assessing the risk of invasive spread in fragmented landscapes€¦ · · 2004-09-09assessing the...
TRANSCRIPT

Risk Analysis, Vol. 24, No. 4, 2004
Assessing the Risk of Invasive Spreadin Fragmented Landscapes
Kimberly A. With∗
Little theoretical work has investigated how landscape structure affects invasive spread, eventhough broad-scale disturbances caused by habitat loss and fragmentation are believed tofacilitate the spread of exotic species. Neutral landscape models (NLMs), derived from perco-lation theory in the field of landscape ecology, provide a tool for assessing the risk of invasivespread in fragmented landscapes. A percolation-based analysis of the potential for invasivespread in fragmented landscapes predicts that invasive spread may be enormously enhancedbeyond some threshold level of habitat loss, which depends upon the species’ dispersal abili-ties and the degree of habitat fragmentation. Assuming that invasive species spread primarilythrough disturbed areas of the landscape, poor dispersers may spread better in landscapes inwhich disturbances are concentrated in space, whereas good dispersers are predicted to spreadbetter in landscapes where disturbances are small and dispersed (i.e., fragmented landscape).Assessing the risk of invasive spread in fragmented landscapes ultimately requires under-standing the relative effects of landscape structure on processes that contribute to invasivespread—dispersal (successful colonization) and demography (successful establishment). Col-onization success is predicted to be highest when >20% of the landscape has been disturbed,particularly if disturbances are large or aggregated in space, because propagules are more likelyto encounter sites suitable for colonization and establishment. However, landscape patternbecomes less important for predicting colonization success if species are capable of occasionallong-distance dispersal events. Invasive species are also more likely to persist and achievepositive population growth rates (successful establishment) in landscapes with clumped dis-turbance patterns, which can then function as population sources that produce immigrantsthat invade other landscapes. Finally, the invasibility of communities may be greatest in land-scapes with a concentrated pattern of disturbance, especially below some critical threshold ofbiodiversity. Below the critical biodiversity threshold, the introduction of a single species cantrigger a cascade of extinctions among indigenous species. The application of NLMs may thusoffer new insights and opportunities for the management and restoration of landscapes so asto slow the spread of invasive species.
KEY WORDS: Community invasibility; complex systems theory; critical biodiversity; demography;dispersal; landscape connectivity; landscape ecology; neutral landscape models; percolation theory;thresholds
1. INTRODUCTION
Although habitat loss, fragmentation, and in-vasive species collectively pose the greatest threats
∗ Division of Biology, Kansas State University, Manhattan, KS66506, USA; [email protected].
to biodiversity,(1) there has been little theoreticalor empirical research that has addressed how thealteration of landscape structure might promote in-vasive spread.(2) There are several reasons why habi-tat loss and fragmentation might be expected toenhance the spread of invasive species. Humans are
803 0272-4332/04/0100-0803$22.00/1 C© 2004 Society for Risk Analysis

804 With
the primary drivers of land-use change and have alsobeen directly or indirectly responsible for transport-ing species beyond their native range. Human land-use activities may thus enhance the invasibility ofecological communities,(3) particularly through land-scape transformation that results in the destructionand fragmentation of native habitats. Fragmentationis characterized as a “landscape-level” disturbance,(4)
and disturbance is almost unanimously acknowledgedto influence invasive spread.(5) Thus, fragmentationought to enhance the spread of invasive species, andif so, it might be possible to manage landscapes so asto minimize the risk of invasive spread.
Landscape structure may affect any or all ofthe stages of the invasion process that include(1) introduction, (2) colonization, (3) establishment,(4) dispersal, which interacts with landscape structureto create, (5) spatially distributed populations, whichmay set the stage for invasive spread.(2) Although thearrival of an exotic species to a landscape generallyresults from the intentional or accidental introduc-tion by humans, which involves overcoming a geo-graphic barrier, landscape structure may still affectintroduction if it governs human land-use activitiesand enhances the probability of introduction. Estab-lishment depends upon successful colonization, whichmay be a function of propagule pressure or numberof introductions to a locale. Establishment requiresattaining positive population growth rates, whichmay be affected by landscape structure if populationvital rates are affected by fragmentation (e.g., in-creased competition, predation, or parasitism at habi-tat edges may negatively impact birth rates, b).(6) Themovement of organisms or their dispersal vectors canbe affected by the spatial arrangement of habitat orresources,(7–10) and in combination with the aboveprocesses, can lead to spatially structured populationsthat may set the stage for invasion (e.g., nascent foci,Reference 11).
My objective in this article was to explore howalteration of landscape structure in the form ofhabitat loss and fragmentation contributes to therisk of invasive spread. Little theoretical or em-pirical work has directly investigated the effectsof landscape structure—or spatial pattern moregenerally—on invasive spread.(2) Therefore, much ofthe research I highlight in this article was actuallydeveloped to investigate other questions related tofragmentation effects on dispersal, population persis-tence, and community dynamics. Invasive spread is aresult of successful dispersal and positive populationgrowth rates within landscapes, and a general under-
standing of how landscape structure affects these pro-cesses in other species should give us some insight intohow fragmentation might promote invasive spread.The key is to redefine the landscape context of theseother studies in terms of the availability of habitat thatis suitable for invasive spread. Thus, applications thatinvestigated the effect of habitat abundance on popu-lation persistence to demonstrate that extinction riskwas reduced beyond some threshold level of avail-able habitat, can also be interpreted in the context ofidentifying the threshold level of habitat required forpopulation establishment of invasive species (e.g., dis-turbed areas of landscapes) that may promote inva-sive spread.
