auk 119-3-715
TRANSCRIPT

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 1/20
715
The Auk 119(3):715–734, 2002
PHYLOGENY OF THE TYRANT FLYCATCHERS (TYRANNIDAE)
BASED ON MORPHOLOGY AND BEHAVIOR
JEFFREY S. BIRDSLEY1
W. K. Kellogg Biological S tation and D epartment of Z oology, M ichigan S tate University, 3700 East Gull Lake Drive,
Hickory Corners, Michigan 49060, USA
ABSTRACT.—Previously published morphological and behavioral data for the tyrant fly-
catchers (Tyrann idae) were reanalyzed using cladistic techniques. Several add itional char-
acters, includ ing two p utative synapom orphies of the Tyrannidae were incorporated. Nearly
all of the 100 traditional tyran nid genera were includ ed. Results of the analysis of this set
of 68 characters support three previously proposed tyrannid assemblages: the kingbird as-
semblage and slightly restricted Empidonax an d Myiarchus assemblages. Characters of the
nasal septum that past workers have considered conservative and phylogenetically infor-
mative are su pp orted as synap omorphies of the kingbird and restricted Empidonax assem-
blages. Several sma ll monophyletic groups of genera are su pp orted. The monop hyly of nei-
ther the Elaenia assemblage nor the flatbill and tody-tyrant assemblage is supported in anymost-parsimonious tree, but such trees are not significantly better than trees supporting
their mon ophyly. A Tyrannidae exclusive of the Cotingidae an d Pipridae is monop hyletic in
some most-par simonious trees, but not in oth ers in wh ich flatbills and tody-tyrants are basal
to a clade containing Cotingidae, Pipridae, and the remaining tyrann id genera. There is some
evidence that characters describing plumage color pattern are more homoplasious than the
other character suites I examined. Received 5 Au gust 2000, accepted 9 February 2002.
RESUMEN .—Utilizand o tecnicas cladı sticas rean alice dato s mor folog icos y condu ctua les
pr eviamen te pu blicados pa ra la fam ilia Tyra nn idae. Incorp ore varios caracteres adicionales
incluyendo dos sinapomorfias putativas. Incluı casi todos los generos de los aproximad a-
men te 100 gener os tra dicionales d e la familia Tyra nn idae. Los resultad os del an alisis de u n
set de 68 caracteres apoyan tres gru pos d e tiranid os previamente p ropuestos: el grupo Tyran-nu s y los grupos Emipidonax y Myiarchus. Los caracteres relacionados con el septum nasal
que han sido considerados previamente como conservadores y filogeneticamente informa-
tivos, son apoyados como sinapomorfias del grup o Tyrannus y del grup o restringido Empi-
donax. Varios pequ en os gr up os d e gen eros m onofileticos son ap oyados. N i la m onofilia d el
g r u p o Elaenia ni la de los picochato (e.g. Plathyrhinchus) y mosqueritos (e.g. Todirostrum , He-
mitriccus) es a poyad a en cualqu iera d e los a rb oles ma s pa rsimon iosos, per o estos arb oles no
son significativamente mejores que los que apoyan su monofilia. Un Tyrannidae exclusivo
de la fam ilia Cotingid ae y P iprid ae es m onofiletico en a lgun o d e los ar boles ma s p arsim o-
niosos, pero no en otros en los que los picochatos y m osqueritos son basales a un clado que
contiene a Coting idae, Pipr idae y a los d ema s gener os d e tira n idos. Existe cierta evid encia
qu e los carater es que d escrib en los p atr one s de coloracion d el plum aje son mas hom op las icos
que el set d e caracteres qu e exam inee.
THE TYRANT FLYCATCHERS (Tyran nidae) are a
large, primarily Neotropical family of subos-
cine passerine birds in the su perfamily Tyran-
noidea w hich also includ e the cotingas (Cotin-
g id a e) a n d m a n ak in s (P ip r id a e). A g r eat
behavioral and ecological diversity within that
superfam ily presents r ich opportun ities for
comparative tests of evolutionary and ecologi-
1 Present add ress: Departm ent of Biological Sci-ence, Florida State University, Tallahassee, Florida
32306, USA. E-ma il: bird [email protected] u
cal hy po theses (e.g. Pru m 1990a, 1994, 1997).
Because w ell-sup ported phylogenies are fun-
damental to such tests, the evolutionary rela-
tionships of these birds have received much re-
cent attention (W. E. Lanyon 1984, 1985, 1986,
1988a, b , c; McKitrick 1985; S. M. Lanyon 1985;
Prum and Lanyon 1989; Prum 1990b, 1992; Mo-
bley an d Pru m 1995; Bostw ick 2000). W. E. Lan -
yon’s work in par ticular h as resulted in an ex-
t en s iv e co m p ar at iv e m o rp h olo gica l a n d
behavioral data set for the Tyrannidae.The relationsh ips am ong t he 90 to 100 genera
of tyrant flycatchers were historically based on

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 2/20
716 [Auk , Vol. 119JEFFREY S. BIRDSLEY
the shap e of the bill , wing and tail, and color
and p attern of plum age. Warter’s (1965) sur vey
of cranial osteological variation in the Tyran-
noidea revealed variation in the nasal capsule
a n d s ep t u m , p a lat in e a n d p a lat om a xi lla r y
bones, interorbital septum, and general shapeof the craniu m. Warter felt that the nasa l region
provided useful taxonomic characters an d rec-
ognized several basic states of the configu ra-
tion of the nasal septum . Building u pon War-
ter’s (1965) work, W. E. Lanyon (1984, 1985,
1986, 1988a, b, c) completed a nearly exhaus-
tive survey of tyrannoid cranial variation and
used states of the nasal septu m to hyp othesize
m onophyly of five separate tyrannid assem -
blages that collectively included nearly all tra-
ditional tyrannid genera.Ames (1971) documented great variation in
suboscine syringeal m u sculature and supp ort
elements. However, Ames did not use differ-
ential stains and so could not readily distin-
guish between ossified and cartilaginous sup-
p ort element s. Lanyon (1984, 1985, 1986, 1988a,
b , c) s t u d ie d a n e ar ly e xh au s t iv e s a m p le o f
tyrannoid syringes which he stained to distin-
guish the ossified (primarily tracheal) series of
elem ents from the cartilaginous (prim arily
bronchial) series. This allowed recognition of
homologous elements. Lanyon used syringeal
characters to group taxa within his five assem-
b la g es a n d u s e d ch a r act er s o f t h e cr a n iu m ,
nesting behavior, plumage, and egg coloration
wherever necessary to establish relationships
within syringeal group s and occasionally at
higher levels. That procedu re is the equivalent
of weighting those syringeal characters more
heavily than others, effectively disallowing
convergence in them. Nasal septu m characters
were effectively weighted even more heavily
than syringeal characters because Lanyon as-sum ed them to be synapom orphies of his five
assem blages and disallowed convergence in
them .
Lanyon (1984, 1985, 1986, 1988a, b , c) d evel-
oped hyp otheses of generic relationsh ips based
on shared similarities, but because he did not
do so w ithin a clad istic framework, some of his
group s were su pp orted by plesiomorph ies. For
example, in the Elaenia group Lanyon (1988a)
used the poorly ossified condition of the ante-
rior segm ent of the nasal septum to supp ort aclade composed of Capsiempis, Phaeomyias, and
Nesotriccus. He then used the fully ossified con-
dition to sup port a clade com posed of Serpop-
haga and Anairetes. Only on e of those character
states is the d erived state and the other is the
plesiomorph ic, or ancestral, state in that grou p.
Logically, only the derived state may argue for
close relationship of the taxa that possess it.A cladistic reanalysis of Lanyon’s (1984,
1985, 1986, 1988a, b, c) data was performed
plus any other d ata that could be found in the
literature. I w eighted characters equally and
analyzed them together in one character matrix
containing nearly all tyrannid genera, includ-
ing several that Lanyon could n ot p lace in any
of his assemblages. That provided a test of the
monophyly of each of the five tyran nid assem-
blages Lanyon proposed and provided a test of
h o m olo g y o f e a ch o f t h e fi ve n a sa l s e p t u m
states that supp orted them . M onophyly of the
Tyrannidae has been questioned on the basis of
DNA–DNA hybridization analyses that place
several ‘‘mionectid’’ flycatchers as the sister
group to the rest of the tyrann oids (cotingids
piprids rem aining tyrannids) (Sibley and
Ah lquist 1985). The present an alysis provides a
test of the monop hyly of the Tyrann idae as tra-
ditionally constituted (e.g. American Ornithol-
ogist s’ Union [AOU] 1998).
I comp ared reliability of morp hological, be-
havioral, an d ecological characters to recovertyrannid phylogeny. Although past workers
have arg ued (e.g. Atz 1970) that beh avioral and
ecological characters lack phy logenetic infor-
mation, several studies have found such char-
acters to be at least as reliable as morp hological
and molecular characters (McLennan and Mat-
tern 2001, Paterson et al. 1995, DeQu eiroz an d
Wimberger 1993, but see Lee et al. 1996). Sig-
nificantly different levels of homoplasy among
four suites of tyrannid characters was tested
for: cranial, syringeal, plum age, and behavioraland ecological char acters.
ME T HODS
All 95 genera that served as terminal taxa in Lan-
yon’s (1984, 1985, 1986, 1988a, b, c) five assemblages
were included and the n omenclature follows h is.
Lanyon provided character su pp ort for m onophyly
of 64 of th ose gen era. Lanyon (1986, 1988a) sp lit M e-
cocerculus an d Myiophobus into three group s each and
placed them in d ifferent p arts of his phylogenies yet
retaining their generic names until more compara-
tive data are available. Here these taxa are split asLanyon d id. Lanyon (1988b) argu ed for merging ’’T-
erenotriccus’’ erythrurus into Myiobius a n d h i s r e c-

