atp hydrolysis-dependent conformational changes in … we determined that atp hydrolysis-dependent...
TRANSCRIPT

126 Journal of Lipid Research Volume 53, 2012
Copyright © 2012 by the American Society for Biochemistry and Molecular Biology, Inc.
This article is available online at http://www.jlr.org
regulated by various mechanisms ( 1, 2 ). Mutations in the ATP binding cassette protein A1 ( ABCA1 ) gene lead to Tang-ier disease, which is characterized by an accumulation of excess cellular cholesterol and low plasma high-density lipoprotein (HDL) ( 3–5 ). The link between the ABCA1 gene and Tangier disease indicates that ABCA1 plays a piv-otal role in cholesterol homeostasis by generating HDL, the only pathway that eliminates excess cholesterol from nonhepatic cells. ABCA1 mediates apolipoprotein A-I (apoA-I) binding to the cell surface and the loading of cel-lular cholesterol and phospholipids onto apoA-I to form pre- � HDL ( 6–9 ). Recently, it was reported that the capac-ity of macrophages to mediate cholesterol effl ux to HDL is strongly and inversely associated with both carotid intima-media thickness and the likelihood of angiographic coro-nary artery disease, independent of HDL cholesterol levels ( 10 ). However, despite the physiological importance of this pathway, the mechanism by which ABCA1 mediates cholesterol effl ux remains unclear ( 11 ).
ABCA1 has two intracellular nucleotide-binding domains (NBD) ( Fig. 1 ), whose amino acid sequences are highly conserved among ABC proteins. Chambenoit et al. reported that substituting the Walker A lysine residue in either NBD with methionine (K939M or K1952M) abolishes the ability of apoA-I to bind to ABCA1-expressing cells and inhibits cholesterol secretion to apoA-I ( 12 ). Because these lysine residues are essential for ATPase activity ( 13 ) and thus for the transport activity of ABC proteins, it was predicted that
Abstract ATP-binding cassette protein A1 (ABCA1) plays a major role in cholesterol homeostasis and high-density lipo-protein (HDL) metabolism. Although it is predicted that apolipoprotein A-I (apoA-I) directly binds to ABCA1, the physiological importance of this interaction is still contro-versial and the conformation required for apoA-I binding is unclear. In this study, the role of the two nucleotide-binding domains (NBD) of ABCA1 in apoA-I binding was determined by inserting a TEV protease recognition sequence in the linker region of ABCA1. Analyses of ATP binding and occlu-sion to wild-type ABCA1 and various NBD mutants revealed that ATP binds equally to both NBDs and is hydrolyzed at both NBDs. The interaction with apoA-I and the apoA-I-dependent cholesterol effl ux required not only ATP binding but also hydrolysis in both NBDs. NBD mutations and cellular ATP depletion decreased the accessibility of antibodies to a hemagglutinin (HA) epitope that was inserted at position 443 in the extracellular domain (ECD), suggesting that the conformation of ECDs is altered by ATP hydrolysis at both NBDs. These results suggest that ATP hydrolysis at both NBDs induces conformational changes in the ECDs, which are associated with apoA-I binding and cholesterol effl ux. — Nagao, K., K. Takahashi, Y. Azuma, M. Takada, Y. Kimura, M. Matsuo, N. Kioka, and K. Ueda. ATP hydrolysis-dependent conformational changes in the extracellular domain of ABCA1 are associated with apoA-I binding. J. Lipid Res. 2012. 53: 126–136.
Supplementary key words: ATP binding cassette protein A1 • apolipo-proteins • cholesterol effl ux • HDL • transport
Cholesterol is an essential component of cellular mem-branes, but excess cholesterol accumulation is harmful to cells. Therefore, cellular cholesterol levels are extensively
This work was supported by a grant-in-aid for scientifi c research (S) from the Ministry of Education, Culture, Sports, Science and Technology of Japan; by the Program for Promotion of Basic and Applied Research for Innovations in Bio-oriented Industry (BRAIN) of Japan; by a grant from the Takeda Scientifi c Foundation; and by the World Premier International Research Center Initiative (WPI) of the Ministry of Education, Culture, Sports, Science and Technology of Japan.
Manuscript received 25 August 2011 and in revised form 23 October 2011.
Published, JLR Papers in Press, October 25, 2011 DOI 10.1194/jlr.M019976
ATP hydrolysis-dependent conformational changes in the extracellular domain of ABCA1 are associated with apoA-I binding
Kohjiro Nagao , * Kei Takahashi , †,1 Yuya Azuma , †,2 Mie Takada , † Yasuhisa Kimura , † Michinori Matsuo , † Noriyuki Kioka , † and Kazumitsu Ueda 3, * ,†
† Laboratory of Cellular Biochemistry, Division of Applied Life Sciences, Graduate School of Agriculture and * Institute for Integrated Cell-Material Sciences (iCeMS), Kyoto University , Kyoto 606-8502, Japan
Abbreviations: ECD, extracellular domain; GFP, green fl uorescent protein; HA, hemagglutinin; HEK, human embryonic kidney; NBD, nucle-otide-binding domain; PC, phosphatidylcholine; sulfo-NHS-biotin, sulfo- N -hydroxysuccinimidobiotin; TEV, tobacco etch virus; TM, transmembrane helix.
1 Present address of K. Takahashi: National Research Institute of Brewing, Higashi-Hiroshima 739-0046, Japan
2 Present address of Y. Azuma: Japan Tobacco Inc., Central Pharma-ceutical Research Institute, Osaka 569-1125, Japan
3 To whom correspondence should be addressed. e-mail: [email protected]
The online version of this article (available at http://www.jlr.org) contains supplementary data in the form of four fi gures.
by guest, on June 11, 2018w
ww
.jlr.orgD
ownloaded from
.html http://www.jlr.org/content/suppl/2011/10/25/jlr.M019976.DC1Supplemental Material can be found at:

Conformational changes of ABCA1 for apoA-I binding 127
and Alexa 546-conjugated apoA-I were prepared as previously re-ported ( 24 ). The remaining chemicals were purchased from Sigma, Amersham Biosciences, Wako Pure Chemical Industries, and Nacalai Tesque.
Cell culture HEK293 cells were grown in a humidifi ed incubator (5% CO 2 )
at 37°C in Dulbecco’s modifi ed Eagle’s medium (DMEM) supple-mented with 10% heat-inactivated fetal bovine serum (FBS).
Plasmids The expression vectors for wild-type ABCA1, ABCA1-K939M,
ABCA1-K1952M, and ABCA1-K939M,K1952M that were tagged with GFP at the C terminus were generated as previously de-scribed ( 7, 13 ). The infl uenza virus hemagglutinin (HA) epitope sequence (coding YPYDVPDYA) was introduced between G207 and D208 as previously reported ( 25 ). To introduce an HA tag or a Flag tag at several sites, a MulI site (acgcgt) was introduced just after K136, N349, R443, K568, G1421, K1490, D1567, and E1622 of ABCA1-GFP using a QuickChange mutagenesis kit (Strata-gene). Oligos containing an HA tag, 5 ′ -cgcgttacccatacgatgttcca-gattacgcca-3 ′ and 5 ′ -cgcgtggcgtaatctggaacatcgtatgggtaa-3 ′ , or a Flag tag, 5 ′ -cgcgtgactacaaagacgatgacgacaaga-3 ′ and 5 ′ -cgcgtctt-gtcgtcatcgtctttgtagtca-3 ′ were introduced into the MulI site. The TEV protease recognition sequence (coding ENLYFQG) was in-troduced between R1272 and R1273 of ABCA1-GFP by PCR using primers 5 ′ -agacgaaacagggaaaacctgtacttccagggtcgggccttcgg-3 ′ and 5 ′ -ccgaaggcccgaccctggaagtacaggttttccctgtttcgtct-3 ′ . The mutated DNA was confi rmed by sequencing.
Transfection and generation of stable ABCA1 transformants
HEK293 cells were transfected with each expression vector using Lipofectamine and Plus reagent (Invitrogen) according to the manufacturer’s instructions. To establish stable transfor-mants, the cells were selected with G418, and single colonies were isolated.
Western blotting Cells were washed with PBS and lysed in lysis buffer A (20 mM
Tris-Cl, pH 7.5, 1 mM EDTA, 10% glycerol, and 1% Triton X-100) containing the following protease inhibitors: 100 � g/ml (p-amidinophenyl)methanesulfonyl fl uoride, 10 � g/ml leupep-tin, and 2 � g/ml aprotinin. The samples were electrophoresed on SDS-polyacrylamide gels, blotted, and probed with the indi-cated antibodies.
Cellular lipid release assay Cells were subcultured in poly-L-lysine-coated 24-well plates at
a density of 3 × 10 5 cells in DMEM containing 10% FBS. After 24 h incubation, the cells were washed twice with DMEM, and then incubated in DMEM containing 0.02% BSA and 5 � g/ml recom-binant apoA-I. After 24 h incubation, the cholesterol content in the medium was determined using a colorimetric enzyme assay or a fl uorescence enzyme assay ( 26, 27 ).