2. EVALUATING THE POTENTIAL RISKFOR INVASIVE SPREAD
As a first approximation, the risk of inva-sive spread in different landscapes can be mod-eled as a percolation process using neutral land-scape models (NLMs). NLMs were developed in land-scape ecology as an extension of percolation theory,which studies flows through spatially heterogeneousmedia.(12) Neutral landscapes were first presented asa null model of what landscape patterns are expectedin the absence of any organizing or formative pro-cess that typically gives rise to landscape pattern,by assuming a random distribution of land cover orland use.(13–14) Since their introduction, NLMs haveevolved greater spatial complexity(15–18) and havebeen used to develop spatially explicit theory of howlandscape structure (and spatial pattern more gener-ally) affects a wide range of ecological phenomena,(19)
including the spread of disturbances,(20) the disper-sal of organisms,(7,9,10,21) extinction risk for species infragmented landscapes,(22) and coexistence of speciesin space.(23–25)
Complex landscape patterns can be generatedacross a gradient of habitat availability and fragmen-tation severity (Fig. 1). Because many invasive species,such as exotic plants, spread through disturbed ar-eas where native habitat has been cleared, the avail-able habitat for these species is represented by thedistribution of disturbances in space (gray cells inFig. 1). Disturbances can be simulated as a randomprocess, or as a fractal distribution in which the de-gree of spatial contagion (H) can be adjusted to pro-duce disturbances that are either aggregated (H =1.0) or dispersed across the landscape (H = 0.0). Forexample, disturbances may occur at a local scale and

Risk of Invasive Spread in Fragmented Landscapes 805
Fig. 1. Effect of landscape structure on the potential for invasive spread. Neutral landscape models, derived from percolation theory, are usedto generate complex spatial patterns representing a gradient of landscape disturbance and fragmentation (random is the most fragmented;H refers to the degree of spatial autocorrelation of the fractal landscape patterns with H = 1.0 representing the highest degree of spatialcontagion). The spread of a hypothetical species constrained to disperse through adjacent cells of disturbed habitat (gray) is highlighted inblack. The species is said to “percolate” if it is capable of spreading across the entire landscape (black cluster spans the landscape).
be distributed as many small clearings throughout thelandscape (e.g., random landscapes, Fig. 1). Alterna-tively, disturbances may be concentrated in space andoccur as a few large clearings on the landscape (e.g.,fractal, H = 1.0 landscapes, Fig. 1).
2.1. Identifying Critical Thresholdsin Invasive Spread
The probability of invasive spread is determinedby the presence of a spanning cluster of available habi-tat (disturbed areas in this case), which would permitthe species to spread (or percolate) across the entirelandscape. The critical level of disturbance at whichthis occurs depends upon the spatial pattern of thedisturbance and the dispersal abilities of the species.For example, consider a species that has localized dis-
persal and is constrained to spread through adjacentcells of disturbed habitat (i.e., cells that are connectedalong an edge). If disturbances are small and local-ized (random), such a species would not be able tospread across the landscape until >50% of the land-scape had been disturbed (invasive spread throughdisturbed habitat depicted in black in Fig. 1). Alter-natively, if disturbances are large and concentratedin space (fractal, H = 1.0), invasive spread can occurwhen as little as 30% of the landscape has been dis-turbed, owing to the greater connectivity of habitat(Fig. 1).
Invasive spread is predicted to occur as a thresh-old response to habitat destruction (Fig. 2A). Byconvention, the critical level of disturbance at whichinvasive spread occurs is taken as a ≥50% probabilityof percolation across the landscape. Thus, the critical

806 With
Fig. 2. The probability of invasive spread as a function of distur-bance in different landscapes (cf. Fig. 1) for species constrained todisperse through adjacent cells of disturbed habitat (A) and thosecapable of crossing a single cell of unsuitable habitat (i.e., specieshas gap-crossing abilities, B). Invasive spread is defined here in apercolation context, as the probability that a species can percolateacross the landscape (cf. Fig. 1).
threshold of invasive spread occurs when 57% of thelandscape has been disturbed if disturbances are smalland localized (random curve), but at only 43% whendisturbances are large or concentrated in space (H =1.0 curve, Fig. 2A). In other words, invasive spread oc-curs at a lower level of disturbance when disturbancesare large or clumped in distribution on the landscape.
If the species has better dispersal abilities and isable to cross gaps of unsuitable habitat (single cellsof nondisturbed habitat in this case), then criticalthresholds of invasive spread are shifted (Fig. 2B).Now, invasive spread is more likely to occur on land-scapes in which disturbances are small and localized(a reversal in the ordering of random and H = 1.0response curves) when only 26% of the landscape hasbeen disturbed (random curve, Fig. 2B). This is be-
cause the more dispersed pattern of disturbance facil-itates invasive spread by providing “stepping stones”to dispersal for species capable of crossing gaps(Fig. 3). In contrast, invasive spread on landscapes inwhich disturbances are concentrated in space wouldnot occur until nearly half (48%) of the landscape hadbeen disturbed (Fig. 2B). The threshold for invasivespread thus changes little in landscapes with a con-centrated pattern of disturbance (at least for thesetwo types of species), but is reduced significantly forbetter dispersers in landscapes with small, localizeddisturbances. Increasing the dispersal neighborhoodof the species would result in further shifts in the crit-ical threshold of invasion, such that spread occurs atincreasingly lower levels of habitat disturbance.(2)
Thus, based on this application of percolation the-ory, land management to reduce the risk of invasivespread for species with limited gap-crossing abilitiesshould strive to concentrate disturbances within aparticular region of the landscape (i.e., restrict habi-tat clearing to one or a few regions of the land-scape). Land management to reduce invasive spreadfor species with localized dispersal should instead cre-ate a dispersed or fragmented pattern of disturbanceacross the landscape. Because these species cannotcross gaps of unsuitable habitat (nondisturbed sites),fragmentation would serve as a barrier to control theirspread. A similar recommendation has been madepreviously to control the spread of disturbances suchas fire across landscapes.(20) The notion of creating“fire-breaks” to control invasive spread is also be-hind the practice by the U.S. Forest Service of cre-ating “barrier zones” at invasion fronts to slow thespread of gypsy moths (Lymantria disparare) throughforested landscapes of the eastern United States.(26)
Activities targeting the eradication or at least suppres-sion of gypsy moths are concentrated along these inva-sion fronts, thus creating a barrier (absence of mates)that slows their rate of spread. The efficacy of manag-ing landscapes to control invasive spread has not yetbeen demonstrated to my knowledge, but the resultsof these percolation-based studies suggest that differ-ent land-management strategies may be required tocontrol the invasive spread of species that differ indispersal ability.
2.2. When is Landscape Pattern Importantfor Predicting Invasive Spread?
The importance of landscape structure for inva-sive spread is expected to diminish as the dispersalrange of the species increases. Although a species

Risk of Invasive Spread in Fragmented Landscapes 807
Fig. 3. Effect of landscape structure on the potential for invasive spread in a species that has gap-crossing abilities (i.e., can cross a singlecell of unsuitable habitat). Note that invasive spread is possible in fragmented (random) landscapes when as little as 30% of the landscapeis disturbed (cf. Fig. 2B) because the more dispersed pattern of disturbance acts as “stepping stones” to dispersal.
may generally exhibit local dispersal, it is the infre-quent long-distance dispersal events that ultimatelygovern the rate of invasive spread.(27,28) For exam-ple, landscape pattern was unimportant for predictingthe spread of pine trees (Pinus pinaster) invading thefynbos of South Africa in a simulation model wherea small percentage of seeds (0.1%) were capable ofdispersing long distances (1–10 km) by the wind.(29)
Landscape pattern—the specific arrangement of dis-turbances in space—may in fact be important for pre-dicting the rate of invasive spread only when somethreshold of disturbance has been crossed.
Tree migration rates reflect the potential for rangeexpansion, and thus represent a form of invasion on aregional or continental scale. In one simulation study,landscape structure affected tree migration rates onlywhen suitable habitat (forest) fell below 10–25% (i.e.,75–90% of the landscape had been deforested), de-
pending upon the specific pattern of disturbance.(30)
Tree migration rates were slowest in landscapes inwhich disturbances were aggregated in space, thus cre-ating large gaps between forest patches.(30) Landscapepattern may not always be important for predictingthe risk of invasive spread, but identifying when it isand thus the extent to which landscapes can be man-aged to minimize invasive spread is an important re-search question that needs to be addressed.
2.3. Edge Effects on the Potentialfor Invasive Spread
Assessing the risk of invasive spread in landscapesalso depends on whether species are moving primar-ily within or between habitat types.(20) Thus far, it hasbeen assumed that the invasive species are movingprimarily through disturbed areas, but some species

808 With
are then able to infiltrate native habitats (moving be-tween habitat types), especially along habitat edges.For example, the invasion of dry sclerophyll bush-land in Australia by Pittosporum undulatum has beenenhanced along suburban edges because these areasattract birds such as pied currawongs (Strepera grac-ulina), which eat the fruits of P. undulatum and arethe primary dispersal agent for this plant.(31) Land-management activities that minimize habitat frag-mentation of native habitats would reduce such edgeeffects and decrease the potential for biotic exchangesbetween different habitats or land uses.
3. EFFECT OF LANDSCAPE STRUCTUREON THE INVASION PROCESS
Although percolation models are process basedin that they incorporate local dispersal neighborhoodsin assessing the risk of spread across spatially struc-tured landscapes, they do not explicitly incorporatethe effects of landscape structure on the specific pro-cesses that contribute to invasive spread, such asdispersal (successful colonization) and demography(successful population establishment). Their maincontribution has thus been to provide an assessmentof the potential for invasive spread in different land-scapes. Although the effect of landscape structure onthe process of invasive spread has not yet been ad-dressed formally,(2) results from other theoretical in-vestigations into how landscape structure affects dis-persal and demography at least provide some insightsinto how the invasion process may be affected by habi-tat fragmentation.
3.1. Assessing the Risk of Fragmentationon the Dispersal Phase of Invasive Spread
Consider a species that colonizes disturbed ar-eas but is capable of moving through other habitats.What is the probability that individuals will be suc-cessful in locating habitat suitable for colonizationin landscapes that differ in the amount and distribu-tion of disturbances? If dispersal occurs as a randomsearch (random direction and distance moved witheach step), then landscape pattern is not importantand only the amount of habitat and the dispersal abil-ity (m, the number of steps or dispersal attempts) ofthe organism affect colonization success. The proba-bility of colonization success can be defined as
Pr(success) = 1 − (1 − ε)(u + ph)m, (1)
where γ is the probability the individual does not dis-perse; u = 1 − h, where h is the amount of habitaton the landscape; and p is the proportion of suitablesites already colonized.(32) If ε is set to 0 to force dis-persal and p = 0 (all suitable sites are available forcolonization), then this expression simplifies to
Pr(success) = 1 − (u)m. (2)
Landscape structure is important for predictingcolonization success when individuals are constrainedto move through the adjacent cells and must interactwith the spatial patterning of habitat (i.e., search oc-curs at a local scale relative to landscape pattern). Theprobability of dispersal success on fractal landscapescannot be derived analytically from first principlesand may lack a closed-form solution; thus, it was nec-essary to obtain the probability of dispersal successas an empirical fit to simulation results.(9,33) Dispersalwas simulated as a modified-percolation process onfractal landscapes, in which individuals initiate dis-persal from a habitat cell (e.g., a natal territory orpropagules produced by a plant on a disturbed site)and move with random directionality through the ad-jacent cells up to m-steps in search of a suitable sitefor colonization. Colonization success on fractal land-scapes is assessed as the proportion of individuals thatlocate suitable habitat, and can be approximated bythe equation
Pr(success) = 1 − (1 − ε)[(1 − h′)mβ1 + (ph′)mβ2]
, (3)
where γ , p, and m are as in Equation (1); h′ = a +bh, where h is the abundance of habitat and a and bare fitted parameters that vary with the spatial con-tagion H of the landscape. The parameters β1 and β2
are also fitted; β1 varies with H, and β2 varies withboth H and h. The mathematical form of this functionwas selected for its relative simplicity and consistencywith the analytical form of Equation (1); this equationprovided the best fit among several other functionsgenerated.(22)
Colonization success declined precipitously be-low a threshold level of disturbance (Fig. 4).(9) In otherwords, as the availability of habitat suitable for colo-nization (disturbed areas) falls below a threshold ofabout 10%, the probability that individuals will en-counter such habitat declines rapidly (Fig. 4A). In thisscenario, colonization success is expected to be higherin landscapes in which disturbances are large or con-centrated in space (H = 1.0) because dispersers origi-nate from disturbed areas and are thus more likely toland or encounter suitable habitat when dispersal is

Risk of Invasive Spread in Fragmented Landscapes 809
Fig. 4. (A) The effect of habitat fragmentation on colonizationsuccess—the probability that a species will locate suitable habitat—as a function of landscape disturbance. The hypothetical species de-picted here is able to move through unsuitable habitat in its searchfor a suitable colonization site (i.e., disturbed areas), but is con-strained to move through the adjacent cells up to the dispersalrange of the species (m). Landscapes are as in Figs. 1 and 3 (afterReference 9). (B) Added risk of fragmentation on the dispersalphase of invasive spread. Added risk is assessed as the percent dif-ference in colonization success between landscapes (fractal, H =0.0 and fractal, H = 1.0 depicted in A).
localized, especially if the species is capable of mov-ing through the areas of unsuitable habitat. Spread isthus more likely in these landscapes than in ones inwhich disturbances are small and localized (H = 0.0)owing to higher colonization success (Fig. 4A). Again,species with long-distance dispersal may not exhibitsuch dispersal thresholds and may be more affectedby the amount of suitable habitat available for colo-nization, than the spatial arrangement of that habitaton the landscape (Equation (1)). The importance oflandscape pattern for colonization success (and thusinvasive spread) ultimately depends on how the scaleof dispersal interacts with the scale of landscape dis-turbances.
Added risk is the increase in risk that resultsfrom some impact on a population.(34) The addedrisk of fragmentation on the dispersal aspects of in-vasive spread can be calculated as the percent differ-ence in colonization success between two landscapes
(Fig. 4B). In this scenario, it is at low levels of dis-turbance that landscape pattern—the distribution ofdisturbances in space—is expected to have the great-est effect on colonization success, and by inference, oninvasive spread. Beyond this, the added risk of frag-mentation on invasive spread declines linearly, suchthat the extent of disturbance (percent of landscapedisturbed) becomes more important than the patternof disturbance.
Many models that simulate tree migration infragmented landscapes also found thresholds in mi-gration rate at low levels of available habitat (e.g.,References 30, 35–37). With and King(9) suggestedthat such thresholds in dispersal or colonization suc-cess might be more related to lacunarity thresholdsthan percolation thresholds. Lacunarity is derivedfrom fractal geometry and is a measure of the variabil-ity in gap-size distributions (interpatch distances) onthe landscape.(38) At low levels of available habitat onthe landscape (<10%), there is a dramatic increase inthe lacunarity index (i.e., a lacunarity threshold) thatcorresponds with the threshold in dispersal or col-onization success (Fig. 4A).(9) Thus, invasive spreadmight be limited more by the size and distribution ofgaps among patches of suitable habitat than by theconnectivity of suitable habitat patches (as measuredby percolation thresholds). However, this remains ahypothesis to be tested. Empirical evidence for move-ment thresholds at low levels of habitat (<20%) hasbeen found for some terrestrial insects within exper-imental landscapes inspired by NLMs (tenebrionidbeetles on random landscapes, Reference 8; early in-star crickets, Acheta domestica, on fractal landscapes,Reference 10).
3.2. Assessing the Risk of Fragmentationon the Demographic Phase of Invasive Spread
Invasive spread involves more than dispersal toand successful colonization of new sites; the speciesmust also be able to persist and achieve positive pop-ulation growth (λ > 1) in the new habitat. Most pop-ulation models ignore the effect of landscape struc-ture on demography, despite the fact that edge ef-fects (a consequence of landscape fragmentation)may alter species interactions in ways that negatively(or sometimes positively) affect demographic rates(reproductive output, survivorship). For example,With and King(6) devised a functional relationship be-tween patch structure and reproductive success forvarious neotropical migratory songbirds as part of aspatially structured demographic model developed to

810 With
Fig. 5. Functional relationship between avian reproductive successand patch geometry (assessed by an edge index derived as the nor-malized perimeter-to-area ratio of the patch) for two hypotheticalspecies that vary in edge-sensitivity. Patch geometry is influenced bylandscape structure (the amount and spatial arrangement of habi-tat). This function thus provides an explicit link between spatialpattern and demography (after Reference 6).
assess population viability in fragmented landscapes(Fig. 5). Reproductive success in a given patch, Si, canbe obtained as
Si = Smax1
1 + (ei
/kθ
) , (4)
where Smax is the maximum probability of reproduc-tive success in the absence of edge effects (normal-ized patch edge-to-area ratio → 0.0). This was set atSmax = 0.8 in this analysis (i.e., not all eggs hatch, evenin the absence of predation or brood parasitism). Theparameter ei is the normalized edge-to-area ratio ofpatch i and k is the value of ei where Si = 0.5Smax.The parameter θ determines the rate at which repro-ductive success declines as a function of increasingedge (i.e., edge effects). Some species were “edge-sensitive” and exhibited a steep decline in reproduc-tive output in small or irregularly shaped patches thatwere dominated by edge (high edge index), which isconsistent with the results of field studies document-ing a decline in nesting success in such patches ow-ing to increased nest predation or brood parasitismby brown-headed cowbirds, Moluthrus ater (e.g.,Reference 39). Although the focus here is on birdspecies that exhibit negative edge effects, some birdsare actually “edge species,” which exhibit a nestingpreference for, and thus have higher reproductive suc-cess along, habitat edges. It is not clear whether exoticbird species would necessarily exhibit such positive
edge effects, given that their breeding habitat is likelyto be disturbed areas and thus they should still per-form better in large disturbance patches (low edge in-dex) producing some degree of (negative) edge sensi-tivity. Nevertheless, such positive edge responses canbe easily incorporated as another type of functionalresponse to patch geometry (landscape structure) inthe model. The unique aspect of this model is that itexplicitly incorporates the effects of spatial pattern ondemography.
The demographic consequences of landscapestructure were assessed as the expected number offemale offspring produced per female, b, for the pop-ulation across the entire landscape as
b =∑n
i=1
∑Nij=1 Fji∑n
i=1 Ni, (5)
where Fji is the number of female offspring producedin territory j of patch i, Ni is the number of territo-rial females in patch i, and n is the total number ofpatches. The number of female fledglings (Fji) is theproduct of the number of territories in patch i (Ti),species-specific clutch size (here a uniform distribu-tion of 4–5 eggs), the probability of reproductive suc-cess for patch i (Si, Equation (4)), and the fledglingsex ratio (1:1). A simple two-stage life-table combin-ing fecundity (b) and survivorship (juvenile, s0, andadult, s) was then constructed for each species (e.g.,Fig. 5) in a given landscape (e.g., Fig. 1). From the lifetable, we calculated the finite rate of population in-crease (λ) given by the solution to the characteristicequation:(40)
λα − sλα−1 − blα = 0, (6)
for α ≥ 1 and 0 < s < 1, where lα is survivorship atthe age of first breeding, s is the annual probabilityof survivorship for breeding adults (>1 year), and bis derived from the population across the entire land-scape (Equation (5)). The population is stable whenλ = 1.0, declining when λ < 1.0, and increasing whenλ > 1.
