author's personal copy - home.gwu.eduhome.gwu.edu/~rpyron/publications/pyron_wiens_2011.pdf ·...
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
A large-scale phylogeny of Amphibia including over 2800 species, and a revisedclassification of extant frogs, salamanders, and caecilians
R. Alexander Pyron a,⇑, John J. Wiens b
a Dept. of Biological Sciences, The George Washington University, 2023 G St. NW, Washington, DC 20052, United Statesb Dept. of Ecology and Evolution, Stony Brook University, Stony Brook, NY 11794-5245, United States
a r t i c l e i n f o
Article history:Received 7 May 2011Revised 10 June 2011Accepted 12 June 2011Available online 23 June 2011
Keywords:AmphibiaAnuraApodaCaudataLissamphibiaGymnophionaPhylogenySupermatrixSystematics
a b s t r a c t
The extant amphibians are one of the most diverse radiations of terrestrial vertebrates (>6800 species).Despite much recent focus on their conservation, diversification, and systematics, no previous phylogenyfor the group has contained more than 522 species. However, numerous studies with limited taxon sam-pling have generated large amounts of partially overlapping sequence data for many species. Here, wecombine these data and produce a novel estimate of extant amphibian phylogeny, containing 2871 spe-cies (�40% of the known extant species) from 432 genera (�85% of the �500 currently recognized extantgenera). Each sampled species contains up to 12,712 bp from 12 genes (three mitochondrial, ninenuclear), with an average of 2563 bp per species. This data set provides strong support for many groupsrecognized in previous studies, but it also suggests non-monophyly for several currently recognized fam-ilies, particularly in hyloid frogs (e.g., Ceratophryidae, Cycloramphidae, Leptodactylidae, Strabomanti-dae). To correct these and other problems, we provide a revised classification of extant amphibians fortaxa traditionally delimited at the family and subfamily levels. This new taxonomy includes several fam-ilies not recognized in current classifications (e.g., Alsodidae, Batrachylidae, Rhinodermatidae, Odontoph-rynidae, Telmatobiidae), but which are strongly supported and important for avoiding non-monophyly ofcurrent families. Finally, this study provides further evidence that the supermatrix approach provides aneffective strategy for inferring large-scale phylogenies using the combined results of previous studies,despite many taxa having extensive missing data.
� 2011 Elsevier Inc. All rights reserved.
1. Introduction
With over 6800 known species (AmphibiaWeb; http://www.amphibiaweb.org/, accessed April, 2011; hereafter ‘‘AW’’) the extantamphibians (frogs, salamanders, and caecilians) are one of the mostdiverse radiations of terrestrial vertebrates. The number of knownextant amphibians has increased rapidly in recent years, with over2700 species (�40%) described in the last 26 years (Duellman,1999; Lannoo, 2005). This newly discovered diversity includes doz-ens of new species from known genera in poorly studied tropical re-gions such as Madagascar (Vieites et al., 2009), but also new generain relatively well-explored regions such as the southeastern UnitedStates (Camp et al., 2009), and even new families such as Nasikabatr-achidae (Biju and Bossuyt, 2003). Unfortunately, much extantamphibian diversity is currently under extreme threat from pres-sures such as habitat loss, global climate change, and infectious dis-ease, and many species have gone extinct in the last few decades(Blaustein and Wake, 1990; Stuart et al., 2004).
A phylogenetic framework is critical for discovering, under-standing, and preserving extant amphibian diversity, but a large-scale phylogeny for extant amphibians is presently lacking. How-ever, recent molecular and combined-data studies have madeimportant contributions to higher-level phylogeny (Frost et al.,2006; Roelants et al., 2007; Wiens, 2007a, 2011) and to the phylog-eny of many major groups, such as caecilians (San Mauro et al.,2009; Zhang and Wake, 2009b), hyloid frogs (Darst and Cannatella,2004), ranoid frogs (e.g., Bossuyt et al., 2006; Wiens et al., 2009),microhylid frogs (van der Meijden et al., 2007), bufonid frogs(Pauly et al., 2004; Pramuk et al., 2008; Van Bocxlaer et al.,2009), centrolenid frogs (Guayasamin et al., 2009), dendrobatidfrogs (Grant et al., 2006; Santos et al., 2009), hemiphractid frogs(Wiens et al., 2007a), hylid frogs (Faivovich et al., 2005, 2010;Wiens et al., 2005b, 2010), terraranan frogs (Hedges et al., 2008;Heinicke et al., 2009), and salamanders (Kozak et al., 2009; Vieiteset al., 2011; Wiens et al., 2005a, 2007b; Zhang and Wake, 2009a).
The largest estimate of extant amphibian phylogeny to date isthat of Frost et al. (2006). Those authors reconstructed amphibianphylogeny based on relatively intensive sampling of species (522)and characters (up to 4.9 kb of sequence data from 2 mitochondrialand 5 nuclear genes [mean = 3.5 kb], and 152 morphological
1055-7903/$ - see front matter � 2011 Elsevier Inc. All rights reserved.doi:10.1016/j.ympev.2011.06.012
⇑ Corresponding author.E-mail addresses: [email protected] (R. Alexander Pyron), [email protected].
sunysb.edu (John J. Wiens).
Molecular Phylogenetics and Evolution 61 (2011) 543–583
Contents lists available at ScienceDirect
Molecular Phylogenetics and Evolution
journal homepage: www.elsevier .com/ locate /ympev

Author's personal copy
characters). Those authors also proposed extensive changes in tax-onomy, especially for taxa delimited at the family and genus level.However, that study has also been criticized on numerous grounds,including concerns about taxon sampling and methodologicalstrategies (Marjanovic and Laurin, 2007; Pauly et al., 2009; Wiens,2007b, 2008). For example, those authors collected up to �4900characters per species, but their analysis is apparently based on15,320 characters, suggesting that their controversial approach tosequence alignment (POY) dominates their results (Wiens, 2008).Although some of the changes made by Frost et al. (2006) havebeen widely adopted, others are more controversial, such as thepartitioning of Bufo and Rana (Marjanovic and Laurin, 2007; Paulyet al., 2009; AW). Indeed, many of these changes are no longer sup-ported, even in Frost’s (2011) taxonomic database of extantamphibians (e.g., the families Amphignathodontidae, Batrachoph-rynidae, Cryptobatrachidae, and Thoropidae recognized by Frostet al. (2006)). Much of the most unstable taxonomy involves thefamily-level assignment of many of the genera of hyloid frogs, par-ticularly those traditionally assigned to the family Leptodactylidae.
Clearly, extant amphibian phylogeny and classification is still inneed of additional study. Fortunately, the numerous studies refer-enced above (and many others) have produced a massive amountof data that are potentially suitable for a combined, supermatrixapproach (e.g., de Queiroz and Gatesy, 2007; Driskell et al., 2004;Pyron et al., 2011; Thomson and Shaffer, 2010; Wiens et al.,2005b). This includes thousands of species represented in GenBankfor numerous nuclear and mitochondrial genes, often with sub-stantial overlap of genes among species.
Here, we present a large-scale estimate of amphibian phylog-eny, including 2871 species (42% of the 6807 known, extantamphibian species) from 432 of the 504 currently recognized gen-era (86%), and representatives from every currently delimited, ex-tant family and subfamily. This is 5.5 times more species andnearly twice as many genes as the largest previous study (Frostet al., 2006). The data matrix includes up to 12,712 bp for each spe-cies from 12 genes (three mitochondrial, nine nuclear). Impor-tantly, rather than simply reanalyzing published data forrelatively well-studied families (e.g., dendrobatids, hylids), we ad-dress the monophyly and relationships of many smaller groupsthat have not been the subject of focused studies (e.g., Ceratophryi-dae, Cylcoramphidae), as well as relationships among families. Weproduce a revised classification of extant amphibians, focusing ontaxa traditionally ranked as families and subfamilies. This studyalso provides additional support for the value of the supermatrixapproach to large-scale phylogenetic inference (e.g., de Queirozand Gatesy, 2007; Driskell et al., 2004; Pyron et al., 2011; Thomsonand Shaffer, 2010; Wiens et al., 2005b).
2. Materials and methods
2.1. Taxonomic reference
This analysis has been several years in the making. Our initialtaxonomy was based on the September 2009 update of theAmphibiaWeb (AW) database. However, when we refer to currentnumbers, these are taken from the April, 2011 update. The AW listis fairly current in terms of recently described species, but moreconservative than the Amphibian Species of the World (Frost,2011; hereafter ‘‘ASW’’) regarding some of the more controversialof the recent taxonomic changes (e.g., Bufo and Rana maintain sim-ilar composition as they did prior to Frost et al., 2006). We notesome instances where recent updates have modified our originalclassification. Note that even when not made explicit, we refer inall instances to the known extant diversity of Lissamphibia, giventhat the clade Amphibia includes numerous extinct stem-group
members that are not lissamphibians. The gymnophionans, cau-dates, and anurans also contain numerous extinct taxa, many ofwhich are grouped in separate genera, subfamilies, and familiesthat are not addressed in our analyses or included in our discussionof phylogeny. See Marjanovic and Laurin (2007), Carroll (2009),and Pyron (2011) for an overview of these taxa, their phylogeneticaffinities, and the origins of Amphibia and Lissamphibia.
2.2. Molecular data
We identified 12 candidate loci that have been broadly sampledand successfully used in amphibian phylogenetics at both lowerand higher taxonomic levels. These 12 genes included nine nucleargenes: C-X-C chemokine receptor type 4 (CXCR4), histone 3a(H3A), sodium–calcium exchanger (NCX1), pro-opiomelanocortin(POMC), recombination-activating gene 1 (RAG1), rhodopsin(RHOD), seventh-in-absentia (SIA), solute-carrier family 8(SLC8A3), and tyrosinase (TYR). Three mitochondrial genes werealso included: cytochrome b (cyt-b), and the large and small sub-units of the mitochondrial ribosome genes (12S/16S; omittingthe adjacent tRNAs as they were difficult to align and representedonly a small amount of data). This selection of genes includes al-most all of those genes used in the higher-level analyses by Frostet al. (2006) and Roelants et al. (2007), and most of those used inother large-scale studies (Faivovich et al., 2005; Grant et al.,2006; Wiens et al., 2009, 2010). However, we did not include thenuclear gene 28S (used by Frost et al. (2006)), as previous analysesof this gene region alone suggest that it contains relatively fewinformative characters and supports some relationships that aregrossly inconsistent with other studies (see Wiens et al., 2006).We conducted GenBank searches by family and subfamily to gath-er all available sequences, using a minimum-length threshold of200 bp (a somewhat arbitrary threshold of 1.5% of the total matrixlength, to avoid including very short [e.g., <50 bp] fragments), andstopping in August of 2010. Only species in the taxonomic data-base were included in the sequence matrix, which excludednumerous named taxa of ambiguous status, and many sequenceslabeled ‘sp.’ We removed a few (<10) taxa with identical sequencedata for all genes (arbitrarily retaining the first in alphabetical or-der), to avoid potentially misidentified or otherwise confoundedspecimens or sequences.
For the protein-coding genes, alignment was relatively straight-forward. Conceptual translations were used to ensure an openreading frame, and sequences were aligned using the translation-alignment algorithm in the program Geneious v4.8.4 (GeneMattersCorp.), with the default cost matrix (Blosum62) and gap penalties(open = 12, extension = 3). For the ribosomal RNA sequences (12Sand 16S sequences), alignment was more challenging. Preliminaryglobal alignments using the MUSCLE (Edgar, 2004) and CLUSTAL(Larkin et al., 2007) algorithms under a variety of gap-cost param-eters yielded low-quality results (i.e., alignments with large num-bers of gaps and little overlap of potentially homologouscharacters).
We subsequently employed a two-step strategy for these data.First, we identified sequence clusters of similar length and cover-age from the global alignment. These were subsequently alignedseparately using the MUSCLE algorithm with the default high-accuracy parameters, which have been shown to outperform CLUS-TAL in a variety of settings (Edgar, 2004). These alignments weresubsequently refined using the MUSCLE refinement algorithm,and then adjusted manually and trimmed for quality and maxi-mum coverage (i.e., end sequences with low overlap and poorapparent alignment were deleted using the alignment editor inGeneious). These length and position-based sequence groups werethen aligned to each other using the profile-profile alignmentalgorithm in MUSCLE. The resulting final global alignment was
544 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
manually adjusted and trimmed again for maximum quality andcoverage, and refined a second time. Final adjustments were madeby eye for a small number of sequences. This process was repeatedfor 12S and 16S separately. For 12S, four clusters were producedand combined using profile alignment, with five clusters for 16S.
The final concatenated alignment consists of 12,712 bp for 2871species. These species included 42% of the currently recognizedamphibian species (6807; this and subsequent numbers from AW2011), representing 41 caecilian species in 23 genera (22% of 189known extant species and 70% of 33 genera), 436 salamander spe-cies in 68 genera (72% of 608 species and 99% of 69 genera), and2394 frog species from 341 genera (40% of 6010 species and 85%of 402 genera). We selected Homo as an outgroup because datawere available for Homo from all 12 genes, and the sister groupto Amphibia is Amniota (e.g., Alfaro et al., 2009; Hugall et al.,2007; Pyron, 2010). The matrix contains data from 2572 speciesfor 16S (90%, 1986 bp), 2312 species for 12S (81%, 1357 bp), 1241for cyt-b (43%, 1140 bp), 1049 for RAG-1 (37%, 2700 bp), 606 forTYR (21%, 1132 bp), 589 for RHOD1 (21%, 315 bp), 478 for SIA(17%, 397 bp), 445 for POMC (16%, 666 bp), 352 for H3A (12%,328 bp), 284 for CXCR4 (10%, 753 bp), 220 for NCX1 (8%,1338 bp), and 175 for SLC8A3 (6%, 1132 bp). The mean length perspecies is 2563 bp (20% of the total length of the matrix,12,712 bp), with a range from 249 to 11,462 bp (2–90%), and isavailable in Dryad repository doi:10.5061/dryad.vd0m7. GenBanknumbers are listed in Appendix S1.
Clearly, many taxa had large amounts of missing data (some>97%), and on average each species had 80% missing cells. How-ever, several lines of evidence suggest that these missing dataare not problematic. First, two genes (12S/16S) were shared bythe vast majority of taxa (90% and 81%, respectively), providinga ‘‘backbone’’ for the placement of most taxa based on overlap-ping sequence data. Simulations suggest that this sampling designcan be critically important for allowing the accurate placement oftaxa with extensive missing data, as opposed to having all genesbe randomly sampled across species with limited overlap (Wiens,2003). Second, a large body of empirical and theoretical studiessuggests that highly incomplete taxa can be accurately placedin model-based phylogenetic analyses (and with high levels ofbranch support), especially if a large number of characters havebeen sampled (recent review in Wiens and Morrill, 2011). Finally,several recent empirical studies have shown that the supermatrixapproach (with extensive missing data in some taxa) yieldsgenerally well-supported large-scale trees that are in generalhighly congruent with both existing taxonomies and previousphylogenetic estimates (e.g., Driskell et al., 2004; McMahon andSanderson, 2006; Pyron et al., 2011; Thomson and Shaffer,2010; Wiens et al., 2005b).
2.3. Phylogenetic analyses
We performed phylogenetic inference using maximum likeli-hood (ML) and assessed support using non-parametric bootstrap-ping (BS). We assume that Bayesian analysis would yield verysimilar results (but would be very difficult to implement for so manytaxa), and we strongly prefer model-based methods to parsimony(for reasons described in Felsenstein (2004)). We performed ML treeinference and non-parametric bootstrapping using the programRAxMLv7.0.4 (Stamatakis, 2006) with the 12-gene concatenatedmatrix (species-tree methods were not practical given the largenumber of taxa). We used the GTRGAMMA model for all genes andpartitions because GTR is the only substitution model implementedin RAxML, and all other substitution models are encompassed with-in the GTR model (Felsenstein, 2004). The GTRGAMMA model inRAxML is recommended over the GTR + C + I because the largenumber of rate categories for C (25, as opposed to the usual 4)
should adequately account for potentially invariant sites withoutthe need for an extra parameter (Stamatakis, 2006). Even thoughthe GTR + C + I model is implemented in some versions of RAxML,its use is explicitly not recommended (Stamatakis, 2006).
Previous phylogenetic analyses of these data show thatGTR + C + I is generally the best-fitting model for these genes(e.g., Roelants et al., 2007; Wiens et al., 2005a,b, 2009, 2010). Theseprevious analyses also suggest that the protein-coding genesshould be partitioned by codon position, whereas the ribosomalgenes (12S, 16S) should be partitioned by stems and loops, withseparate partitions within and between genes. These secondarystructures were identified and coded following the protocol usedby Wiens et al. (2005b), based on predicted features from Pseuda-cris regilla (12S; AY819376) and Rana temporaria (16S; AY326063)from the European Ribosomal RNA database (http://bioinformat-ics.psb.ugent.be/webtools/rRNA/). The placement of stems andloops appears to be conserved across most sites, at least withinfrogs (Wiens et al., 2005b). Our final analysis was partitioned bygene, codon position (for protein-coding genes), and stems andloops (for ribosomal genes).
To find the optimal ML tree, we used a searching strategy thatcombined the rapid bootstrapping algorithm (100 non-parametricbootstrap replicates) with the thorough ML search option (20 inde-pendent searches, starting from every fifth bootstrap replicate).Similar analyses were performed numerous (>10) times as newtaxa and sequences were added to the near-final matrix. Theseanalyses collectively represent hundreds of independent searchesfrom random starting points. All of these preliminary analysesshowed high congruence with the final ML topology. This concor-dance strongly suggests that our final ML estimate represents theoptimal topology for these data (or close to it). Given that BS valuesgenerally appear to be conservative (Felsenstein, 2004), we consid-ered clades with values of 70% or greater to be well-supported.These analyses were performed on a 240-core Dell PowerEdgesupercomputing system at the High Performance Computing Cen-ter at the City University of New York, and were completed in16 days using 24 nodes of the CUNY cluster.
2.4. Taxonomic revision
A major goal of this study was to revise the higher-level taxon-omy of extant amphibians to correspond with the new phylogeny,given several major problems that were discovered. In the Resultssection below, we compare our results with existing phylogeniesand classifications, and describe our proposed solutions to taxo-nomic problems as we describe them (rather than having a separatesection discussing taxonomy). In general, we attempt to alter exist-ing classifications (e.g., AW, ASW) as little as possible, and only whenexisting groups are not monophyletic. Furthermore, we recognizenew groups only if they are strongly supported. Given the size ofthese phylogenies, we do not detail every congruence and discor-dance between our phylogeny and all previous studies (especiallywithin families). Instead, we emphasize necessary taxonomicchanges revealed by our study. We also focus our phylogenetic com-parisons on the largest of the previous analyses (in terms of taxo-nomic scope and number of species sampled), those of Frost et al.(2006), Roelants et al. (2007), and Wiens (2007a, 2011). The genericcomposition of all families and subfamilies in our revised classifica-tion is provided in Appendix B. Genus names follow our original tax-onomic database from the 2009 AW update.
In some cases, our analyses show that higher taxa (i.e., familiesand subfamilies) are not monophyletic, but not all genera havebeen included in our tree. Some of these genera are effectively‘‘orphaned’’ because it is no longer clear to which higher taxon theybelong. We generally denote these as incertae sedis within thehigher taxon in which they were embedded in previous
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 545