Biotinylation of cell surface proteins Cell monolayers were kept on ice for 10 min. The cells were
washed with ice-cold PBS+ (phosphate-buffered saline contain-ing 0.1 mg/ml CaCl 2 and 0.1 mg/ml MgCl 2 6H 2 O) and incubated with 0.5 mg/ml sulfo-NHS-biotin solubilized in PBS+ for 30 min on ice in the dark. The cells were washed with TBS (20 mM Tris-Cl, pH 7.5, 150 mM NaCl, 0.1 mg/ml CaCl 2 and 0.1 mg/ml MgCl 2 6H 2 O) to remove unbound sulfo-NHS-biotin and lysed in lysis buffer B (20 mM Tris-Cl, pH 7.5, 1% Triton X-100, 0.1% SDS, and 1%
ABCA1 induces a local and transient modifi cation in the spatial arrangement of lipid species on the outer mem-brane leafl et and that this membrane modifi cation gener-ates apoA-I-docking sites on the cell surface ( 8, 14–16 ).
ABCA1 has two large extracellular domains (ECD) after transmembrane helix 1 (TM1) and TM7 ( Fig. 1 ). Two in-tramolecular disulfi de bonds are formed between the two ECDs of ABCA1, and these disulfi de bonds are necessary for apoA-I binding and HDL formation ( 17 ). Several groups have used cross-linking experiments to show that apoA-I directly binds to ABCA1 ( 6, 8, 18–21 ). Because a 3-Å cross-linker can cross-link apoA-I with ABCA1 ( 19 ) and the K939M mutant cannot bind or be cross-linked to apoA-I ( 22 ), it is likely that apoA-I interacts with specifi c confor-mations of the ECDs that are linked by the two disulfi de bonds and formed in an ATP-dependent manner. How-ever, the importance of a direct apoA-I-ABCA1 interaction in HDL formation is still controversial, and it is unclear what conformation of ABCA1 mediates apoA-I binding and how this specifi c conformation is formed. In this study, we analyzed ATP binding and hydrolysis for each NBD of ABCA1 and examined the contribution of the NBDs to apoA-I binding and HDL formation. On the basis of these results, we determined that ATP hydrolysis-dependent con-formational changes in the ECDs of ABCA1 are associated with apoA-I binding and cholesterol effl ux by ABCA1.
MATERIALS AND METHODS
Materials The mouse anti-ABCA1 monoclonal antibody KM3110 was
generated against the 20 C-terminal amino acids of ABCA1 ( 23 ). The rat anti-ABCA1 monoclonal antibody KM3073 was generated against ECD1 of ABCA1 ( 23 ). An anti-ABCA1 NBD2 rabbit poly-clonal antibody was generated against the purifi ed NBD2 of ABCA1 ( 13 ). Anti-GFP and anti-HA (F-7) antibodies were purchased from Santa Cruz Biotechnology. Anti-Flag rabbit polyclonal anti-body was purchased from Sigma. Tobacco etch virus (TEV)-derived ProTEV protease was obtained from Promega. Recombinant apoA-I
Fig. 1. Secondary structure of ABCA1 . The HA tag-insertion sites and Walker A lysine mutations are indicated by open circles and open squares, respectively. The TEV protease recognition sequence was introduced between R1272 and R1273, as indicated by the fi lled triangle. The two cysteine residues in ECD1 and three cysteine residues in ECD2 involved in S-S bond formation between the ECDs are indicated by fi lled circles.
by guest, on June 11, 2018w
ww
.jlr.orgD
ownloaded from
.html http://www.jlr.org/content/suppl/2011/10/25/jlr.M019976.DC1Supplemental Material can be found at:
128 Journal of Lipid Research Volume 53, 2012
samples. This washing procedure was repeated. The pellets were resuspended in lysis buffer B (20 mM Tris-Cl, pH 7.5, 1% Triton X-100, 0.1% SDS, and 1% sodium deoxycholate), and ABCA1 was immunoprecipitated with KM3110. The samples were electro-phoresed on 7% SDS-polyacrylamide gels and then analyzed by autoradiography. The same membranes were analyzed by Western blotting with the indicated antibodies.
Nucleotide trapping Membrane fractions were prepared from HEK293 cells that sta-
bly or transiently expressed ABCA1-GFP. The membrane fractions were incubated with 5 � M [ � 32 P]8N 3 ATP or [ � 32 P]8N 3 ATP, 3 mM MgCl 2 , 2 mM ouabain, 0.1 mM EGTA, and 40 mM Tris-Cl, pH 7.5 in a total volume of 10 � l at 37°C. The specifi c activity of [ � 32 P]8N 3 ATP was 1.6-fold higher than that of [ � 32 P]8N 3 ATP. The reactions were stopped by adding 400 � l of ice-cold 40 mM Tris-Cl buffer containing 0.1 mM EGTA and 1 mM MgCl 2 . Supernatants containing unbound ATP were removed from the membrane pel-lets after centrifugation (15,000 rpm, 5 min, 2°C), and the proce-dure was repeated. The pellets were resuspended in 8 � l of TE buffer containing 1 mM MgCl 2 and irradiated for 5 min (254 nm, 8.2 mW/cm 2 ) on ice. The samples were electrophoresed on 7% SDS-polyacrylamide gels and then analyzed by autoradiography. The same membranes were analyzed by Western blotting using the anti-ABCA1 antibody KM3073 or an anti-GFP antibody.
TEV protease digestion Immunoprecipitated ABCA1 was digested with TEV protease
(0.2 units/ � l) at 30°C for 30 min according to the manufactur-er’s instructions.
sodium deoxycholate) containing protease inhibitors. Immobi-lized monomeric avidin gel (Pierce) was added to the cell lysates to precipitate the biotinylated proteins, which were electrophore-sed on a 7% SDS-polyacrylamide gel and immunodetected.
ApoA-I binding Cells grown on collagen-coated coverslips were incubated with
DMEM containing 0.02% BSA and Alexa 546-conjugated apoA-I (5 � g/ml) for 15 min at 37°C. The cells were then washed, fi xed with 4% paraformaldehyde at room temperature for 30 min, and observed with a confocal microscope (LSM 510; Carl Zeiss).
Anti-HA or anti-Flag antibody staining Cells grown on collagen-coated coverslips were incubated with
anti-HA antibody (1 � g/ml) or anti-Flag antibody (1:1000) in DMEM containing 0.02% BSA for 15 min at 37°C. The cells were then washed and fi xed with 4% paraformaldehyde at room tem-perature for 30 min. After blocking with PBS+ containing 1% BSA, the cells were incubated with an Alexa546-conjugated sec-ondary antibody and observed with a confocal microscope.
Cellular ATP depletion Cellular ATP was depleted by incubating with 10 mM sodium
azide and 10 mM 2-deoxy D-glucose in glucose-free DMEM con-taining 0.02% BSA for 10 min.
ATP binding Membrane fractions were prepared from HEK293 cells that sta-
bly or transiently expressed ABCA1-GFP. Membranes were incu-bated on ice with 20 � M [ � 32 P]8N 3 ATP, 3 mM MgCl 2 , 2 mM ouabain, 0.1 mM EGTA, and 40 mM Tris-Cl (pH 7.5) in a total volume of 10 � l for 10 min and then irradiated for 5 min (254 nm, 8.2 mW/cm 2 ) on ice. Unbound ATP was removed by adding 400 � l ice-cold 40 mM Tris-Cl buffer containing 0.1 mM EGTA and 1 mM MgCl 2 and then centrifuging (15,000 rpm, 5 min, 2°C) the
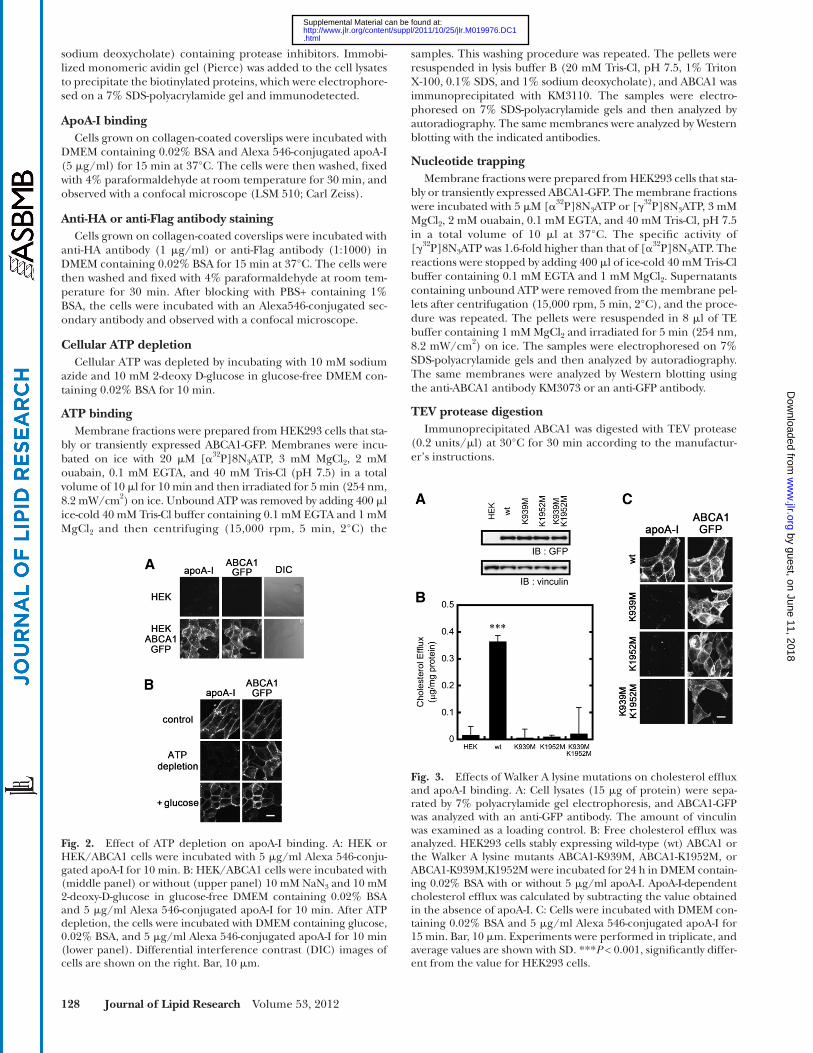
Fig. 2. Effect of ATP depletion on apoA-I binding. A: HEK or HEK/ABCA1 cells were incubated with 5 � g/ml Alexa 546-conju-gated apoA-I for 10 min. B: HEK/ABCA1 cells were incubated with (middle panel) or without (upper panel) 10 mM NaN 3 and 10 mM 2-deoxy-D-glucose in glucose-free DMEM containing 0.02% BSA and 5 � g/ml Alexa 546-conjugated apoA-I for 10 min. After ATP depletion, the cells were incubated with DMEM containing glucose, 0.02% BSA, and 5 � g/ml Alexa 546-conjugated apoA-I for 10 min (lower panel). Differential interference contrast (DIC) images of cells are shown on the right. Bar, 10 � m.