Although the goal of this analysis was to deter-mine the potential of different landscapes to supportpersistent and increasing populations (i.e., populationsources) for birds of conservation concern, the modelstructure is general enough that the results couldalso be interpreted from the opposite perspective ofidentifying landscape configurations that would sup-port increasing populations of potentially invasivespecies, which would increase the risk of their spread.Negative ecological interactions at edges, such as

Risk of Invasive Spread in Fragmented Landscapes 811
Fig. 6. (A) Effect of habitat fragmentation on population growthrates (λ) as a function of landscape disturbance. The hypotheticalspecies depicted here has low edge sensitivity, in which reproductivesuccess is reduced only in patches dominated by edge effects (e.g.,small areas of disturbed habitat). The horizontal line is a thresholdof population establishment (λ = 1.0). Below this threshold, popu-lations are declining (λ < 1.0), populations are unlikely to becomeestablished, and the risk of spread is low. Above this threshold,populations are increasing (λ > 1.0), which increases the risk ofinvasive spread. Landscapes are as depicted in Figs. 1 and 3 (afterReference 6). (B) Added risk of fragmentation on the demographicphase of invasive spread. Added risk was assessed as the differencein population growth rates (λ) between landscapes for the speciesdepicted in (A).
increased competition, predation, or parasitism, mayaffect reproductive success even in invasive species.As an example, consider a species with low edgesensitivity, in which reproductive success is not re-duced except in fragments dominated by edge effects(e.g., small disturbed areas surrounded by habitat un-suitable for colonization). Landscapes in which dis-turbances were small and localized (random) wouldnot be able to support persistent populations of thisspecies as long as <30% of the landscape was dis-turbed (Fig. 6A). Conversely, if disturbances are evenslightly clustered in space (fractal, H = 0.0), the land-scape could support persistent and increasing pop-ulations when as little as 5% of the landscape hadbeen disturbed (Fig. 6A). These results are not gen-eral and are specific to this particular scenario (i.e., aspecies with low edge sensitivity), but demonstrate
how such spatially structured demographic modelsmight be used to assess the effect of landscape struc-ture on demography and thus on the potential for in-vasive spread.
The added risk of fragmentation on the demo-graphic aspects of invasive spread can again be as-sessed as the difference between population growthrates (λ) in different landscape scenarios (random andfractal, H = 0.0). The added risk of fragmentation oninvasive spread is greatest at low levels of disturbance(5–20%; Fig. 6B). It is in this range that the specificpattern of disturbance has the greatest effect on theability of the population to achieve positive popula-tion growth rates, and thus where there is the greatestpotential risk of spread. Once 30% of the landscapehas been disturbed, however, this species is able toachieve positive population growth regardless of thepattern of disturbance (i.e., landscape configuration).Again, this should not be viewed as a general result,as it is specific to the particulars of how the model isparameterized for this species (e.g., edge sensitivity;Equation (4), Fig. 5). Landscape structure may affectthe demographic aspects of invasive spread even athigh levels of disturbance for species with greater edgesensitivity (i.e., habitat fragmentation affects popula-tion growth rates of edge-sensitive species even whenthe landscape is mostly suitable for colonization; Ref-erence 6).
3.3. Assessing the Relative Effects of Dispersaland Demography on Invasive Spread
Invasive spread requires both successful dispersal(colonization) and positive population growth rates(establishment) in new habitats. Assessing the rel-ative contributions of dispersal versus demographyto invasive spread is complicated by the fact thatlandscape structure may affect different stages of theinvasion process in contrasting ways. For example,Bergelson et al.(41) demonstrated that the weedSenecio vulgaris was able to disperse farther whenbare-ground areas were uniformly distributed acrossthe landscape (i.e., landscape is fragmented). In con-trast, population growth rates of S. vulgaris wereenhanced when disturbed areas were aggregated inspace (i.e., not fragmented). More seedlings were ableto establish successfully when suitable sites for colo-nization were clumped because a greater concentra-tion of seeds was able to accumulate in these sites(i.e., higher propagule pressure). This illustrates atrade-off that may exist for many species in frag-mented landscapes. Dispersal may be facilitated in a

812 With
fragmented landscape because colonization sites arewell distributed across the landscape and the speciescan move farther or “percolate” across the entirelandscape (e.g., random landscapes in Fig. 3). In con-trast, the population persistence and growth rates areenhanced in landscapes with more aggregated habi-tat (e.g., fractal, H = 0.0 landscapes in Fig. 6A). Thedemographic aspects of the invasion process, particu-larly in terms of how landscape structure affects pop-ulation vital rates, have generally received less atten-tion than dispersal and are in need of further study.Land management to control invasive spread may becomplicated if landscape structure affects the disper-sal and demography of an invasive species in contrast-ing ways. Deciding which land-management scenariowill best control the invasive species in this situationwill depend on whether dispersal or demography con-tributes more to invasive spread.
4. THE EFFECT OF LANDSCAPE STRUCTUREON THE INVASIBILITY OF COMMUNITIES
The attributes of communities may influence theirsusceptibility to invasion.(5) The invasibility of com-munities thus needs to be considered when assessingthe risk of invasive spread. Although there has beenconsiderable debate as to whether species-rich com-munities are more resistant to invasion than species-poor communities (e.g., References 5, 42–46), theremay nevertheless exist a critical level of biodiver-sity(47) that contributes to system resilience and makescommunities less susceptible to local perturbations,such as species introductions. For example, criticalbiodiversity may be evident in studies that have doc-umented a threshold effect of species richness on sys-tem stability or resilience. Productivity in a grasslandsystem declined precipitously below a threshold num-ber of species (S = 9; Reference 48). Species-richcommunities maintained productivity in the face ofa severe drought, and were able to recover more fullyafter this type of disturbance than species-poor com-munities.
Recently, With and King(49) adopted a complexsystems approach(50) to assess how landscape struc-ture affected community organization and critical bio-diversity thresholds. Briefly, this approach involvesgenerating simple rules governing how species evolve(mutation events that lead to increased competitive-ness for a particular niche or a shift to a new niche),interact (competition for existing niche space), andspread across landscapes (see Reference 47 for a fulldescription of the model). These local rules of inter-
action give rise to global system properties such ascommunity structure and resilience. Although notspecifically designed to assess the invasibility of com-munities, this research may nevertheless have implica-tions for understanding the resilience of communitiesto perturbations, such as the introduction of an exoticspecies.
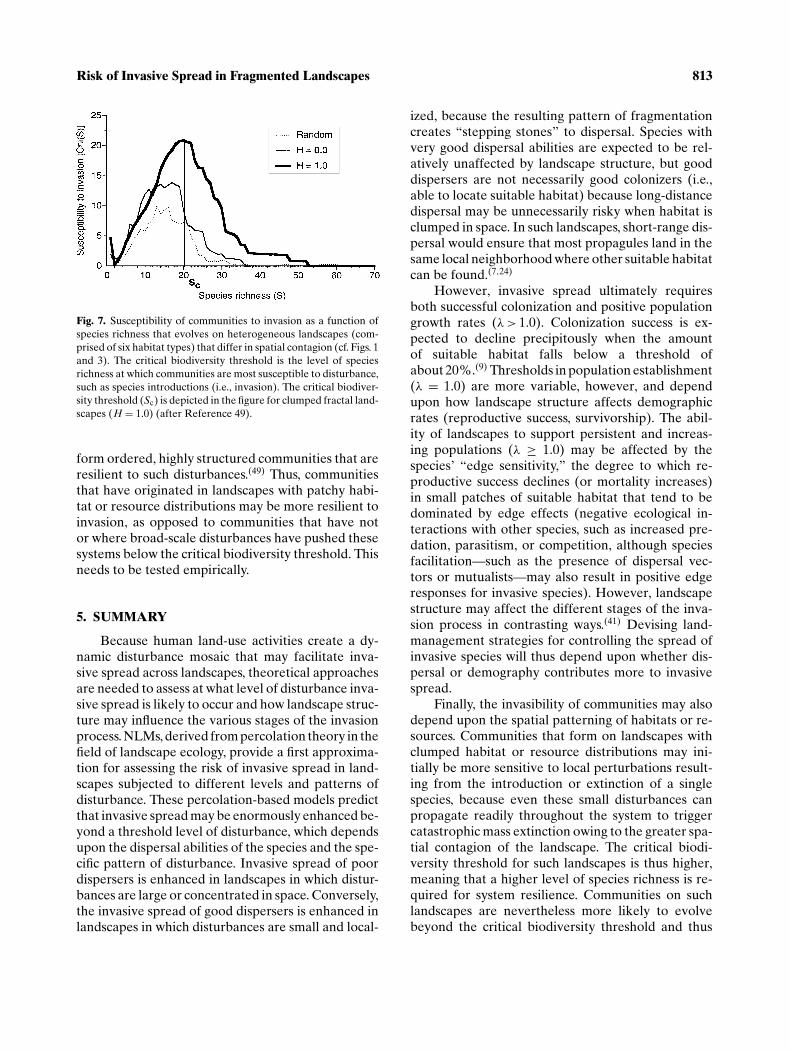
The critical biodiversity threshold is the level ofspecies richness at which the system is most suscepti-ble to small perturbations, such as the introduction orextinction of a single species, whose effects are thenable to propagate throughout the entire system andtrigger a mass extinction event.(47) Beyond this thresh-old, species spontaneously form ordered communitieswith well-defined spatial structure such that local per-turbations are no longer able to trigger system-widecatastrophes such as mass extinctions. The susceptibil-ity to extinction, χ(S), is the average extinction sizeas a function of diversity (S) and is defined as
χ(S) ≡ 1S
∫ ∞
t0E(t, S′(t))θ(S′, S) dt
{θ = 0 : S′ ≥ S
θ = 1 : S′ < Ss ′(t0) > s,
(7)
where E is the sum total of extinctions integrated overa range of species S′.(47) The level of species rich-ness (S) at which the susceptibility to extinction, χ(S),is greatest defines the critical biodiversity threshold(Sc, Fig. 7). Spatial contagion was found to enhancethe susceptibility of communities to extinction, andthus the critical biodiversity threshold increased inlandscapes with high spatial contagion (random: Sc =15; fractal, H = 1.0: Sc = 20; Reference 49). Land-scapes with clumped habitat or resource distributionsmight initially be more susceptible to invasion un-til this threshold is reached, because extinctions re-sulting from the introduction of a novel species canpropagate more readily owing to the greater spatialcontagion of habitat. A greater level of species rich-ness might therefore be required before the system isable to resist invasion.
Thus, the landscape conditions that gener-ally favor invasive spread (clumped distribution ofhabitat suitable for colonization and populationestablishment) appear to enhance the invasibility ofcommunities to invasion. Although landscapes withhigh spatial contagion may initially be more sensitiveto local perturbations (invasion, extinction) than ran-dom landscapes, they nevertheless were more likely toevolve beyond the critical biodiversity threshold and
Risk of Invasive Spread in Fragmented Landscapes 813
Fig. 7. Susceptibility of communities to invasion as a function ofspecies richness that evolves on heterogeneous landscapes (com-prised of six habitat types) that differ in spatial contagion (cf. Figs. 1and 3). The critical biodiversity threshold is the level of speciesrichness at which communities are most susceptible to disturbance,such as species introductions (i.e., invasion). The critical biodiver-sity threshold (Sc) is depicted in the figure for clumped fractal land-scapes (H = 1.0) (after Reference 49).
form ordered, highly structured communities that areresilient to such disturbances.(49) Thus, communitiesthat have originated in landscapes with patchy habi-tat or resource distributions may be more resilient toinvasion, as opposed to communities that have notor where broad-scale disturbances have pushed thesesystems below the critical biodiversity threshold. Thisneeds to be tested empirically.
