arsenic transformation and plant growth promotion ...soils.ifas.ufl.edu/lqma/publication/xu...
TRANSCRIPT

lable at ScienceDirect
Chemosphere 144 (2016) 1233e1240
Contents lists avai
Chemosphere
journal homepage: www.elsevier .com/locate/chemosphere
Arsenic transformation and plant growth promotion characteristicsof As-resistant endophytic bacteria from As-hyperaccumulator Pterisvittata
Jia-Yi Xu a, b, 1, Yong-He Han a, 1, Yanshan Chen a, Ling-Jia Zhu a, Lena Q. Ma a, c, *
a State Key Laboratory of Pollution Control and Resource Reuse, School of the Environment, Nanjing University, Jiangsu 210046, Chinab Geological Survey of Jiangsu Province, Jiangsu 210018, Chinac Soil and Water Science Department, University of Florida, Gainesville, FL 32611, USA
h i g h l i g h t s
* Corresponding author. State Key Laboratory of PoReuse, School of the Environment, Nanjing University
E-mail address: [email protected] (L.Q. Ma).1 The authors contributed equally to this work.
http://dx.doi.org/10.1016/j.chemosphere.2015.09.1020045-6535/Published by Elsevier Ltd.
g r a p h i c a l a b s t r a c t
� 43 As-resistant endophytic bacteriawere identified from Pteris vittata.
� All endophytes showed plant growthpromotion characteristics.
� The roots isolates showed highertolerance to AsV.
� The leaflet and stem endophytesshowed higher tolerance to AsIII.
� Bacterial As resistance was positivelycorrelated to their ability in AsVreduction.
a r t i c l e i n f o
Article history:Received 13 June 2015Received in revised form18 September 2015Accepted 30 September 2015Available online xxx
Keywords:Endophytic bacteriaPteris vittataArsenic resistanceOxidationReduction
a b s t r a c t
The ability of As-resistant endophytic bacteria in As transformation and plant growth promotion wasdetermined. The endophytes were isolated from As-hyperaccumulator Pteris vittata (PV) after growingfor 60 d in a soil containing 200 mg kg�1 arsenate (AsV). They were isolated in presence of 10 mM AsVfrom PV roots, stems, and leaflets, representing 4 phyla and 17 genera. All endophytes showed at leastone plant growth promoting characteristics including IAA synthesis, siderophore production and Psolubilization. The root endophytes had higher P solubilization ability than the leaflet (60.0 vs.18.3 mg L�1). In presence of 10 mM AsV, 6 endophytes had greater growth than the control, suggestingAs-stimulated growth. Furthermore, root endophytes were more resistant to AsV while the leaflet en-dophytes were more tolerant to arsenite (AsIII), which corresponded to the dominant As species in PVtissues. Bacterial As resistance was positively correlated to their ability in AsV reduction but not AsIIIoxidation. The roles of those endophytes in promoting plant growth and As resistance in P. vittatawarrant further investigation.
Published by Elsevier Ltd.
llution Control and Resource, Jiangsu 210046, China.
1. Introduction
Arsenic (As), a toxic metalloid, is ubiquitous in the environment,which results from both natural and anthropogenic sources(Abernathy et al., 2003). Due to its toxicity and carcinogenicity, Ascontamination in soils poses a potential risk to public health (Zhao

J.-Y. Xu et al. / Chemosphere 144 (2016) 1233e12401234
et al., 2010). Therefore, it is necessary to develop remediationstrategies to reduce its effects on ecosystems and humans.
Phytoremediation is a promising technology to remove As fromsoils, which is cost-effective and environment-friendly comparedto traditional remediation methods. Arsenic hyperaccumulatorPteris vittata (PV) has a potential to be used for phytoremediation inAs-contaminated soils, which accumulates up to 2.3% As in thefronds (Ma et al., 2001). However, most hyperaccumulators areunsuitable for phytoremediation due to their low biomass and lowAs uptake (Lebeau et al., 2008). So plant-associated microbes haveattracted attention for their roles in promoting plant growth andenhancing metal resistance in contaminated soils (Glick, 2010).
Endophytic bacteria (endophytes) colonize the internal planttissues with little negative effects on their host (Schulz and Boyle,2006). They help to promote plant growth under adverse condi-tions and have potential to improve phytoremediation (Ryan et al.,2008). Their beneficial effects on plant growth have been attrib-uted to their ability in indole acetic acid (IAA) synthesis, siderophoreproduction, and phosphate solubilization (Rajkumar et al., 2009).While IAA helps plant growth, siderophores solubilize Fe from soilsto enhance plant Fe uptake. Many endophytic bacteria from hyper-accumulators have shown plant growth promotion (PGP) charac-teristics. For example, Luo et al. (2011) isolated 30 endophytes fromCd-hyperaccumulator Solanumnigrum fromamine tailing,with 70%strains producing IAA at 1.60e122 mg L�1 and 8 strains solubilizingphosphate at 22e400 mg L�1. Visioli et al. (2014) isolated 5 endo-phytes from Ni-hyperaccumulator Noccaea caerulescens, showingability of siderophore production. However, endophytes isolatedfromPVgrown in clean soils by Zhu et al. (2014) showed limited PGPcharacteristics. It is possible that PGPability of a plant is related to itsmetal resistance. Therefore it is necessary to isolate and characterizeAs-resistant endophytic bacteria fromPVgrown inAs-contaminatedsoils.