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 3/20
July 2002] 717Phylogeny of Tyrannidae
FIG . 1. Character state tree for character 11. Char-
acters and states are described in Appendix 1.
ommendation are followed here, but My iobius eryth-
rurus is kept as a terminal taxon separate from the
remaining Myiobius. Se ve r al t r a d it io n a l t y r an n i d
taxa h ave a nasal septum morphology that is either
unknown or is sufficiently unique that Lanyon (1986,
1988a) refrained from placing th em in any of his as-
semblages, maintaining them incertae sedis: Phyllom-
yias fasciatu s, P. griseiceps, P. gris eocapilla, a n d t he
monotypic genera Colonia, Culicivora, Machetornis,
Muscigralla, a n d Tachuris. A l l o f t h es e t a x a a r e i n -
clud ed in th is analysis except for Phyllomyias griseo-
capilla and Culicivora. For those two taxa, neither cra-
nial nor syringeal data were available. Also included
is the genus Neopipo, traditionally placed in the Pi-
pridae, because Mobley and Prum (1995) hypothe-
sized it to belong in the Tyrannidae on the basis of
syringeal and plumage data.
Character data were extracted prim arily from Lan-
yon (1984, 1985, 1986, 1988a, b, c). These referencesinclude photographs of representative syringes for
all terminal taxa and photogr aph s of crania for most.
Because Lanyon reported on one assemblage at a
time, he often did not explicitly describe, for a given
character, the d istribution of states across all tyran-
nid genera and in outgroups. In those cases, family-
wide assessments and outgroup comparisons were
mad e using Lanyon’s photograp hs and other litera-
ture sources describing tyrannoid crania (Warter
1965) and syringes (Ames 1971; McKitrick 1985;
Prum and Lanyon 1989; Prum 1990b, 1992; Mobley
and Prum 1995). Lanyon d id not make extensive use
of syringeal musculature characters so those were
extracted from Ames (1971). Because Ames (1971)
did not u se stains on ossified and cartilaginous sy-
ringeal sup port elements p recedence was given to all
more-recent information on those elements. Hilty
and Brown (1986), Stiles and Skutch (1989), Ridgely
and Tudor (1994) and similar guides for plumage
characters were consulted. Foraging behavior char-
acters are from Fitzpatrick (1980). For nest a nd egg
characters, an exhaustive literature search was con-
du cted, drawing information from many sources and
add ing two characters that Lanyon did not use: pres-
ence of a visor over the entrance of enclosed nests(character 66) and nests used as a dormitory (char-
acter 67). In add ition, crania, skins, and Lanyon’s
cleared-and-stained syringeal specimens were ex-
amined at the American Museum of Natural History
for any character data that were incomplete or in-
conclusive in the literature.
Lanyon (1984, 1986, 1988a) hypothesized the
Tyrannidae to be monophyletic based on the pres-
ence of internal syringeal cartilages. Prum (1990b)
questioned the homology of all internal syringeal
cartilages and recommended using unique, detailed
morphologies of internal cartilages as characters,
wh ile refraining from the hyp othesis of homology of internal cartilages as broadly d efined. Pru m (1990b)
hypothesized monophyly of the Tyrannidae on the
basis of the presence of Mm. obliqui ventrales, a pair
of intrinsic syringeal mu scles. In th is analysis ’’ Mm.
obliqui ventrales’’ is included as a character but not
‘‘internal cartilages,’’ except as several characters
that describe unique details of some internal carti-
lages. An add itional, heretofore unrecognized, p o-
tential synapomorphy of the tyrannids is included :B1 and B2 syringeal sup port elements w hich ar e con-
nected at their ventral tips. That connection is absent
in cotingids and piprids (Prum 1992; R. O. Prum
pers. comm .).
Morphological and behavioral variation were cod-
ed as 50 binary characters, 15 unordered multistate
characters, and 3 ordered mu ltistate characters (Ap-
pend ix 1). Characters 6 and 13 were ordered because
states shared significant detail but some states had
a d d i t io n a l d e t a il s t h a t a p p e a r ed t o b e f u r t h e r d e -
rived. For character 11 a state tree (Fig. 1) is defined
reflecting a trend toward increasing num bers of dou -
ble, complete syringeal A elements an d increasing
degree of ossification of those elements. That was
justified because such a trend has ap parently oc-
curred in piprids (Prum 1992) and in some falconi-
forms (Griffiths 1994). Also, the ontogenetic devel-
opment of those elements proceeds from an entirely
cartilaginous state to an at least partly ossified state
(Ames 1971).
PAUP* 4.0b8 (Swofford 2000) was u sed to p erform
heuristic searches for the most-parsimonious trees.
To increase the probability of find ing globally most-
parsimonious trees, I repeated this search using all
possible combinations of branch-swapp ing and step-wise-addition options. For each search I set a maxi-
mu m of 2,000 trees to be h eld in m emory. I also per-
formed 1,000 replicates with the random add ition
option and one tree held in m emory. Multistate taxa
were interpreted as polymorphisms. I gave all char-
acters equal weight. I felt this w as justified in the ab-
sence of objective criteria for w eighting some char-
acters m ore h eavily than others. Equal w eighting
schemes d o, however, carry the assumption of low
rates of change in all characters (Felsenstein 1982). I
rooted all trees to Fur narioidea and a llowed the oth-
er potential outgroups, Cotingidae and Pipridae, to
‘‘float’’ in the analysis. This allowed a test of them o n op h y ly o f t h e Ty r an n id a e a s t r ad i tio n al ly
constituted.

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 4/20
718 [Auk , Vol. 119JEFFREY S. BIRDSLEY
I used consensus trees to summarize the resulting
set of most-parsimonious trees. I computed a strict
consensus tree w hich identifies groups found in all
most-parsimon ious trees. I also compu ted an Adam s
consensus tree (Adams 1972) which better illustrates
patterns and trends among alternative most-parsi-
monious tr ees by identifying group s (not necessarily
monophyletic) that consistently nest within larger
groups in all trees. For example, if a set of nested
groups exists in all most-parsimonious trees but in
some of them is ‘‘invaded’’ by one taxon or a small
group of taxa, the Adams consensus preserves the
nested structure and places the problematic group
outside of it in a polytomous position. The strict con-
s en s u s s im p l y co ll ap s e s t h e e n t ir e n e st e d g r ou p .
Consensus trees should be interpreted with care as
they may not be the m ost parsimonious hypotheses
suggested by the data (Swofford 1991).
To evaluate the evidential sup port for relation-ships d epicted in the strict consensus tr ee I used So-
renson’s (1996) TreeRot program and PAUP*to com-
pu te a decay ind ex (Brem er 1988, Kallersjo et al.
1992). I ran 10 replicates with TBR branch-swap pin g,
random addition sequence and a maximum of 800
trees held in memory. I also ran 100 replicates with
r a nd o m a d d i tio n s eq u en ce a n d o ne t r ee h eld i n
memory. The decay index gives the number of extra
steps required to show a node as unsupported. For
example, if a node that occurs in all most-parsimo-
nious trees is not present in trees that are one step
longer, that node gets a d ecay ind ex of 1. Higher in-
dex values indicate more robust clades.
I used two approaches to evaluate Lanyon’s (1984,
1985, 1986, 1988a, b, c) hyp othesized assemblages
within the context of my d ata m atrix. First, for those
assemblages not sup ported in the strict consensus of
most-parsimonious trees, I d etermined the nu mber
of extra steps required to force their monop hyly. To
do this I conducted heuristic searches constraining
mon ophy ly of each assemblag e in turn . For each I ran
10 replicates w ith TBR branch-swapping, random
addition, and a maximum of 800 trees held in mem-
ory. I also ran 1,000 replicates with one tree h eld in
mem ory. I u sed Temp leton’s test (Temp leton 1983) asimplemented in PAUP*to determine whether the re-
sulting trees w ere significantly d ifferent from the mo st-
pa rsimon ious trees. Second, I used MACCLADE 3.04
(Madd ison and Maddison 1992) to examine evolu-
tion of nasal septum characters on the most-parsi-
monious trees. I evaluated the homology of septum
characters that Lanyon (1984, 1985, 1986, 1988a, b, c)
used to define his assemblages.
Finally, I u sed MACCLADE to calculate rescaled
consistency ind ices (RC) for each cha racter and to av-
erage them across 2,000 most-parsimonious trees
representing all known tree islands. I u sed a single
factor ANOVA to determ ine if mean RC was signifi-cantly d ifferent amon g four suites of characters: cra-
nial (characters 1–10), syringeal (characters 12–45),
plum age (characters 46–59), and behavior/ ecology
(characters 60–67). I did not include character 11 be-
cause MacClade does n ot include characters of user-
defined type in these calculations.
RESULTS AN D D ISCUSSION
Phylogenetic analysis of 68 characters (a total
of 101 apomorp hies) and 106 taxa (Appen d ix 2)
identified more th an 2,000 most-parsimonious
trees of length 514 and a consistency index (CI)
of 0.44. I fou nd several ‘‘islands’’ of m ost-par-
simonious trees using different combinations
of PAUP’s search options. I found add itional
tree islands in the course of computing decay
indices. The strict consensus of all most-parsi-
monious trees (Figs. 2–4) indicates support for
the monop hyly of many small group s of generaand of three assem blages w hich correspond
more or less to Lanyon’s (1984, 1985, 1986)
kingbird, Myiarchus, and Empidonax assemblag-
es. The monophyly of neither an Elaenia assem-
blage (Lanyon 1988a) nor a flatbill and tod y-ty-
rant assemblage (Lanyon 1988c) is su pp orted.
An Adam s consensu s tree (Figs. 5 and 6) is bet-
ter resolved. Each clade depicted in the Adams
tree is present in at least some most-parsimo-
nious trees, but the tree as a w hole may not be
consistent with any single most-parsimonioustree. Despite this, for the purposes of the fol-
lowing discussion it i llustrates m any results
which are not apparent in the strict consensus
(Figs. 2–4). Here I discuss my results as com-
p ared to each of Lanyon’s (1984, 1985, 1986,
1988a, b, c) proposed assemblages and as com-
pared to the traditional concept of the family
Tyrannidae. Character numbers refer to char-
acters in the p resent analysis.
Tyrannidae monophyly. A traditional Tyran-
nidae exclusive of Cotingidae and Pipridae is
supp orted in som e m ost-parsim onious trees.
However, there are no unam biguous synapo-
morphies. Neither of the two putative synapo-
morph ies of the Tyrann idae provides unequiv-
ocal support for m onophyly of the fam ily as
traditionally defined. In all most-parsimonious
trees, intrinsic syringeal muscles Mm. obliqui
ventrales that insert ventrally (character 43,
s t at e 1) a r e a s y n ap o m o r p hy o f a r e st r ict ed
Tyrannidae whose exact m em bership varies
from tree to tree but n ever includes the tody-
t y ra n t g r ou p o r t h e Cnipodectes–Onychorhyn-chus– Myiobius clade. The ventrally connected
B1 and B2 elements (character 19) proved dif-

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 5/20
July 2002] 719Phylogeny of Tyrannidae
FIG . 2 . St r ic t c on s en s u s t r e e d e r iv ed f r om a l l
most-parsimonious trees. Decay indices are shown
when greater than 1. The two taxa at the bottom de-
note the assemblages shown in Figures 3 and 4.
FIG . 3. Strict consensus tree of the kingbird as-
semblage. Decay indices are shown when greater
than 1.
fi c u l t t o e v a l u a t e a s a s y n a p o m o r p h y o f t h e
Tyrannidae. The basal or next-to-basal group s
in each m ost-parsim onious tree p ossess this
character, but the outgr oup (Furn arioidea) was
coded ’’?’’ (unknown). Therefore character 19
d o es n o t p r ov id e a n u n e qu i vo ca l s yn a p o-
m orphy of the Tyrannidae in any m ost-parsi-
mon ious trees. To assess th e level at wh ich it is
informative, one would need more information
on this trait in the Furnarioidea and Old World
suboscines.
Flatbill and tody-ty rant assemblage. Lanyon
(1988c) defined a flatbill and tody-tyrant as-
semblage based on the possession of: unossi-fied nasal septa (character 1); cranial interor-
bital septa in which the supraorbital fenestra is
redu ced (character 9); and enclosed, p endant
nests (character 64). This assem blage is not
s u p p o r t e d i n a n y m o s t-p a r s im o n io u s t r ee s.
Four extra steps are required to force its mono-
phyly, but such trees are not significantly dif-
ferent from most-parsimonious trees (P 0.64;
21 n 27 characters requiring differing
num bers of steps on th e trees being compared ).
Within his assemblage, Lanyon (1988c) defin ed
a fl at bill g rou p ( Rhynchocyclus, Tolmomyias ,
Onychorhynchus, and Platyrinchus) based on the
p ossession of at least one ossified, dou ble, com-
plete syringeal A element (character 11). This
state is present in m any tyrannids outside the
flatbills and tody-tyrants and is not a synapo-
m orphy of these flatbills in any m ost-parsi-
mon ious trees. Lanyon’s (1988c) hyp othesized
sister relationship between Onychorhynchus and
Platyrinchus is not supp orted in any m ost-par-simonious trees. Two seemingly u nique syrin-
geal characters (23 and 31) shared by Ony-

