caudal peduncle; four to fivelongitudinal rows of red dots on anterior half offlank, and red chevron-like oblique bars with ver-tex anteriorly directed on posterior half of flank;dark blue humeral blotch. Dorsum light brown.Venter white. Side of head metallic greenish bluewith small red spots; infraorbital region yellow.Jaws black. Iris brown, anterior and posterior por-tion green. Dorsal fin light blue with dark redreticulation. Anal fin light blue to pale yellow ondistal portion, with grey dots or short bars on basaland posterior portions; narrow dark reddish greydistal stripe. Caudal fin pale blue with five or sixdark red bars not extending to ventralmost portionof fin; usually black ventral stripe. Pectoral fin hya-line. Pelvic fin light blue with dark reddish greyanterior margin.Female: Side of body light grey with small dark
reddish brown spots arranged both in overlappedlongitudinal and oblique rows, often alternatingwith bright greenish blue small spots; dark greyhumeral blotch; dark chromatophores concentratedon longitudinal zone between flank and dorsum,often forming short transverse dark brown barswhen fish exposed to strong sunlight. Dorsum lightbrown. Venter white. Side of head pale blue withsmall dark brown spots; infraorbital region pale yel-low. Jaws black. Iris brown. Dorsal fin pale yellowwith grey dots. Anal fin pale yellow, basal portionlight blue; distal margin dark grey. Caudal fin paleyellow with dark brown dots on upper two thirds offin. Pectoral fin hyaline. Pelvic fin pale yellow withdark grey anterior margin.
DistributionLower Tapajós and lower Trombetas river basins,
and adjacent areas in eastern Amazonas river basin(Fig. 19).
HabitatMargins of clear water streams with moderate cur-
rent, sand bottom, at depths of about 20-40 cm(Fig. 33).
Owiyeye, new subgenus
Type species: Rivulus amanapira Costa, 2004.
DiagnosisDistinguished from all other subgenera of Rivulus
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
Fig. 33. Brazil: Pará: Santarém. clear water stream, habitatof Rivulus dibaphus. Photo by W. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006157
in having the following apomorphic features:absence of dermosphenotic (3.1), frontal scalestransversely arranged (79.0) (Fig. 34A), frontalsquamation S-patterned (80.4; also occurring inAnablepsoides) (Fig. 34A), and a transverse stripethrough chin (98.2; also occurring in Anablep-soides).
EtymologyOwiyeye is the popular name used byTucanos Indi-
ans from the upper rio Negro, Brazil, for R.amanapira (Costa 2004f). This name had alreadybeen reported byWallace (2002) in notes made dur-ing his journey to the Amazon (1850-1852). Gen-der masculine.
Included speciesRivulus altivelis Huber, 1992, R. amanapira Costa,
2004, R. kirovskyi Costa, 2004, R. rectocaudatus Fels& de Rham, 1981, R. romeri Costa, 2003, R. tecmi-nae Thomerson, Nico & Taphorn, 1992, R. uaktiCosta, 2004 and R. uatuman Costa, 2004.
DistributionAmazonas and upper Orinoco river basins, in
Venezuela, Colombia, Brazil and Peru.
Rivulus romeri Costa, 2003
Rivulus romeri Costa, 2003a (type locality: brooktributary to Igarapé Iauiari, about 0º14.31’N,68°03.48’W, which is a tributary of the Uaupésriver, itself a tributary of the Negro river, Ama-zonian basin, Amazonas, Brazil; holotype: MCP29751).
Material examinedBrazil: Estado do Amazonas: MCP 29751, holo-
type; MCP 29752, 1 paratype; UFRJ 5447, 2paratypes; UFRJ 5448, 4 paratypes (c&s); brooktributary to Igarapé Iauiari, a tributary of theUaupés river, itself a tributary of the Negro river,Amazonian basin; U. Römer, 21 Feb. 1995.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anterior por-tion of trunk slightly deeper than wide, jaws short,snout blunt, tip of anal fin pointed in male, caudalfin rounded in male, pelvic-fin tip reaching urogen-ital papilla in male, dorsal-fin origin on verticalbetween base of 6th and 8th anal-fin rays, dorsal-finrays 7-8, anal-fin rays 9-10, frontal squamation S-patterned, frontal scales arranged transversely, canalpreopercular absent, contact organs absent, longitu-dinal series of scales 29-30, gill rakers of firstbranchial arch 1 + 6, longitudinal rows of red dotson flank in male, transverse dark brown bar throughthe chin, and no distinctive black spot on dorsalportion of caudal-fin.
DistributionUpper rio Negro drainage (Fig. 10).
HabitatShallow pools within dense forest (U. Römer, pers.
comm.).
Rivulus kirovskyi Costa, 2004
Rivulus kirovskyi Costa, 2004d: 10 (type locality:
Wilson J. E. M. Costa
Fig. 34. Diagrammatic representation of the frontal squamation patterns in the genus Rivulus: A. R. romeri; B. R. litteratus;C. R. scalaris. Scale bar = 1 mm.

aqua vol. 11 no. 4 - 2006 158
Reserva Ducke, temporary pools near igarapéAcará, lower rio Negro drainage, rio Amazonasbasin, about 3°0’S 60°30’W, Manaus, Estado doAmazonas, Brazil; holotype: UFRJ 5935 [trans-ferred to INPA]).
Rivulus duckensis Hrbek, de Deus & Farias, 2004:571 (Reserva Adolpho Ducke, rivulet emptyinginto Igarapé Barro Branco, Manaus, Amazonas,Brazil, 2º53’S 59º58’W; holotype: INPA 21444).
Material examinedBrazil: Estado do Amazonas: UFRJ 5935, holo-
type; UFRJ 5936, 1 paratype (c&s); Reserva Ducke,temporary pools near igarapé Acará, lower rio Negrodrainage, rio Amazonas basin, Manaus; A. Kirovsky,22 Feb. 1996. UFRJ 5937, 1 paratype, (c&s); UFRJ5938, 1 paratype; same locality and collector, 25Jan. 1996. UFRJ 5939, 2 paratypes; same localityand collector, 1 Feb. 1996. INPA 1850, 2 paratypes;same locality; J. Zuanon, 22 Nov. 1986.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anterior por-tion of trunk slightly wider than deep, jaws short,snout blunt, tip of anal fin rounded in male, caudalfin rounded in male, pelvic-fin tip reaching anteriorportion of anal-fin base in male, dorsal-fin origin onvertical between base of 8th and 9th anal-fin rays,dorsal-fin rays 6-7, anal-fin rays 9-10, frontal squa-mation S-patterned, frontal scales arranged trans-versely, canal preopercular absent, contact organs onflank scales in male, longitudinal series of scales 32,gill rakers of first branchial arch 1 + 7, transverseblack bar through chin, and no black spot on dor-sal portion of caudal-fin.
DistributionMiddle rio Amazonas basin (Fig. 23).
HabitatShallow pools in forest (A. Kirovsky, pers. comm.)
Rivulus uatuman Costa, 2004
Rivulus uatuman Costa, 2004e: 2 (type locality: Bal-bina, temporary pool near stream close to RioUatumã, rio Amazonas basin, 1°56’19.6”S59°59’27.30.6”W, altitude 43 m, Estado do Ama-zonas, Brazil; holotype: UFRJ 6022).
Material examinedBrazil: Estado do Amazonas: UFRJ 6022, holo-
type; UFRJ 6023, 3 paratypes; UFRJ 6024, 2paratypes (c&s); Balbina, temporary pool nearstream close to Rio Uatumã, rio Amazonas basinaltitude 43 m); W. J. E. M. Costa, R. Paiva and D.Ramos, 12 Jun. 2004.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anteriorportion of trunk slightly deeper than wide, jawsshort, snout blunt, tip of anal fin rounded in male,caudal fin rounded in male, pelvic-fin tip reachinganterior portion of anal-fin base in male, dorsal-finorigin on vertical through base of the last anal-finray or slightly posterior to it, dorsal-fin rays 6-7,anal-fin rays 9-10, frontal squamation S-patterned,frontal scales arranged transversely, canal preoper-cular absent, contact organs on flank scales inmale, longitudinal series of scales 31-33, gill rakersof first branchial arch 1 + 6, longitudinal rows ofred dots on flank in male, transverse dark brownbar through the chin, and no distinctive black spoton dorsal portion of caudal fin. Also distinguishedfrom all other species of Owiyeye by having uniquecolour patterns of unpaired fins in male, consistingof dorsal fin dark grey on basal portion and bright
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
Fig. 35. Rivulus uatuman, UFRJ 6022, male, holotype,16.6 mm SL (one day after collection); Brazil: Amazonas:Balbina. Photo by W. J. E. M. Costa.
Fig. 36. Brazil: Amazonas: Balbina; temporary swampwithin forest, the habitat of Rivulus uatuman. Photo by W.J. E. M. Costa.

aqua vol. 11 no. 4 - 2006159
yellow on distal portion, and caudal fin dark greywith white posterior margin (Fig. 35).
DistributionMiddle rio Amazonas basin (Fig. 23).
HabitatIsolated shallow temporary pools within forest,
bottom litter, and water dark tea coloured (Fig. 36).
Rivulus uakti Costa, 2004
Rivulus uakti Costa, 2004f: 2 (type locality: SãoGabriel da Cachoeira, stream near igarapé Iá, km9.4 of the road São Gabriel da Cachoeira to Cucuí,upper rio Negro drainage, rio Amazonas basin,0°3’51.1”S 66°59’46.9”W, altitude 99 m, Estadodo Amazonas, Brazil; holotype: UFRJ 5925).
Material examinedBrazil: Estado do Amazonas: UFRJ 5925, holo-
type; UFRJ 5926, 36 paratypes; UFRJ 5927, 4paratypes (c&s); MCP 34858, 2 paratypes; SãoGabriel da Cachoeira, stream near igarapé Iá, km9.4 of the road São Gabriel da Cachoeira to Cucuí,upper rio Negro drainage, rio Amazonas basin; W. J.E. M. Costa, S. Lima and L. Silva, 30 Aug. 2003.UFRJ 5928, 9; São Gabriel da Cachoeira, upper rioNegro drainage, rio Amazonas basin, stream tribu-
tary to igarapé Miuá, km 13.6 of the road SãoGabriel da Cachoeira to Cucuí, 0°2’58.7”S66°57’48.6”W; W. J. E. M. Costa, S. Lima and L.Silva, 30 Aug. 2003.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anteriorportion of trunk slightly deeper than wide, jawsshort, snout blunt, tip of anal fin slightly pointedin male, caudal fin rounded in male, pelvic-fin tipreaching middle of anal-fin base in male, dorsal-finorigin on vertical between base of 8th and 9th anal-fin rays, dorsal-fin rays 7-8, anal-fin rays 11-12,frontal squamation S-patterned, frontal scalesarranged transversely, canal preopercular absent,contact organs on flank scales in male, longitudinalseries of scales 32-33, gill rakers of first branchialarch 1 + 7-8, oblique rows of red dots on flank inmale, transverse dark brown bar through the chin,and dark grey to black spot on dorsal portion ofcaudal-fin in both sexes. Also differs from all othercongeners of the Owiyeye in possessing two uniquecolour patterns: dorsal fin with bright blue distalzone in male, and caudal spot forming a black barposteriorly bordered by pale yellow bar in female(Figs 37-38).
DistributionUpper rio Negro basin (Fig. 22).
HabitatModerately sunny places of shallow streams within
Wilson J. E. M. Costa
Fig. 38. Rivulus uakti, UFRJ 5926, female, paratype, 27.9mm SL (two days after collection); Brazil: Amazonas: SãoGabriel da Cachoeira. Photo by W. J. E. M. Costa.
Fig. 37. Rivulus uakti, UFRJ 5925, male, holotype, 24.0mm SL (two days after collection); Brazil: Amazonas: SãoGabriel da Cachoeira. Photo by W. J. E. M. Costa.
Fig. 39. Brazil: Amazonas: São Gabriel da Cachoeira; creekat the forest border, the habitat of Rivulus uakti. Photo byW. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006 160
the forest, sandy bottom and reddish hyaline water(Fig. 39).
Rivulus amanapira Costa, 2004
Rivulus amanapira Costa, 2004f: 7 (type locality:São Gabriel da Cachoeira, pools near igarapé
Palestina, Airport road, upper rio Negro drainage,rio Amazonas basin, 0°9’19.2”S 66°59’58.9”W,altitude 110 m, Estado do Amazonas, Brazil; holo-type: UFRJ 5929).
Material examinedBrazil: Estado do Amazonas: UFRJ 5929, holo-
type; UFRJ 5930, 11 paratypes; UFRJ 5931, 3paratypes (c&s); MCP 34859, 2 paratypes; SãoGabriel da Cachoeira, pools near igarapé Palestina,Airport road, upper rio Negro drainage; W. J. E. M.Costa, S. Lima and L. Silva, 30 Aug. 2003.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anterior por-tion of trunk slightly deeper than wide, jaws short,snout blunt, tip of anal fin slightly pointed in male,caudal fin truncate in male, pelvic-fin tip reachinganterior portion of anal-fin base in male, dorsal-finorigin on vertical between base of 11th and 13th anal-fin rays, dorsal-fin rays 8, anal-fin rays 14-15,frontal squamation S-patterned, frontal scalesarranged transversely, canal preopercular absent,contact organs on flank scales in male, longitudinalseries of scales 48-49, gill rakers of first branchialarch 1 + 8, longitudinal rows of red dots on flank inmale, transverse dark brown bar through the chin,and ovoid dark grey to black spot on dorsal portionof caudal fin in both sexes. Additionally distin-guished from all other members of Owiyeye in hav-ing entire marginal region of caudal fin bright yel-low in male (Fig. 40).
DistributionUpper rio Negro drainage (Fig. 23).
Habitat notesSmall shallow temporary pools in the forest,
about 0.5-2.0 m in diameter and about 15 cm deep(Fig. 41).
Rivulus tecminae Thomerson,Nico & Taphorn, 1992
Rivulus tecminae Thomerson, Nico & Taphorn,1992: 290 (type locality: open savanna pools about500 m from left bank, río Guayapo, about 83 kmabove confluence with río Sipapo, Orinoco basin,Amazonas Federal Territory, Venezuela, 4°16’N67°20’W; holotype: MCNG 23886).
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
Fig. 40. Rivulus amanapira, UFRJ 5929, male, holotype,38.6 mm SL (one day after collection); Brazil: Amazonas:São Gabriel da Cachoeira. Photo by W. J. E. M. Costa.
Fig. 41. Brazil: Amazonas: São Gabriel da Cachoeira; shal-low temporary pool within the forest, habitat of Rivulusamanapira. Photo by W. J. E. M. Costa.
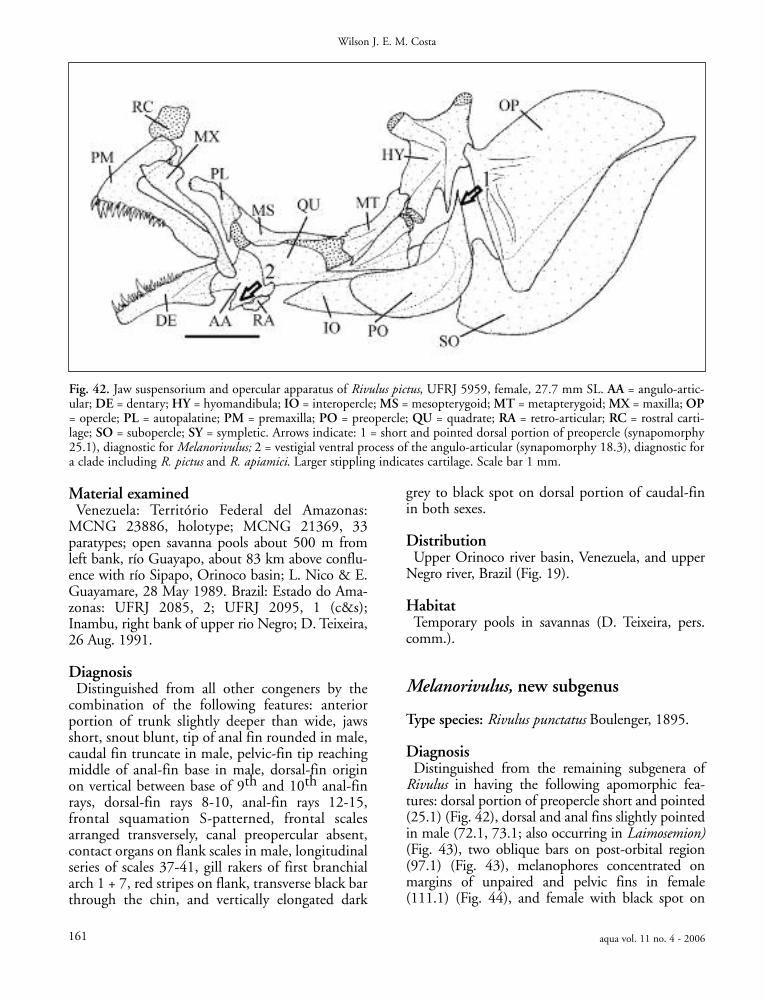
aqua vol. 11 no. 4 - 2006161
Material examinedVenezuela: Território Federal del Amazonas:
MCNG 23886, holotype; MCNG 21369, 33paratypes; open savanna pools about 500 m fromleft bank, río Guayapo, about 83 km above conflu-ence with río Sipapo, Orinoco basin; L. Nico & E.Guayamare, 28 May 1989. Brazil: Estado do Ama-zonas: UFRJ 2085, 2; UFRJ 2095, 1 (c&s);Inambu, right bank of upper rio Negro; D. Teixeira,26 Aug. 1991.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anteriorportion of trunk slightly deeper than wide, jawsshort, snout blunt, tip of anal fin rounded in male,caudal fin truncate in male, pelvic-fin tip reachingmiddle of anal-fin base in male, dorsal-fin originon vertical between base of 9th and 10th anal-finrays, dorsal-fin rays 8-10, anal-fin rays 12-15,frontal squamation S-patterned, frontal scalesarranged transversely, canal preopercular absent,contact organs on flank scales in male, longitudinalseries of scales 37-41, gill rakers of first branchialarch 1 + 7, red stripes on flank, transverse black barthrough the chin, and vertically elongated dark
grey to black spot on dorsal portion of caudal-finin both sexes.
DistributionUpper Orinoco river basin, Venezuela, and upper
Negro river, Brazil (Fig. 19).
HabitatTemporary pools in savannas (D. Teixeira, pers.
comm.).
Melanorivulus, new subgenus
Type species: Rivulus punctatus Boulenger, 1895.
DiagnosisDistinguished from the remaining subgenera of
Rivulus in having the following apomorphic fea-tures: dorsal portion of preopercle short and pointed(25.1) (Fig. 42), dorsal and anal fins slightly pointedin male (72.1, 73.1; also occurring in Laimosemion)(Fig. 43), two oblique bars on post-orbital region(97.1) (Fig. 43), melanophores concentrated onmargins of unpaired and pelvic fins in female(111.1) (Fig. 44), and female with black spot on
Wilson J. E. M. Costa
Fig. 42. Jaw suspensorium and opercular apparatus of Rivulus pictus, UFRJ 5959, female, 27.7 mm SL. AA = angulo-artic-ular; DE = dentary; HY = hyomandibula; IO = interopercle; MS = mesopterygoid; MT = metapterygoid; MX = maxilla; OP= opercle; PL = autopalatine; PM = premaxilla; PO = preopercle; QU = quadrate; RA = retro-articular; RC = rostral carti-lage; SO = subopercle; SY = sympletic. Arrows indicate: 1 = short and pointed dorsal portion of preopercle (synapomorphy25.1), diagnostic for Melanorivulus; 2 = vestigial ventral process of the angulo-articular (synapomorphy 18.3), diagnostic fora clade including R. pictus and R. apiamici. Larger stippling indicates cartilage. Scale bar 1 mm.