Besides PGP traits, endophytes exhibit high resistance to metalstress, which helps to alleviate metal phytotoxicity and improveplant growth (Sessitsch et al., 2013). Since hyperaccumulators havehigh metal concentrations in their biomass, endophytes probablyhave developed mechanisms to adapt to high metal stress (Idriset al., 2004). For instance, Long et al. (2011) found endophytefrom Sedum alfredii tolerated Zn and Cd while endophytes fromhyperaccumulators Thlaspi goesingense, Alyssum bertolonii, and S.nigrum showed high tolerance to Ni and Cd (Barzanti et al., 2007;Idris et al., 2004; Luo et al., 2011).
However, only one study focused on endophytes from PVgrowing in a clean soil (Zhu et al., 2014). It is known that microbialcomposition and diversity are affected by soil As contamination.Furthermore, endophytes from hyperaccumulators growing incontaminated soils are probably more tolerant to metals than thosefrom clean soils. Zhu et al. (2014) found that the ability of PV en-dophytes to reduce AsV and oxidize AsIII was related to their Astolerance. Nevertheless, the study had limited number of PV en-dophytes. In addition, rhizosphere bacteria of PV have beeninvestigated for their As resistance and As transformation, somerhizobacteria showed a dual function of AsIII oxidation and AsVreduction (Ghosh et al., 2011; Huang et al., 2010; Wang et al., 2012).However, there is limited information on endophytes associatedwith PV and their roles in As tolerance and transformation.
The objectives of this study were to: (1) isolate and characterizeAs-resistant endophytic bacteria from PV grown in a soil containing200 mg kg�1 AsV, and (2) explore the relationship between bac-terial ability in As resistance and As transformation. The resultsfrom these investigations are important for better understandingAs tolerance and transformation mechanisms in As-resistantendophytes and help to improve phytoremediation of As-contaminated soils.
2. Materials and methods
2.1. Plant cultivation in As-spiked soils
To obtain endophytic bacteriawith high As-resistance, PV plantswere grown in a soil containing 200 mg kg�1 AsV for 60 d (Xu et al.,2014). The plants were propagated using spores from University ofFlorida, Gainesville, USA. The spores were sown onto a moist soil(40% soil, 40% peat moss, and 20% quartz sand) in seedling boxeswith a transparent cover to maintain moisture. After spores weregerminated and developed to sporelings with 2e3 fronds, theywere transplanted into plastic pots with 10 cm diameter. They werecultivated until they had 5�6 fronds and ~10 cm in height beforetransplant.
Surface soil (0e15 cm) was collected from Nanjing, China. Thesoil properties were as follows: pH 7.02, 0.98% OC, 6.79 mg kg�1As,673 mg kg�1 Mn, and 27.5 g kg�1 Fe (Xu et al., 2014). The soil wasair-dried, sieved through a 2-mm sieve and mixed thoroughly withsolution containing Na2HAsO4$7H2O to reach 200 mg kg�1 As(SigmaeAldrich,�98%). Basal fertilizers (120mg kg�1 N as NH4NO3,and 30 mg kg�1 P and 75.5 mg kg�1 K as K2HPO4) and 1% organicmatter were mixed in the soil. After aging for two weeks at ~50%field capacity, PV plants were transplanted into one per pot with 3replicates. The plants grew in a growth chamber with 25 �C averagetemperature, 70% humidity, 16 h photoperiod and light density of350 mmol m�2 s�1. The plants were watered daily as required andgrew for 60 d before being harvested.
2.2. Isolation of As-resistant endophytic bacteria
Three PV plants were collected and washed thoroughly withMilli-Q water, and then separated into the roots, stems and leaflets,which are referred to as R, S and L for their location. The endophyticbacteria were isolated according to Zhu et al. (2014). The tissueswere surface-sterilized by sequential immersion in 70% ethanol and2% sodium hypochlorite solution for 30 min, and then rinsedthoroughly with sterile distilled water. To validate surface disin-fection, 100 mL of final washing water was plated onto Lur-iaeBertani (LB) agar media, followed by incubation at 28 �C for 3d and the plates showed no microbe.
The surface-sterilized plant tissues were macerated in a sterilemortar containing 10 mL sterile phosphate buffered saline (pH 7.2),and ground with a sterile pestle. Meanwhile, sterile quartz sandwas added to improve cell wall disruption. Tissue extracts (100 mL)and their serial dilutions were plated onto modified LB agar media(Ghosh et al., 2011) supplemented with 10 mM As (Na2HA-sO4$7H2O). After incubation at 28 �C for 7 d, colonies with differentmorphologies were picked out and purified by re-streaking ontothe modified LB agar media. The isolate colony was ensured toreach morphologically homogenous. All purified bacteria werestored in 15% glycerol at �80 �C until analyses.
2.3. Identification of As-resistant endophytic bacteria
The bacterial isolates were cultured overnight in LB broth andthen extracted for genomic DNA by an EZNA® Bacterial DNA Kit(Omega Bio�Tek, USA). The quantity and quality of the extractedDNAwere determined byaNanodropND�2000 Spectrophotometer(NanoDrop Co., USA). Further identificationwas carried out throughPCR amplification of 16S rRNA following Zhu et al. (2014). Theamplificationprimerswere 27F (50�GAGTTTGATCACTGGCTCAG�30)and 1492R (50�TACGGCTACCTTGTTACGACTT�30) (Zhu et al., 2014).PCR products were examined electrophoretically in a 1.5% (w/v)agarose gel.
The amplified products of ~1400 bp 16S rRNA were sequenced

J.-Y. Xu et al. / Chemosphere 144 (2016) 1233e1240 1235
by Sangon Biotechnology Company (Shanghai, China). To checksimilarities, the isolate sequence was compared with nucleotidesequences in the NCBI database using the Basic Local AlignmentSearch Tool (BLAST). The neighbor-joining phylogenetic tree wasconstructed by software MEGA5.1 with 1000 replicates to producebootstrap values. The sequences of all isolates in this study weresubmitted to the NCBI GenBank database. The accession numbersare from KP072747 to KP072769 and from KP125967 to KP125976.