Author's personal copy
classifications. Resolving the placement of these taxa in the treeand classification will require additional data and analyses. Weconsider this a more conservative strategy than simply placingthem in the nominate group without data. While in theory thisstrategy creates more ‘‘instability’’ by removing taxa from namedgroups, it highlights the need for their study in future analyses,and does not promote the taxonomic burden of heritage (Pyronand Burbrink, 2009) in further propagating classifications withhigh probability of error based solely on the status quo.
3. Results
A summary of the ML tree based on the rapid-bootstrapping anal-ysis from RAxMLv7.0.4 (lnL = �1704992.20) is shown in Fig. 1. Thisphylogeny is generally well-supported, with 64% of nodes having BSproportions >70. Our analyses support the monophyly of frogs, cae-cilians, and salamanders, respectively, and weakly support a sister-group relationship between frogs and salamanders, as found in mostother studies of extant lissamphibian phylogeny (Frost et al., 2006;Pyron, 2011; Roelants et al., 2007; San Mauro, 2010; Wiens, 2011;Zhang et al., 2005). An alternative grouping of Gymnophiona andCaudata (i.e., Procera) has been supported by some studies (e.g.,Feller and Hedges, 1998), and in re-analyses of others (e.g., Zhanget al., 2005; San Mauro et al., 2005; see Marjanovic and Laurin,2007; Pyron, 2011). Many of the family-level relationships withinthese three groups remain unchanged from recent estimates (Frostet al., 2006; Pyron, 2011; Roelants et al., 2007; San Mauro, 2010;Wiens, 2007a, 2011). However, we find some significant deviationsfrom previous phylogenies and taxonomies, which we describe be-low along with proposed solutions.
Within caecilians (Fig. 2A), our results agree with other recentstudies in supporting clades corresponding to Rhinatrematidae,Ichthyophiidae, and Caeciliidae (Frost et al., 2006; Roelants et al.,2007; San Mauro et al., 2009; Zhang and Wake, 2009b). The tradi-tional family-level classification of caecilians (still used by AW,2011) is not supported, given that Uraeotyphlidae renders Ichthy-ophiidae paraphyletic and that Scolecomorphidae and Typhlonect-idae render Caeciliidae paraphyletic. Furthermore, we find (Fig. 2A)the caeciliid subfamily Typhlonectinae (recognized by ASW) to beparaphyletic with respect to Caeciliinae (Cthonerpeton and Typhlo-nectes are not sister taxa, although this is weakly supported), andalso makes Caeciliinae non-monophyletic (i.e., the caeciliines Cae-cilia and Oscaecilia are in a strongly supported clade with Cthonerp-eton and Typhlonectes, which excludes all other caeciliine generasuch as Dermophis, Gegeneophis, and Siphonops). Typhlonectinaeis synonymized with Caeciliinae in our classification. Within Cae-ciliidae, we concur with ASW in recognizing the strongly supportedsubfamily Scolecomorphinae for Crotaphatrema and Scolecomor-phus, which is the sister group to all other caeciliids. Althoughwe could recognize Caeciliinae as the sister group to Scolecomor-phinae, this clade is only weakly supported, despite strong supportfor each subfamily-level clade. Thus, we recognize these two cladesas separate subfamilies. We restore the subfamily Herpelinae (Lau-rent, 1984) for the clade comprising Herpele and Boulengerula. Werecognize the other strongly supported clade as Caeciliinae (Figs. 1,2A; Appendix B). This arrangement accommodates all taxa in-cluded in our tree, though many other genera have not yet beensampled. The following genera are thus considered Caeciliidaeincertae sedis: Atretochoana, Brasilotyphlus, Idiocranium, Indotyphlus,Microcaecilia, Mimosiphonops, Nectocaecilia, Parvicaecilia, Potomo-typhlus, and Sylvacaecilia.
Within salamanders, the family and subfamily-level relation-ships are mostly consistent with most recent model-based molec-ular analyses (Roelants et al., 2007; Wiens, 2011; Wiens et al.,2005a; Zhang and Wake, 2009a) and current classifications (AW,
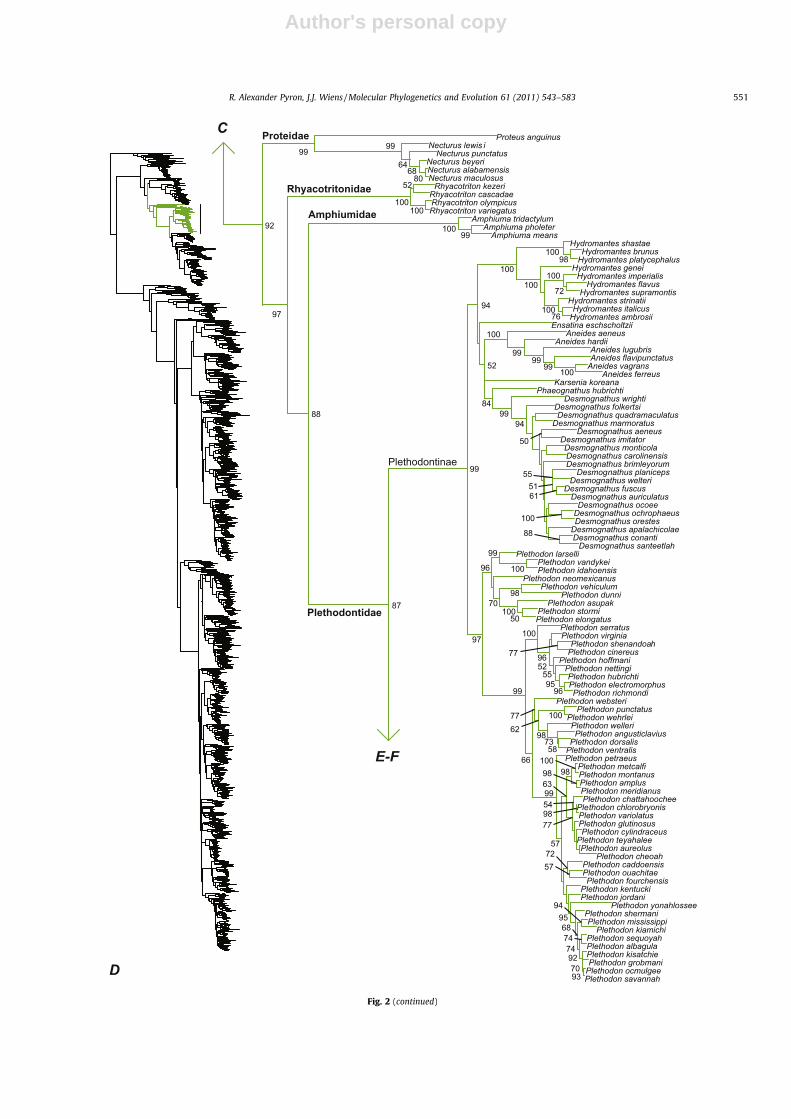
ASW). However, our results differ strongly from Frost et al.(2006) with respect to relationships among the salamander fami-lies. Frost et al. (2006) recover a clade comprising Sirenidae, Dica-mptodontidae, Ambystomatidae, and Salamandridae. In contrast,we find strong support for a sister-group relationship betweenSirenidae and all salamanders exclusive of Cryptobranchidae andHynobiidae (Figs. 1 and 2B and C), as do Wiens et al. (2005a), Roe-lants et al. (2007), and Wiens (2007a, 2011). While some classifica-tions recognize only two subfamilies in Plethodontidae(Hemidactylinae and Plethodontinae; AW), we follow most recentauthors (Chippindale et al., 2004; Kozak et al., 2009; Vieites et al.,2011; Wiens, 2007a) and ASW in recognizing four subfamilies inPlethodontidae (Bolitoglossinae, Hemidactylinae, Plethodontinae,and Spelerpinae; Figs. 1 and 2D–F). We do not recognize a separatesubfamily for Protohynobius (contra AW), given that it is likelynested in Hynobiinae (Peng et al., 2010).
Within frogs, strongly-supported higher-level groups and rela-tionships (e.g., among the non-neobatrachian frogs, Neobatrachia,Hyloidea, and Ranoidea) are consistent with most recent studies(Frost et al., 2006; Roelants et al., 2007; Wiens, 2007a, 2011), withsome notable exceptions. We find (Fig. 2G) strong support for plac-ing Discoglossoidea (Alytidae + Bombinatoridae + Discoglossidae)as the sister group to all other frogs exclusive of Ascaphidae + Leio-pelmatidae (see also Roelants et al., 2007), whereas Frost et al.(2006) and Wiens (2007a, 2011) placed Pipoidea (Pipidae + Rhin-ophrynidae) in this position. Within Neobatrachia (Fig. 1), we cor-roborate the placement of Heleophrynidae as the sister taxon to allother neobatrachian frogs (Frost et al., 2006; Roelants et al., 2007;Wiens, 2007a, 2011). We find a weakly supported sister-group rela-tionship between the clade Sooglossidae + Nasikabatrachidae andall neobatrachian frogs to the exclusion of Heleophrynidae (Fig. 1).In contrast, both Roelants et al. (2007) and Wiens (2007a, 2011; Soo-glossidae only) placed this clade as the sister-taxon to Ranoidea,whereas Frost et al. (2006) found it to be the sister-group of Hyloidea(including Myobatrachidae + Calyptocephalellidae).
Within Hyloidea, our results suggest that several families cur-rently recognized by both AW and ASW are not monophyletic.All of these problematic taxa were previously placed in the familyLeptodactylidae (subdivided extensively by Frost et al., 2006), andinclude Ceratophryidae, Cycloramphidae, Leptodactylidae, andStrabomantidae. Below we describe the specific problems andour proposed taxonomic solutions. These solutions include recog-nizing several additional families relative to current classifications(Alsodidae, Batrachylidae, Odontophrynidae, Rhinodermatidae,Telmatobiidae) and synonymizing one (Strabomantidae withCraugastoridae). These newly recognized families are either re-def-initions of previously recognized families (Rhinodermatidae,Telmatobiidae), or elevation of existing taxa presently below fam-ily rank (Alsodinae, Batrachylinae, Odontophrynini) to the rank offamilies.
Most of these problematic taxa are contained in a weakly sup-ported clade (Fig. 2Z) that includes the currently recognized familiesCeratophryidae, Cycloramphidae, and Hylodidae (note that thecontent of these families and their subfamilies are the same in bothAW and ASW). Under these classifications, Ceratophryidae ispresently divided into the subfamilies Batrachylinae (Atelognathus,Batrachyla), Ceratophryinae (Ceratophrys, Chacophrys, Lepidobatra-chus), and Telmatobiinae (Telmatobius, including Batrachophrynusin AW). Cycloramphidae comprises Alsodinae (Alsodes, Eupsophus,Hylorina, Insuetophrynus, Limnomedusa, Macrogenioglottus, Odon-tophrynus, Proceratophrys, Thoropa) and Cycloramphinae (Crossodac-tylodes, Cycloramphus, Rhinoderma, Zachaenus), and one genus ofuncertain placement (Rupirana). Hylodidae contains Crossodactylus,Hylodes, and Megaelosia. With respect to this classification, we findstrong support for monophyly of Hylodidae (Fig. 2Z), but the familiesCeratophryidae and Cycloramphidae are non-monophyletic, with
546 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
Scaphiopodidae
Heleophrynidae
RhinodermatidaeBrevicipitidae
Gastrophryninae
Salamandrininae
Pipidae
Leptopelinae
Ranixalidae
Ptychadenidae
Buergeriinae
Brachycephalidae
Scolecomorphinae
Hynobiidae
Otophryninae
Microhylinae
Eleutherodactylinae
Dyscophinae
Bufonidae
Cryptobranchidae
Hemiphractidae
Hyperoliidae
Cycloramphidae
Pleurodelinae
Phrynomerinae
Scaphiophryninae
Dicroglossinae
Sirenidae
Limnodynastinae
Cophylinae
Sooglossidae
Ceuthomantidae
Ceratobatrachidae
Alytidae
Hyalinobatrachinae
Rhinatrematidae
Kalophryninae
OdontophrynidaeAlsodidae
Conrauidae
Nasikabatrachidae
BolitoglossinaeLeiopelmatidae
Rhacophorinae
Pelobatidae
Hylinae
Discoglossidae
Phyllomedusinae
Leuiperinae
Batrachylidae
Rhyacotritonidae
Phyzelaphryninae
Ascaphidae
Hylodidae
Phrynobatrachidae
Salamandrinae
Arthroleptinae
Ambystomatidae
Mantellinae
Hemidactylinae
Petropedetidae
Telmatobiidae
Boophinae
Ichthyophiidae
Rhinophrynidae
Nyctibatrachidae
Asterophryninae
Ranidae
Cacosterninae
Melanobatrachinae
Dicamptodontidae
Astylosterninae
Craugastorinae
Hoplophryninae
Micrixalidae
Pelodytidae
Laliostominae
Calyptocephalellidae
Pelodryadinae
AllophrynidaeCentroleninae
Spelerpinae
Paratelmatobiinae
Caeciliinae
Dendrobatidae
Holoadeninae
Megophryidae
Pyxicephalinae
Leptodactylinae
Herpelinae
Plethodontinae
Ceratophryidae
Pristimantinae
AmphiumidaeProteidae
Hemisotidae
Bombinatoridae
Occidozyginae
Strabomantinae
Myobatrachinae
97
100
100
96
92
99
93
97
63
99
91
92 99
98
56
99
98
71
98
100
98
88
54
86
85
100
76
77
100
100
71
100
100
83
100
96
87
100
100
54
98
79
98
57
96
74
98
99
100
60
50
97
98
100
54
98
100
100
97
98100
88
100
97
59
53
100
0.2 subst./site
Ranoidea
Hyloidea
Caeciliidae
Salamandridae
Plethodontidae
Myobatrachidae
Eleutherodactylidae
Craugastoridae
Hylidae
Centrolenidae
Leptodactylidae
Arthroleptidae
Microhylidae
Pyxicephalidae
Dicroglossidae
Rhacophoridae
Mantellidae
Gymnophiona
Caudata
Anura
Neobatrachia
Fig. 1. Skeletal representation of the 2871-species tree from maximum likelihood analysis, with tips representing families and subfamilies based on our taxonomic revision.Numbers at nodes are BS proportions greater than 50%. The full version of this tree is presented in Fig. 2, with multiple panels indicated by bold italic letters.
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 547

Author's personal copy
Dermophis mexicanus
Dermophis oaxacae
Dermophis parviceps
Gymnopis multiplicata
Schistometopum thomense
Schistometopum gregorii
Geotrypetes seraphini
Siphonops annulatus
Siphonops paulensis
Siphonops hardyi
Luetkenotyphlus brasiliensis
Grandisonia sechellensis
Grandisonia larvata
Grandisonia brevis
Grandisonia alternans
Praslinia cooperi
Hypogeophis rostratus
Gegeneophis seshachari
Gegeneophis ramaswamii
Oscaecilia ochrocephala
Caecilia volcani
Caecilia tentaculata
Typhlonectes natans
Chthonerpeton indistinctum
Boulengerula taitana
Boulengerula uluguruensis
Boulengerula boulengeri
Herpele squalostoma
Scolecomorphus vittatus
Scolecomorphus uluguruensis
Crotaphatrema tchabalmbaboensis
Ichthyophis bannanicus
Caudacaecilia asplenia
Ichthyophis glutinosus
Ichthyophis orthoplicatus
Ichthyophis tricolor
Ichthyophis bombayensis
Uraeotyphlus narayani
Rhinatrema bivittatum
Epicrionops niger
Epicrionops marmoratus
100
98
100
59
98
97
52
62
99
100
69
100
51
99
56
62
94
92
55
99
100
100
70
98
88100
95100
100
100
100
100
54
73Rhinatrematidae
Ichthyophiidae
Scolecomorphinae
Herpelinae
Caeciliidae
Gymnophiona
Caeciliinae
A
Fig. 2. Large-scale maximum likelihood estimate of amphibian phylogeny, containing 2871 species represented by up to 12,871 bp of sequence data from 12 genes (threemitochondrial, nine nuclear). Numbers at nodes are maximum likelihood BS proportions greater than 50%. A skeletal version of this tree at the subfamily level is presented inFig. 1. Bold italic letters indicate figure panels.
548 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
Hynobius takedaiHynobius nigrescens
Hynobius tokyoensisHynobius abeiHynobius lichenatus
Hynobius stejnegeriHynobius nebulosus
Hynobius tsuensisHynobius okiensis
Hynobius dunniHynobius quelpaertensis
Hynobius yangiHynobius leechii
Hynobius maoershanensisHynobius chinensisHynobius guabangshanensisHynobius yiwuensis
Hynobius hidamontanusHynobius katoi
Hynobius amjiensisHynobius naeviusHynobius sonaniHynobius arisanensisHynobius formosanus
Hynobius glacialisHynobius fuca
Hynobius kimuraeHynobius boulengeri
Hynobius retardatusBatrachuperus karlschmidti
Batrachuperus tibetanusBatrachuperus londongensis
Batrachuperus pinchoniiBatrachuperus yenyuanensis
Liua shihiLiua tsinpaensis
Pseudohynobius shuichengensisPseudohynobius flavomaculatus
Paradactylodon persicusParadactylodon gorganensis
Paradactylodon mustersiRanodon sibiricus
Salamandrella keyserlingiiPachyhynobius shangchengensis
Onychodactylus japonicusOnychodactylus fischeri
Andrias davidianusAndrias japonicus
Cryptobranchus alleganiensis
100
100
100
58
99
95
86
54
95
6198
59
10061
50
9399
74
100
75
97
93100
98
54
100
100
79
100
74
100
100100
Cryptobranchidae
Hynobiidae
99 Caudata
BC-F
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 549

Author's personal copy
Paramesotriton fuzhongensisParamesotriton deloustali
Paramesotriton hongkongensisParamesotriton chinensis
Paramesotriton zhijinensisParamesotriton caudopunctatusCynops orientalis
Cynops orphicusCynops cyanurusCynops ensicauda
Cynops pyrrhogasterPachytriton brevipesPachytriton labiatusLaotriton laoensis
Ichthyosaura alpestrisEuproctus montanus
Euproctus platycephalusTriturus cristatus
Triturus kareliniiTriturus carn ifex
Triturus dobrogicusTriturus pygmaeus
Triturus marmoratusCalotriton asper
Calotriton arnoldiNeurergus microspilotusNeurergus crocatusNeurergus kaiseriNeurergus strauchiiOmmatotriton ophryticus
Ommatotriton vittatusLissotriton montandoniLissotriton vulgaris
Lissotriton helveticusLissotriton italicus
Lissotriton boscaiNotophthalmus viridescens
Notophthalmus perstriatusNotophthalmus meridionalisTaricha granulosa
Taricha torosaTaricha rivularis
Tylototriton kweichowensisTylototriton verrucosusTylototriton shanjingTylototriton taliangensis
Tylototriton wenxianensisTylototriton asperrimusEchinotriton chinhaiensisEchinotriton andersoniPleurodeles nebulosusPleurodeles poiretiPleurodeles waltlLyciasalamandra billae
Lyciasalamandra antalyanaLyciasalamandra atifiLyciasalamandra flavimembris
Lyciasalamandra helverseniLyciasalamandra fazilae
Lyciasalamandra luschaniSalamandra algira
Salamandra salamandraSalamandra infraimmaculataSalamandra lanzai
Salamandra corsicaSalamandra atra
Mertensiella caucasicaChioglossa lusitanica
Salamandrina perspicillataSalamandrina terdigitata
Ambystoma jeffersonianumAmbystoma laterale
Ambystoma maculatumAmbystoma talpoideum
Ambystoma macrodactylumAmbystoma barbouriAmbystoma texanum
Ambystoma mabeeiAmbystoma ordinarium
Ambystoma andersoniAmbystoma mexicanumAmbystoma dumeriliiAmbystoma tigrinumAmbystoma californiense
Ambystoma opacumAmbystoma gracile
Ambystoma cingulatumDicamptodon copeiDicamptodon aterrimus
Dicamptodon ensatusDicamptodon tenebrosus
Siren lacertinaSiren intermedia
Pseudobranchus axanthusPseudobranchus striatus
7198
100
85
98
97
95
8688
65
68826582
59
65 5797
9193
100
74
85100
1009166
100100
97
9988100
100
100
100
99100
82
100100
99
99816464
91
100100
91
100
10064
74100
92
6790
100
100
100
58
62100
93686370
100
8775
100
100
100
97
Sirenidae
Dicamptodontidae
Ambystomatidae
Salamandridae
SalamandrininaeSalamandrinae
Pleurodelinae
Paramesotriton guangxiensis98
B
D-F
C
Fig. 2 (continued)
550 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583
Author's personal copy
Plethodon savannahPlethodon ocmulgeePlethodon grobmaniPlethodon kisatchiePlethodon albagulaPlethodon sequoyah
Plethodon kiamichiPlethodon mississippi
Plethodon shermaniPlethodon yonahlossee
Plethodon jordaniPlethodon kentucki
Plethodon fourchensisPlethodon ouachitaePlethodon caddoensis
Plethodon cheoahPlethodon aureolus
Plethodon teyahaleePlethodon cylindraceus
Plethodon glutinosusPlethodon variolatusPlethodon chlorobryonis
Plethodon chattahoocheePlethodon meridianusPlethodon amplusPlethodon montanusPlethodon metcalfi
Plethodon petraeusPlethodon ventralisPlethodon dorsalis
Plethodon angusticlaviusPlethodon welleri
Plethodon wehrleiPlethodon punctatus
Plethodon websteriPlethodon richmondi
Plethodon electromorphusPlethodon hubrichti
Plethodon nettingiPlethodon hoffmani
Plethodon cinereusPlethodon shenandoah
Plethodon virginiaPlethodon serratus
Plethodon elongatusPlethodon stormi
Plethodon asupakPlethodon dunni
Plethodon vehiculumPlethodon neomexicanus
Plethodon idahoensisPlethodon vandykei
Plethodon larselliDesmognathus santeetlah
Desmognathus conantiDesmognathus apalachicolaeDesmognathus orestesDesmognathus ochrophaeusDesmognathus ocoee
Desmognathus auriculatusDesmognathus fuscus
Desmognathus welteriDesmognathus planiceps
Desmognathus brimleyorumDesmognathus carolinensisDesmognathus monticola
Desmognathus imitatorDesmognathus aeneus
Desmognathus marmoratusDesmognathus quadramaculatus
Desmognathus folkertsiDesmognathus wrighti
Phaeognathus hubrichtiKarsenia koreana
Aneides ferreusAneides vagransAneides flavipunctatusAneides lugubris
Aneides hardiiAneides aeneus
Ensatina eschscholtziiHydromantes ambrosiiHydromantes italicus
Hydromantes strinatiiHydromantes supramontis
Hydromantes flavusHydromantes imperialis
Hydromantes geneiHydromantes platycephalusHydromantes brunus
Hydromantes shastaeAmphiuma means
Amphiuma pholeterAmphiuma tridactylum
Rhyacotriton variegatusRhyacotriton olympicusRhyacotriton cascadae
Rhyacotriton kezeriNecturus maculosusNecturus alabamensisNecturus beyeri
Necturus punctatusNecturus lewis i
Proteus anguinus
92
97
88
99
97
99
66
99
9568
74927093
74
94
7257
63
77
57
5498
98 98100
7762
987358
100
100
9652
5595
96
77
96
70100
50
98
99
100
94
52
8499
94
88
100
555161
50
100
9999
99 100
100
100
10076
100
72
10098
10099
100100
52
9999
6468
80
Proteidae
Rhyacotritonidae
Amphiumidae
Plethodontidae87
Plethodontinae
D
C
E-F
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 551