aqua vol. 11 no. 4 - 2006 162
upper portion of caudal fin not close to fin margin,overlapping caudal fin bars (113.2) (Fig. 44). Allincluded species except R. modestus have obliquechevron-like rows of red dots or bars on flank inmales (90.2) (Fig. 43).
EtymologyFrom the Latin, melania (black pigmentation on
the skin) and rivulus (stream), referring to the blackmargins of unpaired and pelvic fins, a conditionunique among congeners. Gender masculine.
Included speciesRivulus apiamici Costa, 1989, R. cyanopterus Costa,
2005, R. dapazi Costa, 2005, R. decoratus Costa,1989, R. egens Costa, 2005, R. litteratus Costa,2005, R. modestus Costa, 1991, R. paracatuensisCosta, 2003, R. parnaibensis Costa, 2003, R. pictusCosta, 1989, R. pinima Costa, 1989, R. punctatusBoulenger, 1895, R. rossoi Costa, 2005, R. rutili-caudus Costa, 2005, R. scalaris Costa, 2005, R. vio-laceus Costa, 1991, R. vittatus Costa, 1989, and R.zygonectes Myers, 1927.
DistributionParaná-Paraguay-Uruguay, upper Tapajós, upper
and middle Xingu, upper and middle Tocantins,São Francisco and Parnaíba river basins, in centraland north-eastern Brazil, south-eastern Bolivia,Paraguay and northern Argentina. Greatest diversi-fication concentrated in the central BrazilianPlateau (Costa, 2005a).
Rivulus modestus Costa, 1991
Rivulus modestus Costa, 1991: 329 (type locality: rioMutum, rio Tapajós basin, Mato Grosso, Brazil;holotype: MNRJ 11670).
Material examinedBrazil: Estado de Mato Grosso: MNRJ 11670,
holotype; MNRJ 11671, 5 paratypes; UFRJ 2102,6; UFRJ 2103, 5 (c&s); small stream tributary of rioMutum, at crossing with road BR-364, 51 km fromVilhena, rio Juruena drainage, rio Tapajós basin; K.Tanizaki, M. T. Lacerda, S. O. Kullander & A.Hogerborn-Kullander, 16 Oct. 1989.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anterior por-
tion of trunk deeper than wide, jaws short, snoutblunt, tip of anal fin slightly pointed in male, caudalfin rounded in male, pelvic-fin tip reaching anteriorportion of anal-fin base in male, dorsal-fin origin onvertical between base of 8th and 9th anal-fin rays,dorsal-fin rays 10-11, anal-fin rays 13-14, frontalsquamation F-patterned, frontal scales arranged cir-cularly, canal preopercular absent, contact organsabsent, longitudinal series of scales 31-33, gill rakersof first branchial arch 1 + 7-8, red marks of flankabsent, lower jaw black, and round black spot ondorsal portion of caudal-fin in female.
DistributionUpper rio Juruena drainage (Fig. 10).
Habitat notesShallow creeks with slow current, in moderately
sunny region.
Rivulus zygonectes Myers, 1927
Rivulus zygonectes Myers, 1927: 127 (type locality:Vereda Extrema, into Cannabrava [now CanaBrava, at rio Tocantins left bank], Goyas [nowEstado de Goiás], Brazil; lectotype: CAS 76314,designated by Huber, 1992: 464).
Material examinedBrazil: Tocantins-Araguaia basin: Estado de Goiás:
CAS 76314, lectotype; Vereda Extrema into CanaBrava; C. Ternetz, 14 Jan. 1924. UFRJ 1575, 3;stream 15 km E of Aruanã; W. J. E. M. Costa et al.,29 Aug. 1993. UFRJ 1748, 6; stream tributary torio Verde, 30 km E of São Miguel do Araguaia; W.J. E. M. Costa et al., 25 Aug. 1993. UFRJ 1567, 5;stream 98 km W of Jussara; W. J. E. M. Costa et al.,30 Aug. 1993. UFRJ 1482, 22; UFRJ 2108, 2(c&s); stream tributary to rio Verde, 32 kmN of SãoMiguel do Araguaia; W. J. E. M. Costa et al., 25Aug. 1993. MZUSP 35416, 20; MZUSP 38334,55; pool in Aruanã; W. J. E. M. Costa et al., 28 Jan.1986. Estado do Tocantins: UFRJ 2106, 6; UFRJ2107, 4 (c&s); stream 20 km S of Santa Rosa; W. J.E. M. Costa et al., 15 Feb. 1994. MZUSP 38375, 2;stream tributary to córrego Dona Francisquinha,Porto Nacional; W. J. E. M. Costa et al., 31 Jan.1986. UFRJ 1712, 7; stream tributary to rio Verde,50 km N of São Miguel do Araguaia; W. J. E. M.Costa et al., 25 Aug. 1993. UFRJ 1597, 2; stream 12km N of Sandolândia; W. J. E. M. Costa et al., 27
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin

aqua vol. 11 no. 4 - 2006163
Aug. 1993. MZUSP 37212, 28 (6 c&s); roadbetween road BR-153 and Formoso do Araguaia, 29km S of Gurupi; P. S. Santos-Filho, Jun. 1976.MZUSP 45223, 9; rio Água Fria, road Araguaçu-Barreira do Piqui, 27 km N of Araguaçu; F. C. T.Lima, 21-26 Feb. 1993. UFRJ 1374, 13; shallowlagoon 2 km W of rio das Mortes at the road ÁguaBoa-Cocalinhos; W. J. E. M. Costa et al., 20 Feb.1993. UNT 555, 5; river at the road BR-153between Filadelphia and Araguaina; E. L. Beerli, 2Nov. 2003. UNT 556, 3; rio Brejão, Araguaina; E.L. Beerli & L, M, Lima, 22 Nov. 2003. UNT 2045,1; córrego Gorgulho, Porto Nacional; NEAMB, 14Sep. 2001. UNT 2046, 1; córrego Gorgulho, PortoNacional; NEAMB, 14 Sep. 2001. UNT 2047, 3;córrego Marimbondo, Tupirantins; NEAMB, 27Oct. 2000. UNT 2048, 1; córrego Barreiro, Guaraí;NEAMB, 20 Oct. 2000. UNT 2049, 2; córregoGorgulho, Porto Nacional; NEAMB, 14 Sep. 2001.UNT 2050, 1; córrego Gorgulho, Porto Nacional;NEAMB, 14 Sep. 2001. UNT 2051, 1; córregoLageado, Paranã; NEAMB, 22 Mar. 1999. UNT2052, 9; córrego Água Suja, Tupirantins; NEAMB,27 Oct. 2000. UFRJ 5150, 1; floodplains of leftbank of rio Tocantins, about 1200 m from the rivermargin, SSE Sampaio, 5°23’00”S 47°51’41”W; G.C. Brasil, 8 Jun. 2000. Estado do Maranhão: UFRJ5152, 16; swamp about 500 m from the right mar-gin of rio Tocantins, 5°12’33”S 48°27’59”W; G. C.Brasil, 8 Jun. 2000. Rio Xingu basin, Estado deMato Grosso: UFRJ 1386, 1; stream 67 km N ofParanatinga; W. J. E. M. Costa et al., 10 Feb. 1993.UFRJ 1173, 5; stream at the road BR-080, 9 km Eof São José do Xingu; W. J. E. M. Costa et al., 16Feb. 1993. UFRJ 1417, 1; stream at the road BR-080, 2 km E of São José do Xingu;W. J. E. M. Costaet al., 18 Feb. 1993. UFRJ 1389, 6; stream at theroad BR-080, 41 km E of São José do Xingu; W. J.E. M. Costa et al., 17 Feb. 1993. UFRJ 1420, 6;stream at the road BR-080, 42 km E of São José doXingu;W. J. E. M. Costa et al., 17 Feb. 1993. UFRJ1392, 7; stream at the road BR-080, 36 km E of SãoJosé do Xingu; W. J. E. M. Costa et al., 17 Feb.1993. UFRJ 1357, 20; stream at the road Para-natinga-Canarana, 158 km of Paranatinga; W. J. E.M. Costa et al., 12 Feb. 1993. UFRJ 1414, 20;UFRJ 2109; stream at the road BR-080, 7 km E ofrio Xingu margin; W. J. E. M. Costa et al., 16 Feb.1993. UFRJ 6265, 1; stream near rio Xingu,Altamira; W. J. E. M. Costa et al., 16 Jun. 2004. RioTapajós basin, Estado de Mato Grosso: MZUSP45304, 10; upper rio Preto, road Cuiabá-Santarém;
N. Menezes et al., 24 Oct. 1992. MZUSP 45321, 2;stream tributary to rio Preto, road to São Francisco;N. Menezes et al., 24 Oct. 1992.
DiagnosisDistinguished from all other congeners by the com-
bination of the following features: anterior portion oftrunk slightly deeper than wide, jaws short, snoutblunt, tip of anal fin slightly pointed in male, caudalfin rounded in male, pelvic-fin tip reaching anteriorportion of anal-fin base in male, dorsal-fin origin onvertical between base of 8th and 10th anal-fin rays,dorsal-fin rays 9-11, anal-fin rays 13-15, frontalsquamation F-patterned, frontal scales arranged cir-cularly, canal preopercular absent, contact organsabsent, longitudinal series of scales 33-35, gill rakersof first branchial arch 1 + 7-8, oblique rows of reddots sometimes united forming chevron-like bars onflank in male, lower jaw black, and round black spoton dorsal portion of caudal fin in female.
DistributionTocantins, Araguaia, Xingu and Tapajós river
basins (Fig. 22).
Habitat notesShallow creeks with slow current, in gallery forest
of savanna-like region.
Rivulus violaceus Costa, 1991
Rivulus violaceus Costa, 1991: 331 (type locality: riodas Mortes, rio Araguaia-Tocantins basin, MatoGrosso, Brazil; holotype: MNRJ 11672).
Material examinedBrazil: Estado de Mato Grosso, rio das Mortes
basin: MNRJ 11672, holotype; MNRJ 11673, 5paratypes; UFRJ 2104, 20; rio das Mortes whencrossed by BR-163 at km 313; K. Tanizaki, M. T.Lacerda, S. O. Kullander & A. Hogerborn-Kullan-der, 19 Oct. 1989. UFRJ 143, 16; UFRJ 2105, 7(c&s); small stream tributary to rio Perdidos, closeto BR-070, about 10 km W of Primavera do Leste;K. Tanizaki, M. T. Lacerda, S. O. Kullander & A.Hogerborn-Kullander, 19 Oct. 1989. UFRJ 1207,4; rio Suspiro, 57 km S of Paranatinga; W. J. E. M.Costa, C. P. Bove, R. D. Cunha & C. Muratori, 9Feb. 1993. UFRJ 4284, 13; idem; M. Britto, C.Moreira & R. Cunha, 28 Oct. 1996. UFRJ 4283,3; stream at the road Primavera do Leste-Para-
Wilson J. E. M. Costa