Fig. 1. Neighbor-joining phylogenetic tree showing the relationships among the 16S rRNA s200 mg kg�1 AsV. Bootstrap numbers are given at the node. R represents roots; S represen
2.4. Determination of plant growth promoting properties
The bacterial isolates were activated at 28 �C and 200 rpm byshaking incubators. To evaluate growth, the optical density at600 nm (OD600) of cultures was measured by a UVevis spectro-photometer (UV�2550, Shimadzu, Japan). After incubation, OD600of the cultures was ~1.5 and then adjusted to the same level forinoculation. All experiments were performed on sterile conditions
equences of representative endophytes from PV after growing 60 d in a soil containingts stems; and L represents leaflets.

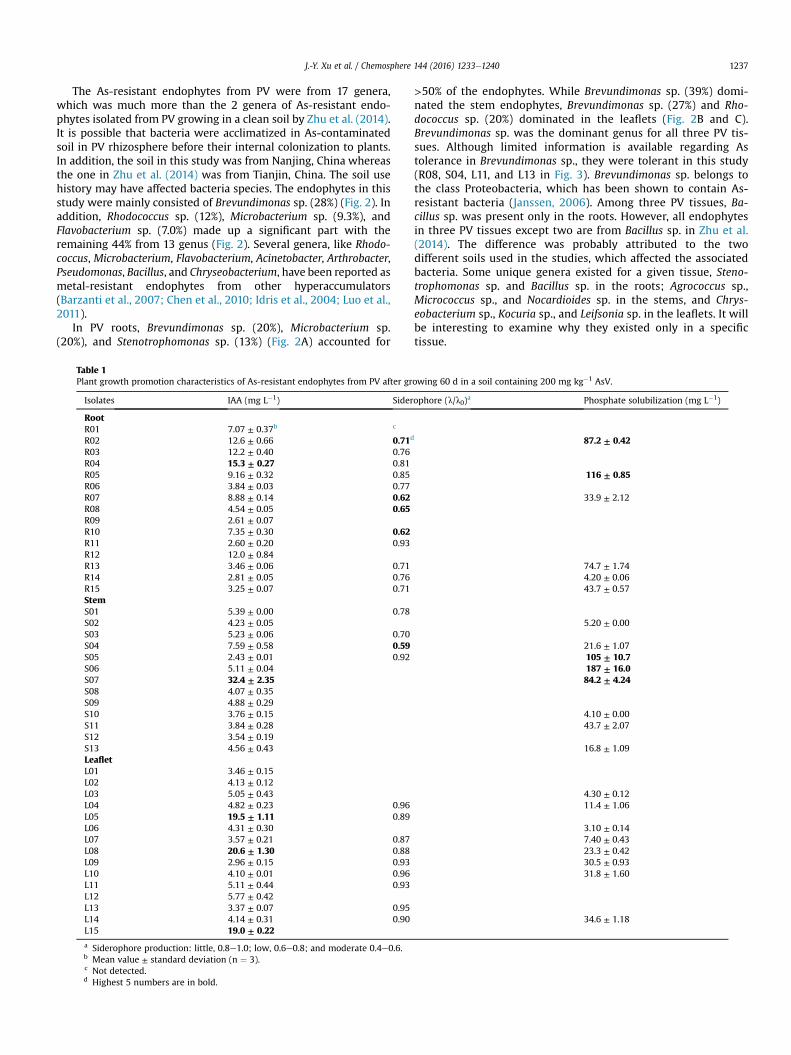
Fig. 2. Diversity of cultivable bacteria in the roots (A), stems (B), and leaflets (C) of PVafter growing 60 d in a soil containing 200 mg kg�1 AsV. Numbers indicate the relativeabundance, expressed as a percentage of total endophytes.
J.-Y. Xu et al. / Chemosphere 144 (2016) 1233e12401236
and in triplicate. Treatment without bacterial isolates was includedas a control.
National Botanical Research Institute's phosphate (NBRIP)growth medium was used to determine phosphate solubilizingability of the isolates (Luo et al., 2011). The NBRIP medium wascomposed of 5 g L�1 glucose, 5 g L�1 MgCl2$6H2O, 0.25 g L�1
MgSO4$7H2O, 0.2 g L�1 KCl, 0.1 g L�1 (NH4)2SO4, and amended with5 g L�1 Ca3(PO4)2 (pH 6.8e7.0). 20 mL NBRIP medium was inocu-lated with 0.2 mL cultures followed by incubation in a shaker at28 �C, 200 rpm for 8 d. The culture supernatant was obtained bycentrifuging at 4000 g for 15 min and the solubilized phosphatewas measured spectrophotometrically at 880 nm using the modi-fied molybdenum blue method.
Production of IAA was analyzed in sucrose minimal salts (SMS)medium (Sheng et al., 2008). 0.2 mL isolate was cultured in 20 mLSMS medium consisting of 10 g L�1 sucrose, 2 g L�1 K2HPO4, 1 g L�1
(NH4)2SO4, 0.5 g L�1 MgSO4, 0.5 g L�1 yeast extract, 0.5 g L�1 CaCO3,0.1 g L�1 NaCl, and 0.5 g L�1
L-tryptophan (pH 7.2). After incubationat 28 �C, 200 rpm for 4 d, 1 mL supernatant was collected bycentrifugation at 4000 g for 15 min and then mixed thoroughlywith 2 mL Salkowski's reagent (50 mL of 35% HClO4 þ 1mL of 0.5 MFeCl3). The mixture was incubated at room temperature for 40 minuntil a stable pink color occurred, followed by measuring absor-bance at 530 nm.