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 6/20
720 [Auk , Vol. 119JEFFREY S. BIRDSLEY
FIG . 4. Strict consensus tree of the restricted Em -
pidonax assemblage. Decay indices are shown when
greater than 1.
FIG . 5 . A d a m s co n se n su s t r e e d e r iv e d f r om a ll
most-parsimonious trees. The taxon at the bottom
denotes the assemblage shown in Figure 6.
chorhynchus an d Platyrinchus m ay be derived
independ ently in these two taxa. A sister rela-
tionship between Rhynchocyclus and Tolmomyias
is supp orted in all most-parsimonious trees by
the presence of narrow, linear internal syrin-
geal cartilages (character 29), a retort-shap ed
pend ant nest (character 65) wh ich is u sed as a
dormitory (character 67). In all most-parsimo-
nious trees this relationship is ad ditionally
sup p orted by at least two of the follow ing char-
acters: 1, 11, 43, 49, 64.
Lanyon (1988c) could not determine the re-
lationships of Cnipodectes w i t h in t h e a ss em -
blage, m aintaining it in a polytom y with his
flatbill clade and tody-tyrant clade. My resu lts
suggest that Cnipodectes is the sister group to a
clade com posed of Onychorhynchus and Myio-bius (including erythrurus). This r elationsh ip is
supp orted in all m ost-parsim onious trees by
th e pr esence of long r ictal bristles (cha racter 57)
and ru fou s tails (char acter 53; lost with in Myio-
bius). I discuss the hy p othesized sister relation-
ship between Onychorhynchus and Myiobius be-
low with the Empidonax assemblage.
Lanyon (1988c) defined a tody-tyrant g roup
(Todirostrum , Poecilotriccus [including Taenio-
triccus], Hemitriccus [including Myiornis], Lop-
hotriccus [including Atalotriccus], and Oncosto-
ma) based on th e p ossession of two characters:
1) a horseshoe-shap ed, cartilaginous bronchial
plate, and 2) delicate, rod -like intern al cartilag-
es located near the caudal ends of the horse-
shoe. In the present study, I com bined these
into one character state (character 27, state 1)
wh ich sup ports monophyly of Lanyon’s tod y-
tyrant group in som e, but not all m ost-parsi-
m o n io u s t r ee s. La n yo n (19 88c) p r o p o se d a
cl ad e co m p o se d o f Hemitriccus, Lophotriccus,
and Oncostoma based on the pr esence of a pes-
sulus that extends anteriorly to divide four or
more A elements dorsally (character 16). This
clade is sup ported in all m ost-parsim onioustrees by character 16 and, additionally in some
most-parsimonious tr ees, by characters 43 and

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 7/20
July 2002] 721Phylogeny of Tyrannidae
FIG . 6. Adams consensus tree of an ‘‘elaeniid and
empid’’ clade.
64. Lophotriccus an d Oncostoma are supported
as sister gr oups by a laterally compressed syr-
inx and trachea (character 26) in all most-par-
simonious trees.
There is some su pp ort for a basal placement
o f t h e fl at b il ls a n d t o d y -t y r an t s w i t h in t h e
Tyrannidae and perhaps w ithin the Tyrannoi-
dea. In all most-parsimonious trees the flatbills
and tody-tyrants are basal to a large clade con-
taining the m ajority of tyrann ids plus, in some
m o s t-p a r s im o n io u s t r ee s, co t in g id s a n d p i -
p r id s . Th i s l a r g e c la d e i s s u p p o r t ed b y t h e
presence of ossified nasal septa (character 1).
This is at least par tly congr uen t with the resu lts
of Sibley and Ahlquist’s (1985) DNA–DNA hy-
bridization analyses in that the tody-tyrants To-
dirostrum and Hemitriccus were among th e taxa
they placed in their family ‘‘Mionectidae’’ as
the sister group to the rest of the tyrannoids
(cotingids piprids remaining tyrannids).Kingbird assemblage. Resu lt s su p p or t a
kingbird assem blage com pletely consistent
with that of Lanyon (1984) (Fig. 3). It is sup-
ported in all most-parsimonious tr ees by char-
acters 3, 9, 10, 44, and 55. My hyp othesis of re-
lationships within this assemblage is, however,
less well resolved than Lanyon’s (1984). This is
due to the fact that Lanyon (1984) used alter-native d erived states of the ventral connection
of the B 1 and B 2 syringeal support elem ents
(ch a r act er 19) t o s u p p o r t t w o m a jo r cl ad e s
within the assemblage: Phelpsia, Pitangus, Phi-
lohydor , Legatus, and Myiozetetes have flattened
connections (state 1) w hereas the rem aining
seven genera in the assemblage have rounded
connections (state 2). My family-wide analysis
indicates that flattened connections, which oc-
cur in n early all tyrann ids, are ancestral in the
kingbird assemblage and thu s do not argu e forrelationships within it. My results do not sup-
p ort m onoph yly of Lanyon’s clade comp osed of
the five g enera named above. However, round -
ed connections (state 2) app ear to be u nique to
Myiodynastes, Conopias, Megarynchus, Tyrannop-
sis, Tyrannus, Empidonomus, and Griseotyrannus.
This clade is supported in all m ost-parsim o-
nious trees by characters 19 and 32. Lanyon’s
hypothesized relationships am ong these gen-
era are also sup ported in all m ost-parsim oni-
ous trees. Character 62 sup ports a sister-group
relationship between Myiodynastes and Cono-
pias. Characters 20, 42, and 61 supp ort a clade
composed of Megarynchus, Tyrannopsis, Tyran-
nu s, Empidonomus, and Griseotyrannus. Charac-
ter 32 supports a sister-group relationship be-
tween Megarynchus and Tyrannopsis. Character
59 sup ports a clade composed of Tyrannus, Em-
pidonomus, and Griseotyrannus.
Character 11 supp orts sister-group relation-
ships between Pitangus and Philohydor (two or
more double, complete, ossified A elements)
and between Legatus an d Myiozetetes (incom-plete A3 elements fused to complete A2). Char-
acter 45 (intrinsic syringeal muscles Mm. obli-
qui laterales) s u p p o r t s t h e Legatus– Myiozetetes
relationship in some most-parsimonious trees,
but in the rem ainder of trees, this is ambiguous
due to uncertainty about the state of this char-
acter in Phelpsia.
Empidonax assemblage. A restricted Empido-
nax assemblage (Fig. 4) is supported contain-
ing the same genera that Lanyon (1986, 1988b)
placed in it except for the four g enera that com-p rise h is Myiophobus group: Myiophobus,
Pyrrhomyias, Hirundinea, a n d Myiobius. Seven

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 8/20
722 [Auk , Vol. 119JEFFREY S. BIRDSLEY
extra steps are requ ired to force a monop hyletic
Empidonax assemblage including the Myiopho-
bus group. However, such trees are not signifi-
cantly different from most-parsimonious trees
(P 0.36; 14 N 26 char acters requiring d if-
fering nu mb ers of steps on the trees being com-p a r e d ). In a ll m o s t-p a r s im o n io u s t r ee s t h e
monophyly of the restricted Empidonax assem-
blage is supported by the presence of a nasal
septum with basal trabecular plate and ante-
rior notch (character 4), a wide posterior fork
on the trabecular plate (character 7), and ‘‘near-
ground generalist’’ foraging behavior (charac-
ter 61). In all m ost-parsim onious trees, the
‘‘ Myiophobus group’’ genera are distantly relat-
ed to th is restricted Empidonax assemblage.
Within the Empidonax assemblage, these re-sults sup port Lanyon’s (1986) hypoth esis of a
basal position of the chat-tyran ts Ochthoeca, Sil-
vicultrix and Colorhamphus. These resu lts do not
supp ort the m onophyly of Lanyon’s (1986)
Ochthoeca group composed of these three gen-
era plus Arundinicola, Fluvicola, and Alectrurus.
Lanyon (1986) used char acter 11, state 0 (no A
elements completely encircle the bronchi) to
s u p p o r t h i s Ochthoeca group, but m y fam ily-
wide analysis indicates that th is state is ances-
tral in the Empidonax assemblage and does not
argue for a close relationship of these taxa.
The m onophyly of a large clade ( Alectrurus
through Lathrotriccus in Fig. 4) comp osed of all
of the restricted Empidonax assemblage genera
except Ochthoeca, Silvicultrix and Colorhamphus
is supp orted in all most-parsimonious trees by
characters 22, 49, or both. Within this large
clade, a group (Cnemotriccus to Lathrotriccus in
Fig. 4) similar to Lanyon’s (1986) Empidonax
group is supported in all m ost-parsim onious
trees by character 49, and additionally in some
of those trees by characters 13, 22, and 61. Sa- yornis, which possesses the uniquely modified
syringeal A2 elem ents (character 13) of this
group is, in some most-parsimonious trees, the
basal member of the group. In other m ost-par-
simonious t rees, however, Sayornis groups with
other taxa that lack wing bars (character 49)
and are ‘‘near-ground generalist’’ foragers
(character 61). In all most-parsimonious trees,
characters 24, 25, and 52 supp ort Gubernetes
and Muscipipra as sister group s.
Few a d d i t io n a l r el at io n sh i p s a r e w e ll r e -solved in the strict consensus of this assem -
blage (Fig. 4), but up on examination of the set
of m ost-parsim onious trees and of the Adam s
consensus (Fig. 6), some patterns are evident.
For example, all most-parsimonious trees con-
tain a clade corresp ond ing rou ghly to Lanyon’s
(1986) Muscisaxicola g r ou p co n ta in in g t h e
ground-tyrants, shrike-tyrants, bush-tyrantsand allies. Hymenops, which shares some plum -
age and habitat characters with this group, is,
in some most-parsimonious trees, placed w ith-
in this clade ( Muscisaxicola through Cnemarchus
in Fig. 6); hence, a Muscisaxicola g r ou p i s n o t
depicted in the strict consensus (Fig. 4). This
group, with or without Hymenops, is supp orted
by characters 60 (habitation of open grassland
or m arsh) and 61 (ground specialist foraging
behavior). In a greemen t w ith Lanyon’s (1986)
concept of this group, Muscisaxicola always isplaced basally, as illustrated in Figure 6. Pat-
terned wings (character 48) and notched or at-
t en u a te d p r i m a rie s (ch a r act er 59) s u p p o r t
mon ophy ly of the rest of the grou p exclusive of
Muscisaxicola. ‘‘Perch-to-grou nd ’’ foraging be-
havior (character 61) supports a group exclu-
sive of Muscisaxicola an d Lessonia.
A clade composed of Agriornis, Myiotheretes,
Polioxolmis and Cnemarchus is supported in all
m ost-parsim onious trees by the presence of
streaked throat plum age (character 50). Lan-
yon’s (1986) hyp othesized bush-tyrant clade
composed of Myiotheretes, Polioxolmis and Cne-
marchus is supported in all most-parsimonious
trees by an ossified interorbital septum (char-
acter 9) and a transition from open grassland
to forest–wood land habitat (char acter 60). Lan-
yon (1986) was unable to resolve relationships
am ong these genera, but these results supp ort
Polioxolmis an d Cnemarchus a s s is t er g r ou p s
based on the relatively narrow dorsal ends of
their B1 syringeal su pp ort elements (character
22, state 0), and the p resence of well developed Mm. obliqui ventrales (char acter 44). Lanyon
(1988a) used char acter 44 in the Elaenia assem-
blage but app arently did not r ecognize it in the
Empidonax assemblage. Lanyon (1986) hypoth-
esized a clade composed of Arundinicola, Flu-
vicola, Alectrurus su pp orted by char acters 8, 56,
and 60. In the present study, these characters
support such a clade in some most-parsimoni-
ous trees. The remainder of most-parsimonious
t r ee s h y p o t he si ze a n ( Arundinicola (Fluvicola
( Alectrurus, Hymenops))) cl ad e. Hymenopsshares plum age and habitat characters with the
other three genera, but Lanyon (1986) placed