aqua vol. 11 no. 4 - 2006 164
natinga; M. Britto, C. Moreira & R. Cunha, 28Oct. 1996.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anterior por-tion of trunk slightly deeper than wide, jaws short,snout blunt, tip of anal fin slightly pointed in male,caudal fin rounded in male, pelvic-fin tip reachingurogenital papilla in male, dorsal-fin origin on verti-cal between base of 8th and 9th anal-fin rays, dorsal-fin rays 10-11, anal-fin rays 14-15, frontal squama-tion F-patterned, frontal scales arranged circularly,canal preopercular absent, contact organs absent,longitudinal series of scales 30-32, gill rakers of firstbranchial arch 1 + 7-8, oblique chevron-like red barson flank in male, lower jaw black, and round blackspot on dorsal portion of caudal fin in female.
DistributionUpper rio das Mortes drainage (Fig. 10).
Habitat notesShallow creeks with slow current, in gallery forest
of savanna-like region.
Rivulus litteratus Costa, 2005
Rivulus sp.: Lacerda, 1989: 25 (Ponte Branca, RioAraguaia, central Brazil).
Rivulus pictus (non Costa): Costa et al., 2003: 143(misidentification of specimens from Alto Ara-guaia).
Rivulus litteratus Costa, 2005a: 75 (type locality:Município de Alto Araguaia, Córrego do Sapo,upper Rio Araguaia basin, road MT-100, 31 km Sof Alto Araguaia, 17º33’38.5”S 53º18’33.1”W,altitude 750 m, Estado de Mato Grosso, Brazil;holotype: UFRJ 5956).
Material examinedBrazil: Estado de Mato Grosso: Município de Alto
Araguaia, upper rio Araguaia basin: UFRJ 5956,holotype; Município de Alto Araguaia, córrego doSapo, upper rio Araguaia basin, road MT-100, 31km S of Alto Araguaia; W. J. E. M. Costa, B. B.Costa and C. P. Bove, 14 Jan. 2004.
DiagnosisDistinguished from all other congeners by the
combination of the following features: anterior por-tion of trunk slightly deeper than wide, jaws short,snout blunt, tip of anal fin slightly pointed in male,caudal fin rounded in male, pelvic-fin tip reachinganterior portion of anal-fin base in male, dorsal-finorigin on vertical between base of 7th 8th l-fin rays,dorsal-fin rays 9-10, anal-fin rays 13-14, frontalsquamation F-patterned, frontal scales arranged cir-cularly, canal preopercular absent, contact organsabsent, longitudinal series of scales 31-32, gill rakersof first branchial arch 1 + 7-8, overlapped red marksof variable shape on flank in male, lower jaw darkgrey, and round black spot on dorsal portion of cau-dal-fin in female. Also distinguished from the
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin
Fig. 43. Rivulus litteratus, UFRJ 5956, male, holotype,26.9 mm SL (one day after collection); Brazil: MatoGrosso: Alto Araguaia. Photo by W. J. E. M. Costa.
Fig. 44. Rivulus litteratus, UFRJ 5957, female, paratype,25.6 mm SL (one day after collection); Brazil: MatoGrosso: Alto Araguaia. Photo by W. J. E. M. Costa.
Fig. 45. Brazil: Mato Grosso: Alto Araguaia; savanna creekclose to córrego do Sapo, habitat of Rivulus litteratus.Photo by W. J. E. M. Costa.

aqua vol. 11 no. 4 - 2006165
remaining species of Melanorivulus by possessing acolour pattern consisting of red marks variable inshape and highly overlapped (Fig. 43).
DistributionUpper rio Araguaia drainage (Fig. 23).
Habitat notesShallow creeks with slow current, in gallery forest
of savanna-like region (Fig. 45).
DiscussionThe present study confirms a weakly supported
genus Rivulus containing clades corroborated bothby morphological (e.g. Huber 1992, Costa 1998a,the present study) and molecular data (e.g. Murphyet al. 1999, Hrbek & Larson 1999), and recognizedas subgenera. No unambiguous, apomorphic mor-phological condition was found to be shared by allspecies presently assigned to Rivulus and molecularstudies indicate that Rivulus is a paraphyletic assem-blage (Murphy et al. 1999, Hrbek & Larson 1999).However, a group comprising all species of Rivulusexcept the subgenus Rivulus is well supported bymorphology, their members having a uniquelyderived pattern of arrangement of rostral neuro-masts (i.e. transversely placed), and a long neuralprezygapophysis in caudal vertebrae as already dis-cussed in former studies (e.g. Costa 1998a), alsooccurring in Kryptolebias.The subgenus Rivulus, as herein defined, is also cor-
roborated by a molecular analysis (Murphy et al.1999: 295). In other molecular analyses, R. roloffi isconsidered to be more closely related to other nomi-nal species of Rivulus than to R. cylindraceus (e.g.Hrbek & Larson 1999). As R. roloffi, which was notaccurately examined in the present study (only exam-ined at the field just after collection), have more thanhalf caudal fin scaled and a dark green humeral spot,which constitute two synapomorphies of the sub-genus Rivulus, R. roloffi is inserted in this subgenus.Although weakly supported by morphological fea-
tures, Cynodonichthys as proposed here is also cor-roborated by molecular data (Murphy et al. 1999:295). However, some molecular analyses do notsupport monophyly of this assemblage (e.g. Hrbek& Larson 1999). Further analyses should be con-ducted in order to test the monophyly of this geo-graphically widespread assemblage.Three species of Cynodonichthys from the Brazilian
Amazonas basin were found in collections examinedin the present study: R. urophthalmus, R. taeniatus
and R. micropus. A fourth species, R. xanthonotus, hasbeen reported for the Brazilian Amazon. However,this species, which seems to be similar to R. uroph-thalmus, is only known from its poor original descrip-tion (Ahl 1926), where the type locality is inaccu-rately described (i.e. “Amazon Strom”). Huber (1992)reported that types exhibit an uncommon posteriorlypositioned dorsal fin, thus constituting a validspecies. No specimen potentially identifiable as R.xanthonotus was examined in the present study.Rivulus urophthalmus was found in an extensive
region encompassing the lower Amazonas basin andtributaries, and adjacent coastal areas to East, includ-ingMaranhão, the type locality R. auratus. This latterspecies is therefore considered a synonym of R. uroph-thalmus, as already proposed by Huber (1992).Rivulus poey was described by Steindachner (1876)
based on specimens from Belém. The brief descrip-tion includes characters congruent with R. urophthal-mus, previously described from the same locality. Asthe types of R. poey are unknown, this highly proba-ble synonymy cannot be confirmed. In order to solvethis problem, a neotype from the city of Belém isherein designated for R. poey, becoming a clear syn-onym of R. urophthalmus, as already proposed byother authors (e.g. Garman 1895, Huber 1991).A geographically widespread species of the central
and western Brazilian Amazon is here tentativelyidentified as R. taeniatus, a species first describedfrom the Colombian Amazon and still poorlyknown. However, the species collected in some dis-tant points of the Brazilian Amazon also fits wellwith other nominal species from the Peruvian andEcuadorian Amazon, such as R. rubrolineatus and R.limoncochae, which possibly are synonyms of R. tae-niatus. These species have been distinguished mainlyon the basis of the number of rows of red dots on thecaudal peduncle (i.e. three or five) (e.g. Huber1992). However, this character was variable withinpopulations both of R. urophthalmus and R. taenia-tus (see descriptions above).Steindachner (1863) described R. micropus based
upon a single specimen collected in the rio Negro,Brazilian Amazon. The identity of this species wasobscure until publication of photos and new mor-phological data on the holotype (Huber 1991).Henn (1916) described a similar species, R. compres-sus, from Manaus, a city at the confluence of rioNegro and rio Solimões, central Brazilian Amazon.At the beginning of the description of R. compressus,Henn (1916: 111) noted that: “This may be R.micropus Steidachner, but it seems to differ in the
Wilson J. E. M. Costa

aqua vol. 11 no. 4 - 2006 166
more forward position of the dorsal, the longerhead, etc.”. However, Huber (1992) examined typesof both species and concluded that these diagnosticcharacters are similar in both nominal species. Thissynonymy follows Costa (2003b), supported bymaterial collected in the type locality region, havingpointed snout and posteriorly positioned dorsal finas observed in holotypes of R. micropus (Huber,1991: fig. 2) and R. compressus (Henn, 1916: fig. 1).Anablepsoides was first proposed to include only R.
atratus (Huber, 1992). However, morphologicaldata (Costa 1998a, the present study) consistentlycorroborated a clade containing both R. atratus andR. ornatus. This hypothesis has not yet been testedby molecular analyses, since both included specieswere not simultaneously included in these studies(e.g. Murphy et al. 1999, Hrbek & Larson 1999).Examination of types revealed that R. ornatus and
R. obscurus, respectively, refer to the male and femaleof the same species, thus constituting synonyms(Costa 2003b). Rivulus ornatus has been identifiedas R. obscurus (e.g. Huber 1992), whereas an unde-scribed species from Peru has been misidentified asR. ornatus (e.g. Fels & de Rham 1982, Huber 1992).The specimen mentioned by Garman (1895: 139)under the description of R. ornatus, as possiblybelonging to another species, is in fact a member ofthe poeciliid genus Fluviphylax Whitley (Costa1996, Costa & Le Bail 1999).Benirivulus is erected to R. beniensis, which does
not fit with any other subgenus of Rivulus. The pre-sent phylogenetic hypothesis indicates that Benirivu-lus is the sister group to a clade including three sub-genera, Laimosemion, Owiyeye, and Melanorivulus,each of them corroborated by both morphologicaland molecular data. The only species of Benirivulus,R. beniensis, was poorly known until now, and con-sequently was not included in molecular analyses.Myers (1927b) erected R. beniensis on the basis ofspecimens collected in the drainage of the Beni-Mamoré drainage, rio Madeira basin, and previouslymisidentified by Pearson (1925) as R. strigatus.Based on putative minor differences in dorsal-finorigin position, Myers (1927b) recognized two sub-species from distinct localities, R. beniensis beniensisand R. beniensis lacustris. Seegers (1985) described R.bolivianus also from the río Mamoré drainage.Examination of material from distant localitiesalong the rio Madeira basin, including the Mamoré,Beni and Guaporé river drainages, revealed that R.beniensis is a widespread species, indistinguishablefrom R. beniensis lacustris and R. bolivianus, thus
confirming the synonymy proposed by Costa(2003b).Huber (1999) erected Laimosemion to include
species of the “R. geayi superspecies” as proposed byHuber (1992). Molecular studies (Murphy et al.1999, Hrbek et al. 2004) corroborated monophyly ofa group comprising species of the “R. geayi super-species”, “R. breviceps superspecies”, and “R. frenatussuperspecies”, which is in accordance to the presentstudy. Two species of Laimosemion are found in theBrazilian Amazonas river: R. strigatus and R. dibaphus.Rivulus strigatus was described by Regan (1912)
based on a single specimen donated by Arnold. Thismaterial was obtained from an aquarium fish ship-ment from the city of Pará (now Belém, capital ofEstado do Pará) (Arnold 1913). Huber (1992) men-tioned rio da Prata, a tributary of rio Jari, Pará, Brazil,as the true type locality of R. strigatus, based on amis-interpreted citation by von Ihering (1931). The lat-ter author, under a brief diagnosis of R. strigatus,cited: “Distribuição: (typo) Amazonas, sem mais indi-cação além do nome do colleccionador Arnold, mas cujomaterial provem principalmente do rio da Prata” (Dis-tribution: (type) Amazonas, without further indica-tion besides the collector name Arnold, but thatmaterial mainly originates from rio da Prata) (vonIhering 1931: 263). In fact, von Ihering referred tothe río de La Plata basin, from where R. punctatusBoulenger was often misidentified as R. strigatus inthe past (e.g. Ringuelet et al. 1967), not to the smallrio da Prata in Pará, unknown for most ichthyolo-gists and still almost inaccessible at present. The geo-graphic position of the “rio da Prata” mentioned byvon Ihering may be clarified when examining otherparts of his paper. For example, among other locali-ties, he cited the occurrence of Phalloceros caudimac-ulatus (Hensel) for the “rio da Prata, Paraguay” (vonIhering 1931: 246).Huber (1992) considered both R. strigatus and R.
dibaphus as synonyms of R. geayi, a species firstdescribed from Amapá, northern Brazil, but alsooccurring in the Guianas (e.g. Huber 1991, 1992).According to Huber (1991), the three nominalspecies would have near type localities, but in fact thetype locality of R. geayi is about 480 km and 560 kmin a straight line from the type locality of R. strigatusand R. dibaphus, respectively, and type locality of R.strigatus is about 710 km from the type locality of R.dibaphus. Comparison of material of R. strigatus withmaterial from the type locality region of R. geayirevealed that R. strigatus is a distinct and valid species,easily distinguished from R. geayi by having a
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin

aqua vol. 11 no. 4 - 2006167
rounded caudal fin in the male (vs. subtruncate) andfrontal squamation F-patterned (vs. usually E-pat-terned). Rivulus dibaphus is also a valid species.Examination of the type material of R. dibaphus andmaterial collected in the type locality region revealedthat it may be readily distinguished from R. strigatusand R. geayi by the unique colour pattern on the cau-dal peduncle in the male (Figs 31-32) and that thefrontal squamation is always E-patterned.Owiyeye corresponds to the “R. rectocaudatus species
group” diagnosed by Thomerson et al. (1992) andthe “R. rectocaudatus superspecies” diagnosed byHuber (1992). This clade is well supported both bymorphological (the present study) and moleculardata (Murphy et al. 1999, Hrbek & Larson 1999,Hrbek et al. 2004). Owiyeye and Anablepsoides sharetwo derived conditions (frontal squamation S-pat-terned and a transverse stripe through chin), but it isparsimoniously considered to a member of a cladealso including Laimosemion and Melanorivulus,mainly diagnosed by the absence of second pharyn-gobranchial teeth. Seven species of Owiyeye are foundin the Brazilian Amazon, all described in recent years:R. romeri, R. kirovskyi, R. uatuman, R. uakti, R.amanapira, and R. tecminae. Rivulus kirovskyi and R.duckensis are identical and were collected at the samelocality. Both original descriptions were published in2004, but the name R. kirovskyi has chronologicalpriority over R. duckensis, since the former was pub-lished on 30 April, the latter on 20 August.Melanorivulus corresponds to the “R. punctatus
superspecies” diagnosed by Huber (1992) or the“Rivulus punctatus species-complex” diagnosed byCosta (1995a). This clade is strongly supported bymorphology, and is also corroborated by molecularstudies (e.g. Murphy et al. 1999, Hrbek & Larson1999). It includes four species endemic to theBrazilian Amazonas basin: R. modestus, R. zygonectes,R. violaceus, and R. litteratus.As already noted in previous studies (e.g. Costa
1998a, Murphy et al. 1999, Hrbek & Larson 1999),species of Rivulus endemic to the Amazonas basindo not form a monophyletic assemblage. Speciesfrom southern Amazonas tributaries, running on theBrazilian Shield (i.e., belonging to Melanorivulus)are more closely related to species endemic to basinsto South and East (i.e., Paraná, Paraguay, São Fran-cisco and Parnaíba river basins). On the other hand,species occurring in northern and western Ama-zonas tributaries arising from the Andes or theGuyana Shield (i.e., belonging to Cynodonichthys,Laimosemion and Owiyeye) are more closely related
to species endemic to the Orinoco and Guianasbasin. This is probably a historic consequence of thepast isolation of rivers draining the Brazilian Shieldbefore the formation of the present Amazonas basin,as described in geological studies (e.g. Beurlen 1970,Lundberg et al. 1998).Field observation on the habitat of Amazonian
species of Rivulus revealed that different lineagesoccupy distinct kinds of habitat. This is most clearfor the clade Benirivulus + Laimosemion + Owiyeye +Melanorivulus, in which members of each genus arefound in a particular kind of aquatic environment,thus suggesting divergent specializations during theevolution of the group.In Benirivulus, individuals were collected near the
margin of stagnant streams and lakes (about 50-150cm deep), with dense aquatic vegetation, at the for-est border (Fig. 25). Species of Laimosemion, Owiy-eye and Melanorivulus were collected in shallowerplaces (about 5-40 cm). Possibly, the ecological pref-erence noted for Benirivulus is the primitive condi-tion for the clade Benirivulus + Laimosemion +Owiyeye + Melanorivulus, since species of Anablep-soides, the putative sister group to the above men-tioned clade, live in similar habitats.Amazonian species of Laimosemion were uniquely
found in clear water streams, with moderate currentand sandy bottom (Figs 29 and 33). No otherspecies from the Amazonas basin inhabits a similarenvironment, but R. xiphidius and R. geayi, twospecies of Laimosemion from Guyana, seem to live inthe same environmental conditions (Huber 1979).However, R. agilae, another species of Laimosemionfrom Guyana, has been reported as living in sunnyplaces (Huber 1979).Only species of Owiyeye were found in extremely
shallow temporary pools (about 5-10 cm deep) withtea-coloured water (Figs 36 and 41). Since somespecies of Owiyeye were uniquely found in tempo-rary pools, they possibly are semiannual fishes. Anexception is R. uakti, which inhabit marginal zonesof creeks at the forest border (Fig. 39).Species of Melanorivulus are found in sunny creeks
(Fig. 45) in savanna-like environments. However,this is not a unique condition among AmazonianRivulus, since R. urophthalmus, R. taeniatus, and R.micropus are also found in sunny creeks, often nearthe border of forests (Figs 11, 17 and 20). Theseconditions are interpreted as independently derived,because Melanoorus and Cynodonichthys are notclosely related taxa, and basal rivulids (i.e. basalspecies of Kryptolebias and Prorivulus) inhabit dark
Wilson J. E. M. Costa

aqua vol. 11 no. 4 - 2006 168
forested habitats (Costa 2004a, 2004c, Vermeulen& Hrbek 2005).
AcknowledgementsThanks are due to E. Araujo, F. Autran, S. Barrera,
D. Belote, C. P. Bove, G. C. Brasil, M. Britto, C.Campinha, B. B. Costa, R. D. Cunha, R. D’Arrigo,M. I. Landim, S. M. Q. Lima, C. Moreira, R. Paiva,A. Pinto, D. Ramos, A. Sarraf, K. Tanizaki, E.Vicente, and L. Villa-Verde, for help in collectingtrips; to H. Berkenkamp, G. Brasil, H. Britski, J.Carvalho, D. Catania, J. L. Figueiredo, K. Hartel, J.Huber, M. Kottelat, S. Kullander, M. Lacerda, P.Lucinda, N. Menezes, D. Moraes Jr., G. Nunan, O.Oyakawa, L. Parenti, C. Pereira, L. Py-Daniel, U.Römer, J. Sarmiento, D.Taphorn, D.Teixeira, and J.Zuanon, for the loan, exchange, or donation of spec-imens, or for support during visits to their institu-tions. Collecting trips were supported by Fundação oBoticário de Proteção à Natureza. This study wasfunded by CNPq-MCT and FAPERJ. Material wascollected with permits 02001.001660/98 and02.022005956/02 from IBAMA (Instituto Brasileirodo Meio Ambiente e dos Recursos Naturais Ren-ováveis – Ministério do Meio Ambiente, dos Recur-sos Hídricos e da Amazônia Legal).
ReferencesARNOLD, J. P. 1913. Rivulus strigatus, sp. n. Rgn. Wochen-
schrift für Aquarien-und Terrarienkunde 10: 353-355.BERKENKAMP, H. O. &. ETZEL, V. 1992. Die Rivulus-Artenvon Panama 2: Wiederbeschreibung von Rivulus brunneusMeek & Hildebrand, 1913 und Beschreibung von Rivulusbirkhahni spec. nov. von Chiquiri Grande, West-Panama.Deutsche Killifisch Gemeinschaft Journal 24: 97-104.
BERKENKAMP, H. O. &. ETZEL, V. 1993a. Die Rivulus-Artenvon Panama 3: Rivulus kuelpmanni spec. nov., eine neueBachlingsart von Punta Peña, West-Panama. Deutsche Killi-fisch Gemeinschaft Journal 25: 17-24.
BERKENKAMP, H. O. &. ETZEL, V. 1993b. Die Rivulus-Artenvon Panama 4: Rivulus frommi spec. nov., eine neue Bach-lingsart von El Valle, Zentral-Panama. Deutsche KillifischGemeinschaft Journal 25: 81-89.
BERKENKAMP, H. O. &. ETZEL, V. 1995. Die Rivulus-Artenvon Panama 5: Rivulus monikae spec. nov., eine weitere neueBachlingsart aus den bewaldeten und bergigen Teilen derProvinz Coclé, Zentral-Panama. Deutsche Killifisch Gemein-schaft Journal, 27: 1-7.
BERKENKAMP, H. O. &. ETZEL, V. 1997. Die Rivulus-Artenvon Panama 6: Zur Identität von Rivulus montium Hilde-brand, 1938 mit der Beschreibung einer neuen Art, Rivulusvillwocki spec. nov. Deutsche Killifisch Gemeinschaft Journal29: 125-136.
BERKENKAMP, H. O. &. ETZEL, V. 1999. Die Rivulus-Arten
von Panama 7: Rivulus wassmanni spec. nov., eine neueBachlingsart der Rivulus birkhahni-kuelpmanni-Gruppe ausder Provinz Bocas del Toro, Nordost-Panamá. AquaristikAktuell 1999: 62-66.
BEURLEN, K. 1970. Geologie von Brasilien. Bornträger,Berlin, 444 pp.
BUSSING, W. A. 1980. Status of the cyprinodontid fish genusRivulus in Costa Rica, with descriptions of new endemicspecies. Brenesia 17: 327-364.
COSTA, W. J. E. M. 1989. Descrição de cinco novas espéciesde Rivulus das bacias dos rios Paraná e São Francisco(Cyprinodontiformes, Rivulidae). Revista Brasileira deZoologia 6: 523-533.
COSTA, W. J. E. M. 1990. Análise filogenética da famíliaRivulidae (Cyprinodontiformes, Aplocheiloidei). RevistaBrasileira de Biologia 50: 65-82.
COSTA, W. J. E. M. 1991a. Redescrição do gênero Rivulus(Cyprinodontiformes: Rivulidae), com notas sobre R. stel-lifer e R. compactus e a descrição de duas novas espécies doBrasil central. Revista Brasileira de Biologia 51: 327-333.
Costa, W. J. E. M. 1991b. Description of two new species ofthe genus Rivulus (Cyprinodontiformes: Rivulidae) fromeastern South American coastal plains. Revue Suisse de Zoolo-gie 98: 581-587.
COSTA, W. J. E. M. 1995a. Revision of the Rivulus punctatusspecies-complex (Cyprinodontiformes: Rivulidae). Ichthy-ological Exploration of Freshwaters 3: 207-226.
COSTA, W. J. E. M. 1995b. Pearl killifishes – the Cynolebiati-nae: systematics and biogeography of the neotropical annual fishsubfamily. TFH, Neptune City, 128 pp.
COSTA, W. J. E. M. 1996. Relationships, monophyly andthree new species of the neotropical miniature poeciliidgenus Fluviphylax (Cyprinodontiformes: Cyprinodon-toidei). Ichthyological Exploration of Freshwaters 7: 111-130.
COSTA, W. J. E. M. 1998a. Phylogeny and classification ofRivulidae revisited: evolution of annualism and miniatur-ization in rivulid fishes (Cyprinodontiformes: Aplocheiloi-dei). Journal of Comparative Biology 3: 33-92.
COSTA, W. J. E. M. 1998b. Phylogeny and classification ofthe Cyprinodontiformes (Euteleostei: Atherinomorpha): areappraisal. In: Malabarba, L. R., Reis, R. E., Vari, R. P.,Lucena, Z. M. S. & Lucena, C. A. S. (Eds.). Phylogeny andclassification of neotropical fishes, Edipucrs, Porto Alegre, pp.537-560.
COSTA, W. J. E. M. 2001. The neotropical annual fish genusCynolebias (Cyprinodontiformes: Rivulidae): phylogeneticrelationships, taxonomic revision and biogeography. Ichthy-ological Exploration of Freshwaters 12: 333-383.
COSTA, W. J. E. M. 2003a. Family Rivulidae (South Ameri-can annual fishes). In: R. E. Reis, Kullander, S. O. & Fer-raris, C. J., Jr. (eds) Check list of the freshwater fishes of Southand Central America, Edipucrs, Porto Alegre, Pp 526-548.
COSTA, W. J. E. M. 2003b. A new miniature rivuline fishfrom the upper Negro river basin, northern Brazil(Teleostei, Cyprinodontiformes, Rivulidae). Arquivos doMuseu Nacional, Rio de Janeiro 61: 175-178.
COSTA, W. J. E. M. 2003c. Rivulus paracatuensis n. sp.(Cyprinodontiformes: Rivulidae): a new rivuline species
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin

aqua vol. 11 no. 4 - 2006169
from the Rio São Francisco basin, Brazil. aqua Journal ofIchthyology and Aquatic Biology 7: 39-43.
COSTA,W. J. E.M. 2004a. Relationships and redescription ofFundulus brasiliensis (Cyprinodontiformes: Rivulidae), withdescription of a new genus and notes on the classification ofthe Aplocheiloidei. Ichthyological Exploration of Freshwaters15: 105-120.
COSTA,W. J. E.M. 2004b. Kryptolebias, a substitute name forCryptolebias Costa, 2004 and Kryptolebiatinae, a substitutename for Cryptolebiatinae Costa, 2004 (Cyprinodontif-ormes: Rivulidae). Neotropical Ichthyology 2: 107-108.
COSTA, W. J. E. M. 2004c. A new killifish genus and speciesfrom the coastal plains of northeastern Brazil (Teleostei:Cyprinodontiformes: Rivulidae). Zootaxa 642: 1-10.
COSTA, W. J. E. M. 2004d. Rivulus kirovskyi, a new killifishfrom the central Amazon, Brazil (Cyprinodontiformes:Rivulidae). Neotropical Ichthyology 2: 9-12.
COSTA,W. J. E. M. 2004e. Rivulus uatuman sp. n. (Teleostei:Cyprinodontiformes: Rivulidae): a new miniature killifishfrom the central Amazon. Zootaxa 696: 1-8.
COSTA, W. J. E. M. 2004f. Rivulus uakti sp. n. and R.amanapira sp. n. (Teleostei: Cyprinodontiformes: Rivuli-dae): two new species from the upper Rio Negro, BrazilianAmazon. Zootaxa 465: 1-12.
COSTA, W. J. E. M. 2004g. Rivulus simplicis n. sp. (Cyprin-odontiformes: Rivulidae): a new killifish from the coastalplains of south-eastern Brazil. aqua Journal of Ichthyologyand Aquatic Biology 8: 103-108.
COSTA, W. J. E. M. 2005a. Seven new species of the killifishgenus Rivulus (Cyprinodontiformes: Rivulidae) from theParaná, Paraguay and upper Araguaia river basins, centralBrazil. Neotropical Ichthyology 3: 69-82.
COSTA, W. J. E. M. 2005b. The Neotropical annual killifishgenus Pterolebias Garman (Teleostei: Cyprinodontiformes:Rivulidae): phylogenetic relationships. descriptive morphol-ogy, and taxonomic revision. Zootax 1067: 1-36.
COSTA, W. J. E. M. & BRASIL, G. C. 1991. Description of anew species of Rivulus (Cyprinodontiformes: Rivulidae)from the coastal plains of eastern Brazil. Ichthyological Explo-ration of Freshwater 1: 379-383.
COSTA, W. J. E. M. & LE BAIL, P.-Y. 1999. Fluviphylaxpalikur: a new poeciliid from the rio Oiapoque basin,northern Brazil (Cyprinodontiformes: Cyprinodon-toidei), with comments on miniaturization in Fluviphy-lax and other Neotropical freshwater fishes. Copeia 1999:1027-1034.
ETZEL, V. & BERKENKAMP, H. O. 1992. Die Rivulus-Artenvon Panama 1: Untersuchungen zur identität von Rivuluschucunaque Breder, 1925. Deutsche Killifisch GemeinschaftJournal 24: 29-32.
FELS, J. F. & HUBER, J. H. 1985. Descriptión d’une espècenouvelle de Rivulus du Pérou, R. derhami n. sp. et nouvelledescription de R. beniensis Myers, 1927 (Pisces, Cyprin-odontidae). Revue Française d’Aquariologie 12: 33-38.
FELS, J. F. & DE RHAM, P. 1982. Récentes collectes de Rivu-lus (Cyprinodontidés) au Pérou, avec description de sixespèces nouvelles, deuxième partie. Revue Française d’Aquar-iologie 8: 97-106.
FELSENSTEIN, J. 1985. Confidence limits on phylogenies: anapproach using the bootstrap. Evolution 39: 783-791.
FOWLER, H. W. 1944. Freshwater fishes from North-westernColombia. Proceedings of the Academy of Natural Sciencies ofPhiladelphia 96: 227-248.
GARMAN, S. 1895.The cyprinodonts. Memoirs of the Museumof Comparative Zoology 19: 1-179.
HENN, A. W. 1916. IX – On various South American poe-ciliid fishes. Annals of the Carnegie Museum 10: 93-142 + 4plates.
HOEDEMAN, J. J. 1958. The frontal scalation pattern in somegroups of toothcarps (Pisces, Cyprinodontiformes). Bulletinof Aquatic Biology 1: 23-28.
HOEDEMAN, J. J. 1959. Rivulid fishes of Suriname andother Guyanas, with a preliminary review of the genusRivulus. Studies on the Fauna of Suriname and OtherGuyanas 3: 44-98.
HOEDEMAN, J. J. 1961. Studies on Cyprinodontiform fishes,8: Preliminary key to the species and subspecies of the genusRivulus. Bulletin of Aquatic Biology 2: 65-74.
HRBEK, T., DE DEUS, C. P. & FARIAS, I. P. 2004. Rivulusduckensisi (Teleostei; Cyprinodontiformes): new speciesfrom the Tarumã basin of Manaus, Amazonas, Brazil,and its relationships to other neotropical Rivulidae.Copeia 2004: 569-576.
HRBEK, T. & LARSON, A. 1999. The diapause in the killifishfamily Rivulidae (Atherinomorpha, Cyprinodontiformes): amolecular phylogenetic and biogeographic perspective. Evo-lution 53: 1200-1216.
HUBER, J. H. 1979. A propos de quatre nouvelles collectionsde Rivulus des Guyanes, avec description de Rivulus xiphid-ius n. sp. et Rivulus amphoreus n. sp. Revue Françaised’Aquariologie 6: 65-72.
HUBER, J. H. 1991. Revue des espèces de Rivulus de Guyanefrançaise, avec descriptions de Rivulus cladophorus n. sp. etde R. igneus n. sp., et nouvelles descriptions de R. uroph-thalmus Günther et de R. micropus Steindachner. RevueFrançaise d’Aquariologie 18: 65-74.
HUBER, J. H. 1992. Review of Rivulus: ecobiogeografia - rela-tionships. Cybium, Société Française d’Ichtyologie, Paris,572 + 13 pp.
HUBER, J. H. 1999. Updates to the phylogeny and sys-tematics of the Neotropical cyprinodont genus Rivulusand its allied (Cyprinodontiformes: Rivulinae). Cybium23: 29-52.
KITCHING, I. J., FOREY, P. L., HUMPHRIES, C. J. &WILLIAMS,D. M. 1998. Cladistics, the theory and practice of parsimonyanalysis (2nd edition), Oxford University Press, Oxford-New York-Tokyo, 228 pp.
LASSO, C., TAPHORN, D. C. & THOMERSON, J. E. 1992.Rivulus gransabanae, a new species of killifish fromVenezuela (Cyprinodontiformes: Rivulidae). IchthyologicalExploration of Freshwaters 2: 297-303.
LUNDBERG, J. G., MARSHALL, L. G., GUERRERO, J., HOR-TON, B., MALABARBA, M. C. S. L. & WESSELINGH, F.1998. The stage for Neotropical fish diversification: a his-tory of tropical South American rivers. In: Malabarba,L.R., Reis, R. E., Vari, R. P., Lucena, Z. M. S. & Lucena,
Wilson J. E. M. Costa