Fig. 3. Effects of Walker A lysine mutations on cholesterol effl ux and apoA-I binding. A: Cell lysates (15 � g of protein) were sepa-rated by 7% polyacrylamide gel electrophoresis, and ABCA1-GFP was analyzed with an anti-GFP antibody. The amount of vinculin was examined as a loading control. B: Free cholesterol effl ux was analyzed. HEK293 cells stably expressing wild-type (wt) ABCA1 or the Walker A lysine mutants ABCA1-K939M, ABCA1-K1952M, or ABCA1-K939M,K1952M were incubated for 24 h in DMEM contain-ing 0.02% BSA with or without 5 � g/ml apoA-I. ApoA-I-dependent cholesterol effl ux was calculated by subtracting the value obtained in the absence of apoA-I. C: Cells were incubated with DMEM con-taining 0.02% BSA and 5 � g/ml Alexa 546-conjugated apoA-I for 15 min. Bar, 10 � m. Experiments were performed in triplicate, and average values are shown with SD. *** P < 0.001, signifi cantly differ-ent from the value for HEK293 cells.
by guest, on June 11, 2018w
ww
.jlr.orgD
ownloaded from
.html http://www.jlr.org/content/suppl/2011/10/25/jlr.M019976.DC1Supplemental Material can be found at:
Conformational changes of ABCA1 for apoA-I binding 129
ATP were examined. When Alexa 546-labeled apoA-I was added to the medium, apoA-I bound to ABCA1-express-ing HEK293 cells within 10 min but not to the parental HEK293 cells, as previously reported ( 24, 28 ) ( Fig. 2A ). When the intracellular ATP levels were reduced by incu-bating the cells with sodium azide and 2-deoxy-D-glucose for 10 min, apoA-I binding was abolished, although ABCA1 expression on the plasma membrane was not af-fected ( Fig. 2B ). Incubating the cells in glucose-contain-ing medium for 10 min after ATP depletion restored apoA-I binding to levels comparable to before ATP deple-tion ( Fig. 2B ). These results suggest that apoA-I binding
Statistical analysis Values are presented as the means ± SD. The statistical signifi -
cance of differences between mean values was analyzed using the nonpaired t -test. Multiple comparisons were performed using Dunnett’s test following ANOVA. A P value < 0.05 was considered statistically signifi cant.
RESULTS
ApoA-I binding to ABCA1 is ATP-dependent To characterize the interaction between apoA-I and
ABCA1-expressing cells, the effects of depleting cellular
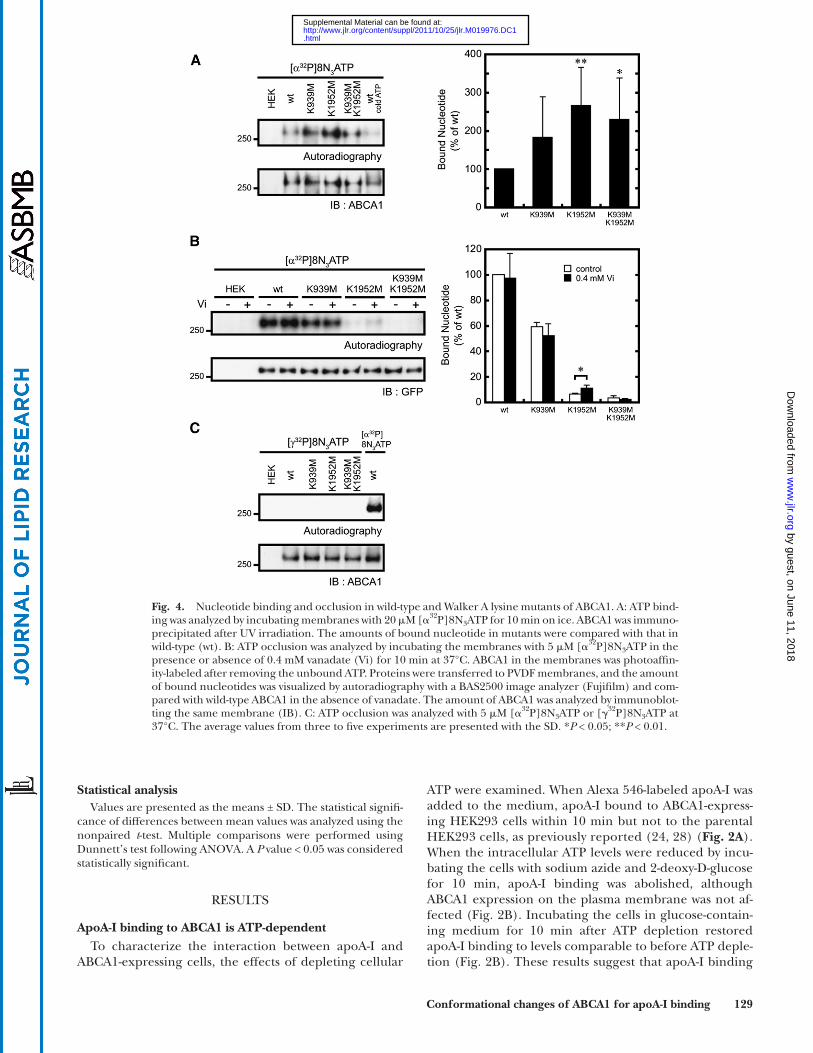
Fig. 4. Nucleotide binding and occlusion in wild-type and Walker A lysine mutants of ABCA1. A: ATP bind-ing was analyzed by incubating membranes with 20 � M [ � 32 P]8N 3 ATP for 10 min on ice. ABCA1 was immuno-precipitated after UV irradiation. The amounts of bound nucleotide in mutants were compared with that in wild-type (wt). B: ATP occlusion was analyzed by incubating the membranes with 5 � M [ � 32 P]8N 3 ATP in the presence or absence of 0.4 mM vanadate (Vi) for 10 min at 37°C. ABCA1 in the membranes was photoaffi n-ity-labeled after removing the unbound ATP. Proteins were transferred to PVDF membranes, and the amount of bound nucleotides was visualized by autoradiography with a BAS2500 image analyzer (Fujifi lm) and com-pared with wild-type ABCA1 in the absence of vanadate. The amount of ABCA1 was analyzed by immunoblot-ting the same membrane (IB). C: ATP occlusion was analyzed with 5 � M [ � 32 P]8N 3 ATP or [ � 32 P]8N 3 ATP at 37°C. The average values from three to fi ve experiments are presented with the SD. * P < 0.05; ** P < 0.01.
by guest, on June 11, 2018w
ww
.jlr.orgD
ownloaded from
.html http://www.jlr.org/content/suppl/2011/10/25/jlr.M019976.DC1Supplemental Material can be found at:
130 Journal of Lipid Research Volume 53, 2012
A Walker A lysine mutation abolishes ATP hydrolysis but not ATP binding
It has been reported that the two NBDs in some ABC proteins do not have equivalent roles in the ATP hydroly-sis cycle and substrate transport ( 29–32 ). To analyze how the two NBDs of ABCA1 interact with ATP and how they are involved in the functions of ABCA1, the effects of Walker A lysine mutations in each NBD on ATP binding were examined ( Fig. 4A ). Membranes prepared from cells expressing the wild-type or mutant ABCA1 were incubated with 20 � M [ � 32 P]8N 3 ATP for 10 min on ice and then UV irradiated. Photoaffi nity-labeled ABCA1 was immunopre-cipitated from wild-type ABCA1-expressing cells but not from the parental HEK293 cells. This labeling was inhib-ited by 100-fold excess of nonlabeled ATP, indicating that the photoaffi nity labeling was specifi c. A Walker A lysine mutation did not abolish ATP binding to ABCA1, suggest-ing that the Walker A lysine residues are not essential for ATP binding to ABCA1.
Next, nucleotide binding under ATP-hydrolysis condi-tions was analyzed. ABCA1 was strongly photoaffi nity-labeled as previously reported ( 7 ). These results indicate that the nucleotide was occluded in the NBD of ABCA1 at 37°C when membranes expressing wild-type ABCA1 were
is ABCA1- and ATP-dependent. The apoA-I on the cell surface may include both the direct binding to ABCA1 and the secondary association of lipidated apoA-I to the cell surface.