Author's personal copy
Oedipina uniformisOedipina pacificensis
Oedipina gracilisOedipina leptopoda
Oedipina poelziOedipina cyclocauda
Oedipina pseudouniformisOedipina collaris
Oedipina grandisOedipina stenopodia
Oedipina alleniOedipina savagei
Oedipina complexOedipina parvipes
Oedipina maritimaOedipina elongataOedipina carablanca
Oedipina gephyraOedipina kasiosOedipina quadra
Bradytriton silusNototriton abscondensNototriton gameziNototriton picadoiNototriton guanacaste
Nototriton richardiNototriton lignicolaNototriton limnospectator
Nototriton barbouriNototriton brodiei
Cryptotriton veraepacisCryptotriton nasalis
Cryptotriton alvarezdeltoroiNyctanolis pernix
Dendrotriton rabbiChiropterotriton chondrostega
Chiropterotriton terrestrisChiropterotriton priscus
Chiropterotriton magnipesChiropterotriton cracensChiropterotriton multid entatus
Chiropterotriton arboreusChiropterotriton lavae
Chiropterotriton orculusChiropterotriton dimidiatus
Thorius dubitusThorius troglodytes
Thorius minutissimusBatrachoseps nigriventris
Batrachoseps simatusBatrachoseps gregariusBatrachoseps attenuatusBatrachoseps pacificus
Batrachoseps majorBatrachoseps gavilanensisBatrachoseps gabrieli
Batrachoseps relictusBatrachoseps kawia
Batrachoseps regiusBatrachoseps diabolicus
Batrachoseps campiBatrachoseps wrighti
Eurycea latitansEurycea tridentiferaEurycea pterophilaEurycea neotenesEurycea nana
Eurycea sosorumEurycea troglodytesEurycea rathbuniEurycea waterlooensis
Eurycea chisholmensisEurycea tonkawaeEurycea naufragia
Eurycea quadridigitataEurycea aquatica
Eurycea junaluskaEurycea bislineata
Eurycea wilderaeEurycea cirrigera
Eurycea longicaudaEurycea lucifuga
Haideotriton wallaceiEurycea tynerensisEurycea spelaea
Eurycea multiplicataUrspelerpes brucei
Pseudotriton montanusPseudotriton ruber
Stereochilus marginatusGyrinophilus palleucusGyrinophilus gulolineatus
Gyrinophilus porphyriticusHemidactylium scutatum
87
83
98
93
62
77
5250
72
7467
100
6699
8698
100
967765
6153
95
94
58
61
9164 98
6777
10094
92
100
5310084
53
6299
96
100
100
98
97
99
69
86100
9985100
53886576
100
100
61
93
100
10068
100
71 100
100
100
Hemidactylinae
Spelerpinae
Bolitoglossinae
Plethodontidae cont.
Bolitoglossinae cont.
E
Fig. 2 (continued)
552 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
Bolitoglossa carriBolitoglossa conantiBolitoglossa diaphora
Bolitoglossa dunniBolitoglossa flavimembris
Bolitoglossa morioBolitoglossa synoria
Bolitoglossa celaqueBolitoglossa longissima
Bolitoglossa porrasorumBolitoglossa decoraBolitoglossa lincolni
Bolitoglossa frankliniBolitoglossa riletti
Bolitoglossa hermosaBolitoglossa zapoteca
Bolitoglossa macriniiBolitoglossa oaxacensis
Bolitoglossa helmrichiBolitoglossa rostrata
Bolitoglossa engelhardtiBolitoglossa dofleini
Bolitoglossa alvaradoiBolitoglossa palmata
Bolitoglossa equatorianaBolitoglossa peruviana
Bolitoglossa altamazonicaBolitoglossa medemi
Bolitoglossa adspersaBolitoglossa simaBolitoglossa biseriata
Bolitoglossa paraensisBolitoglossa colonnea
Bolitoglossa schizodactylaBolitoglossa robusta
Bolitoglossa minutulaBolitoglossa sooyorumBolitoglossa mar morea
Bolitoglossa epimelaBolitoglossa cerroensis
Bolitoglossa pesrubraBolitoglossa gracilis
Bolitoglossa subpalmataBolitoglossa mombachoensisBolitoglossa striatula
Bolitoglossa yucatanaBolitoglossa lignicolor
Bolitoglossa mexicanaBolitoglossa odonnelli
Bolitoglossa flaviventrisBolitoglossa platydactyla
Bolitoglossa occidentalisBolitoglossa rufescens
Bolitoglossa hartwegiPseudoeurycea brunnataPseudoeurycea goebeli
Pseudoeurycea exspectataPseudoeurycea anitae
Pseudoeurycea cochranaePseudoeurycea melanomolga
Pseudoeurycea gadoviiPseudoeurycea robertsi
Pseudoeurycea altamontanaPseudoeurycea longicauda
Pseudoeurycea tenchalliPseudoeurycea smithiPseudoeurycea papenfussi
Ixalotriton parvusIxalotriton niger
Pseudoeurycea rexPseudoeurycea conanti
Pseudoeurycea obesaPseudoeurycea werleri
Pseudoeurycea mystaxPseudoeurycea nigromaculata
Pseudoeurycea saltatorPseudoeurycea juarezi
Pseudoeurycea ruficaudaPseudoeurycea unguidentis
Pseudoeurycea leprosaPseudoeurycea firscheini
Pseudoeurycea lynchiLineatriton lineolus
Lineatriton orchileucosPseudoeurycea gigantea
Pseudoeurycea naucampatepetlPseudoeurycea belliiPseudoeurycea maxima
Pseudoeurycea bonetiPseudoeurycea scandensPseudoeurycea galeanae
Pseudoeurycea cephalicaParvimolge townsendi
88
99
52
96
90
8452
100
100
58
100
99
52 100
100
100
92
90
99
94
93
80
68
100
80
9558
100
59
92
8669
88 94
100
96
70
57
88
69
886097
94 100
59
87
71 100
70
8974 86
74
9065 87
96
91
68
Bolitoglossinae cont.
F
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 553

Author's personal copy
Oreolalax nanjiangensisOreolalax popeiOreolalax omeimontisOreolalax chuanbeiensisOreolalax multipunctatus
Oreolalax liangbeiensisOreolalax majorOreolalax xiangchengensis
Oreolalax jingdongensisOreolalax rugosus
Oreolalax schmidtiOreolalax pingiiOreolalax lichuanensisOreolalax rhodostigmatusScutiger tuberculatusScutiger muliensisScutiger mammatus
Scutiger boulengeriScutiger glandulatusScutiger chintingensis
Leptobrachium chapaenseLeptobrachium huashen
Leptobrachium ailaonicumVibrissaphora echinataLeptobrachium leishanense
Leptobrachium liuiLeptobrachium boringii
Leptobrachium promustacheLeptobrachium mouhotiLeptobrachium ngoclinhense
Leptobrachium hainanenseLeptobrachium xanthospilum
Leptobrachium banaeLeptobrachium montanum
Leptobrachium gunungenseLeptobrachium smithiLeptobrachium hasseltii
Leptolalax bourretiLeptolalax pelodytoidesLeptolalax liuiLeptolalax oshanensis
Leptolalax arayaiLeptolalax pictus
Brachytarsophrys platyparietusBrachytarsophrys feae
Xenophrys shapingensisXenophrys nankiangensisXenophrys omeimontis
Xenophrys spinataXenophrys minorXenophrys majorMegophrys lekaguli
Megophrys nasutaXenophrys baluensis
Ophryophryne hansiOphryophryne microstoma
Pelobates varaldiiPelobates cultripes
Pelobates fuscusPelobates syriacus
Pelodytes punctatusPelodytes ibericus
Pelodytes caucasicusSpea intermontana
Spea bombifronsSpea hammondii
Spea multiplicataScaphiopus holbrookiiScaphiopus hurterii
Scaphiopus couchiiXenopus amietiXenopus longipesXenopus ruwenzoriensis
Xenopus largeniXenopus wittei
Xenopus fraseriXenopus pygmaeusXenopus andreiXenopus boumbaensisXenopus victorianusXenopus petersii
Xenopus laevisXenopus gilli
Xenopus vestitusXenopus clivii
Xenopus muelleriXenopus borealis
Silurana tropicalisSilurana epitropicalis
Hymenochirus boettgeriPipa parva
Pipa pipaPipa carvalhoi
Rhinophrynus dorsalisDiscoglossus jeanneae
Discoglossus galganoiDiscoglossus pictus
Discoglossus sardusDiscoglossus montalentii
Alytes muletensisAlytes dickhilleni
Alytes maurusAlytes obstetricans
Alytes cisternasiiBombina variegataBombina pachypus
Bombina bombinaBombina orientalis
Bombina fortinuptialisBombina microdeladigitoraBombina lichuanensisBombina maxima
Barbourula busuangensisBarbourula kalimantanensis
Leiopelma pakekaLeiopelma hamiltoniLeiopelma archeyi
Leiopelma hochstetteriAscaphus trueiAscaphus montanus
99
98
92
100
96
99
100
100
100
90
100
80
66
100100100
97
9985
57
100
95
100
7859100
93
99100
79
100
100
90
100
675899
100100 100
10097
99
81 100
100
94100
99100
95
100
100
10059100
100 100
100 100
8677
100
99
100100
100100
100
61
50
100
58
69
9597
100
77
100
100
100
100
100
100 958486
100100
7264
100
100
60100
9962
86
100
9899
99
99
100
98
Ascaphidae
LeiopelmatidaeBombinatoridae
Alytidae
DiscoglossidaeRhinophrynidae
Pipidae
Scaphiopodidae
Pelodytidae
Pelobatidae
Megophryidae
Anura
H-AE
G
Fig. 2 (continued)
554 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
Hyperolius puncticulatusHyperolius montanusHyperolius zonatusHyperolius concolorHyperolius molleriHyperolius thomensisHyperolius cinnamomeoventris
Hyperolius torrentisHyperolius chlorosteusHyperolius baumanniHyperolius picturatusHyperolius cystocandicansHyperolius frontalis
Hyperolius castaneusHyperolius lateralis
Hyperolius alticolaHyperolius ocellatusHyperolius mosaicus
Hyperolius kivuensisAlexteroon obstetricans
Hyperolius tuberilinguisHyperolius marmoratusHyperolius viridiflavusHyperolius angolensisHyperolius tuberculatus
Hyperolius phantasticusHyperolius glandicolorHyperolius argus
Hyperolius fusciventrisHyperolius guttulatus
Hyperolius pardalisHyperolius nasutusHyperolius acuticeps
Hyperolius pusillusHyperolius horstockii
Hyperolius semidiscusHeterixalus variabilisHeterixalus tricolorHeterixalus andrakata
Heterixalus carboneiHeterixalus betsileoHeterixalus rutenbergi
Heterixalus luteostriatusHeterixalus punctatusHeterixalus alboguttatusHeterixalus boettgeriHeterixalus madagascariensis
Tachycnemis seychellensisAfrixalus paradorsalisAfrixalus dorsalis
Afrixalus laevisAfrixalus fornasini
Afrixalus stuhlmanniAfrixalus delicatus
Afrixalus knysnaeMorerella cyanophthalmaOpisthothylax immaculatusAcanthixalus spinosusAcanthixalus sonjae
Kassina maculataPhlyctimantis verrucosusPhlyctimantis leonardiSemnodactylus wealiiKassina senegalensisCryptothylax greshoffii
Arthroleptis poecilonotusArthroleptis adelphus
Arthroleptis krokosuaArthroleptis variabilis
Arthroleptis stenodactylusArthroleptis affinisArthroleptis nikeaeArthroleptis reichei
Arthroleptis tanneriArthroleptis francei
Arthroleptis wahlbergiiArthroleptis aureoli
Arthroleptis sylvaticusArthroleptis taeniatusArthroleptis schubotzi
Arthroleptis xenodactyloidesArthroleptis xenodactylus
Cardioglossa gracilisCardioglossa schioetzi
Cardioglossa manengoubaCardioglossa oreasCardioglossa pulchraCardioglossa occidentalis
Cardioglossa leucomystaxCardioglossa gratiosa
Cardioglossa elegansLeptopelis concolorLeptopelis bocagiiLeptopelis palmatusLeptopelis kivuensis
Leptopelis natalensisLeptopelis vermiculatus
Leptopelis modestusLeptopelis argenteus
Leptopelis brevirostrisAstylosternus batesiAstylosternus schioetziAstylosternus diadematusTrichobatrachus robustusNyctibates corrugatus
Scotobleps gabonicusLeptodactylodon bicolor
Probreviceps macrodactylusProbreviceps uluguruensis
Probreviceps durirostrisSpelaeophryne methneri
Callulina krefftiCallulina kisiwamsitu
Breviceps fichusBreviceps mossambicusBreviceps fuscus
Hemisus marmoratusSooglossus thomassetiSooglossus sechellensis
Sechellophryne pipilodryasSechellophryne gardineri
Nasikabatrachus sahyadrensisHeleophryne purcelliHeleophryne regis
Hadromophryne natalensis
98
56
97
98
99
97
82
84
100
6171
100100
5064
92
72
868054
100
99
7794848897
98
56
97
56
96
10098
67
10093
55
100100
99 99 100
100
100
100
100
8263
63
99
100
100
85
69
75100
68
10099
55
86
100
100
51100
100
100
71
99
5673
87
94
80
68
5998100
98
100
100
10060
9897
100
100
98
100
10099
100
100
99Heleophrynidae
Nasikabatrachidae
Sooglossidae
Hemisotidae
Brevicipitidae
Astylosterninae
Leptopelinae
Arthroleptinae
Arthroleptidae
Hyperoliidae
G
HIJ-OP-AE
100
77
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 555

Author's personal copy
Callulops pulliferCallulops eurydactylus
Callulops slateriAsterophrys turpicola
Xenorhina oxycephalaXenorhina varia
Xenorhina bouwensiXenorhina lanthanites
Barygenys flavigularisHylophorbus picoidesHylophorbus tetraphonusHylophorbus wondiwoi
Hylophorbus nigrinusCallulops robustusHylophorbus rufescens
Sphenophryne cornutaLiophryne dentata
Xenorhina obesaOxydactyla crassa
Liophryne rhododactylaOreophryne atrigularisOreophryne wapoga
Oreophryne brachypusOreophryne unicolor
Oreophryne pseudasplenicolaOreophryne asplenicola
Cophixalus balbusOreophryne sibilans
Oreophryne wairaOreophryne clamata
Cophixalus tridactylusCophixalus humicola
Aphantophryne pansaAlbericus laurini
Choerophryne rostelliferBarygenys exsul
Cophixalus sphagnicolaCopiula obsti
Copiula pipiensAustrochaperina derongo
Copiula majorLiophryne schlaginhaufeni
Genyophryne thomsoniMelanobatrachus indicus
Kalophrynus pleurostigmaKalophrynus intermedius
Kalophrynus baluensisMicrohyla ornata
Microhyla fissipesMicrohyla okinavensis
Microhyla borneensisMicrohyla heymonsiMicrohyla rubraMicrohyla pulchraMicrohyla butleri
Calluella guttulataGlyphoglossus molossus
Chaperina fuscaMicryletta inornata
Ramanella obscuraRamanella variegata
Uperodon systomaKaloula taprobanica
Kaloula conjunctaKaloula pulchra
Metaphrynella sundanaDyscophus antongiliiDyscophus guineti
Dyscophus insularisScaphiophryne menabensisScaphiophryne madagascariensisScaphiophryne boribory
Scaphiophryne marmorataScaphiophryne gottlebei
Scaphiophryne calcarataScaphiophryne spinosaScaphiophryne brevis
Paradoxophyla tiaranoParadoxophyla palmata
Rhombophryne laevipesRhombophryne alluaudi
Rhombophryne testudoRhombophryne minuta
Rhombophryne serratopalpebrosaRhombophryne coronata
Rhombophryne coudreauiStumpffia helenae
Plethodontohyla notostictaPlethodontohyla guentheriPlethodontohyla fonetana
Plethodontohyla mihanikaCophyla berara
Cophyla phyllodactylaStumpffia roseifemoralis
Stumpffia grandisStumpffia tetradactyla
Stumpffia psologlossaStumpffia gimmeli
Stumpffia pygmaeaStumpffia tridactyla
Platypelis barbouriPlatypelis pollicaris
Platypelis tuberiferaPlatypelis mavomavo
Platypelis millotiPlatypelis grandis
Plethodontohyla ocellataPlethodontohyla brevipes
Plethodontohyla bipunctataPlethodontohyla tuberata
Plethodontohyla inguinalisAnodonthyla hutchisoniAnodonthyla boulengerii
Anodonthyla nigrigularisAnodonthyla moramora
Anodonthyla montanaAnodonthyla rouxae
Hoplophryne uluguruensisHoplophryne rogersi
Gastrophryne olivaceaGastrophryne carolinensis
Gastrophryne elegansHypopachus variolosus
Elachistocleis ovalisDermatonotus muelleri
Hamptophryne bolivianaDasypops schirchi
Ctenophryne geayiNelsonophryne aequatorialis
Chiasmocleis shudikarensisChiasmocleis hudsoni
Synapturanus mirandaribeiroiOtophryne pyburni
Phrynomantis micropsPhrynomantis bifasciatus
Phrynomantis annectens
97
99
6693
7898 100
90
68
97 74 67
100100100
100100
58
6794 100
100
76
78
100100
7388 99
87
100
100
92 95 100
81100 100
80
9985 98
65100
7263
95
100
90
51 95
6080
78
100
9876
9964
100
100
847154 82
61
85100
100
9595 90 100
88100
96
100100
729485 100
9910071
99100
Microhylidae
Asterophryninae
Melanobatrachinae
Kalophryninae
Dyscophinae
Microhylinae
Scaphiophryninae
Cophylinae
Hoplophryninae
Gastrophryninae
Phrynomerinae
Otophryninae
I
Fig. 2 (continued)
556 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
Platymantis cryptotisPlatymantis pelewensis
Platymantis papuensisPlatymantis weberi
Platymantis wuenscheorumPlatymantis bimaculatus
Platymantis vitiensisDiscodeles guppyi
Platymantis punctatusCeratobatrachus guentheri
Batrachylodes vertebralisPlatymantis naomiiPlatymantis mimulusPlatymantis dorsalis
Platymantis corrugatusPlatymantis hazelaePlatymantis montanus
Ingerana baluensisNyctibatrachus majorNyctibatrachus aliciae
Lankanectes corrugatusArthroleptella subvoceArthroleptella bicolorArthroleptella landdrosiaArthroleptella drewesiiArthroleptella villiersiArthroleptella lightfootiNatalobatrachus bonebergi
Amietia vertebralisAmietia angolensisStrongylopus grayiiAmietia fuscigula
Cacosternum boettgeriCacosternum capense
Cacosternum nanumCacosternum platysMicrobatrachella capensisPoyntonia paludicola
Strongylopus fasciatusStrongylopus bonaespeiTomopterna tandyi
Tomopterna cryptotisTomopterna damarensis
Tomopterna marmorataTomopterna lugangaTomopterna tuberculosa
Tomopterna delalandiiTomopterna krugerensisTomopterna natalensis
Anhydrophryne rattrayiPyxicephalus edulisPyxicephalus adspersus
Aubria subsigillataPetropedetes newtoniPetropedetes parkeri
Petropedetes cameronensisPetropedetes palmipes
Petropedetes martiensseniPetropedetes yakusini
Conraua crassipesConraua goliathConraua robusta
Phrynobatrachus africanusPhrynobatrachus cricogaster
Phrynobatrachus acridoidesPhrynobatrachus auritus
Phrynobatrachus natalensisPhrynobatrachus disparPhrynobatrachus levelevePhrynobatrachus calcaratusPhrynobatrachus mababiensis
Phrynobatrachus dendrobatesPhrynobatrachus krefftii
Phrynobatrachus sandersoniMicrixalus fuscus
Micrixalus kottigeharensisPtychadena mahnerti
Ptychadena porosissimaPtychadena bibroni
Ptychadena aequiplicataPtychadena cooperiPtychadena oxyrhynchus
Ptychadena telliniiPtychadena anchietae
Ptychadena longirostrisPtychadena subpunctataPtychadena taenioscelisPtychadena pumilioPtychadena newtoniPtychadena mascareniensis
Hildebrandtia ornata
99
50
54
100
100
97
99100
94
69
5769
76
95
9791
89
100
7498
91
74
61
94
100
59
100
9183
55
8088
9978
100
100
96
92
83
5696
95
66
100
76
100
94100
100
100
100
100
99
99
98
62
94
100
100
9999
100
99
82
75
8098
88
9398
10093
Ptychadenidae
Micrixalidae
Phrynobatrachidae
Conrauidae
Petropedetidae
Cacosterninae
Pyxicephalinae
Nyctibatrachidae
Ceratobatrachidae
Pyxicephalidae
JK-O
H
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 557