Chrome azuroleS (CAS) analytical method was used to deter-mine siderophore production (Chen et al., 2010). Briefly, 0.2 mLisolate was cultivated at 28 �C, 200 rpm for 2 d in the modifiedsucrose aspartic acid medium containing 20 g L�1 sucrose, 2 g L�1
aspartic acid, 1 g L�1 K2HPO4 and 0.5 g L�1 MgSO4$7H2O (pH 7.0).After centrifuging at 4000 g for 15 min, 1 mL supernatant wascollected and mixed thoroughly with 1 mL CAS solution. The su-pernatant in uninoculated media was used as a control. Followingby 3 h incubation in the dark at room temperature, the absorbanceat 630 nmwas detected and the value was compared with OD630 ofthe control reference (l/l0), which represented siderophoreproduction.
2.5. Arsenic resistance and transformation by endophytes
All cultivable isolates were tested for their As resistance byinoculation in modified LB liquid media spiked with 10 mM AsV(Na2HAsO4$7H2O) or AsIII (NaAsO2). After incubation at 28 �C and200 rpm for 2 d, the cultures were collected and determined fortheir OD600. The mediawithout microbes were used as a control. Asresistance by endophytes was indicated by bacterial growth in As-media compared to that in the control.
To further examine As transformation by endophytes, 10 endo-phytes from roots (R), stems (S), and leaflets (L) (R01, R07, R15, S09,S10, S12, S13, L01, L02, and L11) with different As tolerance werecultured in modified LB liquid media with 10 mM AsV or AsIII at28 �C and 200 rpm for 2 d. The supernatants of cultures werecollected and analyzed for AsV and AsIII using high performanceliquid chromatography (HPLC; Waters 2695, USA) coupled withinductively coupled plasma mass spectrometry (ICP�MS; Perki-nElmer NexION 300X, USA). The media without microbes wereused as control and all experiments were conducted in triplicate.
2.6. Statistical analysis
Statistical analysis was performed according to the linear modelprocedure of the statistical analysis system (SAS, 1985). The datawere processed using analysis of variance (ANOVA). To examine thestatistical significance of differences (P < 0.05) between means, theTukey test was performed. Simple linear correlations between Asresistance and transformation were established by SigmaPlot 12.5.
3. Results and discussion
3.1. High diversity of As-resistant endophytic bacteria from PV
To obtain As-resistant endophytic bacteria from PV, only endo-phytes survived at 10 mM AsV were isolated and characterized.Based on their morphological characteristics, 43 As-resistant en-dophytes were isolated, with even distribution among PV roots,stems, and leaflets (15, 13 and 15) (data not shown). Though thetotal number of endophytes was similar to Zhu et al. (2014) whoisolated 39 endophytes from PV growing in a clean soil, the endo-phyte distribution in tissues was different. Zhu et al. (2014) showedthat the roots have the most endophytes at 20, with 9 and 12 en-dophytes from the stems and leaflets.
The endophytes were identified by 16S rRNA sequencing andanalyzed for their phylogenetic relationship. For As-resistant endo-phytes from PV, a high diversity was observed including 4 phyla and17 genera (Fig.1). Proteobacteria (47%) and Actinobacteria (42%) werethe most dominant, followed by Bacteroidetes (9.3%) and Firmicutes(2.3%) (Fig. 1). Similarly, the 30 Cd-resistant endophytes from Cdhyperaccumulator S. nigrumwere also from these 4 phyla, includingActinobacteria (43%), Bacteroidetes (27%), Proteobacteria (23%), andFirmicutes (7%) (Luo et al., 2011). The Ni-resistant endophytes fromthe roots of Ni hyperaccumulator N. caerulescens consisted of Fir-micutes (41%), Actinobacteria (35%), Proteobacteria (12%), and Bac-teroidetes (12%) (Visioli et al., 2014). These results suggested mostmetal-resistant endophytes belong to these 4 phyla including Pro-teobacteria, Actinobacteria, Bacteroidetes, and Firmicutes though theproportion varied with metal hyperaccumulators.
J.-Y. Xu et al. / Chemosphere 144 (2016) 1233e1240 1237
The As-resistant endophytes from PV were from 17 genera,which was much more than the 2 genera of As-resistant endo-phytes isolated from PV growing in a clean soil by Zhu et al. (2014).It is possible that bacteria were acclimatized in As-contaminatedsoil in PV rhizosphere before their internal colonization to plants.In addition, the soil in this study was from Nanjing, China whereasthe one in Zhu et al. (2014) was from Tianjin, China. The soil usehistory may have affected bacteria species. The endophytes in thisstudy were mainly consisted of Brevundimonas sp. (28%) (Fig. 2). Inaddition, Rhodococcus sp. (12%), Microbacterium sp. (9.3%), andFlavobacterium sp. (7.0%) made up a significant part with theremaining 44% from 13 genus (Fig. 2). Several genera, like Rhodo-coccus, Microbacterium, Flavobacterium, Acinetobacter, Arthrobacter,Pseudomonas, Bacillus, and Chryseobacterium, have been reported asmetal-resistant endophytes from other hyperaccumulators(Barzanti et al., 2007; Chen et al., 2010; Idris et al., 2004; Luo et al.,2011).