Two intact NBDs are required for apoA-I binding and cholesterol effl ux
The contribution of the two NBDs of ABCA1 to apoA-I binding and cholesterol effl ux by ABCA1 was analyzed. Wild-type ABCA1 mediated apoA-I-dependent cholesterol effl ux. However, cholesterol effl ux was impaired in cells expressing ABCA1-K939M,K1952M, a mutant in which the Walker A lysines in both NBDs are replaced with methi-onines, or the single Walker A lysine mutants ABCA1-K939M or ABCA1-K1952M, in which the Walker A lysine in NBD1 or NBD2 is replaced with methionine ( Fig. 3B ). Furthermore, these mutations did not signifi cantly affect the expression levels ( Fig. 3A ), and at least a fraction of the mutants were expressed on the plasma membrane ( Fig. 3C ). ApoA-I binding was also impaired when either Walker A lysine was replaced with methionine ( Fig. 3C ). These results suggest that the two intact NBDs are required for apoA-I binding and apoA-I-dependent cholesterol effl ux by ABCA1.
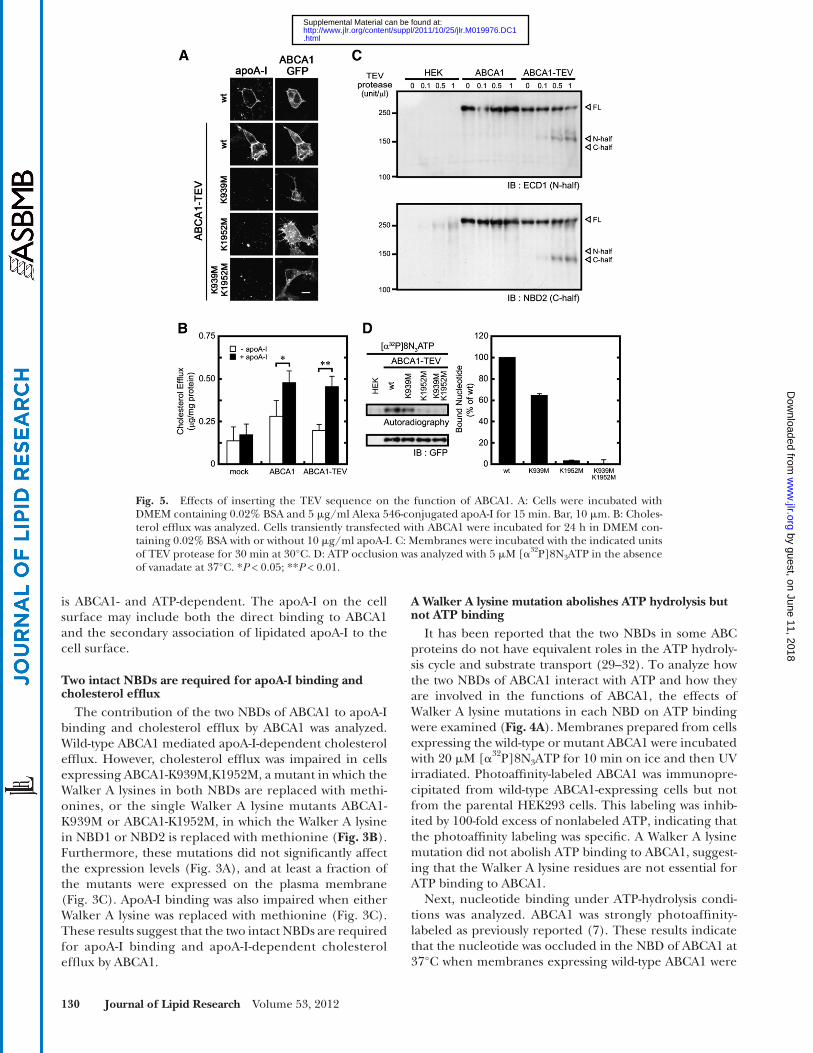
Fig. 5. Effects of inserting the TEV sequence on the function of ABCA1. A: Cells were incubated with DMEM containing 0.02% BSA and 5 � g/ml Alexa 546-conjugated apoA-I for 15 min. Bar, 10 � m. B: Choles-terol effl ux was analyzed. Cells transiently transfected with ABCA1 were incubated for 24 h in DMEM con-taining 0.02% BSA with or without 10 � g/ml apoA-I. C: Membranes were incubated with the indicated units of TEV protease for 30 min at 30°C. D: ATP occlusion was analyzed with 5 � M [ � 32 P]8N 3 ATP in the absence of vanadate at 37°C. * P < 0.05; ** P < 0.01.
by guest, on June 11, 2018w
ww
.jlr.orgD
ownloaded from
.html http://www.jlr.org/content/suppl/2011/10/25/jlr.M019976.DC1Supplemental Material can be found at:

Conformational changes of ABCA1 for apoA-I binding 131
was after R1272 and R1273 (supplementary Fig. I). To cleave ABCA1 specifi cally at this site, a TEV protease rec-ognition sequence (ENLYFQG; TEV protease cleaves be-tween Q and G) was introduced between R1272 and R1273 ( Fig. 1 and supplementary Fig. I). Insertion of the TEV sequence did not affect the cellular localization of ABCA1 or the ability of ABCA1 to bind apoA-I and mediate apoA-I-dependent cholesterol effl ux ( Fig. 5A , B ). TEV protease-mediated digestion of ABCA1-TEV-GFP yielded a 160 kDa N-terminal fragment and 140 kDa C-terminal fragment ( Fig. 5C ). ABCA1-GFP, which does not contain an inserted TEV sequence, was not digested by the TEV protease ( Fig. 5C ), indicating that the digestion by TEV protease is specifi c and that the two NBDs of ABCA1 can be separated by TEV protease digestion.
First, ATP binding to each NBD was analyzed ( Fig. 6A ). ATP bound equally to both NBDs in ABCA1 as previously reported ( 32 ). Walker A lysine mutations in both NBDs did not abolish ATP binding to each NBD. We consistently observed stronger ATP binding to NBD2 in ABCA1-K939M,K1952M-TEV-GFP, although the reason for this higher-affi nity interaction is unknown. Next, nucleotide binding to each NBD under ATP-hydrolysis conditions was analyzed. The full-length wild-type and mutant forms of ABCA1 harboring the TEV sequence exhibited a nucle-otide occlusion pattern ( Fig. 5D ) that was similar to that of the TEV-less ABCA1 ( Fig. 4B ), confi rming that insertion of the TEV sequence does not affect the nucleotide-bind-ing properties of ABCA1. When wild-type ABCA1 was incu-bated with [ � 32 P]8N 3 ATP and cleaved by the TEV protease after UV irradiation, strong nucleotide occlusion was ob-served in NBD2 (C-half) and weak occlusion was observed in NBD1 (N-half) ( Fig. 6B ). However, there was no photo-affi nity-labeled band when [ � 32 P]8N 3 ATP was used. The K939M mutation reduced the photoaffi nity labeling of
incubated with 5 � M [ � 32 P]8N 3 ATP for 10 min at 37°C and then UV irradiated on ice after the free nucleotides were removed ( Fig. 4B ). The addition of vanadate did not affect the photoaffi nity labeling of ABCA1 as previously reported ( 7 ). The K939M mutation decreased the photoaffi nity label-ing by approximately 40%, and vanadate did not affect the labeling. On the other hand, the K1952M mutation drasti-cally decreased the photoaffi nity labeling by approximately 95%. Interestingly, the addition of vanadate signifi cantly (1.7-fold) enhanced the photoaffi nity labeling of ABCA1-K1952M, suggesting that vanadate stabilizes the nucleotide at NBD1 after ATP hydrolysis. There was no photoaffi nity-labeled band with ABCA1-K939M,K1952M.
When [ � 32 P]8N 3 ATP was used for the photoaffi nity-labeling experiments under ATP-hydrolysis conditions, no photoaffi nity-labeled band was observed with the wild-type or any of the ABCA1 mutants ( Fig. 4C ), although ABCA1 was strongly labeled when [ � 32 P]8N 3 ATP was used as a control. These results suggest that the occluded nucleo-tide in the NBD of wild-type ABCA1, ABCA1-K939M, and ABCA1-K1952M is not ATP but ADP, that both NBDs oc-clude ADP after ATP hydrolysis, and that the features of the two NBDs of ABCA1 are quite different.