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 9/20
July 2002] 723Phylogeny of Tyrannidae
Hymenops in his Knipolegus group w ith wh ich it
also shares plumage and syringeal characters.
Arundinicola, Fluvicola, and Alectrurus, at least,
ap pea r to be closely related, as dep icted in Fig-
ure 6.
The d ouble, comp lete, ossified syr ingeal A el-em ents (character 11) and concealed crown
patch (character 46) present in Lanyon’s (1986,
1988b) Myiophobus group are not present in the
rest of the Empidonax assemblage and argue for
the p lacement of these genera elsewhere. In th e
set of m ost-parsim onious trees, Myiophobus,
Pyrrhomyias, a n d Hirundinea a r e a ll ie d w i t h
several different group s outside th e Empidonax
assem blage. A sister-group relationship be-
tween Myiobius (including erythrurus) and Ony-
chorhynchus is supp orted by the presence of atleast two d ouble, complete, ossified syrin geal A
elements (character 11) and an ossified inter-
orbital septum (character 9). Myiobius also has
the long r ictal br istles (char acter 57) and eryth-
rurus has th e ru fous tail (char acter 53) foun d in
Onychorhynchus and Cnipodectes. Past workers
(Ames 1971, Traylor 1977, Traylor and Fitzpat-
rick 1982) have hypothesized a close associa-
tion between Myiobius and Onychorhynchus
based on these syringeal and plumage charac-
ters. Myiobius builds a pendant, enclosed nest
(character 64) as do Onychorhynchus and Cni-
podectes, but in the present analysis this char-
acter does not su pp ort the relationship of Myio-
bius to th ese genera because the globular nests
(state 1) of Myiobius and elong ate nests (state 2)
of Onychorhynchus and Cnipodectes were coded
as alternative u nord ered st ates. Characters 1, 4,
19, and, in some most-parsimonious trees, 64
s u p p o r t Myiobius (formerly Terenotriccus ) eryth-
rurus as sister group to the other Myiobius spe-
cies. This is congr uent w ith Lanyon’s (1988b)
merger of erythrurus into Myiobius. Characters40, 48, and, in some m ost-parsimonious trees,
22 supp ort Pyrrhomyias an d Hirundinea as sister
g r ou p s in a g re em en t w i th La ny on (1986,
1988b). Neopipo i s s u p p o r t ed a s t h ei r s is t er
grou p by characters 11, 43, and 54. This agrees
w ith on e of Mobley an d Pru m’s (1995) hypot h-
eses. However, cran ial data for Neopipo are still
needed to evaluate th is hypothesis.
The nasa l septa of ’’ Myiophobus’’ (lintoni, och-
raceiventris, phoenicomitra, a n d roraimae) h a v e
trabecular plates that are elevated above thebase of the septum (character 4, state 2) which
caused Lanyon (1986, 1988a) to remove th em
from Myiophobus and place them in the Elaenia
assemblage. He refrained from creating new
generic names u ntil more comparative data are
available. These results do not give strong sup-
port for the removal of lintoni an d ochraceiven-
tris. Myiophobus, now includin g only the sp eciescryptoxanthus, fasciatus, flavicans, inornatus, and
pulcher , is s u p p o rt ed a s t h e s is ter g r ou p t o
’’ Myiophobus’’ lintoni an d ochraceiventris in
some m ost-parsimonious trees by character 49
and , add itionally in some trees by 17. However,
in no m ost-parsim onious trees are ’’ Myioph-
obus’’ phoenicomitra and roraimae closely related
to other Myiophobus. These results supp ort Lan-
yon’s (1986, 1988a) conclusion that Myiophobus
as trad itionally d elimited (Traylor 1979, Sibley
and Monroe 1990) is not monophyletic, with atleast phoenicomitra an d roraimae belonging else-
wh ere. In all most-parsimonious trees, ’’ Myio-
phobus’’ phoenicomitra and roraimae are the sister
group of a clade composed of Machetornis and
Muscigralla. The relationship is supported by
char acters 8, 9, or both. The interorbital sep tu m
(character 9) requires further study in these
taxa, particularly in Machetornis for w hich I fol-
lowed Warter (1965) and coded as having two
fenestrae open (state 0). The one specimen of
Machetornis examined (AMNH 6657), however,
had the infraorbital fenestra enlarged and su-
praorbital fenestra reduced (state 1), suggest-
ing some unknown degree of variation.
My iarchus assemblage. Th e m o no p hy ly o f
Lanyon’s (1985) Myiarchus assem blage is sup -
ported in some, but n ot all, most-parsimonious
trees. This clade is supported by the presence
of cavity nesting (character 62). A restricted as-
semblage composed only of Rhytipterna, Casior-
nis, Sirystes, and Myiarchus is supported in all
most-parsimonious trees by th e pr esence of in-
ternal cartilages that are robust ‘‘J’’ or ‘‘L’’shaped (character 32) and are attached to the
ventral side of the tracheobronchial junction
(character 39). In som e m ost-parsim onious
trees it is additionally su pp orted by an ossified
interorbital septum (character 9). A Deltarhyn-
chus– Ramphotrigon cla d e is s u p p o rt ed in a ll
most-parsimonious trees by characters 37 and
49, and , in some most-parsimonious trees, ad-
ditionally by character 11. Lanyon (1985) de-
scribed extrem e sim ilarity am ong the crania
and syringes of Myiarchus, Sirystes, and Casior-nis. He was unable to develop a hypothesis of
their relationships. M y results suggest that

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 10/20
724 [Auk , Vol. 119JEFFREY S. BIRDSLEY
Sirystes and Myiarchus are sister groups. This
relationship is sup ported by the presence of
dor sally divergent B1 and B2 syring eal sup p ort
elements (character 21), a character that Lanyon
(1984) used in the kingbird assemblage but ap-
parently d id not recognize in the m yiarchines. Elaenia assemblage. Th es e r es u lt s d o n ot
support m onophyly of the Elaenia assemblage
(Lanyon 1988a). Seven extra st eps are r equired
to force its monophyly, but such trees are not
significantly different from most-parsimonious
trees (P 0.19; 14 n 25 characters requir-
ing d iffering nu mbers of steps on th e trees be-
ing comp ared). Some m ost-parsimonious tr ees
include a large clade containing m ost of the
genera of the Elaenia assem blage plus the re-
stricted Empidonax assemblage (Fig. 6). In ther em a i nd e r o f m o s t-p a r s im o n io u s t r ee s t h is
clade also includes a group of ten taxa ( Myio-
triccus through Muscigralla in Fig. 5), most of
which Lanyon (1988a) placed in his Elaenia as -
semblage. In all most-parsimonious trees, the
fu s io n o f t r a ch e al A e le m en t s i n to a d r u m
(character 14) sup ports a ‘‘tyrannu let’’ group
(Fig. 6) corresponding roughly to Lanyon’s
(1988a) Elaenia group in his Elaenia assemblage.
I t s m o n o p h y l y i s n o t s u p p o r t e d i n t h e s t r i c t
consensus (Fig. 2) because Inezia, Pseudocolop-
teryx, and Polystictus are includ ed in the grou p
in some most-parsimonious trees. That group
is characterized in all most-parsimonious trees
b y t h e a p p a r en t ly h o m olo g ou s p r e se nce o f
well-developed Mm. obliqui ventrales (character
44), a sup erciliary eye stripe (character 55),
perch-gleaning foraging behavior (character
61), and unmarked eggs (character 68).
These results sup port the monophy ly of sev-
eral of Lanyon’s (1988a) sm aller clades w ithin
the Elaenia assemblage. A clade composed of
Elaenia, Myiopagis, and Tyrannulus is supportedby character 14 plus either 46 or 55. Sister-
group relationships between Stigmatura an d
Pseudelaenia (character 6), between Pseudotric-
cus and Corythopis (char acters 5, 18, 33), be-
tween Serpophaga an d Anairetes (char acter 14
plus either 7 or 43), and between M ionectes an d
Leptopogon (characters 6, 42, 66, and, in some
m ost-parsim onious trees, 68) are supp orted.
Tyranniscus is supported as the sister group to
’’ Mecocerculus’’ calopterus an d minor by charac-
ters 14 and 35 and, in some m ost-parsimonioust r ee s, ch a r act er 6. T h is s u p p o r t s La n yo n’s
(1988a) grouping of these taxa and his conclu-
sion that the genus Mecocerculus as traditionally
delimited (Traylor 1979, Sibley and Monroe
1990) is polyphyletic. In some most-parsimo-
nious trees, ’’ Mecocerculus’’ (hellmayri, poecilo-
cercus, and stictopterus) is the sister group to the
clade com posed of Tyranniscus and ’’ Mecoce-rculus’’ calopterus and minor based on the pres-
ence of an ad d itional, po sterior trabecular plate
i n t h e n a sa l s ep t u m (ch a r act er 6). La n yo n
(1988a) left one species (leucophrys) in Mecocer-
culus. In no m ost-par simoniou s trees is it close-
ly related to the other species traditionally
placed in Mecocerculus.
Additional tyrannid genera. This analysis re-
sulted in hypotheses of relationships for most
of the problematic genera that Lanyon (1986,
1988a) maintained incertae sedis. A clade com-p o s e d o f Tachuris , Phyllomy ias fasciatu s an d P.
griseiceps i s s u p p o r t ed b y t h e l o ss o f t w o d e -
rived characters: the elevated trabecular plate
in the nasal septum (character 4; cranial data
for P. griseiceps lacking) and a Myiarchus-like
configuration of the syringeal B1 and B2 ele-
men ts (char acter 22). They also shar e a derived
syringeal character with Stigmatura an d Pseu-
delaenia: a cartilaginou s bronchial plate with ro-
bust internal cartilages (character 27) w hich
supp orts a clade com posed of these five taxa.
La nyo n (1988a ) t hou g ht t he s yr in g es of
Phyllomyias fasciatus and P. griseiceps to be very
different from each other and speculated that
Phyllomyias (composed of fasciatus, griseiceps,
an d griseocapilla) is polyphyletic. I exam ined
Lanyon’s cleared-and -stained syring eal speci-
m ens (three of fasciatus a n d o n e o f griseiceps)
and found them to be quite similar. The syrinx
of griseiceps gives the im pression of being a
more ossified version of the fasciatus syringes.
Their cartilaginous bronchial p lates are very
similar in detail and the internal cartilages of both species are attached to n arrow caud al ex-
tensions of th e bronchial plates. Besides the d e-
gree of ossification, the only conspicuous dif-
ference between th e syringes of the two sp ecies
is the shorter internal cartilages of griseiceps
which lack the large, am orphous ventral and
caudal extension present in fasciatus. Phyllom-
yias fasciatus and P. griseiceps are supported as
sister grou ps in all most-parsimonious trees by
character 61 (fruit/ up ward hover-glean forag-
ing behavior). I suggest that Phyllomyias fascia-tus and P. griseiceps are indeed closely related,
p ossibly sister sp ecies (pend ing examin ation of