5. SUMMARY
Because human land-use activities create a dy-namic disturbance mosaic that may facilitate inva-sive spread across landscapes, theoretical approachesare needed to assess at what level of disturbance inva-sive spread is likely to occur and how landscape struc-ture may influence the various stages of the invasionprocess. NLMs, derived from percolation theory in thefield of landscape ecology, provide a first approxima-tion for assessing the risk of invasive spread in land-scapes subjected to different levels and patterns ofdisturbance. These percolation-based models predictthat invasive spread may be enormously enhanced be-yond a threshold level of disturbance, which dependsupon the dispersal abilities of the species and the spe-cific pattern of disturbance. Invasive spread of poordispersers is enhanced in landscapes in which distur-bances are large or concentrated in space. Conversely,the invasive spread of good dispersers is enhanced inlandscapes in which disturbances are small and local-
ized, because the resulting pattern of fragmentationcreates “stepping stones” to dispersal. Species withvery good dispersal abilities are expected to be rel-atively unaffected by landscape structure, but gooddispersers are not necessarily good colonizers (i.e.,able to locate suitable habitat) because long-distancedispersal may be unnecessarily risky when habitat isclumped in space. In such landscapes, short-range dis-persal would ensure that most propagules land in thesame local neighborhood where other suitable habitatcan be found.(7,24)
However, invasive spread ultimately requiresboth successful colonization and positive populationgrowth rates (λ > 1.0). Colonization success is ex-pected to decline precipitously when the amountof suitable habitat falls below a threshold ofabout 20%.(9) Thresholds in population establishment(λ = 1.0) are more variable, however, and dependupon how landscape structure affects demographicrates (reproductive success, survivorship). The abil-ity of landscapes to support persistent and increas-ing populations (λ ≥ 1.0) may be affected by thespecies’ “edge sensitivity,” the degree to which re-productive success declines (or mortality increases)in small patches of suitable habitat that tend to bedominated by edge effects (negative ecological in-teractions with other species, such as increased pre-dation, parasitism, or competition, although speciesfacilitation—such as the presence of dispersal vec-tors or mutualists—may also result in positive edgeresponses for invasive species). However, landscapestructure may affect the different stages of the inva-sion process in contrasting ways.(41) Devising land-management strategies for controlling the spread ofinvasive species will thus depend upon whether dis-persal or demography contributes more to invasivespread.
Finally, the invasibility of communities may alsodepend upon the spatial patterning of habitats or re-sources. Communities that form on landscapes withclumped habitat or resource distributions may ini-tially be more sensitive to local perturbations result-ing from the introduction or extinction of a singlespecies, because even these small disturbances canpropagate readily throughout the system to triggercatastrophic mass extinction owing to the greater spa-tial contagion of the landscape. The critical biodi-versity threshold for such landscapes is thus higher,meaning that a higher level of species richness is re-quired for system resilience. Communities on suchlandscapes are nevertheless more likely to evolvebeyond the critical biodiversity threshold and thus

814 With
should be able to resist invasions unless disturbancesare widespread (rather than local) and species rich-ness is seriously eroded. Unfortunately, the natureof human disturbances on landscapes has been bothwidespread and has seriously eroded biodiversity.Theory suggests that not only should communities insuch landscapes be more susceptible to invasion, butalso that the alteration of landscape structure by hu-man land-use activities may well be facilitating thespread of invasive species. Future research shouldassess the feasibility of managing or restoring land-scapes to control the spread of invasive species and toenhance the ability of communities to resist invasion.
ACKNOWLEDGMENTS
I thank Mark C. Andersen for inviting me to con-tribute to this special issue on how new approachesfrom theoretical ecology and ecological risk assess-ment can be applied to predict the risk of invasivespread. My research on applications of NLMs to prob-lems in conservation biology has been supported bygrants from the National Science Foundation. I thankMark Andersen and three anonymous reviewers fortheir comments on the manuscript.
REFERENCES
1. Wilcove, D. S., Rothstein, D., Dubow, J., Phillips, A., & Losos,E. (1998). Assessing the relative importance of habitat de-struction, alien species, pollution, over-exploitation, and dis-ease. BioScience, 48, 607–616.
2. With, K. A. (2002). The landscape ecology of invasive spread.Conservation Biology, 16, 1192–1203.
3. Hobbs, R. J. (2000). Land-use changes and invasion. In H. A.Mooney & R. J. Hobbs (Eds.), Invasive Species in a ChangingWorld (pp. 55–64). Washington, DC: Island Press.
4. Hobbs, R. J., & Huenneke, L. F. (1992). Disturbance, diver-sity, and invasion: Implications for conservation. ConservationBiology, 6, 324–337.
5. Fox, M. D., & Fox, B. D. (1986). The susceptibility of commu-nities to invasion. In R. H. Groves & J. J. Burdon (Eds.), Ecol-ogy of Biological Invasions: An Australian Perspective (pp.97–105). Canberra, Australia: Australian Academy of Science.
6. With, K. A., & King, A. W. (2001). Analysis of landscapesources and sinks: The effect of spatial pattern on avian de-mography. Biological Conservation, 100, 75–88.
7. Lavorel, S., Gardner, R. H., & O’Neill, R. V. (1995). Disper-sal of annual plants in hierarchically structured landscapes.Landscape Ecology, 10, 277–289.
8. Wiens, J. A., Schooley, R. L., & Weeks, Jr., R. D. (1997).Patchy landscapes and animal movements: Do beetles per-colate? Oikos, 78, 257–264.
9. With, K. A., & King, A. W. (1999). Dispersal thresholds infractal landscapes: A consequence of lacunarity thresholds.Landscape Ecology, 14, 73–82.
10. With, K. A., Cadaret, S. J., & Davis, C. (1999). Movement re-sponses to patch structure in experimental fractal landscapes.Ecology, 80, 1340–1353.
11. Moody, M. E., & Mack, R. N. (1988). Controlling the spreadof plant invasions: The importance of nascent foci. Journal ofApplied Ecology, 25, 1009–1021.
12. Stauffer, D., & Aharony, A. (1992). Introduction to Percola-tion Theory, 2nd ed. London, UK: Taylor and Francis.
13. Gardner, R. H., Milne, B. T., Turner, M. G., & O’Neill, R. V.(1987). Neutral models for the analysis of broad-scale land-scape pattern. Landscape Ecology, 1, 19–28.