In PV roots, Brevundimonas sp. (20%), Microbacterium sp.(20%), and Stenotrophomonas sp. (13%) (Fig. 2A) accounted for
Table 1Plant growth promotion characteristics of As-resistant endophytes from PV after gr
Isolates IAA (mg L�1) Sider
RootR01 7.07 ± 0.37b c
R02 12.6 ± 0.66 0.71d
R03 12.2 ± 0.40 0.76R04 15.3 ± 0.27 0.81R05 9.16 ± 0.32 0.85R06 3.84 ± 0.03 0.77R07 8.88 ± 0.14 0.62R08 4.54 ± 0.05 0.65R09 2.61 ± 0.07R10 7.35 ± 0.30 0.62R11 2.60 ± 0.20 0.93R12 12.0 ± 0.84R13 3.46 ± 0.06 0.71R14 2.81 ± 0.05 0.76R15 3.25 ± 0.07 0.71StemS01 5.39 ± 0.00 0.78S02 4.23 ± 0.05S03 5.23 ± 0.06 0.70S04 7.59 ± 0.58 0.59S05 2.43 ± 0.01 0.92S06 5.11 ± 0.04S07 32.4 ± 2.35S08 4.07 ± 0.35S09 4.88 ± 0.29S10 3.76 ± 0.15S11 3.84 ± 0.28S12 3.54 ± 0.19S13 4.56 ± 0.43LeafletL01 3.46 ± 0.15L02 4.13 ± 0.12L03 5.05 ± 0.43L04 4.82 ± 0.23 0.96L05 19.5 ± 1.11 0.89L06 4.31 ± 0.30L07 3.57 ± 0.21 0.87L08 20.6 ± 1.30 0.88L09 2.96 ± 0.15 0.93L10 4.10 ± 0.01 0.96L11 5.11 ± 0.44 0.93L12 5.77 ± 0.42L13 3.37 ± 0.07 0.95L14 4.14 ± 0.31 0.90L15 19.0 ± 0.22
a Siderophore production: little, 0.8e1.0; low, 0.6e0.8; and moderate 0.4e0.6.b Mean value ± standard deviation (n ¼ 3).c Not detected.d Highest 5 numbers are in bold.
>50% of the endophytes. While Brevundimonas sp. (39%) domi-nated the stem endophytes, Brevundimonas sp. (27%) and Rho-dococcus sp. (20%) dominated in the leaflets (Fig. 2B and C).Brevundimonas sp. was the dominant genus for all three PV tis-sues. Although limited information is available regarding Astolerance in Brevundimonas sp., they were tolerant in this study(R08, S04, L11, and L13 in Fig. 3). Brevundimonas sp. belongs tothe class Proteobacteria, which has been shown to contain As-resistant bacteria (Janssen, 2006). Among three PV tissues, Ba-cillus sp. was present only in the roots. However, all endophytesin three PV tissues except two are from Bacillus sp. in Zhu et al.(2014). The difference was probably attributed to the twodifferent soils used in the studies, which affected the associatedbacteria. Some unique genera existed for a given tissue, Steno-trophomonas sp. and Bacillus sp. in the roots; Agrococcus sp.,Micrococcus sp., and Nocardioides sp. in the stems, and Chrys-eobacterium sp., Kocuria sp., and Leifsonia sp. in the leaflets. It willbe interesting to examine why they existed only in a specifictissue.
owing 60 d in a soil containing 200 mg kg�1 AsV.
ophore (l/l0)a Phosphate solubilization (mg L�1)
87.2 ± 0.42
116 ± 0.85
33.9 ± 2.12
74.7 ± 1.744.20 ± 0.0643.7 ± 0.57
5.20 ± 0.00
21.6 ± 1.07105 ± 10.7187 ± 16.084.2 ± 4.24
4.10 ± 0.0043.7 ± 2.07
16.8 ± 1.09
4.30 ± 0.1211.4 ± 1.06
3.10 ± 0.147.40 ± 0.4323.3 ± 0.4230.5 ± 0.9331.8 ± 1.60
34.6 ± 1.18

J.-Y. Xu et al. / Chemosphere 144 (2016) 1233e12401238
3.2. As-resistant endophytes with plant growth promotingcharacteristics
Endophytes with PGP characteristics benefit plant growth andincrease their metal resistance growing in contaminated sites (Longet al., 2011; Sheng et al., 2008). Endophytes are mostly from soilswith some from phyllosphere (Rajkumar et al., 2009). In this study,all endophytes showed at least one PGP property and 33% showedall three traits (Table 1). All endophytes showed ability to synthe-size IAA while 58% and 51% of endophytes showed ability to pro-duce siderophore and solubilize Ca3(PO4)2.
Theendophytesproduced2.43e32.4mgL�1 IAA,withS07 fromPVstem producing the highest (Table 1). These were greater than thosereported by Zhu et al. (2014), showing 0.2e10.8 mg L�1 IAA. Though58% of the endophytes produced siderophore, the levels were low at0.59e0.96, with the highest from S04 at 0.59, which was moderate.However, the endophytes showedno ability inproducing siderophorein Zhu et al. (2014). The data suggested that As-resistant endophytesproduced some IAA, but were not efficient producers of siderophore.