ATP hydrolysis occurs in both NBDs of ABCA1 To analyze the interaction of each NBD with nucleotides
in detail, we fi rst performed a limited digestion with trypsin to separate the two NBDs. However, limited trypsin diges-tion of ABCA1 occurs at two sites: one site is between TM6 and NBD1 (site A), and the other is in the linker region between the two halves of ABCA1 (site B) ( 13 ). Thus, it was diffi cult to quantitatively analyze ATP binding to each NBD. Therefore, we developed a new strategy to separate the two NBDs of ABCA1. First, Edman sequencing showed that the trypsin cleavage site in the linker region (B site)
Fig. 6. Nucleotide binding and occlusion in each NBD of ABCA1. A: Membranes were incubated with 20 � M [ � 32 P]8N 3 ATP for 10 min on ice and UV irra-diated. After immunoprecipitating with KM3110, ABCA1 was treated with or without 0.2 units/ � l TEV protease for 30 min at 30°C. B: Membranes were in-cubated with 5 � M [ � 32 P]8N 3 ATP or [ � 32 P]8N 3 ATP for 10 min at 37°C. After immunoprecipitating with KM3110, ABCA1 was treated with 0.2 units/ � l TEV protease for 30 min at 30°C. The proteins were trans-ferred to PVDF membranes and analyzed by autora-diography and immunoblotting (IB). Experiments were performed in duplicate and yielded similar results.
by guest, on June 11, 2018w
ww
.jlr.orgD
ownloaded from
.html http://www.jlr.org/content/suppl/2011/10/25/jlr.M019976.DC1Supplemental Material can be found at:
132 Journal of Lipid Research Volume 53, 2012
To examine the accessibility of the anti-HA antibody, HEK293 cells expressing the HA-insertion mutants were incubated with the anti-HA antibody at 37°C for 15 min in DMEM containing 0.02% BSA. Then cells were fi xed and
NBD2 with [ � 32 P]8N 3 ATP, apparently consistent with the decrease in the photoaffi nity labeling of the full-length K939M mutant, although immunoprecipitation and sub-sequent protease digestion make it diffi cult to quantita-tively compare the mutants ( Fig. 6B ). Nevertheless, there was no signal for the K939M mutant with [ � 32 P]8N 3 ATP. In addition, there were no detectable nucleotide occlu-sions at either NBD in the K1952M mutant. These results suggest that ATP is hydrolyzed at both NBDs of ABCA1 and that ADP is occluded in both NBDs after ATP hydroly-sis. Furthermore, a Walker A lysine mutation has a greater impact on ADP occlusion in NBD2 than in NBD1.
Accessibility of an anti-HA antibody to the HA tag inserted in the ECDs
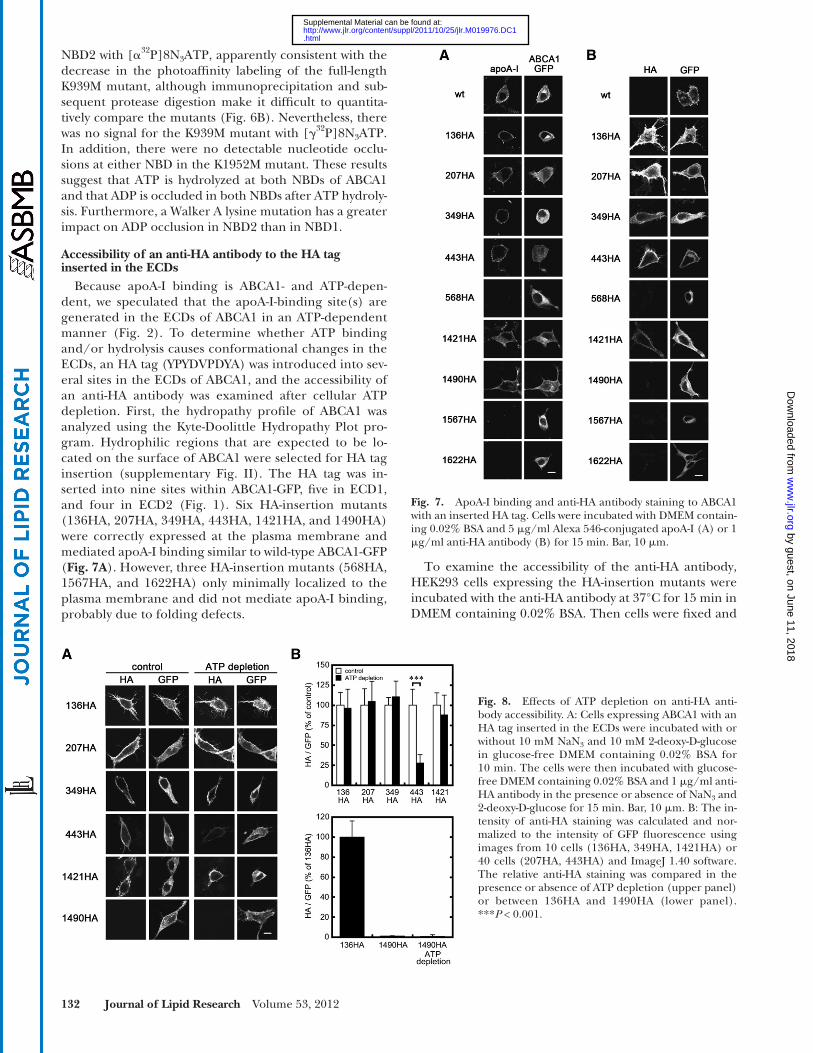
Because apoA-I binding is ABCA1- and ATP-depen-dent, we speculated that the apoA-I-binding site(s) are generated in the ECDs of ABCA1 in an ATP-dependent manner ( Fig. 2 ). To determine whether ATP binding and/or hydrolysis causes conformational changes in the ECDs, an HA tag (YPYDVPDYA) was introduced into sev-eral sites in the ECDs of ABCA1, and the accessibility of an anti-HA antibody was examined after cellular ATP depletion. First, the hydropathy profi le of ABCA1 was analyzed using the Kyte-Doolittle Hydropathy Plot pro-gram. Hydrophilic regions that are expected to be lo-cated on the surface of ABCA1 were selected for HA tag insertion (supplementary Fig. II). The HA tag was in-serted into nine sites within ABCA1-GFP, fi ve in ECD1, and four in ECD2 ( Fig. 1 ). Six HA-insertion mutants (136HA, 207HA, 349HA, 443HA, 1421HA, and 1490HA) were correctly expressed at the plasma membrane and mediated apoA-I binding similar to wild-type ABCA1-GFP ( Fig. 7A ). However, three HA-insertion mutants (568HA, 1567HA, and 1622HA) only minimally localized to the plasma membrane and did not mediate apoA-I binding, probably due to folding defects.
Fig. 7. ApoA-I binding and anti-HA antibody staining to ABCA1 with an inserted HA tag. Cells were incubated with DMEM contain-ing 0.02% BSA and 5 � g/ml Alexa 546-conjugated apoA-I (A) or 1 � g/ml anti-HA antibody (B) for 15 min. Bar, 10 � m.
Fig. 8. Effects of ATP depletion on anti-HA anti-body accessibility. A: Cells expressing ABCA1 with an HA tag inserted in the ECDs were incubated with or without 10 mM NaN 3 and 10 mM 2-deoxy-D-glucose in glucose-free DMEM containing 0.02% BSA for 10 min. The cells were then incubated with glucose-free DMEM containing 0.02% BSA and 1 � g/ml anti-HA antibody in the presence or absence of NaN 3 and 2-deoxy-D-glucose for 15 min. Bar, 10 � m. B: The in-tensity of anti-HA staining was calculated and nor-malized to the intensity of GFP fl uorescence using images from 10 cells (136HA, 349HA, 1421HA) or 40 cells (207HA, 443HA) and ImageJ 1.40 software. The relative anti-HA staining was compared in the presence or absence of ATP depletion (upper panel) or between 136HA and 1490HA (lower panel). *** P < 0.001.
by guest, on June 11, 2018w
ww
.jlr.orgD
ownloaded from
.html http://www.jlr.org/content/suppl/2011/10/25/jlr.M019976.DC1Supplemental Material can be found at:
Conformational changes of ABCA1 for apoA-I binding 133
whereas staining of the other fi ve mutants (136HA, 207HA, 349HA, 1421HA, and 1490HA) was not affected. These re-sults suggest that the accessibility of the anti-HA antibody to the HA tag at 443 is specifi cally altered by ATP depletion.