aqua vol. 11 no. 4 - 2006 170
C. A. S. (Eds.). Phylogeny and classification of neotropicalfishes, Edipucrs, Porto Alegre, pp. 13-48.
MEEK, S. E. 1904. The freshwater fishes of Mexico North ofthe Isthmus of Tchuantepec. Publications of the FieldColumbian Museum, Zoology 5: 1-252.
MENEZES, N. A. &WEITZMAN, S. H. 1990.Two new speciesof Mimagoniates (Teleostei: Characidae: Glandulocaudi-nae), their phylogeny and biogeography and a key to glan-dulocaudin fishes of Brazil and Paraguay. Proceedings of theBiological Society of Washington 103: 380-426.
MURPHY, W. J. & COLLIER, G. E. 1996. Phylogenetic rela-tionships within the aplocheiloid fish genus Rivulus(Cyprinodontiformes, Rivulidae): Implications forCaribbean and Central American Biogeography. MolecularBiology and Evolution 14: 642-649.
MURPHY, W. J. & COLLIER, G. E. 1997. A molecularphylogeny for aplocheiloid fishes (Atherinomorpha,Cyprinodontiformes): the role of vicariance and the originsof annualism. Molecular Biology and Evolution 14: 790-799.
MURPHY, W. J., THOMERSON, J. E. & COLLIER, G. E. 1999.Phylogeny of the neotropical killifish family Rivulidae(Cyprinodontiformes, Aplocheiloidei) inferred from mito-chondrial DNA sequences. Molecular and Phylogenetic Evo-lution 13: 289-301.
MYERS, G. S. 1927a. An analysis of the genera of neotropicalkillifishes allied to Rivulus. Annals and Magazine of NaturalHistory, ser. 9 19: 115-129.
MYERS, G. S. 1927b. Descriptions of new South Americanfresh-water fishes collected by Dr. Carl Ternetz. Bulletin ofthe Museum of Comparative Zoology, Harvard 68: 107-135.
PARENTI, L. R. 1981. A phylogenetic and biogeographicanalysis of cyprinodontiform fishes (Teleostei, Atheri-nomorpha). Bulletin of the American Museum of NaturalHistory 168: 335-557.
PEARSON, N. E. 1925. The fishes of the Rio Beni basin,Bolivia, collected by the Mullford Expedition. Indiana Uni-versity Studies 64: 1-51.
POEY, F. 1860. Poissons de Cuba, espèces nouvelles. Memoriassobre la Historia Natural de la Isla de Cuba, 2: 115-336.
RAMOS, T. C. 1996. Tree Gardener (computer program), Ver-sion 2.3beta. Univ. São Paulo, São Paulo.
REGAN, C. T. 1907. VI – Diagnoses of new Central-Ameri-can freshwater fishes of the families Cyprinodontidae andMugilidae. Annals and Magazine of Natural History, ser. 719: 64-65.
REGAN, C. T. 1912. A revision of the poeciliid fishes of thegenera Rivulus, Pterolebias, and Cynolebias. Annals and Mag-azine of Natural History, ser. 8 10: 494-508.
REIS, R. E. 1997. Revision of the neotropical catfish genusHoplosternum (Ostariophysi: Siluriformes: Callichthyidae),with the description of two new genera and three newspecies. Ichthyological Exploration of Freshwaters, 7: 299-326.
RINGUELET, R. A., ARAMBURU, R. H. & ARAMBURU, A.1967. Los peces argentinos de agua dulce. Comisión de Inves-
tigaciones Científicas de la Provincia de Buenos Aires, LaPlata, 602 pp.
SEEGERS, L. 1984. Zur Revision der Rivulus-Arten Südost-Brasiliens, mit einer Neubeschreibung von Rivulus luelingin. sp. und Rivulus caudomarginatus n. sp. (Pisces: Cyprin-odontidae: Rivulinae). Zoologische Beiträge 28: 271-320.
SEEGERS, L. 1988. Ein neuer Rivulide aus Bolivien: Rivulusbolivianus n. sp. (Pisces: Cyprinodontiformes: Rivulidae).Bonner Zoologische Beitrage 39: 171-177.
STEINDACHNER, F. 1863. Beiträge zur Kenntniss der Sci-aenoiden Brasiliens und der Cyprinodonten Mejicos.Sitzungsberichte der kaiserlichen Akademie der WissenschaftenClasse K. 48: 162-185, plates 1-4.
SWOFFORD, D. L. 1998. PAUP*. Phylogenetic analysis usingparsimony (*and other methods), Beta version 4.0b2. Sinauer,Sunderland, Massachusetts.
TAYLOR, W. R. & VAN DYKE, G. C. 1985. Revised proce-dures for staining and clearing small fishes and other verte-brates for bone and cartilage study. Cybium 9: 107-109.
THOMERSON, J. E., BERKENKAMP, H. O. &TAPHORN, D. C.1991a. Rivulus lyricauda, a new species from the GuyanaShield in eastern Venezuela (Cyprinodontiformes: Rivuli-dae). Ichthyological Exploration of Freshwaters 1: 289-294.
THOMERSON, J. E., NICO, L. G. &TAPHORN, D. C. 1991b.Rivulus immaculatus, a new killifish from Venezuela(Cyprinodontiformes, Rivulidae). Copeia 1991: 323-328.
THOMERSON, J. E., NICO, L. G. & TAPHORN, D. C. 1992.Rivulus tecminae, a new killifish from Amazonas Territory,Venezuela (Cyprinodontiformes: Rivulidae). IchthyologicalExploration of Freshwaters 2: 289-296.
THOMERSON, J. E. & TAPHORN, D. C. 1992. Two newannual killifishes from Amazonas Territory, Venezuela(Cyprinodontiformes: Rivulidae). Ichthyological Explorationof Freshwaters 3: 377-384.
WALLACE, A. R. 2002. Peixes do Rio Negro/Fishes of the RioNegro. EDUSP, São Paulo, 517 pp.
WEITZMAN, S. H. & WOURMS, J. P. 1967. South Americancyprinodont fishes allied to Cynolebias with the descriptionof a new species of Austrofundulus from Venezuela. Copeia1967: 89-100.
WIENS, J. J. (ed.). 2000. Phylogenetic analysis of morphologicaldata, Smithsonian Institution Press, Washington and Lon-don, 220 pp.
VERMEULEN, F. B. M. &HRBEK,T. 2005 Kryptolebias sepia n.sp. (Actinopterygii: Cyprinodontiformes: Rivulidae), a newkillifish from the Tapanahony River drainage in southeastSurinam. Zootaxa 928: 1-20.
VERMEULEN, F. B. M. & ISBRÜCKER, I. J. H. 2000. Rivulustorrenticola n. sp. (Actinopterygii: Cyprinodontiformes:Rivulidae), a new killifish from highlands in the GuyanaShield. Beaufortia 50: 185-190.
VON IHERING, R. 1931. Cyprinodontes brasileiros (peixes“guarús”): sistemática e informações biologicas. Archivos doInstituto Biologico 4: 242-280.
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin

aqua vol. 11 no. 4 - 2006171
Appendix 1The list below includes material examined, except belonging to
species of the Brazilian Amazonas basin, which are listed in the“Taxonomic accounts” above. Data on material is organized inthe following sequence: catalogue number, number of speci-mens, locality. Abbreviations are: c&s, specimens cleared andstained for bone and cartilage, H, holotype, and P, paratype(s).Institutional acronyms are listed in material and methods.
Aplocheilidae: Aplocheilus panchax: UFRJ 3140, 2; UFRJ3141, 2 (c&s); Indonesia: Sulawesi: Desa Radda. Notho-branchiidae: Scriptaphyosemion guignardi: UFRJ 3883, 8; UFRJ4110, 4 (c&s); Guinea: Dalaba. Rivulidae: Aphyolebias boticarioi:UFRJ 5986, H; UFRJ 5987, 11 P; UFRJ 5988, 5 P; Brazil:Estado do Acre: Porto Acre. Gnatholebias hoignei: MCNG 1116,18; Venezuela: Portuguesa: La Trinidad. UFRJ 6116, 6; UFRJ6117, 5 (c&s); Venezuela: Portuguesa: Papelón. Kryptolebiasbrasiliensis: UFRJ 3458, 32; UFRJ 3682, 1 (c&s); UFRJ 4603,2 ex. (c&s); UFRJ 5332, 6 ex. (c&s); Brazil: Rio de Janeiro:Magé. Micromoema xiphophora: MCNG 26440, 20; Venezuela:Amazonas: Isla Ratón. UFRJ 3165, 1 (c&s); aquarium material.Moema apurinan: UFRJ 5980, H; UFRJ 5981, 9 P; UFRJ 5982,7 P (c&s); Brazil: Estado do Acre: Porto Acre. Neofundulusparaguayensis: UFRJ 3647, 10; UFRJ 3648, 4 (c&s); Brazil:Mato Grosso do Sul, about 70 km NW from de Aquidauana.Pituna compacta: UFRJ 3563, 33; UFRJ 3564, 4 (c&s); Brazil:Tocantins: Barreira do Piqui. Prorivulus auriferus: UFRJ 5932,H; UFRJ 5933, 4P; UFRJ 5934, 3P (c&s); Brazil: Bahia:Valença. Rachovia maculipinnis: MCNG 35565, 2; Venezuela:Portuguesa: Sabaneta. UFRJ 6118, 7; UFRJ 6119, 4 (c&s);Venezuela: Portuguesa: Papelón. R. stellifer: MCNG 25828, 9;Venezuela: Portuguesa: La Capilla. UFRJ 245, 5 (c&s);Venezuela: Cojedes: 1 km N of Caño Benito. Renova oscari:MCNG 35926, 2; Venezuela: Amazonas: Isla Ratón. UFRJ4606, 3 (c&s): aquarium material. Rivulus amphoreus: UFRJ3889, 2, UFRJ 4606, 3 (c&s); Surinam: Tafelberg. R. apiamici:MZUSP 39976, H; MZUSP 39977, 3P (c&s); UFRJ 5971, 4;UFRJ 5972, 3 (c&s); Brazil: Mato Grosso do Sul; Bataguaçu. R.bahianus: UFRJ 3167, 44; UFRJ 4602, 2 (c&); UFRJ 277, 1(c&s); Brazil: Bahia: Busca-Vida. R. brunneus: MZUSP uncata-logued, 10 (2 c&s); Panama: Isla de Barro Colorado. R. chucu-naque: USNM 293487, 68 (1c&s); Panama. R. cladophorus:UFRJ 643, 4P (2c&s); Guyana: Fourgassier. R. cryptocallus:UFRJ 359, 3; UFRJ 2126, 1 (c&s); Martinique: Ravine Vilaine.R. cyanopterus: UFRJ 5911, H; UFRJ 5913, 36P; UFRJ 5914,5P (c&s); Brazil: Mato Grosso: Jaciara. R. cylindraceus: MHNCuncat., 6; Cuba: Zapata. USNM uncat., 1 (c&s); Cuba: LaHabana. R. dapazi: UFRJ 5915, H; UFRJ 5916, 3P; UFRJ5917, 2P (c&s); Brazil: Mato Grosso do Sul: Sonora. R. decora-tus: MZUSP 39982, H; MZUSP 39983, 4P; UFRJ 2135, 3(c&s); Brazil: Bahia: Ibiraba. R. derhami: UFRJ 392, 2 (c&s);Peru:TingoMaria. R. depressus: UFPB 2213, H; UFPB 1749, 17(2 c&s); UFRJ 2118, 1 (c&s); Brazil: Bahia: Porto Seguro. R.elongatus: MUSM 336, 6; MZUSP 26211, 3; Peru: Ucayali:Pucallpa. R. egens: UFRJ 5973, H; UFRJ 5974, 17P; UFRJ5975, 3P (c&s); Brazil: Mato Grosso do Sul: Camapuã. R. erberi:UFRJ 358, 3; Ecuador: Napo: Coca. R. fuscolineatus: USNM219778, 10P; Costa Rica: Guanacaste: Tilaran. R. geayi:MZUSP uncatalogued Brazil: Amapa: Serra do Navio. R. har-
aldsiolii: UFRJ 125, 6; UFRJ 6295, 2 (c&s); Brazil: Santa Cata-rina: Joinville. R. hartii: MCZ 26092, 3; Trinidad: Port of Spain.MZUSP 37204, 3; MZUSP 38472, 2 (c&s); Venezuela: NuevaEsparta: Isla de Margarita. R. hildebrandi: USNM 92958, 1P;Panama: Chiriqui: Boquete. R. holmiae: USNM 66302, 1P;Guiana: Holmia. R. igneus: UFRJ 3888, 6; UFRJ 4595, 2 (c&s);Suriname: Creek Colibri. R. immaculatus: USNM 308411, 2P;Venezuela: Bolivar: Elena de Uairen. R. janeiroensis: UFRJ 5333,8; UFRJ 130, 2 P (c&s); UFRJ 5416, 7 (c&s); Brazil: Rio deJaneiro: Magé. R. luelingi: UFRJ 161, 8, UFRJ 127, 5 (c&s);Brazil: Santa Catarina: Araquari. R. lungi: ZSM 27825, 15;Brazil: Amapá: rio Flechal. R. nicoi: MCNG 23891, H; MCNG23892, 1P; MCNG 23893, 1P; Venezuela: Amazonas: río Ven-tuari floodplains. R. nudiventris: MZUSP 40283, H; MZUSP40284, 3P; MNRJ 11740, 2P (c&s); Brazil: Espírito Santo:Itapemirim. R. paracatuensis: MCP 29639, H; MCP 29640, 1P;UFRJ 2290, 3P (c&s); Brazil: Minas Gerais: Brasilândia deMinas. R. parnaibensis: MCP 29639, H; UFRJ 4962, 25P;UFRJ 5449, 4P (c&s); Brazil: Piauí: São Dimas. R. pictus:MNRJ 11550, H; MNRJ 11551, 2P; UFRJ 5959, 2 ex. (c&s);Brazil: Distrito Federal: Planaltina. R. pinima: MZUSP 39978,H; MZUSP 39984, 2P; UFRJ 5960, 20; UFRJ 5961, 4 (c&s);Brazil: Goiás: Rio Verde. R. punctatus: UFRJ 975, 9; UFRJ2110, 4 (c&s); Brazil: Mato Grosso do Sul: Aquidauana. R.rossoi: UFRJ 5976, H; UFRJ 5977, 7P; UFRJ, 5978, 4P (c&s);Brazil: Mato Grosso do Sul: Campo Grande. R. rubrolineatus:MZUSP 26371, 3; MUSM 1465, 6; Peru: Loreto: Jenaro Her-rera. R. rutilicaudus: UFRJ 5965, H; UFRJ 5966, 12P; UFRJ5967, 3P (c&s); Brazil: Goiás: Serranópolis. R. santensis: UFRJ5441, 11; UFRJ 6294, 5 (c&s); Brazil: São Paulo: Boracéia. R.simplicis: UFRJ 5940, H; UFRJ 5976, 3P; UFRJ 5942, 5P(c&s); Brazil: Rio de Janeiro: Parati. R. scalaris: UFRJ 5968, H;UFRJ 5969, 19P; UFRJ 5970, 4P (c&s); Brazil: Mato Grosso doSul: Costa Rica. R. stagnatus: UFRJ 3890, 3; UFRJ 4605, 4(c&s); Suriname: Wageningen. R. tenuis: UFRJ 4600, 4; UFRJ4601, 2 (c&s); Guatemala: Alta Verapaz: Sebol. R. vittatus:MZUSP 39981, H;MZUSP 39981, 6P; UFRJ 2206, 15; UFRJ5962, 4 (c&s); Brazil: Goiás: Cachoeira Alta. R. xiphidius: UFRJ4608, 3 (c&s); Guyana (specimens born in aquarium).Trigonectes rubromarginatus: UFRJ 3553, 13; UFRJ 3554, 3(c&s); Brazil: Tocantins: Barreira do Piqui.
Wilson J. E. M. Costa