In addition, over half of the endophytes were able to solubilizesparingly-soluble P mineral Ca3(PO4)2 at 3.1e187 mg L�1, of which 3endophytes solubilized over 100 mg L�1 P, including S06, R05 and
R15
R01
R06
R12
R10
R08
R14
R05
R07
R02
R09
R03
R13
R04
R11
Rel
ativ
e gr
owth
as %
of c
ontro
l
020406080
100120140160180
S12
S04
S05
S07
S03
S13
S09
S08
S10
S02
S11
S01
S06
Rel
ativ
e gr
owth
as %
of c
ontro
l
0
20
40
60
80
100
120
140
L08
L11
L10
L04
L12
L15
L07
L13
L03
L09
L14
L06
L05
L01
L02
Rel
ativ
e gr
owth
as %
of c
ontro
l
0
20
40
60
80
100
120
A
C
E
Fig. 3. Arsenic resistance of 43 endophytic bacteria from PV roots, stems, and leaflets in modfor 2 d at 28 �C, 200 rpm. R represents roots; S represents stems; and L represents leaflets
S05 (Table 1). Theywere considered efficient P solubilizer (Rajkumaret al., 2009). Phosphate solubilizing endophytes were from all threetissues, with PV roots, stems, and leaflet endophytes solubilizing 60,59, and 18mg L�1 P (Table 1). The results suggested that endophytesfrom PV roots and stems had greater ability in P solubilization,which was probably associated with P-solubilizing bacteria in therhizosphere. Most P in soils is insoluble, resulting in low P avail-ability to plants (Richardson et al., 2009). Under P deficiency, Psolubilizing bacteria are common in the rhizosphere (Mander et al.,2012; Oliveira et al., 2009). By internal colonization, endophytesprobably enhance P availability to the host plant (Compant et al.,2010; Rajkumar et al., 2009). Overall, the endophytes in this studyhad more prominent PGP characteristics than those from Zhu et al.(2014). Microbial PGP ability may be related to their metal resis-tance. In our study, PV plants and their endophytes grew in As-containing soil for 60 d, probably resulting in better PGP ability.
3.3. Root endophytes more resistant to AsV and leaflet endophytesmore tolerant to AsIII
As-resistant endophytes were isolated from PV by incubation inmodified LB agar media containing 10 mM AsV. To further measure
R15
R12
R01
R09
R13
R05
R10
R06
R08
R11
R14
R04
R02
R03
R07
0
20
40
60
80
100S0
9S0
4S1
0S0
8S0
3S1
1S0
5S0
7S0
2S0
1S0
6S1
2S1
3
0
20
40
60
80
100
120
L09
L08
L03
L06
L04
L05
L10
L13
L07
L01
L14
L12
L11
L15
L02
0
20
40
60
80
100
B
D
F
ified LB media with 10 mM AsV (ACE) and AsIII (BDF). The endophytes were incubated. The bars indicate the standard error of triplicates.

J.-Y. Xu et al. / Chemosphere 144 (2016) 1233e1240 1239
their As tolerance, all 43 endophyteswere inoculated to liquidmediasupplementedwith 10mMAsIII or AsV. Compared to agar media, Asbioavailability is higher in liquid media (Huang et al., 2010).
All 43 endophytes survived in liquid media with 10 mM AsV(Fig. 3ACE). Among these, 6 endophytes (R01, R06, R12, R15, S12,and L08) had greater growth than the control. It was unclear whatpromoted endophyte growth under As stress. Arsenic has beenreported to promote PV growth, which has been attributed to As-induced P uptake (Xu et al., 2014). It was possible that some en-dophytes living in As-hyperaccumulators have adapted to high Asstress (Idris et al., 2004). However, some endophytes were stillnegatively impacted by As, with 51% endophytes showing 30%growth reduction, and 12% endophytes showing >50% in presenceof AsV.
Similarly, all PV endophytes tolerated 10mMAsIII in liquidmedia(Fig. 3BDF). However, unlike AsV, all endophytes were inhibited by10 mM AsIII with some reduction as much as 90%. Among endo-phytes, S09, L09 and R15 exhibited the greatest resistance, with<20% growth reduction. This was because AsIII is more toxic tobacteria than AsV as AsIII binds to thiol groups and deactivatesenzyme activity (Kaltreider et al., 2001). As a result, 19% endophytesshowing 30% growth reduction, and 40% showing >50% growthreduction in presence of AsIII. When looking at the ability of bac-terial resistance to AsV and AsIII, no relationship was found (Fig. 3).While some endophytes (R15, S4, and L8) were tolerant to both AsVand AsIII, some (R3, S1, and L2) were intolerant to either AsV or AsIII.
In addition to their growth reduction under As stress, wecompared their As tolerance among endophytes from differenttissues. Based on their average growth (89 vs. 69 and 68%), theendophytes from PV roots were more tolerant to AsV than thosefrom PV stems and leaflets (Fig. 3ACE). However, in 10mMAsIII, theleaflet endophytes had higher resistance than the roots and stems(59 vs. 50 and 52%) (Fig. 3BDF), which was similar to Zhu et al.(2014). This was probably attributed to As species in PV tissues,with AsV being dominant in the roots while AsIII in the fronds(Mathews et al., 2010; Xu et al., 2014).
Fig. 4. Arsenic resistance of 10 endophytic bacteria in the media with 10 mM AsV (A) and A10 mM AsV or AsIII. These 10 isolates were incubated for 2 d at 28 �C, 200 rpm. R representsof triplicates. Lowercase letters indicate significant difference among different isolates by T
3.4. AsV reduction by endophytes was positively related to Asresistance
10 endophytes from PV roots, stems and leaflets with differentAs tolerance were selected to examine their ability in As trans-formation of 10 mM AsV or AsIII (Fig. 4AB). Six endophytes hadhigher resistance to AsV than AsIII while S9, S10, L01 and L02 werethe opposite. Particularly, R15 from PV root grew well in bothmedia, showing resistance to both AsIII and AsV.
Compared to the control, all endophytes were able to transformAs but with varying ability. Among the 10 endophytes, R15 was themost effective in reducing AsV while L01 was the most effective inoxidizing AsIII, both at 15e17% (Fig. 4CD). While S09, S10, L02, andL01 were ineffective in reducing AsV, R07, S12 and S13 were inef-fective in oxidizing AsIII, all being<1% (Fig. 4C D). Interestingly, S09,S10, L02, and L01 were not AsV reducers and they were moreresistant to AsIII than AsV. The data implied that their As resistancemay be related to their ability in AsV reduction.