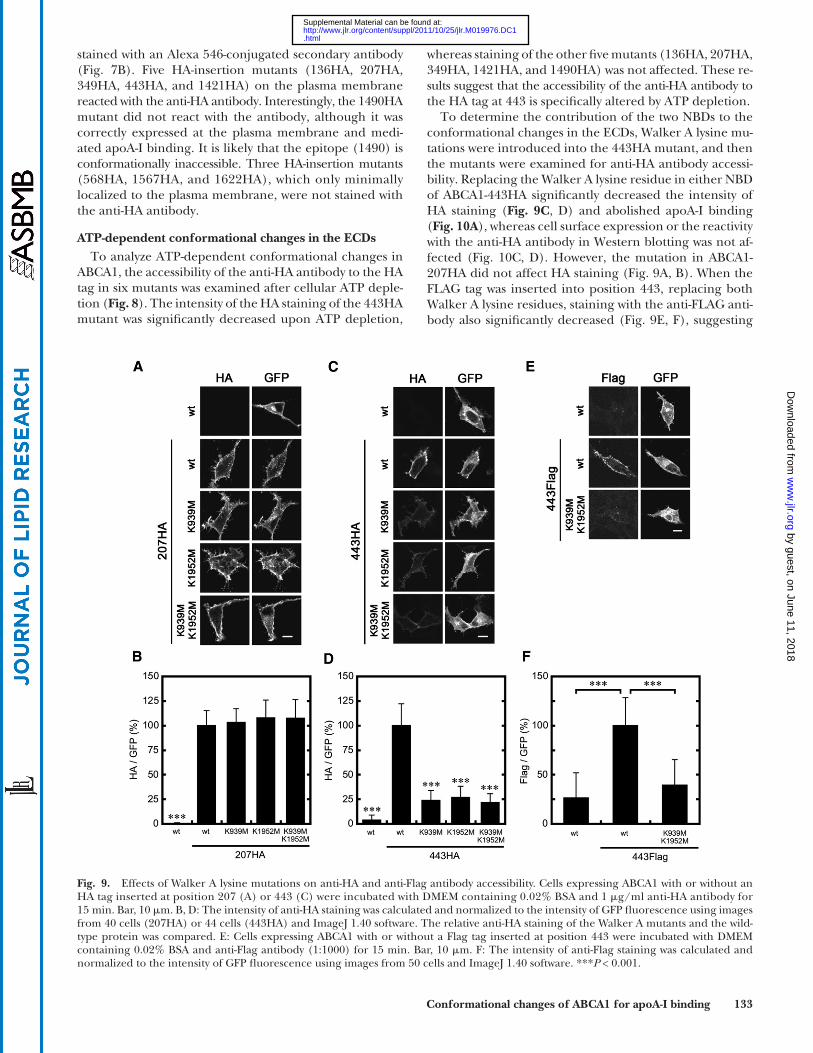
To determine the contribution of the two NBDs to the conformational changes in the ECDs, Walker A lysine mu-tations were introduced into the 443HA mutant, and then the mutants were examined for anti-HA antibody accessi-bility. Replacing the Walker A lysine residue in either NBD of ABCA1-443HA signifi cantly decreased the intensity of HA staining ( Fig. 9C , D ) and abolished apoA-I binding ( Fig. 10A ), whereas cell surface expression or the reactivity with the anti-HA antibody in Western blotting was not af-fected ( Fig. 10C, D ). However, the mutation in ABCA1-207HA did not affect HA staining ( Fig. 9A, B ). When the FLAG tag was inserted into position 443, replacing both Walker A lysine residues, staining with the anti-FLAG anti-body also signifi cantly decreased ( Fig. 9E, F ), suggesting
stained with an Alexa 546-conjugated secondary antibody ( Fig. 7B ). Five HA-insertion mutants (136HA, 207HA, 349HA, 443HA, and 1421HA) on the plasma membrane reacted with the anti-HA antibody. Interestingly, the 1490HA mutant did not react with the antibody, although it was correctly expressed at the plasma membrane and medi-ated apoA-I binding. It is likely that the epitope (1490) is conformationally inaccessible. Three HA-insertion mutants (568HA, 1567HA, and 1622HA), which only minimally localized to the plasma membrane, were not stained with the anti-HA antibody.
ATP-dependent conformational changes in the ECDs To analyze ATP-dependent conformational changes in
ABCA1, the accessibility of the anti-HA antibody to the HA tag in six mutants was examined after cellular ATP deple-tion ( Fig. 8 ). The intensity of the HA staining of the 443HA mutant was signifi cantly decreased upon ATP depletion,
Fig. 9. Effects of Walker A lysine mutations on anti-HA and anti-Flag antibody accessibility. Cells expressing ABCA1 with or without an HA tag inserted at position 207 (A) or 443 (C) were incubated with DMEM containing 0.02% BSA and 1 � g/ml anti-HA antibody for 15 min. Bar, 10 � m. B, D: The intensity of anti-HA staining was calculated and normalized to the intensity of GFP fl uorescence using images from 40 cells (207HA) or 44 cells (443HA) and ImageJ 1.40 software. The relative anti-HA staining of the Walker A mutants and the wild-type protein was compared. E: Cells expressing ABCA1 with or without a Flag tag inserted at position 443 were incubated with DMEM containing 0.02% BSA and anti-Flag antibody (1:1000) for 15 min. Bar, 10 � m. F: The intensity of anti-Flag staining was calculated and normalized to the intensity of GFP fl uorescence using images from 50 cells and ImageJ 1.40 software. *** P < 0.001.
by guest, on June 11, 2018w
ww
.jlr.orgD
ownloaded from
.html http://www.jlr.org/content/suppl/2011/10/25/jlr.M019976.DC1Supplemental Material can be found at:
134 Journal of Lipid Research Volume 53, 2012
In this study, we examined the contribution of the two NBDs to ABCA1 function.
First, the Walker A lysine in each NBD was replaced with methionine. This amino acid substitution in either NBD abolished apoA-I binding and apoA-I-dependent choles-terol effl ux by ABCA1, but it did not affect ATP binding to the NBDs. The amino acid substitution of glutamate in the Walker B motif, which is involved in ATP hydrolysis but not in ATP binding ( 33 ), in either NBD abolished apoA-I binding and apoA-I-dependent cholesterol effl ux by ABCA1 (supplementary Fig. III). These results suggest that the two intact NBDs are required for ABCA1 function, that ATP binding to NBDs is not suffi cient, and that ATP hydroly-sis is required for apoA-I binding and apoA-I-dependent cholesterol effl ux by ABCA1.
To analyze in detail the interaction between each NBD and nucleotides, we identifi ed a trypsin digestion site (R1272 and R1273) in the linker region between the two halves of ABCA1 and inserted a TEV protease recognition sequence that can specifi cally cleave ABCA1. TEV pro-tease-mediated cleavage of ABCA1 after photoaffi nity la-beling with [ � 32 P]8N 3 ATP and [ � 32 P]8N 3 ATP, together with the effects of mutations on the photoaffi nity labeling of full-length ABCA1, revealed the following characteris-tics of each NBD of ABCA1: i ) ATP binds equally to both NBDs; ii ) ATP is hydrolyzed at both NBDs; iii ) strong ADP occlusion occurs in NBD2 after hydrolysis, even in the ab-sence of vanadate, a phosphate analog; iv ) weak ADP oc-clusion occurs in NBD1 after hydrolysis in the presence of
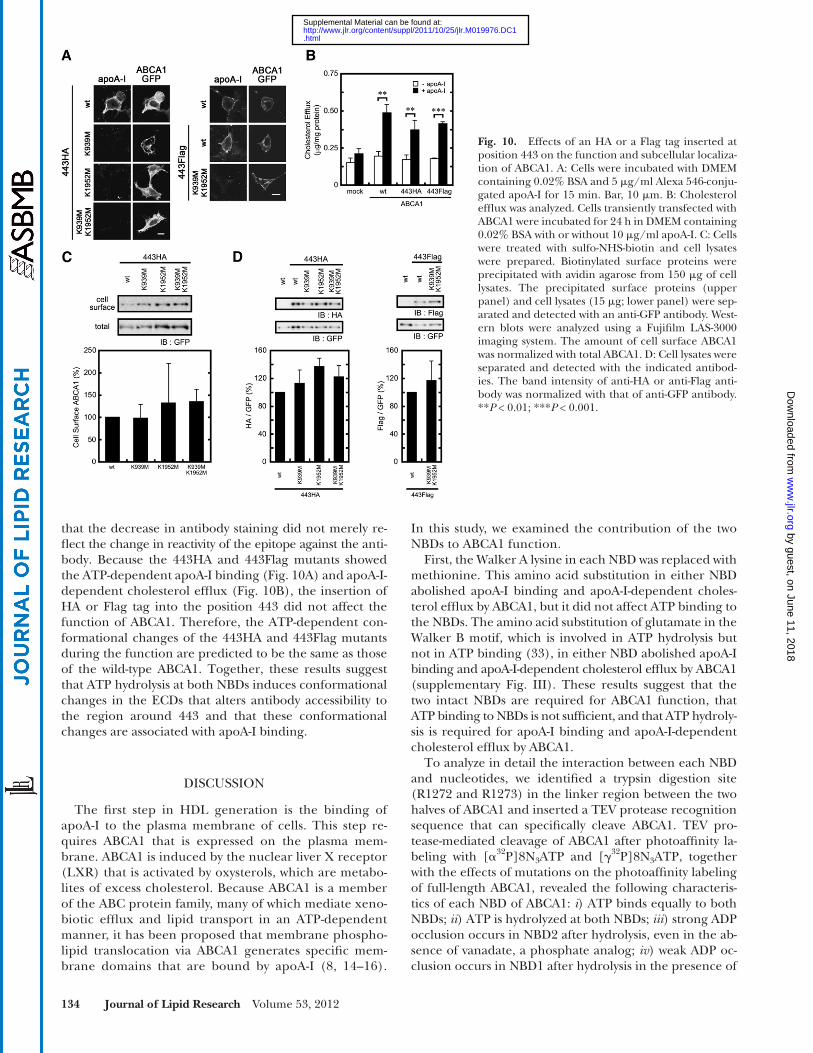
that the decrease in antibody staining did not merely re-fl ect the change in reactivity of the epitope against the anti-body. Because the 443HA and 443Flag mutants showed the ATP-dependent apoA-I binding ( Fig. 10A ) and apoA-I-dependent cholesterol effl ux ( Fig. 10B ), the insertion of HA or Flag tag into the position 443 did not affect the function of ABCA1. Therefore, the ATP-dependent con-formational changes of the 443HA and 443Flag mutants during the function are predicted to be the same as those of the wild-type ABCA1. Together, these results suggest that ATP hydrolysis at both NBDs induces conformational changes in the ECDs that alters antibody accessibility to the region around 443 and that these conformational changes are associated with apoA-I binding.
DISCUSSION
The fi rst step in HDL generation is the binding of apoA-I to the plasma membrane of cells. This step re-quires ABCA1 that is expressed on the plasma mem-brane. ABCA1 is induced by the nuclear liver X receptor (LXR ) that is activated by oxysterols, which are metabo-lites of excess cholesterol. Because ABCA1 is a member of the ABC protein family, many of which mediate xeno-biotic effl ux and lipid transport in an ATP-dependent manner, it has been proposed that membrane phospho-lipid translocation via ABCA1 generates specifi c mem-brane domains that are bound by apoA-I ( 8, 14–16 ).