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 11/20
July 2002] 725Phylogeny of Tyrannidae
crania of P. griseiceps an d P. griseocapilla and sy-
ringes of P. griseocapilla).
Machetornis an d Muscigralla are supported as
sister g roup s by three characters: the loss of a
Myiarchus-like configuration of the syringeal
B1 and B2 elements (cha racter 22), ha bitation of open grassland or m arsh (character 60), and
ground specialist foraging behavior (character
61). Colonia and Sublegatus are supp orted as sis-
ter groups by character 55 and, in some most-
parsimonious trees, add itionally by characters
9, 30, and 46.
Homology of nasal septum characters. My re-
sults p rovide a test of the homology of each of
t h e s t at es o f t h e n a sa l s ep t u m t h at La n yo n
(1984, 1985, 1986, 1988a, c) used to define his
assemblages. Lanyon (1988c) used the unossi-fied n asal septum (character 1) as one of three
characters d efining h is flatbill and tody-tyrant
assemblage. When reconstructed on all most-
parsim onious trees, the unossified septum is
homologous in Cnipodectes, Onychorhynchus, To-
dirostrum, Poecilotriccus, Hemitriccus, Lophotric-
cus, a n d Oncostoma, and, in som e m ost-parsi-
monious trees, Rhynchocyclus, Tolmomyias , and
Platyrinchus as well. The unossified septum in
Tachuris , wh ich Lanyon (1988c) d id n ot include
in his flatbill and tody-tyrant assem blage, is
convergent in all most-parsimonious trees. The
unossified septum is symplesiomorphic in all
most-parsimonious trees because, except for
the h ighly ossified n asal capsu les of Dendroco-
laptes, furnarioids have unossified septa. That
character, therefore, does not provide a syna-
pomorp hy for a flatbill and tod y-tyrant assem-
blage. However, informative characters are p o-
t en t ia lly e xi st en t i n t h e m o r p h olo g y o f t h e
ca r t il ag in ou s p a r t s o f t h e n a sa l ca p s u le s o f
these birds. At least som e of the tody-tyrants
probably have cartilaginous trabecular plateswithin their cartilaginous septa as suggested
by th e ossification of those stru ctures in occa-
sional specimens (Lanyon 1988c).
Lanyon (1984) defined his kingbird assem-
blage par tly by the presence of an ossified n asal
septum that lacks both a trabecular p late and
an internal support rod. In all m ost-parsim o-
n io u s t r ee s, a r ed u c ed i n te rn a l s u p p o r t r o d
(character 3) is hom ologous in all kingbird as-
semblage taxa. In all m ost-parsimonious trees,
character 3 is convergent in Attila, in Colonia,and in Machetornis. The absence of a trabecular
plate (character 4) is symplesiomorphic in all
most-parsimonious trees and does not provide
a synap omorphy of the kingbird assemblage.
Lanyon (1985) defined his Myiarchus assem-
blage (without Attila) partly by th e presence of
an ossified nasal septum with a conspicuous
internal sup port rod (character 3). Most tyran-nids possess this character. My fam ily-wide
analysis indicates that it is symplesiomorphic
and does not provide a synapom orphy of the
Myiarchus assemblage. My results indicate that
there are no nasal septum characters sup port-
ing this group, with or w ithout Attila. Lanyon
(1985) recogn ized that these gen era (except At -
tila) h ave ossified nasal capsu les (character 8)
wh ich su rround the septum , but he did not ex-
plicitly include this in his list of characters. In
a ll m o s t-p a r s im o n io u s t r ee s i n t h e p r e se n tanalysis, character 8 is hom ologous in Del-
tarhynchus, Ramphotrigon, Rhytipterna, Sirystes,
Casiornis, and Myiarchus, though in some most-
parsimonious trees it is not a synapom orphy. I
examined the same four crania of Attila spadi-
ceus a n d t w o o f A. cinnamomeus that Lanyon
(1985) did and, contrary to his report, found
o n e ( A. cinnamomeus, A M N H 11616) w i t h a
com pletely ossified nasal capsule. A larger
sample of Attila crania may show a significant
tendency for nasal capsules to ossify and thus
may strengthen the evidence of a relationship
between Attila and the Myiarchus assemblage.
Lanyon (1986) defined his Empidonax assem-
blage solely by the p resence of an ossified n asal
septum with basal trabecular plate and ante-
rior notch (character 4) and posterior forking
(character 7). In all most-parsimonious trees, a
basal trabecular plate and anterior notch is ho-
mologous in all genera in the restricted Empi-
donax assemblage (Fig. 4). In all m ost-parsi-
monious trees, this character is convergent in
Myiobius a nd in t he Pyrrhomyias– Hirundineaclade. This type of nasal septum is also present
in Myiophobus (composed of cryptoxanthus, fas-
ciatus, flavicans, inornatus and pulcher ) and is, in
some most-parsimonious trees, hypothesized
t o b e h o m o l o g o u s w i t h t h o s e i n Pyrrhomyias
an d Hirundinea. Lanyon (1988a) described the
surp rising app earance of this type of nasal sep-
tum in six species of Elaenia, providing further
evidence that this seemingly unique character
has arisen convergently. In these Elaenia spe-
cies and in Pyrrhomyias, Hirundinea, and Myio-bius, the trabecular plate is not conspicuously
forked posteriorly (character 7) as it is in the re-

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 12/20
726 [Auk , Vol. 119JEFFREY S. BIRDSLEY
stricted Empidonax assem blage and in Myiop-
hobus. In all m ost-parsim onious trees, a wide
posterior fork is homologous in all genera in
the restricted Empidonax assemblage (except
lost in Cnemarchus). In all most-parsimonious
trees it is convergent in Myiophobus. Nasal sep tawith basal trabecular plates and anterior notch-
es (character 4) and wide posterior forking
(character 7) app ear to b e synap omorphies of a
restricted Empidonax assemblage.
Lanyon (1988a) defined his Elaenia assem-
blage solely by the p resence of an ossified n asal
septum with a trabecular plate elevated above
the ventral edge of the septum (character 4). In
all most-parsimonious trees, an elevated tr a-
becular plate is hom ologous in all genera of
‘‘tyrannulets’’ and allies (Pseudotriccus throughTyranniscus in Fig. 6). I t is also hom ologous
w i t h in a s m a lle r g r o u p ( Myiotriccus through
Muscigralla in Fig. 5). In all most-p arsimon ious
trees, the elevated trabecular plate is a syna-
pom orphy of a clade containing som e or all
Elaenia assem blage genera and the restricted
Empidonax assemblage g enera. In all most-par-
simonious trees, the basal p lates of the restrict-
ed Empidonax assemblage are hypothesized to
have evolved from elevated plates. In all most-
parsimonious t rees, the narrow, slender p oste-
rior forks (char acter 7) found in several Elaenia
assemblage genera arise two or three indep en-
dent times, but are always homologous in Cap-
siempis, Phaeomyias, and Nesotriccus, and in Ser-
pophaga and Anairetes, a n d i n Pseudocolopteryx
and Polystictus. Lanyon (1988a) examined one
cranium each of Phyllomyias fasciatus and Tachu-
ris and both lacked trabecular plates (the sep-
t u m o f Tachuris was simply unossified). In all
m o s t-p a r s im o n io u s t r ee s, t h es e a r e r e co n -
structed as a single derived loss of ossified, el-
evated trabecular plates. Levels of homoplasy in character suit es. Cra-
nial characters (m ean RC 0.39), syring eal
characters (mean RC 0.47), p luma ge char ac-
ters (mean RC 0.33), and behavioral and eco-
logical characters (mean RC 0.52) d id n ot d if-
fer significantly in levels of homop lasy (P
0.57). The b ehavioral and ecological char acters
in th is stud y contained a level of phylogenetic
information sim ilar to those t hat Hu ghes (1996)
used for the Cuculidae. These results are in ac-
c o r d w i t h a g r o w i n g n u m b e r o f s t u d i e s t h a tfind significant phylogenetic structure in be-
havioral and ecological characters (e.g. Pru m
1990a, Winkler and Sheldon 1993, Paterson et
al. 1995, Hu ghes 1996, McLenn an and Mattern
2001). Amon g th e su ites of morph ological char-
acters, plumage characters exhibited the most
homoplasy, though not significantly so. Only
one plum age character—long rictal bristles(char acter 57)—exhibited n o hom oplasy on an y
most-parsimonious tree. A pointed crest (char-
acter 58) exhibited no homoplasy on some, but
not all most-parsimonious trees. The ten plum -
age characters that d escribe color patterns tend
to be m ore hom oplasious (m ean R C 0.26)
than those that describe feather form, but this
is still not significantly different from other
character suites (P 0.36). Omland and Lan-
yon (2000) found similarly high levels of ho-
moplasy in plumage characters in orioles andcautioned against heavy use of such characters
in phylogenetic analysis. It app ears likely that
convergence, parallelism , and reversal have
been common in the evolution of avian plum-
age color p atterns, perh aps du e to evolutionary
conservatism of the u nderlying d evelopm ental
pathways coupled with lability of the on–off
switches of those pathways (Price and Pavelka
1996).
Summary. I p erformed a cladistic analysis
of all cur rently available morp hological and be-
havioral data for the Tyrannidae. Most charac-
ters were drawn from Lanyon (1984, 1985,
1986, 1988a, b, c). My an alysis d iffers from Lan-
yon’s in five imp ortan t ways: first, I ad ded sev-
eral characters. Second, I examined furnarioid
o u t gr ou p s i n a d d i t io n t o t h e t y r a n n oid o u t-
groups Lanyon considered. Third, I assessed
character states across all tyran nid genera and
in outgroups, allowing simultaneous analysis
of all taxa and tests of monoph yly of the Tyran-
nid ae and of each of Lanyon’s prop osed assem -
blages. Fourth, I gave characters equal weight,allowing tests of the homology of cranial and
syringeal characters that Lanyon used to define
g r ou p s . Fift h , I w or ke d w i th in a cla d is tic
framework that explicitly roots trees to an out-
g r ou p . Th i s e n su r e s p l es io m or p h i es a r e n o t
used as character supp ort for clades.
My r e su l t s p r o vid e e qu i vo ca l s u p p o r t fo r
monophyly of the Tyrann idae. An equally par-
simonious hypothesis su ggests that th e flatbill
and tody-tyrant genera are m ore distantly re-
lated to other tyrann ids than are cotingids andp i p r id s . R es u l ts s u p p o r t m o n op h y ly o f t h e
kingbird assemblage, restricted Myiarchus and

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 13/20
July 2002] 727Phylogeny of Tyrannidae
Empidonax assemblages, an d several lower-lev-
el group ings. Monophyly of neither the Elaenia
nor the flatbill and tody-tyrant assem blage is
supp orted. Nasal septu m characters th at past
workers have considered conservative and ph y-
lo g en e tica lly i n fo r m at iv e a r e s u p p o r t e d a ssynapom orphies of the kingbird assem blage
and of a restricted Empidonax assemblage. A
nasal capsule character provides equivocal
s u p p o r t f o r a Myiarchus assemblage without
Attila. M an y n o d es h av e m i n im a l s u p p o rt .
Much more data are needed to resolve relation-
ships in the Tyrannidae, especially at higher
levels. In accord with previous studies, plum-
age coloration patterns exhibited relatively
high levels of homoplasy.
ACKNOWLEDGMENTS
This paper is based on a dissertation submitted in
par tial fulfillment of the requiremen ts for a Ph.D. in
the Departm ent of Zoology, Michigan State Univer-
sity. I thank D. Straney, R. Prum , W. Lanyon, D.
Brooks, R. Leschen, M. Donoghue, J. White, and T.
Getty for helpful d iscussion and encouragement.
This paper benefited from comments by B. Livezey,
R. Zink, and an anonymou s reviewer. I thank G. Bar-
rowclough for access to th e collections at the Am er-
ican Museu m of Natural H istory. This work was sup -
ported by the W. K. Kellogg Biological Station
Gradu ate Research Training Group (RTG) funded by
NSF DIR-09113598, by a Michigan State University
College of Natural Science Doctoral Recruiting Fel-
low s h ip , b y t h e F r an k M . C h a p m a n Fu n d o f t h e
A m e r ic an M u se u m o f N a t u r a l H i s t or y, a n d b y a
Grant-in-Aid-of-Research from Sigma Xi. This is
contribution no. 868 of the W. K. Kellogg Biological
Station.
LITERATURE CITED
ADAM S, E. N. 1972. Consensus techniques and the
comparison of taxonomic trees. Systematic Zo-
ology 21:390–397.
AMERICAN O RNITHOLOGISTS’ UN I O N . 1998. Check-
list of North American Birds, 7th ed. American
Ornithologists’ Union, Washington, D.C.
AME S, P. L. 1971. The morphology of the syrinx in
passerine birds. Bulletin of the Peabody Muse-
um of Natural H istory, no. 37.
BAUM E L, J. J. 1979. Ost eologia. Pag es 53–121 in No-
min a An atomica Avium (J. J. Baum el, A. S. King,
A. M. Lucas, J. E. Breazile, and H. E. Evans,
Eds.). Academic Press, London.BOSTWICK, K. S. 2000. Display behaviors, m echanical
sounds, an d evolutionary relationships of the
C lu b - w in g e d M a n a ki n ( M achaeropterus delicio-
sus). Auk 117:465–478.
BREMER, K. 1988. The limits of amino acid sequence
data in angiosperm phylogenetic reconstruction.
Evolution 42:795–803.
DEQ UEIROZ , A., AN D P . H . WIMBERGER. 1993. The
usefulness of behavior for phylogeny estima-
tion—levels of homoplasy in behavioral and
mor ph ological characters. Evolution 47:46–60.
FELSENSTEIN , J. 1982. Numerical methods for infer-
ring evolutionary trees. Quarterly Review of Bi-
ology 57:379–404.
FITZPATRICK, J. W. 1980. Foraging behavior of Neo-
tropical tyrant flycatchers. Condor 82:43–57.
GRIFFITHS, C. S. 1994. Monophyly of the Falconifor-
mes based on syringeal m orphology. Auk 111:
787–805.
H ILTY, S. L., AN D W. L. BROWN . 1986. A Guide to the
Birds of Colombia. Princeton University Press,Princeton, New Jersey.
H UGHE S, J. M. 1996. Phylogenetic an alysis of the Cu -
culidae (Aves: Cuculiformes) using behavioral
and ecological char acters. Auk 113:10–22.
KALLERSJO , M., J. S. FARRIS, A . G . KLUGE, AN D C.
BUL T. 1992. Skewness an d permu tation. Cladis-
tics 8:275–288.
KIN G, A. S. 1979. System a r espirat orium . Pages 227–
265 in Nomina Anatomica Avium (J. J. Baum el,
A. S. King, A. M . Lucas, J. E. Breaz ile, and H. E.
Evans, Eds.). Academic Press, London.
LANYON , S. M. 1985. Molecular perspective on high-
er-level relationships in the Tyrannoidea (Aves).
Systematic Zoology 34:404–418.
LANYON , W. E. 1984. A phylogeny of the kingbirds
and their allies. American Museum Novitates,
no. 2797.
LANYON , W. E. 1985. A phylogeny of the myiarchine
flycatchers. Ornithological Monographs 36:361–
380.
LANYON , W. E. 1986. A p hylogeny of the th irty-thr ee
g e n e r a i n t h e Empidonax assemblage of tyrant
flycatchers. American Museum Novitates, no.
2846.
LANYON
, W. E. 1988a. A phylogeny of the thirty-twogenera in the Elaenia assemblage of tyrant fly-
catchers. Am erican Museu m N ovitates, no. 2914.
LANYON , W. E. 1988b. The phylogenetic affinities of
the flycatcher genera Myiobius D a r w i n a n d Te-
renotriccus Ridgeway. American Museum Novi-
tates, n o. 2915.
LANYON , W. E. 1988c. A p hylogeny of the flatbill and
tody-tyrant assemblage of tyrant flycatchers.
American Museum Novitates, no. 2923.
LEE, P. L. M., D. H. C LAYTON , R. GRIFFITHS, AN D R.
D. M. P A GE . 1996. Does behavior reflect phylog-
eny in swiftlets (Aves: Apodidae)? A test u sing
cytochrome b mitochondrial DNA sequences.Proceedings of the National Academy of Scienc-
es USA 93:7091–7096.