The endophytes with limited ability in As transformation alsoshowed limited growth in presence of 10 mM AsV or AsIII thanothers (Fig. 4AB). Among 10 endophytes, R15 had the highest abilityin both AsV reduction and AsV resistance (Fig. 4AC). Similarly, en-dophytes with high AsV reduction grew well in AsV, which wassupported by their positive correlation (R2 ¼ 0.68), which is atypical mechanism for microbial As-resistance. In As-resistantbacteria, ArsC mediates the reduction of AsV to AsIII in the cyto-plasm and subsequently AsIII is extruded from cells by ArsB,helping As detoxification (Rosen, 2002; Stolz et al., 2006). Thus, thehigher ability of PV endophytes in AsV reduction, themore resistantthey were to AsV. However, there was no correlation between AsIIIoxidation and AsIII resistance (data not shown). The results weredifferent from Zhu et al. (2014) who reported both AsV reductionand AsIII oxidation by PV endophytes had positive relationshipwithAs tolerance. However, in that research only few PV endophyteswere used for the correlation analysis. Among the 10 endophytes,R15, R01, and L11 exhibited a dual function of AsV reduction and
sIII (B), and their AsV reduction (C) and AsIII oxidation (D) by 10 isolates incubated inroots; S represents stems; and L represents leaflets. The bars indicate the standard errorukey test at P < 0.05.

J.-Y. Xu et al. / Chemosphere 144 (2016) 1233e12401240
AsIII oxidation (Fig. 4CD). It was probably because many AsIII-oxidizing bacteria had an ars operon to oxidize and reduce As(Kashyap et al., 2006; Macur et al., 2004). In addition, the threemost efficient AsIII oxidizers (L01, S09 and L02) were the leasteffective in AsV reduction. Among the three most effective AsVreducers (R15, S13 and R01), only S13 was the least effective AsIIIoxidizer. Hence, it didn't seem therewas a relationship between theability of As reduction and As oxidation in those endophytes.
4. Conclusion
In this study, we isolated and characterized 43 As-resistantendophytic bacteria from the roots, stems, and leaflets of As-hyperaccumulator PV. A high diversity was observed including 4phyla and 17 genera, with Proteobacteria and Actinobacteria beingdominant. Among endophytes, Brevundimonas sp. was the mainspecies in all three PV tissues. All endophytes had at least one PGPcharacteristics. Compared to the leaflets, the endophytes from PVroots and stems had higher ability of phosphate solubilization. Inaddition, all endophytes showed resistance to As. Particularly, 6endophytes had greater growth in presence of 10 mM AsV than thecontrol, probably due to As-stimulated growth. Furthermore, theendophytes from PV roots were more resistant to AsV while thosefrom leaflets were more tolerant to AsIII. Bacterial resistance to AsVwas positively correlated to their ability in AsV reduction but notAsIII oxidation. In addition, bacterial ability in As reduction wasprobably not related to their ability in As oxidation. Our data sug-gest that As-resistant endophytes may help plant growth in PV,thereby increasing its efficiency in phytoremediation of As-contaminated sites.
Acknowledgments
This work was supported in part by the National Natural ScienceFoundation of China (No. 21277070) and the Program for Post-graduates Research and Innovation in Jiangsu Province (No.KYLX_0052).
References
Abernathy, C.O., Thomas, D.J., Calderon, R.L., 2003. Health effects and risk assess-ment of arsenic. J. Nutr. 133, 1536Se1538S.
Barzanti, R., Ozino, F., Bazzicalupo, M., Gabbrielli, R., Galardi, F., Gonnelli, C.,Mengoni, A., 2007. Isolation and characterization of endophytic bacteria fromthe nickel hyperaccumulator plant Alyssum bertolonii. Microb. Ecol. 53,306e316.
Chen, L., Luo, S., Xiao, X., Guo, H., Chen, J., Wan, Y., Li, B., Xu, T., Xi, Q., Rao, C., Liu, C.,Zeng, G., 2010. Application of plant growth�promoting endophytes (PGPE)isolated from Solanum nigrum L. for phytoextraction of Cd�polluted soils. Appl.Soil Ecol. 46, 383e389.
Compant, S., Cl�ement, C., Sessitsch, A., 2010. Plant growth�promoting bacteria inthe rhizo� and endosphere of plants: their role, colonization, mechanismsinvolved and prospects for utilization. Soil Biol. Biochem. 42, 669e678.
Ghosh, P., Rathinasabapathi, B., Ma, L.Q., 2011. Arsenic�resistant bacteria solubilizedarsenic in the growth media and increased growth of arsenic hyperaccumulatorPteris vittata L. Bioresour. Technol. 102, 8756e8761.
Glick, B.R., 2010. Using soil bacteria to facilitate phytoremediation. Biotechnol. Adv.28, 367e374.
Huang, A., Teplitski, M., Rathinasabapathi, B., Ma, L.Q., 2010. Characterization ofarsenic-resistant bacteria from the rhizosphere of arsenic hyperaccumulatorPteris vittata L. Can. J. Microbiol. 56, 236e246.
Idris, R., Trifonova, R., Puschenreiter, M., Wenzel, W.W., Sessitsch, A., 2004. Bacterial
communities associated with flowering plants of the Ni hyperaccumulatorThlaspi goesingense. Appl. Environ. Microbiol. 70, 2667e2677.
Janssen, P.H., 2006. Identifying the dominant soil bacterial taxa in libraries of 16SrRNA and 16S rRNA genes. Appl. Environ. Microb. 72, 1719e1728.
Kaltreider, R.C., Davis, A.M., Lariviere, J.P., Hamilton, J.W., 2001. Arsenic alters thefunction of the glucocorticoid receptor as a transcription factor. Environ. HealthPerspect. 109, 245.
Kashyap, D.R., Botero, L.M., Franck, W.L., Hassett, D.J., McDermott, T.R., 2006.Complex regulation of arsenite oxidation in Agrobacterium tumefaciens.J. Bacteriol. 188, 1081e1088.
Lebeau, T., Braud, A., Jezequel, K., 2008. Performance of bioaugmentation�assistedphytoextraction applied to metal contaminated soils: a review. Environ. Pollut.153, 497e522.
Long, X., Chen, X., Chen, Y., Woon�Chung, W.J., Wei, Z., Wu, Q., 2011. Isolation andcharacterization endophytic bacteria from hyperaccumulator Sedum alfrediiHance and their potential to promote phytoextraction of zinc polluted soil.World J. Microbiol. Biotechnol. 27, 1197e1207.
Luo, S.L., Chen, L., Chen, J.L., Xiao, X., Xu, T.Y., Wan, Y., Rao, C., Liu, C.B., Liu, Y.T., Lai, C.,Zeng, G.M., 2011. Analysis and characterization of cultivable heavy metal-resistant bacterial endophytes isolated from Cd�hyperaccumulator Solanumnigrum L. and their potential use for phytoremediation. Chemosphere 85,1130e1138.
Ma, L.Q., Komar, K.M., Tu, C., Zhang, W., Cai, Y., Kennelley, E.D., 2001. A fern thathyperaccumulates arsenic. Nature 409, 579�579.
Macur, R.E., Jackson, C.R., Botero, L.M., McDermott, T.R., Inskeep, W.P., 2004. Bac-terial populations associated with the oxidation and reduction of arsenic in anunsaturated soil. Environ. Sci. Technol. 38, 104e111.
Mander, C., Wakelin, S., Young, S., Condron, L., O'Callaghan, M., 2012. Incidence anddiversity of phosphate�solubilising bacteria are linked to phosphorus status ingrassland soils. Soil Biol. Biochem. 44, 93e101.
Mathews, S., Ma, L.Q., Rathinasabapathi, B., Natarajan, S., Saha, U.K., 2010. Arsenictransformation in the growth media and biomass of hyperaccumulator Pterisvittata L. Bioresour. Technol. 101, 8024e8030.
Oliveira, C.A., Alves, V.M.C., Marriel, I.E., Gomes, E.A., Scotti, M.R., Carneiro, N.P.,Guimar~aes, C.T., Schaffert, R.E., S�a, N.M.H., 2009. Phosphate solubilizing mi-croorganisms isolated from rhizosphere of maize cultivated in an oxisol of theBrazilian Cerrado Biome. Soil Biol. Biochem. 41, 1782e1787.
Rajkumar, M., Ae, N., Freitas, H., 2009. Endophytic bacteria and their potential toenhance heavy metal phytoextraction. Chemosphere 77, 153e160.
Richardson, A.E., Hocking, P.J., Simpson, R.J., George, T.S., 2009. Plant mechanisms tooptimise access to soil phosphorus. Crop Pasture Sci. 60, 124e143.
Rosen, B.P., 2002. Biochemistry of arsenic detoxification. FEBS Lett. 529, 86e92.Ryan, R.P., Germaine, K., Franks, A., Ryan, D.J., Dowling, D.N., 2008. Bacterial en-
dophytes: recent developments and applications. FEMS Microbiol. Lett. 278,1e9.
Sas, S., 1985. SAS User's Guide: Statistics. SAS Institute Inc, Cary, NC.Schulz, B., Boyle, C., 2006. What are Endophytes?, Microbial Root Endophytes.
Springer, pp. 1e13.Sessitsch, A., Kuffner, M., Kidd, P., Vangronsveld, J., Wenzel, W.W., Fallmann, K.,
Puschenreiter, M., 2013. The role of plant�associated bacteria in the mobili-zation and phytoextraction of trace elements in contaminated soils. Soil Biol.Biochem. 60, 182e194.
Sheng, X.F., Xia, J.J., Jiang, C.Y., He, L.Y., Qian, M., 2008. Characterization of heavymetal-resistant endophytic bacteria from rape (Brassica napus) roots and theirpotential in promoting the growth and lead accumulation of rape. Environ.Pollut. 156, 1164e1170.
Stolz, J.F., Basu, P., Santini, J.M., Oremland, R.S., 2006. Arsenic and selenium in mi-crobial metabolism. Annu. Rev. Microbiol. 60, 107e130.
Visioli, G., D'Egidio, S., Vamerali, T., Mattarozzi, M., Sanangelantoni, A.M., 2014.Culturable endophytic bacteria enhance Ni translocation in the hyper-accumulator Noccaea caerulescens. Chemosphere 117, 538e544.
Wang, X., Rathinasabapathi, B., de Oliveira, L.M., Guilherme, L.R., Ma, L.Q., 2012.Bacteria-mediated arsenic oxidation and reduction in the growth media ofarsenic hyperaccumulator Pteris vittata. Environ. Sci. Technol. 46, 11259e11266.
Xu, J.Y., Li, H.B., Liang, S., Luo, J., Ma, L.Q., 2014. Arsenic enhanced plant growth andaltered rhizosphere characteristics of hyperaccumulator Pteris vittata. Environ.Pollut. 194, 105e111.
Zhao, F.J., McGrath, S.P., Meharg, A.A., 2010. Arsenic as a food chain contaminant:mechanisms of plant uptake and metabolism and mitigation strategies. Annu.Rev. Plant Biol. 61, 535e559.
Zhu, L.J., Guan, D.X., Luo, J., Rathinasabapathi, B., Ma, L.Q., 2014. Characterization ofarsenic�resistant endophytic bacteria from hyperaccumulators Pteris vittataand Pteris multifida. Chemosphere 113, 9e16.