aqua vol. 11 no. 4 - 2006 172
Appendix 2Characters (in brackets) and character states (in parentheses)
used to erect the phylogenetic hypothesis among species of Rivu-lus and other rivulids are listed below, with the respective refer-ence to papers where the character is first described or discussed.Distribution of character states among terminal taxa is presentedin the data matrix in Appendix 3.
Superficial dermal bones and neurocranium[1] Lachrymal (Parenti 1981; Costa 1998a, 1998b) (CI: 1.00;
RI: 1.00): (0) flat, posterior rim wide; (1) slightly twisted, poste-rior rim reduced, bone formed mainly by canal; (2) very twistedand narrow, slender, canal vestigial.[2] Ventral portion of lachrymal (Costa 1998a) (CI: 0.50; RI:
0.66): (0) short; (1) expanded.[3] Dermosphenotic (Costa 2004d): (CI: 1.00; RI: 1.00): (0)
present; (1) absent.[4] Vomerine teeth (Costa 1998a) (CI: 0.40; RI: 0.80): (0) usu-
ally 1-4, sometimes 5-6; (1) 6-12; (2) teeth absent [not ordered].[5] Anterior retrorse process of lateral ethmoid (Costa 1990)
(CI: 1.00; RI: 1.00): (0) short; (1) moderate to elongate.[6] Lateral process of sphenotic (Costa 2005b) (CI: 0.50; RI:
0.50): (0) narrow; (1) wide.[7] Anterolateral process of parasphenoid (Costa 1998a) (CI:
0.16; RI: 0.78): (0) short, free; (1) long, attached to pterosphe-noid.[8] Posterior portion of parasphenoid (Costa 2005b) (CI: 0.50;
RI: 0): (0) wide; (1) narrow.[9] Lateral border of frontal (Costa 1998a) (CI: 1.00; RI:
1.00): (0) well-ossified, approximately straight; (1) poorly ossi-fied, concave.Jaws, jaw suspensorium and opercular apparatus[10] General shape of the premaxilla and dentary (Costa
1998a) (CI: 0.25; RI: 0): (0) elongate, snout profile sharplypointed; (1) short, snout profile blunt.[11] General shape of ascending process of premaxilla (modi-
fied from Parenti, 1981, Costa 1998b) (CI: 1.00; RI: 1.00): (0)curved, posterior portion medially directed; (1) approximatelystraight.[12] Ascending process of premaxilla (Costa 2005b) (CI: 0.50;
RI: 0): (0) wide; (1) narrow.[13] Ventral process of maxilla (Parenti 1981, Costa 1998a)
(CI: 1.00; RI: 1.00): (0) slightly curved, anterior marginrounded; (1) bent, anterior margin triangular.[14] Maxilla (modified from Parenti 1981) (CI: 0.40; RI:
0.72): (0) approximately straight; (1) slightly twisted; (2) greatlytwisted.[15] Rostral cartilage (Costa 1998a) (CI: 0.25; RI: 0.40): (0)
approximately rounded; state 1: longitudinal length longer thantransversal length.[16] Posterior portion of rostral cartilage (Costa 2004a) (CI:
0.20; RI: 0.69): (0) not distinctively narrowed; (1) distinctivelynarrowed.[17] Coronoid process of dentary (Costa 1998a) (CI: 1.00; RI:
1.00): (0) broad; (1) narrow.[18] Ventral process of angulo-articular (modified from Costa
1990, 1998a) (CI: 0.66, RI: 0.81): (0) large and broad; (1) large,somewhat narrowed; (2) moderate, narrow and pointed; (3) ves-tigial; (4) long and somewhat narrowed [not ordered].
[19] Curvature of ventral process of angulo-articular (Costa1998a) (CI: 1.00, RI: 1.00): (0) straight; (1) curved.[20] External medial teeth of premaxilla and dentary (Costa
1998a) (CI: 0.33; RI: 0.50): (0) approximately directed as otherteeth; (1) laterally displaced, strongly contrasting to other teeth.[21] Ventral portion of palatine (Costa 1998a) (CI: 1.00; RI:
1.00): (0) long, overlapping dorsal portion of quadrate; state 1:short, not or slightly contacting quadrate.[22] Metapterygoid (modified from Costa 1998a) (CI: 0.66;
RI: 0.90): (0) about rectangular, dorsal and ventral portions wideand approximately equal in width; (1) dorsal portion slightlyconstricted; (2) about triangular, dorsal portion strongly con-stricted.[23] Posterior process of quadrate (Costa 1998a) (CI: 0.50; RI:
0.50): (0) short, about 50% of quadrate length; (1) long, about70% of quadrate length.[24] Preopercle (Costa 1990) (CI: 1.00; RI: 1.00): (0) robust,
L-shaped, with a well-developed anteromedian rim; (1) thin, C-shaped, with a reduced anteromedian rim.[25] Dorsal arm of preopercle (Costa 1990) (CI: 0.50; RI:
0.85): (0) broad; (1) narrow and pointed.Hyoid and branchial arches[26] Anterior process of urohyal (Costa 1998a) (CI: 1.00; RI:
1.00): (0) short; (1) elongate.[27] Dorsal process of urohyal length (CI: 1.00; RI: 1.00): (0)
present; (1) absent; (?) very short to absent.[28] Dorsal process of urohyal length (Costa 1998a) (CI: 0.25;
RI: 0.85): (0) short; (1) elongate; (?) process absent.[29] Basihyal (Costa 1998a) (CI: 0.50; RI: 0): (0) shorter than
space occupied by basibranchials; (1) longer than space occupiedby basibranchials.[30] Basihyal cartilage extent, as percentage of basihyal length
(modified from Costa 1998a) (CI: 0.40; RI: 0.62): (0) 50-70 %;(1) 20-40 %; (2) 10-15 %.[31] Interhyal (Parenti 1981) (CI: 1.00; RI: 1.00): (0) ossified;
(1) cartilaginous.[32] Interhyal (Costa 2005b) (CI: 1.00; RI: 1.00): (0) large; (1)
minute.[33] Number of branchiostegal rays (Costa 2004e) (CI: 0.33;
RI: 0): (0) 6; (1) 5.[34] Main axis of first epibranchial (Costa 1998a) (CI: 1.00;
RI: 1.00): (0) approximately straight; (1) curved.[35] Subdistal process of second epibranchial (Costa 2004a)
(CI: 1.00; RI: 1.00): (0) present; (1) absent.[36] Uncinate process of third epibranchial (modified from
Costa 1998a, b) (CI: 1.00; RI: 1.00): (0) long; (1) moderate; (2)short.[37] Interarcual cartilage (modified from Parenti 1981) (CI:
1.00; RI: 1.00): (0) not reduced; (1) reduced (CI: 1.00; RI:1.00).[38] Number and arrangement of second pharyngobranchial
teeth (Costa 2004a) (CI: 0.50; RI: 0.88): (0) numerous teetharranged in two rows; (1) few teeth arranged in single row; (2)teeth absent [not ordered].[39] Proximal edge of first hypobranchial (Costa 1998a) (CI:
0.33; RI: 0.33): (0) plain, terminating in single cartilage unitedto second basibranchial; (1) bifid, terminating in cartilage unitedto second basibranchial and another smaller cartilage united tofirst basibranchial.
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin

aqua vol. 11 no. 4 - 2006173
[40] Distal edge of first hypobranchial (Costa 2004a) (CI:1.00; RI: 1.00): (0) articular face restricted to cartilaginous headof first ceratobranchial; (1) articular face anteriorly expanded.[41] Orientation of anterior tip of fifth ceratobranchial (Costa
2005b) (CI: 0.50; RI: 0.50): (0) anterior; (1) anterolateral.Vertebrae and caudal skeleton[42] Pointed, anteriorly directed process on first vertebra
(modified from Costa 1990) (CI: 1.00; RI: 1.00): (0) absent;(1) present.[43] Epipleural ribs (Parenti 1981, Costa 1998a) (CI: 0.50;
RI: 0.66): (0) rod-like; (1) bifid.[44] Neural prezygapophyses (Costa 1990) (CI: 0.33; RI:
0.84): (0) short; (1) long.[45] Hypurals (modified from Costa 1998a) (CI: 0.42; RI:
0.33): (0) two dorsal plates and one ventral plate separated bygap; (1) two plates separated by wide gap; (2) two plates in closeproximity, sometimes ankylosed; (3) single plate.[46] Proximal end of parahypural (Costa 1998b) (CI: 1.00;
RI: 1.00): (0) robust with paired dorsal processes overlappingpreural centrum; (1) shortened and laminar, without dorsalpaired process, not contacting preural centrum.[47] Hemal spine of preural centrum two (Costa 1998b) (CI:
0.50; RI: 0): (0) distinctively wider than hemal spines anteriorto it; (1) slightly wider or equal in width to hemal spines ante-rior to it.[48] Number of vertebrae (modified from Costa 1990) (CI:
0.18; RI: 0.52): (0) 29-32; (1): 33-35; (2) 36-38.[49] Number of caudal-fin rays (modified from Costa 1990)
(CI: 0.18; RI: 0.59): (0) 26-31; (1) 32-36; (2) 24-25 [notordered].Dorsal- and anal-fin skeleton[50] First dorsal-fin ray (modified from Parenti, 1981) (CI:
1.00; RI: 1.00): (0) single long first ray connected to two prox-imal radials; (1) long fin ray connected to two proximal radials,preceded by one or two short fin rays.[51] Anterior proximal radials of dorsal and anal fins (Costa
2005b) (CI: 1.00; RI: 1.00): (0) slender; (1): wide.[52] Number of anal-fin rays (modified from Costa 1998a)
(CI: 0.28; RI: 0.44): (0) 9-15; (1) 16-20; (2) 25-26.[53] Orientation of anterior proximal radials of anal fin
(modified from Costa 1998a) (CI: 1.00; RI: 1.00): (0) anteri-orly or dorsally directed; (1) posteriorly directed.Shoulder and pelvic girdle[54] Pectoral-fin insertion (Costa 1998b) (CI: 1.00; RI:
1.00): (0) lateral; (1) ventrolateral.[55] Supracleithrum and posttemporal (modified from Par-
enti 1981, Costa 1998b) (CI: 0.66; RI: 0.50): (0): separated;(1) co-ossified, limits almost inconspicuous; (2) fused to forma single structure.[56] Keel along supracleithrum-posttemporal (Costa 2005b)
(CI: 1.00; RI: 1.00): (0) absent; (1) present.[57] Ventral process of posttemporal (Parenti 1981, Costa
1998a, b) (CI: 0.20; RI: 0.81): (0) present; (1) absent.[58] Posterior flange of cleithrum (Costa 1998a) (CI: 1.00;
RI: 1.00): (0) present; (1) absent.[59] First postcleithrum (Parenti 1981) (CI: 1.00; RI: 1.00):
(0) present; (1) absent.[60] Fourth pectoral radial (Costa 1998a) (CI: 0.33; RI:
0.75): (0) not expanded; (1) ventrally expanded.
[61] Ischial process of pelvic girdle (Costa 2005b) (CI: 1.00;RI: 1.00): (0) prominent; (1) vestigial.[62] Number of pectoral-fin rays (modified from Costa
1998a): (0) 13-15; (1) 16-17.[63] Number of pelvic-fin rays (Costa 1990) (CI: 1.00; RI:
1.00): (0) six; (1) seven; (2) eight.External morphology of body and head[64] Mouth (Costa 1998b) (CI: 1.00; RI: 1.00): (0) terminal;
(1) superior.[65] Orbital rim (modified from Parenti 1981) (CI: 1.00; RI:
1.00): (0) free dorsally, attached ventrally; (1) completelyattached.[66] Branchiostegal and opercular membranes (Parenti
1981) (CI: 1.00; RI: 1.00): (0) separated by long fold; (1)continuous.[67] Skin fold on corner of preopercular region (Costa
2005b) (CI: 1.00; RI: 1.00): (0) present; (1) absent.External morphology of fins[68] Pectoral fin (Costa 1990) (CI: 0.50; RI: 0.75): (0)
rounded; (1) pointed.[69] Pectoral-fin length (modified from Parenti 1981) (CI:
0.33; RI: 0.75): (0) 17.0–23.8 % SL; (1) 24.1–31.1 % SL.[70] Extent of pelvic fin in males (Costa 1998a) (CI: 0.25; RI:
0.25): (0) short, tip not surpassing anterior portion of anal fin;(1) long, its tip reaching the central or the posterior portion ofthe anal fin.[71] Pelvic-fin (Costa 2005b) (CI: 1.00; RI: 1.00): (0) bases
separated or in contact; (1) bases united; (2) pelvic fins unitedalong proximal portion of medial margin.[72] Dorsal fin (modified from Costa 1998a) (CI: 0.33; RI:
0.84): (0) short, tip rounded; (1) somewhat elongated, tippointed; (2) long, tip sharply pointed.[73] Anal fin (modified from Costa 1998a) (CI: 0.22; RI:
0.75): (0) short, tip rounded; (1) somewhat elongated, tippointed; (2) long, tip sharply pointed.[74] Filaments on tip of dorsal and anal fins in males (Costa
1998a) (CI: 0.50; RI: 0.66): (0) absent; (1) present.[75] Filaments on posterior border of caudal fin in males
(Costa 1998a) (CI: 0.33; RI: 0.60): (0) absent; (1) present.[76] Caudal-fin length in males (Costa 1990) (CI: 0.40; RI:
0.70): (0) 29.0-41.5; (1) 42.0-49.0; (2) 52.5-81.0.[77] Caudal-fin shape in males (CI: 0.33; RI: 0.40): (0)
rounded; (1) subtruncate; (2) truncate; (3) acuminate [notordered].[78] Dorsal and ventral extensions on caudal fin in males
(Costa 1998a) (CI: 0.50; RI: 0): (0) absent; (1) present.Squamation[79] General arrangement of frontal scales (Hoedeman 1958)
(CI: 0.50; RI: 0.85): (0) transverse; (1) circular; (2) irregular[not ordered].[80] Predominant frontal squamation-pattern (Hoedeman
1958) (CI: 0.28; RI: 0.52): (0) G; (1) E; (2) D; (3) F; (4) S [notordered].[81] Arrangement of E-scales (Costa 1990) (CI: 1.00; RI:
1.00): (0) overlapped; (1) not overlapped.[82] Caudal-fin squamation in older males (Costa 1990,
1998a) (CI: 0.25; RI: 0.57): (0) approximately on anterior10-30 % of fin; (1) approximately on 40 % of fin; (2) approx-imately on 50-80 % of fin.
Wilson J. E. M. Costa

aqua vol. 11 no. 4 - 2006 174
[83] Anal-fin base squamation in males (Costa 2005b) (CI:1.00; RI: 1.00): (0) no scales on anal-fin base; (1) 1-7 rows ofscales on anal-fin base.Laterosensory system[84] Cephalic canals (modified from Parenti 1981) (CI:
0.75; RI: 0.92): (0) anterior and posterior infraorbital, preop-ercular and posterior mandibular closed, supraorbital, rostral,anterior mandibular open with skin trenches around neuro-masts; (1) anterior and posterior infraorbital, and preopercu-lar closed, supraorbital, rostral, anterior and posteriormandibular open with skin trenches around neuromasts; (2)anterior and posterior infraorbital, dorsal preopercular, supra-orbital, rostral, anterior and posterior mandibular open withskin trenches around neuromasts; (3) no vestige of canals, allneuromasts completely exposed.[85] Number of anterior supraorbital neuromasts (Costa
1990) (CI: 0.50; RI: 0.33): (0) 3; (1) 4; (2) 6–7.[86] Arrangement of anterior supraorbital neuromasts (Costa
2004d) (CI: 1.00; RI: 1.00): (0) continuous row; (1) posterior-most neuromast separated by space covered by scale.[87] Arrangement of anterior and posterior rostral neuromast
(CI: 1.00; RI: 1.00): (0) longitudinal; (1) transverse.Contact organs[88] Contact on flank scales in males (Costa 2005b) (CI:
0.14; RI: 0.66): (0) absent; (1) present.Male colour patterns[89] Red marks on male flank (CI: 0.16; RI: 0.44): (0) absent;
(1) present.[90] Kind of red marks on male flank (CI: 0.44; RI: 0.69): (0)
vertical rows of dots, sometimes forming stripes; (1) verticalrows of dots on anterior portion, chevron-like bars on poste-rior; (2) dots on chevron-like series on entire flank, sometimesforming oblique bars; (3) overlapping longitudinal and obliquerows; (4) red marks irregularly arranged, not forming any dis-tinguishable pattern; (?) red marks absent [not ordered].[91] Dark pigmentation pattern on humeral region (modified
from Costa 1998a) (CI: 0.33; RI: 0.50): (0) not distinctivelycoloured; (1) forming distinct blotch.[92] Dark pigmentation pattern on upper portion of caudal-
fin base (Costa 2005b) (CI: 0.50; RI: 0.40): (0) not distinc-tively coloured; (1) forming distinct blotch.[93] Dark pigmentation on anterior half of flank when fish is
exposed to sunlight (modified from Costa 1998a) (CI: 0.20;RI: 0.80): (0) not distinctively marked; (1) forming grey verti-cal bars; (2) forming dark grey broad stripe; (3) forming darkgrey oblique bars [not ordered].[94] Dark pigmentation between dorsum and flank (modified
from Costa, 1998a) (CI: 1.00; RI: 1.00): (0) not distinctivelymarked; (1) forming longitudinal dark zone.[95] Dark pigmentation pattern on iris (Parenti 1981) (CI:
1.00; RI: 1.00): (0) no distinctive mark; (1) bar crossing eye.[96] Pigmentation pattern on suborbital region (Costa 1998a)
(CI: 0.66; RI: 0.75): (0) no distinctive mark; (1)grey to black suborbital bar; (2) red suborbital spot [notordered].[97] Pigmentation pattern on post-orbital and preopercular
region (Costa 1998a) (CI: 0.40; RI: 0.66): (0) no distinctivepattern; (1) two oblique bars; (2) post-orbital vertical bar; (3)two oblique stripes [not ordered].
[98] Dark pigmentation pattern on lower jaw (Costa 2004d)(CI: 0.33; RI: 0.84): (0) not distinctively concentrated; (1) jawdark grey to black; (2) transverse stripe through chin [notordered].[99] Dark blue to dark purplish blue iridescence on opercular
and infraorbital region (Costa 1998a) (CI: 0.25; RI: 0.57): (0)absent; (1) present.[100] Dark pigmentation on distal portion of male anal fin
(Costa 1998a) (CI: 0.20; RI: 0.50): (0) not distinctive concen-trated; (1) concentrated to form stripe.[101] Yellow to orange pigmentation pattern on ventral por-
tion of caudal fin (modified from Costa 1998a) (CI: 0.10; RI:0.55). (0) not distinctive concentrated; (1) concentrated toform stripe; (?) variable.[102] Yellow to orange pigmentation pattern on dorsal por-
tion of caudal fin (modified from Costa 1998a) (CI: 0.11; RI:0.50). (0) not distinctive concentrated; (1) concentrated toform stripe; (?) variable.[103] Dark pigmentation of ventral margin of male caudal fin
(modified from Costa 1998a) (CI: 0.33; RI: 0.75): (0) not dis-tinctive concentrated; (1) concentrated to form stripe.[104] Red pigmentation pattern on subventral portion of cau-
dal fin (CI: 0.33; RI: 0.66): (0) not distinctive concentrated; (1)concentrated to form stripe.[105] Red pigmentation on caudal fin (modified from Costa
1998a) (CI: 0.33; RI: 0.66): (0) not distinctive arranged; (1)forming bars.[106] Dark pigmentation on dorsal portion of caudal fin
(Costa 1998a) (CI: 0.50; RI: 0.75): (0) not distinctive concen-trated; (1) concentrated to form dark brown stripe; (?) variable.[107] Melanophore pattern on pectoral fin (Costa 1998a)
(CI: 0.33; RI: 0): (0) no distinctive marks; (1) spots; (2) bars[unordered].[108] Colour of pectoral fin (CI: 0.33; RI: 0.50): (0) hyaline;
(1) yellow.Female colour patterns[109] Dark pigmentation pattern on flank and fins (Costa
1998a) (CI: 0.50; RI: 0): (0) forming bars, stripes or spots,according to the pattern occurring in males; (1) flank almostplain, pigmentation reduced to minute dots, not presenting thegeneral pattern as in males.[110] Red marks on flank (CI: 0.25; RI: 0.84): (0) absent; (1)
present.[111] Melanophore pattern on unpaired fin margins (Costa
1995a) (CI: 1.00; RI: 1.00): (0) not distinctively concentrated;(1) forming black zones of marginal regions.[112] Dark pigmentation pattern on upper portion of caudal-
fin base (Costa 1990) (CI: 0.14; RI: 0.71): (0) no distinctivemark; (1) one black spot.[113] Kind of caudal blotch (CI: 0.66; RI: 0.85): (0) black
blotch close to fin margin, surrounded by light area to form anocellate spot; (1) black blotch close to or contacting fin margin,anteriorly bordered by triangular light spot; (2) black blotchapart from fin margin, with light area surrounding anterior,dorsal and posterior margin; (?) caudal blotch absent.
Killifish genus Rivulus (Rivulidae) from the Brazilian Amazonas river basin