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 14/20
728 [Auk , Vol. 119JEFFREY S. BIRDSLEY
MADDISON , W. P., AN D D. R. MADDISON . 1992.
MacClade: Analysis of Phylogeny and Character
Evolution , version 3.04. Sinau er A ssociates, Sun -
derland, Massachusetts.
MAR I NI, M. A. 1992. Foraging behavior and diet of
the Helmeted Manakin. Condor 94:151–158.
MCKITRICK, M. C. 1985. Monophyly of th e Tyrann i-
d a e (Av es ): C om p a r i so n o f m o r p h o lo g y a n d
DNA. Systematic Z oology 34:35–45.
MCLE N N A N , D. A., AN D M. Y. M ATTERN . 2001. The
phylogeny of Gasterosteidae: combining beh av-
ioral and morp hological d ata sets. Cladistics 17:
11–27.
MOBLEY, J. A., AN D R. O. PRU M. 1995. Phylogenetic
relationships of the Cinnamon Tyrant, Neopipo
cinnamomea, to the tyrant flycatchers (Tyranni-
dae). Condor 97:650–662.
O M L AND , K. E., AN D S . M . LANYON . 2000. Recon-
structing plumage evolution in orioles ( Icterus):Repeated convergence and reversal in patterns.
Evolution 54:2119–2133.
PATERSON , A. M., G. P. W ALLIS, A N D R. D. GRA Y.
1995. Penguins, petrels, and parsimony: Does
cladistic analysis of behavior reflect seabird
phylogeny? Evolution 49:974–989.
PRICE, T., AN D M. PAVELKA. 1996. Evolution of a col-
our pattern: H istory, development, and selec-
tion. Jour nal of Evolutiona ry Biology 9:451–470.
PRU M, R. O. 1990a. Phylogenetic analysis of the evo-
lution of d isplay behavior in the Neotropical
man akins (Aves: Pipridae). Ethology 84:202–231.
PRU M, R. O. 1990b. A test of the monophyly of the
man akins (Pipridae) and of the cotingas (Cotin-
gidae) based on morp hology. Occasional Papers
of the Museum of Zoology, University of Mich-
igan, no. 723.
PRU M, R. O. 1992. Syrin geal m orp hology, phylogeny,
a n d e vo lu t io n o f t h e N e ot r o p ic al m a n a k in s
(Aves: Pipridae). American Mu seum Novitates,
no. 3043.
PRU M, R. O. 1994. Phylogenetic analysis of the evo-
lution of alternative social behavior in the m an-
akins (Aves: Pipridae). Evolution 48:1657–1675.
PRU M, R. O. 1997. Phylogen etic tests of alter native in-
tersexual selection mechanisms: Trait macro-
evolution in a polygynous clade (Aves: Pipri-
da e). Amer ican N atu ralist 149:668–692.
PRU M, R. O., A N D W. E. LANYON . 1989. Monophyly
and phylogeny of the Schiffornis group (Tyran-
noid ea). Con dor 91:444–461.
RIDGELY, R. S., AN D G. TUDOR. 1994. The Birds of
South Am erica, vol. 2. The Subo scine Passerines.
University of Texas Press, Austin.
SIBLEY, C. G., AN D J. E. AHLQUIST. 1985. Phylogeny
and classification of New World suboscine pas-serine birds (Passeriformes: Oligomyod i: Tyran-
nid es). Ornith ological Monograp hs 36:396–428.
SIBLEY, C. G., AN D B. L. MO N R O E, JR. 1990. Distri-
bution and Taxonomy of Bird s of the World. Yale
University Press, New Haven, Connecticut.
SKUTCH , A. F. 1960. Life histories of Central Ameri-
can birds II. Cooper Ornithological Society,
Berkeley, California.
SN O W, D. W. 1982. The Cotingas. Cornell University
Press, Ith aca, New York.
SORENSON , M. D. 1996. TREEROT. University of
Michigan, An n Arbor.
STILES, F. G., AN D A. F. SKUTCH . 1989. A Guide to the
Birds of Costa Rica. Cornell University Press,
Itha ca, New York.
SWOFFORD, D. L. 1991. When are phylogeny esti-
mates from molecular and morphological data
incongru ent? Pages 295–333 in Phylogenetic
Analysis of DNA Sequences (M.M. Miyamoto
and J. Cracraft, Eds.). Oxford University Press,
New York .SWOFFORD, D. L. 2000. PAUP*. Phy logen etic An alysis
Using Parsimony (*and Other Methods), version
4. Sinauer Associates, Sun derland , Massachu setts.
TEMPLETON , A. R. 1983. Phylogenetic inference from
restriction endonuclease cleavage site maps with
particular reference to the evolution of humans
and the ap es. Evolution 37:221–244.
TRAYLOR, M. A., JR. 1977. A classification of the ty-
rant flycatchers (Tyrannidae). Bulletin of the
Museum of Comparative Zoology 148:129–184.
TRAYLOR, M. A., JR. 1979. Family Tyrannidae. Pages
1–245 in Check-list of Birds of the World, vol.
VIII (M. A. Traylor, Jr., Ed.). Museu m of Com-
parative Zoology, Cam bridge, Massachu setts.
TRAYLOR, M. A., JR., AN D J. W. FITZPATRICK. 1982. A
survey of th e tyra nt flycatchers. Living Bird 19:
7–50.
W ARTER, S. L. 1965. The cranial osteology of the New
World Tyrannoidea and its taxonomic implica-
tions. Ph.D. dissertation, Louisiana State Uni-
versity, Baton Rouge.
W INKLER, D. W., AN D F. H. SHELDON . 1993. Evolut ion
of nest construction in swa llows (Hirun dinidae):
A molecular phylogenetic perspective. Proceed-
ings of the National Academy of Sciences USA90:5705–5707.
Associate Editor: R. Zink
A PPENDIX 1. Descriptions of characters used in an al-
ysis. Characters 6, 11, and 13 are ordered . Character
11 is ordered by a character state tree d escribed in
Figure 1. The remaining characters are unordered.
Cranial
Nomenclature follows Baumel (1979) when possi-ble. Otherwise, it follows Lanyon (1984, 1985, 1986,
1988a, b, c).

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 15/20
July 2002] 729Phylogeny of Tyrannidae
1. Septum nasale.—(0) Ossified to varying degrees;
(1) unossified in the flatbill and tody-tyrant assem-
blage (Lanyon 1988c), in Tachuris (Lanyon 1988a),
and in most furnarioids (pers. obs.). In these taxa
there is usually just a shallow dorsal remnant of a
septum in cleaned museum specimens.
2. A nterior segment of septum nasale osseum .—(0)
Well ossified a nterior to the intern al sup port rod ; (1)
poorly ossified anterior to sup port rod.
3. Internal support rod .—(0) Conspicuous internal sup-
port r od w ithin the septum nasale osseum; (1) no con-
spicuous internal support rod. In the kingbird assem-
blage (Lanyon 1984) and in the genera Attila, Colonia,
an d Machetornis (J. S. Birdsley pers. obs.), there is only
a pattern or grain running anterodorsally to postero-
ventrally on the surface of the septum in the position
w h er e t he r od is lo cat ed in t he sep ta of o th er
tyrannoids.
4. Transverse trabecular plate wit hin septu m nasal os-seum.—(0) No tran sverse trabecu lar plate; (1) a tran s-
verse trabecular plate is at the base (ventral edg e) of
the septu m an d th ere is a distinctive notch in the sep-
tum anterior to the trabecular plate; (2) a transverse
trabecular p late is elevated above the ventral edge of
the septu m, so that there appear s to be a mid-sagittal
r id g e o n t h e v en t r a l s i d e o f t h e t r a be cu l a r p l at e.
There is no anterior notch associated with this type
of trabecular plate; (3) a transverse trabecular plate
is barely elevated above the ventral edge of the sep-
tum and occupies only a very short segment of the
anterior part of the septum. It is very broad, trun-
cated posteriorly, and has no anterior notch. Present
in Colonia and hypothesized by Lanyon (1986) to be
unique among tyrannids.
5. Trabecular plate reduced , anterior .—(0) The elevat-
ed type trabecular plate (if present) is relatively
broad an d occupies the length of the septum anterior
to the internal supp ort rod ; (1) the elevated trabec-
ular plate is narrow and occupies only a small part
of the most anterior section of the septum.
6. Trabecular plate divided .—(0) The tr abecular plate
(if present) exists only anterior to the internal sup-
port rod; (1) the elevated typ e trabecular p late is di-
vided into a small section posterior to the internalsupport rod and larger section anterior to the rod.
Both sections ar e relatively slim; (2) the elevated tra -
becular plate is divided into a bu lbous posterior sec-
tion and a th in an terior section; (3) the elevated tr a-
becular plate is divided into bulbous posterior and
bulbous an terior sections.
7. Trabecular plate forked posteriorly.—(0) Trabecu lar
plate (if present) is truncate or tapered posteriorly;
(1) trab ecular p late is w idely forked p osteriorly (Lan-
yon 1986, 1988b). (2) trabecular plate is narrow ly
forked posteriorly, ap pearing as a slender tu ning
fork (Lanyon 1988a).
8. Nasal capsule.—(0) Unossified; (1) fully ossifiedinclud ing alinasal w alls and concha nasalis [turbi-
nals in Lanyon (1985)].
9. Septum interorbitale.—(0) Two op en fenestrae; (1)
infraorbital fenestra enlarged and supraorbital fe-
nestra reduced (Lanyon 1988a, c); (2) both fenestrae
reduced or absent; septum ossified.
10. M edial ridge in front al region of craniu m.—(0) Ab-
sent; (1) p resent.
Syringeal
Nomenclature of syringeal supp ort elements and
cartilages follows Ames (1971) and Lanyon (1984,
1985, 1986, 1988a, b, c). Nomenclature of syringeal
mu sculature follows King (1979) wh ich is also con-
sistent with the above references.
11. Double, complete A elements.—(0) None of the
(prim arily tr acheal) ‘‘A’’ series of syringea l sup por t
rings completely encircle the bronchi; (1) one A ele-
ment is double (associated with the bronchi rather
than the trachea) and complete (completely encir-
cling each bronchus). They are medially cartilagi-
nous; (2) two A elements are double, complete, and
medially cartilaginous; (3) two or more A elements
are double, complete, and med ially ossified; (4) one
A element is double, complete, and medially ossified;
(5) incomplete A3 elements contribute to support of
each bronchu s throu gh fusion to the comp lete A2 el-
ements; (6) incomplete A2 elements contribute to
sup port of each bronchus th rough fusion to the com-
plete A3 elements.
12. Cartilaginous dorsal segments of A1 elements.—(0)
A1 elements are nearly fully ossified, sometimeswith small, cartilaginous dorsal tips; (1) A1 elements
terminate dorsally in large cartilaginous segments.
13. M edial segments of A 2 elements m odified .—(0) Me-
d i a l s e g m e n ts o f A 2 e le m en t s (w h e n d o u b le a n d
complete) are smooth; (1) cartilaginous medial seg-
ments of A2 elements are enlarged and bulbous cau-
dally, presumably for the attachment of the internal
cartilages (Lanyon 1986); (2) cartilaginous med ial
segments of A2 elements are enlarged and bulbous
as in state (1) but are d isconnected from the rem ain-
ing (ossified) portion of the A 2 element.
14. Tracheal ‘‘drum’’.—(0) Tracheal A elements are
largely independent of one another; (1) two or more
t r a ch e a l A e le m en t s a r e fu s e d t o fo r m a t r a ch e a l
dr um . The dru m is poorly fused an d includ es A2 and
higher elements; (2) the dru m is robust an d includes
A2 and higher elements; (3) the d rum is robust and
includes A3 and higher elements, but not A2.
15. Nodule on lateral surface of A1 and A2.—(0) Lat -
eral surfaces of A1 and A2 are smooth; (1) ossified
nodule on the lateral surface of each A1 and A2 el-
e m en t , w i t h c ar t il ag i no u s c on n e ct io n s b e tw e en
them.
16. Pessulus divides tracheal A elements dorsally.—(0)
Tracheal A elements are complete d orsally; (1) ossi-fied p essulus extends anteriorly on the d orsal side of
the trachea to divide four or more A elements.

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 16/20
730 [Auk , Vol. 119JEFFREY S. BIRDSLEY
17. Pessulus extends from the first single, complete A
element .—(0) Pessulus (when present) is not attached
to the first single, dorsally complete A element; (1)
pessulus is an ossified caud al extension of the dorsal
side of the first single, dorsally complete A element.
18. Pessulus large, concave.—(0) Pessulus (wh en pr e-
sent) is convex dorsally; (1) the ossified pessulus is
relatively large and concave dorsally. Its greatest
breadth is at the p oint wh ere the internal cartilages
attach.
19. B1-2 connected ventrally.—(0) The bronchial,
cartilaginous B1 and B2 elements are ind ependen t at
their ven tral tip s; (1) B1 and B2 are connected at their
ventral tips. The connection is flattened or acute; (2)
B1 and B2 are connected at their ventral t ips. The
connection is symmetrically rounded.
20. B1-2 connections are close medially.—(0) The left
and right ventr al end s of B1 and B2 are relatively far
apar t; (1) the bronchi are close together in the regionof B1 and B2, so that the left and right ventral con-
nections of those elements nearly touch one an other
medially.
21. B1-2 diverge dorsally.—(0) B1 and B2 are near ly
parallel dorsally; (1) B1 and B2 diverge dorsally so
that th eir dorsal tips lie relatively far a par t.
22. B1-2 shaped as in Myiarchus.—(0) B1 is narrow
dor sally or B2 is narr ow ven trally; (1) B1 and B2 have
a d i s ti n ct co n fi gu r a t io n a s t y p i fie d b y Myiarchus
(Lanyon 1985): B1 is narrow except for a very broad
dorsal end. B2 is n arrow except for a broad, trian-
gular ventr al end w hich is barely attached to th e ven-
tral end of B1.
23. B1 breadth.—(0) Breadth of the lateral region of
B1 is less than or equal to the breadth at the ventral
and dorsal tips; (1) B1 is broader laterally than at the
tips.
24. B1 bulbous dorsally.—(0) B1 flat dorsally; (1) B1
bulbous or swollen at dorsal tips.
25. B2 Y-shaped ventrally.—(0) Ventral end of B2 is
rounded or truncate; (1) ventral end of B2 is forked
or s ha p ed like th e lett er ‘‘Y’’.
26. Syrinx and trachea laterally compressed .—(0) Tra -
chea a nd tracheobronchial junction are not both lat-
erally compressed; (1) in Lophotriccus and Oncostoma(Lanyon 1988c), the entire trachea and tracheobron-
chial junction are laterally compressed so that they
are unusually narrow when viewed dorsally or ven-
trally, and broad when viewed laterally. Similar mor-
phology occurs in Machetornis where the tracheo-
bronchial junction, but not the trachea, is laterally
compressed (J. S. Birdsley pers. obs.).
27. Cartilaginous bronchial plate.—(0) Dorsal tips of
the incomplete bronchial A elements are not con-
nected with one another or with the pessulus; (1) a
horseshoe-shaped cartilaginous plate connects the
dorsal t ips of the incomplete bronchial A elements
and the pessulus. It forms the dorsal or craniodorsalmargins of the medial tympaniform membranes.The
internal cartilages are very small, delicate rods lo-
cated near the caudal ends of the horseshoe, some-
times attached to i t ; (2) a cartilaginous plate as in
state (1), but the in ternal cartilages are robust.
28. Cartilaginous tracheal plate.—(0) Lower tr acheal
A elements are ossified dorsally; (1) cartilaginous
pessu lus extends cranially so that the lower tracheal
A elements up to A6 or A8 are cartilaginous dorsally.
29–39. Int ernal cart ilages.—The internal tympani-
form mem branes in the syringes of all tyrann ids and
s o m e c ot in g id s c on t a in c ar t il ag in o u s s t r u ct u r e s
which are often attached to A elements. Because
there is extreme variation in th e shap e, position, and
attachment of internal cartilages, Prum and Lanyon
(1989) hypothesized th em to be derived in cotingids
independently of those in tyrannids. Prum (1990b)
questions their homology even within tyrannids, and
recommend s using u nique, detailed morp hologies of
internal cartilages as characters, while refraining
from the hypothesis of homology of internal carti-lages as broadly defined. Here I follow that recom-
mendation and the precedent of Lanyon (1984, 1985,
1986, 1988a, b, c) in coding shared, detailed mor-
phologies of tyrann id internal cartilages as indepen-
dent, d erived characters.
29. Narrow , linear .—(0) Absent; (1) cartilages are
narr ow an d basically linear.
30. Triangular .—(0) The base (the cranial end) is
narrower than or scarcely greater than the w idth of
the re st of the cartilage; (1) the cartilag e is trian gu lar,
w i th t h e b a se m u ch b ro ad e r t h a n t h e r e st o f t h e
cartilage.
31. Spatulate.—(0) Absent; (1) cartilages are broad
and shaped like a sp atula or child’s mitten w ith a
thickened dorsal edge.
32. ‘‘J’’ or ‘‘L’’ shaped.—(0) Absen t; (1) cartilag es are
thin and bent in a ‘‘J’’ or ‘‘L’’ sha pe; (2) cartilages are
robu st a nd bent in a ‘‘J’’ or ‘‘L’’ shap e.
33. Forked .—(0) Absent; (1) cartilages are forked
caudally.
34. Ventral extension.—(0) Absent; (1) cartilages
have poorly staining, amorphous ventral extensions
from their caudal halves; (2) cartilages have well-
formed, squarish ventral extensions from their cau-
dal halves.35. Broad , amorphous caudal segments.—(0) Absent;
(1) cartilages are narrow cranially but broaden and
become amorphous caudally.
36. Narrow , curved .—(0) Absent; (1) cartilages are
relatively n arrow and curved (medially concave).
37. Straight , expanded caudally.—(0) Absent; (1) car-
tilages are relatively long, straight, and expand ed
caudally. They are somewhat twisted, with the cra-
n i a l h a l f fl a t te n ed a t r ig h t a n g le s t o t h e i n te r n al
t y m p a n i fo r m m e m b r a ne a n d t h e c a u d a l h a l f fl a t-
tened w ithin the plane of the membrane.
38. At tached to tracheal drum and incomplete A ele-
ments.—(0) Cartilages are attached either to an in-complete A element or to a tracheal drum, but not
both; (1) cartilages are attached to the dorsal end of

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 17/20
July 2002] 731Phylogeny of Tyrannidae
an incomplete A element (either A1 or A2) as well as
to a tracheal drum .
39. Attached to ventral side of tracheobronchial junc-
tion .—(0) Cartilages are attached to the dorsal side
of various structures in the tracheobronchial junc-
tion; (1) cartilages are attached to ventral or m edio-
ventral segments of A2 or A3.
40. Narrow strand of cartilage between B2 and B3.—
(0) Absent; (1) in addition to the large internal car-
tilages, there is a narrow strand of cartilage in the
internal tym paniform membrane uniquely p laced
between the ventral end s of B2 and B3.
41. Cartilaginous plug of tissue between A 2 ele-
ments.—(0) The space just caudal to the tracheobron-
chial junction is vacant; (1) there is a plug of tissue
just caudal to the tracheobronchial junction and be-
tween the cartilaginous m edial segments of the A2
elements. This p lug stains weakly for cartilage.
42. Mm. tracheolaterales.—(0) Cover only the lat-eral portions of the trachea; (1) right and left Mm.
tracheolaterales widen ventrally and meet above
A20, somet imes lower, and com pletely cover the ven-
tral and ventrolateral surface of the trachea from
there p osteriad, diverging just before their insertion
in the region of the syrinx; (2) M m. tracheolaterales
cover the ventr al and ventrolateral surface of the tra-
chea as in state (1) but do not diverge at their inser-
tion. Instead, they converge to a point and insert
midventrally on the lower A elements.
43. Mm. obliqui ventrales.—(0) Intrinsic syringeal
muscles (if present) are different from Mm. obliqui
ventrales in detail of fiber direction and form of in-
sertion (Prum and Lanyon 1989; Prum 1990b, 1992);
(1) Mm. obliqui ventrales insert on the ventral or lat-
eral side of the syrinx (Ames 1971, McKitrick 1985,
Mobley and Prum 1995). (2) M m. obliqui ventrales in -
sert on the d orsal side of the syrinx (Ames 1971).
44. Mm . obliqui ventrales well developed.—(0) M m.
obliqui ventrales (if present) are relatively undevel-
oped, ap pearing flat; (1) Mm. obliqui ventrales are well
developed , appearing as round , bulging mu scle mas-
ses. Present in at least some mem bers of the kingbird
an d Elaenia assemb lages (Lanyon 1984, 1988a) and in
Hymenops, Polioxolmis, and Cnemarchus (pers. obs.).45. Mm. obliqui laterales.—(0) Absent; (1) syrinx
has a pair of laterally-originating and laterally-in-
serting intrinsic syringeal mu scles. In m ost taxa pos-
sessing th em, these mu scles exist in ad dition to M m.
obliqui ventrales, but app arently in some tody-tyrants,
Mm. obliqui laterales are the only intrinsic syringeal
mu scles (Ames 1971).
Plumage
46. Crown patch.— (0 ) C r o w n p lu m a g e i s m o n o -
chromatic; (1) crown plumage contains a patch of white, red, orange, or yellow which is usually con-
cealed or semiconcealed.
47. Sexually dichromatic plumage.—(0) Sexes h ave
similar or identical plumage color and patterns; (1)
sexes have markedly different plumage color and
patterns.
48. Wing patterned .—(0) Remiges ar e sim ilarly col-
ored on their p roximal and distal halves; (1) remiges
are black or fuscous d istally, and buff or cinnamon-
rufous proximally; (2) remiges as in state (1) except
they are white proximally.
49. Wing bars.—(0) Ab sent; (1) ru fous-buff; (2) yel-
low; (3) white.
50. Throat plumage. —(0 ) U n st r ea k ed ; ( 1) d a r k
streaks against a white background .
51. Outer rectrix pale.—(0) Outer rectrices are sim-
ilarly colored on their inner and outer webs; (1) outer
webs of the outer rectrices are pale (white to cinna-
mon–rufous), contrasting with darker inner webs.
52. Tail length.—(0) Short to m od erate; (1) elongat-
ed, forked.53. Tail rufous.—(0) Tail is various colors but not
entirely rufous; (1) entire ta il is ru fous.
54. Underparts rufous.—(0) Underparts various col-
ors but not entirely rufous; (1) underparts entirely
rufous.
55. Superciliary .—(0) No contrasting stripe over or
through the eye; (1) white, yellow, or cinnamon su-
perciliary stripe over or through the eye.
56. Plumage black and white.—(0) Plumage has at
least some color or is solid black, brown, or white;
(1) plumage is black and white or brown and white.
57. R ictal bristles.—(0) Short or absent; (1) as long
or nearly as long as the bill.
58. Pointed crest .—(0) Feathers of the crown form a
rounded crest; (1) feathers of the crown form a point-
ed, prominent crest.
59. Primaries notched .—(0) Outer primaries not
notched at their tips; (1) outer one, two, or three pri-
mar ies are notched or attenuated in at least the male
of the species.
Behavior and Ecology
60. Habitat .—(0) Inhabit forests and wood lands; (1)
inhabit open grassland or marsh; (2) inhabit opencliffs, canyons, and roadcuts.
61. Foragin g behavior .—Fitzp atrick (1980) charac-
terized the foraging behavior of nearly every tyran-
nid genus and recognized 10 ‘‘foraging mode’’ cat-
e g or ie s i n t h e fa m i ly . H e r e I t r e at e a ch o f t h e s e
foraging mode categories as alternative unordered
derived states of a single character. I consulted Snow
(1982), Hilty and Brown (1986), and Marini (1992) for
behavior of outgroups.
(0) Fruit/ up ward hover-glean generalists feed
heavily on fru its dur ing most or all of the year
but also upward hover-glean and perch-glean
insects.( 1) F ru i t / h a w k g e ne r al is t s co m b in e f r u g iv or y
with aerial-hawking of flying insects. They

8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 18/20
732 [Auk , Vol. 119JEFFREY S. BIRDSLEY
use these capture techniques in roughly equal
proportions.
(2) Outw ard hover-glean generalists often glean
insects from foliage surfaces via an outward or
downward ‘‘sally’’ or capture flight. The bird
may slow down and hover while capturing the
insect or snap it u p w hile in d irect flight, often
striking the foliage and continuing to a new
p e r ch . U p w a r d h o ve r- gl ea n i n g a n d a er ia l -
hawking are ad ditional significant elements of
their foraging repertoires.
(3) Enclosed perch hawk generalists have foraging
repertoires composed of roughly equal prop or-
tions of aerial-hawking, up ward hover-glean-
ing, and up ward-striking.
(4) Near-ground generalists usually hu nt from
perches near the ground and use a w ide vari-
ety of capture techniques including perch-to-
g r ou n d a n d p e r ch - to -w a t er s a ll ie s, u p w a r dand outward hover-gleaning and striking, and
aerial-hawking.
(5) Perch-glean sp ecialists captur e m ost of their
prey while p erched, without sallying. Hover-
gleaning and frugivory are minor components
of their rep ertoire.
(6) Up ward -strike sp ecialists captu re m ost of their
prey in an explosively rapid upward sally to
the underside of a leaf, snatching or scooping
an insect off the u nder-surface, usu ally without
hovering.
(7) Aerial-hawk specialists capture most of theirprey out of the air.
(8) Perch-to-ground specialists hun t from low
p e r ch e s, u s u a l ly s a ll yi n g t o t h e g r ou n d t o
catch prey. Aerial-hawking may be a minor
component of their repertoire.
(9) Ground specialists stand, w alk, or run on the
ground and search for their prey which they
pick off the ground or low vegetation. Aerial-
hawking from the groun d m ay be a minor com-
ponent of their repertoire.
62. Cavity nesting.—(0) Nests are in open situa-
tions; (1) nests are located in tree cavities or holes in
earthen banks.
63. Enclosed nest within hanging vegetation.—(0)
Nest, if hanging, is built around a branch and sus-
pended from it; (1) enclosed, globular nest with a
s id e e nt r an ce i s b u i lt w i th i n a h a n gi ng m a ss o f
vegetation.
64. Enclosed nest pendant .—(0) Nest is not su spend -
ed from a branch; (1) enclosed, globular nest with a
side or bottom entran ce is suspend ed from a branch;
(2) enclosed, elongate (1 m or m ore) nest with a side
entrance is suspended from a branch.
65. Enclosed , pendant nest retort-shaped .—(0) Nes t, if han ging, has an entr ance hole at or above the bottom
of the n est chamber; (1) enclosed, p endan t, globular
nest shaped like a chemists’ retort with a downwar d-
pointing entrance spout.
66. Enclosed nest with visor over entrance.—(0) Nest,
if enclosed, has an un sheltered entran ce hole; (1) en-
closed nest has a short roof or visor over the side
entrance.
67. Nest used as dormitory.—(0) Nests are used only
during breeding; (1) adults sleep in their nests at all
seasons. Presum ed p resent only in Rhynchocyclus and
Tolmomyias (Skutch 1960).
Eggs
68. Eggs.—( 0) M a rk ed w i t h d a r k b lo t ch e s a n d
streaks; (1) plainly colored, usually some shade of
white.
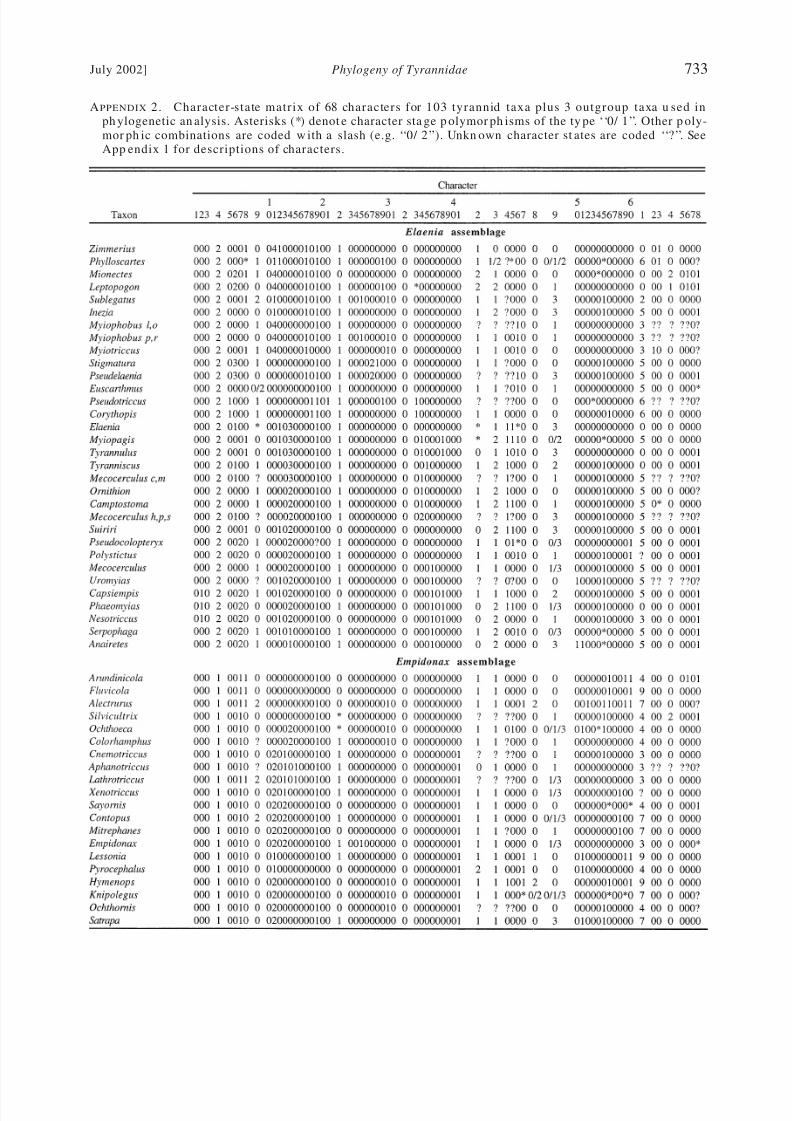
8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 19/20
July 2002] 733Phylogeny of Tyrannidae
APPENDIX 2. Character-state matrix of 68 characters for 103 tyrannid taxa plus 3 outgroup taxa u sed inph ylogenetic an alysis. Asterisks (*) denot e character sta ge p olymor ph isms of the ty pe ‘‘0/ 1’’. Other p oly-mor ph ic combinations are coded w ith a slash (e.g. ‘‘0/ 2’’). Unkn own character st ates are coded ‘‘?’’. SeeApp endix 1 for descriptions of characters.
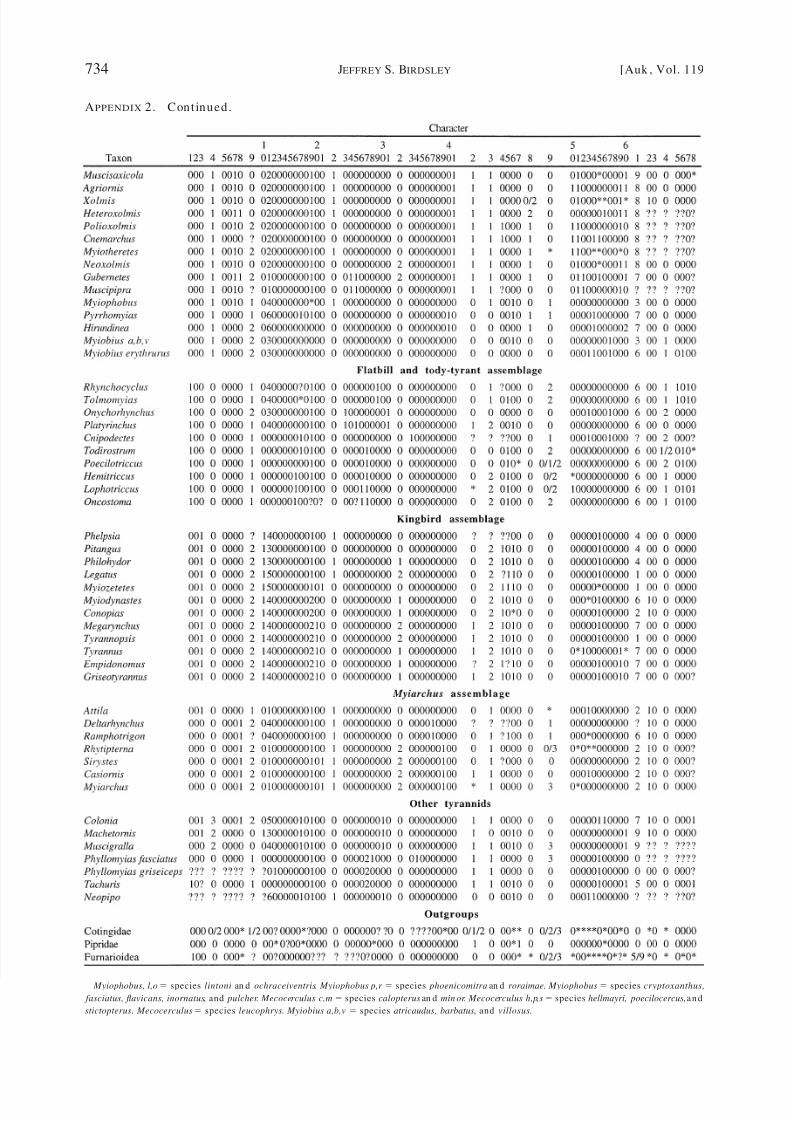
8/6/2019 Auk 119-3-715
http://slidepdf.com/reader/full/auk-119-3-715 20/20
734 [Auk , Vol. 119JEFFREY S. BIRDSLEY
APPENDIX 2 . C on t in u e d .
Myiophobus, l,o species lintoni an d ochraceiventris. Myiophobus p,r species phoenicomitra an d roraimae. Myiophobus species cryptoxanthus,
fasciatus, flavicans, inornatus, and pulcher. Mecocerculus c,m species calopterus an d min or. Mecocerculus h,p,s species hellmayri, poecilocercus, a n d
stictopterus. Mecocerculus species leucophrys. Myiobius a,b,v species atricaudus, barbatus, and villosus.