14. Gardner, R. H., & O’Neill, R. V. (1991). Pattern, processand predictability: The use of neutral models for landscapeanalysis. In M. G. Turner & R. H. Gardner (Eds.), Quantita-tive Methods in Landscape Ecology (pp. 289–307). New York:Springer-Verlag.
15. O’Neill, R. V., Gardner, R. H., & Turner, M. G. (1992). Ahierarchical neutral model for landscape analysis. LandscapeEcology, 7, 55–61.
16. Lavorel, S., Gardner, R. H., & O’Neill, R. V. (1993). Analysisof patterns in hierarchically structured landscapes. Oikos, 67,521–528.
17. With, K. A. (1997). The application of neutral landscape mod-els in conservation biology. Conservation Biology, 11, 1069–1080.
18. Keitt, T. (2000). A spectral analysis of neutral landscapes.Landscape Ecology, 15, 479–493.
19. With, K. A., & King, A. W. (1997). The use and misuse ofneutral landscape models in ecology. Oikos, 79, 219–229.
20. Turner, M. G., Gardner, R. H., Dale, V. H., & O’Neill, R. V.(1989). Predicting the spread of disturbance across heteroge-neous landscapes. Oikos, 55, 121–129.
21. With, K. A., & Crist, T. O. (1995). Critical thresholds inspecies’ responses to landscape structure. Ecology, 76, 2446–2459.
22. With, K. A., & King, A. W. (1999). Extinction thresholds forspecies in fractal landscapes. Conservation Biology, 13, 314–326.
23. Palmer, M. W. (1992). The coexistence of species in fractallandscapes. American Naturalist, 139, 375–397.
24. Lavorel, S., O’Neill, R. V., & Gardner, R. H. (1994). Spatio-temporal dispersal strategies and annual plant species coexis-tence in a structured landscape. Oikos, 71, 75–88.
25. Keitt, T. H. (1997). Stability and complexity on a lattice: Co-existence of species in an individual-based food web model.Ecological Modelling, 102, 243–258.
26. Sharov, A. A., & Liebhold, A. M. (1998). Model of slow-ing the spread of gypsy moth (Lepidoptera: Lymantri-idae) with a barrier zone. Ecological Applications, 8, 1170–1179.
27. Kot, M., Lewis, M. A., & van den Driesshe, P. (1996). Dispersaldata and the spread of invading organisms. Ecology, 77, 2027–2042.
28. Neubert, M. G., & Caswell, H. (2000). Demography anddispersal: Calculation and sensitivity analysis of inva-sion speed for structured populations. Ecology, 81, 1613–1628.
29. Higgins, S. I., & Richardson, D. M. (1999). Predicting plantmigration rates in a changing world: The role of long-distancedispersal. American Naturalist, 153, 464–475.
30. Collingham, Y. C., & Huntley, B. (2000). Impacts of habitatfragmentation and patch size upon migration rates. EcologicalApplications, 10, 131–144.
31. Rose, S. (1997). Influence of suburban edges on invasionof Pittosporum undulatum into the bushland of northernSydney, Australia. Australian Journal of Ecology, 22, 89–99.
32. Lande, R. (1987). Extinction thresholds in demographic mod-els of territorial populations. American Naturalist, 130, 624–635.
33. King, A. W., & With, K. A. (2002). Dispersal success onspatially structured landscapes: When do spatial pattern and

Risk of Invasive Spread in Fragmented Landscapes 815
dispersal behavior really matter? Ecological Modelling, 147,23–39.
34. Burgman, M. A., Ferson, S., & Akcakaya, H. R. (1993). RiskAssessment in Conservation Biology. London, UK: Chapmanand Hall.
35. Schwartz, M. W. (1992). Modelling effects of habitat fragmen-tation on the ability of trees to respond to climatic warming.Biodiversity and Conservation, 2, 51–61.
36. Malanson, G. P., & Cairns, D. M. (1997). Effects of dispersal,population delays, and forest fragmentation on tree migrationrates. Plant Ecology, 131, 67–79.
37. Collingham, Y. C., Hill, M. O., & Huntley, B. (1996). The mi-gration of sessile organisms: A simulation model with mea-surable parameters. Journal of Vegetation Science, 7, 831–846.
38. Plotnick, R. E., Gardner, R. H., & O’Neill, R. V. (1993). La-cunarity indices as measures of landscape texture. LandscapeEcology, 8, 201–211.
39. Robinson, S. K., Thompson III, F. R., Donovan, T. M.,Whitehead, D. R., & Faaborg, J. (1995). Regional forest frag-mentation and the nesting success of migratory birds. Science,267, 1987–1990.
40. Lande, R. (1988). Demographic models of the Northern Spot-ted Owl (Strix occidentalis). Oecologia, 75, 601–607.
41. Bergelson, J., Newman, J. A., & Floresroux, E. M. (1993).Rates of weed spread in spatially heterogeneous environ-ments. Ecology, 74, 999–1011.
42. Elton, C. S. (1958). The Ecology of Invasions by Animals andPlants. London, UK: Methuen.
43. Burke, M. J. W., & Grime, J. P. (1996). Experimental study ofplant community invasibility. Ecology, 77, 776–790.
44. Tilman, D. (1997). Community invasibility, recruitment limi-tation, and grassland biodiversity. Ecology, 78, 81–92.
45. Symstad, A. J. (2000). A test of the effects of functional grouprichness and composition on grassland invasibility. Ecology,81, 99–109.
46. Dukes, J. S. (2001). Biodiversity and invasibility in grasslandmicrocosms. Oecologia, 126, 563–568.
47. Kaufman, J. H., Brodbeck, D., & Melroy, O. R. (1998). Criticalbiodiversity. Conservation Biology, 12, 521–532.
48. Tilman, D., &. Downing, J. A. (1994). Biodiversity and stabilityin grasslands. Nature, 367, 363–365.
49. With, K. A., & King, A. W. (in press). The effect of landscapestructure on community self-organization and critical biodi-versity. Ecological Modelling.
50. Levin, S. A. (1998). Ecosystems and the biosphere as complexadaptive systems. Ecosystems, 1, 431–436.