aqua vol. 11 no. 4 - 2006175
Wilson J. E. M. Costa1–
1011
–20
21–3
031
–40
41–5
051
–60
61–7
071
-80
81-90
91-100
101-11
011
1-11
3
A.pa
ncha
x00
0000
0000
0000
0000
0000
0000
0000
0000
0000
0000
0000
0000
0100
2010
0000
0000
0000
0000
0000
0000
0000
000?
0000
0001
0000
0000
0000
00?
S.gu
ignar
di10
0000
0001
0100
0000
0000
0000
0000
0000
0002
0000
1011
1011
0101
2000
0001
0110
1000
0000
0010
0000
0100
0011
0000
0010
0011
0100
0000
00?
K.br
asili
ensis
2000
1000
1101
1110
1000
1001
00?002
0000
0101
1000
0111
0001
0001
0000
1000
0111
1000
0000
0000
1100
0200
000?
1000
0101
0100
1000
0000
00?
P.au
rifer
us20
0010
1011
1112
1010
0010
0100
0001
1110
1101
1001
0011
1011
0001
0010
1000
0111
10?0
0000
0000
1100
0200
000?
0000
0001
0000
0000
0100
00?
A.bo
ticar
ioi
2100
1010
1111
1110
1100
1201
0001
0111
0012
0111
1100
2111
0100
0111
0111
0001
1111
1002
2111
1111
1002
0001
1300
0010
0000
1001
0010
1000
?G
.hoi
gnei
2000
1001
1010
1100
1101
1101
0001
0111
0012
12?1
0100
3112
0112
1111
0111
1011
1111
1122
2112
1013
1012
0000
0?00
0010
0000
0000
0100
0000
?M
.xip
hoph
ora
2000
1000
1111
1210
1101
1111
0001
1111
0012
0111
0100
1110
0100
0111
1111
0001
1111
1002
2001
3111
1002
0000
1000
0012
1000
1101
0000
0100
?M
.apu
rinan
200?10
1110
1111
1011
0012
0100
0101
1100
1201
1111
0011
1101
0101
1101
1100
0111
1110
0221
1200
1110
0200
0110
0000
1010
0010
0100
0000
00?
N.p
arag
uaye
nsis
2000
1110
1111
1110
1100
1001
0001
0111
0012
0111
0100
1111
0100
0111
0110
0001
1110
1001
1000
0011
1102
1000
1010
0010
0000
1000
0010
0000
?P.
com
pacta
2100
1100
1111
1110
1300
1101
1001
0111
0012
0111
0100
2110
0100
0111
0111
0001
1110
1001
1000
0013
1003
2001
0?00
0011
0000
0000
0010
0000
?P.
longip
inni
s20
0010
1011
1112
1012
0112
1100
0111
1100
1211
1101
0011
1001
1111
1101
1110
1111
1011
1221
1210
1110
1200
000?
?000
1000
0100
0000
2000
00?
R.m
acul
ipin
nis
2100
1010
1111
1110
1100
1101
0101
0111
0012
0111
0100
2110
0100
0111
0111
0001
1110
1001
1011
1013
1202
0000
0?00
0011
2000
?100
0000
1000
?R.
stelli
fer21
0010
1011
1111
1011
0011
0101
0101
1100
1201
1101
0121
1101
0101
1101
1000
0111
1000
0110
0010
1112
0200
000?
0000
1100
0100
0000
0000
011
R.osc
ari
2001
1010
1111
1210
1100
1111
0001
0111
0012
0111
1100
1110
1100
0111
0111
0001
1111
1002
2011
0013
1002
0000
1000
0012
0000
1001
0000
0101
1R.
aman
apira
2010
1000
1111
1210
1100
1001
0000
0111
0012
0201
0101
1112
0100
0111
1110
0001
1110
0001
1000
2004
1003
0011
1000
1100
0200
1100
0000
0001
1R.
amph
oreu
s20
0010
1011
1112
1114
0010
0100
0102
1100
1201
1101
0111
1111
0101
1101
1001
0111
1000
0000
0000
1111
0200
1110
0000
0000
0000
0000
0001
011
R.ap
iam
ici20
0210
0011
1112
1013
0010
0110
0001
1101
1202
1101
0111
1011
0001
1111
1000
0111
1000
0110
0000
1110
0300
1012
0000
0011
0000
0010
0001
112
R.at
ratu
s20
0010
1011
1112
0111
0110
0100
0001
1100
1201
1101
0111
1021
0001
1111
1000
0111
1000
0000
0130
1410
0200
100?
0310
0002
0000
0000
1000
00?
R.ba
hian
us20
0010
1011
1112
1111
0010
0100
0002
1100
1201
1101
0111
1001
0001
1101
1000
0111
1000
0000
0000
1111
0200
1010
0010
0000
0111
1000
0000
011
R.be
nien
sis20
0010
0011
1112
1111
0010
0100
0101
1100
1201
1101
0111
1021
0001
1111
1000
0111
1000
0000
0000
1110
0200
1110
0010
0001
0100
0000
0000
011
R.br
unne
us20
0010
1011
1112
1111
0010
0100
0101
1100
1201
1101
0111
1001
0001
1101
1000
0111
1000
0000
0000
1211
0200
1010
0000
0001
1011
0100
0101
011
R.cla
doph
orus
2002
1000
1111
1210
1100
1001
0000
0111
0012
0211
0111
1110
0100
0111
1110
0001
1110
0000
0000
1013
1002
0011
1110
1000
0100
0010
0000
0100
?R.
cryp
toca
llus
2000
1010
1111
1211
1400
1001
0001
0211
0012
0111
0101
1111
1100
0111
0110
0001
1110
0000
0000
0011
1102
0011
1001
1000
0000
1100
0000
0101
1R.
cylin
drac
eus
2000
1000
1111
1200
1100
1001
0000
0111
0012
0111
0100
3110
0100
0111
0110
0001
1110
0000
0000
0011
1202
1000
1400
0000
0100
0000
0000
0001
0R.
depr
essus
2000
1010
1111
1211
1110
1001
0000
0111
0012
0111
0101
1111
0100
0111
1110
0001
1110
0000
0000
0011
1002
0010
1400
1000
0101
1110
0100
0001
1R.
diba
phus
2002
1000
1111
1210
1100
1001
001?01
1100
1202
1101
1111
1001
0001
1111
1000
0111
1000
0110
0010
1110
0200
1111
1010
0001
0000
1010
0001
00?
R.ha
rtii
2000
1010
1111
1210
1400
1001
0001
0211
0012
0111
0101
1111
1101
0111
0110
0101
1110
0000
0000
0012
1102
0011
1001
1000
0010
1100
0000
0101
1R.
igneu
s20
0110
1011
1112
1014
0010
0100
0102
1100
1201
1101
0111
1211
0101
1101
1001
0111
1000
0000
0000
1211
0200
1110
0100
0000
1000
0000
0001
00?
R.ja
neiro
ensis
2000
1010
1111
1211
1110
1001
0000
0111
0012
0111
0101
?111
0100
0111
1110
0001
1110
0000
0000
0011
1002
0010
1400
1000
0101
1110
0100
0001
1R.
kiro
vsky
i20
1210
0011
1112
1011
0010
0100
0001
1100
1202
1101
0111
1021
0001
1111
1000
0111
1000
0000
0000
0410
0301
1110
0010
0002
0000
0000
0000
00?
R.lit
terat
us20
0210
0011
1112
1013
0010
0110
0001
1100
1202
1101
0111
1011
0001
1111
1000
0111
1000
0110
0000
1310
0300
1014
0000
0011
0000
0010
0001
112
R.lu
eling
i20
0010
1011
1112
1111
1010
0100
0101
1100
1201
1101
0111
1001
0001
1111
1000
0111
1000
0000
0000
1110
0200
1114
0010
0001
0111
1001
0000
012
R.m
icrop
us20
0010
1011
1112
1114
0010
0100
0101
1100
1201
1101
0111
1101
0001
1101
1000
0111
1000
0000
0000
1111
0200
1110
0010
0000
1000
0000
0001
011
R.m
odest
us20
0210
0011
1112
1011
0010
0110
0001
1100
1202
1101
0111
1011
0001
1111
1000
0111
1000
0110
0000
1310
0300
100?
0000
0011
0000
0000
0000
112
R.or
natu
s20
0010
1011
1112
0111
0110
0100
0001
1100
1201
1101
0111
1021
0001
1111
1000
0111
1000
0000
0130
1410
0200
1013
0310
0002
1000
0010
0000
00?
R.pi
ctus
2002
1000
1111
1210
1300
1001
1000
0111
0112
0201
0101
1110
1100
0111
1110
0001
1110
0001
1000
0011
1003
0010
1200
0000
1100
0000
1000
0111
2R.
punc
tatu
s20
0210
0011
1112
1011
0010
0110
0001
1100
1202
1101
0111
1011
0001
1111
1000
0111
1000
0110
0000
1310
0300
1012
0200
0011
0000
0000
0001
112
R.ro
mer
i20
1210
0011
1112
1011
0010
0100
0001
1110
1202
1101
0111
1021
0001
1111
1000
0111
1000
0020
0000
0410
0301
1010
0010
0002
0000
0000
0000
00?
R.sa
nten
sis20
0010
1011
1112
1011
1010
0100
0?01
1100
1201
1101
01?111
1100
0111
1110
0001
1110
0000
0000
0011
1002
0010
1400
1000
0101
1110
0100
0001
1R.
striga
tus
2002
1000
1111
1210
1100
1001
001?01
1100
1202
1101
1111
1001
0001
1111
1000
0111
1000
0110
0000
1310
0200
1111
1010
0001
0010
1010
0001
00?
R.ta
enia
tus
2000
1010
1111
1211
1100
1001
0001
0211
0012
0111
0101
1111
0100
0111
0110
0001
1110
0000
0000
0011
1002
0011
1000
1000
0010
0100
0001
0101
1R.
tecm
inae
2012
1000
1111
1210
1100
1001
0000
0111
0012
0211
0101
1110
2100
0111
1110
0001
1110
0100
0000
2004
1003
0011
1000
1100
0200
1100
0000
0001
1R.
tenui
s20
0010
1011
1112
1111
0010
0100
0101
1100
1201
1101
0111
1001
0001
1101
1000
0111
1000
0000
0000
1111
0200
1010
00?000
0110
1101
0001
0101
1R.
uakt
i20
1?10
0011
1112
1011
0010
0100
0001
1100
1202
1101
0111
1021
0001
1111
1000
0111
1001
0010
0000
0410
0300
1114
0011
0002
0000
0000
0000
011
R.ua
tum
an20
1210
0011
1112
1011
0010
0100
0001
1110
1202
1101
0111
1021
0001
1111
1000
0111
1000
0000
0000
0410
0301
1110
0010
0002
0000
0000
0000
00?
R.ur
opht
halm
us20
0010
1011
1112
1111
0010
0100
0102
1100
1201
1101
0111
1101
0001
1101
1000
0111
1000
0000
0000
1110
0200
1010
0110
0000
1011
0000
0101
011
R.vi
olace
us20
0210
0011
1112
1011
0010
0110
0001
1100
1202
1101
0111
1011
0001
1111
1000
0111
1000
0110
0000
1310
0300
1012
0000
0011
0011
0010
0001
112
R.zy
gone
ctes
2002
1000
1111
1210
1100
1001
1000
0111
0012
0211
0101
1110
1100
0111
1110
0001
1110
0001
1000
0013
1003
0010
1202
0000
1100
0000
0000
0111
2T.
rubr
omar
ginat
us20
0111
1010
1111
1014
0010
0100
0101
1100
1201
1101
0011
1211
0101
1101
1100
0111
1001
0220
0000
1211
0210
0010
0000
1000
0010
0000
0000
00?
Appendix 3Matrix of 113 characters for 49 aplocheiloid species. Characters and states are according to Appendix 2. Autapomorphies were
excluded from the analysis; 0 = plesiomorphic state; 1-4 = apomorphic states; ? = not pertinent or unknown state.

Fifteen years ago, the idea of publishing a new scientificjournal on aquatic biology with a strong focus on fisheswas conceived, and in March 1994 the first issue of “aqua– Journal of Ichthyology and Aquatic Biology” saw thelight of day. One of the guiding considerations lay in thefact that many fish species and other aquatic animals areprimarily distinguished by their colour pattern and acolour photograph of a life specimen may explain morethan a thousand words. Still, aquatic biologists did nothave adequate access to a scientific journal, which wouldpublish in colour without imposing charges that mostauthors could not afford. The scientific community enthu-siastically welcomed aqua and many of the world’s leadingichthyologists submitted their contributions. In responseto the ever-increasing number of papers received, aquaincreased its output from originally 16 to 48 pages perissue, and from two to six issues per year.With a new Scientific Editor on board, the Managing
Editor decided to review the first 12 years of aqua’s exis-tence. A total of 168 papers have been published in vol-umes 1 to 11. Out of these, two papers were on decapodcrustaceans, one on ascidians, two on amphibians and rep-tiles, and two on dolphins. The remaining 161 papers, or96 % of all contributions, were on fish. Further researchrevealed that non-ichthyologists perceived aqua as anichthyological journal, while ichthyologists were notalways certain about its profile. As a consequence, wedecided to change the name to “aqua – International Jour-nal of Ichthyology”. This change will become effective as
Subscription Notice
As of volume 12, the subscription rate per volume of aqua (= 4 issues, each issue comprising at least 48 pages) will be:
Personal subscription: Euro 75 (including priority mail)
Institutional subscription: Euro 140 (including priority mail)
In Italy the costs of mail services have increased – actually tippled – during the last three years and also printing costs havegone up. For these reasons the subscription rates had to be adjusted and we trust that our readership will understand.
aqua vol. 11 no. 4 - 2006 176
of volume 12. Although there are abundant examples ofname changes in scientific publishing, any editor would bereluctant to alter the name of a well-established journal.However, we feel it is timely to reflect in a new title whathas for a long period been reality on the printed page. Youwill also find some changes in our Guidelines for Authors,in the journal’s layout and in its letter font, all being partof an updated profile of aqua.In our efforts at adapting to constantly changing require-
ments in the scientific fields covered by aqua, we are sup-ported by the members of our Editorial Board. As of issue11 (3) a new Board has been formed and we may againdraw on the expertise of eminent scientists from variouscountries, who agreed to make available their expertise ina wide range of specialisations, including emerging fields,such as molecular genetics, which is of increasing impor-tance in fish systematics and phylogeny. We express ourgratitude to the members of the outgoing Board for thetime and effort they have spent advising us on editorialmatters. We warmly welcome the new members and lookforward to an inspiring and fruitful cooperation.The changes described here above are part of our contin-
uous efforts at providing the international community ofichthyologists with an efficiently published journal, meet-ing high scientific and technical standards. We look for-ward to reading your comments.
The Editors
A note from the Editors:As of volume 12, aqua will change its name

BLABER, S. J. M. 1980. Fish of the Trnity inlet systemof North Queensland, with notes on the ecologyof fish faunas of tropical Indo-Pacific estuaries.Australian Journal of Marine and FreshwaterResearch 31:137-46.
DAY, J. H., BLABER, S. J. M., & WALLACE, J. H. 1981.Estuarine fishes. In: Estuarine Ecology with Parti-cular Reference to Southern Africa. (Ed. J.H. Day.):197-221. A. A. Balkema, Rotterdam.
DIMMICH, W. W. 1988. Ultrastructure of North Ame-rican cyprinid maxillary barbels. Copeia 1988 (1):72-79.
TREWAVAS, E. 1983. Tilapiine Fishes of the GeneraSarotherodon, Oreochromis and Danakilia.British Museum (Natural History), London, 583 pp.
7. Submission of manuscript and illustrations: Themanuscript and illustrations must be submitted digi-tally to the Scientific Editor:
Dr. Friedhelm KruppSenckenberg Research Institute
Senckenberganlage 2560325 Frankfurt am Main, Germany
E-mail: [email protected]: +49-69-7542.255, Fax: +49-69-7542.253
to whom all subsequent correspondence shall beaddressed. Texts, tables, and graphs should be inMicrosoft-compatible electronic form.
After the paper has been accepted for publication, illus-trations as high-resolution TIF files or original pho-tographs (ideally transparencies; otherwise glossyprints, preferably in the size in which they will appear -the type area of aqua is 158 x 224 mm, one column is76 mm wide) must be sent to:
Aquapress, The Managing EditorVia G. Falcone 11,
27010 Miradolo Terme (Pavia), ItalyE-mail: [email protected]
Authors should retain copies of all materials for refer-ence.
Proofs of accepted papers will be sent as PDF files by e-mail attachment to the corresponding author.
8. Evaluation of manuscripts: manuscripts will be eval-uated by the editors and referees. Papers are accepted onthe understanding that they have not and will not bepublished elsewhere.
9. Reprints: Authors will receive one free copy of theissue in which their paper appears and an e-print inPDF format. Additional copies may be ordered fromAquapress.
1. Manuscript preparation: manuscripts must be sub-mitted in English. In exceptional cases aqua may pro-vide translations of manuscripts written in French, Ger-man, Italian, or Spanish.
Manuscripts must be word-processed in MicrosoftWORD and submitted in an electronic form. Generic,specific, and sub-specific names must be italicised. Allpapers must conform to the International Code of Zoo-logical Nomenclature. Authors are strongly advised tofollow the format set out in previous publications ofaqua.
2. Title: the title must be short and should preciselyidentify the main topic of the article. Names of generaor species are followed by the systematic group to whichthey belong. Author name(s) are given in full beneaththe title, followed by the complete mailing and e-mailaddress(es).
3. Abstract: the abstract should not exceed 250 wordsand draw attention to the principal conclusions. Itshould not contain any uncommon abbreviations or lit-erature citations. The inclusion of abstracts in other lan-guages is optional.
5. Subject matter: the text of the manuscript is usuallyarranged in four main sections: Introduction, Materialsand methods (including a key to abbreviations),Results, and Discussion. Other subdivisions may bechosen depending on the material presented. Acknowl-edgements should be placed between the text and refer-ences.
Scientific names of genera, species, and subspeciesshould be followed by the name(s) of author(s) and theyear of publication on first mention. A description of anew taxon must contain the following sections: Mater-ial, Diagnosis, Description, and Affinities. Holotypeand paratypes must be clearly identified, the institutionin which they have been deposited named, and the cat-alogue numbers given. Private collections are notacceptable as repositories for holotypes.
Synonyms must be arranged in chronological order.Identification keys must be dichotomous.
The metric system and SI units must be used.Temperatures are given in °C. Fractions should not beused.
6. References to literature: the name-year system mustbe used. The list of references should be placed at theend of the paper, alphabetically arranged according toauthor name. Only those publications cited in the papermay be included. Titles of journals must be given infull.
Examples of correct reference formats:
Guidelines for Authors

aquaJournal of Ichthyology and Aquatic Biology
Vol. 11 (4), October 2006
Contents:
Wilson J. E. M. Costa: Relationships and taxonomy of the killifish genus Rivulus(Cyprinodontiformes: Aplocheiloidei: Rivulidae) from the Brazilian Amazonas river basin,with notes on historical ecology ................................................................................................... 133-175
A note from the Editors ............................................................................................................... 176
Papers appearing in this journal are indexed in: Zoological Record;Biolis – Biologische Literatur Information Senckenberg;
www.aquapress-bleher.it; www.aquageo.com; www.Joachim-Frische.com
Cover photo: Rivulus urophthalmus, UFRJ 6264, male, 29.5 mm SL (one day after collection); Brazil: Pará: Altamira.Photo by W. J. E. M. Costa.
Brazil: Pará: Primavera; pool close to creek near forest border, typical habitat of Rivulus urophthalmus.Photo by W. J. E. M. Costa.