Fig. 10. Effects of an HA or a Flag tag inserted at position 443 on the function and subcellular localiza-tion of ABCA1. A: Cells were incubated with DMEM containing 0.02% BSA and 5 � g/ml Alexa 546-conju-gated apoA-I for 15 min. Bar, 10 � m. B: Cholesterol effl ux was analyzed. Cells transiently transfected with ABCA1 were incubated for 24 h in DMEM containing 0.02% BSA with or without 10 � g/ml apoA-I. C: Cells were treated with sulfo-NHS-biotin and cell lysates were prepared. Biotinylated surface proteins were precipitated with avidin agarose from 150 � g of cell lysates. The precipitated surface proteins (upper panel) and cell lysates (15 � g; lower panel) were sep-arated and detected with an anti-GFP antibody. West-ern blots were analyzed using a Fujifi lm LAS-3000 imaging system. The amount of cell surface ABCA1 was normalized with total ABCA1. D: Cell lysates were separated and detected with the indicated antibod-ies. The band intensity of anti-HA or anti-Flag anti-body was normalized with that of anti-GFP antibody. ** P < 0.01; *** P < 0.001.
by guest, on June 11, 2018w
ww
.jlr.orgD
ownloaded from
.html http://www.jlr.org/content/suppl/2011/10/25/jlr.M019976.DC1Supplemental Material can be found at:

Conformational changes of ABCA1 for apoA-I binding 135
hydrolysis cycles and that they are not due to temporal ATP binding. The extracellular domain of ABCA1 is formed with ECD1 (598 aa) and ECD2 (288 aa) ( 25 ), which are connected by two intramolecular disulfi de bonds ( 17 ). We hypothesize that lipid accumulation within the extra-cellular domain via ATP hydrolysis-dependent lipid transport causes conformational changes that generate apoA-I-binding site(s) on the surface of the extracellular domain and that apoA-I bound to the site(s) is directly loaded with lipids by ABCA1 ( 42 ). Because antibody ac-cessibility was different for only one of six sites in the ECDs, apoA-I may bind to specifi c regions within the ECDs, al-though we found no signifi cant effect on apoA-I binding or HA staining when the 443HA mutant was preincubated with the anti-HA antibody or apoA-I (supplementary Fig. IV). These results suggest that the apoA-I binding site is not very close to position 443, although the exposure of posi-tion 443 to the surface of the ECD is accompanied by the exposure of the apoA-I binding site. Recently, we reported that lysine residues in the ECDs of ABCA1 contribute to the interaction with apoA-I and that the electrostatic inter-action between ABCA1 and apoA-I is predicted to be the fi rst step in HDL formation ( 21 ). ATP hydrolysis may cause conformational changes that generate a lysine cluster on the surface of the extracellular domain as apoA-I-binding site(s). This study facilitates our understanding of the mechanism of HDL formation by ABCA1.
REFERENCES
1 . Goldstein , J. L. , R. A. DeBose-Boyd , and M. S. Brown . 2006 . Protein sensors for membrane sterols. Cell . 124 : 35 – 46 .
2 . Nagao , K. , M. Tomioka , and K. Ueda . 2011 . Function and regula-tion of ABCA1--membrane meso-domain organization and re-orga-nization. FEBS J. 278 : 3190 – 3203 .
3 . Bodzioch , M. , E. Orso , J. Klucken , T. Langmann , A. Bottcher , W. Diederich , W. Drobnik , S. Barlage , C. Buchler , M. Porsch-Ozcurumez , et al . 1999 . The gene encoding ATP-binding cassette transporter 1 is mutated in Tangier disease. Nat. Genet. 22 : 347 – 351 .
4 . Brooks-Wilson , A. , M. Marcil , S. Clee , L. Zhang , K. Roomp , M. van Dam , L. Yu , C. Brewer , J. Collins , H. Molhuizen , et al . 1999 . Mutations in ABC1 in Tangier disease and familial high-density li-poprotein defi ciency. Nat. Genet. 22 : 336 – 345 .
5 . Rust , S. , M. Rosier , H. Funke , J. Real , Z. Amoura , J. Piette , J. Deleuze , H. Brewer , N. Duverger , P. Denefl e , et al . 1999 . Tangier disease is caused by mutations in the gene encoding ATP-binding cassette transporter 1. Nat. Genet. 22 : 352 – 355 .
6 . Oram , J. F. , R. M. Lawn , M. R. Garvin , and D. P. Wade . 2000 . ABCA1 is the cAMP-inducible apolipoprotein receptor that mediates choles-terol secretion from macrophages. J. Biol. Chem. 275 : 34508 – 34511 .
7 . Tanaka , A. R. , S. Abe-Dohmae , T. Ohnishi , R. Aoki , G. Morinaga , K. I. Okuhira , Y. Ikeda , F. Kano , M. Matsuo , N. Kioka , et al . 2003 . Effects of mutations of ABCA1 in the fi rst extracellular domain on subcellular traffi cking and ATP binding/hydrolysis. J. Biol. Chem. 278 : 8815 – 8819 .
8 . Wang , N. , D. Silver , P. Costet , and A. Tall . 2000 . Specifi c binding of ApoA-I, enhanced cholesterol effl ux, and altered plasma mem-brane morphology in cells expressing ABC1. J. Biol. Chem. 275 : 33053 – 33058 .
9 . Yokoyama , S. 2000 . Release of cellular cholesterol: molecular mechanism for cholesterol homeostasis in cells and in the body. Biochim. Biophys. Acta . 1529 : 231 – 244 .
10 . Khera , A. V. , M. Cuchel , M. de la Llera-Moya , A. Rodrigues , M. F. Burke , K. Jafri , B. C. French , J. A. Phillips , M. L. Mucksavage , R. L. Wilensky , et al . 2011 . Cholesterol effl ux capacity, high-density lipo-protein function, and atherosclerosis. N. Engl. J. Med. 364 : 127 – 135 .
11 . Nagao , K. , Y. Kimura , M. Mastuo , and K. Ueda . 2010 . Lipid outward translocation by ABC proteins. FEBS Lett. 584 : 2717 – 2723 .
vanadate; and v ) Walker A lysine mutations in either NBD affect ADP occlusion in both NBDs.
The roles of the two NBDs in the ATP hydrolysis cycle have been reported for several ABC proteins. The two NBDs of MDR1 (ABCB1) are virtually equivalent ( 34 ), al-though some nonequivalency has also been reported ( 35 ). ATP hydrolysis occurs alternatively ( 34 ), and ADP is oc-cluded equally in both NBDs in the presence of vanadate ( 36 ). However, in other ABC proteins, the two NBDs are proposed to have nonequivalent roles, one catalytic and the other regulatory. In the case of MRP1 (ABCC1), ATP is hydrolyzed only at NBD2, while NBD1 plays a regulatory role ( 29 ). Similar models have been proposed for CFTR (ABCC7) ( 30 ), SURs (ABCC8, C9) ( 31 ), and ABCA4 ( 32 ). MRP1 mainly transports hydrophilic compounds that are conjugated with glutathione ( 37, 38 ). SUR1 and CFTR are not active transporters but function as a regulator and a channel ( 39 ). In ABCA4, only NBD2 binds and hydrolyzes ATP, whereas NBD1, containing a bound ADP, plays a cru-cial, noncatalytic role in ABCA4 function ( 32 ). Although ABCA4 and ABCA1 have about 40% homology and are predicted to have similar secondary structures, the direc-tions of transport are proposed to be opposite ( 40 ). Be-cause the two NBDs of ABCA1 hydrolyze ATP and must be intact for the function of ABCA1, the ATP hydrolysis cycle for ABCA1 may more closely resemble that of MDR1 than that of the other ABC proteins.
Because apoA-I binding is ATP hydrolysis-dependent, we assumed that ATP hydrolysis causes conformational changes in the ECDs to generate apoA-I-binding site(s). To determine whether this is the case, an HA tag was intro-duced into nine hydrophilic regions in the ECDs, and the accessibility of an anti-HA antibody was examined before and after cellular ATP depletion. We found that HA stain-ing of the 443HA mutant was signifi cantly decreased by ATP depletion, suggesting that the region around 443 moves drastically in an ATP-dependent manner. Replac-ing the Walker A lysine residue in either NBD of ABCA1-443HA signifi cantly decreased the intensity of HA staining ( Fig. 9C, D ) and abolished apoA-I binding, whereas the mutation in ABCA1-207HA did not affect HA staining ( Fig. 9A, B ). When the FLAG tag was inserted into position 443, replacing both Walker A lysine residues, staining with the anti-FLAG antibody also signifi cantly decreased ( Fig. 9E, F ), suggesting that the decrease in antibody staining did not merely refl ect the change in reactivity of the epitope against the antibody. Together, these results sug-gest that ATP hydrolysis at both NBDs induces conforma-tional changes in the ECD and alters antibody accessibility to the region around 443.