Author's personal copy
Limnonectes finchiLimnonectes ingeri
Limnonectes malesianusLimnonectes shompenorum
Limnonectes macrodonLimnonectes paramacrodon
Limnonectes poilaniLimnonectes blythii
Limnonectes grunniensLimnonectes ibanorum
Limnonectes palavanensisLimnonectes parvus
Limnonectes leporinusLimnonectes visayanusLimnonectes macrocephalus
Limnonectes woodworthiLimnonectes modestus
Limnonectes heinrichiLimnonectes magnusLimnonectes arathooniLimnonectes microtympanumLimnonectes acanthi
Limnonectes leytensisLimnonectes kuhliiLimnonectes bannaensis
Limnonectes fujianensisLimnonectes fragilis
Limnonectes asperatusLimnonectes gyldenstolpei
Limnonectes dabanusLimnonectes hascheanus
Limnonectes limborgiLimnonectes laticeps
Limnonectes kadarsaniLimnonectes microdiscus
Paa arnoldiPaa chayuensisPaa maculosa
Paa medogensisPaa conaensis
Nanorana parkeriNanorana pleskei
Nanorana ventripunctataPaa liui
Paa yunnanensisPaa bourreti
Chaparana fansipaniChaparana aenea
Chaparana unculuanusChaparana quadranus
Paa taihangnicusPaa liebigii
Paa spinosaPaa exilispinosa
Paa jiulongensisPaa yei
Paa boulengeriPaa verrucospinosa
Paa shiniPaa robertingeri
Paa fasciculispinaChaparana delacouri
Fejervarya granosaFejervarya syhadrensis
Fejervarya caperataFejervarya rufescens
Fejervarya kudremukhensisFejervarya kirtisinghei
Fejervarya greeniiFejervarya pierrei
Fejervarya muddurajaFejervarya limnocharis
Fejervarya sakishimensisFejervarya orissaensis
Fejervarya iskandariFejervarya triora
Fejervarya vittigeraFejervarya cancrivora
Sphaerotheca brevicepsSphaerotheca dobsonii
Hoplobatrachus rugulosusHoplobatrachus tigerinus
Hoplobatrachus crassusHoplobatrachus occipitali s
Euphlyctis ehrenbergiiEuphlyctis cyanophlyctis
Euphlyctis hexadactylusNannophrys marmorata
Nannophrys ceylonensisOccidozyga magnapustulosa
Occidozyga martensiiOccidozyga laevis
Occidozyga baluensisOccidozyga lima
Occidozyga borealisIngerana tenasserimensis
Indirana semipalmataIndirana beddomii
88
99
99
100
100
63
6791
71100 100
100
100
56 100
10078
10069
5765
80100 63
86
6674 100 100
100
100
100
89908099
100
91
8051
57 88100
72100
93
99
99100
100
82
100
100
95
10097
51 58 59
56100
88
54100
8598
100100
100
9797
100 93
87 100
100
100
78
100
100
99Ranixalidae
Occidozyginae
Dicroglossinae
Dicroglossidae
K
Fig. 2 (continued)
558 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583
Author's personal copy
Gephyromantis striatusGephyromantis malagasius
Gephyromantis ventrimaculatusGephyromantis horridus
Gephyromantis klemmeriGephyromantis ambohitra
Gephyromantis rivicolaGephyromantis silvanus
Gephyromantis webbiGephyromantis decaryi
Gephyromantis leucocephalusGephyromantis eiselti
Gephyromantis enkiGephyromantis blanci
Gephyromantis asperGephyromantis boulengeri
Gephyromantis leucomaculatusGephyromantis zavona
Gephyromantis salegyGephyromantis tandroka
Gephyromantis moseriGephyromantis redimitus
Gephyromantis cornutusGephyromantis tschenki
Gephyromantis granulatusGephyromantis pseudoasper
Gephyromantis luteusGephyromantis pliciferGephyromantis sculpturatus
Gephyromantis azzurraeGephyromantis corvus
Mantidactylus femoralisMantidactylus mocquardi
Mantidactylus ambreensisMantidactylus ulcerosus
Mantidactylus op iparisMantidactylus charlotteae
Mantidactylus biporusMantidactylus lugubris
Mantidactylus argenteusMantidactylus grandidieri
Boehmantis microtympanumSpinomantis peraccae
Spinomantis elegansSpinomantis aglavei
Mantella betsileoMantella viridis
Mantella expectataMantella ebenaui
Mantella maneryMantella laevigata
Mantella milotympanumMantella crocea
Mantella aurantiacaMantella pulchra
Mantella madagascariensisMantella nigricans
Mantella haraldmeieriMantella baroni
Mantella cowaniiMantella bernhardi
Wakea madinikaBlommersia kely
Blommersia sarotraBlommersia grandisonae
Blommersia domergueiBlommersia blommersae
Blommersia witteiGuibemantis liberGuibemantis bicalcaratus
Guibemantis albolineatusGuibemantis depressiceps
Guibemantis tornieriBoophis viridis
Boophis rappiodesBoophis sibilans
Boophis microtympanumBoophis occidentalisBoophis albilabris
Boophis vittatusBoophis marojezensisBoophis boehmei
Boophis goudotiiBoophis madagascariensis
Boophis luteusBoophis douliotiBoophis tephraeomystax
Boophis xerophilusBoophis idae
Boophis paulianiAglyptodactylus laticeps
Aglyptodactylus madagascariensisLaliostoma labrosum
100
60
100
52
86
73
5885 100
64
5595
5264
97
99
57
99
62
88
7664
519461 96
9090
10094
99
100
96
91
1009473
74100
93
1009997
100
9392
99
95
99100
96
99100
100
100
100
55 100
100
100
9387
82
87
7894
100
9999
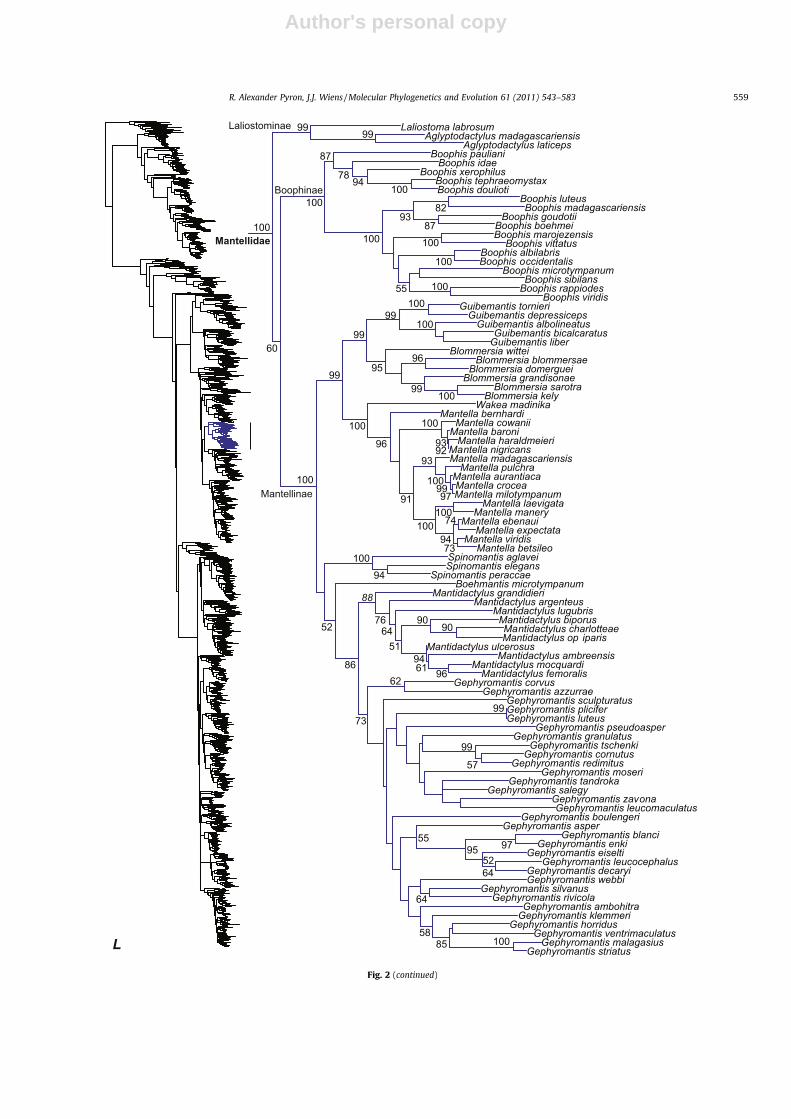
Mantellidae
Laliostominae
Boophinae
Mantellinae
L
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 559
Author's personal copy
Philautus schmardaPhilautus cavirostris
Philautus lunatusPhilautus stuarti
Philautus popularisPhilautus femoralisPhilautus poppiaePhilautus mooreorum
Philautus steineriPhilautus microtympanumPhilautus papillosus
Philautus hoffmanniPhilautus asankai
Philautus pleurotaeniaPhilautus ocularisPhilautus tanu
Philautus mittermeieriPhilautus decoris
Philautus simbaPhilautus leucorhinus
Philautus wynaadensisPhilautus zorro
Philautus tuberohumerusPhilautus bombayensis
Philautus menglaensisPhilautus gryllus
Philautus longchuanensisPhilautus ponmudi
Philautus tinniensPhilautus signatus
Philautus chariusPhilautus griet
Philautus travancoricusPhilautus neelanethrus
Philautus nerostagonaPhilautus graminirupes
Philautus bobingeriPhilautus anili
Philautus glandulosusPhilautus beddomii
Kurixalus hainanusKurixalus odontotarsu sPhilautus carinensis
Kurixalus idiootocusKurixalus eiffingeriPhilautus banaensis
Theloderma molochPhilautus petersi
Philautus mjobergiPhilautus surdusPhilautus acutirostris
Philautus aurifasciatusPhilautus abditus
Philautus ingeriPhilautus quyetiGracixalus gracilipes
Kurixalus jinxiuensisRhacophorus puerensisRhacophorus dugriteiRhacophorus hungfuensis
Rhacophorus minimusRhacophorus huiRhacophorus omeimontis
Rhacophorus taronensisRhacophorus moltrechti
Rhacophorus arboreusRhacophorus schlegeliiRhacophorus nigropunctatus
Rhacophorus chenfuiRhacophorus maximus
Rhacophorus dennysiRhacophorus feae
Rhacophorus lateralisRhacophorus reinwardtii
Rhacophorus kioRhacophorus bipunctatusRhacophorus rhodopus
Rhacophorus calcaneusRhacophorus malabaricusRhacophorus orlovi
Rhacophorus annamensisPolypedates megacephalus
Polypedates mutusPolypedates leucomystaxPolypedates crucigerPolypedates maculatus
Polypedates collettiPolypedates fastigo
Polypedates equesFeihyla palpebralis
Chiromantis xerampelinaChiromantis rufescens
Chiromantis doriaeChiromantis vittatus
Ghatixalus variabilisTheloderma asperum
Theloderma bicolorTheloderma rhododiscus
Theloderma corticaleNyctixalus pictus
Nyctixalus spinosusLiuixalus romeriPhilautus ocellatusPhilautus hainanus
Buergeria robustaBuergeria buergeri
Buergeria japonicaBuergeria oxycephlus
100
100
78
73
62
67
100
98
55
89
99
53
5491 100
100
100
89
5992
7352 100
70100 100
98
9610080
52
8868
86 68
100
98
96
69
62
92
100
5561
6053
89
77
92
7493
9998
5287 99
99
66
94100
8899
5899
7698
85
99
99
96100
58
100
83 98
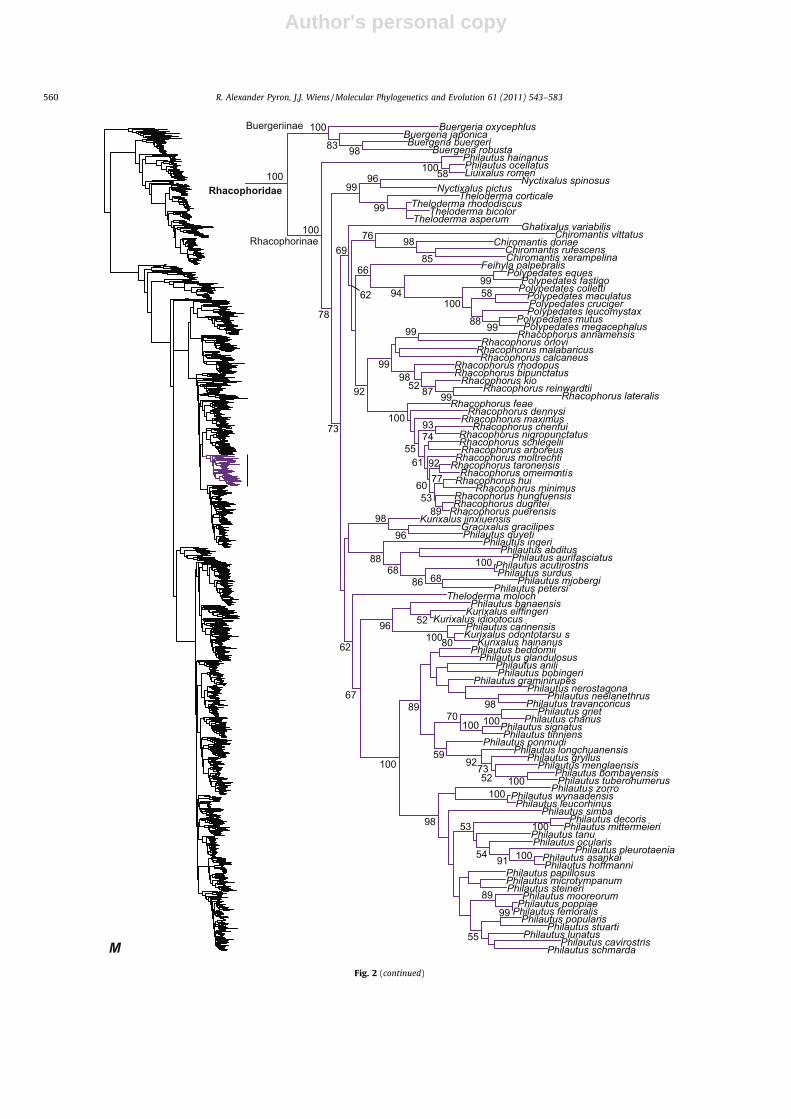
Buergeriinae
Rhacophoridae
Rhacophorinae
M
Fig. 2 (continued)
560 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
Rana laterimaculataRana baramica
Rana glandulosaRana banjarana
Rana signataRana picturata
Rana siberuAmnirana lepus
Amnirana albolabrisAmnirana galamensisHylarana nicobariensis
Rana eschatiaRana parvaccola
Rana labialisRana raniceps
Rana megalonesaRana chalconota
Rana mocquardiiRana spinulosa
Rana maosonensisRana latouchii
Rana cubitalisRana nigrovittataRana faberRana daemeli
Rana jimiensisRana arfaki
Rana milletiRana gracilis
Rana malabaricaRana temporalis
Rana aurantiacaRana miopus
Rana lateralisRana guentheri
Rana macrodactylaRana taipehensis
Rana erythraeaRana luctuosa
Rana rugosaRana emeljanovi
Rana tientaiensisRana minima
Rana luzonensisRana igorota
Rana sanguineaRana bergeri
Rana shqipericaRana cerigensis
Rana bedriagaeRana ridibundaRana kurtmuelleriRana epeiroticaRana cretensis
Rana lessonaeRana esculenta
Rana pereziRana saharica
Rana nigromaculataRana plancyiRana hubeiensis
Rana porosaRana fukienensis
Amolops liangshanensisAmolops loloensisAmolops jinjiangensis
Amolops mantzorumAmolops kangtingensis
Amolops granulosusAmolops lifanensisAmolops viridimaculatus
Amolops chunganensisAmolops bellulus
Amolops marmoratusAmolops panhai
Amolops larutensisAmolops cremnobatus
Amolops rickettiAmolops wuyiensis
Amolops daiyunensisAmolops hongkongensis
Amolops torrentisAmolops hainanensis
Amolops spinapectoralisMeristogenys phaeomerus
Meristogenys jerboaMeristogenys poecilusMeristogenys orphnocnemis
Meristogenys whiteheadiMeristogenys kinabaluensis
Huia cavitympanumHuia sumatrana
Huia masoniiRana alticola
Rana curtipesHuia melasma
Staurois tuberilinguisStaurois parvusStaurois natator
Staurois latopalmatus
100
96
56
71
59
100
67
60
76
60 99
95
10099 98
56
9599
98
97
100
96
100 82
10095
94
100 98
100 100
97
100
807653
8570
71
100
100
1009565
90
9490
9892
93 9891100
71100
97100
96
9399
97100
100100
67
82 969661
53
58
58 100100
100
100 100
Ranidae
Ranidae cont.N
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 561

Author's personal copy
Rana macroglossaRana taylori
Rana brownorumRana spectabilis
Rana forreriRana omiltemana
Rana tlalociRana neovolcanica
Rana berlandieriRana blairi
Rana sphenocephalaRana onca
Rana yavapaiensisRana magnaocularis
Rana montezumaeRana dunni
Rana chiricahuensisRana pipiens
Rana capitoRana sevosa
Rana areolataRana palustris
Rana vaillantiRana juliani
Rana bwanaRana palmipes
Rana warszewitschiiRana vibicaria
Rana maculataRana psilonota
Rana zweifeliRana tarahumarae
Rana pustulosaRana sierramadrensis
Rana clamitansRana okaloosaeRana heckscheri
Rana virgatipesRana grylio
Rana septentrionalisRana catesbeiana
Rana sylvaticaRana longicrus
Rana zhenhaiensisRana omeimontis
Rana chaochiaoensisRana japonica
Rana arvalisRana chensinensis
Rana kukunorisRana huanrensis
Rana piricaRana dybowskii
Rana ornativentrisRana amurensis
Rana kunyuensisRana iberica
Rana italicaRana temporaria
Rana pyrenaicaRana tsushimensis
Rana okinavanaRana holsti
Rana macrocnemisRana graecaRana dalmatina
Rana latasteiRana asiaticaPseudoamolops sauteri
Rana zhengiRana johnsi
Rana tagoiRana cascadaeRana muscosaRana aurora
Rana boyliiRana pretiosaRana luteiventris
Rana shuchinaeRana weiningensis
Odorrana aureolaRana livida
Rana chloronotaRana hosii
Rana morafkaiRana banaorum
Rana tiannanensisRana megatympanum
Rana narinaRana amamiensisRana supranarina
Rana utsunomiyaorumRana swinhoana
Rana versabilisOdorrana nasica
Odorrana tormotaOdorrana schmackeri
Rana hejiangensisRana bacboensisRana ishikawaeRana cucae
Rana compotrixRana vitrea
Rana archotaphusRana daorumRana iriodes
Odorrana jingdongensisRana margaretae
Rana grahamiOdorrana junlianensis
Rana andersoniiRana hmongorum
Odorrana chapaensisRana khalam
Odorrana absitaRana adenopleura
Rana chapaensisRana pleuraden
76
67
98
100
100
99
66
54
7772
9099
9588
9899
79
100100 100
100100
63
72
97100
100
99
95100
10098
73
67
10084
100100
52
92
78
85
8177
9384
9786
57
5599
63
917081
73
99
92
818892
100
100
73
99
8863
5193 99
52
100
97
10086
10051
97
93
94100
98 98
94
8866
90
68
94
100100
Ranidae cont.
O
Fig. 2 (continued)
562 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
Ischnocnema hoehneiIschnocnema guentheriIschnocnema parva
Ischnocnema holtiIschnocnema juipocaBrachycephalus ephippiumCeuthomantis smaragdinus
Crinia tinnulaCrinia parinsignifera
Crinia deserticolaCrinia signiferaCrinia ripariaCrinia nimbusAssa darlingtoni
Geocrinia victorianaParacrinia haswelliMetacrinia nichollsiMyobatrachus gouldii
Pseudophryne bibroniiPseudophryne coriaceaUperoleia laevigataUperoleia littlejohni
Spicospina flammocaeruleaTaudactylus acutirostris
Mixophyes fasciolatusMixophyes schevilliMixophyes coggeri
Mixophyes carbinensisMixophyes balbus
Rheobatrachus silusLimnodynastes dumeriliiLimnodynastes interiorisLimnodynastes dorsalis
Limnodynastes terraereginaeLimnodynastes fletcheri
Limnodynastes depressusLimnodynastes tasmaniensisLimnodynastes peronii
Limnodynastes convexiusculusLimnodynastes lignarius
Limnodynastes salminiPhiloria sphagnicolus
Adelotus brevisHeleioporus australiacusLechriodus fletcheriPlatyplectrum ornatum
Platyplectrum spenceriNeobatrachus sudelliNeobatrachus pictusNeobatrachus pelobatoidesNotaden bennettii
Notaden melanoscaphusTelmatobufo bullockiTelmatobufo venustusCalyptocephallela gayi
57
97 97
10093 66
100
98
74
71
99
100
81
100
100
67
89
100
100
100
100
100
100
76
100
100
6864
100
93
1006597
78
54
10065
10098
100
99
100
100
94
98
Calyptocephallelidae
Ceuthomantidae
Brachycephalidae
Limnodynastinae
Myobatrachinae
Myobatrachidae
A-O
Q-ST-AEP
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 563

Author's personal copy
Eleutherodactylus pantoniEleutherodactylus pentasyringos
Eleutherodactylus cundalliEleutherodactylus glaucoreius
Eleutherodactylus griphusEleutherodactylus gossei
Eleutherodactylus junoriEleutherodactylus fuscus
Eleutherodactylus alticolaEleutherodactylus orcutti
Eleutherodactylus andrewsiEleutherodactylus nubicolaEleutherodactylus jamaicensis
Eleutherodactylus cavernicolaEleutherodactylus grabhami
Eleutherodactylus sisyphodemusEleutherodactylus luteolusEleutherodactylus toa
Eleutherodactylus rivularisEleutherodactylus riparius
Eleutherodactylus cuneatusEleutherodactylus turquinensis
Eleutherodactylus weinlandiEleutherodactylus rhodesiEleutherodactylus lentus
Eleutherodactylus grahamiEleutherodactylus pictissimusEleutherodactylus monensis
Eleutherodactylus probolaeusEleutherodactylus acmonis
Eleutherodactylus ricordiiEleutherodactylus bresslerae
Eleutherodactylus darlingtoniEleutherodactylus leoncei
Eleutherodactylus armstrongiEleutherodactylus alcoae
Eleutherodactylus limbatusEleutherodactylus jaumeiEleutherodactylus iberia
Eleutherodactylus etheridgeiEleutherodactylus orientalis
Eleutherodactylus cubanusEleutherodactylus varleyiEleutherodactylus intermedius
Eleutherodactylus gundlachiEleutherodactylus atkinsi
Eleutherodactylus thomasiEleutherodactylus blairhedgesi
Eleutherodactylus pinarensisEleutherodactylus pezopetrus
Eleutherodactylus rogersiEleutherodactylus tonyi
Eleutherodactylus goiniEleutherodactylus casparii
Eleutherodactylus planirostrisEleutherodactylus guanahacabibes
Eleutherodactylus greyiEleutherodactylus emiliaeEleutherodactylus dimidiatusEleutherodactylus maestrensis
Eleutherodactylus albipesEleutherodactylus schmidti
Eleutherodactylus caribeEleutherodactylus eunaster
Eleutherodactylus amadeusEleutherodactylus corona
Eleutherodactylus heminotaEleutherodactylus glaphycompus
Eleutherodactylus dolomedesEleutherodactylus bakeri
Eleutherodactylus thorectesEleutherodactylus glanduliferoides
Eleutherodactylus jugansEleutherodactylus oxyrhyncus
Eleutherodactylus apostatesEleutherodactylus rufifemoralis
Eleutherodactylus furcyensisEleutherodactylus paulsoni
Eleutherodactylus glanduliferEleutherodactylus sciagraphus
Eleutherodactylus brevirostrisEleutherodactylus ventrilineatus
Eleutherodactylus zugiEleutherodactylus klinikowskii
Eleutherodactylus pipilansEleutherodactylus nitidus
Eleutherodactylus marnockiiEleutherodactylus symingtoni
Eleutherodactylus zeusEleutherodactylus schwartziEleutherodactylus coquiEleutherodactylus portoricensis
Eleutherodactylus wightmanaeEleutherodactylus gryllusEleutherodactylus cochranaeEleutherodactylus hedrickiEleutherodactylus brittoni
Eleutherodactylus antillensisEleutherodactylus locustus
Eleutherodactylus eneidaeEleutherodactylus cooki
Eleutherodactylus flavescensEleutherodactylus martinicensis
Eleutherodactylus amplinymphaEleutherodactylus johnstoneiEleutherodactylus barlagnei
Eleutherodactylus pinchoniEleutherodactylus principalis
Eleutherodactylus auriculatusEleutherodactylus bartonsmithi
Eleutherodactylus mariposaEleutherodactylus ronaldi
Eleutherodactylus glamyrusEleutherodactylus eileenae
Eleutherodactylus audantiEleutherodactylus haitianus
Eleutherodactylus abbottiEleutherodactylus parabates
Eleutherodactylus pituinusEleutherodactylus minutus
Eleutherodactylus pooleiEleutherodactylus guantanamera
Eleutherodactylus ionthusEleutherodactylus varians
Eleutherodactylus leberiEleutherodactylus melacara
Eleutherodactylus lamprotesEleutherodactylus fowleri
Eleutherodactylus wetmoreiEleutherodactylus sommeri
Eleutherodactylus auriculatoidesEleutherodactylus patriciae
Eleutherodactylus unicolorEleutherodactylus richmondi
Eleutherodactylus hypostenorEleutherodactylus parapelatesEleutherodactylus ruthae
Eleutherodactylus bothroboansEleutherodactylus chlorophenaxEleutherodactylus nortoni
Eleutherodactylus inoptatusEleutherodactylus counouspeus
Diasporus diastemaPhyzelaphryne miriamae
Adelophryne gutturosa
89
90
85
100
97
65
95
100
64
8692
60
72
90
100 1007774
83
100
98
100
97
61
718010098
60 100
70
50
98 100
100 100
100
10099
86
1006455
7159
70
6251
100
97100
98100
90 100
99
100 100
100
79
59
90
55
81
100
92
6684
92 8964
93
97 89
10093
10097
100
100
98
83 93
8199
100
64
54
51
90
8299
100100
100
60
100100
92100
52
93 10082
100
100100
100
Eleutherodactylinae
Phyzelaphryninae
Eleutherodactylidae
Q
Fig. 2 (continued)
564 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
Oreobates madidiOreobates heterodactylus
Oreobates cruralisOreobates ibischi
Oreobates discoidalisOreobates sanderiOreobates granulosus
Oreobates sanctaecrucisOreobates choristolemma
Oreobates saxatilisOreobates quixensis
Oreobates lehriLynchius nebulanastes
Lynchius parkeriLynchius flavomaculatus
Phrynopus juninensisPhrynopus kauneorum
Phrynopus tautzorumPhrynopus barthlenaePhrynopus horstpauli
Phrynopus pesantesiPhrynopus bufoides
Phrynopus brackiBarycholos pulcher
Barycholos ternetziNoblella lochites
Holoaden luederwaldtiHoloaden bradeiBryophryne cophites
Psychrophrynella iatamasiPsychrophrynella wettsteini
Noblella peruvianaCraugastor talamancae
Craugastor crassidigitusCraugastor fitzingeri
Craugastor raniformisCraugastor tabasarae
Craugastor longirostrisCraugastor cuaquero
Craugastor andiCraugastor melanostictus
Craugastor emcelaeCraugastor ranoidesCraugastor rugulosusCraugastor fleischmanni
Craugastor rupiniusCraugastor megacephalus
Craugastor angelicusCraugastor punctariolusCraugastor obesus
Craugastor sandersoniCraugastor rhodopis
Craugastor lokiCraugastor mexicanus
Craugastor bransfordiiCraugastor podiciferusCraugastor lineatus
Craugastor laticepsCraugastor sartori
Craugastor pygmaeusCraugastor montanus
Craugastor spatulatusCraugastor bocourti
Craugastor stuartiCraugastor uno
Craugastor alfrediCraugastor tarahumaraensisCraugastor augusti
Craugastor daryiHaddadus binotatus
Strabomantis anomalusStrabomantis bufoniformisStrabomantis necerus
Strabomantis biporcatusStrabomantis sulcatus
Hypodactylus brunneusHypodactylus elassodiscus
Hypodactylus peraccaiHypodactylus dolops
86
85
72
100
76
87
65
100
100100
98
100
100
100
100
5859
95
7688
93
77
100 100
100
100100
77
89
81
85
52
98
100
7698 80
90
77
89
77100
98
85
100
100
69
99
10069
99100
8497
100
10067
100
100100
98
100
61 100
Strabomantinae
Craugastorinae
Holoadeninae
81
Pristimantinae
Pristimantinae cont.
Craugastoridae
R
incertae sedis
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 565

Author's personal copy
Pristimantis llojsintutaPristimantis croceoinguinisPristimantis imitatrix
Pristimantis lirellusPristimantis marmoratus
Pristimantis inguinalisPristimantis pulvinatus
Pristimantis altamazonicusPristimantis platydactylus
Pristimantis diadematusPristimantis ockendeni
Pristimantis unistrigatusPristimantis ardalonychus
Pristimantis cajamarcensisPristimantis ceuthospilus
Pristimantis walkeriPristimantis luteolateralis
Pristimantis parvillusPristimantis chalceus
Pristimantis simonbolivariPristimantis orestes
Pristimantis rivetiPristimantis versicolor
Pristimantis phoxocephalusPristimantis spinosus
Pristimantis cryophiliusPristimantis wiensiPristimantis petrobardus
Pristimantis quaquaversusPristimantis rhabdocnemus
Pristimantis melanogasterPristimantis simonsiiPristimantis rhodoplichusPristimantis dissimulatus
Pristimantis calcarulatusPristimantis appendiculatus
Pristimantis pycnodermisPristimantis orcesi
Pristimantis glandulosusPristimantis inusitatusPristimantis acerus
Pristimantis acuminatusPristimantis schultei
Pristimantis bromeliaceusPristimantis zeuctotylus
Pristimantis galdiPristimantis subsigillatusPristimantis nyctophylax
Pristimantis cruciferPristimantis truebaePristimantis gentryi
Pristimantis curtipesPristimantis buckleyi
Pristimantis vertebralisPristimantis devillei
Pristimantis surdusPristimantis quinquagesimus
Pristimantis duellmaniPristimantis thymalopsoides
Pristimantis ocreatusPristimantis thymelensis
Pristimantis pyrrhomerusPristimantis leoni
Pristimantis celatorPristimantis verecundusPristimantis supernatis
Pristimantis chloronotusPristimantis eriphus
Pristimantis rozeiPristimantis urichi
Pristimantis prolatusPristimantis buccinator
Pristimantis citriogasterPristimantis malkini
Pristimantis condorPristimantis conspicillatus
Pristimantis achatinusPristimantis lymani
Pristimantis bipunctatusPristimantis skydmainos
Pristimantis fenestratusPristimantis koehleri
Pristimantis chiastonotusPristimantis samaipatae
Pristimantis terraebolivarisPristimantis toftae
Pristimantis rhabdolaemusPristimantis pluvicanorus
Pristimantis danaePristimantis sagittulus
Pristimantis stictogasterPristimantis aniptopalmatus
Pristimantis reichleiPristimantis peruvianus
Pristimantis euphronidesPristimantis shrevei
Pristimantis capriferPristimantis latidiscus
Pristimantis colomaiPristimantis cruentus
Pristimantis cremnobatesPristimantis caryophyllaceus
Pristimantis ridensPristimantis actitesPristimantis w-nigrum
Pristimantis lanthanitesPristimantis labiosus
Pristimantis crenunguisPristimantis bisignatus
Pristimantis fraudatorPristimantis mercedesae
Pristimantis ashkapara
75
98
75
90
99
78
98
100
100
98
77100
96
87
9598
100
57
68
79
93
100 100
100
100
81100
86
63
9981
71
88
86
94
8695
8455 88
100
82
56
8576
58
84100
60
100
1009965
100100
88
94
90
100
100 85100
100
100100
64
99
66
99100
9990
70
9999
9950 80
99
8883
8994
83
100
100
100
100
9367
62 100
93
100 100
100
10092
Pristimantinae cont.
S
Fig. 2 (continued)
566 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583
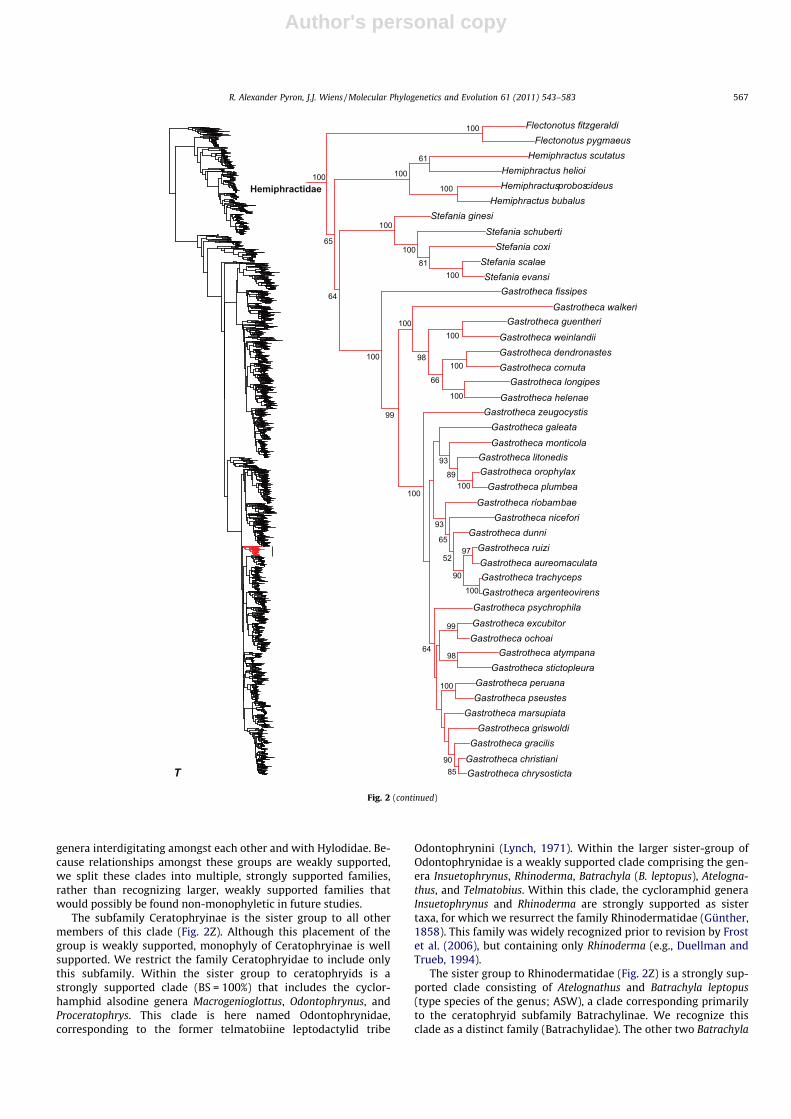
Author's personal copy
genera interdigitating amongst each other and with Hylodidae. Be-cause relationships amongst these groups are weakly supported,we split these clades into multiple, strongly supported families,rather than recognizing larger, weakly supported families thatwould possibly be found non-monophyletic in future studies.
The subfamily Ceratophryinae is the sister group to all othermembers of this clade (Fig. 2Z). Although this placement of thegroup is weakly supported, monophyly of Ceratophryinae is wellsupported. We restrict the family Ceratophryidae to include onlythis subfamily. Within the sister group to ceratophryids is astrongly supported clade (BS = 100%) that includes the cyclor-hamphid alsodine genera Macrogenioglottus, Odontophrynus, andProceratophrys. This clade is here named Odontophrynidae,corresponding to the former telmatobiine leptodactylid tribe
Odontophrynini (Lynch, 1971). Within the larger sister-group ofOdontophrynidae is a weakly supported clade comprising the gen-era Insuetophrynus, Rhinoderma, Batrachyla (B. leptopus), Atelogna-thus, and Telmatobius. Within this clade, the cycloramphid generaInsuetophrynus and Rhinoderma are strongly supported as sistertaxa, for which we resurrect the family Rhinodermatidae (Günther,1858). This family was widely recognized prior to revision by Frostet al. (2006), but containing only Rhinoderma (e.g., Duellman andTrueb, 1994).
The sister group to Rhinodermatidae (Fig. 2Z) is a strongly sup-ported clade consisting of Atelognathus and Batrachyla leptopus(type species of the genus; ASW), a clade corresponding primarilyto the ceratophryid subfamily Batrachylinae. We recognize thisclade as a distinct family (Batrachylidae). The other two Batrachyla
Gastrotheca chrysostictaGastrotheca christianiGastrotheca gracilis
Gastrotheca griswoldiGastrotheca marsupiata
Gastrotheca pseustesGastrotheca peruana
Gastrotheca stictopleuraGastrotheca atympana
Gastrotheca ochoaiGastrotheca excubitorGastrotheca psychrophila
Gastrotheca argenteovirensGastrotheca trachycepsGastrotheca aureomaculataGastrotheca ruizi
Gastrotheca dunniGastrotheca nicefori
Gastrotheca riobambaeGastrotheca plumbea
Gastrotheca orophylaxGastrotheca litonedis
Gastrotheca monticolaGastrotheca galeata
Gastrotheca zeugocystisGastrotheca helenae
Gastrotheca longipesGastrotheca cornutaGastrotheca dendronastesGastrotheca weinlandii
Gastrotheca guentheriGastrotheca walkeri
Gastrotheca fissipesStefania evansi
Stefania scalaeStefania coxi
Stefania schubertiStefania ginesi
Hemiphractus bubalusHemiphractus proboscideusHemiphractus helioi
Hemiphractus scutatusFlectonotus pygmaeus
Flectonotus fitzgeraldi
100
65
64
100
99
100
64
9085
100
98
99
93
65
52
90
100
97
93
89100
100
98
66
100
100
100
100
10081
100
100
100
61
100
Hemiphractidae
T
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 567
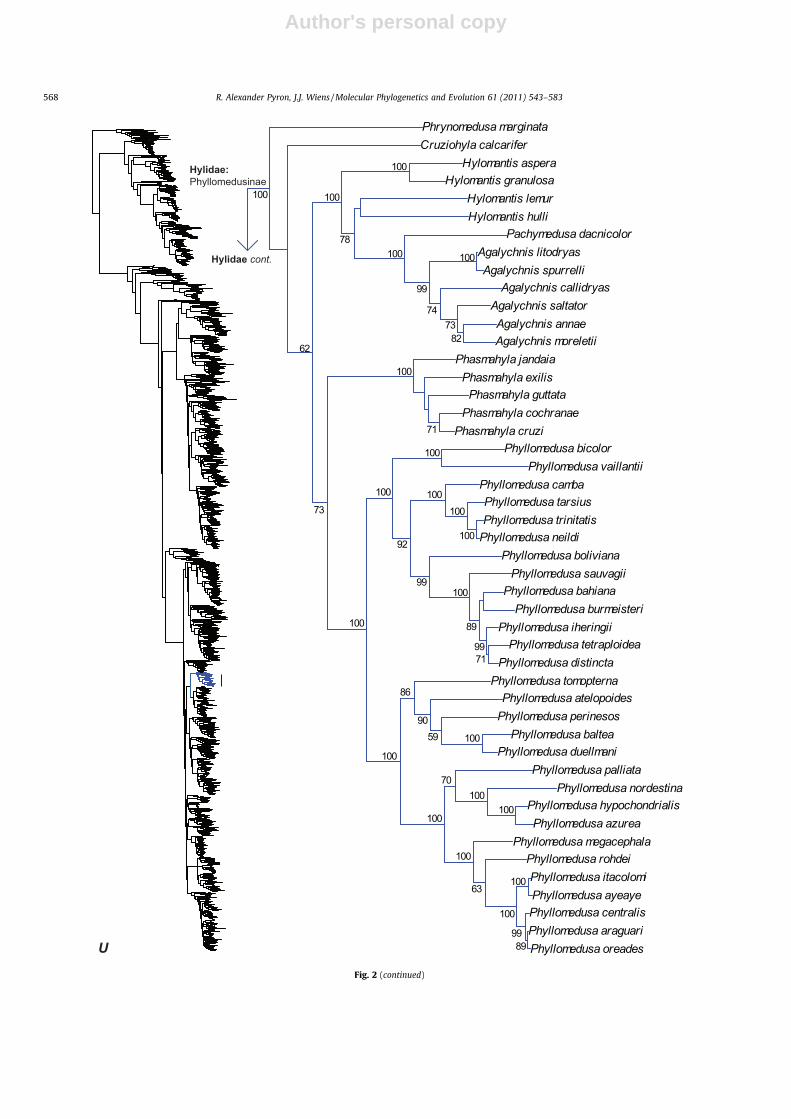
Author's personal copy
Phyllomedusa oreadesPhyllomedusa araguariPhyllomedusa centralisPhyllomedusa ayeayePhyllomedusa itacolomi
Phyllomedusa rohdeiPhyllomedusa megacephala
Phyllomedusa azureaPhyllomedusa hypochondrialis
Phyllomedusa nordestinaPhyllomedusa palliata
Phyllomedusa duellmaniPhyllomedusa baltea
Phyllomedusa perinesosPhyllomedusa atelopoides
Phyllomedusa tomopternaPhyllomedusa distincta
Phyllomedusa tetraploideaPhyllomedusa iheringii
Phyllomedusa burmeisteriPhyllomedusa bahiana
Phyllomedusa sauvagiiPhyllomedusa boliviana
Phyllomedusa neildiPhyllomedusa trinitatisPhyllomedusa tarsius
Phyllomedusa cambaPhyllomedusa vaillantii
Phyllomedusa bicolorPhasmahyla cruzi
Phasmahyla cochranaePhasmahyla guttata
Phasmahyla exilisPhasmahyla jandaia
Agalychnis moreletiiAgalychnis annae
Agalychnis saltatorAgalychnis callidryas
Agalychnis spurrelliAgalychnis litodryas
Pachymedusa dacnicolorHylomantis hulliHylomantis lemur
Hylomantis granulosaHylomantis aspera
Cruziohyla calcariferPhrynomedusa marginata
100
62
73
100
100
100
100
63
100
9989
100
70100
100
86
9059 100
100
92
99100
89
9971
100100
100
100
100
71
100
78100
99
7473
82
100
100Hylidae:Phyllomedusinae
U
Hylidae cont.
Fig. 2 (continued)
568 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
Litoria mainiLitoria maculosa
Litoria longipesLitoria vagitusLitoria cultripes
Litoria manyaLitoria cryptotis
Litoria alboguttataLitoria brevipes
Litoria verrucosaLitoria platycephala
Litoria novaehollandiaeLitoria australis
Litoria dahliiLitoria mooreiLitoria cyclorhyncha
Litoria aureaLitoria raniformis
Litoria xanthomeraLitoria chlorisLitoria gracilenta
Litoria kumaeLitoria gilleniLitoria caerulea
Litoria splendidaLitoria cavernicolaLitoria wilcoxii
Litoria jungguyLitoria booroolongensis
Litoria lesueuriiLitoria andiirrmalin
Litoria genimaculataLitoria eucnemis
Litoria exophthalmiaLitoria pearsonianaLitoria barringtonensis
Litoria nudidigitaLitoria phyllochroa
Litoria citropaLitoria daviesaeLitoria subglandulosa
Litoria spenceriLitoria rheocola
Litoria nyakalensisLitoria nannotis
Litoria dayiLitoria impura
Litoria thesaurensisLitoria pulcher
Litoria papuaLitoria cheesmani
Litoria foriculaLitoria semipalmatus
Litoria humeralisLitoria zweifeli
Litoria narinosusLitoria kubori
Litoria brevipalmataLitoria infrafrenata
Litoria duxLitoria paraewingi
Litoria verreauxiiLitoria revelata
Litoria littlejohniLitoria ewingii
Litoria jervisiensisLitoria electrica
Litoria rubellaLitoria dentata
Litoria congenitaLitoria amboinensisLitoria darlingtoni
Litoria peroniiLitoria tyleri
Litoria rothiiLitoria adelaidensis
Litoria burrowsiLitoria inermis
Litoria pallidaLitoria tornieri
Litoria freycinetiLitoria latopalmata
Litoria coplandiLitoria watjulumensis
Litoria nasutaLitoria nigrofrenata
Litoria personataLitoria microbelos
Litoria dorsalisLitoria longirostris
Litoria meirianaLitoria arfakiana
Litoria wollastoniLitoria spartacus
Litoria angianaLitoria micromembrana
Litoria modicaLitoria leucova
Litoria proraLitoria nigropunctata
Litoria majikthiseLitoria iris
Litoria havinaLitoria multiplica
Litoria pronimiaLitoria bicolorLitoria fallaxLitoria olongburensis
100
76
95
79
9274
9474
7953
100
100
86
80 100
96100
100
98
10075
87
5482
55
100100
100
97
54
91100 93
100
100
89100
9556
9994
100
100
100
98
93
53
58
10081
9155
56
100
72
58
10092
99
72
10093
93
100
100
69 86
94
76
78
87
67 98
79
91
65
83 100
100
Hylidae:Pelodryadinae
Hylidae cont.
V
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 569

Author's personal copy
species included in our analysis are placed in a clade with Alsodes,Eupsophus, and Hylorina (see below). The sister group to the cladeof Rhinodermatidae + Batrachylidae is the ceratophryid genus Tel-matobius (Fig. 2Z). We resurrect the family Telmatobiidae (Miran-da-Ribeiro, 1920) for this strongly supported clade (containingonly Telmatobius, which likely contains Batrachophrynus; ASW),
which was previously recognized as a ceratophryid subfamily(Telmatobiinae) by ASW and AW. Next up the tree is a moderatelywell supported clade (BS = 79%) consisting of two cycloramphidgenera, the cycloramphine genus Cycloramphus and the alsodinegenus Thoropa. We recognize this clade as the family Cycloramphi-dae (a greatly restricted version relative to current classifications).
Hypsiboas pulchellusHypsiboas cordobaeHypsiboas prasinus
Hypsiboas cainguaHypsiboas bischoffiHypsiboas marginatusHypsiboas guentheri
Hypsiboas andinusHypsiboas riojanusHypsiboas marianitae
Hypsiboas balzaniHypsiboas latistriatusHypsiboas polytaeniusHypsiboas leptolineatus
Hypsiboas joaquiniHypsiboas semiguttatusHypsiboas ericae
Hypsiboas lundiiHypsiboas pardalis
Hypsiboas faberHypsiboas crepitans
Hypsiboas rosenbergiHypsiboas albomarginatusHypsiboas rufitelusHypsiboas pellucens
Hypsiboas albopunctatusHypsiboas multifasciatusHypsiboas lanciformis
Hypsiboas calcaratusHypsiboas dentei
Hypsiboas fasciatusHypsiboas raniceps
Hypsiboas heilpriniHypsiboas picturatus
Hypsiboas lemaiHypsiboas benitezi
Hypsiboas ornatissimusHypsiboas nympha
Hypsiboas microdermaHypsiboas roraima
Hypsiboas siblesziHypsiboas geographicus
Hypsiboas semilineatusHypsiboas boans
Hypsiboas cinerascensAplastodiscus leucopygius
Aplastodiscus cavicolaAplastodiscus callipygius
Aplastodiscus albosignatusAplastodiscus cochranae
Aplastodiscus perviridisAplastodiscus weygoldtiAplastodiscus arildaeAplastodiscus eugenioi
Aplastodiscus albofrenatusBokermannohyla circumdataBokermannohyla hylax
Bokermannohyla astarteaBokermannohyla martinsi
Hyloscirtus pachaHyloscirtus pantostictusHyloscirtus lindae
Hyloscirtus tapichalacaHyloscirtus charazaniHyloscirtus armatus
Hyloscirtus lasciniusHyloscirtus palmeri
Hyloscirtus phyllognathusHyloscirtus colymba
Hyloscirtus simmonsiMyersiohyla kanaima
Myersiohyla inparquesi
98
63
97
97
76
51
55
66
94
100
100
100
87
70100
100
7999
100
100
100
91 100
100
94
6590
100
99
100
84
100
57 83100
72
93
99100
91100
100100
82
909050
100
99
100
100
80
8192
100
94
79 100100
100
100
65
71Hylidae:Hylinae
Hylinae cont.
W
Fig. 2 (continued)
570 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
Osteocephalus oophagusOsteocephalus taurinus
Osteocephalus leprieuriiOsteocephalus planiceps
Osteocephalus alboguttatusOsteocephalus verrucigerOsteocephalus buckleyi
Osteocephalus cabreraiOsteocephalus mutabor
Tepuihyla edelcaeOsteopilus wilderi
Osteopilus marianaeOsteopilus crucialis
Osteopilus brunneusOsteopilus pulchrilineatus
Osteopilus dominicensisOsteopilus septentrionalis
Osteopilus vastusPhyllodytes luteolus
Trachycephalus hadrocepsTrachycephalus venulosusTrachycephalus resinifictrixTrachycephalus nigromaculatus
Trachycephalus imitatrixTrachycephalus mesophaeusTrachycephalus coriaceus
Trachycephalus jordaniArgenteohyla siemersi
Nyctimantis rugicepsAparasphenodon brunoi
Corythomantis greeningiPhyllodytes auratus
Itapotihyla langsdorffiiDendropsophus rubicundulus
Dendropsophus sanborniDendropsophus minusculus
Dendropsophus branneriDendropsophus berthalutzae
Dendropsophus bipunctatusDendropsophus nanus
Dendropsophus walfordiDendropsophus riveroi
Dendropsophus sartoriDendropsophus robertmertensi
Dendropsophus microcephalusDendropsophus rhodopeplus
Dendropsophus lealiDendropsophus minutus
Dendropsophus bifurcusDendropsophus sarayacuensis
Dendropsophus leucophyllatusDendropsophus triangulum
Dendropsophus ebraccatusDendropsophus elegans
Dendropsophus miyataiDendropsophus schubarti
Dendropsophus aperomeusDendropsophus anceps
Dendropsophus pelidnaDendropsophus labialis
Dendropsophus carnifexDendropsophus giesleri
Dendropsophus brevifronsDendropsophus parviceps
Dendropsophus koechliniDendropsophus seniculus
Dendropsophus marmoratusDendropsophus allenorum
Xenohyla truncataPseudis paradoxaPseudis bolbodactyla
Pseudis tocantinsPseudis fusca
Pseudis minutaPseudis cardosoi
Pseudis carayaPseudis limellum
Pseudis laevisScarthyla goinorum
Scinax ruberScinax x-signatus
Scinax nasicusScinax fuscovarius
Scinax cruentommusScinax boesemani
Scinax staufferiScinax elaeochroa
Scinax squalirostrisScinax crospedospilus
Scinax garbeiScinax jolyi
Scinax proboscideusScinax nebulosus
Scinax rostratusScinax sugillatus
Scinax boulengeriScinax acuminatus
Scinax uruguayusScinax catharinae
Scinax berthaeSphaenorhynchus dorisae
Sphaenorhynchus lacteusSphaenorhynchus orophilus
56
100
65
69
90
100
8587
100
97
95
99
100
5753
86 90
94
66
1008451
10076
81100
91
53
54
97
89
55
66
7393
94
100
85
9081 100
8897
8283
100
59
100
100
100
75
100
99
99
87 94
80
99
100
76
100
97
95
97
95
96
8796 100
100
80
5188
100
9660
65
52
100
100
10097
Hylinae cont.
Hylinae cont.X
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 571

Author's personal copy
The sister group to Cycloramphidae consists of Hylodidae(strongly supported as monophyletic), and a clade consisting ofsome of the former alsodine cycloramphid genera (Alsodes, Eupso-phus, Hylorina, Limnomedusa) and two species of the ceratophryidgenus Batrachyla. Support for the monophyly of this group isrelatively weak (54%) including Limnomedusa but is strong (91%)
without it. We tentatively recognize this clade as a distinct family(Alsodidae), comprising Alsodes, Eupsophus, Hylorina, and Limnome-dusa. This is the only family that we recognize (that was not in-cluded in previous classifications) that is not strongly supported,but we prefer this taxonomy as opposed to recognizing a mono-typic family containing only Limnomedusa. While we generally
Hyla plicataHyla euphorbiacea
Hyla eximiaHyla walkeri
Hyla wrightorumHyla arenicolor
Hyla immaculataHyla japonicaHyla avivocaHyla chrysoscelis
Hyla versicolorHyla femoralis
Hyla andersoniiHyla cinerea
Hyla gratiosaHyla squirella
Hyla molleriHyla intermedia
Hyla orientalisHyla arborea
Hyla savignyiHyla sarda
Hyla meridionalisHyla annectans
Hyla tsinlingensisHyla chinensis
Smilisca phaeotaSmilisca puma
Smilisca cyanostictaSmilisca fodiens
Smilisca sordidaSmilisca sila
Smilisca baudiniiAnotheca spinosa
Triprion petasatusTriprion spatulatusIsthmohyla rivularis
Isthmohyla ticaIsthmohyla zeteki
Isthmohyla pseudopumaTlalocohyla loquaxTlalocohyla godmani
Tlalocohyla pictaTlalocohyla smithii
Charadrahyla nephilaCharadrahyla taeniopus
Megastomatohyla mixePtychohyla zophodesPtychohyla leonhardschultzeiPtychohyla euthysanota
Ptychohyla hypomykterPtychohyla dendrophasma
Duellmanohyla rufioculisDuellmanohyla soralia
Bromeliohyla bromeliaciaPtychohyla spinipollex
Ecnomiohyla miotympanumEcnomiohyla miliaria
Ecnomiohyla mineraPlectrohyla calthulaPlectrohyla pentheterPlectrohyla bistincta
Plectrohyla ameibothalamePlectrohyla cycladaPlectrohyla arborescandens
Plectrohyla siopelaPlectrohyla matudai
Plectrohyla guatemalensisPlectrohyla chrysopleura
Plectrohyla glandulosaExerodonta xera
Exerodonta smaragdinaExerodonta chimalapa
Exerodonta melanommaExerodonta sumichrasti
Exerodonta abdivitaExerodonta perkinsi
Pseudacris nigritaPseudacris kalmi
Pseudacris triseriataPseudacris feriarum
Pseudacris maculataPseudacris clarkiiPseudacris fouquetteiPseudacris brimleyi
Pseudacris brachyphonaPseudacris streckeriPseudacris illinoensis
Pseudacris ornataPseudacris crucifer
Pseudacris ocularisPseudacris regilla
Pseudacris cadaverinaAcris crepitans
Acris blanchardiAcris gryllus
100
92
94
86
100
77
100
98
8095 95
968373
8161100
100
100 99
100
92100
9252
100100
99
94
100100
9995
97 96
98
87
84
83100
100
100
100
99
95 100
100
100
99100
89100
8889
100
100
96
5674
83 84
80
92
68
100
76 10064
100
100
98
97
100
71
10067
52 100100
100
100
100100
100
100
100 90Hylidae cont.
Y
Fig. 2 (continued)
572 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
refrain from addressing generic-level taxonomy here, the genusBatrachyla represents a special case, being split between two fam-ilies. As Batrachyla taeniata, Batrachyla antartandica, and Hylorinasylvatica are strongly placed within Eupsophus, we synonymizethose three species with Eupsophus. We keep the other two
Batrachyla species (Batrachyla fitzroya and Batrachyla nibaldoi) inBatrachyla with the type species B. leptopus in Batrachylidae.
In summary, our new taxonomy replaces the non-monophyleticinterdigitating families Ceratophryidae and Cycloramphidae with asomewhat larger set of families that are each generally strongly
Eupsophus contulmoensisEupsophus roseus
Eupsophus insularisEupsophus nahuelbutensisEupsophus migueli
Batrachyla antartandicaBatrachyla taeniata
Hylorina sylvaticaEupsophus vertebralis
Eupsophus calcaratusEupsophus emiliopuginiAlsodes barrioi
Alsodes monticolaAlsodes kaweshkariAlsodes gargolaAlsodes tumultuosusAlsodes australis
Alsodes nodosusAlsodes vanzolinii
Limnomedusa macroglossaHylodes ornatus
Hylodes sazimaiHylodes phyllodes
Hylodes perplicatusHylodes meridionalis
Megaelosia goeldiiHylodes dactylocinus
Crossodactylus schmidtiCrossodactylus caramaschii
Thoropa taophoraThoropa miliaris
Cycloramphus acangatanCycloramphus boraceiensis
Telmatobius huayraTelmatobius hintoniTelmatobius gigasTelmatobius culeusTelmatobius bolivianus
Telmatobius yuracareTelmatobius sibiricusTelmatobius simonsi
Telmatobius marmoratusTelmatobius vilamensis
Telmatobius sanborniTelmatobius verrucosusTelmatobius espadai
Telmatobius truebaeTelmatobius vellardi
Telmatobius nigerTelmatobius zapahuirensis
Atelognathus patagonicusAtelognathus jeinimenensis
Batrachyla leptopusRhinoderma darwinii
Insuetophrynus acarpicusProceratophrys renalis
Proceratophrys boieiProceratophrys laticeps
Proceratophrys cururuOdontophrynus moratoi
Proceratophrys concavitympanumProceratophrys goyana
Proceratophrys avelinoiProceratophrys bigibbosa
Proceratophrys appendiculataProceratophrys melanopogon
Proceratophrys schirchiProceratophrys cristiceps
Odontophrynus achalensisOdontophrynus occidentalis
Odontophrynus americanusOdontophrynus cultripes
Odontophrynus carvalhoiMacrogenioglottus alipioi
Ceratophrys ornataCeratophrys cranwelli
Lepidobatrachus laevisChacophrys pierottii
Ceratophrys cornuta
53
54
91
95
9599
50
97
90
61
56
6175
68
89
97
9254
92
79
100
100
100
57
88
9396
68
50
73 8571
76
96 100
96
100
100
67
77 99
59
99
5057
85
100
66
99
61
100
65100
82Ceratophryidae
Odontophrynidae
Rhinodermatidae
Batrachylidae
Telmatobiidae
Cycloramphidae
Hylodidae
Alsodidae
Z
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 573

Author's personal copy
Leptodactylus plaumanniLeptodactylus gracilis
Leptodactylus albilabrisLeptodactylus longirostris
Leptodactylus bufoniusLeptodactylus fuscus
Leptodactylus mystaceusLeptodactylus didymus
Leptodactylus notoaktitesLeptodactylus elenae
Leptodactylus spixiLeptodactylus mystacinus
Leptodactylus pentadactylusLeptodactylus knudseniLeptodactylus labyrinthicus
Leptodactylus fallaxLeptodactylus vastus
Leptodactylus rhodonotusLeptodactylus griseigularis
Leptodactylus discodactylusLeptodactylus diedrus
Leptodactylus podicipinusLeptodactylus validusLeptodactylus pallidirostris
Leptodactylus wagneriLeptodactylus melanonotus
Leptodactylus riveroiLeptodactylus leptodactyloides
Leptodactylus ocellatusLeptodactylus chaquensis
Leptodactylus silvanimbusLeptodactylus rhodomystax
Adenomera heyeriAdenomera hylaedactyla
Adenomera andreaeLithodytes lineatus
Physalaemus barrioiPhysalaemus gracilisPhysalaemus riograndensis
Physalaemus biligonigerusPhysalaemus ephippifer
Physalaemus cuvieriPhysalaemus albonotatus
Physalaemus signiferEupemphix nattereri
Engystomops randiEngystomops montubio
Engystomops coloradorumEngystomops guayaco
Engystomops pustulatusEngystomops freibergi
Engystomops petersiEngystomops pustulosus
Edalorhina pereziPleurodema bibroniPleurodema thaul
Pleurodema bufoninumPleurodema marmoratum
Pleurodema brachyopsPseudopaludicola falcipes
Paratelmatobius gaigeaeParatelmatobius poecilogaster
Paratelmatobius cardosoiScythrophrys sawayae
79
93
100
56
84
6294
70
5666
88
10095
52
80
54
58
69100
92
93
9354
65
99
99
98
100
98
9996
80
100
100
100
99
100100
100
94
9899
91
9994
94
9393
100
Leptodactylidae
Paratelmatobiinae
Leiuperinae
Leptodactylinae
AA
Fig. 2 (continued)
574 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
supported. However, we acknowledge that the family-level place-ment of the cycloramphid genera Rupirana (no subfamily assignedby AW or ASW), Zachaenus, and Crossodactylodes (both previouslyCycloramphinae; AW; ASW) are uncertain in our classification(Hyloidea incertae sedis), given the lack of molecular data for these
genera and the apparent problems in the previous taxonomy(Figs. 1 and 2Z). Assuming that they are cycloramphids as the fam-ily is defined here (Figs. 1 and 2; Appendix B) may be incorrect.
We also find that the currently recognized families Leiuperidaeand Leptodactylidae are also problematic (Fig. 2AA). Our results
Centrolene altitudinaleCentrolene notostictum
Centrolene buckleyiCentrolene hesperium
Centrolene venezuelenseCentrolene pipilatum
Centrolene hybridaCentrolene bacatum
Centrolene geckoideumCentrolene antioquiense
Centrolene peristictumCentrolene savagei
Centrolene daidaleumNymphargus siren
Nymphargus megacheirusNymphargus rosadus
Nymphargus puyoensisNymphargus cochranae
Nymphargus wileyiNymphargus griffithsi
Nymphargus posadaeNymphargus bejaranoiNymphargus pluvialis
Nymphargus mixomaculatusCentrolene grandisonae
Rulyrana adiazetaRulyrana susatamai
Rulyrana spiculataRulyrana flavopunctata
Sachatamia albomaculataSachatamia punctulata
Sachatamia ilexEspadarana prosobleponEspadarana callistomma
Espadarana andinaChimerella mariaelenaeCochranella euknemos
Cochranella macheCochranella granulosa
Cochranella nolaCochranella litoralis
Teratohyla spinosaTeratohyla midas
Teratohyla pulverataVitreorana helenae
Vitreorana oyampiensisVitreorana gorzulae
Vitreorana castroviejoiVitreorana antisthenesiVitreorana eurygnatha
Ikakogi tayronaHyalinobatrachium ibama
Hyalinobatrachium pallidumHyalinobatrachium duranti
Hyalinobatrachium orocostaleHyalinobatrachium fragile
Hyalinobatrachium orientaleHyalinobatrachium fleischmanni
Hyalinobatrachium tatayoiHyalinobatrachium carlesvilai
Hyalinobatrachium mondolfiiHyalinobatrachium crurifasciatum
Hyalinobatrachium eccentricumHyalinobatrachium ignioculus
Hyalinobatrachium tayloriHyalinobatrachium iaspidienseHyalinobatrachium colymbiphyllum
Hyalinobatrachium chirripoiHyalinobatrachium talamancae
Hyalinobatrachium pellucidumHyalinobatrachium bergeri
Hyalinobatrachium aureoguttatumHyalinobatrachium valerioi
Celsiella revocataCelsiella vozmedianoi
Allophryne ruthveni
100
100
70
60
80
100
61
100
8389
6799
86
95
100
100
100
98
8189 85
76
5790
84 80
100
76
85
10097
88
100100
61100
96
100
96100
5099
95
66 100
5096
100
98
72
61
83
93100
99
58
95
88
8292
10088
85
10070
100
62100
100
Allophrynidae
Centrolenidae
Hyalinobatrachinae
Centroleninae
AB
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 575

Author's personal copy
Dendrobates ventrimaculatusDendrobates variabilis
Dendrobates amazonicusDendrobates uakarii
Dendrobates reticulatusDendrobates duellmani
Dendrobates fantasticusDendrobates vanzoliniiDendrobates imitator
Dendrobates flavovittatusDendrobates biolat
Dendrobates lamasiDendrobates claudiaeDendrobates minutus
Dendrobates fulguritusDendrobates bombetesDendrobates virolinensis
Dendrobates mysteriosusDendrobates captivus
Dendrobates pumilioDendrobates vicentei
Dendrobates speciosusDendrobates arboreusDendrobates lehmanniDendrobates sylvaticusDendrobates histrionicus
Dendrobates granuliferusDendrobates leucomelasDendrobates tinctorius
Dendrobates auratusDendrobates truncatus
Dendrobates galactonotusDendrobates castaneoticus
Dendrobates quinquevittatusDendrobates steyermarki
Phyllobates terribilisPhyllobates aurotaenia
Phyllobates bicolorPhyllobates lugubrisPhyllobates vittatus
Hyloxalus awaHyloxalus toachi
Hyloxalus infraguttatusHyloxalus elachyhistusHyloxalus insulatus
Hyloxalus idiomelusHyloxalus chlorocraspedusHyloxalus azureiventris
Hyloxalus nexipusHyloxalus sylvaticus
Hyloxalus pulcherrimusHyloxalus anthracinus
Hyloxalus delatorreaeHyloxalus pulchellus
Hyloxalus vertebralisHyloxalus sauliHyloxalus bocagei
Hyloxalus maculosusHyloxalus sordidatusHyloxalus leucophaeus
Hyloxalus subpunctatusAmeerega petersiAmeerega cainarachi
Ameerega smaragdinaAmeerega rubriventrisAmeerega maceroAmeerega pictaAmeerega hahneliAmeerega pulchripectaAmeerega trivittata
Ameerega pongoensisAmeerega parvulaAmeerega bilinguis
Ameerega bassleriAmeerega braccata
Ameerega flavopictaAmeerega silverstonei
Ameerega simulansAmeerega altamazonica
Colostethus argyrogasterColostethus fugax
Colostethus fraterdanieliColostethus panamansis
Colostethus inguinalisColostethus imbricolus
Colostethus prattiColostethus latinasus
Epipedobates tricolorEpipedobates anthonyiEpipedobates machalillaEpipedobates espinosai
Epipedobates boulengeriSilverstoneia flotator
Silverstoneia nubicolaAllobates conspicuus
Allobates trilineatusAllobates caeruleodactylus
Allobates gasconiAllobates juaniiAllobates femoralis
Allobates zaparoAllobates nidicola
Allobates brunneusAllobates talamancae
Allobates undulatusMannophryne venezuelensis
Mannophryne trinitatisMannophryne herminae
Aromobates nocturnusAromobates saltuensis
Anomaloglossus beebeiAnomaloglossus roraima
Anomaloglossus praderioiAnomaloglossus degranvillei
Anomaloglossus tepuyensisAnomaloglossus baeobatrachusAnomaloglossus stepheni
Rheobates palmatus
100
97
71
100
99
72
58
64
69
59
85
89
84 93
100
10081 100
10084
71
10094615795
95
98
10096
100
6272
70
100
10075
100
99
100
91100
6580
100 100
100100
100 100
80
100 99
57
100
100
98
83100
89
86
88
85
82
97
85100
100
10097
100
100100
598795
77
100
50
100
10099
9188
8583
100
99 100 100
100
100
8710064
100
98
Dendrobatidae
AC
Fig. 2 (continued)
576 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583
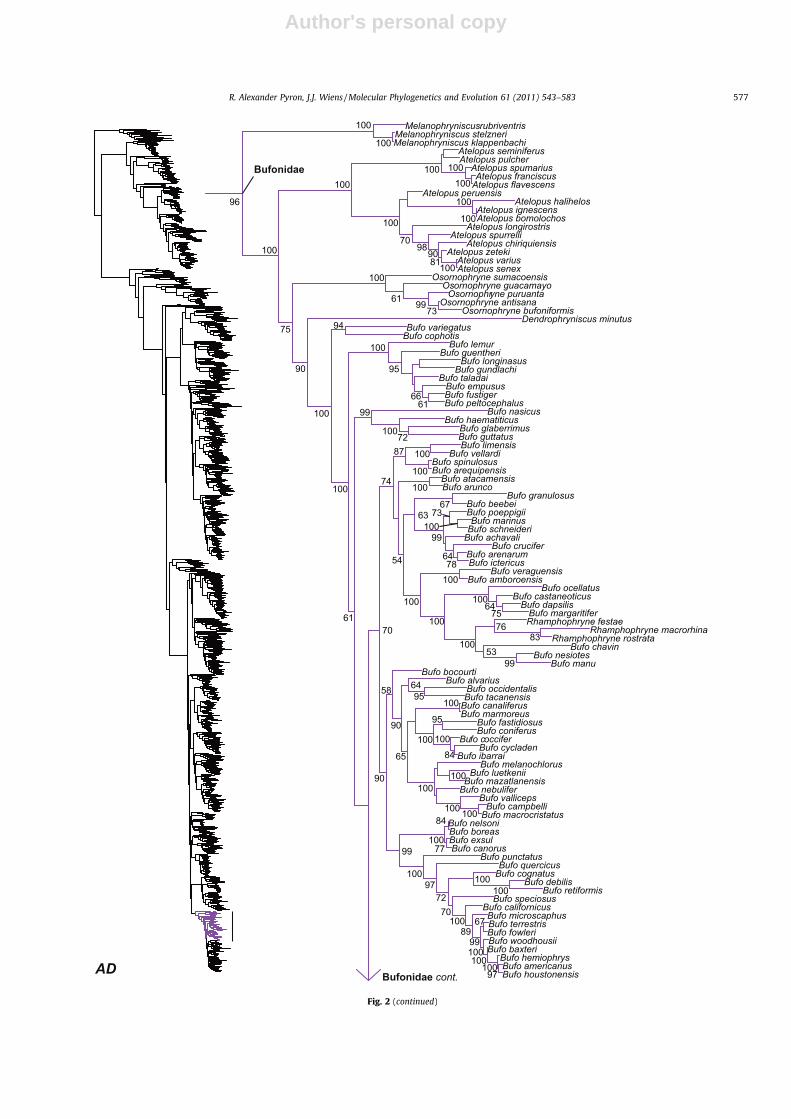
Author's personal copy
Bufo houstonensisBufo americanusBufo hemiophrys
Bufo baxteriBufo woodhousiiBufo fowleriBufo terrestrisBufo microscaphus
Bufo californicusBufo speciosus
Bufo retiformisBufo debilis
Bufo cognatusBufo quercicus
Bufo punctatusBufo canorusBufo exsulBufo boreasBufo nelsoni
Bufo macrocristatusBufo campbelli
Bufo vallicepsBufo nebuliferBufo mazatlanensis
Bufo luetkeniiBufo melanochlorus
Bufo ibarraiBufo cycladen
Bufo cocciferBufo coniferusBufo fastidiosus
Bufo marmoreusBufo canaliferusBufo tacanensisBufo occidentalis
Bufo alvariusBufo bocourti
Bufo manuBufo nesiotes
Bufo chavinRhamphophryne rostrata
Rhamphophryne macrorhinaRhamphophryne festaeBufo margaritifer
Bufo dapsilisBufo castaneoticus
Bufo ocellatusBufo amboroensis
Bufo veraguensisBufo ictericusBufo arenarum
Bufo cruciferBufo achavaliBufo schneideriBufo marinus
Bufo poeppigiiBufo beebei
Bufo granulosusBufo aruncoBufo atacamensis
Bufo arequipensisBufo spinulosus
Bufo vellardiBufo limensisBufo guttatusBufo glaberrimus
Bufo haematiticusBufo nasicus
Bufo peltocephalusBufo fustigerBufo empusus
Bufo taladaiBufo gundlachi
Bufo longinasusBufo guentheri
Bufo lemurBufo cophotisBufo variegatus
Dendrophryniscus minutusOsornophryne bufoniformis
Osornophryne antisanaOsornophryne puruanta
Osornophryne guacamayoOsornophryne sumacoensis
Atelopus senexAtelopus varius
Atelopus zetekiAtelopus chiriquiensis
Atelopus spurrelliAtelopus longirostris
Atelopus bomolochosAtelopus ignescens
Atelopus halihelosAtelopus peruensis
Atelopus flavescensAtelopus franciscus
Atelopus spumariusAtelopus pulcherAtelopus seminiferus
Melanophryniscus klappenbachiMelanophryniscus stelzneri
Melanophryniscus rubriventris
96
100
75
90
100
100
6170
90
99
10097
7270
10089
99100100
10097
67
100100
10077
84
58
90
65
100
100 100
100
100 100
84
95
100
6495
74
54
100
100
10053
99
7683
10064
75
100
63
99
6478
73100
67
100
87
100
100
99
10072
100
95
6661
94
100
61 9973
100
100
7098
9081100
100
100
100 100100
100
100
Bufonidae
Bufonidae cont.AD
Fig. 2 (continued)
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 577

Author's personal copy
Bufo parietalisBufo brevirostris
Bufo melanostictusBufo atukoraleiBufo scaber
Bufo himalayanusBufo stuarti
Bufo crocusBufo stomaticus
Bufo hololiusBufo dhufarensisBufo koynayensis
Adenomus kelaartiiBufo brongersmai
Nectophrynoides minutusNectophrynoides tornieri
Nectophrynoides viviparusChuramiti maridadi
Bufo variabilisBufo viridis
Bufo balearicusBufo pewzowiBufo oblongusBufo siculus
Bufo boulengeriSchismaderma carens
Pedostibes tuberculosusAnsonia minuta
Ansonia platysomaAnsonia hanitschi
Ansonia spinuliferAnsonia malayanaAnsonia longidigita
Ansonia leptopusAnsonia muelleri
Ansonia fuligineaPelophryne signata
Pelophryne brevipesPelophryne misera
Didynamipus sjostedtiAnsonia ornata
Bufo galeatusBufo philippinicus
Bufo macrotisBufo celebensis
Bufo divergensBufo biporcatus
Sabahphrynus maculatusPedostibes hosiiPedostibes rugosusBufo juxtasper
Bufo asperBufo tibetanusBufo tuberculatusBufo stejnegeri
Bufo japonicusBufo torrenticola
Bufo bankorensisBufo gargarizansBufo aspinius
Bufo cryptotympanicusBufo tuberospinius
Bufo bufoBufo verrucosissimus
Bufo raddeiBufo calamita
Leptophryne borbonicaNectophryne batesii
Nectophryne afraWolterstorffina parvipalmata
Werneria mertensianaBufo xerosBufo garmaniBufo gutturalis
Bufo gracilipesBufo kisoloensis
Bufo camerunensisBufo tuberosus
Bufo latifronsBufo vertebralis
Bufo maculatusBufo regularis
Bufo lemairiiBufo brauni
Bufo poweriBufo pantherinus
Bufo pardalisBufo steindachneri
Bufo mauritanicusCapensibufo roseiCapensibufo tradouwi
Stephopaedes loveridgeiStephopaedes anotisBufo lindneri
Mertensophryne micranotisBufo taitanus
Bufo uzunguensisBufo damaranus
Bufo dombensisBufo fenoulheti
Bufo gariepensisBufo robinsoni
Bufo amatolicusBufo inyangae
Bufo angusticeps
99
73
9862
10099
10055
100
59100
97
98100 95
10081
59
55
91
60
9894
97 74
94100
70
100 92
100
5268
90
98
90
100
100
97
81
97
100100
98 73
100
89
100
96
82
91
100100
91
82
55
80
8157
100
99
9984
100 100
100
89
100
100
56Bufonidae cont.
AE
Fig. 2 (continued)
578 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
(Fig. 2AA) show that Leptodactylidae is paraphyletic with respectto Leiuperidae, given that the leptodactylid genera Paratelmatobiusand Scythrophrys are more closely related to leiuperids than theyare to other leptodactylids (although this is not strongly sup-ported). We solve this taxonomic problem by expanding Leptodac-tylidae to again include all leiuperid genera, and recognizingLeiuperinae and Leptodactylinae as subfamilies. This arrangementtreats Leptodactylidae as the strongly supported clade (BS = 79%)of mostly foam-nesting frogs that has long been recognized asbehaviorally and morphologically distinctive (as the leptodactylineleptodactylids; Lynch, 1971; Duellman and Trueb, 1994). We con-sider this preferable to recognizing a new family for Paratelmatobi-us and Scythrophrys, genera for which we recognize a newsubfamily (Paratelmatobiinae subfam. nov.; Appendix A), to avoida paraphyletic Leptodactylinae.
According to our results, the family Strabomantidae (sensu AW,ASW) is also paraphyletic (Fig. 2R). Strabomantidae is a recentlydescribed family (Hedges et al., 2008) that includes many speciesthat were traditionally classified as Eleutherodactylus (in the familyLeptodactylidae; see ASW). Specifically, we find that the strabo-mantid genera Hypodactylus and Strabomantis are more closely re-lated to craugastorids (Haddadus, Craugastor) than they are to theother sampled strabomantid genera (e.g., Barycholos, Bryophryne,Holoaden, Lynchius, Noblella, Oreobates, Phrynopus, Pristimantis,Psychrophrynella). To resolve this problem (and to reduce the pro-liferation of terraranan families), we subsume Strabomantidae intoCraugastoridae, given that Craugastoridae occurs first in Hedgeset al. (2008), though we recognize that this is an arbitrary criterion.Within Craugastoridae, we recognize a subfamily Craugastorinae(Fig. 2R) that corresponds to the previously recognized family(e.g., Hedges et al., 2008; AW, ASW).
The former members of Strabomantidae are apportioned amongthree strongly supported clades (which we recognize as subfamilieswithin Craugastoridae) and one weakly supported clade (which weconsider incertae sedis). One of the strongly supported clades con-sists solely of the genus Strabomantis, for which we recognizea restricted subfamily Strabomantinae (Appendix B). Another com-prises the genera Barycholos, Bryophryne, Holoaden, Noblella, andPsychrophrynella, for which we use the previously recognized sub-family Holoadeninae (e.g., ASW). The third strongly supported cladecomprises the genera Lynchius, Oreobates, Phrynopus, and Pristiman-tis. We recognize this clade as a new subfamily, Pristimantinae sub-fam. nov. (see Appendix A for formal definition). The weaklysupported clade is the genus Hypodactylus. Rather than recognizinga new subfamily for this genus (or allowing it to render other sub-families non-monophyletic), we consider this genus incertae sedis,along with several other genera of the former Strabomantidae thatwere not included in this (or other molecular phylogenies), includ-ing Atopophrynus, Dischiodactylus, Geobatrachus, Mucubatrachus,Niceforonia, and Paramophrynella. In theory, some of these closely re-lated subfamilies could be combined (e.g., Craugastorinae and Strab-omantinae or Holoadeninae and Pristimantinae), but the resultingsubfamilies would then not be strongly supported.
As in most previous studies (Darst and Cannatella, 2004; Roe-lants et al., 2007; Wiens, 2007a, 2011), we find strong supportfor monophyly of Hyloidea, but relationships among the familiesare weakly supported (Figs. 1 and 2P-AE). Therefore, we do notpresent extensive comparison of among-family relationships be-tween our study and previous studies. Nevertheless, there aresome intriguing patterns that occur in multiple studies, such asplacement of bufonids with dendrobatids (e.g., this study, Frostet al., 2006; Roelants et al., 2007; Wiens, 2011). We also corrobo-rate the monophyly and current composition of Brachycephalidae(Hedges et al., 2008), Hemiphractidae (Wiens et al., 2007a,b;Wiens, 2011), Hylidae (with subfamilies Hylinae, Phyllomedusinae,and Pelodryadinae; Wiens et al., 2010), Bufonidae (Pramuk et al.,
2008; Van Bocxlaer et al., 2009), Allophrynidae and Centrolenidae.Within Centrolenidae (Fig. 2AB), we support the subfamilies Cen-troleninae and Hyalinobatrachinae, and we place the previouslyincertae sedis genus Ikakogi in Centroleninae with strong (70%)bootstrap support. We do not recognize a family Aromobatidaeseparate from Dendrobatidae, as there is no phylogenetic justifica-tion for splitting Dendrobatidae (Fig. 2AC). We follow Santos et al.(2009) in recognizing a single family Dendrobatidae, including theformer aromobatids and containing no subfamilies (see also AW).
Our higher-level phylogeny within Ranoidea is generally similarto other recent estimates (e.g., Frost et al., 2006; Roelants et al., 2007;Wiens, 2011), and many of these relationships are well supported inour study and previous studies (e.g., among brevicipitids, hemisot-ids, microhylids, hyperoliids, arthroleptids). Our results also supportcurrent delimitation of subfamilies (Figs. 1 and 2H–I; see AppendixB) in Microhylidae (van der Meijden et al., 2007) and Arthroleptidae(including Astylosterninae; see ASW). Some previous taxonomies(e.g., Bossuyt et al., 2006; Wiens et al., 2009) and current taxonomies(including AW) recognize a large (>1400 species) family Ranidae(with 13 subfamilies) as the sister group to all other ranoids. Otherrecent authors have considered these subfamilies distinct families(Bossuyt and Roelants, 2009; Frost et al., 2006; Roelants et al.,2007). We follow this latter arrangement here, recognizing the fam-ilies Ptychadenidae, Micrixalidae, Phrynobatrachidae, Conrauidae,Petropedetidae, Pyxicephalidae, Nyctibatrachidae, Ceratobatrachi-dae, Ranixalidae, Dicroglossidae, Ranidae, and Rhacophoridae, andMantellidae, all of which are strongly supported (Figs. 1 and 2J–O).Within these families, subfamilies follow Frost (2011): within Pyxi-cephalidae, we recognize Cacosterninae and Pyxicephalinae;Dicroglossidae comprises Occidozyginae and Dicroglossinae; Rhac-ophoridae consists of Buergeriinae and Rhacophorinae; and Mantel-lidae is composed of Laliostominae, Boophinae, and Mantellinae(Figs. 1 and 2J–O). Relationships among many of these families re-main poorly supported, as in previous studies (Roelants et al.,2007; Wiens et al., 2009). Given the extensive sampling of taxa inthis clade, resolving their relationships may instead require addingmany more characters (i.e., more genes).
4. Discussion
4.1. Systematics of extant lissamphibians
Here, we provide a large-scale estimate of amphibian phylogeny,containing over 2800 species (42% of the known extant diversity).This phylogeny is largely congruent with those of several recentmolecular analyses (Frost et al., 2006; Roelants et al., 2007; Wiens,2007a, 2011). Nevertheless, our increased sampling of taxa andcharacters (especially among former leptodactylid frogs) revealsthat several currently recognized families are not monophyletic ascurrently delimited (i.e., Ceratophryidae, Cycloramphidae, Lepto-dactylidae, Strabomantidae). To correct these problems, we presenta new classification of extant amphibians. We have been as conser-vative as possible in our taxonomic changes, such that deviationsfrom current taxonomy are made only when necessitated by non-monophyly of higher taxa, and in such a way that newly recognizedtaxa are strongly supported (with the exception of Alsodidae as de-scribed above). It is possible that future phylogenies might revealsome of our changes to be unnecessary (e.g., in cases where therewas weak support for non-monophyly of families, future analysesmight strongly resolve them as monophyletic). However, by focus-ing on only strongly supported clades, the higher taxa that we recog-nize should not be misleading, even as new data are added. We arealso conservative in that we only address taxonomy at the subfamilylevel and above, given that our sampling of genera is mostly com-plete whereas our sampling of species is not. Nevertheless, our study
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 579

Author's personal copy
corroborates previous studies showing the paraphyly of numerousgenera (e.g., Bufo, Rana) and shows many additional genera to benon-monophyletic (e.g., Batrachyla). Importantly, our study alsoprovides a framework to which additional sequences can be readilyadded. Our data matrix is available at Dryad repository doi:10.5061/dryad.vd0m7. It should thus be relatively straightforward to addmore taxa to this tree.
4.2. Supermatrices and large-scale phylogenetic inference
This analysis corroborates several recent studies suggesting thatthe supermatrix approach is a powerful strategy for large-scalephylogenetic inference (e.g., Driskell et al., 2004; McMahon andSanderson, 2006; Thomson and Shaffer, 2010; Pyron et al., 2011;Wiens et al., 2005b). For example, even though each taxon had(on average) 80% missing data, we found that most species wereplaced in the families and genera expected based on previous tax-onomy, often with very strong support. In cases where we foundthat currently recognized families were not monophyletic, thiswas not due to species being randomly placed on the tree, butrather due to differences in the placement of strongly supportedclades of species and genera. Moreover, all of our 67 families andall but two of the 50 subfamilies are strongly supported.
Despite the overall strong support for most of the tree (i.e., 64%of all nodes have BS >70), certain clades remain poorly supported.Therefore, a potential criticism of the supermatrix approach is thatthis poor support may have arisen due to missing data in manytaxa. However, several lines of evidence suggest that this is notthe case. First, many previous studies have shown that there is typ-ically little relationship between the support for clades and theamount of missing data in the included species (e.g., Pyron et al.,2011; Wiens et al., 2005b, and for an analysis of multiple datasetssee Wiens and Morrill, 2011). Second, we find that most of theclades that are weakly supported in our study are also weakly sup-ported in other studies that have more limited missing data (andthe same is true for strongly supported clades). For example, intheir study of extant amphibian phylogeny based on multiple nu-clear and mitochondrial genes (with little missing data), Roelantset al. (2007) showed weak support for relationships among hyloidfamilies, microhylid subfamilies, and the families of former ranids,but strong support for relationships among the major clades offrogs, salamanders, and caecilians. These same patterns of weakand strong support appear in our study (Fig. 1). Previous studiessuggest that these patterns of support are more likely to reflectunderlying branch lengths, such as weak support for shortbranches (Wiens et al., 2008), rather than impacts of missing data.
Our results and those of other studies suggest that extensivemissing data need not be an impediment to constructing large-scale phylogenies with the supermatrix approach. Nevertheless,one aspect of our sampling design may have greatly facilitatedthe integration of the different genes used in this study. Specifi-cally, 90% of the species had sequence data for the 16S gene and81% had data for 12S. Thus, most species had comparable datafor at least one gene, which may have greatly facilitated placingthem on the tree with only a limited amount of character data(Wiens et al., 2005b). Although it is possible to reconstruct a phy-logeny when missing and non-missing data are randomly distrib-uted among characters, the number of genes and charactersrequired to obtain accurate phylogenies may be much higher thanwhen the non-missing data are all in the same characters (e.g.,Wiens, 2003). A clear lesson for future studies is that adding taxato this large-scale phylogeny will be greatly facilitated if research-ers include these same 12S and 16S genes, along with widely sam-pled nuclear genes such as RAG-1. Increasing sampling of the othernuclear genes may be advantageous as well.
Acknowledgments
We thank the many researchers who made this study possiblethrough their detailed studies of amphibian phylogeny with a (mostly)shared set of molecular markers, and uploading their sequence data toGenBank. We would like to thank F.T. Burbrink (CUNY-CSI), J. Lom-bardo (CUNY-CSI), and T.J. Guiher (CUNY-CSI) for computational assis-tance, and D.R. Frost (AMNH) and A.M. Bauer (Villanova) for advice ontaxonomy. This research was supported in part by US National ScienceFoundation grants to R.A.P. (DBI-0905765), J.J.W. (EF 0334923), and theCUNY HPCC (CNS-0958379 and CNS-0855217). We would like to thankA. Larson, and two anonymous reviewers for helpful comments thatsubstantially improved this manuscript.
Appendix A. Description of new subfamilies
Pristimantinae subfam. nov. (family Craugastoridae)Type: genus and species Pristimantis galdi Jiménez de la
Espada, 1870.Content: four genera, >465 species; Lynchius, Oreobates,
Phrynopus, Pristimantis.Phylogenetic Definition: this subfamily consists of the most
recent common ancestor of Lynchius, Oreobates, Phrynopus,and Pristimantis, and all descendants thereof.
Distribution: these frogs are primarily restricted to the Andesand Amazon Basin of South America (Hedges et al., 2008).
Remarks: The four genera of this subfamily are grouped withstrong support (81% BS proportion; Fig. 2R), and we findweak support for their placement as the sister group toHoloadeninae (Fig. 2R).
Paratelmatobiinae subfam. nov. (family Leptodactylidae)Type: genus and species Paratelmatobius lutzii Lutz and
Carvalho 1958.Content: two genera, eight species; Paratelmatobius and
Scythrophrys.Phylogenetic Definition: this subfamily consists of the most
recent common ancestor of Paratelmatobius andScythrophrys, and all descendants thereof.
Distribution: these frogs are primarily restricted to southernand eastern Brazil (AW; ASW).
Remarks: The two genera of this subfamily are grouped withstrong support (93% BS proportion), and we find weaksupport for placing them as the sister group of Leiuperinae(Fig. 2AA).
Appendix B. Generic content of families and subfamilies
The list below accounts for all extant amphibian genera currentlyrecognized by AW and ASW, with family and subfamily classificationupdated to reflect our phylogenetic estimate and taxonomic changes.The 72 genera not sampled in our phylogeny are either included intheir previously delimited groups (when these were found to bestrongly supported), or considered incertae sedis (along with anysampled taxa explicitly delimited as such) when their placementwas ambiguous due to non-monophyly of previously recognizedhigher taxa. As noted above, Lissamphibia also contains numerousextinct taxa, including gymnophionans, caudates, and anurans,which are not considered here (Marjanovic and Laurin, 2007).
B.1. Gymnophiona
Caeciliidae: incertae sedis: (Atretochoana, Brasilotyphlus, Idiocra-nium, Indotyphlus, Microcaecilia, Mimosiphonops, Nectocaecilia, Par-
580 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
vicaecilia, Potomotyphlus, Sylvacaecilia); Caeciliinae (Caecilia,Chthonerpeton, Dermophis, Gegeneophis, Geotrypetes, Grandisonia,Gymnopis, Hypogeophis, Luetkenotyphlus, Oscaecilia, Praslinia,Typhlonectes, Schistometopum, Siphonops), Herpelinae (Herpele,Boulengerula), Scolecomorphinae (Crotaphotrema, Scolecomorphus);Ichthyophiidae: Caudacaecilia, Ichthyophis, Uraeotyphlus; Rhinatr-ematidae: Epicrionops, Rhinatrema.
B.2. Caudata
Ambystomatidae: Ambystoma; Amphiumidae: Amphiuma;Cryptobranchidae: Andrias, Cryptobranchus; Dicamptodontidae:Dicamptodon; Hynobiidae: Batrachuperus, Hynobius, Liua, Onycho-dactylus, Pachyhynobius, Paradactylodon, Protohynobius, Pseudohyno-bius, Ranodon, Salamandrella; Plethodontidae: Bolitoglossinae(Batrachoseps, Bolitoglossa, Bradytriton, Chiropterotriton, Cryptotriton,Dendrotriton, Ixalotriton, Lineatriton, Nototriton, Nyctanolis, Oedipina,Parvimolge, Pseudoeurycea, Thorius), Hemidactylinae (Hemidactyli-um), Plethodontinae (Aneides, Desmognathus, Ensatina, Hydromantes,Karsenia, Phaeognathus, Plethodon), Spelerpinae (Eurycea, Gyrinophi-lus, Haideotriton, Pseudotriton, Stereochilus, Urspelerpes); Proteidae:Necturus, Proteus; Rhyacotritonidae: Rhyacotriton; Salamandri-dae: Pleurodelinae (Calotriton, Cynops, Echinotriton, Euproctus,Ichthyosaura, Laotriton, Lissotriton, Neurergus, Notophthalmus,Ommatotriton, Pachytriton, Paramesotriton, Pleurodeles, Taricha, Trit-urus, Tylototriton), Salamandrinae (Chioglossa, Lyciasalamandra, Mer-tensiella, Salamandra), Salamandrininae (Salamandrina); Sirenidae:Siren, Pseudobranchus.
B.3. Anura
Incertae sedis in Hyloidea (Crossodactylodes, Rupirana,Zachaenus).
Allophrynidae: Allophryne; Alsodidae: Alsodes, Eupsophus, Lim-nomedusa; Alytidae: Alytes; Arthroleptidae: Arthroleptinae (Athro-leptis, Cardioglossa), Astylosterninae (Astylosternus, Leptodactylodon,Nyctibates, Scotobleps, Trichobatrachus), Leptopelinae (Leptopelis);Ascaphidae: Ascaphus; Batrachylidae: Atelognathus, Batrachyla;Bombinatoridae: Barbaroula, Bombina; Brachycephalidae: Brachy-cephalus, Ischnocnema; Brevicipitidae: Balebreviceps, Breviceps,Callulina, Probreviceps, Spelaeophryne; Bufonidae: Adenomus, Altiph-rynoides, Andinophryne, Ansonia, Atelopus, Bufo, Bufoides, Capensi-bufo, Churamiti, Crepidophryne, Dendrophryniscus, Didynamipus,Frostius, Laurentophryne, Leptophryne, Melanophryniscus, Merten-sophryne, Metaphryniscus, Nectophryne, Nectophrynoides, Nimbaph-rynoides, Oreophrynella, Osornophryne, Parapelophryne, Pedostibes,Pelophryne, Pseudobufo, Rhamphophryne, Sabahphrynus, Schismader-ma, Spinophrynoides, Stephopaedes, Truebella, Werneria, Wolterstorffi-na; Calyptocephalellidae: Calyptocephalella, Telmatobufo;Centrolenidae: Centroleninae (Centrolene, Chimerella, Cochranella,Espadarana, Ikakogi, Nymphargus, Rulyrana, Sachatamia, Teratohyla,Vitreorana), Hyalinobatrachinae (Celsiella, Hyalinobatrachium); Cer-atobatrachidae: Batrachylodes, Ceratobatrachus, Discodeles, Inger-ana, Palmatorappia, Platymantis; Ceratophryidae: Ceratophrys,Chacophrys, Lepidobatrachus; Ceuthomantidae: Ceuthomantis; Con-rauidae: Conraua; Craugastoridae: incertae sedis (Atopophrynus,Dischiodactylus, Geobatrachus, Hypodactylus, Mucubatrachus, Nicef-oronia, Paramophrynella), Craugastorinae (Craugastor, Haddadus),Holoadeninae (Barycholos, Bryophryne, Euparkerella, Holoaden,Noblella, Psychrophrynella), Pristimantinae (Lynchius, Oreobates,Phrynopus, Pristimantis), Strabomantinae (Strabomantis); Cycloram-phidae: Cycloramphus, Thoropa; Dendrobatidae: Allobates, Ameere-ga, Anomaloglossus, Aromobates, Colostethus, Dendrobates,Epipedobates, Hyloxalus, Mannophryne, Phyllobates, Rheobates,Silverstoneia; Dicroglossidae: Dicroglossinae (Chaparana, Chirixalus,Euphlyctis, Fejervarya, Hoplobatrachus, Limnonectes, Nannophrys,
Nanorana, Paa, Sphaerotheca), Occidozyginae (Ingerana, Occidozyga);Discoglossidae: Discoglossus; Eleutherodactylidae: Eleutherodac-tylinae (Diasporus, Eleutherodactylus), Phyzelaphryninae (Adeloph-ryne, Phyzelaphryne); Heleophrynidae: Hadromophryne,Heleophryne; Hemiphractidae: Cryptobatrachus, Flectonotus, Gas-trotheca, Hemiphractus, Stefania; Hemisotidae: Hemisus; Hylidae:Hylinae (Acris, Anotheca, Aparasphenodon, Aplastodiscus,Argenteohyla, Bokermannohyla, Bromeliohyla, Charadrahyla, Corytho-mantis, Dendropsophus, Diaglena, Duellmanohyla, Ecnomiohyla, Exer-odonta, Hyla, Hyloscirtus, Hypsiboas, Isthmohyla, Itapotihyla,Megastomatohyla, Myersiohyla, Nyctimantis, Osteocephalus, Osteopi-lus, Phyllodytes, Plectrohyla, Pseudacris, Pseudis, Ptychohyla, Scarthyla,Scinax, Smilisca, Sphaenorhynchus, Tepuihyla, Tlalocohyla, Trachy-cephalus, Triprion, Xenohyla), Pelodryadinae (Litoria), Phyllomedusi-nae (Agalychnis, Cruziohyla, Hylomantis, Pachymedusa, Phasmahyla,Phrynomedusa, Phyllomedusa); Hylodidae: Crossodactylus, Hylodes,Megaelosia; Hyperoliidae: Acanthixalus, Afrixalus, Alexteroon, Arl-equinus, Callixalus, Chlorolius, Chrysobatrachus, Cryptothylax, Heterix-alus, Hyperolius, Kassina, Kassinula, Morerella, Opisthothylax,Paracassina, Phlyctimantis, Semnodactylus, Tachycnemis; Leiopel-matidae: Leiopelma; Leptodactylidae: Leptodactylinae (Adenomera,Hydrolaetere, Leptodactylus, Lithodytes), Leiuperinae (Edalorhina,Engystomops, Eupemphix, Physalaemus, Pleurodema, Pseudopaludico-la, Somuncuria), Paratelmatobiinae (Paratelmatobius, Scythrophrys);Mantellidae: Boophinae (Boophis), Laliostominae (Aglyptodactylus,Laliostoma), Mantellinae (Blommersia, Boehmantis, Gephyromantis,Guibemantis, Mantella, Mantidactylus, Spinomantis, Tsingymantis,Wakea); Megophryidae: Borneophrys, Brachytarsophrys, Leptobrach-ella, Leptobrachium, Leptolalax, Megophrys, Ophryophryne, Oreolalax,Scutiger, Vibrissaphora, Xenophrys; Micrixalidae: Micrixalus; Micro-hylidae: incertae sedis (Gastrophrynoides, Paramophrynella),Asterophryninae (Albericus, Aphantophryne, Asterophrys, Austrocha-perina, Barygenys, Callulops, Choerophryne, Cophixalus, Copiula, Geny-ophryne, Hylophorbus, Liophryne, Mantophryne, Oreophryne,Oxydactyla, Pherohapsis, Sphenophryne, Xenorhina), Cophylinae(Anodonthyla, Cophyla, Madecassophryne, Platypelis, Plethodontohyla,Rhombophryne, Stumpffia), Dyscophinae (Dyscophus), Gastrophryni-nae (Adelastes, Altigius, Arcovomer, Chiasmocleis, Ctenophryne, Dasy-pops, Dermatonotus, Elachistocleis, Gastrophryne, Hamptophryne,Hyophryne, Hypopachus, Melanophryne, Myersiella, Nelsonophryne,Relictivomer, Stereocyclops, Syncope), Hoplophryninae (Hoplophryne,Parhoplophryne), Kalophryninae (Kalophrynus), Melanobatrachinae(Melanobatrachus), Microhylinae (Calluella, Chaperina, Glyphoglos-sus, Kaloula, Metaphrynella, Microhyla, Micryletta, Ramanella, Uper-odon), Otophryninae (Otophryne, Synapturanus), Phrynomerinae(Phrynomantis), Scaphiophryne (Paradoxophyla, Scaphiophryne);Myobatrachidae: Limnodynastinae (Adelotus, Heleioporus, Lechrio-dus, Limnodynastes, Mixophyes, Neobatrachus, Notaden, Philoria,Platyplectrum, Pseudophryne), Myobatrachinae (Arenophryne, Assa,Crinia, Geocrinia, Metacrinia, Myobatrachus, Paracrinia, Rheobatra-chus, Spicospina, Taudactylus, Uperoleia); Nasikabatrachidae: Nasik-abatrachus; Nyctibatrachidae: Lankanectes, Nyctibatrachus;Odontophrynidae: Macrogenioglottus, Odontophrynus, Proceratoph-rys; Pelobatidae: Pelobates; Pelodytidae: Pelodytes; Petropedeti-dae: Petropedetes; Phrynobatrachidae: Phrynobatrachus; Pipidae:Hymenochirus, Pipa, Pseudhymenochirus, Silurana, Xenopus; Ptycha-denidae: Hildebrandtia, Lanzarana, Ptychadena; Pyxicephalidae:Cacosterninae (Amietia, Anhydrophryne, Arthroleptella, Cacosternum,Ericabatrachus, Microbatrachella, Natalobatrachus, Nothophryne,Poyntonia, Strongylopus, Tomopterna), Pyxicephalinae (Aubria,Pyxicephalus); Ranidae: Amnirana, Amolops, Huia, Hylarana, Meris-togenys, Odorrana, Pseudoamolops, Pterorana, Rana, Staurois; Ranix-alidae: Indirana; Rhacophoridae: Buergeriinae (Buergeria),Rhacophorinae (Chiromantis, Dendrobatorana, Feihyla, Ghatixalus,Gracixalus, Kurixalus, Liuixalus, Nyctixalus, Philautus, Polypedates,Rhacophorus, Theloderma), Rhinodermatidae: Insuetophrynus,
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 581

Author's personal copy
Rhinoderma; Rhinophrynidae: Rhinophrynus; Scaphiopodidae:Scaphiopus, Spea; Sooglossidae: Sechellophryne, Sooglossus; Telma-tobiidae: Batrachophrynus, Telmatobius.
Appendix C. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ympev.2011.06.012.
References
Alfaro, M.E., Santini, F., Brock, C., Alamillo, H., Dornburg, A., Rabosky, D.L., Carnevale,G., Harmon, L.J., 2009. Nine exceptional radiations plus high turnover explainspecies diversity in jawed vertebrates. Proc. Natl. Acad. Sci. USA 106, 13410–13414.
Biju, S.D., Bossuyt, F., 2003. New frog family from India reveals an ancientbiogeographical link with the Seychelles. Nature 425, 711–714.
Blaustein, A.R., Wake, D.B., 1990. Declining amphibian populations – a globalphenomenon. Trends Ecol. Evol. 5, 203–204.
Bossuyt, F., Roelants, K., 2009. Anura. In: Hedges, S.B., Kumar, S. (Eds.), The Timetreeof Life. Oxford University Press, New York, pp. 357–364.
Bossuyt, F., Brown, R.M., Hillis, D.M., Cannatella, D.C., Milinkovitch, M.C., 2006.Phylogeny and biogeography of a cosmopolitan frog radiation: late Cretaceousdiversification resulted in continent-scale endemism in the family Ranidae.Syst. Biol. 55, 579–594.
Camp, C.D., Peterman, W.E., Milanovich, J.R., Lamb, T., Maerz, J.C., Wake, D.B., 2009.A new genus and species of lungless salamander (family Plethodontidae) fromthe Appalachian highlands of the south-eastern United States. J. Zool. 279, 86–94.
Carroll, R.L., 2009. The Rise of Amphibians: 365 Million Years of Evolution. JohnsHopkins University Press, Baltimore.
Chippindale, P.T., Bonett, R.M., Baldwin, A.S., Wiens, J.J., 2004. Phylogenetic evidencefor a major reversal of life-history evolution in plethodontid salamanders.Evolution 58, 2809–2822.
Darst, C.R., Cannatella, D.C., 2004. Novel relationships among hyloid frogs inferredfrom 12S and 16S mitochondrial DNA sequences. Mol. Phylogenet. Evol. 31,462–475.
de Queiroz, A., Gatesy, J., 2007. The supermatrix approach to systematics. TrendsEcol. Evol. 22, 34–41.
Driskell, A.C., Ane, C., Burleigh, J.G., McMahon, M.M., O’Meara, B.C., Sanderson, M.J.,2004. Prospects for building the Tree of Life from large sequence databases.Science 306, 1172–1174.
Duellman, W.E., 1999. Patterns of Distribution of Amphibians: A Global Perspective.Johns Hopkins University Press, Baltimore.
Duellman, W.E., Trueb, L., 1994. Biology of Amphibians. Johns Hopkins UniversityPress, Baltimore.
Edgar, R.C., 2004. MUSCLE: multiple sequence alignment with high accuracy andhigh throughput. Nucl. Acids Res. 32, 1792–1797.
Faivovich, J., Haddad, C.F.B., Garcia, P.C.A., Frost, D.R., Campbell, J.A., Wheeler, W.C.,2005. Systematic review of the frog family Hylidae, with special reference toHylinae: phylogenetic analysis and taxonomic revision. Bull. Am. Mus. Natl.Hist. 294, 6–228.
Faivovich, J., Haddad, C.F.B., Baeta, D., Jungfer, K.H., Alvares, G.F.R., Brandao, R.A.,Sheil, C., Barrientos, L.S., Barrio-Amoros, C.L., Cruz, C.A.G., Wheeler, W.C., 2010.The phylogenetic relationships of the charismatic poster frogs,Phyllomedusinae (Anura, Hylidae). Cladistics 26, 227–261.
Feller, A.E., Hedges, S.B., 1998. Molecular evidence for the early history of livingamphibians. Mol. Phylogenet. Evol. 9, 509–516.
Felsenstein, J., 2004. Inferring Phylogenies. Sinauer Associates, Sunderland, Mass.Frost, Darrel, R., 2011. Amphibian Species of the World: an Online Reference.
Version 5.5 (31 January, 2011). American Museum of Natural History, NewYork, USA. Electronic Database accessible at http://research.amnh.org/vz/herpetology/amphibia/.
Frost, D.R., Grant, T., Faivovich, J., Bain, R.H., Haas, A., Haddad, C.F.B., De Sa, R.O.,Channing, A., Wilkinson, M., Donnellan, S.C., Raxworthy, C.J., Campbell, J.A.,Blotto, B.L., Moler, P., Drewes, R.C., Nussbaum, R.A., Lynch, J.D., Green, D.M.,Wheeler, W.C., 2006. The amphibian tree of life. Bull. Am. Mus. Natl. Hist. 297,8–370.
Grant, T., Frost, D.R., Caldwell, J.P., Gagliardo, R., Haddad, C.F.B., Kok, P.J.R., Means,D.B., Noonan, B.P., Schargel, W.E., Wheeler, W.C., 2006. Phylogeneticsystematics of dart-poison frogs and their relatives (Amphibia:Athesphatanura: Dendrobatidae). Bull. Am. Mus. Natl. Hist. 299, 6–262.
Guayasamin, J.M., Castroviejo-Fisher, S., Trueb, L., Ayarzaguena, J., Rada, M., Vila, C.,2009. Phylogenetic systematics of Glassfrogs (Amphibia: Centrolenidae) andtheir sister taxon Allophryne ruthveni. Zootaxa 2100, 1–97.
Günther, A.C.L.G., 1858. On the systematic arrangement of the tailless batrachiansand the structure of Rhinophrynus dorsalis. Proc. Zool. Soc. Lond. 1858, 339–352.
Hedges, S.B., Duellman, W.E., Heinicke, M.P., 2008. New World direct-developingfrogs (Anura: Terrarana): molecular phylogeny, classification, biogeography,and conservation. Zootaxa 1737, 1–182.
Heinicke, M.P., Duellman, W.E., Trueb, L., Means, D.B., MacCulloch, R.D., Hedges, S.B.,2009. A new frog family (Anura: Terrarana) from South America and an
expanded direct-developing clade revealed by molecular phylogeny. Zootaxa2211, 1–35.
Hugall, A.F., Foster, R., Lee, M.S.Y., 2007. Calibration choice, rate smoothing, and thepattern of tetrapod diversification according to the long nuclear gene RAG-1.Syst. Biol. 56, 543–563.
Kozak, K.H., Mendyk, R.W., Wiens, J.J., 2009. Can parallel diversification occur insympatry? Repeated patterns of body-size evolution in coexisting clades ofNorth American salamanders. Evolution 63, 1769–1784.
Lannoo, M.J., 2005. Amphibian Declines: The Conservation Status of United StatesSpecies. University of California Press, Berkeley.
Larkin, M.A., Blackshields, G., Brown, N.P., Chenna, R., McGettigan, P.A., McWilliam,H., Valentin, F., Wallace, I.M., Wilm, A., Lopez, R., Thompson, J.D., Gibson, T.J.,Higgins, D.G., 2007. ClustalW and ClustalX version 2.0. Bioinformatics 23, 2947–2948.
Laurent, R.F., 1984. Heterogeneidad de la familia Caeciliidae (Amphibia–Apoda).Acta Zool. Lilloana 37, 199–200.
Lynch, J.D., 1971. Evolutionary relationships, osteology, and zoogeography ofleptodactyloid frogs. Misc. Pub. Mus. Natl. Hist. Kansas 58, 1–238.
Marjanovic, D., Laurin, M., 2007. Fossils, molecules, divergence times, and the originof lissamphibians. Syst. Biol. 56, 369–388.
McMahon, M.M., Sanderson, M.J., 2006. Phylogenetic supermatrix analysis ofGenBank sequences from 2228 papilionoid legumes. Syst. Biol. 55, 818–836.
Miranda-Ribeiro, A.D., 1920. Algumas consideracões sobre Holoaden lüderwaldti egeneros correlatos. Rev. Mus. Paulista, São Paulo 12, 319–320.
Pauly, G.B., Hillis, D.M., Cannatella, D.C., 2004. The history of a Nearcticcolonization: molecular phylogenetics and biogeography of the NearcticToads (Bufo). Evolution 58, 2517–2535.
Pauly, G.B., Hillis, D.M., Cannatella, D.C., 2009. Taxonomic freedom and the role ofofficial lists of species names. Herpetologica 65, 115–128.
Peng, R., Zhang, P., Xiong, J.-L., Gu, H.-J., Zeng, X.-M., Zou, F.-D., 2010. Rediscovery ofProtohynobius puxiongensis (Caudata: Hynobiidae) and its phylogenetic positionbased on complete mitochondrial genomes. Mol. Phylogenet. Evol. 56, 252–258.
Pramuk, J.B., Robertson, T., Sites, J.W., Noonan, B.P., 2008. Around the world in 10million years: biogeography of the nearly cosmopolitan true toads (Anura:Bufonidae). Glob. Ecol. Biogeogr. 17, 72–83.
Pyron, R.A., 2010. A likelihood method for assessing molecular divergence timeestimates and the placement of fossil calibrations. Syst. Biol. 59, 185–194.
Pyron, R.A., 2011. Divergence time estimation using fossils as terminal taxa and theorigins of Lissamphibia. Syst. Biol. 60, 466–481.
Pyron, R.A., Burbrink, F.T., 2009. Systematics of the Common Kingsnake(Lampropeltis getula; Serpentes: Colubridae) and the burden of heritage intaxonomy. Zootaxa 2241, 22–32.
Pyron, R.A., Burbrink, F.T., Colli, G.R., de Oca, A.N.M., Vitt, L.J., Kuczynski, C.A., Wiens,J.J., 2011. The phylogeny of advanced snakes (Colubroidea), with discovery of anew subfamily and comparison of support methods for likelihood trees. Mol.Phylogenet. Evol. 58, 329–342.
Roelants, K., Gower, D.J., Wilkinson, M., Loader, S.P., Biju, S.D., Guillaume, K., Moriau,L., Bossuyt, F., 2007. Global patterns of diversification in the history of modernamphibians. Proc. Natl. Acad. Sci. USA 104, 887–892.
San Mauro, D., 2010. A multilocus timescale for the origin of extant amphibians.Mol. Phylogenet. Evol. 56, 554–561.
San Mauro, D., Vences, M., Alcobendas, M., Zardoya, R., Meyer, A., 2005. Initialdiversification of living amphibians predated the breakup of Pangaea. Am. Nat.165, 590–599.
San Mauro, D., Gower, D.J., Massingham, T., Wilkinson, M., Zardoya, R., Cotton, J.A.,2009. Experimental design in caecilian systematics: phylogenetic informationof mitochondrial genomes and nuclear RAG1. Syst. Biol. 58, 425–438.
Santos, J.C., Coloma, L.A., Summers, K., Caldwell, J.P., Ree, R., Cannatella, D.C., 2009.Amazonian amphibian diversity is primarily derived from Late Miocene Andeanlineages. PLoS Biol. 7, 448–461.
Stamatakis, A., 2006. RAxML-VI-HPC: maximum likelihood-based phylogeneticanalyses with thousands of taxa and mixed models. Bioinformatics 22, 2688–2690.
Stuart, S.N., Chanson, J.S., Cox, N.A., Young, B.E., Rodrigues, A.S.L., Fischman, D.L.,Waller, R.W., 2004. Status and trends of amphibian declines and extinctionsworldwide. Science 306, 1783–1786.
Thomson, R.C., Shaffer, H.B., 2010. Sparse supermatrices for phylogenetic inference.taxonomy, alignment, rogue taxa, and the phylogeny of living turtles. Syst. Biol.59, 42–58.
Van Bocxlaer, I., Biju, S.D., Loader, S.P., Bossuyt, F., 2009. Toad radiation reveals into-India dispersal as a source of endemism in the Western Ghats–Sri Lankabiodiversity hotspot. BMC Evol. Biol. 9, 131.
van der Meijden, A., Vences, M., Hoegg, S., Boistel, R., Channing, A., Meyer, A., 2007.Nuclear gene phylogeny of narrow-mouthed toads (Family: Microhylidae) and adiscussion of competing hypotheses concerning their biogeographical origins.Mol. Phylogenet. Evol. 44, 1017–1030.
Vieites, D.R., Wollenberg, K.C., Andreone, F., Kohler, J., Glaw, F., Vences, M., 2009.Vast underestimation of Madagascar’s biodiversity evidenced by an integrativeamphibian inventory. Proc. Natl. Acad. Sci. USA 106, 8267–8272.
Vieites, D.R., Roman, S.R., Wake, M.H., Wake, D.B., 2011. A multigenic perspective onphylogenetic relationships in the largest family of salamanders, thePlethodontidae. Mol. Phylogenet. Evol. 59, 623–635.
Wiens, J.J., 2003. Missing data, incomplete taxa, and phylogenetic accuracy. Syst.Biol. 52, 528–538.
Wiens, J.J., 2007a. Global patterns of diversification and species richness inamphibians. Am. Nat. 170, S86–S106.
582 R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583

Author's personal copy
Wiens, J.J., 2007b. Review of ‘‘The amphibian tree of life’’ by Frost et al. Quart. Rev.Biol. 82, 55–56.
Wiens, J.J., 2008. Systematics and herpetology in the age of genomics. Bioscience 58,297–307.
Wiens, J.J., 2011. Re-evolution of lost mandibular teeth in frogs after more than 200million years, and re-evaluating Dollo’s law. Evolution 65, 1283–1296.
Wiens, J.J., Morrill, M.C., 2011. Missing data in phylogenetic analysis: reconcilingresults from simulations and empirical data. Syst. Biol. doi:10.1093/sysbio/syr025.
Wiens, J.J., Bonett, R.M., Chippindale, P.T., 2005a. Ontogeny discombobulatesphylogeny: paedomorphosis and higher-level salamander relationships. Syst.Biol. 54, 91–110.
Wiens, J.J., Fetzner, J.W., Parkinson, C.L., Reeder, T.W., 2005b. Hylid frog phylogenyand sampling strategies for speciose clades. Syst. Biol. 54, 719–748.
Wiens, J.J., Graham, C.H., Moen, D.S., Smith, S.A., Reeder, T.W., 2006. Evolutionaryand ecological causes of the latitudinal diversity gradient in hylid frogs: treefrogtrees unearth the roots of high tropical diversity. Am. Nat. 168, 579–596.
Wiens, J.J., Kuczynski, C.A., Duellman, W.E., Reeder, T.W., 2007a. Loss and re-evolution of complex life cycles in marsupial frogs: does ancestral traitreconstruction mislead? Evolution 61, 1886–1899.
Wiens, J.J., Parra-Olea, G., Garcia-Paris, M., Wake, D.B., 2007b. Phylogenetic historyunderlies elevational biodiversity patterns in tropical salamanders. Proc. Roy.Soc. Lond. B – Biol. Sci. 274, 919–928.
Wiens, J.J., Kuczynski, C.A., Smith, S.A., Mulcahy, D.G., Sites, J.W., Townsend, T.M.,Reeder, T.W., 2008. Branch lengths, support, and congruence: testing thephylogenomic approach with 20 nuclear loci in snakes. Syst. Biol. 57, 420–431.
Wiens, J.J., Sukumaran, J., Pyron, R.A., Brown, R.M., 2009. Evolutionary andbiogeographic origins of high tropical diversity in Old World frogs (Ranidae).Evolution 63, 1217–1231.
Wiens, J.J., Kuczynski, C.A., Hua, X., Moen, D.S., 2010. An expanded phylogeny oftreefrogs (Hylidae) based on nuclear and mitochondrial sequence data. Mol.Phylogenet. Evol. 55, 871–882.
Zhang, P., Wake, D.B., 2009a. Higher-level salamander relationships and divergencedates inferred from complete mitochondrial genomes. Mol. Phylogenet. Evol.53, 492–508.
Zhang, P., Wake, M.H., 2009b. A mitogenomic perspective on the phylogeny andbiogeography of living caecilians (Amphibia: Gymnophiona). Mol. Phylogenet.Evol. 53, 479–491.
Zhang, P., Zhou, H., Chen, Y.Q., Liu, Y.F., Qu, L.H., 2005. Mitogenomic perspectives onthe origin and phylogeny of living amphibians. Syst. Biol. 54, 391–400.
R. Alexander Pyron, J.J. Wiens / Molecular Phylogenetics and Evolution 61 (2011) 543–583 583