avaliaÇÃo da estrutura da comunidade de …
TRANSCRIPT

Fernando de Freitas
AVALIAÇÃO DA ESTRUTURA DA COMUNIDADE DE BACTERIOPLÂNCTON NA
REGIÃO DA RESERVA BIOLÓGICA MARINHA DO ARVOREDO E ENTORNO
Trabalho de Conclusão do Curso de
Graduação em Ciências Biológicas do Centro
de Ciências Biológicas da Universidade
Federal de Santa Catarina como requisito
para a obtenção do Título de Bacharel em
Ciências Biológicas.
Orientadora: Profa. Dra. Maria Luiza Schmitz
Fontes
Florianópolis
2019

2

3

4
Fernando de Freitas
Avaliação da estrutura da comunidade de bacterioplâncton na região da
Reserva Biológica Marinha do Arvoredo e entorno
Este Trabalho Conclusão de Curso foi julgado adequado para obtenção do Título de
Bacharel em Ciências Biológicas e aprovado em sua forma final pelo Curso de
Ciências Biológicas da Universidade Federal de Santa Catarina
Florianópolis, 05 de julho de 2019.
________________________
Prof. Carlos Roberto Zanetti
Coordenador do Curso
Banca Examinadora:
________________________
Prof.ª Dr.ª Maria Luiza Schmitz Fontes
Orientadora
________________________
Prof.ª Dr.ª Andrea Santarosa Freire
Universidade Federal de Santa Catarina
________________________
Prof.ª Dr.ª Melissa Carvalho
Universidade Federal de Santa Catarina

5
For those who wander.

6
AGRADECIMENTOS
Aos meus pais Olindina e João, por tudo nessa vida;
À minha irmã Rosana, por me motivar nessa corrida.
À grande Malu, por ter sido uma mãezona;
Aos meus amigos Rafa, Cadinho e Gui, pelos churras bacanas.
À UFSC, por ser minha segunda casa nesses anos;
À Maynooth University, pelo período insano.
Aos meus amigos do Vadio’s TeAm (Mineiro, Patola, Boca, Luizão,
Paquiderme e Apito) por todos os momentos compartilhados durante a
graduação;
Ao Otávio, Carlos e Parrudo, amigos que me incentivaram perto do final,
juntamente com o Lucão.
Aos veteranos Renan Pacheco e Gabi Alanís, pelas dicas de como seriam as
provas e trabalhos;
Ao grande Chico, que sempre mandava todo mundo pra Casa do Car****.
Aos amigos de longa data Cilhex, Serrano, Pedro e Jão;
Ao inefável professor Zanetti, que sempre me estendeu a mão.
Aos professores Elisandro, Selvino e Paulinho por serem mais zoeiros;
Ao professor Falkenberg, que não era zombeteiro.
Ao professor Renato, sempre tão pertinente;

7
Ao professor Kay pelas piadas indecentes.
Ao professor James McInerney, por me apresentar a Bioinformática;
Ao professor Luciano Alejandro, por me ajudar na matemática.
À professora Carla, Letícia e ao Mateus, pelos momentos que a monitoria de
Biofísica me deu;
Ao Alex, que nos dados estatísticos mexeu (e, obviamente, ao Projeto MAArE,
que os dados para análise forneceu);
À turma lipídio (11.2), que sempre manteve sua personalidade;
A todos os demais colegas, amigos, professores e familiares, pela
cumplicidade.
Também ao Iron Maiden, Mumford & Sons, Of Monsters and Men e Kiko
Loureiro;
Pois suas músicas foram essenciais para completar este TCC ligeiro.
Ao Douglas Adams, pelas histórias fantásticas;
E também, é claro, pelo Guia do Mochileiro das Galáxias.
Ao StoneD YoDa por me fazer rir quando precisava;
À Maghla e à Fleeur, também pelas risadas.
À CAPEs, pelo Ciência Sem Fronteiras. Lá, conheci estas pessoas de grande
coração: Ao Tony, Emily, Rafa, Gaudério, Mary, Renato, Manicaca, Sean e
Tiagão.

8

9
‘Would you tell me, please, which way I ought to go from here?'
'That depends a good deal on where you want to get to,' said the Cat.
'I don't much care where’ - said Alice.
'Then it doesn't matter which way you go,' said the Cat.
'- so long as I get SOMEWHERE,' Alice added as an explanation.
'Oh, you're sure to do that,' said the Cat, 'if you only walk long enough.’
― Lewis Carroll, Alice in Wonderland.

10
LISTA DE FIGURAS
Figura 1………………………………………………………………………………………21
Figura 2………………………………………………………………………………………23
Figura 3………………………………………………………………………………………24
Figura 4………………………………………………………………………………………28
Figura 5………………………………………………………………………………………29
Figura 6………………………………………………………………………………………31
Figura 7..…………………………………………………………………………………….32
Figura 8..…………………………………………………………………………………….33
Figura 9..…………………………………………………………………………………….36
Apêndice 8.1..………………………………...…………………………………………….48
Apêndice 8.2.……….………………………...…………………………………………….49

11
LISTA DE TABELAS
Tabela 1………………………………………………………………….………………..25
Tabela 2…………………………………………………………………….……………..26
Tabela 3…………………………………………………………………….…………..…34
Tabela 4…………………………………………………………………….…………..…34
Tabela 5…………………………………………………………………….…………..…35
Tabela 6…………………………………………………………………….…………..…35
Tabela 7 …………………………………………………………………….………….…36

12
LISTA DE ABREVIATURAS E SIGLAS
AEA Agência Europeia do Meio Ambiente
ANOVA Análise de variância
APM-MOD Matéria orgânica dissolvida de alto peso molecular
ACAS Água Central do Atlântico Sul ou SACW South Atlantic Central Water
BIOM Biological Observation Matrix
BMP Brazilian Microbiome Project
BPM- MOD Matéria Orgânica dissolvida de baixo peso molecular
CB Corrente do Brasil
FAPEU Fundação de Amparo à Pesquisa e Extensão Universitária
GOM Grupo de Oceanografia Microbiana
ICMBio Instituto Chico Mendes de Conservação da Biodiversidade
MAArE Projeto de Monitoramento Ambiental da Reserva Biológica Marinha do
Arvoredo e Entorno
NASA National Aeronautics and Space Administration
MOD Matéria orgânica dissolvida
OD Oxigênio dissolvido
OTU Operational Taxonomic Unit
PEAR Paired-End reAd mergeR
PPW Plumas de Água da Plata
PRIMER Plymouth Routines In Multivariate Ecological Research
QIIME (pronuncia-se ‘Tchaime’) Quantitative Insights Into Microbial Ecology
REBIO Arvoredo Reserva Biológica Marinha do Arvoredo
Tukey's HSD Honestly significant difference
UC Unidade de Conservação
USEARCH - Ultra-fast sequence analysis
ZC Zona Costeira

13
SUMÁRIO
RESUMO ……………………………………………………..……………….……14
ABSTRACT…………………………………………………..………………….….15
INTRODUÇÃO…………………………………………………..……………….....16
OBJETIVOS…………………………………………………..………………….…20
Principal…………………………………………………..…………….……..20
Específicos…………………………………………………..………………..20
MATERIAL E MÉTODOS………………………………………..………..……...20
Área de estudo, amostragem e extração de DNA………………………..20
Sequenciamento do Gene 16s rRNA e análise de dados……………… 22
RESULTADOS & DISCUSSÃO…………………………………………………..24
Resultado do Workflow de bioinformática…………………………………24
Metadados das amostras…………………………………………….……..25
Distribuição Bacteriana (Diversidade Taxonômica)…………………...…27
Verão…………………………………………………..……………….28
Inverno…………………………………………………..………..……31
Análises estatísticas………….…………..…………………………………33
Análises estatísticas de Filo………………………….…………...…33
Análises estatísticas de Família…………………….…………….…35
Distribuição espacial……………………………………………….…36
CONCLUSÕES………………………………………………...……………......…40
REFERÊNCIAS…………………………………………………..……….….….....42
APÊNDICE…………………………………………………..……….….…........…48
Principais Táxons.......................…………………………………………..48
Análises Estatísticas………………………………………………………..49

14
RESUMO
Nas últimas décadas, o estudo de microrganismos se intensificou devido a sua
importância nos processos ecológicos que afetam direta ou indiretamente a vida no
planeta Terra. O bacterioplâncton, um conjunto de microorganismos abundante na
coluna d’água, possui um papel fundamental na ciclagem de matéria orgânica dos
oceanos, influenciando mudanças climáticas e alterando a distribuição da biomassa
em redes tróficas. Mesmo com este papel imprescindível, o conhecimento e
entendimento destes seres em ambientes marinhos são escassos, especialmente
em território brasileiro. Dado o exposto, o presente estudo teve como objetivo
principal caracterizar a comunidade do bacterioplâcton na região da REBIO Arvoredo
e entorno através do uso de um Workflow com ferramentas de bioinformática. Os
objetivos específicos foram: 1. verificar se há diferença na estrutura da comunidade
do bacterioplâncton entre verão e inverno; 2. verificar se o filo Cyanobacteria
predomina no verão. Os filos Proteobacteria, Cyanobacteria e Bacteroidetes
contribuíram com um total de 84,6% e 87% dos filos encontrados nas amostras de
verão e inverno, respectivamente. A estrutura da comunidade do bacterioplâncton
mostrou-se diferente entre verão e inverno. No geral, Cyanobacteria foi o segundo
filo mais abundante tanto no período de verão quanto no inverno. No entanto, ao
contrário do esperado, Cyanobacterias foram os microrganismos mais abundantes
na superfície das estacões 16 e 14 (estacões mais offshore e dentro da REBIO
Arvoredo, respectivamente), atingindo 66% e 48% do bacterioplâncton total. Por
outro lado, no verão, a sua contribuição chegou no máximo a 36% da comunidade
total. Cyanobacteria, no entanto, predominou no inverno do que no verão e em
profundidades acima de 5 metros, principalmente próximo à costa.
Palavras-chave: Bacterioplâncton; comunidade; zonas costeiras; Proteobacteria;
Cyanobacteria; REBIO Arvoredo; MAArE.

15
ABSTRACT
In the last decades, the study of microorganisms has been intensified due to its
importance in ecological processes that directly or indirectly affect life on planet
Earth. Bacterioplankton, a group of microorganisms abundant in the water column,
plays a fundamental role in the cycling of organic matter in the oceans, influencing
climate changes and altering the distribution of biomass in food webs. Even with this
essential role, the knowledge and understanding of these organisms in marine
environments are scarce, especially in Brazilian coast. Given the above, the main
objective of this study is to characterize the bacterioplankton community in the region
of REBIO Arvoredo and surroundings through the use of a Workflow with
bioinformatics tools. The specific objectives are: 1. to verify if there is difference in the
community structure of bacterioplankton between summer and winter; 2. verifiy if
Cyanobacteria predominates in the summer. Proteobacteria, Cyanobacteria and
Bacteroidetes represented 84.6% and 87% of total filos in summer and winter,
respectively. The bacterioplankton structure differed between summer and winter
compostion. Cyanobacteria was the second most dominant phylum in both summer
and winter. Therefore, Cyanobacteria were the most abundant microorganisms in the
surface waters of stations 16 and 14 (offshore stations and inside the REBIO
Arvoredo, respectively), reaching up to 66 and 48% of total bacterioplankton. On the
other hand, their maixumum contribution in summer got up to 36%. Cyanobacteria
predominanted in the top 5 meters near the coast, in winter and not in summer, as
hypothesised.
Keywords: Bacterioplankton; community; coastal areas; Proteobacteria;
Cyanobacteria; REBIO Arvoredo; MAArE.

16
1. INTRODUÇÃO
Nas últimas décadas, a busca pelo conhecimento sobre as relações
ecológicas entre o meio ambiente e microrganismos de ambientes aquáticos se
intensificou, impulsionada, entre outros motivos, pela necessidade de compreender
o papel destes organismos nos oceanos (AZAM et al., 1994). Abordagens
promissoras que visam compreender estas relações ecológicas são amplamente
estudadas ao redor do mundo, buscando elucidar a composição, estrutura e
estabilidade da comunidade destes seres, assim como sua função, conexão e
participação nos ecossistemas marinhos (KIRCHMAN et al., 2008). Um dos
conjuntos de organismos aquáticos e microscópicos mais abundantes que vivem
dispersos na coluna d’água são chamados de picoplâncton, organismos entre 0.2 e
2 µm de tamanho (SIEBURTH & SMETACEK, 1978). O bacterioplâncton
componente principal do picoplâncton, pertencentes tanto aos domínios Bacteria e
Archea, possui um importante papel nos ciclos biogeoquímicos de elementos
essenciais, como carbono e nitrogênio (AZAM et al., 1994). Sua importância nas
teias alimentares favoreceu a formulação da alça microbiana (POMEROY, 1974;
AZAM et al., 1983) que, basicamente, estabelece a reciclagem da matéria orgânica
dissolvida (MOD). Logo, estes microrganismos são essenciais para o funcionamento
de sistemas ecológicos aquáticos, pois reciclam a MOD no sistema e assim
fornecem a energia necessária para manutenção e estabilidade de níveis tróficos
superiores (AZAM et al., 1983). Com um tamanho médio entre 0,2 e 2,0 µm, sua
distribuição vai desde águas acumuladas em Bromélias até nas profundezas dos
oceanos (ESTEVES, 2011).
A zona de transição entre componentes puramente terrestres e puramente
marinhos na superfície da Terra é chamada de Zona Costeira (ZC). A riqueza e
diversidade de recursos encontrados nestas zonas atraíram assentamentos
populacionais em todo o mundo (CROSSLAND et al., 2005). De acordo com a
National Aeronautics and Space Administration (NASA), há aproximadamente
620.000 quilômetros de costa em todo o planeta Terra e mais de um terço da
população humana (quase 2,4 bilhões de pessoas) vive numa média de 100
quilômetros de uma zona costeira. A linha costeira da União Europeia, por exemplo,
estende-se por 68.000 quilômetros (AEA – Agência Europeia do Meio Ambiente). Já
o litoral brasileiro possui 7367 quilômetros de extensão (VITTE, 2003). Se incluirmos

17
suas linhas de costa adjacentes (como baías e reentrâncias), esse número pode
ultrapassar 8500 quilômetros. Sua divisão clássica e mais bem aceita é a de Silveira
(1964), que subdivide a costa brasileira em cinco grandes compartimentos: Litoral
Norte, Litoral Nordeste (ou de Barreiras), Litoral Oriental, Litoral Sudeste (ou de
Escarpas Cristalinas) e Litoral Sul ou Subtropical (TESSLER, 2005). Cada região
possui suas características e peculiaridades, abrigando um complexo ecossistema
de relevância ambiental. Por se diferenciarem consideravelmente em seus aspectos
físicos e químicos, alterações na quantidade e diversidade das populações de
microrganismos podem ocorrer nestas áreas (AZAM & FALFATTI, 2007).
Consequentemente, informações sobre microrganismos presentes no ecossistema
costeiro são essenciais para preservar o equilíbrio ambiental nestas regiões de
constante atividade humana e ecológica (GREGORACCI et al., 2012; GONÇALVES,
2009).
A diversidade do bacterioplâncton marinho em ambientes costeiros ao redor
do globo terrestre apresenta padrões similares onde predominam os grupos
taxonômicos Alpha-Proteobacteria, Gamma-Proteobacteria (ambos membros do filo
Proteobacteria) e Bacteroidetes. Estes grupos, somados, representam
aproximadamente 60% da comunidade bacterioplanctônica nos litorais leste da
Austrália, leste da Ásia, sul da África, sul do Atlantico, nordeste, sudeste e oeste da ŕ
América do Norte, oeste da América do sul e Hawaii (POMMIER et al., 2007). Estes
organismos, como citado anteriormente, possuem entre suas funções a capacidade
de reciclar a MOD, tornando-a acessível para diferentes níveis tróficos (ELIFANTZ
et al., 2007). As Alpha-Proteobacteria, por exemplo, contribuem mais eficientemente
na absorção da MOD de baixo peso molecular (BPM- MOD) como glicose e N-
acetilglucosamina (ELIFANTZ et al, 2005, 2007; COTTRELL & KIRCHMAN 2000).
A diversidade do bacterioplâncton marinho em ambientes costeiros brasileiros,
em comparação com trabalhos de ambientes costeiros ao redor do globo, é pouco
conhecida e estudada (GONÇALVES, 2009), grande parte devido a conflitos (ou até
mesmo falta) de interesse (muitas vezes político-econômico) relacionado às
pesquisas de microrganismos aquáticos (GONÇALVES, 2009). Os estudos
concentram-se em regiões de água-doce (como rios e lagos), ou regiões de grande
interesse/movimento humano - como a Baía de Guanabara no Rio de Janeiro
(GONÇALVES, 2009). Esta última possivelmente engloba grande parte dos estudos
sobre bacterioplâncton no Brasil porque é a baía costeira mais proeminente do país,

18
resultado de intensa atividade humana - econômica e comercial (GREGORACCI et
al., 2012; GONÇALVES, 2009; KJERFVE et al., 1997). A maioria dos estudos
realizados na última década nas regiões costeiras Sudeste e Sul do Brasil mostram
a influencia da Água Central do Atlântico Sul (ACAS), trazida pela Corrente do Brasil
(DA SILVEIRA et al., 2000) no verão (CURY et al., 2011; FONTES et al., 2018;
GREGORACCI et al., 2012, SILVEIRA et al., 2011; THOMPSON et al., 2010), e no
Sul, intrusão da massa de água oriunda da pluma do Rio da Plata (FONTES et al.
2018). Gregoracci et al. (2012), Thompson et al. (2010, 2011) e Miloslavich et al.
(2011) observaram em seus respectivos trabalhos que os filos Proteobacteria e
Bacteroidetes, no litoral brasileiro, representam mais da metade dos microrganismos
encontrados na ZC. Seus estudos taxonômicos e de revisão, concentrados na região
da Baía de Guanabara (lat. 22o 509 ́ S e long. 43o 109 ́ W) são alguns dos poucos
estudos sobre a estrutura da comunidade do bacterioplâncton marinho (Miloslavich
et al. 2011 apenas menciona brevemente microrganismos) realizados em território
brasileiro. Já para a região Sul do Brasil, Fontes et al. (2018) encontraram uma
possível diferença entre a abundância e biomassa de bacterioplâncton na região
entre as latitudes (aproximadamente) 26 e 29o S. No entanto, a estrutura da
comunidade não foi descrita naquele momento. Com a finalidade de compreender
melhor a distribuição do bacterioplâncton na região Sul brasileira, este estudo foi
realizado na Reserva Biológica Marinha do Arvoredo.
A Reserva Biológica Marinha do Arvoredo (REBIO Arvoredo) é uma Unidade
de Conservação (UC) federal administrada pelo Instituto Chico Mendes de
Conservação da Biodiversidade (ICMBio) e localiza-se a cinco quilômetros da costa
brasileira, mais precisamente entre os municípios de Florianópolis e Bombinhas no
estado de Santa Catarina, abrangendo uma área de 17.600 ha onde 98% está sob
as águas (MAArE, 2017). Com a necessidade de um programa de monitoramento
ambiental marinho na Região da REBIO Arvoredo (ICMBio), surgiu o Projeto de
Monitoramento Ambiental da Reserva Biológica Marinha do Arvoredo e Entorno
(MAArE), uma parceria realizada entre a Universidade Federal de Santa Catarina
(UFSC), a Fundação de Amparo à Pesquisa e Extensão Universitária (FAPEU) e a
Petrobrás, com duração prevista de 5 anos (2013- 2017). O MAArE foi então
composto de duas grandes áreas para monitorar animais, vegetais, processos
oceanográficos e ecossistemas da Reserva: Indicadores Biológicos e Parâmetros
Oceanográficos (MAArE, 2017). Com o objetivo primordial de levantar informações

19
necessárias para subsidiar a construção de um monitoramento básico dos costões e
águas da Reserva, o projeto MAArE, entre outros dados, coletou informações que
podem elucidar fatores que mais estejam influenciando a estrutura da comunidade
do bacterioplâncton na região da REBIO Arvoredo e entorno (MAArE, 2017).
As Cianobactérias, além de capazes de realizar fotossíntese, também são
componentes importantes na transformação da MOD nos oceanos (KIRCHMAN et
al., 2008). Em território brasileiro, há poucos estudos que elucidam a abundância e
importância destes seres em ambientes marinhos (GONÇALVES, 2009). Apesar de
estudos mais recentes (como os de FONTES et al. - 2018 e BRANDINI et al. - 2018)
demonstrarem como fatores externos (como intrusões de correntes marinhas) são
determinantes para a presença destes microrganismos na costa brasileira, ainda há
um longo caminho a ser percorrido em direção ao entendimento do papel destes
seres nos processos ecológicos oceânicos, especialmente em diferentes períodos
sazonais.
A utilização de programas/protocolos de bioinformática por pesquisadores das
Áreas de Ciências Biológicas tornou-se fundamental devido a complexidade e
amplitude dos dados utilizados em estudos ecológicos (JONES et al., 2006).
Cientistas que buscam uma maneira mais robusta de analisar seus dados
frequentemente aprendem a usar programas ou linguagens de programação como
“R Statistical Package” (http://www.r-project.org) ou Python (JONES et al., 2006).
Contudo, muitos pesquisadores procuram utilizar metodologias de bioinformática
prontas, não adaptando o método de acordo com suas necessidades devido a falta
de conhecimento em informática/linguagens de programação (JONES et al., 2006),
perdendo assim a possibilidade de atingir melhores resultados em suas pesquisas.
Dado o exposto, o presente estudo visa como objetivo principal caracterizar a
comunidade do bacterioplâcton na região da REBIO Arvoredo e entorno; utilizando
um Workflow com ferramentas de bioinformática formulado e otimizado para a
análise dos dados coletados; corroborar com estudos já realizados no Brasil e no
mundo e elucidar a estrutura da comunidade do bacterioplâncton na área estudada.

20
2. OBJETIVOS
2.1 Principal
Caracterizar a comunidade do bacterioplâcton na região da REBIO Arvoredo e
entorno.
2.2 Específicos
- Aprimorar/otimizar o Workflow de bioinformática criado e utilizado para obtenção
dos dados analisados no presente estudo;
- Verificar se há diferença na estrutura da comunidade do bacterioplâncton entre
verão e inverno (2015/2016);
- Verificar se Cianobactérias predominam no verão;
- Verificar se existe diferença espacial na estrutura da comunidade do
bacterioplâncton (em profundidade).
3. MATERIAIS E MÉTODOS
3.1 Área de estudo, amostragem e extração de DNA
Um total de 15 amostras foram coletadas pela embarcação oficial do projeto
MAArE nos períodos de verão (9 amostras) e inverno (6 amostras) do ano de 2016
(inverno – dias 3, 4 e 5 de agosto; verão – dias 16, 17 e 18 de fevereiro). No período
de verão foram coletadas amostras em três profundidades: superfície, meio e fundo
nas estações oceanográficas 10, 14 e 16, enquanto no período de inverno, apenas
amostras de superfície e fundo foram coletadas - também nas estações
oceanográficas 10, 14 e 16 (Figura 1). As amostras da estação 10 foram coletadas

21
nas profundidades de 0, 2 e 4 metros (sup, mid e fun, respectivamente); onde a
profundidade máxima do local era de 7m. As amostras da estação 14 foram
coletadas nas profundidades de 0, 10 e 15 metros (sup, mid e fun, respectivamente),
com profundidade máxima do local =17m. As amostras da estação 16 foram
coletadas nas profundidades de 0, 15 e 45 metros (sup, mid e fun, respectivamente),
com profundidade máxima do local = 53m). Os dados abióticos e bióticos coletados
foram disponibilizados pela pesquisadora Dra. Andrea Santarosa Freire e analisados
pelo Grupo de Oceanografia Microbiana da UFSC durante a realização do projeto de
pesquisa BioEnergia Lagoa. As amostras de água foram coletadas em garrafas van
dorn para análise de nutrientes, filtradas em laboratório com filtros de vidro 0.45 µm
e congeladas a -20 °C. As amostras para extração de DNA do bacterioplancton
foram mantidas refrigeradas até laboratório onde 300-500 mL foram filtradas em
filtros de membrana de porosidade de 0.2 µL, em duplicata, através do PowerSoil®
DNA Isolation Kit (MO BIO Laboratories, Inc., USA), com a adição prévia de um
banho maria em 65 °C; O DNA foi então quantificado por fluorometria (Thermo-
Fisher Qubit® 2.0).
Figura 1: Localização de amostragem. Distribuição das estações oceanográficas do Projeto MAArE©.
A região demarcada em amarelo é referente a REBIO Arvoredo. As amostras analisadas no presente
estudo foram coletadas nas estações 10 (próximo a costa catarinense), 14 (propriamente inserida
dentro da REBIO) e 16 (ponto mais afastado do litoral). As profundidades aproximadas, da costa para
o mar aberto, são respectivamente: entre 7 e 10m, entre 10 e 20m, entre 20 e 50m e maior ou igual
50m. Estações Simples = coletados somente dados físicos; Estações Completas = além dos dados

22
físicos, foram coletadas amostras de amostragem de água, plâncton e sedimento. Fonte: MAArE :
Monitoramento ambiental da Reserva Biológica Marinha do Arvoredo e entorno / Organização:
Bárbara Segal...[et al.]. – 1. ed. - Florianópolis : UFSC/MAArE, 2017. 268 p. : il., gráf., tab. Imagem
retirada da página 178.
3.2 Sequenciamento do Gene 16s rRNA e análise de dados
O sequenciamento foi realizado pela empresa Neoprospecta® através do
sequenciador de nova geração Illumina Miseq. A região de DNA amplificada foi um
fragmento multivariável V3-V4 do gene procariótico 16s rRNA, utilizando um
conjunto de primers U341F e 806R, os quais abrangem muito bem os grupos
Archaea e Bacteria com amplicons de aproximadamente 400bp (TAKAHASHI et al.,
2014). Após o recebimento dos dados brutos de sequenciamento, os arquivos de
leitura foward e reverse foram unidos através do software PEAR (Paired-End reAd
mergeR) versão 0.9.10 (ZHANG et al., 2014). O Workflow utilizado foi uma versão
do Brazilian Microbiome Project (BMP) descrita por PYLRO et al., 2016, modificada
pelo Grupo de Oceanografia Microbiana (GOM – UFSC). O principal software
utilizado para análise foi o QIIME (Quantitative Insights Into Microbial Ecology)
versão 1.9.1 (CAPORASO et al., 2010) seguido do USEARCH7 - Ultra-fast
sequence analysis (EDGAR, 2013). Todos os softwares foram instalados
nativamente e executados através da linha de comando (terminal) no Sistema
Operacional macOS El Captain (10.11.1 – 10.11.6). O Workflow completo pode ser
visualizado no repositório GitHub através do link
“https://github.com/vinisalazar/BioEnergia-Lagoa”. O grau de similaridade das OTUs
(Operational Taxonomic Unit) foi de 97%, as quais foram classificadas
taxonomicamente através da base de dados SILVA (High Quality Ribosomal RNA
Databases) 119 (QUAST et al., 2013). Ao final dos processos do Workflow criado,
foram geradas matrizes no formato .BIOM (do inglês Biological Observation Matrix,
um formato reconhecido como padrão pelos projetos Earth Microbiome Project e
Genomics Standard Consortium). As tabelas .BIOM foram então convertidas para
formato padrão de texto (.txt) e novamente convertidas para .xls ou .ods para
visualização em programas de edição de planilhas como Microsoft Excel ou
LibreOffice Calc. As análises estatísticas das tabelas dos quinze Filos e quinze

23
Famílias dominantes entre verão-inverno e superfície-fundo foram realizadas através
do Software TIBCO Statistica™ versão 13.3. Para maior informação, ver Apêndice
8.1 onde os filos e famílias encontram-se organizados em uma tabela. Os testes
aplicados foram: histograma com teste de Shapiro-Wilk (para verificação da
distribuição normal); análise de variância (ANOVA) para Filos e Famílias X
profundidade e estação; Kruskal-Wallis (equivalente ao ANOVA para dados não
paramétricos – fora da distribuição normal de Shapiro-Wilk); Levene´s (para
confirmar a realização de ANOVAs para todas as amostras) e Tukey HSD (para
verificar possíveis diferenças entre as amostras). A classificação multivariada
(espacial) foi realizada através do Software PRIMER (Plymouth Routines In
Multivariate Ecological Research) versão 6. Nas Figuras 2 e 3 podemos observar
esquemas dos processos de análise de dados para o Workflow e para as análises
estatísticas.
Figura 2: esquema do Workflow de Bioinformática. O QIIME (lê-se ‘tchaimê’) é o programa base no
qual todos os outros processos são inseridos. Pode-se fazer uma analogia ao programa R e suas
bibliotecas. Neste caso, o QIIME seria o terminal base e os programas/scripts inseridos seriam suas
bibliotecas. Ao contrário do R, que possui linguagem própria, o QIIME trabalha com a linguagem de
programação Python (no presente trabalho foi utilizado a versão 2.7).

24
Figura 3: Esquema do processo de análise de dados estatísticos. Primeiramente, para verificar se
as amostras apresentavam uma distribuição normal (necessária para a análise de variância –
ANOVA), aplicou-se o teste de Shapiro-Wilk. Nas amostras que apresentaram distribuição normal,
aplicou-se o teste ANOVA. Os dados que não apresentaram distribuição normal foram submetidos
ao teste de Kruskal-Wallis (equivalente ao teste de variância para dados não paramétricos); para
verificar a possibilidade de aplicação do teste ANOVA nas amostras não paramétricas, os testes de
Levene e Tukey HSD (honestly significant difference) foram realizados.
4. RESULTADOS E DISCUSSÃO
4.1 Resultado do Workflow de bioinformática
O Workflow de bioinformática resultou em 10 arquivos .BIOM (5 para cada
período - inverno e verão), os quais foram convertidos para .ods e visualizados no
software LibreOffice Calc. Os arquivos representam, respectivamente, a
porcentagem de Filo, Classe, Ordem, Família e Gênero encontradas em cada
período.
4.2 Metadados das amostras de verão e inverno
Os metadados das amostras de verão e inverno podem ser observados nas
Tabelas 1 e 2, respectivamente.

25
Tabela 1: Metadados das amostras de verão.
Estação Oceanográfica
Ano Mês Dia Hora:minuto Latitude Longitude Profundidade
Local (m) Profundidade de coleta (m)
Zona Temperatura Salinidade
10 2016 2 18 10:45:00 -27.2572 -48.5795 7.0 0.0 0 26.8946 33.7500
10 2016 2 18 10:46:00 -27.2572 -48.5795 7.0 2.0 1 26.7939 33.7661
10 2016 2 18 10:50:00 -27.2572 -48.5795 7.0 4.0 2 26.7201 33.8105
14 2016 2 17 14:35:00 -27.2797 -48.4229 17.0 0.0 0 25.9175 33.4115
14 2016 2 17 14:50:00 -27.2797 -48.4229 17.0 10.0 1 21.4450 35.1799
14 2016 2 17 14:55:00 -27.2797 -48.4229 17.0 15.0 2 19.5359 37.4484
16 2016 2 16 13:00:00 -27.2798 -48.2838 53.0 0.0 0 27.4038 34.6284
16 2016 2 16 13:16:00 -27.2798 -48.2838 53.0 15.0 1 25.4714 35.4441
16 2016 2 16 13:20:00 -27.2798 -48.2838 53.0 45.0 2 17.2946 35.5927
Oxigênio Dissolvido
(ml/L) DO (%)
AUO (%)
Fosfato (µM)
Silicato (µM)
Amônio (µM)
Nitrito+Nitrato (µM)
NP MST (mg/L) MSI
(mg/L) MSO (mg/L)
Cla SCOR/UNESCO
1966
4.4276 69.7420 30.2580 0.32 7.80 5.71 1.25 21.90 17.2857 13.4286 3.8571 1.91040
3.2839 82.1067 17.8933 0.38 1.78 5.02 0.90 15.42 9.5833 7.1667 2.4167 1.79297
3.8495 90.2753 9.7247 0.27 1.14 3.94 1.17 19.15 8.6154 6.3077 2.3077 2.12148
3.3271 54.5410 45.4590 0.27 2.02 2.20 0.83 11.20 6.5500 4.9500 1.6000 2.48746
2.5890 44.4050 55.5950 0.57 5.75 0.83 2.78 6.33 17.4000 14.4000 3.0000 7.06450
3.7255 77.3270 22.6730 0.96 14.72 13.50 0.49 14.58 9.4000 7.4667 1.9333 13.98269
4.8154 95.4950 4.5050 0.30 1.21 1.50 1.11 8.83 4.2333 3.4000 0.8333 1.69603
4.4097 99.4530 0.5470 0.92 0.57 1.62 0.46 2.28 9.4333 7.9667 1.4667 2.11896
2.6637 49.3641 50.6359 0.31 11.58 0.93 5.04 19.57 5.8800 4.6400 1.2400 6.85008
chla satélite (mg/L)
Fluorescência (CTD)
Secchi (m)
ZEU
coeficiente de
atenuação da luz (m^-
1)
coeficiente de
atenuação da luz -
satélite(m^-1)
Biovol Zooplancton
(mL/m³)
Biomassa úmida
Zooplancton (mg/m³)
Biomassa Seca
Zooplancton (mg/m³)
6.42000 1.87350 1.5 4.05 1.133 0.421 10.02 1854.49 249.90
2.42420
3.00190
1.91970 0.35280 7.5 20.25 0.227 0.158 2.83 543.86 62.31
1.72370

26
3.75500
0.53902 0.28650 16.0 43.20 0.106 0.074 0.48 103.29 7.11
0.82225
1.69960
Tabela 2: Metadados das amostras de inverno.
Estação Oceanográfica
Ano Mês Dia Hora:
minuto Latitude Longitude
Profundidade Local (m)
Profundidade de
coleta (m) Zona Temperatura Salinidade
10 2016 8 5 10:57:00 -27.2572 -48.5795 6.4 0.0 0 17.4298 31.8857
10 2016 8 5 10:57:00 -27.2572 -48.5795 6.4 4.0 2 17.4403 31.9842
14 2016 8 4 10:20:00 -27.2797 -48.4229 17.0 0.0 0 17.2771 32.4213
14 2016 8 4 10:20:00 -27.2797 -48.4229 17.0 14.0 2 17.2890 32.5180
16 2016 8 3 11:47:00 -27.2798 -48.2838 52.0 0.0 0 17.2986 32.6788
16 2016 8 3 11:47:00 -27.2798 -48.2838 52.0 48.0 2 17.7765 33.3583
Oxigênio Dissolvido
(ml/L) DO (%)
AUO (%)
Fosfato (µM)
Silicato (µM)
Amônio (µM)
Nitrito+Nitrato (µM)
NP MST
(mg/L) MSI
(mg/L) MSO (mg/L)
Cla SCOR/UNESCO
1966
4.4446 85.8439 14.156
1 0.37 14.56 1.88 1.66 9.51 27.7692 24.1538 3.6154 5.57020
4.5292 82.0543 17.945
7 0.33 10.77 0.90 0.90 5.47 17.3333 14.9333 2.4000 1.97179
4.8779 81.6509 18.349
1 0.37 9.57 1.10 1.32 6.60 28.4000 15.5333 12.8667 8.97955
4.6083 81.9670 18.033
0 0.35 8.93 1.44 1.91 9.54 19.2667 17.8000 1.4667 1.85328
5.0999 92.9339 7.0661 0.31 10.07 1.29 0.73 6.60 24.9000 21.2000 3.7000 8.61587
4.4295 80.2613 19.738
7 0.36 7.76 0.91 1.35 6.31 29.3000 26.6000 2.7000 1.25995
chla satélite (mg/L)
Fluorescência (CTD)
Secchi (m)
ZEU
coeficiente de
atenuação da luz (m^-
1)
coeficiente de
atenuação da luz -
satélite(m^-1)
Biovol Zooplancton
(mL/m³)
Biomassa úmida
Zooplancton (mg/m³)
Biomassa Seca
Zooplancton (mg/m³)
2.19380 1.8 4.73 0.971
0.24 114.54 12.38
1.70580
2.64000 3.3 8.78 0.523
0.14 92.89 5.66
0.83052
2.06960 3.2 8.64 0.531
0.12 105.19 4.76
1.11360

27
4.3 Distribuição Bacteriana (Diversidade Taxonômica)
O número total de OTUs definido pelo método aplicado no Workflow foi 739
(387 para verão e 352 para inverno). Das amostras totais, Archea representou
uma média de 2,95% (3,1%no verão e 2,8% no inverno), enquanto Bacteria
(maioria) representou uma média de 96,8% (96,6% no verão e 97% no inverno).
Apenas 0,25% das amostras totais (média) não foram identificados pelo método
empregado (0,3% no verão e 0,2% no inverno). Dos 96,8% de bactérias
identificadas, os 3 Filos predominantes foram, respectivamente, Proteobacteria
(média de 48% - 49,6 no verão e 46,4 no inverno), Cyanobacteria (média de 26,95%
- 23,2 no verão e 30,7 no inverno) e Bacteroidetes (média de 10,85% - 11,8 no
verão e 9,9 no inverno). Durante o período de verão, os valores máximos e mínimos
para os três Filos foram: Proteobacteria, verão, 62,6% máx (amostra de meio da
estação 14) e 32,2% mín (amostra de fundo da estação 14); inverno, 62% máx
(amostra de fundo da estação 10) e 23,1% mín (amostra de superfície da estação
16); Cyanobacteria, verão, 36,4% máx (amostra de fundo da estação 14) e 11,3%
mín (amostra de meio da estação 14); inverno, 65,7% máx (amostra de superfície da
estação 16) e 10,6% mín (amostra de fundo da estação 14); Bacteroidetes, verão,
18,2% máx (amostra de superfície da estação 10) e 3,1% mín (amostra de superfície
da estação 14); inverno, 13,7 máx (amostra de superfície da estação 10) e 4,8% mín
(amostra de superfície da estação 16).
As 6 maiores Classes encontradas no período de verão pertencem ao Reino
Bacteria. São, em ordem decrescente: Alpha-Proteobacteria (28,2%), Gamma-
Proteobacteria (17,9%), Chloroplast (14,2%), Flavobacteria (9,4%), Cyanobacteria
(9%) e Acidiomicrobia (8%). Dentre estas 6, duas pertencem ao Filo Proteobacteria
(Alpha & Gamma), duas pertencem ao Filo Cyanobacteria (Chloroplast &
Cyanobacteria), uma ao Filo Bacteroidetes (Flavobacteria) e uma ao Filo
Actinobacteria (Acidiomicrobia). Das 27 Classes restantes, nenhuma apresentou
percentagem maior que 3%; Quanto ao período de inverno, as 6 maiores Classes
foram, em ordem decrescente: Chloroplast (29,3%), Alpha-Proteobacteria (24,1%),
Gamma-Proteobacteria (17,7%), Flavobacteria (9,3%), Acidiomicrobia (6,1%) e
Delta-Proteobacteria (3,6%). Dentre estas 6, 3 pertencem ao Filo Proteobacteria
(Alpha, Delta e Gamma), uma ao Filo Cyanobacteria (Chloroplast), uma ao Filo

28
Bacteroidetes (Flavobacteria) e uma ao Filo Actinobacteria (Acidiomicrobia). Das 22
Classes restantes, nenhuma apresentou percentagem maior que 2,1%.
4.3.a Verão
Nas Figuras 4 e 5, podemos observar, respectivamente, a distribuição dos
Filos e das 15 Famílias predominantes (de um total de 117) para cada estação
oceanográfica.
Figura 4: Principais Filos de microrganismos encontrados na região da REBIO Arvoredo durante o
verão (estações oceanográficas 16, 14 e 10). Sup = Superfície, Mid = Meio e Fun = Fundo.
Proteobacteria foi o Filo predominante, com máximo de 62,6% do total do
bacterioplâncton presente na amostra de meio da estação 14. O mínimo de 32,2%
também foi observado na estação 14 (amostra de fundo). Em em média,
Proteobacteria representou 49,6% do total do bacterioplâncton geral no verão.
O segundo Filo mais abundante na região foi Cyanobacteria, apresentando
uma média de 23,2% do total do bacterioplâncton nesta época do ano, com um valor
máximo de 36,4% do total e mínimo de 11,3 % do total, novamente na mesma
estação 14; no entanto, o máximo foi encontrado na amostra de fundo (15 m) e o
mínimo na profundidade de 10 m (meio). Esses dois filos representaram um total de
72,8%, em media, do total de filos descritos. O terceiro Filo predominante foi
Bacteroidetes, apresentando uma média de 11,8% do total de bacterioplâncton,


30
estação 14 e mín de 6% na amostra de fundo da mesma estação), o cluster SAR86
(10%, com máximo de 11,5% na amostra de meio da estação 14 e mín de 3,4 na
superfície da mesma estação), Rhodospirillaceae (cerca de 6%, com pico de 8,6%
na superficie da estação 14 e mín de 2,9% na superfície da estação 10), cluster SAR
116 (cerca de 4%, com máx de 9,6% no fundo da estação 16 e mín de 1,4% nas
superfícies das estações 14 e 10), Alteromonadaceae (cerca de 4%, com máx de
9,3% na amostra de meio da estação 14 e mín de 1% na superfície da mesma
estação) e o cluster SAR 11 de superfície 1 (cerca de 2%, com máx de 3,3% na
amostra de superfície da estação 14 e mín de 1,7% na amostra de superfície da
estação 16). Somada, a Classe Alpha-Proteobacteria representa 30% das Famílias
predominantes (entre as 15 maiores de 117); enquanto Gamma-Proteobacteria soma
14%.
Quanto a Cyanobacteria (cerca de 26% da média total), as famílias com maior
percentagem foram identificadas como uncultured bacterium e Familyl. Uncultured
bacterium representou uma média de 13% (com máx de 33% na amostra de fundo
da estação 14 e mín de 5% na amostra de meio da estação 16) e Familyl cerca de
11% (com máx de 21% na amostra de fundo da estação 16 e mín de cerca de 2% na
amostra de fundo da estação 14). Na Figura 6 podemos observar os gêneros
predominantes identificados dentro de Familyl.
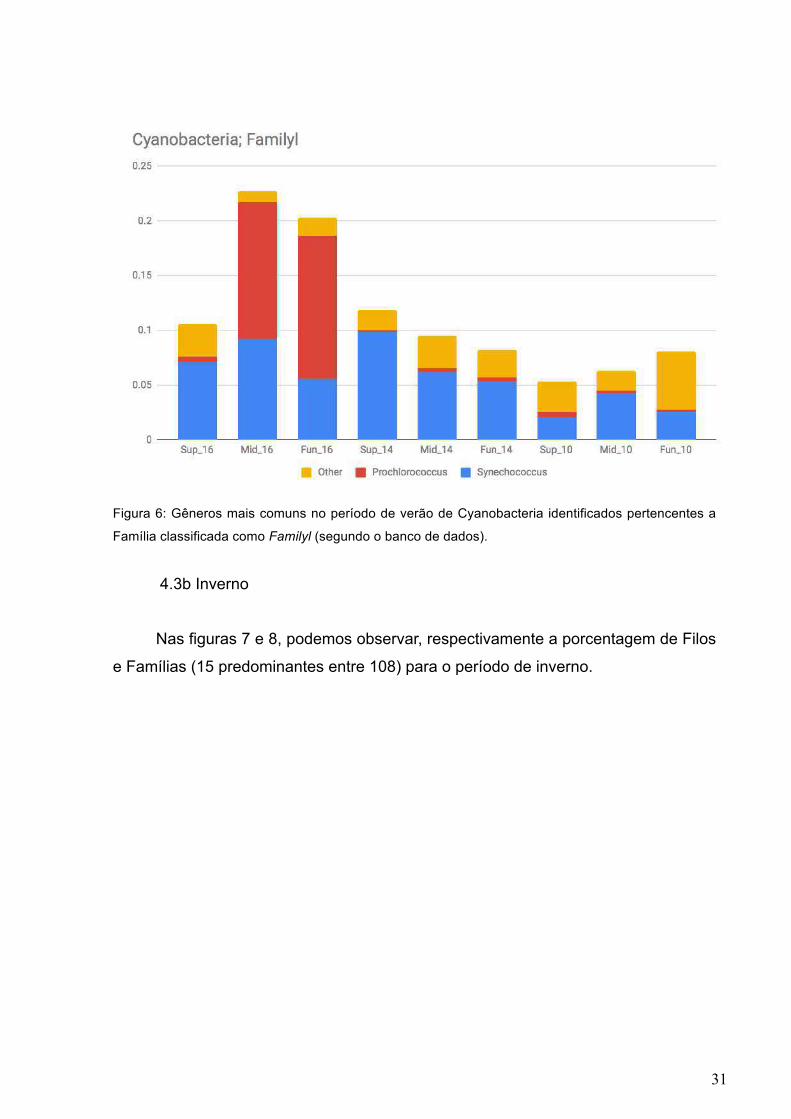
31
Figura 6: Gêneros mais comuns no período de verão de Cyanobacteria identificados pertencentes a
Família classificada como Familyl (segundo o banco de dados).
4.3b Inverno
Nas figuras 7 e 8, podemos observar, respectivamente a porcentagem de Filos
e Famílias (15 predominantes entre 108) para o período de inverno.

32
Figura 7: Principais Filos de microrganismos encontrados na REBIO Arvoredo durante amostragem
de inverno (estações oceanográficas 16, 14 e 10). Sup = Superfície e Fun = Fundo.
Proteobacteria foi novamente o Filo predominante no período de inverno
(46,4% do total), com máximo de 62% no fundo da estação 10 e mínimo de 23,1%
na superfície da estação 16. O segundo Filo mais predominante foi Cyanobacteria,
com média de 30,7% do total (máximo de 65,7% na superfície da estação 16 e
mínimo de 10,6% no fundo da estação 14). Os dois Filos representaram um total de
77,1% da comunidade total de bacterioplâncton, um pouco maior do que o verão. O
terceiro Filo mais abundante foi mais uma vez o Bacteroidetes, com média de 9,9%
do total dos filos presentes, máximo de 13,7% na superfície da estação 10 e mínimo
de 4,8% na superfície da estação 16. Actinobacteria ficou em quarto lugar com uma
média de 6,3% do total (apresentando um valor máximo de 9,6% no fundo da
estação 14 e mínimo de 3,4% no superfície da 16). Em média, Proteobacteria,
Cyanobacteria e Bacteroidetes somaram 87% dos Filos encontrados no período de
inverno; No inverno, os maiores valores de Cyanobacteria foram observados na
superfície da estação 10, enquanto que no verão foi no fundo da estação 14.
No período de inverno, como podemos observar na Figura 6, aquelas famílias
sem classificação segundo a base de dados utilizada dentre Cyanobacteria
mostraram-se predominantes (média de 33% do total, maximo de 71% na superfície
da estação 16 e mínimo de 11% no fundo da estação 14). A classificação máxima
encontrada na base de dados do Workflow foi Filo Cyanobacteria, Classe
Chloroplast.


34
Todos os gráficos de ANOVA e Kruskal-Wallis podem ser observados no
Apêndice.
Os valores do teste de Shapiro-Wilk para Filo (15 predominantes comuns entre
verão e inverno) podem ser observados na Tabela 3.
Tabela 3: Valores de Shapiro-Wilk para os 15 Filos predominantes em comum (verão e inverno). O
valor de p menor que 0,05 representa uma distribuição não normal.
Para os resultados com p abaixo de 0,05, foram aplicados o teste de Levene
(para profundidade e estação - Tabelas 4 e 5 respectivamente).
Tabela 4: Teste de Levene (verão-inverno) realizado nas amostras de Filo (15 predominantes em
comum entre verão e inverno) com valores de p de Shapiro-Wilk menor que 0,05. Os valores de p
abaixo de 0,05 do teste de Levene demonstram que as amostras apresentam variações em suas
populações.

35
Tabela 5: teste de Levene (profundidade) realizado nas amostras de Filo (15 predominantes em
comum entre verão e inverno) com valores de p de Shapiro-Wilk menor que 0,05.
Para Cyanobacteria (populações de verão e inverno), foi aplicado o teste de
Tukey HSD (honestly significant difference).
4.4b Análises estatísticas de Família
Todos os gráficos de ANOVA e Kruskal-Wallis podem ser observados no
Apêndice.
Os valores do teste de Shapiro-Wilk para Família (15 predominantes comuns
entre verão e inverno) podem ser observados na Tabela 6.
Tabela 6: Valores de Shapiro-Wilk para as 15 Famílias predominantes em comum (verão e inverno). O
valor de p menor que 0,05 representa uma distribuição não normal.

36
Para os resultados com p abaixo de 0,05, foram aplicados o teste de Levene
(para profundidade e estação - Tabela 7).
Tabela 7: Teste de Levene (verão-inverno) realizado nas amostras de Família (15 predominantes em
comum entre as amostras de verão e inverno) com valores de p de Shapiro-Wilk menor que 0,05. Os
valores de p abaixo de 0,05 do teste de Levene demonstram que as amostras apresentam variações
em suas populações.
4.4c Distribuição espacial
As distribuições espaciais de Filo e Família (predominantes entre as 15 comuns
entre verão e inverno) podem ser observadas na Figura 9.
Figura 9: Gráfico resultante da análise de MDS para Filos (a) e familias (b). As três primeiras letras
correspondem a profundidade (Sup = Superfície, Mid = Meio e Fun = Fundo); o número que segue
corresponde à estação oceanográfica na qual foram coletadas as amostras; e as letras v e i
correspondem aos períodos de verão e inverno. O grupo Sup16i, tanto para Filo quanto para Família,
encontra-se bastante isolado dos demais (grupo com alto índice de Cyanobacterias não
identificadas). Os grupos Sup10i, Sup10v e Fun10v (alto índice de Proteobacteria) também
encontram-se mais isolados, demonstrando possíveis padrões durante os períodos de verão e
inverno.

37
Ao observarmos os gráficos resultantes do Workflow de bioinformática,
podemos perceber claramente a predominância de três grandes Filos:
Proteobacteria e Cyanobacteria, seguidos de Bacteroidetes nos dois períodos
amostrais. Os dados de Proteobacteria e Bacteroidetes corroboram com pesquisas
realizadas tanto no Brasil (MENEZES et al., 2019; GREGORACCI et al., 2012;
ZINGER et al., 2011; SILVEIRA et al., 2011, THOMPSON et al., 2010) como no
mundo (POMMIER et al., 2007; SUNAGAWANA et al., 2019; FUHRMAN & STEELE,
2008; MILASLAVICH et al., 2011; THOMPSON et al., 2010), demonstrando que a
estrutura da comunidade do bacterioplâncton é similar nas zonas costeiras do
planeta Terra. O Filo Proteobacteria é um dos mais abundantes na superfície do
globo terrestre, apresentando diversos organismos fixadores de nitrogênio
(DELMONT et al., 2018) e especialistas na utilização da MOD de baixo peso
molecular (BPM-MOD). Juntamente com o Filo Bacteroidetes, que é comumente
considerado especializado na degradação da MOD de alto peso molecular – APM-
MOD (FERNÁNDEZ-GÓMEZ et al., 2013), estes grupos são indispensáveis para o
equilíbrio ecossistêmico dos oceanos (AZAM et al., 1983 & 1994).
No entanto, a grande predominância do Filo Cyanobacteria mostrou-se um
pouco incomum (comparado a outras regiões costeiras), pois apenas trabalhos
realizados nas regiões costeiras Sul-Americana demonstraram a altos índices destes
organismos (MILASLAVICH et al., 2011; THOMPSON et al., 2010; CODD et al.,
2005). Conforme observado nas Figuras 4 e 6, o Filo Cyanobacteria foi o segundo
mais dominante tanto para o perído de verão (23,2% do total) quanto para inverno
(30,7% do total). As Cianobactérias, além de capazes de realizar fotossíntese,
também são componentes importantes na transformação da MOD nos oceanos
(KIRCHMAN et al., 2008). Esses microrganismos podem formar densas populações
em períodos específicos do ano, acumulando-se na superfície das águas e
carreadas através de correntes marinhas e de fortes ventos, fato que pode elucidar a
percentual registrado no estudo. Logo, a influência por intrusão das Águas Centrais
do Atlântico Sul (ACAS) – SACW, do inglês, South Atlantic Central Water (DA
SILVEIRA et al., 2000; FONTES et al., 2018) pode ser um dos fatores
influenciadores da dominância deste Filo (FONTES et al., 2018). Por se tratar de
uma região relativamente rasa, a adição de nutrientes pela intrusão da ACAS deve
ter favorecido este grupo taxonômico (principalmente nas águas próximo ao fundo

38
da estação 14). Por outro lado, no período de inverno austral, a intrusão das Plumas
de Água da Plata (PPW – do inglês, Plata Plume Waters) sobre a ACAS pode
explicar a variação de dominância de microrganismos aquáticos (FONTES et al.,
2018), pois ecótipos de procariontes de água quente (como Cyanobacteria) são
dominantes em águas subtropicais (SEYMOUR et al., 2012), corroborando com as
temperaturas registradas nas regiões da REBIO Arvoredo e entorno (temperatura
média de 23 oC - MAArE, 2017) e também com o percentual de organismos
registrados no período de inverno austral.
O alto índice de Cyanobacteria presente nas amostras de superfície tanto no
verão quanto no inverno pode ser justificável pelo fato de que estes organismos
apresentam menor necessidade de nutrientes para manutenção celular se
comparado a outros organismos do fitoplâncton. Todos estes fatos podem explicar
os altos valores de Cyanobacteria encontrados durante o verão nas amostras de
fundo da estação 14, relacionado a intrusão da ACAS e durante o inverno nas
amostras de superfície da estação 16.
Entre as 15 Famílias predominantes para o período de verão, Cyanobacteria e
Proteobacteria juntas representaram, em media, 63% do total. Dentre as Famílias
identificadas, os maiores picos são de Cyanobacteria nas amostras de fundo da
estação 14 (33%) e de meio da estação 16 (26%), onde a Família classificada como
Familyl se destaca. Dentro desta, os gêneros de Synechococcus e Prochlorococcus
são os mais evidentes. Synechococcus é um importante picoplâncton fotossintético
nos oceanos temperado e tropical. Por possuir um mecanismo eficiente para se
adaptar às mudanças na salinidade e na intensidade da luz, estes microrganismos
são abundantes nos oceanos (KIM et al., 2018). Já Prochlorococcus é a
Cyanobacteria autotrófica de menor emissão de oxigênio conhecida (ROCAP et al.,
2003). Ela domina numericamente o fitoplâncton nos oceanos tropicais e
subtropicais e é responsável por uma fração significativa da fotossíntese global nos
oceanos (ROCAP et al., 2003).
Em média, no período de inverno, observou-se que cianobactérias não
cultivadas/desconhecidas representaram 26% das Famílias identificadas, apesar do
percentual individual de cada Família ser menor que Rhodobacteraceae (16%, a
qual foi a família predominante dentre o Filo Proteobacteria, Classe Alfa-
Proteobacteria e Ordem Rhodobacterales). Segundo Madsen (2011), aglomerados
de Cyanobacteria não cultivadas pode ser um indício de dependência nutricional de

39
outros organismos vizinhos e/ou baixos níveis de oxigênio, fato que pode elucidar o
alto percentual de cianobactérias identificadas no período.
Segundo os dados estatísticos (ANOVA, Kruskal-Wallis, Levene ́s e Tukey
HSD), a diferença entre verão e inverno para o filo Cyanobacteria foi significativa.
Mais uma vez, a intrusão da ACAS no período de verão e da PPW no período de
inverno podem explicar a diferença registrada, associadas com presença de frentes
(SEYMOUR et al., 2012; FONTES et al., 2018). Além do mais, seu menor tamanho e
menor demanda de nutrientes favorecem estes grupos tanto em condições
oligotróficas quanto eutróficas (TURA, 2015), fornecendo vantagem sobre outros
microrganismos nas mesmas condições. Estes fatos também corroboram para a
variação espacial (observada na Figuras 8), onde há a possibilidade de inferirmos
pequenos padrões. As amostras de inverno de superfície das estações 16 e 14 são
as mais distantes/isoladas dentre as amostras analisadas. Nelas foram observadas,
conforme visto e comentado anteriormente, grandes percentagens do filo e de
famílias de Cyanobacteria, o que pode evidenciar um distinto grupo tanto para
inverno quanto para verão como também para profundidades próximas à superfície.
No geral, o restante das amostras aparentou uma maior semelhança espacial, onde
as amostras de verão formaram um agrupamento mais junto do que as amostras de
inverno.
Diversos estudos na costa brasileira corroboraram com os dados encontrados
neste trabalho, assim como aqueles realizados em outras zonas costeiras
mundialmente (POMMIER et al., 2007; SUNAGAWANA et al., 2019; FUHRMAN &
STEELE, 2008; MILASLAVICH et al., 2011; THOMPSON et al., 2010), o que reforça
a predominância dos filos Proteobacteria e Bacteroidetes também na costa Sudeste
da América do Sul. Cyanobacteria já tem se mostrado como um filo bastante
relevante, chegando a se equiparar com Proteobacteria, em determinados períodos
do ano (GONÇALVES et al., 2009). Picos de Cyanobacteria encontradas no inverno
reforçam a importância de estudos dos processos oceanográficos físicos sobre o
bacterioplâncton, como observado em Fontes et al. (2018). O mesmo foi descrito por
Seymour et al. (2012) na costa leste da Austrália, onde houve predomínio de
Cyanobacteria na região de frente oceânica.
Rappé & Giavannoni (2003) discutem em seu estudo (The Uncultured
Microbial Majority) sobre a possibilidade de novas descobertas envolvendo bactérias
não cultivadas, tópico relevante para o presente estudo onde grande parte das

40
Cyanobactérias do período de inverno foram identificadas como pertencentes a
outros grupos/uncultured bacteria. Muitos destes microrganismos encontram-se nos
oceanos e são componentes essenciais de ciclos biológicos e geoquímicos (RAPPÉ
& GIAVANNONI, 2003). Contudo, apesar do avanço exponencial das metodologias
de classificação de sequenciamento genômico, a diversidade microbiana destes
clusters identificados como bactérias não cultivadas permanece um grande território
inexplorado, onde a importância majoritária microbiana destes microrganismos “não
cultivados” permanece obscura (RAPPÉ & GIAVANNONI, 2003).
6. CONCLUSÕES
O presente mostrou os grupos Proteobacteria, Cyanobacteria e Bacteroidetes
como predominantes nas regiões amostrais estudadas (tanto no período de verão
como no de inverno). A dominância de Proteobacteria e Bacteroidetes corrobora com
estudos realizados em diferentes ZCs ao redor do globo terrestre, onde estes dois
grupos dominam grande parte da estrutura da comunidade do bacterioplâncton.
Quanto ao grupo Cyanobacteria, sua expressão mostrou-se significativa,
especialmente se levado em consideração a classificação formulada pelo Workflow
do presente estudo, onde estes microrganismos encontram-se entre os três Filos
dominantes nas regiões analisadas. A estrutura da comunidade do bacterioplâncton
varia com o tempo e espaço, sendo que o grupo Cyanobacteria apresentou maior
número e também maior contribuição no período de inverno. Neste período houve
uma grande percentagem de Cyanobacteria não identificada, fato que, com
mencionado anteriormente, é algo relativamente novo e que necessita de estudos
mais detalhados não somente na região da REBIO Arvoredo e entorno, mas também
em outras regiões da ZC brasileira/Leste Sul Americana. Nos períodos de verão e de
inverno, Proteobacteria e Cyanobacteria alternavam-se na disputa pela dominância
nas profundidades superficiais e mais profundas, fato interessante que pode
demonstrar a variação da estrutura da comunidade do bacterioplâncton entre as
diferentes profundidades analisadas no presente estudo. Os resultados do presente
estudo ao redor da REBIO Arvoredo sobre a diversidade taxonômica do
bacterioplâncton apresentaram padrões semelhantes aos descritos para abundância
e biomassa bacteriana, associados também a variação de outros microrganismos

41
(como fitoplâncton) ligados à SACW no verão e influência da PPW no inverno. No
entanto, estudos mais detalhados são necessários para melhor elucidar todos os
fenômenos que ocorrem na região, especialmente os fatores que estão
influenciando a maior presença de Cyanobacterias não cultivadas/desconhecidas no
período de inverno.

42
7. REFERÊNCIAS
AZAM, F. et al., 1983. The ecological role of Water Column Microbes in the Sea.
Mar.Ecol.Prog.Ser., 10: 257-263.
AZAM, F. et al., 1994 Microb Ecol 28: 167.
AZAM, F.; MALFATTI, F., 2007. Microbial structuring of marine ecosystems.
Nature Reviews Microbiology. 5:782-791.
BRANDINI et al., 2018. Ecosystem responses to biogeochemical fronts in the
South Brazil Bight. Progress in Oceanography 164. p52–62.
CAPORASO J.G. et al., 2012. Ultrahigh-throughput microbial community
analysis on the Illumina HiSeq and MiSeq platforms. ISME J.
CODD, G. A. et al., 2005. Cyanobacterial toxins: risk management for health
protection. Toxicology and Applied Pharmacology 203: 264–272.
COTTRELL M T, KIRCHMAN D L., 2000. Natural assemblages of marine
proteobacteria and members of the Cytophaga-Flavobacter cluster consuming
low- and high-molecular-weight dissolved organic matter. Appl Environ
Microbiol;66:1692–1697.
CROSSLAND C.J. et al., 2005. The Coastal Zone — a Domain of Global
Interactions In Coastal Fluxes in the Anthropocene. Global Change — The IGBP
Series. Springer, Berlin, Heidelberg.
DELMONT et al., 2018. Nitrogen-fixing populations of Planctomycetes and
Proteobacteria are abundant in surface ocean metagenomes. Nature
Microbiology 3, 804–813.
DA SILVEIRA , I.C.A. et al., 2000. A Corrente do Brasil ao largo da Costa Leste
Brasileira. Rev. Bras. Oceanogr., 48(2): 171-183.

43
EDGAR, R.C., 2013. UPARSE: highly accurate OTU sequences from microbial
amplicon reads, "Nat. Meth. 10, 996-998".
ELIFANTZ, H. et al., 2005. Assimilation of Polysaccharides and Glucose by
Major Bacterial Groups in the Delaware Estuary. Applied and Environmental
Microbiology Dec 2005, 71 (12) 7799-7805.
ELIFANTZ, H. et al., 2007. Dissolved organic matter assimilation by
heterotrophic bacterial groups in the western Arctic Ocean. Aquatic Microbial
Ecology - AQUAT MICROB ECOL. 50. 39-49. 10.3354/ame01145.
ESTEVES, FA. 2011. Fundamentos de Limnologia. 3. ed. Rio de Janeiro:
Interciência. 826 p.
EUROPEAN ENVIRONMENT AGENCY. Acesso em 15 de abril de 2019. Disponível
em < https://www.eea.europa.eu/>.
FERNÁNDEZ-GÓMEZ, B. et al., 2013. Ecology of marine Bacteroidetes: a
comparative genomics approach. ISME J. 2013 May; 7(5): 1026–1037.
JEFFRIES et al., 2018. Bacterioplankton Dynamics within a Large
Anthropogenically Impacted Urban Estuary. Front. Microbiol., 26 January 2016.
FUERST, J., SAGULENKO, E., 201. Beyond the bacterium: Planctomycetes
challenge our concepts of microbial structure and function. Nature reviews.
Microbiology. 9. 403-13. 10.1038/nrmicro2578.
FUHRMAN, J.A., STEELE, J.A., 2008. Community structure of marine
bacterioplankton: patterns, networks, and relationships to function. Aquat
Microb Ecol 53:69-81.

44
GONÇALVES-ARAUJO, R. et. al., 2012. Brazil-Malvinas confluence: effects of
environmental variability on phytoplankton community structure. Journal of
plankton research, 34(5), 399-415.
GOLÇALVES, K.M., 2009. Caracterização da Dinâmica da Comunidade de
Bacterioplâncton no Estuário do Rio Amazonas (Canal do Norte) - AP.
Dissertação de Mestrado. Universidade Federal do Amapá.
GREGORACCI, G.B. et al., 2012. Structuring of Bacterioplankton Diversity in a
Large Tropical Bay. PLOS ONE 7(2): e31408.
GOM UFSC. Grupo de Oceanografia Microbiana da UFSC. Acesso em 20 maio de
2019. Disponível em <https://gomicrobes.wixsite.com/gomufsc>.
HUSZAR, V. L. M. & SILVA L. H. S., 1999. A estrutura da comunidade
fitoplanctônica no Brasil: cinco décadas de estudos. Limnotemas 2: 1–21.
ICMBio - Instituto Chico Mendes de Conservação da Biodiversidade. Acesso em
15 de abril de 2019. Disponível em <http://www.icmbio.gov.br/portal/> e
<http://www.icmbio.gov.br/portal/unidadesdeconservacao/biomas-
brasileiros/marinho/unidades-de-conservacao-marinho/2276-REBIO-marinha-do-
arvoredo>.
JONES et al., 2006. The New Bioinformatics: Integrating Ecological Data from
the Gene to the Biosphere. Annu. Rev. Ecol. Evol. Syst. 37:519–44.
KIM, Y. et al., 2018. Photosynthetic functions of Synechococcus in the ocean
microbiomes of diverse salinity and seasons. PLoS ONE 13(1).
KIRCHMAN, D.L. Processes in Microbial Ecology. 2ª edição. Oxford University
Press, Oxford, 2008.

45
KJERFVE, B. et al., 1997. Oceanographic characteristics of an impacted coastal
bay: Baía de Guanabara, Rio de Janeiro, Brazil. Continent. Shelf Res., v. 17, n.
13, p. 1609-1643, 1997.
MAArE : Monitoramento ambiental da Reserva Biológica Marinha do Arvoredo e
entorno / Organização: Bárbara Segal...[et al.]. – 1. ed. - Florianópolis :
UFSC/MAArE, 2017. 268 p.
MADSEN, E. L., 2011. Microorganisms and their roles in fundamental
biogeochemical cycles. Current Opinion in Biotechnology, 22:456-464.
MENEZES et al., 2019. Changes in the plankton community according to
oceanographic variability in a shallow subtropical shelf: SW Atlantic.
Hydrobiologia https://doi.org/10.1007/s10750-019-3936-5.
MILOSLAVICH,P. et al., 2011. Marine Biodiversity in the Atlantic and Pacific
Coasts of South America: Knowledge and Gaps. PLoS ONE 6(1): e14631.
NASA / National Aeronautics and Space Administration. Acesso em 15 Abril
2019. Disponível em <https://www.nasa.gov/>.
POMEROY, L. R., 1974. The Ocean's Food Web, A Changing Paradigm.
BioScience, Volume 24, Issue 9, September 1974, Pages 499–504.
POMMIER, T. et al., 2007. Global Patterns of Diversity and Community
Structure in Marine Bacterioplankton. Molecular Ecology. Volume 16: 867–
880.
PYLRO, V. S. et al., 2016. BMPOS: a Flexible and User-Friendly Tool Sets for
Microbiome Studies. Microbial Ecology, Volume 72, n. 2: 443-447.
QUAST C, et al., 2013. The SILVA ribosomal RNA gene database project:
improved data processing and web-based tools. Nucleic Acids Res.
2013;41(Database issue):D590–D596. doi:10.1093/nar/gks1219.

46
RAPPÉ, S. M. & GIAVANNONI, S. J., 2003. The uncultured microbial majority.
Annu. Rev. Microbiol. 2003. 57:369–94.
ROCAP, G., et al., 2003. Genome divergence in two Prochlorococcus ecotypes
reflects oceanic niche differentiation. Nature volume 424, pages1042–1047.
SEYMOUR, J.R. et al., 2012. Contrasting microbial assemblages in adjacent
water masses associated with the East Australian Current. Environ. Microbiol.
Rep., 4 (2012), pp. 548-555.
SILVEIRA, C. B. et al., 2011. Influence of Salinity on Bacterioplankton
Communities from the Brazilian Rain Forest to the Coastal Atlantic Ocean.
PLoS ONE 6(3).
SILVEIRA, J.D. (1964). Morfologia do Litoral. In: Azevedo, A. (ed). Brasil: a
terra e o homem. Companhia Editora Nacional, São Paulo. Volume 1:
253-305.
SUNAGAWA, S. et al,. 2019. Microbial abundance, activity and population
genomic profiling with mOTUs2. Nature Communications 10, Article number: 1014
(2019)
TAKAHASHI, S. et al., 2014. Development of a Prokaryotic Universal Primer for
Simultaneous Analysis of Bacteria and Archaea Using Next-Generation
Sequencing. PLOS ONE 9(8): e105592.
TESSLER, M., & GOYA, S. (2005). Processos Costeiros Condicionantes do
Litoral Brasileiro. Revista Do Departamento De Geografia, 17, 11-23.
THOMPSON, F. L. et al., 2011. Coastal bacterioplankton community diversity
along a latitudinal gradient in Latin America by means of V6 tag
pyrosequencing. Archives of microbiology 193: 105–114.

47
TURA, P. et al., 2015. Levels of 137Cs and 40K in marine superficial sediments
near the Angra Nuclear Power Plant (Angra dos Reis, SE Brazil). Radiochimica
Acta, v. 103, p. 729-735.
VITTE, A. C., 2003. O litoral brasileiro: a valorização do espaço e os riscos
socioambientais. Territorium: Revista Portuguesa de riscos, prevenção e
segurança. Nº 10 (2003) .
WASSMANN, P. & REIGSTAD, M., 2011. Future Arctic Ocean Seasonal Ice Zones
and Implications for Pelagic-Benthic Coupling. Oceanography Vol. 24, No. 3.
ZINGER, L. et al., 2011. Global Patterns of Bacterial Beta-Diversity in Seafloor
and Seawater Ecosystems. PLoS ONE 6(9).
ZHANG, J. et al., 2014. PEAR: a fast and accurate Illumina Paired-End
reAd mergeR. Oxford Journals, Bioinformatics. Mar 1; 30(5): 614–620.

48
8 APÊNDICE
8.1 Principais Táxons
Principais famílias e seus respectivos filos de bacterioplâncton (incluindo Bacteria e
Archaea) identificadas nos períodos de verão e inverno
Domínio Principais Filos
Verão
Principais Filos Inverno
Principais Famílias
Verão
Principais Famílias Inverno
Archaea
Euryarchaeota Euryarchaeota Marine Group II
Marine Group II
Thaumarchaeota Thaumarchaeota
Bacteria
Actinobacteria Actinobacteria OM1 clade OM1 clade
Bacteroidetes Bacteroidetes NS9 marine group Flavobacteriaceae Cryomorphaceae
NS9 marine group Flavobacteriaceae Cryomorphaceae
Chloroflexi Chloroflexi
Cyanobacteria Cyanobacteria
SubsectionI; FamilyI
Chloroplast; Outras ;
uncultured bacterium;
Outras ; uncultured bacterium
Deferribacteres Deferribacteres SAR406 clade
(Marine group A)
SAR406 clade (Marine group A)
Gemmatimonadetes Gemmatimonadete
s
Lentisphaerae Spirochaetae
Proteobacteria Proteobacteria
Alteromonadaceae Rhodobacteraceae Rhodospirillaceae
SAR11 clade; Surface 1
SAR116 clade SAR86 clade
Alteromonadaceae Rhodobacteraceae Rhodospirillaceae
SAR 11 clade; Surface 1
JL-ETNP-Y6 SAR116 clade SAR86 clade
Planctomycetes
Planctomycetes
Tenericutes Tenericutes Verrucomicrobia Verrucomicrobia
Não Classificados;
Outros Não Classificados;
Outros

49
8.2. Análises estatísticas
8.2a Testes Kruskal-Wallis de Filo

50
8.2b ANOVAs de Filo e Família (respectivamente)
8.2c ANOVAs individuais de Filo

51

52