It has been proposed that ATP binding triggers confor-mational changes in ABC proteins ( 41 ). ATP binding alters the conformation from an inward-facing, substrate-bind-ing confi guration to an outward-facing, substrate-releasing confi guration. ATP hydrolysis is proposed to convert the protein to the basal, inward-facing confi guration. Because changes in apoA-I binding and antibody accessibility to 443HA require ATP hydrolysis, it is plausible that the apoA-I-binding site(s) are generated by the buildup of ATP
by guest, on June 11, 2018w
ww
.jlr.orgD
ownloaded from
.html http://www.jlr.org/content/suppl/2011/10/25/jlr.M019976.DC1Supplemental Material can be found at:

136 Journal of Lipid Research Volume 53, 2012
12 . Chambenoit , O. , Y. Hamon , D. Marguet , H. Rigneault , M. Rosseneu , and G. Chimini . 2001 . Specifi c docking of apolipoprotein A-I at the cell surface requires a functional ABCA1 transporter. J. Biol. Chem. 276 : 9955 – 9960 .
13 . Takahashi , K. , Y. Kimura , N. Kioka , M. Matsuo , and K. Ueda . 2006 . Purifi cation and ATPase activity of human ABCA1. J. Biol. Chem. 281 : 10760 – 10768 .
14 . Hamon , Y. , C. Broccardo , O. Chambenoit , M. Luciani , F. Toti , S. Chaslin , J. Freyssinet , P. Devaux , J. McNeish , D. Marguet , et al . 2000 . ABC1 promotes engulfment of apoptotic cells and transbilayer re-distribution of phosphatidylserine. Nat. Cell Biol. 2 : 399 – 406 .
15 . Vedhachalam , C. , P. T. Duong , M. Nickel , D. Nguyen , P. Dhanasekaran , H. Saito , G. H. Rothblat , S. Lund-Katz , and M. C. Phillips . 2007 . Mechanism of ATP-binding cassette transporter A1-mediated cellular lipid effl ux to apolipoprotein A-I and formation of high density lipoprotein particles. J. Biol. Chem. 282 : 25123 – 25130 .
16 . Lin , G. , and J. F. Oram . 2000 . Apolipoprotein binding to protrud-ing membrane domains during removal of excess cellular choles-terol. Atherosclerosis . 149 : 359 – 370 .
17 . Hozoji , M. , Y. Kimura , N. Kioka , and K. Ueda . 2009 . Formation of two intramolecular disulfi de bonds is necessary for ApoA-I-dependent cholesterol effl ux mediated by ABCA1. J. Biol. Chem. 284 : 11293 – 11300 .
18 . Fitzgerald , M. L. , A. L. Morris , A. Chroni , A. J. Mendez , V. I. Zannis , and M. W. Freeman . 2004 . ABCA1 and amphipathic apolipopro-teins form high-affi nity molecular complexes required for choles-terol effl ux. J. Lipid Res. 45 : 287 – 294 .
19 . Chroni , A. , T. Liu , M. L. Fitzgerald , M. W. Freeman , and V. I. Zannis . 2004 . Cross-linking and lipid effl ux properties of apoA-I mutants suggest direct association between apoA-I helices and ABCA1. Biochemistry . 43 : 2126 – 2139 .
20 . Vedhachalam , C. , A. B. Ghering , W. S. Davidson , S. Lund-Katz , G. H. Rothblat , and M. C. Phillips . 2007 . ABCA1-induced cell surface bind-ing sites for ApoA-I. Arterioscler. Thromb. Vasc. Biol. 27 : 1603 – 1609 .
21 . Nagao , K. , Y. Kimura , and K. Ueda . 2011 . Lysine residues of ABCA1 are required for the interaction with apoA-I. Biochim. Biophys. Acta . Epub ahead of print. July 1, 2011; doi: 10.1016/j.bbalip.2011.06.024.
22 . Wang , N. , D. L. Silver , C. Thiele , and A. R. Tall . 2001 . ATP-binding cassette transporter A1 (ABCA1) functions as a cholesterol effl ux regulatory protein. J. Biol. Chem. 276 : 23742 – 23747 .
23 . Munehira , Y. , T. Ohnishi , S. Kawamoto , A. Furuya , K. Shitara , M. Imamura , T. Yokota , S. Takeda , T. Amachi , M. Matsuo , et al . 2004 . Alpha1-syntrophin modulates turnover of ABCA1. J. Biol. Chem. 279 : 15091 – 15095 .
24 . Azuma , Y. , M. Takada , H-W. Shin , N. Kioka , K. Nakayama , and K. Ueda . 2009 . Retroendocytosis pathway of ABCA1/apoA-I contrib-utes to HDL formation. Genes Cells . 14 : 191 – 204 .
25 . Tanaka , A. R. , Y. Ikeda , S. Abe-Dohmae , R. Arakawa , K. Sadanami , A. Kidera , S. Nakagawa , T. Nagase , R. Aoki , N. Kioka , et al . 2001 . Human ABCA1 contains a large amino-terminal extracellular do-main homologous to an epitope of Sjogren’s Syndrome. Biochem. Biophys. Res. Commun. 283 : 1019 – 1025 .
26 . Abe-Dohmae , S. , S. Suzuki , Y. Wada , H. Aburatani , D. E. Vance, and S. Yokoyama. 2000 . Characterization of apolipoprotein-mediated HDL generation Induced by cAMP in a mouse macrophage cell line. Biochemistry . 39 : 11092 – 11099 .
27 . Amundson , D. M. , and M. Zhou . 1999 . Fluorometric method for the enzymatic determination of cholesterol. J. Biochem. Biophys. Methods . 38 : 43 – 52 .
28 . Nagao , K. , Y. Zhao , K. Takahashi , Y. Kimura , and K. Ueda . 2009 . Sodium taurocholate-dependent lipid effl ux by ABCA1: effects of W590S mutation on lipid translocation and apolipoprotein A-I dis-sociation. J. Lipid Res. 50 : 1165 – 1172 .
29 . Gao , M. , H. R. Cui , D. W. Loe , C. E. Grant , K. C. Almquist , S. P. Cole , and R. G. Deeley . 2000 . Comparison of the functional charac-teristics of the nucleotide binding domains of multidrug resistance protein 1. J. Biol. Chem. 275 : 13098 – 13108 .
30 . Aleksandrov , L. , A. A. Aleksandrov , X. B. Chang , and J. R. Riordan . 2002 . The fi rst nucleotide binding domain of cystic fi brosis trans-membrane conductance regulator is a site of stable nucleotide in-teraction, whereas the second is a site of rapid turnover. J. Biol. Chem. 277 : 15419 – 15425 .
31 . Matsuo , M. , K. Tanabe , N. Kioka , T. Amachi , and K. Ueda . 2000 . Different binding properties and affi nities for ATP and ADP among sulfonylurea receptor subtypes, SUR1, SUR2A, and SUR2B. J. Biol. Chem. 275 : 28757 – 28763 .
32 . Ahn , J. , S. Beharry , L. L. Molday , and R. S. Molday . 2003 . Functional interaction between the two halves of the photoreceptor-specifi c ATP binding cassette protein ABCR (ABCA4). Evidence for a non-exchangeable ADP in the fi rst nucleotide binding domain. J. Biol. Chem. 278 : 39600 – 39608 .
33 . Tombline , G. , L. A. Bartholomew , I. L. Urbatsch , and A. E. Senior . 2004 . Combined mutation of catalytic glutamate residues in the two nucleotide binding domains of P-glycoprotein generates a conformation that binds ATP and ADP tightly. J. Biol. Chem. 279 : 31212 – 31220 .
34 . Senior , A. E. , M. K. al-Shawi , and I. L. Urbatsch . 1995 . The catalytic cycle of P-glycoprotein. FEBS Lett. 377 : 285 – 289 .
35 . Takada , Y. , K. Yamada , Y. Taguchi , K. Kino , M. Matsuo , S. J. Tucker , T. Komano , T. Amachi , and K. Ueda . 1998 . Non-equivalent cooper-ation between the two nucleotide-binding folds of P-glycoprotein. Biochim. Biophys. Acta . 1373 : 131 – 136 .
36 . Urbatsch , I. L. , B. Sankaran , S. Bhagat , and A. E. Senior . 1995 . Both P-glycoprotein nucleotide-binding sites are catalytically active. J. Biol. Chem. 270 : 26956 – 26961 .
37 . Borst , P. , R. Evers , M. Kool , and J. Wijnholds . 2000 . A family of drug transporters: the multidrug resistance-associated proteins. J. Natl. Cancer Inst. 92 : 1295 – 1302 .
38 . Leslie , E. M. , R. G. Deeley , and S. P. Cole . 2001 . Toxicological rel-evance of the multidrug resistance protein 1, MRP1 (ABCC1) and related transporters. Toxicology . 167 : 3 – 23 .
39 . Ueda , K. , M. Matsuo , K. Tanabe , N. Kioka , and T. Amachi . 1999 . Comparative aspects of the function and mechanism of SUR1 and MDR1 proteins. Biochim. Biophys. Acta . 1461 : 305 – 313 .
40 . Sun , H. , R. Molday , and J. Nathans . 1999 . Retinal stimulates ATP hydrolysis by purifi ed and reconstituted ABCR, the photoreceptor-specifi c ATP-binding cassette transporter responsible for Stargardt disease. J. Biol. Chem. 274 : 8269 – 8281 .
41 . Higgins , C. F. , and K. J. Linton . 2004 . The ATP switch model for ABC transporters. Nat. Struct. Mol. Biol. 11 : 918 – 926 .
42 . Ueda , K. 2011 . ABC proteins protect the human body and maintain optimal health. Biosci. Biotechnol. Biochem. 75 : 401 – 409 .
by guest, on June 11, 2018w
ww
.jlr.orgD
ownloaded from
.html http://www.jlr.org/content/suppl/2011/10/25/jlr.M019976.DC1Supplemental Material can be found at: